the rat pineal gland comprises an endocannabinoid system
TRANSCRIPT

The rat pineal gland comprises an endocannabinoid system
Introduction
The rhythmic release of norepinephrine (NE) from sympa-thetic nerve fibers into the pineal gland is the essentialstimulus for induction of melatonin biosynthesis in the
mammalian pineal gland at night [1]. Release of NE isstrongly increased after the onset of darkness andNE inducesmelatonin biosynthesis via well-defined, species-specific
transmembrane signaling pathways [2–8]. In rodents, NEcauses a/b-adrenergic receptor-mediated elevation of intra-cellular 3¢,5¢-cyclic adenosine monophosphate- and Ca2+-
levels [9–11]. This increase leads to the transcriptionalactivation of arylalkylamine N-acetyltransferase (AANAT),the key enzyme of melatonin biosynthesis, by protein kinase
A (PKA)-mediated phosphorylation of the transcriptionfactor cyclicAMPresponse element-binding protein (CREB)[2, 12–14]. Moreover, stimulation with NE elicits PKA-dependent phosphorylation of the AANAT protein required
for the interaction between AANAT and 14-3-3 protein[15, 16]. This interaction enhances the affinity of AANATto its substrate serotonin and protects AANAT from
proteasomal proteolysis [3, 16, 17].
Melatonin biosynthesis is turned off before the end of the
night via multiple mechanisms. In rodents, transcription ofthe Aanat gene is down-regulated and this process correlateswith a strong decline of pCREB levels in the second part of
the night [2] and an induction of the inhibitory transcriptionfactor inducible cyclic AMP early repressor (ICER) [2,18–20]. As shown in cultured rat pineal glands, dephosphory-
lation of pCREB is mediated by serine/threonine proteinphosphatase 1 and occurs within several minutes after NEwithdrawal [21]. In addition to transcriptional down-regula-
tion ofAanat, terminationofmelatonin biosynthesis involvespost-translational modifications of the AANAT protein asAANAT is rapidly destructed by proteasomal proteolysisimmediately after light exposure during the dark phase or
after withdrawal of NE [17, 21].In a recent study, we found that NE-dependent
activation of AANAT and melatonin biosynthesis was
attenuated by application of the phytocannabinoids tetra-hydrocannabinol, cannabidiol, or cannabinol in culturedrat pineal glands [22]. These observations raise the
question whether an endocannabinoid system is presentin the pineal gland.
Abstract: In the mammalian pineal gland, the rhythm in melatonin
biosynthesis depends on the norepinephrine (NE)-driven regulation of
arylalkylamine N-acetyltransferase (AANAT), the penultimate enzyme of
melatonin biosynthesis. A recent study showed that phytocannabinoids like
tetrahydrocannabinol reduce AANAT activity and attenuate NE-induced
melatonin biosynthesis in rat pineal glands, raising the possibility that an
endocannabinoid system is present in the pineal gland. To test this
hypothesis, we analyzed cannabinoid (CB) receptors and specific enzymes for
endocannabinoid biosynthesis or catabolism in rat pineal glands and
cultured pinealocytes. Immunohistochemical and immunoblot analyses
revealed the presence of CB1 and CB2 receptor proteins, of N-acyl
phosphatidyl ethanolamine hydrolyzing phospholipase D (NAPE-PLD), an
enzyme catalyzing endocannabinoid biosynthesis and of fatty acid amide
hydrolase (FAAH), an endocannabinoid catabolizing enzyme, in
pinealocytes, and in pineal sympathetic nerve fibers identified by double
immunofluorescence with an antibody against tyrosine hydroxylase. The
immunosignals for the CB2 receptor, NAPE-PLD, and FAAH found in
pinealocytes did not vary under a 12 hr light:12 hr dark cycle. The CB1
receptor immunoreaction in pinealocytes was significantly reduced at the end
of the light phase [zeitgeber time (ZT) 12]. The immunosignal for NAPE-
PLD found in pineal sympathetic nerve fibers was reduced in the middle of
the dark phase (ZT 18). Stimulation of cultured pinealocytes with NE
affected neither the subcellular distribution nor the intensity of the
immunosignals for the investigated CB receptors and enzymes. In summary,
the pineal gland comprises indispensable compounds of the endocannabinoid
system indicating that endocannabinoids may be involved in the control
of pineal physiology.
Marco Koch, Iris Habazettl,Faramarz Dehghani andHorst-Werner Korf
Dr. Senckenbergische Anatomie, Institut fur
Anatomie II, Johann Wolfgang Goethe-
Universitat, Frankfurt am Main, Germany
Key words: cannabinoid receptors, fatty acid
amide hydrolase, N-acyl phosphatidyl
ethanolamine hydrolyzing phospholipase D,
norepinephrine, pineal gland, sympathetic
nerve fibers
Address reprint requests to Horst-Werner Korf,
Dr. Senckenbergische Anatomie, Institut fur
Anatomie II, Johann Wolfgang Goethe-
Universitat Frankfurt, Theodor-Stern-Kai 7,
Frankfurt am Main 60590, Germany.
E-mail: [email protected]
Received February 22, 2008;
accepted April 16, 2008.
J. Pineal Res. 2008; 45:351–360Doi:10.1111/j.1600-079X.2008.00597.x
� 2008 The AuthorsJournal compilation � 2008 Blackwell Munksgaard
Journal of Pineal Research
351

Endocannabinoids form a family of endogenous lipidmessengers which are involved in the regulation of manyphysiological and pathological processes in mammals [23,
24]. The first identified endocannabinoid was arachidonoyl-ethanolamine (AEA; [25]). Like most endocannabinoids,AEA acts upon specific Gi/o-coupled cannabinoid (CB) 1and 2 receptors [26]. In addition, endocannabinoids interact
with binding partners that do not belong to the CB receptorfamily. Thus, AEA also functions as an intracellularmessenger and regulates neurotransmitter- and voltage-
gated ion channels [27]. The lifespan of endocannabinoidsis strongly limited. They are synthesized on demand by amachinery of specific enzymes. The enzyme catalyzing the
formation of AEA is the N-acyl phosphatidyl ethanolaminehydrolyzing phospholipase D (NAPE-PLD; [28]). Aftersynthesis and release, endocannabinoids diffuse passivelyback into the cell or are taken up actively by membrane
transporters [29, 30]. Within the cell endocannabinoids arerapidly degraded by intrinsic enzymes. The specific enzymefor degradation of AEA is the fatty acid amide hydrolase
(FAAH; [31]). To date, it has not yet been establishedwhether CB1 and CB2 receptors, NAPE-PLD, and FAAHare expressed in the mammalian pineal gland. We have
therefore investigated these proteins in rat pineal glandsand pinealocytes by means of immunohistochemistry andimmunoblot to clarify the presence of an endocannabinoid
system in the mammalian pineal gland.
Materials and methods
Animals
All animal experimentation was carried out in accordancewith the Policy on the Use of Animals in NeuroscienceResearch and the Policy on Ethics as approved by the
Society for Neuroscience and by the European Commu-nities Council directive (89/609/EEC). Male Wistar rats(170–200 g; Charles River, Sulzfeld, Germany) were
housed in groups of four animals under 12 hr light:12 hrdark cycle for 2 wk. Dim red light remained on during thedark periods to allow handling of the animals. Animalswere deeply anesthetized with isoflurane and killed by
decapitation or transcardial perfusion at zeitgeber times(ZT) 0, 6, 12, or 18 with lights on at ZT0 and lights off atZT12.
