the megalopa and juvenile development of pachygrapsus transversus (gibbes, 1850) (decapoda,...
TRANSCRIPT

The Megalopa and Juvenile Development of Pachygrapsus transversus (Gibbes, 1850) (Decapoda,Brachyura) Compared with Other Grapsid CrabsAuthor(s): Augusto A. V. Flores, Maria Lucia Negreiros-Fransozo and Adilson FransozoSource: Crustaceana, Vol. 71, No. 2 (Mar., 1998), pp. 197-222Published by: BRILLStable URL: http://www.jstor.org/stable/20105974 .
Accessed: 05/12/2013 07:58
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
BRILL is collaborating with JSTOR to digitize, preserve and extend access to Crustaceana.
http://www.jstor.org
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions

THE MEGALOPA AND JUVENILE DEVELOPMENT OF PACHYGRAPSUS TRANSVERSOS (GIBBES, 1850) (DECAPODA, BRACHYURA) COMPARED
WITH OTHER GRAPSID CRABS
BY
AUGUSTO A. V. FLORES, MARIA LUCIA NEGREIROS-FRANSOZO and ADILSON FRANSOZO
NEBECC (Group of Studies on Crustacean Biology, Ecology and Culture) Departamento de
Zoologia, Instituto de Bioci?ncias and
Centro de Aq?icultura, Universidade Estadual Paulista UNESP, C.P. 510,
18618-000 Botucatu (SP), Brazil
ABSTRACT
Megalopae were reared in the laboratory to the 7th crab stage. The megalopa and 1st crab stage are described and juvenile development was studied with emphasis on pleopodal differentiation.
The megalopal phase, is easily identified, and shares with those of other Grapsinae and Plagusiinae
big size, the presence of many natatory setae, and a series of conspicuous teeth on the inner
margin of the dactyli from the 2nd to 4th walking leg. These features are regarded as adaptive for
settlement in a wave-swept environment, such as the rocky marine intertidal where most of those
species live. Fast development of juvenile pleopods is another characteristic of these subfamilies.
In Pachygrapsus transversus, the sexes can be distinguished from the 2nd crab stage. Gonopod differentiation in males and the basic segmentation of all four pleopod pairs in females are already
concluded at the 5th instar. A review of the available information indicated that settlement of large
megalopae and fast juvenile development, preceding a precocious sexual maturity, are trends in
Grapsinae and Plagusiinae. On the other hand, the Sesarminae pass through a more extensive
juvenile instar sequence and presumably a delayed maturity.
RESUMEN
Individuos en el estadio de megalopa fueron capturados y mantenidos en laboratorio hasta el
7o estadio. La megalopa y el 1er juvenil fueron descritos y el desarrollo juvenil estudiado con
?nfasis en la diferenciaci?n de los ple?podos de estos cangrejos. La fase de megalopa es f?cilmente
reconocida y comparte con otros Grapsinae y los Plagusiinae la gran talla, la presencia de muchas
sedas natatorias y una s?rie de dientes conspicuos en el margen interno de los d?ctilos del 2o al 4o
pere?podo. Estas caracter?sticas se consideran como adaptativas frente a condiciones de exposici?n al oleaje, comunes en el litoral rocoso donde vive la maioria de esas especies. Otra caracter?stica
de esas subfamilias es el r?pido desarrollo de los ple?podos. En Pachygrapsus transversus, los
sexos pueden ser diferenciados desde el 2o estadio juvenil. El desarrollo del gon?podo en machos
y la segmentaci?n b?sica de los cuatro pares de ple?podos en las hembras ya est?n concluidos en
el 5o estadio. Una revisi?n de la escasa informaci?n disponible sugiere que el asentamiento de
? Koninklijke Brill NV, Leiden, 1998 Crustaceana 71 (2)
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions

198 A. A. V. FLORES ET AL.
megalopas de gran talla y el r?pido desarrollo juvenil, antecediendo una maturidad sexual precoz, son una tendencia en Grapsinae y Plagusiinae. Por otro lado, los Sesarminae deben pasar por una
larga secuencia de estadios juveniles y, por lo tanto, alcanzar la maturidad sexual m?s tarde.
INTRODUCTION
Pachygrapsus transversus (Gibbes, 1850) is the common grapsid crab on rocky shores of southern Brazil, but its development is inadequately known. Descrip tions of zoeae are limited to stage I (Lebour, 1944; Ingle, 1987) and stage II
(Cuesta & Rodr?guez, 1994), and only a general illustration is available (Rossig
nol, 1957).
Descriptions of grapsid larvae cover less than 20% of all those known of
Grapsid species (Felder et al., 1985), and these are mainly of Sesarminae and
Varuninae, while the Grapsinae and Plagusiinae remain poorly studied (Wilson,
1980). This biased database is probably related to the fact that plagusiines are
a very restricted group and grapsine larvae are difficult to rear in the labora
tory. Many of the grapsine larval descriptions are limited to the 1st zoea (e.g.,
Paula, 1985; Konishi & Minagawa, 1990), and other rearing efforts failed to
obtain the megalopal stage (Schlotterbeck, 1976; Brossi-Garcia & Rodrigues,
1993). From the scattered complete descriptions of grapsine megalopae, only those of the genus Metopograpsus were obtained by rearing techniques (Kakati,
1982; Pasupathi & Kannupandi, 1986; Fielder & Greenwood, 1983a). Available
megalopal descriptions of the genera Pachygrapsus (Guerao et al., in press) and
Planes (Muraoka, 1973) are based on wild specimens.
Descriptions of early postlarval development in brachyuran crabs are still
scarce, probably due to difficulties in identification (Fransozo & Negreiros
Fransozo, 1987), and in obtaining of juvenile specimens from identified parents
(Hebling et al., 1982). Nevertheless, the existing literature (e.g., Shen, 1935;
Ingle, 1977; Fukuda, 1981; Rieger, 1986) has shown that the acquisition of adult
form, including pleopod sexual differentiation, is highly variable in brachyuran
crabs, including the Grapsidae (Muraoka, 1963; Morita, 1974; Fransozo, 1987;
Hartnoll, 1992; Lee et al., 1994).
In the present paper, the megalopa and the 1st crab instar of P. transversus
are described, and the juvenile development is investigated with emphasis on
the process of pleopod differentiation. The adaptive value of some megalopal features in grapsid crabs are discussed in an ecomorphological approach, and a
preliminary comparative analysis of juvenile development in grapsids is provided.
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions

MEGALOPA AND JUVENILE PACHYGRAPSUS 199
MATERIAL AND METHODS
In April 1995, 17 megalopae were obtained from reefs made by the sabelariid
worm Phragmatopoma lapidosa Kinberg, 1867. Each larva was immediately
placed in an individual 120 ml acrylic vessel, containing pebbles as a hard sub
stratum and partially filled with filtered seawater (34%o S). Every day the water
was changed and the individuals were fed on a combination of live Artemia
nauplii and small pieces of Ulva, shrimp and crab muscle. Temperature was
maintained at 25 ? 1?C and ambient photoperiod. Laboratory rearing was con
tinued until most of the crabs attained the 7th crab stage.
