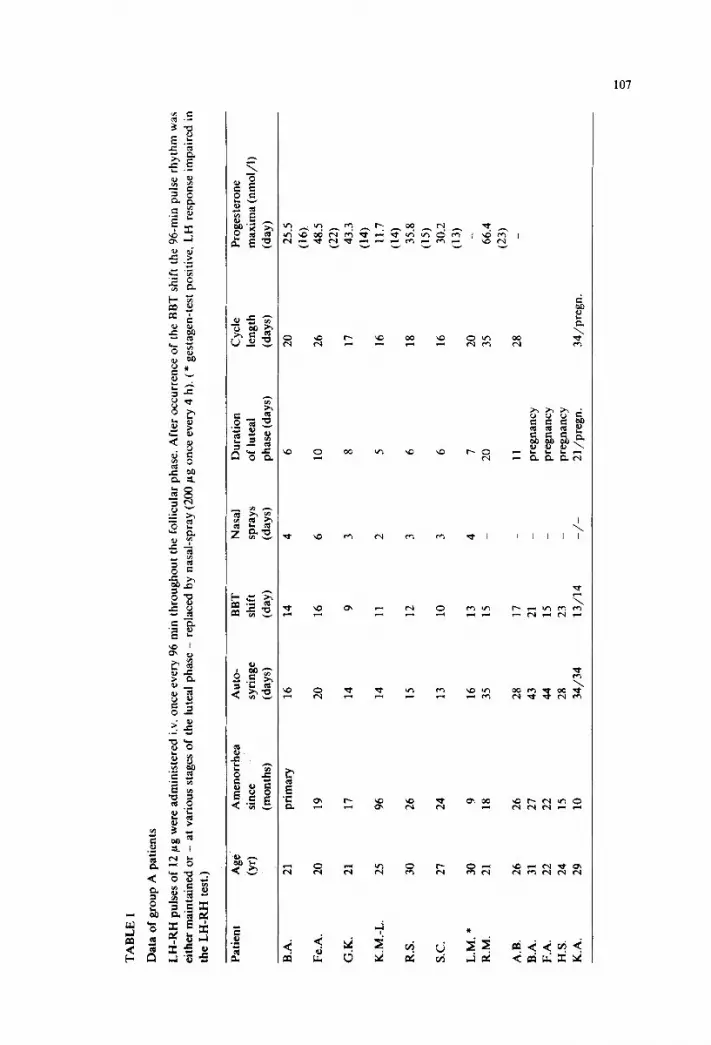
frequency-varied versus unvaried pulsatile lh-rh substitution in hypothalamic amenorrhea
TRANSCRIPT

Europ. J. Obsrer. Gynec. reprod Btol.. 17 (1984) 103- 119
Elsevier
103
EJO 00092
Frequency-varied versus unvaried pulsatile LH-RH substitution in hypothalamic amenorrhea
J.P. Hanker ‘, E. Nieschlag * and H.P.G. Schneider ’
’ Unioersrty Women’.s Hosprtal and
.’ Max Planck Chnrcal Research Unrt for Reproducrrve Medwne. Miinsrer. F. R.G
HANKER, J.P.. NIESCHLAG. E. and SCHNEIDER, H.P.G. (1984): Frequency-varied versus unvaried
pulsatile LH-RH substitution in hypothalamic amenorrhea. Europ. J. Obsrer. Gvnec. reprod. Biol.. /7/,7.3,
103-119.
In 26 patients with hypothalamic amenorrhea cyclic ovarian function was reestablished by pulsatile
LH-RH substitution. Two LH-RH regimens were tested. In group A (n = 13) LH-RH pulses (12 pg i.v.)
were administered by means of a portable mini-pump (Auto-Syringe) at the unvaried frequency of one
pulse every 96 min throughout the follicular phase. In group B (n = 13) LH-RH pulse intervals of 4 h (for
2 consecutive days), 3 h (for 1 day) and % min (for the rest of the follicular phase) were selected for
initiation of follicle growth. After ovulation pump-assisted pulsatile LH-RH substitution was either
maintained at the %-min rhythm or replaced by LH-RH nasal-spray (200 gg/sniff) applied once every 4
h until the onset of menstruation or detection of pregnancy. While in group A patients changes in the
pattern of luteal LH-RH support (pump-assisted versus nasal-spray assisted) resulted in rapid luteolysis.
such effects did not occur in group B patients. It is suggested that LH-RH pulse frequency at the initial
stage of follicular development is of importance for the quality of follicular development and thus luteal
function. Both LH-RH regimens resulted in pregnancies (six times singletons and once twins). In the
meantime, all children have been born and are healthy.
pulsatile LH-RH substitution; hypothalamic amenorrhea
Introduction
In a number of species, including the rat (Gay and Sheth, 1972), sheep (Butler et al., 1972; Katongole et al., 1974), cattle (Rahe et al.. 1980) and human (Nankin and Troen, 1971; Midgley and Jaffe, 1971; Yen et al., 1972; Santen and Bardin, 1973) it has been recognized that the secretion of luteinizing hormone (LH) and follicle-
stimulating hormone (FSH) from the pituitary is a rhythmic pulsatile phenomenon. In the ovariectomized rhesus monkey LH pulses are generated approximately once
every hour and, for this reason, the LH secretion pattern was termed “circhoral” (Dierschke et al., 1970). In the human female pulsatile LH secretion varies with the
stage of the cycle. While LH pulses occurring approximately once every l-2 h (Yen
Reprinr requesrs to: Priv.-Doz. Dr.med. J.P. Hanker.
OO28-2243/84/%03.00 0 1984 Elsevier Science Publishers B.V