Pineal cell and organ culture
After decapitation of the animals at ZT6, their pinealglands were removed and immediately transferred to ice-cold Earl�s balanced salt solution supplemented with 10 mm
HEPES and 7 g/L glucose. For organ cultures, pineal
glands were incubated at 37�C in Dulbecco�s modifiedEagle�s medium containing 5% (v/v) fetal calf serum andsupplemented with 10 mm HEPES, 100 lg/mL ascorbic
acid, 100 lg/mL streptomycin, 100 U/mL penicillin, and2 mm glutamine for 24 hr. For cell cultures, pineal glandswere dissociated by papain digestion and immobilized onto
0.01% (v/v) poly-l-lysine treated glass coverslips. After24 hr, pineal glands and pinealocytes were stimulated with1 lm NE for 6 hr or left untreated as controls.
Antibodies and blocking peptides
The following antibodies and corresponding blockingpeptides were used: rabbit polyclonal antibody against apeptide sequence near the extracellular part of theN-terminus of human CB1 receptor (diluted 1:50 for
immunohistochemistry/1:500 for immunoblot; CaymanChemicals, Ann Arbor, MI, USA; catalog number anti-body 101500/blocking peptide 301500); rabbit polyclonal
antibody against human CB2 receptor amino acid sequence20–33 (NPMKDYMILSGPQK; diluted 1:100/1:1000; Cay-man Chemicals; catalog number antibody 101550/blocking
peptide 301550); rabbit polyclonal antibody against humanNAPE-PLD amino acid sequence 378–390 (LKHGES-RYLNNDD; diluted 1:500/1:5000; Cayman Chemicals;
catalog number antibody 10005430/blocking peptide10009737); rabbit polyclonal antibody against rat FAAHamino acid sequence 561–579 (CLRFMRE-VEQLMTPQKQPS; diluted 1:50/1:500; Cayman Chemi-
cals, catalog number antibody 101600/blocking peptide301600); and sheep polyclonal antibody against tyrosinehydroxylase (TH; diluted 1:2000 for immunohistochemis-
try; Chemicon, Temecula, CA, USA; catalog numberAB1542).
Specificity tests for the antibodies used
The specificity of the antibodies was tested by preabsorbingthem with the corresponding blocking peptides. For
preabsorption, each primary antibody was diluted inphosphate-buffered saline (PBS) and incubated with afivefold excess (by weight) of its blocking peptide for 2 hr at
room temperature with gentle shaking. Thereafter, theantibody-blocking peptide solution was applied to thebrain sections and the subsequent immunohistochemical
procedures followed the protocol described below.
Immunohistochemistry, immunocytochemistry, andconfocal laser scanning microscopy
Animals were deeply anesthetized at the indicated ZT withisoflurane and perfused transcardially with 4% (w/v)
para-formaldehyde, according to standard protocols.Whole brains with the pineal gland attached were removed,postfixed overnight, cryoprotected with a 30% (w/v)
solution of sucrose, cut in the coronal plane on a cryostat(20 lm), and mounted on gelatin-coated slides. Dissociatedand immobilized pinealocytes were fixed with 4% (w/v)
p-formaldehyde in PBS for 10 min. Sections and cellpreparations were boiled in 0.01 m citrate buffer, pH 6,for 15 min in a microwave oven at 700 W and incubatedwith the primary antibodies against the CB1 and CB2
receptor proteins, NAPE-PLD, and FAAH overnight.Binding of the primary antibodies was visualized by meansof the ABC method with a biotin-conjugated anti-rabbit
IgG (diluted 1:100; Sigma-Aldrich, St. Louis, MO, USA)as the second antibody, a horseradish peroxidase (HRP)-conjugated streptavidin complex (diluted 1:100; Sigma-
Aldrich) and 3, 3¢-diamino-benzidine as the chromogen.For immunofluorescence, Alexa Fluor 546 donkey anti-rabbit IgG (diluted 1:500; Invitrogen, Karlsruhe, Germany)
Koch et al.
352

was used to detect immunoreactions for CB1 and CB2receptor proteins, NAPE-PLD, and FAAH. For doubleimmunofluorescence, brain sections including the pineal
organ were first incubated with one of the primaryantibodies against CB1, CB2, NAPE-PLD, or FAAH andthereafter with the primary antibody against TH. Bindingof the antibodies against CB1, CB2, NAPE-PLD, or
FAAH was visualized by means of an Alexa Fluor 488donkey anti-rabbit IgG (diluted 1:200) and binding of theantibody against TH was visualized by an Alexa Fluor 546
donkey anti-sheep IgG (diluted 1:500). The preparationswere coverslipped with Dako fluorescent mounting medium(Dako Diagnostika GmbH, Hamburg, Germany) and
analyzed using a Zeiss LSM 510 confocal imaging systemequipped with a helium/neon laser and an Axiovert 100Zeiss microscope (Zeiss, Halbergmoos, Germany).
Semiquantitative and statistical analysis
The intensity of the immunosignals was semiquantitatively
assessed by measuring the mean optical density (OD) with acomputer-assisted image analysis system (KS400; Zeiss).Images of pineal glands were obtained at 100· magnifica-
tion and were digitized at a resolution of 1024 · 1024pixels. The immunoreactive areas were then segmented,converted to binary images, and automatically measured.
Each analysis was performed with four sections from atleast three independent experiments. The value for themean OD of black (gray value = 0) was set as 100% andall other levels were calculated as percent of this maximum.
The NAPE-PLD immunoreaction in intrapineal nervefibers was determined by measuring the total area (FLD-area) of these fibers in randomly chosen fields of the pineal
glands at 200· magnification. Statistical analysis includedthe one-way ANOVA test followed by Bonferroni post-testfor multiple comparisons using GraphPad Prism (Graph-
Pad Prism Software Inc., San Diego, CA, USA). Data wereexpressed as OD [%] + S.E.M. (for immunosignals in thetotal pineal area) or FLDarea + S.E.M. (for NAPE-PLDimmunoreactive intrapineal nerve fibers) and values were
considered significantly different with P < 0.05.
Immunoblot analysis
For immunoblot analysis protein extracts were obtained bysonication of cultured pineal glands in lysis buffer contain-
ing 9.1 mm NaH2PO4, 1.7 mm Na2HPO4, 150 mm NaCl,0.5% (w/v) Na-deoxycholate, and 0.5% (v/v) nonidet P40(NP40), pH 7.4. Cell debris was removed by centrifugation
for 10 min at 10,000 g. Protein concentrations of thesupernatants were determined according to the method ofBradford [32]. Sample buffer was added to the preparationsand equal amounts of protein (30 lg per lane) were loaded
onto a 10% (w/v) sodium dodecyl sulfate–polyacrylamidegel. After gel electrophoresis, the proteins were blotted ontonitrocellulose membranes. The nitrocellulose membranes
were incubated with the above-described antibodies againstthe CB1 and CB2 receptors, NAPE-PLD and FAAH.Binding of the primary antibodies was visualized by use of
HRP-conjugated secondary antibodies and chemilumines-cence (Ultra Signal; Pierce, Rockford, IL, USA).