Carapace length, width and instar duration were recorded individually in each
stage. Corpses were fixed in 70% ethyl alcohol. To preserve the exuviae, glycerin was added in a 1 : 1 proportion.
The megalopae and 1st crab instar are fully described. In the following stages,
pleopodal differentiation, carapace morphology and the appearence of genital
openings, are detailed. Drawings were made with the aid of an Olympus binoc
ular microscope and a Zeiss stereomicroscope both provided with a camera lu
cida. Terminology of main setal features are based on Pohle & Telford's (1981)
classification.
RESULTS
Megalopa (figs. 1-5)
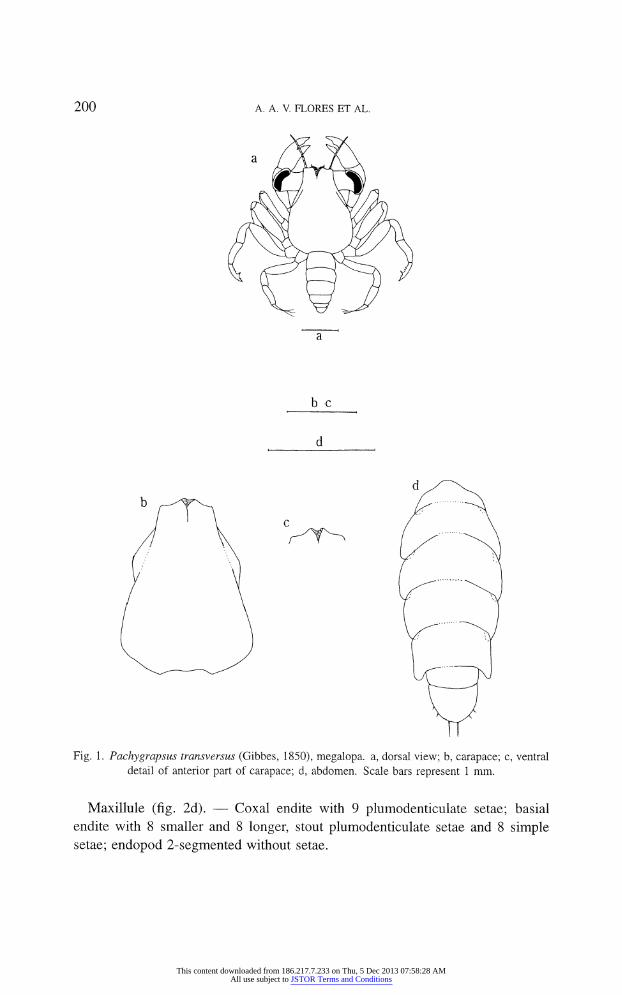
Carapace (fig. la, b, c). ?
Length 2.8 mm, width 1.9 mm. Of subtriangular
shape, maximum width at the branchial region nearly twice as long as anterior
width; smooth surface, without protuberances, knobs or grooves; anterolateral
region with lower expansion reaching orbital region (fig. lb); rostrum ventrally curved with a conspicuous median front groove (fig. lc); eyes large, ocular
peduncles well developed (fig. la).
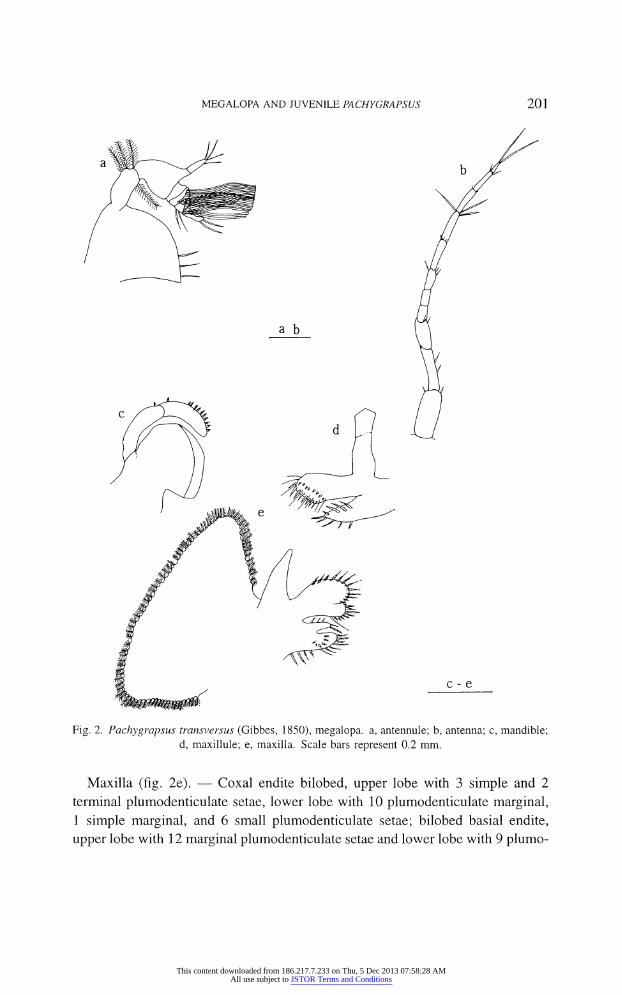
Antennule (fig. 2a). ? Peduncle 3-segmented with 3 plumodenticulate, 4 long
plumose and 0 setae, respectively; endopod unsegmented with 4 terminal simple
setae; exopod 4-segmented with 0, 0, 2, 2 terminal simple setae, and 0, 12, 10, 3
long aesthetascs.
Antenna (fig. 2b). ? Peduncle 3-segmented, with 2, 2, 2 simple setae; flag
ellum 8-segmented with 0, 0, 3, 3, 3 (long), 2, 2 and 2 terminal long, simple setae.
Mandible (fig. 2c). ? Flat with distinct inner margin; 2-segmented palp with
1 and 1 + 10 plumodenticulate setae.
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions
200 A. A. V. FLORES ET AL.
b C
Fig. 1. Pachygrapsus transversus (Gibbes, 1850), megalopa. a, dorsal view; b, carapace; c, ventral
detail of anterior part of carapace; d, abdomen. Scale bars represent 1 mm.
Maxillule (fig. 2d). ? Coxal endite with 9 plumodenticulate setae; basial
endite with 8 smaller and 8 longer, stout plumodenticulate setae and 8 simple
setae; endopod 2-segmented without setae.
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions
MEGALOPA AND JUVENILE PACHYGRAPSUS 201
Fig. 2. Pachygrapsus transversus (Gibbes, 1850), megalopa. a, antennule; b, antenna; c, mandible;
d, maxillule; e, maxilla. Scale bars represent 0.2 mm.