104
et al., 1972; Santen and Bardin, 1973; Backstrom et al., 1982) characterize the follicular phase, a frequency of one LH pulse every 3-6 h is typical for the luteal phase.
Data obtained in the rat (Schuiling and Gnodde, 1976) rhesus monkey (Carmel et al., 1976; Neil1 et al., 1977) and human (Malacara et al., 1972; Seyler and Reichlin, 1974) undoubtedly show that pulsatile release of gonadotropins is the consequence of synchronous rhythmic discharges of LH-RH from the hypothalamus. Accordingly, the diminished LH pulse frequency during the luteal phase must be interpreted as a lower rate of endogenous LH-RH rhythms at this stage of the cycle.
In the human female various dysfunctional states of the hypothalamo-pituitary- gonadal axis - such as hyperprolactinemia or hypothalamic amenorrhea - are characterized by the absence of spontaneous LH episodes (Bohnet et al., 1974,1975, 1976; Schneider and Bohnet, 1974; Schneider et al., 1975, 1977). Thus, a deficient endogenous LH-RH signal has to be assumed as underlying cause of this disorder.
To reestablish ovulation in hypothalamic amenorrhea substitution of the de- capeptide seemed a logical consequence. Any therapy of this kind has to mimic physiological conditions as closely as possible to avoid paradoxic effects of LH-RH on gonadotropin secretion, i.e., desensitization of the gonadotrophs. In the female rhesus monkey it has been shown that LH and FSH secretion are LH-RH pulse frequency-coded (Knobil, 1980). The rhythmic pulsatile pattern of gonadotropin secretion does not, therefore, represent an inherent pulsatile phenomenon of the pituitary but rather an obligatorily intermittent functioning of a suprapituitary control system, that directs gonadotropin release (Pohl and Knobil, 1982).
To reinitiate gonadotropin secretion in patients with hypothalamic amenorrhea chronic-intermittent LH-RH stimulation was performed on them. Two substitution patterns were tested. In the first part of this study LH-RH pulses were administered at a frequency mimicking LH-RH release during the follicular phase of the cycle. In the second part LH-RH pulse frequency was varied in the beginning of the therapy. The rationale for the latter procedure is based upon the fact that follicular develop- ment is normally initiated during the late luteal phase of the preceding cycle (Baird, 1983). At this stage, endogenous LH-RH signals are operating at much lower frequency than during the follicular phase (evidenced by the reduced frequency of spontaneous LH episodes). Therefore, we decided to administer LH-RH pulses once every 4 h for 2 consecutive days, once every 3 h for the following day, and thereafter once every 96 min for the rest of the follicular phase. This pattern was meant to simulate to some degree the gradual acceleration of the frequency of LH-RH pulses occurring during the intermenstruum.
The effects of both LH-RH treatment schedules on luteal function were assessed.
Patients and methods
Twenty-six female outpatients (age 20-35 yr) presenting with longstanding (6-30 months duration, 3 subjects with primary amenorrhea) hypothalamic amenorrhea were selected for this study. Seventeen patients wanted to become pregnant.
Other endocrine disorders such as diabetes mellitus, thyroid or adrenal dysfunc- tion had been excluded. All patients were normoprolactinemic as assessed by

105
metoclopramide (10 mg) tests (Bohnet et al., 1980a,b). In no case was an abnormal X-ray of the sella obtained.
Three women responded to 5 mg of medroxyprogesterone acetate b.i.d. for 10 days by withdrawal-bleedings, whereas 23 failed to do so. In no case did three consecutive trials on clomiphene (50 mg/day for 5 days during 1st and 50 mg b.i.d. for 5 days during 2nd and 3rd) lead to ovulation. Thus, all women were classified as clomid-negative (Hull et al., 1979). In the LH-RH test (25 pg i.v.) the LH responses were impaired (absolute increase < 200 ng/ml, net increase 2 100 ng/ml) in 3 cases and absent (net increase < 100 ng/ml) in 23 (Keller et al., 1975).
The LH-RH treatment protocol was approved by the Human Ethics Committee of the Miinster University Medical Center. Written consent was obtained from all subjects after the procedure had been explained in full detail to them.
Throughout the whole follicular phase pulsatile LH-RH (12 pg/pulse iv.) administration was performed by means of a portable mini-pump (Auto-Syringe). Two groups (A and B) of patients were selected at random.
In group A (ir = 13) LH-RH was injected every 96 min throughout the whole follicular phase. Following occurrence of the hyperthermic shift in the basal body temperature (BBT) the 96-min rhythm of LH-RH pulsing was maintained in 6 women until the onset of menstruation or detection of pregnancy. In 7 women the 96-min rhythm was discontinued 2-5 days after the BBT shift and further substitu- tion was performed by means of an LH-RH nasal-spray (200 pg/sniff). This nasal-spray was applied once every 4 h until menstruation started. Before nasal-spray administrations were started each patient had practised the handling of the spray-de- vice. During therapy each nasal-spray application had to be crossed out on a test protocol.
In group B (n = 13) LH-RH pulses were administered once every 4 h during the first 2 days of treatment. During the 3rd day the frequency was increased to one LH+RH bolus every 3 h and thereafter to one LH-RH pulse every 96 min. In 5 patients the 96-min rhythm was maintained unvariedly until the onset of menstrua- tion (n = 3) or detection of pregnancy (n = 2). In 8 patients the 96-min rhythm of LH-RH pulsing was discontinued 2-6 days after the shift of the BBT had occurred and further LH-RH substitution was executed by nasal-spray (200 pg/sniff). This nasal-spray was applied once every 4 h until menstruation started (n = 7) or pregnancy was detected (n = 1).
In most patients blood samples could be obtained for monitoring of LH, FSH, estradiol (E,) and progesterone (P) secretion during therapy. These were obtained immediately before a pump-delivery or nasal-spray administration, centrifuged (3000 rpm for 20 min at 4’C) and stored at - 23’C until analysis. In 5 patients BBT charts, menstruations or pregnancies served to assess the therapeutic effects of the LH-RH treatment.
All hormones were measured by means of specific radioimmunoassays. For determination of LH the kit supplied by the National Institute of Arthritis, Metabo- lism and Digestive Diseases was used, and for FSH the commercially available kit from IRE, Fleurus, Belgium. The standards were LER 907 for LH and IRE-Stan- dard (1 ng 2 2.3 mIU/ml MRC 69/104) for FSH. The assay procedures for determination of estradiol (E,) and progesterone (P) are given elsewhere (Friedrich et al., 1974); the results are expressed in terms of pmol/l and nmol/l, respectively.