Results
Immunohistochemical analyses of brain sections revealedthe presence of CB1 receptor, CB2 receptor, NAPE-PLD,and FAAH in the rat pineal gland in situ. A clear CB1receptor immunoreaction was detected in virtually all rat
pinealocytes (Fig. 1A). The nucleus remained unstained.Within a single pinealocyte, the CB1 receptor immunosig-nal appeared as fine granules that were predominantly
found in the pineal processes and in the outer areas of thecytoplasm (Fig. 1B). Double immunofluorescence showedthat a CB1 receptor protein immunoreaction was also
present on intrapineal nerve fibers, which displayed apositive TH immunoreaction (Fig. 2A–C). All immunosig-nals were abolished when the sections were incubated withthe antibody preabsorbed with the blocking peptide,
indicating the specificity of the CB1 receptor immunosignalin rat pinealocytes and intrapineal nerve fibers (Fig. 1C).Numerous rat pinealocytes also displayed a CB2 receptor
immunoreaction (Fig. 1D), but when compared with theCB1 receptor immunosignal, the CB2 receptor immuno-reaction appeared weaker. It was evenly distributed in the
outer cytoplasmatic areas of the pinealocytes and appearedin fine granules. The nuclei of the pinealocytes remainedunstained (Fig. 1E). Double immunofluorescence showed
an immunoreaction for the CB2 receptor protein onintrapineal nerve fibers which displayed a positive THimmunoreaction (Fig. 2D–F). All immunosignals wereabolished when the sections were incubated with the
antibody preabsorbed with the blocking peptide, indicatingthe specificity of the CB2 receptor immunosignal in ratpinealocytes and intrapineal nerve fibers (Fig. 1F).
Virtually all pinealocytes showed a strong and homog-enous NAPE-PLD immunoreaction apparently associatedwith the nucleus (Fig. 1G). In addition, a strong NAPE-
PLD immunoreaction was detected in fine intrapineal nervefibers (Fig. 1H) and appeared to be very dense and locallyconcentrated in the varicosities and terminals. As shown bydouble immunofluorescence the NAPE-PLD immunoreac-
tive nerve fibers also contained TH immunoreactivity(Fig. 2G–I). Preabsorption of the NAPE-PLD antibodywith the blocking peptide abolished the described NAPE-
PLD immunreaction in pinealocytes and in intrapinealnerve fibers (Fig. 1I).Application of the FAAH antibody elicited an evenly
distributed, fine granular immunosignal in the cytoplasm ofpinealocytes (Fig. 1J). The nuclei did not show FAAHimmunoreaction. The intensity of the cytoplasmatic FAAH
immunosignals varied among individual pinealocytes. Somepinealocytes were stained more intensively than others(Fig. 1K). Double immunofluorescence revealed that THimmunoreactive nerve fibers also contained a FAAH
immunoreaction (Fig. 2J–L). Preabsorbtion of the FAAHantibody with the blocking peptide abolished the describedFAAH immunoreaction (Fig. 1L).
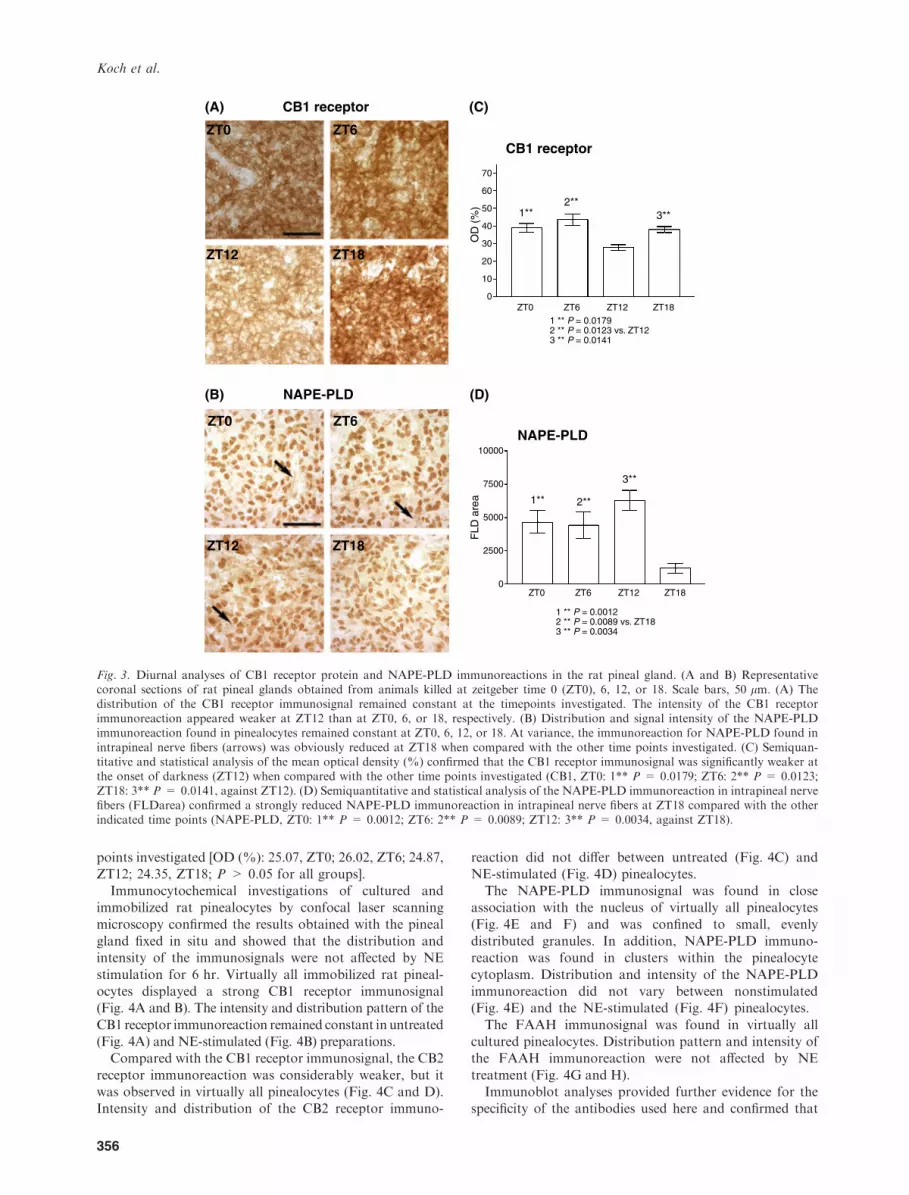
Distribution and intensity of the CB1 receptor, CB2receptor, NAPE-PLD, and FAAH immunoreactions wereanalyzed in the rat pineal gland in situ at ZT0, 6, 12, and 18
with light on at ZT0 and light off at ZT12. The distributionof the CB1 receptor immunosignal remained constantunder diurnal conditions, but the intensity of the CB1
Endocannabinoid system in rat pineal gland
353

receptor immunoreaction was weaker at ZT12 when
compared with ZT0, 6, or 18, respectively (Fig. 3A).Semiquantitative analyses of the mean OD (%) revealedthat the CB1 receptor immunosignal was significantly
weaker at the onset of darkness [OD (%): 28.75, ZT12]when compared with the other investigated time points [OD(%): 39.10, ZT0; 46.66, ZT6; 37.99, ZT18; P < 0.05 versusZT12; Fig. 3C).