Maxilla (fig. 2e). ? Coxal endite bilobed, upper lobe with 3 simple and 2
terminal plumodenticulate setae, lower lobe with 10 plumodenticulate marginal, 1 simple marginal, and 6 small plumodenticulate setae; bilobed basial endite,
upper lobe with 12 marginal plumodenticulate setae and lower lobe with 9 plumo
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions
202 A. A. V. FLORES ET AL.
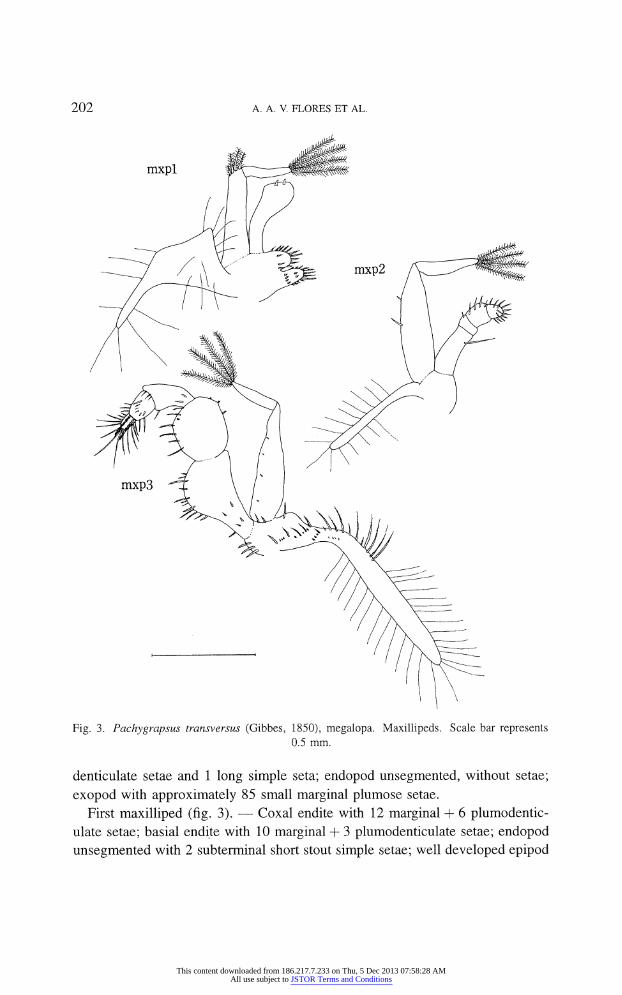
Fig. 3. Pachygrapsus transversus (Gibbes, 1850), megalopa. Maxillipeds. Scale bar represents
0.5 mm.
denticulate setae and 1 long simple seta; endopod unsegmented, without setae;
exopod with approximately 85 small marginal plumose setae.
First maxilliped (fig. 3). ? Coxal endite with 12 marginal + 6 plumodentic
ulate setae; basial endite with 10 marginal + 3 plumodenticulate setae; endopod
unsegmented with 2 subterminal short stout simple setae; well developed epipod
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions
MEGALOPA AND JUVENILE PACHYGRAPSUS 203
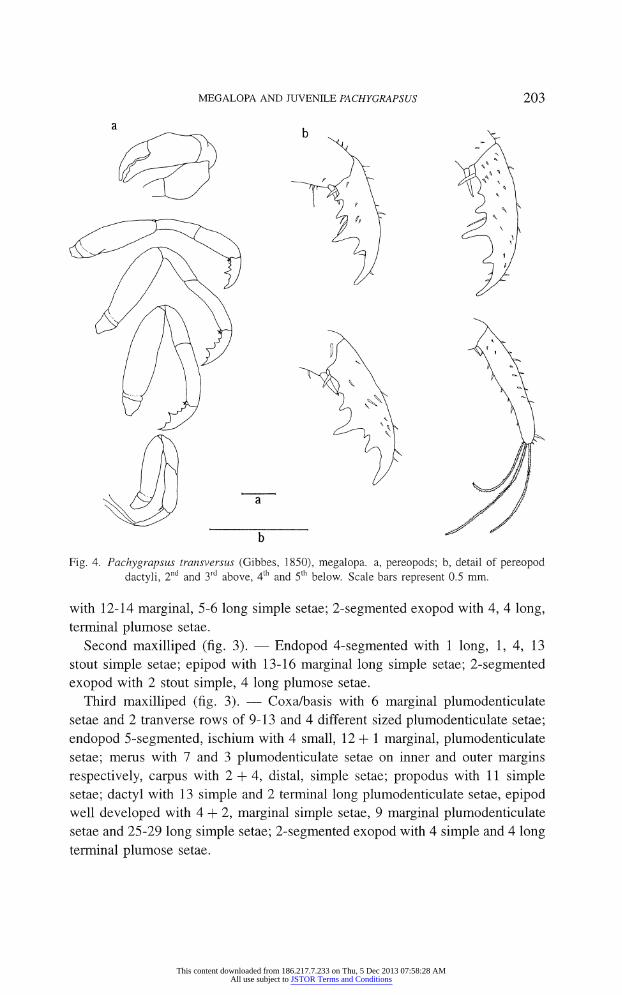
Fig. 4. Pachygrapsus transversas (Gibbes, 1850), megalopa. a, pereopods; b, detail of pereopod
dactyli, 2nd and 3rd above, 4th and 5th below. Scale bars represent 0.5 mm.
with 12-14 marginal, 5-6 long simple setae; 2-segmented exopod with 4, 4 long, terminal plumose setae.
Second maxilliped (fig. 3). ?
Endopod 4-segmented with 1 long, 1, 4, 13
stout simple setae; epipod with 13-16 marginal long simple setae; 2-segmented
exopod with 2 stout simple, 4 long plumose setae.
Third maxilliped (fig. 3). ? Coxa/basis with 6 marginal plumodenticulate
setae and 2 tranverse rows of 9-13 and 4 different sized plumodenticulate setae;
endopod 5-segmented, ischium with 4 small, 12+1 marginal, plumodenticulate
setae; merus with 7 and 3 plumodenticulate setae on inner and outer margins
respectively, carpus with 2 + 4, distal, simple setae; propodus with 11 simple
setae; dactyl with 13 simple and 2 terminal long plumodenticulate setae, epipod well developed with 4 + 2, marginal simple setae, 9 marginal plumodenticulate setae and 25-29 long simple setae; 2-segmented exopod with 4 simple and 4 long terminal plumose setae.