106
Results
The results obtained in group A are summarized in Table I. Fourteen cycles were induced. Hyperthermic shifts of the BBTs occurred 9-23 days after onset of therapy. In 3 cycles the LH-RH substitution rhythm of one pulse every 96 min was maintained until menstruation started and luteal phases lasting 11, 20 and 21 days
600
500
LOO
F 100
F 80 -I
1 60
LO
100 7 a
E 80
i 60
LO
20
0 2 6 10 1.4 18 22 26 30 31 38 13
t+ t
E c”
I% IL
0
b
2.0
1.2
1600 : 1LOO
1200
1000 q a 6
800 d w
600 H
LOO
200
+AUTO-SYRINGE -12,ugLH-RH IV 96mln intervals
Fig. 1. LH, FSH, estradiol (E,) and progesterone (P) levels in a patient (B.A.) with hypothalamic amenorrhea (negative gestagen-, clomiphene- and LH-RH test) of 27 months duration and who conceived
during frequency-unvaried pulsatile LH-RH treatment for a total of 43 days.
TA
BL
E
I
Dat
a of
gro
up
A p
atie
nts
LH
-RH
pu
lses
of
12
pg
wer
e ad
min
iste
red
iv.
once
ev
ery
96 m
in
thro
ugho
ut
the
folli
cula
r ph
ase.
A
fter
oc
curr
ence
of
th
e B
BT
sh
ift
the
96-m
in
puls
e rh
ythm
w
as
eith
er
mai
ntai
ned
or
- at
va
riou
s st
ages
of
th
e lu
teal
ph
ase
- re
plac
ed
by
nasa
l-sp
ray
(200
pg
on
ce
ever
y 4
h).
(* g
esta
gen-
test
po
sitiv
e,
LH
re
spon
se
impa
ired
in
the
LH
-RH
te
st.)
Patie
nt
Age
(W
Am
enor
rhea
A
uto-
B
BT
N
asal
D
urat
ion
Cyc
le
Prog
este
rone
sinc
e sy
ring
e sh
ift
spra
ys
of
lute
al
leng
th
max
ima
(nm
ol/l)
(mon
ths)
(d
ays)
(d
ay)
(day
s)
phas
e (d
ays)
(d
ays)
(d
ay)
B.A
. 21
pr
imar
y 16
14
Fe.A
. 20
19
20
16
G.K
. 21
17
14
9
K.M
.-L.
25
96
14
11
R.S
. 30
26
15
12
SC.
27
24
13
10
L.M
. *
30
9 16
13
R.M
. 21
18
35
15
A.B
. 26
26
28
17
B.A
. 31
27
43
21
F.
A.
22
22
44
15
H.S
. 24
15
28
23
K.A
. 29
10
34
/34
13/1
4
4 6 3 2 3 3 4 _ _ _ _ -/-
6 10 8 5 6 6 7
20
11
preg
nanc
y
preg
nanc
y
preg
nanc
y
2l/p
regn
.
20
26
17
16
18
16
20
35
28
34/p
regn
.
25.5
(16)
. 48
.5
(22)
43
.3
(14)
11
.7
(14)
35
.8
(15)
30
.2
(13)
66.4
(23)

108
e _...._._ 4
FSH fnghlll -
LH
lnglml)
ml
45
40
3.5
. . . .
3.0 ,AOl"l
25 -60
.50
40
2.0 .30
1.5 ,20
1.0 .lO
x5 -0
Fig. 2. LH, FSH, estradiol (E2) and progesterone (P) levels in a patient (R.M.) with hypothalamic
amenorrhea (negative gestagen-, clomiphene- and LH-RH test) of 18 months duration during pulsatile
LH-RH treatment (t . . . t) at unvaried pulse frequency (96-min intervals) for a total of 35 days. All
values are synchronized on the LH surge (0).
FSH
LH lnglmll lng/rnll ‘I)1 I
0 60 lx) lb 240 tlmnl 0 60 (10 tm 2LO t lrnhl
Fig. 3. Short-term LH fluctuations in the patient (R.M.) depicted in Fig. 2. LH concentrations (IO-min
intervals during a 4-h period) were recorded 1 day before (O- 0) pulsatile LH-RH (12 pg i.v. = 1) treatment as well as on days 1 (O------O), 8 (C-----XI) and 15 (m- n ) during therapy.
Fig. 4. Short-term FSH fluctuations in the patient (R.M.) depicted in Fig. 2. FSH concentrations (lo-min
intervals during a 4-h period) were recorded 1 day before (a- 0) pulsatile LH-RH (12 pg iv. = J) treatment as well as on days 1 (O------O), 8 (U-----O) and 15 (m- n ) during therapy.
109
- LH 300
(ng ml)
200
.__ __ __.
E2 150 ipmolll)
1000- 100 800.
6oo,t 80
LOO-
300. 60
200,;
LO
loo 20
o- 0 -8 -6 4 -2 0 +2 4 +6 +0 +l0 +12+X
0 . . . . . . __+
FSH Inglml)
3.5 o p o [nmolll)
30
20
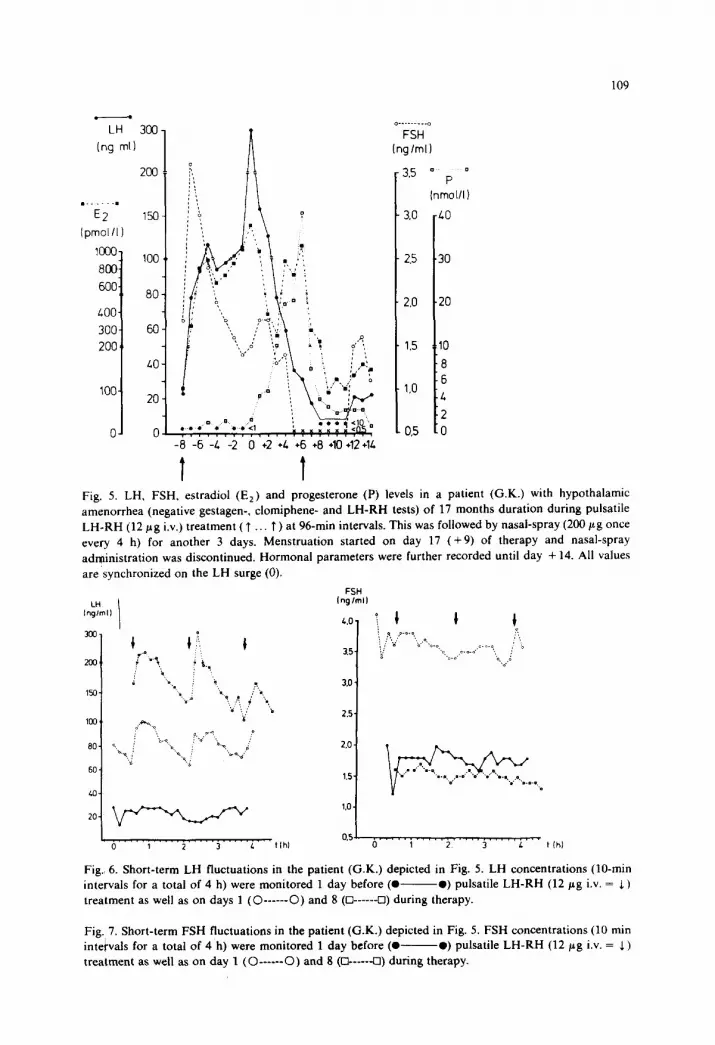
Fig. 5. LH, FSH, estradiol (Ez) and progesterone (P) levels in a patient (G.K.) with hypothalamic
amenorrhea (negative gestagen-. clomiphene- and LH-RH tests) of 17 months duration during pulsatile
LH.RH (12 pg i.v.) treatment ( T . . T ) at 96-min intervals. This was followed by nasal-spray (200 gg once
every 4 h) for another 3 days. Menstruation started on day 17 (+9) of therapy and nasal-spray
administration was discontinued. Hormonal parameters were further recorded until day + 14. All values
are synchronized on the LH surge (0).
FSH
LH I Ing/rldl
0 1 2 3 L tlhl 0 1 2 3 L t Ihl
Fig. 6. Short-term LH fluctuations in the patient (G.K.) depicted in Fig. 5. LH concentrations (IO-min
intervals for a total of 4 h) were monitored 1 day before (0 -0) pulsatile LH-RH (12 pg i.v. = 1)
treatment as well as on days 1 (O------O) and 8 (Cl------Cl) during therapy.
Fig. 7. Short-term FSH fluctuations in the patient (G.K.) depicted in Fig. 5. FSH concentrations (10 min
intervals for a total of 4 h) were monitored 1 day before (0 -0) pulsatile LH-RH (12 pg i.v. = 5)
treatment as well as on day 1 (O------O) and 8 (R-----U) during therapy.