In contrast to the CB1 receptor immunoreaction, the
intensity of the CB2 receptor immunosignal remainedconstant at the different time points as statisticalanalysis of the OD for the CB2 receptor immunoreaction
did not show significant differences [OD (%): 47.88,ZT0; 34.19, ZT6; 31.99, ZT12; 40.54, ZT18; P > 0.05for all groups]. The distribution of the finegranular, cytoplasmatic CB2 receptor immunoreaction
(A) (B) (C)
(D) (E) (F)
(G) (H) (I)
(J) (K) (L)
Fig. 1. Immunohistochemical demonstration of CB1 receptor protein, CB2 receptor protein, NAPE-PLD, and FAAH in coronal brainsections obtained from male rats kept under 12 hr light:12 hr dark cycle with light on at zeitgeber time 0 (ZT0) and light off at ZT12. Scalebars, 50 lm. (A) Virtually all pinealocytes displayed a strong CB1 receptor immunoreaction in the cytoplasm whereas the nuclei did notshow an immunosignal for the CB1 receptor. (B) The staining appeared in the outer areas of the cytoplasm (stars) and in bouton-likestructures (arrows). (C) Sections of rat pineal glands which were incubated with preabsorped CB1 receptor antibodies did not show anyimmunosignal indicating the specificity of the used CB1 receptor antibody. (D) When compared with the CB1 receptor immunoreaction, theintensity of the CB2 receptor staining appeared weaker. It was exclusively found in the cytoplasm of numerous pinealocytes. (E) The CB2receptor immunosignal was primarily associated with the outer region of the cytoplasm and the plasma membrane. (F) Preabsorption withthe corresponding blocking peptide for the CB2 receptor antibody abolished the CB2 receptor immunoreaction. (G) NAPE-PLD immu-noreaction was found in virtually all pinealocytes. (H) The NAPE-PLD immunoreaction was associated with the pinealocyte nuclei and wasalso found in varicosities and terminals of intrapineal nerve fibers running in the perivascular space (arrows). (I) Preabsorption of theNAPE-PLD antibody with the corresponding blocking peptide abolished the NAPE-PLD immunoreaction. (J) FAAH immunoreaction wasevenly distributed in the pineal gland. (K) The FAAH signal was exclusively located in the cytoplasm of numerous pinealocytes (stars).(L) Preabsorption of the FAAH antibody with the corresponding blocking peptide abolished the FAAH immunoreaction.
Koch et al.
354

also remained constant at ZT0, 6, 12 and 18 (data notshown).
Intensity and distribution of the NAPE-PLD immuno-signal in pinealocytes remained constant at ZT0, 6, 12,
or 18 [OD (%): 26.47, ZT0; 33.55, ZT6; 29.24, ZT12;28.88, ZT18; P > 0.05]. At variance, the NAPE-PLDimmunosignal in the intrapineal nerve fibers was weaker
at ZT18 (Fig. 3C). Semiquantitative analysis of the
FLDarea revealed that the NAPE-PLD immunoreactionwithin the intrapineal nerve fibers was significantlyreduced at ZT18 (FLDarea: 1063, ZT18) when comparedwith the other time points investigated (FLDarea: 4407,
ZT0; 3775, ZT6; 6437, ZT12; P < 0.05 versus ZT18;Fig. 3D).The distribution and intensity of the FAAH immuno-
reaction in the pinealocytes remained constant at all time
(A) (B) (C)
(D) (E) (F)
(G) (H) (I)
(J) (K) (L)
Fig. 2. Double immunofluorescence of CB1 receptor protein, CB2 receptor protein, NAPE-PLD, or FAAH (each indicated by greenfluorescence) and tyrosine hydroxylase (TH, indicated by red fluorescence) in coronal brain sections obtained from male rats kept under12 hr light:12 hr dark cycle with light on at zeitgeber time 0 (ZT0) and light off at ZT12. Analyses were performed by confocal laser scanningmicroscopy. Scale bars, 100 lm. (A) Virtually all rat pinealocytes showed a strong cytoplasmatic CB1 receptor immunoreaction (green).(B) Intrapineal nerve fibers were strongly stained with the TH antibody (red). The TH immunoreaction appeared to accumulate at distinctareas within the nerve fibers. (C) In the overlay a colocalization of CB1 receptor protein (green) and TH (red) immunoreactions is indicatedby yellow pixels (arrows). (D) A cytoplasmatic CB2 receptor immunoreaction (green) was detected in virtually all rat pinealocytes. (E andF) Within intrapineal nerve fibers, the CB2 receptor immunoreaction was selectively colocalized with a TH immunoreaction (red) asdemonstrated by the yellow pixels observed in the overlay (arrows). (G) NAPE-PLD immunoreaction (green) was associated with the nucleiof virtually all pinealocytes and was also found within intrapineal nerve fibers. (H) The nerve fibers dispalyed a strong TH immunoreaction(red). (I) At distinct areas, the TH immunoreaction was colocalized with NAPE-PLD immunoreaction as indicated by the yellow pixelsobserved in the overlay (arrows). (J and K) FAAH immunoreaction (green) was detectable in pinealocytes and intrapineal nerve fibers thatalso showed a strong TH immunoreaction. (L) Within the nerve fibers, the FAAH immunoreaction was frequently colocalized with THimmunoreaction (red), as illustrated by the yellow pixels obtained from the overlays (arrows).