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions
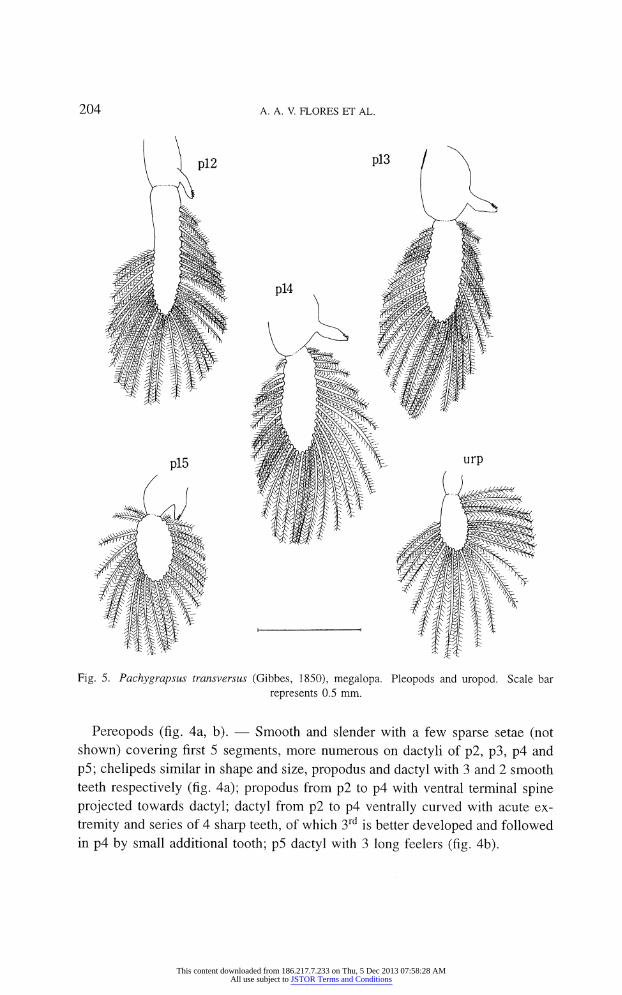
204 A. A. V. FLORES ET AL.
Fig. 5. Pachygrapsus transversus (Gibbes, 1850), megalopa. Pleopods and uropod. Scale bar
represents 0.5 mm.
Pereopods (fig. 4a, b). ? Smooth and slender with a few sparse setae (not
shown) covering first 5 segments, more numerous on dactyli of p2, p3, p4 and
p5; chelipeds similar in shape and size, propodus and dactyl with 3 and 2 smooth
teeth respectively (fig. 4a); propodus from p2 to p4 with ventral terminal spine
projected towards dactyl; dactyl from p2 to p4 ventrally curved with acute ex
tremity and series of 4 sharp teeth, of which 3rd is better developed and followed
in p4 by small additional tooth; p5 dactyl with 3 long feelers (fig. 4b).
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions

MEGALOPA AND JUVENILE PACHYGRAPSUS 205
Abdomen (fig. Id). ?
Elongated, 6 smooth segments wider than long, first
five ventrally articulated by disto-lateral expansions; 6th somite reduced; telson
with 2 pairs of short subterminal setae and 1 pair of long terminal setae.
Pleopods (fig. 5). ? Well developed; endopods from pl2 to pl5 with 4, 4, 4, 3
little hooked setae and 30-32, 32-35, 28-34, 24-26 large plumose natatory setae;
exopod area decreasing from pl2 to pl5.
Uropods (fig. 5). ?
Endopod absent; exopod with 21 large plumose natatory setae.
First crab (figs. 6-9)
Carapace (fig. 6a). ?
Length 2.3 mm, width 2.8 mm; similar to but narrower
than in the adult; second anterolateral tooth still absent; frontal, orbital and lateral
depressions present; gastric, cardiac and branchial regions already differentiated;
eyes relatively large when compared to adult.
Antennule (fig. 7a). ? Peduncle 3-segmented, proximal segment with 18 and
approximately 25 small plumose setae disposed in transverse and marginal rows
respectively; second and third segment with two and three simple setae; 3-seg mented endopod with 3, 2 and 3 terminal, simple setae; 6-segmented exopod with 0, 1, 1, 4, 4, 3 terminal, simple setae, 8, 7, and 3 long aesthetascs present on 2nd, 3rd and 4th segments, respectively.
Antenna (fig. 7b). ? Peduncle 3-segmented, with 7 small, 3 large and 0 plumo
se setae and 0, 3, 3 simple setae; flagellum 8-segmented with 3, 2, 4, 3, 4, 1, 1
and 3 simple setae.
Mandible (fig. 7c). ? With 1 median tooth on differentiated inner margin;
3-segmented palp with 1, 7 small plumose setae and 3 small plumose + 14-16
small plumodenticulate setae.
Maxillule (fig. 7d). ? Coxal endite with 8 + 14 simple, 7 marginal plumose
and 5 terminal stout plumodenticulate setae; basial endite with 1 long plumose, 5 marginal long plumodenticulate and 23 subterminal + 6 stout simple setae;
endopod 2-segmented with 5 long and 4 small plumose + 2 simple setae.
Maxilla (fig. 7e). ? Bilobed coxal endite, upper lobe with 9 marginal plumose
setae, lower lobe with 7, 5 + 10-13 marginal, plumose setae; bilobed basial endite,
upper lobe with 4 small and 14-16 marginal different sized, plumose setae, lower
lobe with 4 small and 5+2 longer marginal plumose setae; endopod unsegmented and smooth; exopod with approximately 120 small marginal plumose setae.
Sternum (fig. 6b). ? First 3 segments fused, other 5 clearly defined; pair of
buttons on the 5th sternite; 8th sternite with slight longitudinal depression; genital
openings still absent.
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions
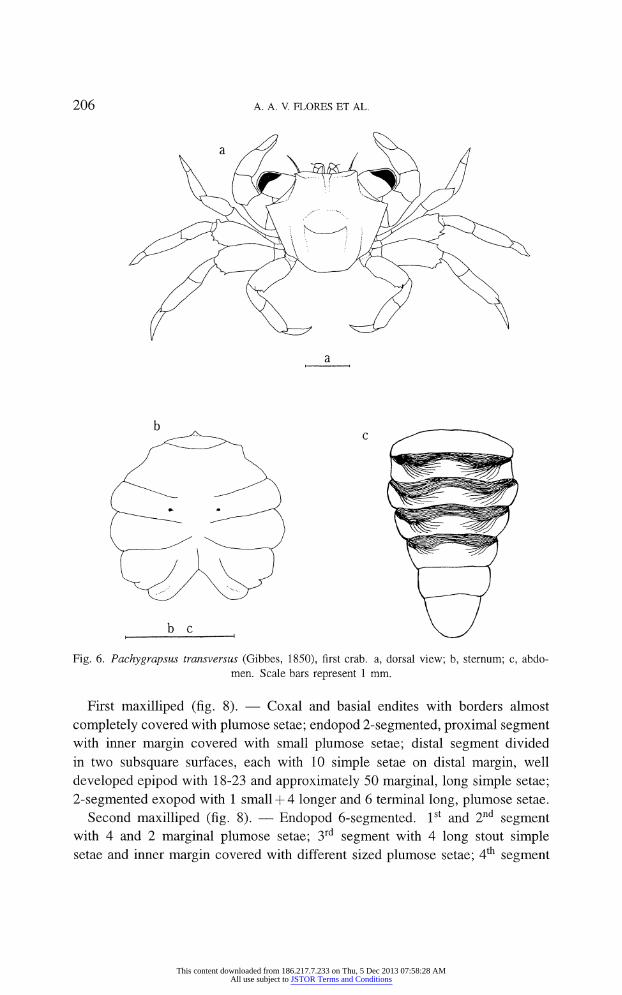
206 A. A. V. FLORES ET AL.
Fig. 6. Pachygrapsus transversus (Gibbes, 1850), first crab, a, dorsal view; b, sternum; c, abdo
men. Scale bars represent 1 mm.