6
I
~6
2 4 6 6012Yl6~iQ22242626B32YB36
I*- day of,!T~-’
.
t
l :prrgtnt*
l m.M)INOE (7,qLHIW cy Lh.Lh-3h-Wmm dw~&
rW.SPRIY 200.4 L~+RH~MI~~ 4h mtuxis
FSH q/ml
’ ‘lo 0 c..... 2 4 6 6 10 12 ,., I4 16 16 ,. 20 22 X 26 26
t4hM 96 mm t bui0-syT1rge
0 2 L 6 6 IO 12 14 16 18 20 22 Z 26 28
96mm t Auto-syrmge
Fig. 8. LH. FSH, estradiol (E,) and progesterone (P) levels in patient K.K.. presenting with hypothalamic amenorrhea (negative gcstagen-, clomiphene- and LH-RH test) of 8 months. who conceived during frequency-varied pulsatile LH-RH substitution.
Fig, 9. LH, FSH, estradiol (E2) and progesterone (Prop) levels in a patient (E.M.) with hypothalamic amenorrhea (gestagen-positive, clomiphene-negative, impaired LH response in the LH-RH test) during frequency-varied pulsatile LH-RH (12 pg i.v.) treatment for a total of 29 days.
were obtained. Four patients out of 9 who wanted to become pregnant conceived: in 3 (K.A., B.A. and F.A.) LH-RH had been substituted every 96 min for a total of 34, 43 and 44 days, respectively. In the meantime, all women have been delivered of healthy children following uneventful pregnancies. A representative case is depicted in Fig. 1. In one case (H.S.) the 96-min rhythm of LH-RH substitution was discontinued 5 days after the BBT rise and no further exogenous support of the corpus luteum was provided. While the BBT remained elevated pregnancy was confirmed 14 days later. After an uneventful pregnancy H.S. was delivered at term of a healthy child.
In 7 patients LH-RH pulsing at 96-min intervals was discontinued 2-5 days after the BBT rise. In these cases, further substitution was performed by LH-RH nasal-spray (200 pg once every 4 h) for another 2-6 days. It was a consistent feature that progesterone levels instantly declined following disruption of the 96-min rhythm

TA
BL
E
II
Dat
a of
gr
oup
B p
atie
nts
LH
-RH
pu
lse
freq
uenc
y w
as
vari
ed
in
the
begi
nnin
g of
th
erap
y (p
ulse
in
terv
als
of
4 h
for
2 da
ys.
3 h
for
1 da
y an
d 96
m
in
for
the
rest
of
th
e fo
llicu
lar
phas
e).
Aft
er
Occ
urre
nce
of t
he
BB
T
shif
t th
e 96
-min
pu
lse
rhyt
hm
was
ei
ther
m
aint
aine
d or
-
at
vari
ous
stag
es
of
the
lute
al
phas
e -
repl
aced
by
na
sal-
spra
y (2
00
pg
once
ever
y 4
h).
(* g
esta
gen-
test
po
sitiv
e,
LH
re
spon
se
impa
ired
in
th
e L
H-R
H
test
.)
Patie
nt
Age
A
men
orrh
ea
Aut
o-
BB
T
Nas
al
Dur
atio
n C
ycle
Pr
oges
tero
ne
(yr)
si
nce
syri
nge
shif
t sp
ray
of l
utea
l le
ngth
m
axim
a (n
mol
/l)
(mon
ths)
(d
ays)
(d
ay)
(day
s)
phas
e (d
ays)
(d
ays)
(d
ay)
B.B
. 24
11
14
8
6 12
20
61
.1
(15)
F.
A.
20
20
15
13
13
15
28
56.9
(17)
H
.A.
23
12
13
10
9 12
22
40
.1
(13)
H
.P.
23
30
17
14
7 10
24
58
.6
(19)
D
.L.
21
29
15
10
4 9
19
18.1
(14)
W
.S.
20
12
15
9 6
12
21
65.8
(15)
E
.M.
* 30
6
29
13
16
29
139.
8
(22)
B
.D.
23
prim
ary
25
18
_ 7
25
18.9
(21)
B
.C.
* 30
6
18
17
14
15
32
79.3
(24)
S.
M.
22
prim
ary
28
16
_ 12
28
46
.8
(22)
S.
Mo.
24
22
40
15
_
K.K
. 29
8
preg
nanc
y 15
13
23
A.-V
-W.
26
29
preg
nanc
y
20
18
28
preg
nanc
y
z