Endocannabinoid system in rat pineal gland
355
points investigated [OD (%): 25.07, ZT0; 26.02, ZT6; 24.87,ZT12; 24.35, ZT18; P > 0.05 for all groups].Immunocytochemical investigations of cultured and
immobilized rat pinealocytes by confocal laser scanning
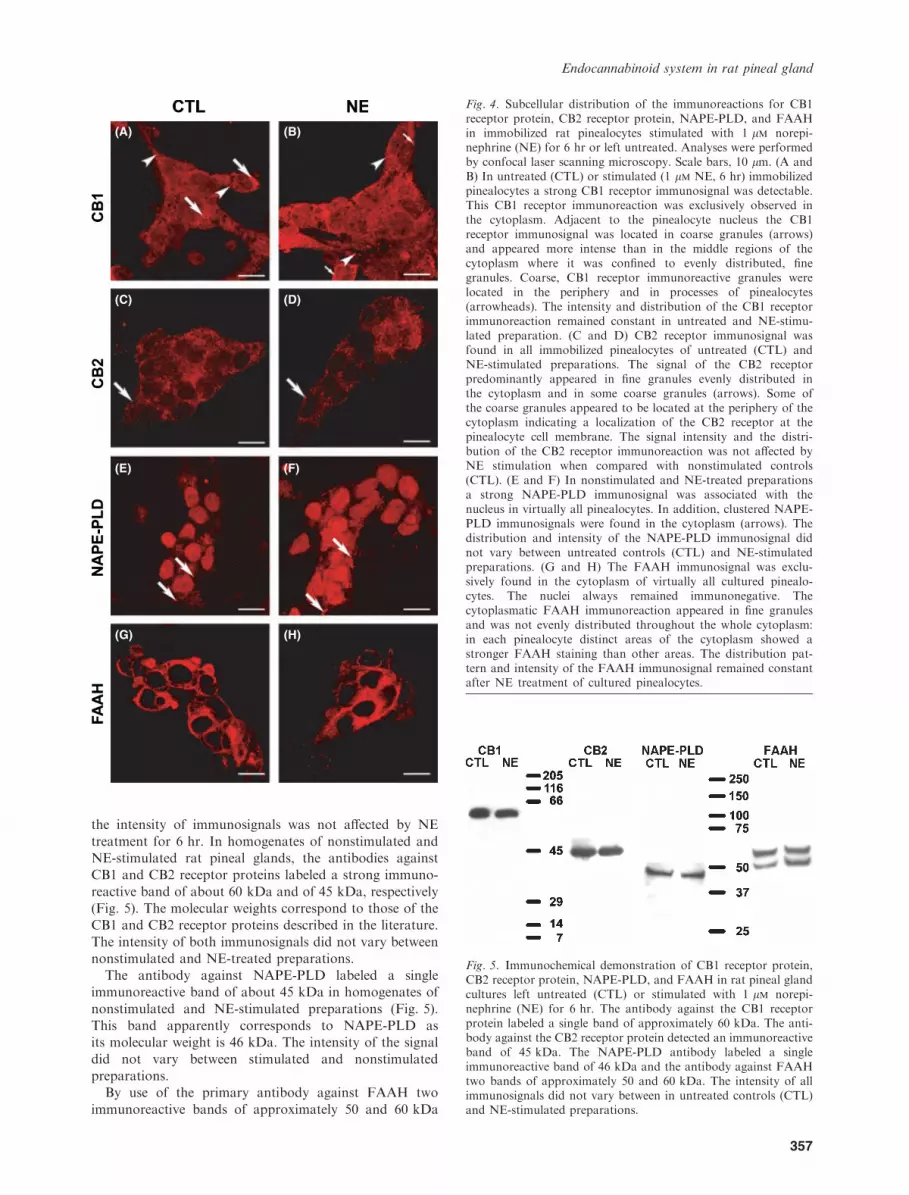
microscopy confirmed the results obtained with the pinealgland fixed in situ and showed that the distribution andintensity of the immunosignals were not affected by NE
stimulation for 6 hr. Virtually all immobilized rat pineal-ocytes displayed a strong CB1 receptor immunosignal(Fig. 4A and B). The intensity and distribution pattern of the
CB1 receptor immunoreaction remained constant in untreated(Fig. 4A) and NE-stimulated (Fig. 4B) preparations.Compared with the CB1 receptor immunosignal, the CB2
receptor immunoreaction was considerably weaker, but itwas observed in virtually all pinealocytes (Fig. 4C and D).Intensity and distribution of the CB2 receptor immuno-
reaction did not differ between untreated (Fig. 4C) andNE-stimulated (Fig. 4D) pinealocytes.The NAPE-PLD immunosignal was found in close
association with the nucleus of virtually all pinealocytes
(Fig. 4E and F) and was confined to small, evenlydistributed granules. In addition, NAPE-PLD immuno-reaction was found in clusters within the pinealocyte
cytoplasm. Distribution and intensity of the NAPE-PLDimmunoreaction did not vary between nonstimulated(Fig. 4E) and the NE-stimulated (Fig. 4F) pinealocytes.
The FAAH immunosignal was found in virtually allcultured pinealocytes. Distribution pattern and intensity ofthe FAAH immunoreaction were not affected by NE
treatment (Fig. 4G and H).Immunoblot analyses provided further evidence for the
specificity of the antibodies used here and confirmed that
CB1 receptor(A) (C)
CB1 receptor
NAPE-PLD(B) (D)
NAPE-PLD
1**
1**2**
3**
2**
3**
10000
0
10
20
30
40
50
60
70
7500
5000
FLD
are
aO
D (
%)
2500
0ZT0 ZT6 ZT12 ZT18
ZT0 ZT6 ZT12 ZT18
1 ** P = 0.00122 ** P = 0.0089 vs. ZT183 ** P = 0.0034
1 ** P = 0.01792 ** P = 0.0123 vs. ZT123 ** P = 0.0141
ZT6ZT0
ZT18ZT12
ZT6ZT0
ZT12 ZT18
Fig. 3. Diurnal analyses of CB1 receptor protein and NAPE-PLD immunoreactions in the rat pineal gland. (A and B) Representativecoronal sections of rat pineal glands obtained from animals killed at zeitgeber time 0 (ZT0), 6, 12, or 18. Scale bars, 50 lm. (A) Thedistribution of the CB1 receptor immunosignal remained constant at the timepoints investigated. The intensity of the CB1 receptorimmunoreaction appeared weaker at ZT12 than at ZT0, 6, or 18, respectively. (B) Distribution and signal intensity of the NAPE-PLDimmunoreaction found in pinealocytes remained constant at ZT0, 6, 12, or 18. At variance, the immunoreaction for NAPE-PLD found inintrapineal nerve fibers (arrows) was obviously reduced at ZT18 when compared with the other time points investigated. (C) Semiquan-titative and statistical analysis of the mean optical density (%) confirmed that the CB1 receptor immunosignal was significantly weaker atthe onset of darkness (ZT12) when compared with the other time points investigated (CB1, ZT0: 1** P = 0.0179; ZT6: 2** P = 0.0123;ZT18: 3** P = 0.0141, against ZT12). (D) Semiquantitative and statistical analysis of the NAPE-PLD immunoreaction in intrapineal nervefibers (FLDarea) confirmed a strongly reduced NAPE-PLD immunoreaction in intrapineal nerve fibers at ZT18 compared with the otherindicated time points (NAPE-PLD, ZT0: 1** P = 0.0012; ZT6: 2** P = 0.0089; ZT12: 3** P = 0.0034, against ZT18).
Koch et al.
356
the intensity of immunosignals was not affected by NE
treatment for 6 hr. In homogenates of nonstimulated andNE-stimulated rat pineal glands, the antibodies againstCB1 and CB2 receptor proteins labeled a strong immuno-
reactive band of about 60 kDa and of 45 kDa, respectively(Fig. 5). The molecular weights correspond to those of theCB1 and CB2 receptor proteins described in the literature.
The intensity of both immunosignals did not vary betweennonstimulated and NE-treated preparations.
The antibody against NAPE-PLD labeled a singleimmunoreactive band of about 45 kDa in homogenates of
nonstimulated and NE-stimulated preparations (Fig. 5).This band apparently corresponds to NAPE-PLD asits molecular weight is 46 kDa. The intensity of the signal
did not vary between stimulated and nonstimulatedpreparations.