First maxilliped (fig. 8). ? Coxal and basial endites with borders almost
completely covered with plumose setae; endopod 2-segmented, proximal segment with inner margin covered with small plumose setae; distal segment divided
in two subsquare surfaces, each with 10 simple setae on distal margin, well
developed epipod with 18-23 and approximately 50 marginal, long simple setae;
2-segmented exopod with 1 small + 4 longer and 6 terminal long, plumose setae.
Second maxilliped (fig. 8). ?
Endopod 6-segmented. 1st and 2nd segment with 4 and 2 marginal plumose setae; 3rd segment with 4 long stout simple setae and inner margin covered with different sized plumose setae; 4th segment
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions

MEGALOPA AND JUVENILE PACHYGRAPSUS 207
Fig. 7. Pachygrapsus transversus (Gibbes, 1850), first crab, a, antennule; b, antenna; c, mandible;
d, maxillule; e, maxilla. Scale bars represent 0.2 mm.
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions
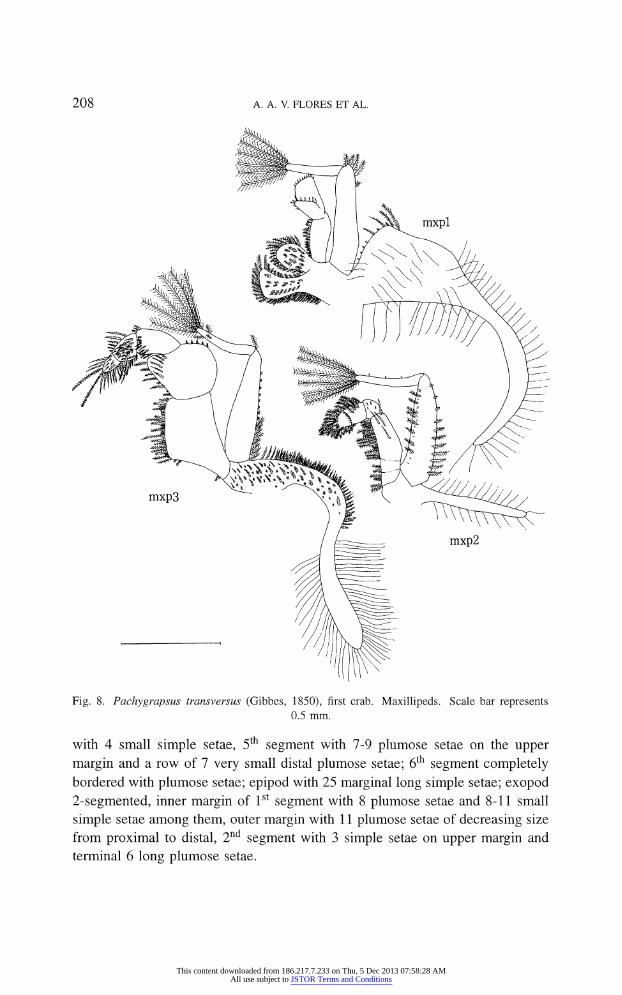
208 A. A. V. FLORES ET AL.
Fig. 8. Pachygrapsus transversus (Gibbes, 1850), first crab. Maxillipeds. Scale bar represents
0.5 mm.
with 4 small simple setae, 5th segment with 7-9 plumose setae on the upper
margin and a row of 7 very small distal plumose setae; 6th segment completely bordered with plumose setae; epipod with 25 marginal long simple setae; exopod
2-segmented, inner margin of 1st segment with 8 plumose setae and 8-11 small
simple setae among them, outer margin with 11 plumose setae of decreasing size
from proximal to distal, 2nd segment with 3 simple setae on upper margin and
terminal 6 long plumose setae.
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions
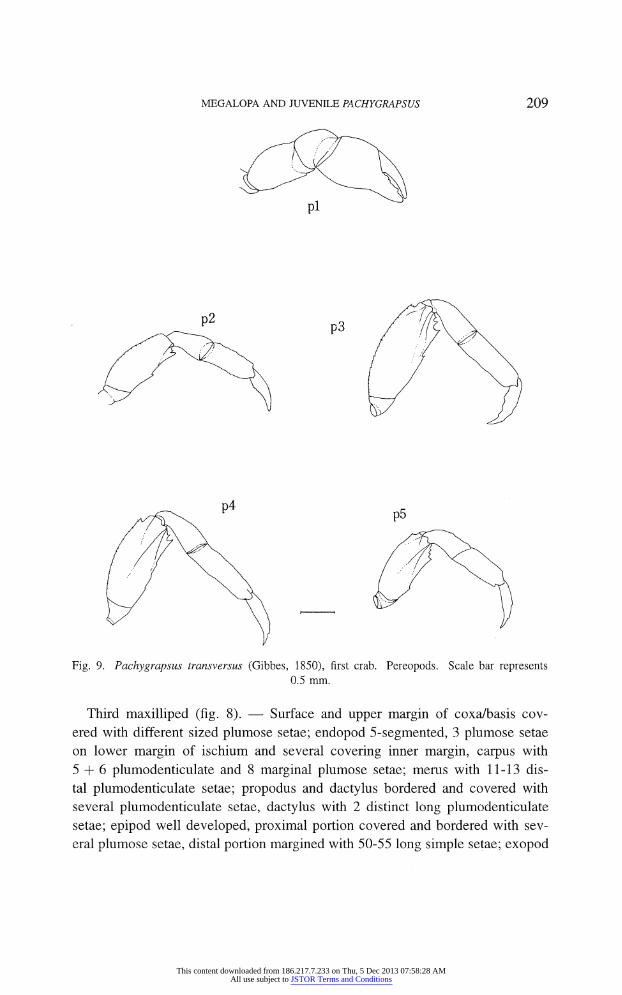
MEGALOPA AND JUVENILE PACHYGRAPSUS 209
Fig. 9. Pachygrapsus transversus (Gibbes, 1850), first crab. Pereopods. Scale bar represents
0.5 mm.