FSH 11. .ng/ml
-. 5 H.A
L
*<lo day 1 5 10 15 20
E2 pmol/l
2000
. hi L i3T 96nin T Lh 1
Auto-Syringe spray
Fig. 10. LH, FSH, estradiol (E2) and progesterone (Prop) levels in a patient (H.A.) with hypothalamic
amenorrhea (negative gestagen-, clomiphene- and LH-RH test) since 12 months during frequency-varied
pulsatile LH-RH treatment.
of pulsatile LH-RH support. Despite administration of LH-RH nasal-spray luteoly- sis proceded and luteal phases lasting 5-10 days only were obtained. In Figs. 2-7 two representative cases are depicted.
Table II summarizes the results of group B. Hyperthermic shifts of the BBTs occurred 8-18 days after pulsatile LH-RH substitution had been started. In 4 patients the 96-min rhythm of LH-RH pulsing was maintained unvariedly after emergence of the BBT rise: in 3 cases menstruations ensued 8, 12 and 16 days later and LH-RH was stopped; one patient conceived (S.M.) and LH-RH pulsing at 96-min intervals was terminated on day 40. After normal pregnancy SM. was delivered at term of a healthy child. In 9 patients the LH-RH substitution rhythm of one pulse every 96 min was discontinued l-6 days after the BBT rise and further luteal support was provided by LH-RH nasal-spray (200 pg once every 4 h). In 7 cases menstruations started after the nasal-spray had been administered for 4-14 days and luteal phases of 9-15 days were recorded. Two women (K.K. and A.-V.W.) conceived. In K.K. nasal-spray was started 2 days following the BBT rise and

113
maintained for another 23 days. This case is presented in detail in Fig. 8. In A.-V.W. the LH-RH nasal-spray was started on day 2 following the rise of her BBT and continued for another 28 days. At this time a positive pregnancy test was obtained and 6 wk later a twin pregnancy detected. Meanwhile, both patients have been delivered: K.K. of a healthy girl and A.-V.W. of healthy heterozygous twins.
While in group A patients the changing pattern of LH-RH substitution (pump-as- sisted at 96-min intervals versus nasal-spray-assisted at 4-h intervals) consistently resulted in rapid declines of progesterone levels, this phenomenon was not observed in group B patients. In the latter corpus luteum function persevered despite the considerable changes in the mode of LH-RH substitution. Other than in group A we observed in group B that the lifespans of the corpora lutea were mostly normal and that progesterone secretion was considerably enhanced. Two representative cases are depicted in Figs. 9 and 10.
Discussion
The human ovulatory cycle is characterized by two patterns of gonadotropin secretion: tonic or cyclic. Irrespective of these, LH and FSH are always secreted in episodic fashion (Yen et al., 1972; Santen and Bardin, 1973; Wildt et al., 1982). The physiological significance of episodic gonadotropin release has only recently been recognized as consequence of an obligatorily intermittent functioning of the hypo- physiotropic control system that directs gonadotropin secretion (Knobil, 1980).
In the primate this control system resides within the bilateral arcuate nuclei of the medio-basal hypothalamus (Plant et al., 1978; 1979). Destruction of this neuronal oscillator or LH-RH pulse generator rapidly abolishes gonadotropin secretion (Plant et al., 1979). While in such animals sustained gonadotropin release can be reinitiated by intermittent administration of exogenous LH-RH pulses, continuous infusion of the decapeptide fails to have this effect (Nakai et al., 1978). In turn, continuous infusion of LH-RH profoundly inhibits gonadotropin release, previously reestab- lished in such animals by pulsatile LH-RH administration (Belchetz et al., 1978).
There is no doubt that the intermittent nature of LH-RH stimulation represents the basic prerequisite for tonic gonadotropin secretion. The mechanism of cyclic gonadotropin release, however, is still controversially discussed. In the arcuate-le- sioned ovariectomized rhesus monkey frequency and amplitude (dose) -unvaried pulsatile LH-RH substitution suffices for the expression of an estradiol-induced positive feedback on LH release (Nakai et al., 1978). These findings lead to the conclusion that no additional central signal, i.e., additional LH-RH release, is required for the initiation of the midcycle LH-surge. Thus, it was suggested that LH-RH has only a permissive, albeit obligatory action on cyclic LH-release (Knobil, 1980). In pituitary stalk-sectioned rhesus monkeys in which Teflon barriers have been placed between the cut ends to secure complete separation of the pituitary from hypothalamic influence, chronic-intermittent LH-RH substitution alone does not result in ovulations. The addition of continuous 24-h LH-RH infusions in the presence of endogenous estradiol levels elevated to 150 pg/ml for 2 consecutive days, however, elicited LH surges in these animals. Consequently it was claimed that an additional LH-RH signal beside the chronic-intermittent one is necessary at midcycle for full expression of the LH surge (Norman et al., 1982).

114
The finding of abolished episodic LH release in hypothalamic amenorrhea (Schneider and Bohnet, 1974; Schneider et al., 1975, 1977) provides indirect evidence for reduced or deficient hypothalamic LH-RH secretion in this functional disorder. To a certain degree hypothalamic amenorrhea can thus be compared with functional states of rhesus monkeys in which the endogenous LH-RH pulse generator has been destroyed. To overcome the lack of the neurohormone in the diseased woman LH-RH substitution is indicated. The principle of chronic-intermittent LH-RH administration for treatment of hypothalamic amenorrhea was first realized by Leyendecker (1979). Since then, several reports have been published dealing with the subject of ovulation-induction and pregnancies by pulsatile LH-RH therapy in patients with amenorrhea of hypothalamic origin (Leyendecker et al., 1980a,b, 1983; Crowley and McArthur, 1980; Keogh et al., 1981; Schoemaker et al., 1981; Skarin et al., 1981; Reid et al., 1981; Berg et al., 1983). While in these studies LH-RH doses ranging from 2.5 to 20 pg per pulse had been utilized the selected pulse intervals of 90-120 min were rather similar and once decided upon, maintained unvariedly. Following ovulation luteal support was either provided by hCG or maintenance of LH-RH pulsing at the frequency by which follicle growth had been induced.
The data presented in this study support the findings of others that by chronic-in- termittent LH-RH substitution follicular growth and ovulation can be achieved in hypothalamic amenorrhea. The dose of 12 pg LH-RH per pulse we used originates from a study in normal volunteers, where we found that the repeated administration of doses higher than that resulted in rapid desensitization of the pituitary gland (Hanker et al., 1980).
In our group A patients ovulatory cycles were induced by LH-RH pulses administered at an unvaried frequency of the latter. While short-term fluctuations of LH and FSH were monitored at certain intervals during therapy, functional changes of the gonadotrophs (i.e., basal and LH-RH inducible gonadotropin release) were observed which resembled those of different stages of a normal follicular phase (Nillius and Wide, 1972; Yen et al., 1972b) and which culminated in LH-surges. No conclusion can be drawn as to whether these LH surges were the consequences of chronic intermittent stimulation with an unvaried amount of LH-RH. Since patients with hypothalamic amenorrhea are not hypothalamo-lesioned, i.e., endogenous LH-RH secretion is not definitely abolished, the possibility cannot be excluded that additional endogenous LH-RH signals were operative at midcycle. We feel that although ovulations and pregnancies - and thus obviously normal cycles - had been induced by frequency-unvaried pulsatile LH-RH administration, this therapeutic schedule of LH-RH does not permit any subtle inference of physiological principles of LH-RH secretion operating during the normal menstrual cycle.
In 3 cases (A.B., K.A. and R.M.) luteal support was provided by maintenance of the 96-min LH-RH pulse rhythm until menstruation ensued and normal lengths of the luteal phases were obtained. While in 3 patients, who conceived during therapy (B.A., F.A. and K.A.), the 96-min rhythm of LH-RH pulsing was extended far beyond the duration of a normal luteal phase, it was discontinued in patient H.S. 5 days after conception. In the latter no adverse effect on corpus luteum function became apparent following disruption of LH-RH support. In contrast to these findings, instant luteolysis occurred in those patients who did not conceive and in