By use of the primary antibody against FAAH two
immunoreactive bands of approximately 50 and 60 kDa
(A) (B)
(C) (D)
(E) (F)
(G) (H)
Fig. 4. Subcellular distribution of the immunoreactions for CB1receptor protein, CB2 receptor protein, NAPE-PLD, and FAAHin immobilized rat pinealocytes stimulated with 1 lm norepi-nephrine (NE) for 6 hr or left untreated. Analyses were performedby confocal laser scanning microscopy. Scale bars, 10 lm. (A andB) In untreated (CTL) or stimulated (1 lm NE, 6 hr) immobilizedpinealocytes a strong CB1 receptor immunosignal was detectable.This CB1 receptor immunoreaction was exclusively observed inthe cytoplasm. Adjacent to the pinealocyte nucleus the CB1receptor immunosignal was located in coarse granules (arrows)and appeared more intense than in the middle regions of thecytoplasm where it was confined to evenly distributed, finegranules. Coarse, CB1 receptor immunoreactive granules werelocated in the periphery and in processes of pinealocytes(arrowheads). The intensity and distribution of the CB1 receptorimmunoreaction remained constant in untreated and NE-stimu-lated preparation. (C and D) CB2 receptor immunosignal wasfound in all immobilized pinealocytes of untreated (CTL) andNE-stimulated preparations. The signal of the CB2 receptorpredominantly appeared in fine granules evenly distributed inthe cytoplasm and in some coarse granules (arrows). Some ofthe coarse granules appeared to be located at the periphery of thecytoplasm indicating a localization of the CB2 receptor at thepinealocyte cell membrane. The signal intensity and the distri-bution of the CB2 receptor immunoreaction was not affected byNE stimulation when compared with nonstimulated controls(CTL). (E and F) In nonstimulated and NE-treated preparationsa strong NAPE-PLD immunosignal was associated with thenucleus in virtually all pinealocytes. In addition, clustered NAPE-PLD immunosignals were found in the cytoplasm (arrows). Thedistribution and intensity of the NAPE-PLD immunosignal didnot vary between untreated controls (CTL) and NE-stimulatedpreparations. (G and H) The FAAH immunosignal was exclu-sively found in the cytoplasm of virtually all cultured pinealo-cytes. The nuclei always remained immunonegative. Thecytoplasmatic FAAH immunoreaction appeared in fine granulesand was not evenly distributed throughout the whole cytoplasm:in each pinealocyte distinct areas of the cytoplasm showed astronger FAAH staining than other areas. The distribution pat-tern and intensity of the FAAH immunosignal remained constantafter NE treatment of cultured pinealocytes.
Fig. 5. Immunochemical demonstration of CB1 receptor protein,CB2 receptor protein, NAPE-PLD, and FAAH in rat pineal glandcultures left untreated (CTL) or stimulated with 1 lm norepi-nephrine (NE) for 6 hr. The antibody against the CB1 receptorprotein labeled a single band of approximately 60 kDa. The anti-body against the CB2 receptor protein detected an immunoreactiveband of 45 kDa. The NAPE-PLD antibody labeled a singleimmunoreactive band of 46 kDa and the antibody against FAAHtwo bands of approximately 50 and 60 kDa. The intensity of allimmunosignals did not vary between in untreated controls (CTL)and NE-stimulated preparations.
Endocannabinoid system in rat pineal gland
357

were detected in homogenates of nonstimulated andNE-stimulated preparations (Fig. 5). The upper immuno-signal presumably corresponds to the predicted molecular
weight of FAAH (63 kDa). The intensity of the FAAHimmunoreaction did not vary between the untreated andNE-stimulated pineal organ cultures.
Discussion
Our immunohistochemical and immunochemical studies
show the presence of endocannabinoid receptor proteinsand enzymes involved in synthesis and metabolism ofendocannabinoids in the rat pineal gland. Immunoblotting
and immunohistochemical controls with preabsorbed anti-bodies indicate the specificity of the immunosignals. Thedata suggest an important functional role of endocannabi-noids for pineal functions in mammals and add the
mammalian pineal organ to other neuroendocrine systemswhose activity is modulated by endocannabinoids, i.e., thehypothalamus-pituitary-adrenal axis and the hypothala-
mus-pituitary-gonadal axis [33, 34].The CB1 receptor protein was found in pinealocytes
and TH immunoreactive intrapineal nerve fibers. These
results extend previous studies demonstrating the presenceof the CB1 receptor on neurons in various brain areas,e.g., in the olfactory bulb, the hippocampal formation, or
the basal ganglia [35–38]. One CB1 receptor antibodydirected against the C-terminal region of the proteindecorated the axons of different neuronal cell types [38],whereas another antibody directed against the N-terminal
region of the CB1 receptor protein also elicited animmunosignal in the perikarya of neocortical, hippocam-pal or cerebellar neurons [36, 37]. We used an antibody
directed against the N-terminal region of the CB1 receptorand found that, within the pinealocytes, the CB1 receptorprotein immunoreaction was predominantly located at the
nuclear envelope, at the periphery of the cytoplasm and inprocesses. These findings indicate that the CB1 receptorprotein immunoreaction is detectable at sites of proteinbiosynthesis (endoplasmatic reticulum and Golgi appara-
tus) and the cell membrane. In addition CB1 receptorprotein immunoreaction was observed on intrapinealnerve fibers that contain TH immunoreactivity and
apparently correspond to postganglionic sympatheticaxons originating from the superior cervical ganglion.These results conform to previous studies showing the
presence of CB1 receptors on postganglionic sympatheticnerve fibers in other organs [39]. Interestingly, endocann-abinoids were shown to suppress the release of NE from
postganglionic sympathetic nerve fibers via their action onCB1 receptors [40, 41] and a similar mechanism mayoperate in intrapineal sympathetic nerve fibers.The intensity of the CB1 receptor protein immunoreac-
tion in the pineal showed a time-dependent variation: it wassignificantly reduced at ZT12, i.e., the onset of darkness. Itis known from other systems that CB1 receptor gene
expression and protein levels can be regulated by hormones,feeding conditions or drug administration. The steroidhormone 17b-estradiol led to an up-regulation of CB1
receptor mRNA and protein levels in different primarytumor colon cancer cells [42]. In mice, caloric restriction
increased CB1 receptor protein levels in the striatum andhypothalamus whereas a high cholesterol-enriched dietdecreased CB1 receptor protein levels in the same brain
areas [43]. In rats, a significant decrease in the CB1 receptorgene expression was found after ethanol administration inthe caudate-putamen, the central amygdala, and the ven-tromedial hypothalamic nucleus [44]. CB1 receptor agonists
are able to control CB1 receptor density: prolongedexposure to CB receptor agonists caused a CB1 receptordesensitization followed by CB1 receptor internalization
[45, 46]. To investigate whether such mechanisms are alsoresponsible for reduction of the CB1 receptor protein levelsin the pineal gland endocannabinoid levels in the pineal
need to be determined by tandem-mass spectroscopy atdifferent time points.In contrast to the CB1 receptor, the CB2 receptor was
first thought to be absent from the healthy brain as it was
only found on activated microglial cells after brain injury[47]. However, this concept has been revised after CB2receptors were found on brainstem neurons [48] and in
nerve fibers of the granule cell layer in the cerebellum [49].In a more systematic analysis, CB2 receptor mRNA andprotein were found in numerous brain areas, e.g., on
hippocampal pyramidal neurons and cerebellar Purkinjecells [50]. By means of the same antibody as used by Gonget al. we observed CB2 immunoreactions in rat pinealocytes
and in intrapineal postganglionic sympathetic nerve fibersand our immunochemical analyses with cultured rat pinealglands revealed an immunoreactive band whose molecularweight is identical to that found by Gong et al. in brain and
spleen. As preabsorption of the antibody with the corre-sponding blocking peptide prevented CB2 receptor immu-nosignals in the rat pineal gland we conclude that the CB2
receptor is functionally expressed in pinealocytes andintrapineal postganglionic sympathetic nerve fibers. Incontrast to the CB1 receptor the signal intensity of the
CB2 receptor protein did not vary under diurnal conditionsin the pineal. This observation suggests that the densities ofthe two receptor subtypes are regulated via different
mechanisms.To investigate the source of endocannabinoids in the
pineal, we analyzed the arachidonoylethanolamine (AEA)synthesizing enzyme NAPE-PLD. Immunoreaction of
NAPE-PLD was present in pinealocytes indicating thatthese cells are capable to synthesize AEA. The distributionand signal intensity of NAPE-PLD within pinealocytes did
not vary upon NE stimulation in vitro or under diurnalconditions. Surprisingly, NAPE-PLD immunoreaction wasalso observed on intrapineal TH immunoreactive nerve
fibers. This result suggests that AEA synthesis also occurswithin sympathetic postganglionic nerve fibers and raisesthe possibility that AEA may be a sympathetic cotransmit-ter. This hypothesis which would add a novel player to the
control mechanisms of sympathetic neuronal activity needsto be verified in future experiments. Notably, the NAPE-PLD immunoreactivity was minimal in the middle of the
dark phase and this might suggest that synthesis of AEAwhich might inhibit NE release via its actions on CB1receptors (see above) is reduced during darkness.