Third maxilliped (fig. 8). ? Surface and upper margin of coxa/basis cov
ered with different sized plumose setae; endopod 5-segmented, 3 plumose setae
on lower margin of ischium and several covering inner margin, carpus with
5 + 6 plumodenticulate and 8 marginal plumose setae; merus with 11-13 dis
tal plumodenticulate setae; propodus and dactylus bordered and covered with
several plumodenticulate setae, dactylus with 2 distinct long plumodenticulate
setae; epipod well developed, proximal portion covered and bordered with sev
eral plumose setae, distal portion margined with 50-55 long simple setae; exopod
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions

210 A. A. V. FLORES ET AL.
2-segmented, proximal segment with 8, 8 small and 1 distal, marginal plumose
setae, distal segment with 3 subterminal and 6 long terminal, plumose setae.
Pereopods (fig. 9). ?
Chelipeds similar to those of adult, but without defined
dentition; walking legs with distribution pattern of setae (not shown) resembling the adult form, grooves in meri already present.
Abdomen (fig. 6c). ? Lateral margins of somites still rounded; 2nd to 5th
somite with characteristic transverse striae which disappear in later instars.
Pleopods. ? Absent. First stage crabs with vestigial, shrivelled pleopods were
captured occasionally in the wild.
Second crab (figs. 10a, e, 11)
Males and females can be already distinguished at this instar. First pair of
pleopods in males from small biramous appendages to longer distally grooved
gonopods (fig. 10a). Male genital openings in lateral ends of sternite 8 (fig. 11).
In females, 2nd to 5th pleopod pairs present. All pairs biramous, with endopod
equal to or longer than exopod (fig. 10e). Female genital openings present on
6th sternite (fig. 11).
Carapace wider (table I), second anterolateral tooth (missing in the previous
stage) present. At this stage the crabs are almost miniature adults.
Further juvenile stages (fig. lOb-d, f-h)
Differentiation of pleopods continues until the 5th crab stage in both males
and females.
Males. ? 3rd instar: pli with distal groove and longitudinal sulcus; pl2 already
present (fig. 10b). 4th instar: pli greatly enlarged and closely resembling that of
adult, terminal region laterally curved, distal groove larger, few setae on upper
Table I Juvenile development of Pachygrapsus transversus (Gibbes, 1850). Carapace
length, carapace length/width ratio and duration of each juvenile stage in labora
tory conditions. Variation expressed as standard deviation, numbers in brackets
indicate number of observed crabs
Juvenile instars
_ JI JH JIII JIV JV JVI JVII
Length (mm) 2.3+0.1 2.6+0.2 3.0+0.2 3.5+0.3 4.1+0.5 4.6+0.5 5.0+0.4
(9) (14) (15) (16) (15) (13) (7) LAV 0.83 0.75 0.72 0.71 0.73 0.72 0.74 Duration (days) 17.9+8.0 17.8+6.2 18.6+4.7 22.5+5.1 20.5+7.5 25.8+4.9 28
(17) (17) (17) (17) (11) (8) (1)
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions
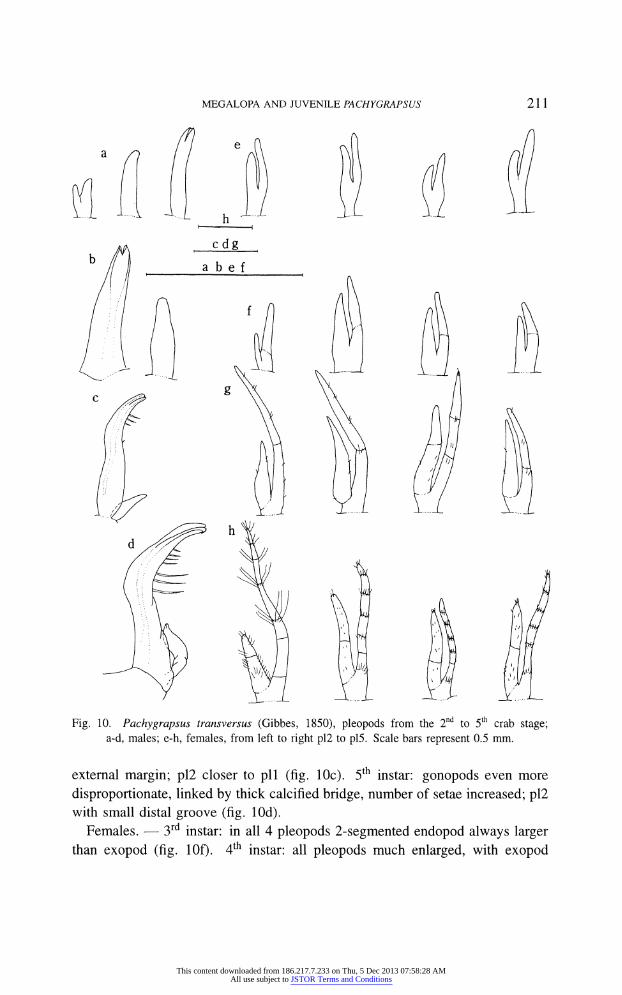
MEGALOPA AND JUVENILE PACHYGRAPSUS 211
Ml} \\ i i I-1
Fig. 10. Pachygrapsus transversus (Gibbes, 1850), pleopods from the 2 to 5 crab stage; a-d, males; e-h, females, from left to right pl2 to pl5. Scale bars represent 0.5 mm.
external margin; pl2 closer to pll (fig. 10c). 5th instar: gonopods even more
disproportionate, linked by thick calcified bridge, number of setae increased; pl2 with small distal groove (fig. lOd).