115
whom the 96-min rhythm of pulsatile LH-RH stimulation was discontinued at different stages of the luteal phase. In none could luteolysis be prevented by LH-RH nasal-spray. Accordingly, short luteal phases were obtained in these patients.
In patients of group B folliculogenesis was induced by LH-RH pulses, the frequency of which had been decreased to 4- and 3-h intervals during the first 3 days of therapy and then changed to a 96-min pulse rhythm. In two women (K.K. and A.-V.W.) who conceived during therapy the 96-min frequency of LH-RH pulses was disrupted 2 days after ovulation and further substitution with the neurohormone performed by means of nasal-spray. In neither patient was luteal function affected by this considerable change of luteal LH-RH support. It is worthwhile recalling that in the non-pregnant patients of group A similar changes of the LH-RH substitution mode had deleterious effects on corpus luteum function. Furthermore, we observed normal luteal phases (in terms of both duration and progesterone secretion) in the majority of group B patients regardless of whether pump-assisted pulsatile LH-RH substitution (pulse intervals 96 min) was maintained throughout the whole luteal phase or discontinued at different stages of the latter and replaced by nasal-spray (pulse intervals 4 h).
It is tempting to ascribe the improved luteal function in patients of group B to the variation of LH-RH pulse frequency in the beginning of therapy. Following the concept of Ross (1976), Channing et al. (1978) and McNatty (1979) the corpus luteum is a direct continuation of follicle development. Therefore, preovulatory hormonal determinants are essential for postovulatory luteal function. Among those, FSH is of special importance during the very early stage of folliculogenesis (Mc- Natty, 1979). In the human female a significant increase of circulating FSH has been shown to precede menstruation by approx. 3 days (Wide et al., 1973). At this time of the menstrual cycle spontaneous LH pulses (reflecting synchronous LH-RH dis- charges from the hypothalamus) are usually generated once every 3 h. The LH pulse frequency gradually accelerates from one pulse every 4-6 h to every 60-90 min from the time of luteolysis up until the later intermenstruum. This alteration in LH frequency is associated with a continuing increase in FSH secretion.
In the arcuate lesioned rhesus monkey it was demonstrated that decreasing the frequency of exogeneous LH-RH pulses from one per hour to one every 3 h leads to declines of plasma LH levels while those of FSH increase (Wildt et al., 1981). These results permit the important conclusion that relatively small changes in the frequency of LH-RH pulses profoundly change LH and FSH concentrations in the peripheral circulation and by this mechanism have major effects on the ratio of FSH to LH.
The mechanism by which low-frequency LH-RH stimulation favors FSH release and the convergence of FSH and LH activity following gradual acceleration of LH+RH pulse frequency to the normal follicular rhythm could serve to explain why luteal function was improved in our group B patients. It is tempting to speculate that variation of LH-RH pulse frequency in the beginning of therapy improved the hormonal determinants of follicular development.
The fact that we could not detect significant differences in the FSH levels among both treatment groups is most likely due to the infrequent blood sampling as well as to the relatively insensitive FSH assays available. Brown (1978) has argued that within the same individual the difference in dose of gonadotropin between no effect

116
and stimulation of follicular development may be as little as 20% and that the “ threshold” dose necessary to maintain follicular development changes with the size of the follicle. Thus, minor changes in the concentrations of FSH in the spontaneous cycle probably have important biological effects on the ovary (Baird, 1983).
Moreover, we do not claim to have simulated the optimal pattern of LH-RH pulses governing initiation of follicle growth. However, we observed that changes in the LH-RH pulse frequency during the early follicular phase do have considerable impact on later luteal function. Thus, LH-RH pulse frequency variation might be of biological significance in the regulation of the normal menstrual cycle.
In the final analysis the question arises of whether the corpus luteum needs any support at all. The work of VandeWiele et al. (1970) provided evidence that the human corpus luteum requires a small but regular input of LH for its maintenance. Recently, however, Asch et al. (1982) demonstrated the occurrence of normal luteal phases in rhesus monkeys in which endogenous LH release had been abolished by hypophysectomy performed immediately after ovulation. In case these data hold for the human corpus luteum, too, the latter has to be considered as autoregulative. Our luteal support policy then needs revision. Consequently, any therapeutic aim will then focus on optimization of the preovulatory hormonal determinants of corpus luteum function. In this context variation of pulse frequency during pulsatile LH-RH stimulation may well be of critical importance.
Acknowledgements
We grately acknowledge the generous gift of LH-RH and LH-RH nasal-spray by Dr. M. von der Ohe, Hoechst AG, F.R.G. We are indebted to Mrs. H. Linnenbaum, Mrs. E. Kropp and Mrs. H. Kellerhoff for their excellent technical assistance.
References
Asch, R.H., Abou-Samra, M., Braunstein, G.D. and Pauerstein, C.J. (1982): Luteal function in hypo- physectomized rhesus monkeys. J. clin. Endocr. Metab., 55, 154-161.
Backstrom, C.T., McNeilly, A.S., Leask, R.M. and Baird, D.T. (1982): Pulsatile secretion of LH, FSH, prolactin, oestradiol and progesterone during the human menstrual cycle. Clin. Endocr., 17, 29-42.
Baird, D.T. (1983): Preovulatory follicle in ewes and women. J. Reprod. Fertil., 69, 343-352. Belchetz, P.E., Plant, T.M., Nakai, Y., Keogh, E.J. and Knobil, E. (1976): Hypophysial responses to
continuous and intermittent delivery of hypothalamic gonadotropin releasing hormone. Science, 202, 631-633.
Berg, D., Mickan, H., Michael, S., Doring, K., Gloning, K., Janicke, F. and Rjosk, H.K. (1983): Ovulation and pregnancy after pulsatile administration of gonadotropin releasing hormone. Arch. Gynak., 233, 205-210.
Bohnet, H.G., Dahlen, H.G. and Schneider, H.P.G. (1974): Hyperprolactinemia and pulsatile LH fluctuation. Acta endocr. (Kbh), Suppl. 184: 109-115.
Bohnet, H.G., Dahlen, H.G. and Schneider, H.P.G. (1975): Einfluss der Prolaktinkonzentration auf die tonische LH-Freisetzung (spiking). Arch. Gym&, 219, 592-593.
Bohnet, H.G., Dahlen, H.G., Wuttke, W. and Schneider, H.P.G. (1976): Hyperprolactinemic anovulatory syndrome. J. clin. Endocr. Metab., 42, 132-143.
Bohnet, H.G., Naber, N.G., Hanker, J.P., Schindler. A.E. and Schneider, H.P.G. (1980a): Modulation of pituitary LH and prolactin pools in amenorrheic women. I. Effects of high dose estrogen or gestagen treatment. Hormone metab. Res. 12, No. 8, 349-420.