Fatty acid amide hydrolase, an enzyme involved inendocannabinoid catabolism, was observed in pinealocytes
Koch et al.
358

and intrapineal sympathetic fibers and the intensity of theFAAH immunoreaction did not vary under diurnalconditions in vivo or after NE treatment in cultured
pinealocytes. Notably, the activity of FAAH was signif-icantly decreased in hippocampus and striatum at nightand this reduction may account for the nocturnal increaseof AEA levels in different brain areas, e.g., the nucleus
accumbens, hippocampus, prefrontal cortex, and thestriatum [51, 52].
In conclusion, our study provides clear experimental
evidence for the presence of a functional endocannabi-noid system in mammalian pinealocytes. Moreover, wehave demonstrated immunosignals for CB1 and CB2
receptors and for enzymes involved in endocannabinoidsynthesis and metabolism on intrapineal sympatheticfibers. These findings indicate that endocannabinoidsmay play an important role for the control of pineal
function at the level of both pinealocytes and sympatheticnerve fibers.
The colocalization of CB receptor proteins and enzymes
involved in AEA metabolism in rat pinealocytes andintrapineal sympathetic nerve fibers suggests that AEAfunctions as paracrine or autocrine messenger within the
pineal gland as has been shown for other systems [53, 54].
References
1. Klein DC, Berg GR, Weller J, Glinsmann W. Pineal
gland: dibutyryl cyclic adenosine monophosphate stimulation
of labeled melatonin production. Science 1970; 167:1738–1740.
2. Maronde E, Pfeffer M, Olcese J et al. Transcription
factors in neuroendocrine regulation: rhythmic changes in
pCREB and ICER levels frame melatonin synthesis. J Neu-
rosci 1999; 19:3326–3336.
3. Schomerus C, Korf HW, Laedtke E et al. Selective adren-
ergic/cyclic AMP-dependent switch-off of proteasomal prote-
olysis alone switches on neural signal transduction: an example
from the pineal gland. J Neurochem 2000; 75:2123–2132.
4. Schomerus C, Korf HW. Mechanisms regulating melatonin
synthesis in the mammalian pineal organ. Ann NY Acad Sci
2005; 1057:372–383.
5. Ackermann K, Bux R, Rub U et al. Characterization of
human melatonin synthesis using autoptic pineal tissue.
Endocrinology 2006; 147:3235–3242.
6. Ribelayga C, Pevet P, Simonneaux V. Hiomt drives the
photoperiodic changes in the amplitude of the melatonin peak
of the Siberian hamster. Am J Physiol Regul Integr Comp
Physiol 2000; 278:R1339–R1345.
7. Johnston JD, Bashforth R, Diack A et al. Rhythmic mel-
atonin secretion does not correlate with the expression of
arylalkylamine N-acetyltransferase, inducible cyclic amp early
repressor, period1 or cryptochrome1 mRNA in the sheep
pineal. Neuroscience 2004; 124:789–795.
8. Liu T, Borjigin J. N-acetyltransferase is not the rate-limiting
enzyme of melatonin synthesis at night. J Pineal Res 2005;
39:91–96.
9. Babila T, Schaad NC, Klein DC. Rat pineal Gsa, Gia and
Goa: relative abundance and development. Brain Res 1992;
572:232–235.
10. Vanecek J, Sugden D, Weller J, Klein DC. Atypical syn-
ergistic alpha 1- and beta-adrenergic regulation of adenosine
3¢,5¢-monophosphate monophosphate and guanosine 3¢,5¢-
monophosphate in rat pinealocytes. Endocrinology 1985;
116:2167–2173.
11. Schomerus C, Laedtke E, Korf HW. Calcium responses of
isolated, immunocytochemically identified rat pinealocytes to
noradrenergic, cholinergic and vasopressinergic stimulations.
Neurochem Int 1995; 27:163–175.
12. Borjigin J, Wang MM, Snyder SH. Diurnal variation in
mRNA encoding serotonin N-acetyltransferase in pineal
gland. Nature 1995; 378:783–785.
13. Tamotsu S, Schomerus C, Stehle JH et al. Norepinephrine-
induced phosphorylation of the transcription factor CREB in
isolated rat pinealocytes: an immunocytochemical study. Cell
Tissue Res 1995; 282:219–226.
14. Roseboom PH, Klein DC. Norepinephrine stimulation of
pineal cyclic AMP response element-binding protein phos-
phorylation: primary role of a beta-adrenergic receptor/cyclic
AMP mechanism. Mol Pharmacol 1995; 47:439–449.
15. Obsil T, Ghirlando R, Klein DC et al. Crystal structure
of the 14-3-3zeta: serotonin N-acetyltransferase complex. A
role for scaffolding in enzyme regulation. Cell 2001; 105:257–
267.
16. Ganguly S, Gastel JA, Weller JL et al. Role of a pineal
cAMP-operated arylalkylamine N-acetyltransferase/14-3-3-
binding switch in melatonin synthesis. Proc Natl Acad Sci
USA 2001; 98:8083–8088.
17. Gastel JA, Roseboom PH, Rinaldi PA et al. Melatonin
production: proteasomal proteolysis in serotonin N-acetyl-
transferase regulation. Science 1998; 279:1358–1360.
18. Stehle JH, Foulkes NS, Molina CA et al. Adrenergic sig-
nals direct rhythmic expression of transcriptional repressor
CREM in the pineal gland. Nature 1993; 365:314–320.
19. Foulkes NS, Borjigin J, Snyder SH, Sassone-Corsi P.
Transcriptional control of circadian hormone synthesis via the
CREM feedback loop. Proc Natl Acad Sci USA 1996;
93:14140–14145.
20. Pfeffer M, Maronde E, Molina CA et al. Inducible cyclic
AMP early repressor protein in rat pinealocytes: a highly
sensitive natural reporter for regulated gene transcription. Mol
Pharmacol 1999; 56:279–289.