Females. ? 3rd instar: in all 4 pleopods 2-segmented endopod always larger
than exopod (fig. lOf). 4th instar: all pleopods much enlarged, with exopod
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions
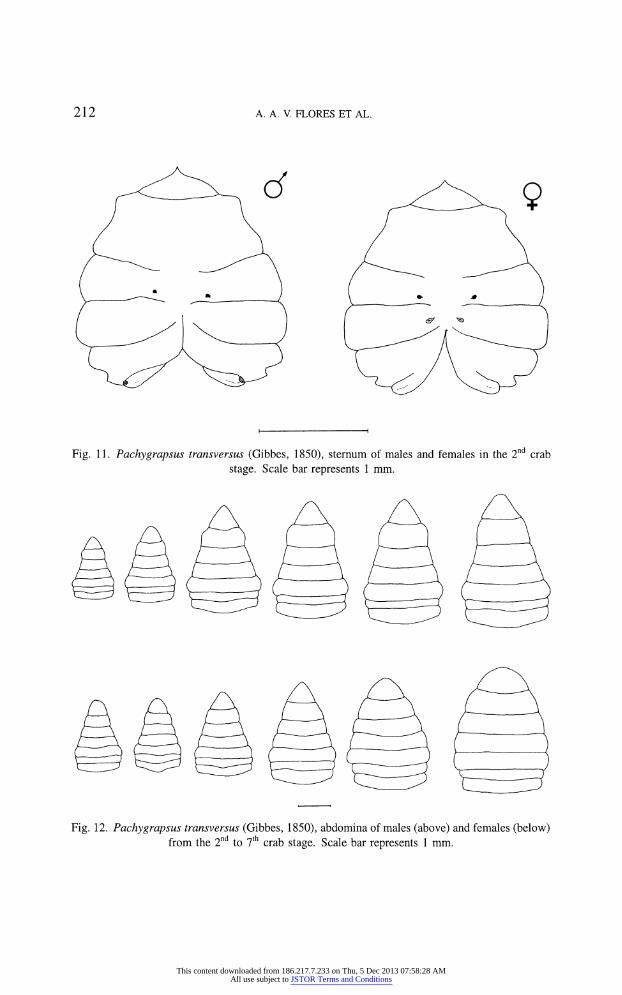
212 A. A. V FLORES ET AL.
Fig. 11. Pachygrapsus transversus (Gibbes, 1850), sternum of males and females in the 2nd crab
stage. Scale bar represents 1 mm.
Fig. 12. Pachygrapsus transversus (Gibbes, 1850), abdomina of males (above) and females (below)
from the 2nd to 7th crab stage. Scale bar represents 1 mm.
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions

MEGALOPA AND JUVENILE PACHYGRAPSUS 213
differentiated from protopod and some sparse setae present, pl4 with 3-segmented
endopod and 2-segmented exopod (fig. 10g). 5th instar: pleopods more setose
and segmented; all endopods 2-segmented; exopods 6, 6, 6 and 5-segmented;
segments with distally concentrated setae; pll longer than rest, with longer setae
(fig. 10h). Females in 4th and 5th instars showed some variability in pleopod segmentation
and setation. One 4th instar female presented a typical 5th instar segmentation, and a 5th instar female with expected setation had segmentation, as in a typical 4th instar.
Abdominal sexual dimorphism can only be clearly observed after the 6th stage.
At the 7th instar, the abdomen of females resembles the pre-puberal shape found
in wild living specimens, while males present a typical adult form (fig. 12).
At the 3rd instar, the carapace shape becomes virtually the same as in the adult,
with a similar length/width ratio. Time elapsed between moults increased with
size, as expected (table I). However, intermoult period and instar size recorded
may vary from the wild.
DISCUSSION
Rossignol (1957) published a dorsal view of a P. transversus megalopa from
Central Western Africa. Measurements and general morphological characteristics
are compatible with the present descriptions, but the carapace ornamentation
shown was not observed in the current study. This difference may correspond to local coloration patterns, as in adult crabs, or it may be a result of genetic
divergence, evidenced by Cuesta & Schubart (in prep.), between populations on
both sides of the Atlantic.
The megalopa of P. transversus can be readily distinguished from other Pachy
grapsus species. A full description of the megalopa of P. crassipes Randall,
1840 is not available, but Hiatt (1948) provided valuable information regard
ing its general morphology. Among other records, the author found a mean
carapace length of 5.6 mm, a much higher size than the 2.8 mm recorded for
P. transversus. Considering that both measurements were obtained from wild
specimens, this difference is sufficient to distinguish these species. The mega
lopa of P. marmoratus (Fabricius, 1787), described by Guerao et al. (in press) can also be easily distinguished from the megalopa of P. transversus, since the
former presents a higher number of teeth (7) on the inner margin of the dactyli
from p2 to p5, and 5 setae in the maxillule endopod, which are absent in the
latter. The morphological similarity between the megalopa of P. transversus
and Planes cyaneus Dana, 1851, described by Muraoka (1973), is equivalent
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions

214 A. A. V. FLORES ET AL.
to that found within the genus Pachygrapsus. Otherwise, those 2 genera differ
remarkably from the megalopal descriptions by Kakati (1982) and Pasupathi &
Kannupandi (1986) of mangrove grapsines of the genus Metopograpsus. The megalopal stage in most brachyuran crabs is considered an obligatory
link between the zoeal planktonic phase and the adult benthonic crabs (Rice,
1981). Therefore, this last larval phase must be able to efficiently locate and
then settle in a favourable environment. In order to accomplish these tasks
some specializations regarding chemical perception, locomotion and substrate
adherence, are expected to have evolved in these larvae. Some morphological characteristics of grapsid megalopae from formal larval descriptions, were chosen