117
Bohnet, H.G., Naber, N.G., Hanker, J.P., Keller, E. and Schneider, H.P.G. (1980b): Modulation of
pituitary LH and prolactin pools in amenorrheic women. II. Effects of low dose estrogen treatment.
Hormone metab. Res., 12, No. 9, 459-464.
Brown, J.B. (1978): Pituitary control of ovarian function - concepts derived from gonadotrophin therapy.
Aust. N. Z. J. Obstet. Gynaec., 18, 47-54.
Butler, W.R., Malven, P.W., Willet, L.B. and Bolt, D.J. (1972): Patterns of pituitary release and cranial
output of LH and prolactin in ovarectomized ewes. Endocrinology, 91, 793-801.
Carmel, P.W., Araki, S. and Ferin, M. (1976): Pituitary stalk portal blood collection in rhesus monkeys:
evidence for pulsatile release of gonadotropin-releasing hormone. Endocrinology, 99, 243-248.
Channing, C.P., Anderson, L.D. and Batta, SK. (1978): Follicular growth and development. Clin. Obstet.
Gynaec., 5 (2) 375-382.
Crowley, W.F.. Jr. and McArthur, J.W. (1980): Simulation of the normal menstrual cycle in Kallmman’s
syndrome by pulsatile administration of luteinizing hormone-releasing hormone (LH-RH). J. clin.
Endocr. Metab., 51, 173-175.
Dierschke, D.J., Bhattacharya, A.N., Atkinson, I.E. and Knobil, E. (1970): Circhoral oscillations of
plasma LH levels in the ovariectomized rhesus monkey. Endocrinology, 87, 850-853.
Friedrich, E., Jaeger-Whitegiver. E.R., Bider, M., Halver-Schmidt, H.. Pente, B., Pallai, P., Keller, E. and
Schindler, A.E. (1974): Standardization of specific radioimmunoassays for plasma estrone. estradiol.
progesterone and androstenedione. J. Steroid B&hem., 5, 305-312.
Gay, V.L. and Sheth, N.A. (1972): Evidence for a periodic release of LH in castrated male and female
rats. Endocrinology, 90, 158-162.
Hanker, J.P., Ende, U., Bohnet, H.G. and Schneider, H.P.G. (1980): Gonadotropin release after
administration of LH-RH during early follicular phase. Int. J. Fertil., 25, 100-105.
Hull, N.G.R., Knuth, U.A., Murray, M.A.F. and Jacobs, H.S. (1979): The practical value of the
progesterone challenge test, serum estradiol estimation or clinical examination in assessment of the
oestrogen state and response to clomiphene in amenorrhea. Brit. J. Obstet. Gynaec., 86, 799-805.
Katongole. C.B., Naftolin, F. and Short, R.V. (1974): Seasonal variations in blood luteinizing hormone
and testosterone levels in rams. J. Endocr., 60, 101-106.
Keller. E.. Dahltn. H.G., Friedrich, E., Bohnet, H.G., Richter, R., Joel, E.W., Schubring, G., Klemt, W..
Staemmler, H.J., Wyss, H.I., Schindler, A.E. and Schneider, H.P.G. (1975): Human pituitary gona-
dotropin index. I. Standardized LRH test criteria for evaluation of functional amenorrhea. J. clin.
Endocr. Metab., 40, 959-969.
Keogh. E.J., Mallal, S.A., Giles, P.F.H. and Evans, D.V. (1981): Ovulation induction with intermittent
subcutaneous LH-RH. Lancet, i, 147-151.
Knobil. E. (1980): Neuroendocrine control of the menstrual cycle. Recent Prog. Hormone Res., 36.
53-88.
Leyendecker, G. (1979): The pathophysiology of hypothalamic ovarian failure. Europ. J. Obstet. Gynec.
reprod. Biol.. 9, 175-189.
Leyendecker, G., Struve, T. and Plotz, E.J. (1980a): Induction of ovulation with chronic-intermittent
(pulsatile) administration of LH-RH in women with hypothalamic and hyperprolactinaemic amenor-
rhea. Arch. Gym%, 229, 177-190.
Leyendecker, G., Wildt, L. and Hansmann, M. (1980b): Pregnancies following chronic-intermittent
(pulsatile) administration of Gn-RH by means of a portable pump (Zyklomat) - A new approach to
the treatment of infertility in hypothalamic amenorrhea. J. clin. Endocr. Metab., 51, 1214-1216.
Leyendecker, G. and Wildt, L. (1983): Induction of ovulation with chronic intermittent (pulsatile)
administration of Gn-RH in women with hypothalamic amenorrhea. J. Reprod. Fertil., 69, 397-409.
Malacara, J.M., Seyler, E., Jr. and Reichlin, S. (1972): Luteinizing hormone-releasing factor activity in
peripheral blood from women during the midcycle luteinizing hormone ovulatory surge. J. clin.
Endocr. Metab., 34, 271-278.
McNatty. K.P. (1979): Follicular determinants of corpus luteum function in the human ovary. In:
Ovarian follicular and corpus luteum function, pp. 465-477. Editors: C.P. Channing, J.M. Marsh, and
W.A. Sadler. Plenum Press, New York.
Midgley. A.R. and Jaffe, R.B. (1971): Regulation of human gonadotropins. X. Episodic fluctuation of LH
during the menstrual cycle. J. clin. Endocr., 33, 962-969.
Nakai. Y.. Plant, T.M., Hess, D.L., Keogh, E.J. and Knobil, E. (1978): On the sites of the negative and