21. Koch M, Mauhin V, Stehle JH et al. Dephosphorylation of
pCREB by protein serine/threonine phosphatases is involved
in inactivation of Aanat gene transcription in rat pineal gland.
J Neurochem 2003; 85:170–179.
22. Koch M, Dehghani F, Habazettl I et al. Cannabinoids
attenuate norepinephrine-induced melatonin biosynthesis in
the rat pineal gland by reducing arylalkylamine N-acetyl-
transferase activity without involvement of cannabinoid
receptors. J Neurochem 2006; 98:267–278.
23. Pagotto U, Marsicano G, Cota D et al. The emerging role
of the endocannabinoid system in endocrine regulation and
energy balance. Endocr Rev 2006; 27:73–100.
24. Di Marzo V, Bisogno T, De Petrocellis L. Endocanna-
binoids and related compounds: walking back and forth
between plant natural products and animal physiology. Chem
Biol 2007; 14:741–756.
25. Devane WA, Hanus L, Breuer A et al. Isolation and struc-
ture of a brain constituent that binds to the cannabinoid
receptor. Science 1992; 258:1946–1949.
26. Piomelli D. The molecular logic of endocannabinoid signal-
ling. Nat Rev Neurosci 2003; 4:873–884.
27. Van Der Stelt M, Di Marzo V. Anandamide as an
intracellular messenger regulating ion channel activity.
Prostaglandins Other Lipid Mediat 2005; 77:111–122.
Endocannabinoid system in rat pineal gland
359

28. Okamoto Y, Morishita J, Tsuboi K et al. Molecular char-
acterization of a phospholipase D generating anandamide and
its congeners. J Biol Chem 2004; 279:5298–5305.
29. Hillard CJ, Edgemond WS, Jarrahian A, Campbell WB.
Accumulation of N-arachidonoylethanolamine (anandamide)
into cerebellar granule cells occurs via facilitated diffusion.
J Neurochem 1997; 69:631–638.
30. Beltramo M, Stella N, Calignano A et al. Functional role
of high-affinity anandamide transport, as revealed by selective
inhibition. Science 1997; 277:1094–1097.
31. Cravatt BF, Giang DK, Mayfield SP et al. Molecular
characterization of an enzyme that degrades neuromodulatory
fatty-acid amides. Nature 1996; 384:83–87.
32. Bradford MM. A rapid and sensitive method for the quan-
titation of microgram quantities of protein utilizing the prin-
ciple of protein-dye binding. Anal Biochem 1976; 72:248–254.
33. Patel S, Roelke CT, Rademacher DJ et al. Endocannabi-
noid signaling negatively modulates stress-induced activation
of the hypothalamic-pituitary-adrenal axis. Endocrinology
2004; 145:5431–5438.
34. Schmid PC, Paria BC, Krebsbach RJ et al. Changes in
anandamide levels in mouse uterus are associated with uterine
receptivity for embryo implantation. Proc Natl Acad Sci USA
1997; 94:4188–4192.
35. Herkenham M, Lynn AB, Johnson MR et al. Character-
ization and localization of cannabinoid receptors in rat brain: a
quantitative in vitro autoradiographic study. J Neurosci 1991;
11:563–583.
36. Tsou K, Brown S, Sanudo-Pena MC, Mackie K, Walker
JM. Immunohistochemical distribution of cannabinoid CB1
receptors in the rat central nervous system. Neuroscience 1998;
83:393–411.
37. Katona I, Sperlagh B, Sik A et al. Presynaptically located
CB1 cannabinoid receptors regulate GABA release from axon
terminals of specific hippocampal interneurons. J Neurosci
1999; 19:4544–4558.
38. Egertova M, Elphick MR. Localisation of cannabinoid
receptors in the rat brain using antibodies to the intracellular
C-terminal tail of CB. J Comp Neurol 2000; 422:159–171.
39. Trendelenburg AU, Cox SL, Schelb V et al. Modulation of
(3)H-noradrenaline release by presynaptic opioid, cannabinoid
and bradykinin receptors and beta-adrenoceptors in mouse
tissues. Br J Pharmacol 2000; 130:321–330.
40. Ishac EJ, Jiang L, Lake KD et al. Inhibition of exocytotic
noradrenaline release by presynaptic cannabinoid CB1 recep-
tors on peripheral sympathetic nerves. Br J Pharmacol 1996;
118:2023–2028.
41. Tam J, Trembovler V, Di Marzo V et al. The cannabinoid
CB1 receptor regulates bone formation by modulating adren-
ergic signaling. FASEB J 2008; 22:285–294.
42. Notarnicola M, Messa C, Orlando A et al. Estrogenic
induction of cannabinoid CB1 receptor in human colon cancer
cell lines. Scand J Gastroenterol 2007; 18:1–7.
43. Hayakawa K, Mishima K, Nozako M et al. High-cholesterol
feeding aggravates cerebral infarction via decreasing the CB1
receptor. Neurosci Lett 2007; 414:183–187.
44. Oliva JM, Ortiz S, Perez-Rial S et al. Time dependent
alterations on tyrosine hydroxylase, opioid and cannabinoid
CB(1) receptor gene expressions after acute ethanol adminis-
tration in the rat brain. Eur Neuropsychopharmacol 2008;
18:373–382.
45. Hsieh C, Brown S, Derleth C, Mackie K. Internalization
and recycling of the CB1 cannabinoid receptor. J Neurochem
1999; 73:493–501.
46. Jin W, Brown S, Roche JP et al. Distinct domains of the CB1
cannabinoid receptor mediate desensitization and internaliza-
tion. J Neurosci 1999; 19:3773–3780.
47. Stella N. Cannabinoid signaling in glial cells. Glia 2004;
48:267–277.
48. Van Sickle MD, Duncan M, Kingsley PJ et al. Identifica-
tion and functional characterization of brainstem cannabinoid
CB2 receptors. Science 2005; 310:329–332.
49. Ashton JC, Friberg D, Darlington CL, Smith PF.
Expression of the cannabinoid CB2 receptor in the rat cere-
bellum: an immunohistochemical study. Neurosci Lett 2006;
396:113–116.
50. Gong JP, Onaivi ES, Ishiguro H et al. Cannabinoid CB2
receptors: immunohistochemical localization in rat brain.
Brain Res 2006; 1071:10–23.
51. Valenti M, Vigano D, Casico MG et al. Differential diurnal
variations of anandamide and 2-arachidonoyl-glycerol levels in
rat brain. Cell Mol Life Sci 2004; 61:945–950.
52. Murillo-Rodriguez E, Desarnaud F, Prospero-Garcia O.
Diurnal variation of arachidonoylethanolamine, palmitoy-
lethanolamide and oleoylethanolamide in the brain of the rat.
Life Sci 2006; 79:30–37.
53. Bacci A, Huguenard JR, Prince DA. Long-lasting self-
inhibition of neocortical interneurons mediated by endocann-
abinoids. Nature 2004; 431:312–316.
54. Kreitzer AC, Regehr WG. Cerebellar depolarization-
induced suppression of inhibition is mediated by endogenous
cannabinoids. J Neurosci 2001; 21:RC174.
Koch et al.
360