118
positive feedback action of estradiol in the control of gonadotropin secretion in the rhesus monkey. Endocrinology, 102,1008-1014.
Nankin, H.R. and Troen, P. (1971): Repetitive luteinizing hormone elevation in serum of normal men. J. clin. Endocr. Metab., 33, 558-566.
Neill, J.D., Patton, J.M.. Dailey, R.A., Tsou, R.C. and Tindall, G.T. (1977): LH-RH in pituitary stalk blood of rhesus monkeys; relationship to level of LH release. Endocrinology, 101, 430-434.
Nillius, S.J. and Wide, L. (1972): Variation in LH and FSH response to LH releasing hormone during the menstrual cycle. J. Obstet. Gynaec. Brit. Cwlth, 79, 865-873.
Norman, R.L., Gliessman, P., Lindstrom, S.A., Hill, J. and Spies, H.G. (1982): Reinitiation of ovulatory cycles in pituitary-stalk sectioned rhesus monkeus: evidence for a specific hypothalamic message for the preovulatory release of luteinizing hormone. Endocrinology, 111, 1874-1882.
Plant, T.M., Krey, L.C., Moossy, J., McCormack, J.T., Hess, D.L. and Knobil, E. (1978): The arcuate nucleus and the control of gonadotropin and prolactin secretion in the female rhesus monkey (Macaca mulatta). Endocrinology, 102, 52-62.
Plant, T.M., Moossy, J., Hess, D.L., Nakai, Y., McCormack, J.T. and Knobil, E. (1979): Further studies on the effects of lesions in the rostra1 hypothalamus on gonadotropin secretion in the female rhesus monkey (Macaca mulatta). Endocrinology, 105, 465-473.
Pohl, C.R. and Knobil, E. (1982): The role of the central nervous system in the control of ovarian function in higher primates. Ann. Rev. Physiol., 44, 583-593.
Rahe, C.H., Owens, R.E., Fleeger, IL., Newton, H.J. and Harms, P.G. (1980): Pattern of plasma luteinizing hormone in the cyclic cow: dependence upon the period of the cycle. Endocrinology, 107, 498-503.
Reid, R.L., Leopold, G.R. and Yen, S.S.C. (1981): induction of ovulation and pregnancy with pulsatile luteinizing hormone releasing factor: Dosage and mode of delivery. J. Fertil. Steril.. 36, 553-559.
Ross, G. (1976): Preovulatory determinants of human corpus luteum function. Europ. J. Obstet. Gynec. reprod. Biol., 6, 147-155.
Santen, R.J. and Bardin, C.W. (1973): Episodic luteinizing hormone secretion in men. Pulse analysis, clinical interpretation, physiologic mechanisms. J. Clin. Invest., 52, 2617-2628.
Schneider, H.P.G., Keller, E., Hanker, J.P. and Bohnet, H.G. (1977): Clinical use of LH-RH in hypothalamic amenorrhea. In: Progress in Reproductive Biology, Vol. 2, pp. 180-194. Karger, Basel.
Schneider, H.P.G., Bohnet, H.G. and Dahlen, H.G. (1975): Hypothalamic amenorrhea: an approach to diagnosis and therapy with LH-releasing hormone. In: Basic applications and clinical uses of hypothalamic hormones. Proceedings of the 1st International Symposion on Basic Applications and Clinical Uses of Hypothalamic Hormones, Madrid. Excerpta Medica Congress Series 374. 261-269.
Schneider, H.P.G. and Bohnet, H.G. (1974): LH-releasing hormone in diagnosis and treatment of hypothalamic amenorrhea. In: Recent Advances in Human Reproduction. Proceedings of the 1st International Congress on Human Reproduction. Rio de Janeiro. Excerpta Medica Congress Series 370.110-118.
Schoemaker, J., Simons, A.H.M., Burger, C.W., Delemarre, H.A., and VanKessel, H. (1981): Induction of ovulation with LH-FSH releasing hormone (Lh-RH). In: Follicular maturation and ovulation. Editors: R. Rolland, E.V. Van Hall, S.G. Hillier, K.P. McNatty and J. Schoemaker. Excerpta Medica Congress Series 560, 373-388.
Schuiling, G.A. and Gnodde, H.P. (1976): Site of origin of the pulsatile secretion of luteinizing hormone in long-term ovariectomized rats. J. Endocr., 70, 97-104.
Seyler, E.L. and Reichlin, S. (1974): Episodic secretion of luteinizing hormone releasing factor (LRF) in the human. J. clin. Endocr. Metab., 39, 471-478.
Skarin, G., Nillius, S.J. and Wide, L. (1981): Intermittent low dose luteinizing hormone-releasing hormone therapy for induction of normal ovulatory menstrual cycles in women with amenorrhea. In: Follicular maturation and ovulation. Editors: R. Rolland, E.V. Van Hall, S.G. Hillier, K.P. McNatty and J. Schoemaker. Excerpta Medica Congress Series 560, 398-406.
VandeWiele, R.L., Bonumil, J., Dyrenfurth, I., Ferin, M., Jewelevicz, R., Warren, M., Rizkailah, J. and Mikhail, G. (1970): Mechanisms regulating the menstrual cycle in women. Recent Progr. Hormone Res., 26, 63-95.
Wide. L., Nillius, S.J., Gemzell, C., and Roos, P. (1973): Radioimmunosorbent assay of follicle-stimulat- ing hormone and luteinixing hormone in serum and urine from men and women. Acta endocr., (Kbh), Suppl. 174, l-58.

119
Wildt, L., Hausler, A., Marshall, G., Hutchinson, J.S., Plant, T.M., Belchetz, P.E. and Knobil, E. (1981):
Frequency and amplitude of gonadotropin-releasing hormone stimulation and gonadotropin secretion
in the rhesus monkey. Endocrinology, 109, 376-385.
Wildt, L., Brensing, K.A. and Leyendecker, G. (1981): The changing pattern of pulsatile gonadotropin
secretion during the luteal and early follicular phase of the cycle. Acta endocr.. Supl. 246, 82-91.
Yen, S.S.C., Tsai, C.C.. Naftolin, F., Vandenberg, G. and Ajabor, L. (1972a): Pulsatile patterns of
gonadotropin release in subjects with and without ovarian function. J. clin. Endocr. Metab.. 34,
671-675.
Yen, S.S.C., Vandenberg, G., Rebar, R. and Ehara, Y. (1972b): Variation of pituitary responsiveness to
synthetic LRF during different phases of the menstrual cycle. J. clin. Endocr. Metab., 35, 931-934.