to assess the power of swimming and substrate grasping. For this purpose, it
was assumed that: first, larger megalopae are more likely capable to challenge
eventually weak water movements; second, number of natatory setae in pleopods are proportional to swimming stroke; and third, inner serratiform margin of
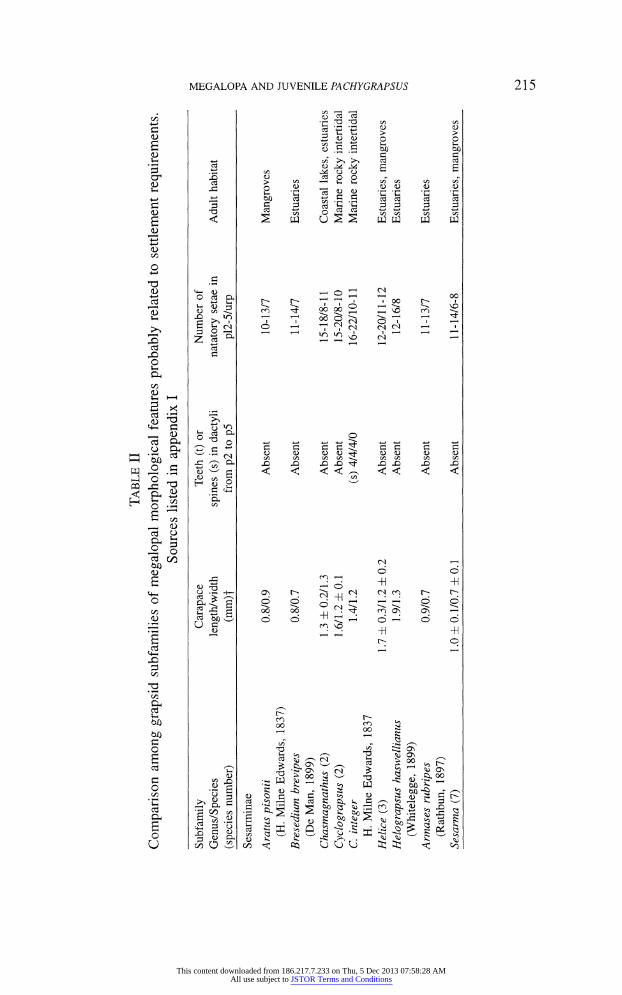
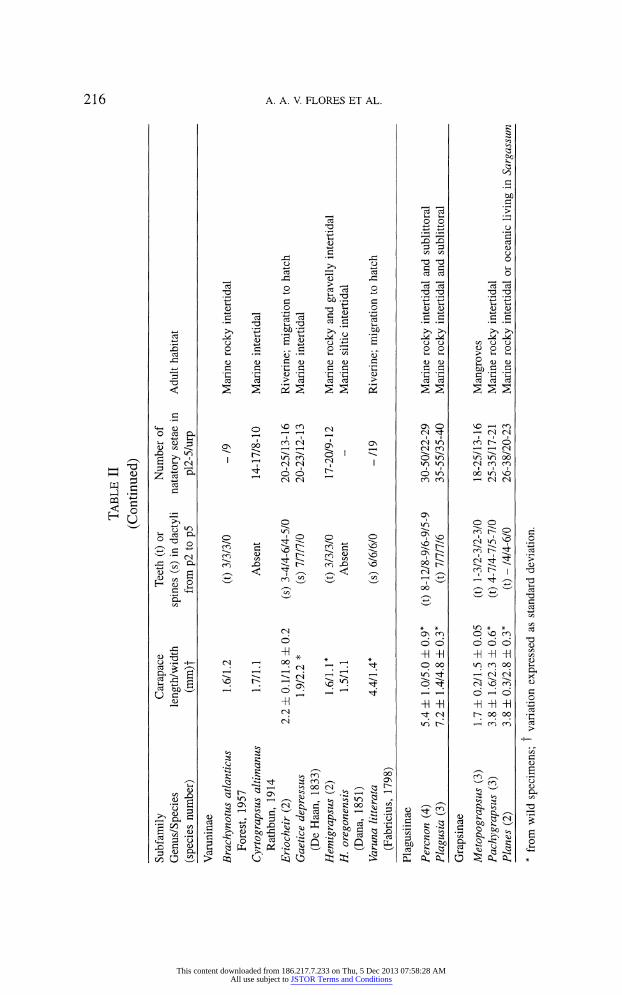
pereopod dactyli are useful to avoid wave dislodging. In table II, these features are presented in a subfamilial order, in which species
belonging to the same genus are grouped together when no relevant differences
were recorded among them. An additional column of species/genus habitat is
also provided.
Perhaps the most obvious trend from this analysis is the morphological di
chotomy between the Sesarminae and a group enclosing the Plagusiinae and
the Grapsinae. Most Sesarmine megalopae settle in relatively sheltered envi
ronments, such as mangroves and other estuaries. With a few exceptions, these
settlers are small and do not possess a high number of natatory setae nor spe
cialized grasping dactyli. On the other hand, plagusiine and grapsine megalopae, are larger and are provided with more natatory setae and armed dactyli. This
trend was quite expected, since this second grouping is almost totally composed
by inhabitants of wave exposed environments.
The grasping structures of dactyli comprise both spines; i.e., stout large out
grows, and teeth; i.e., acute expansions of the segment itself. These projections
may play an equivalent role, but they are certainly not homologous. Among the Brachyura, prominent spines are also present on the dactyli of some majids
(e.g., Kurata, 1969; Christiansen, 1973; Negreiros-Fransozo & Fransozo, 1991;
Hiyodo & Fransozo, 1994), but a series of noticeable teeth is only present in
some grapsids. All the Grapsinae and the Plagusiinae present well developed teeth on the inner margin of dactyli from p2 to p4. This can be an apomor
phic condition distinguishing this group. Small teeth, a second condition that
might have evolved independently, are present in some Varuninae, namely Hemi
grapsus sanguineus (De Haan, 1835); H. nudus (Dana, 1851) and Brachynotus
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions
MEGALOPA AND JUVENILE PACHYGRAPSUS 215
G
G
g
O -T3
O s
Oh 00
p
PQ
03
i?H cd
O
'S) o
'S
& O
G
a.
on
cd Cu O
00
G r-H O
O
G
cd Wh b?
W) G O
G O
cd &.
S o
u
3
O rt ft Vh "? a
' <d co <
>? I
2 9' X ft
vo b cd ft
?^ o ^ c ^
,t? ̂ ft d> w t?
^ ?I 'ft CO
<i b?3 U c
(D
<D CO o
"t? ̂ 5 <*>
3 (D ft
> O bo C
W
?) fl ci ?H TD T3
g "t; t: 3 ?? CD
co -? "? (D .? .5 &rT >-> >-> <L> ^ ^
^ O O ci O O *-h V_l Vh
'ci O ? to . 60 'S 'S
u S S
^ O ^ r?I t?I , 00 oo 2 oo cB ̂ V ^ S !2 !Q ̂
xi xi <
o t? t? 5 CD <D ̂ co vi <i. JO X) ^J"
< < ?
On ? ?
T-H* ?
?
-H ̂
> 2 t? ci
CO CO CD CD
c -c ci ci
B B CO CO w W
8 <N
00 v?
t? t? CD CD CO CO
X> X> X < S
?
-H es m * "H ̂ ^ J
m as ? i-i
-H
o as ?
o
-H
o
?
-H o
CO
CD 00
S ^ (N
<d -^ 2 '? S
t? s
1
2 /-s Si! ON ^ Os ? 22
C^ W. t? ^ ^ ^ ra < ist, is
^ r^ r QQ U O O
^ ^^ ?o
CO S
la s!
1^
CD b? ^ bJQ "S CD S
^> On .?, 00
t?
s sa DS
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions
216 A. A. V. FLORES ET AL.
H-l pq
G #G
G O
U
t?
<
?4-1 cd O cd ft
03
^ cd ft
w -~ (N XS ^ ft "?j co ^
<D N?' C!
? S3 g C ?
Xj
"5 : ft -5 ? s ? a <i b?S U C
CD co _o
xP ft c
?00 co
r^ t? O - CD
O O
13
'S
t?
X
X2 O
b? \o "t? <-l B B
> ci
2 S
o s o? ?
o o
VO
(N d
d ^ -H
^
(N (N
X C ?? I - CD ft 00
t? 'c
I
C ta
o o ?3 tin
a
-S ^
?a i?i S
II bo xj S ci
15 T3 ' E B t?
> ^
as
s I ci g
>> 'H M .Sa ^ ?? O rt?
ci ci
O (N
en ? m X
X? O
C 'G <D >
2
o VO VO v^
4
a en . . ?a OO (N
C
w sr ci h ^ i
s ;g s 3
b ?o t? .?p ^ ci
g o Q
li li O O
X X) t? t?
ts t: CD CD
M M O o o o t-i Vh CD (D
t? t? c -a ci ci
O *n
? v? m m
as v?
^ vo ^ c=
<? r= OO ̂
On m d d -H -H
P oo v? Tt
O] ̂ -H -H
v? C^
t? ? -
2 ? ? I b? P bp
cu a, a*1
t: t: CD CD
CD <D b? q u u ci cd ci
S S S
vo i-H m ^ (N (N
O
o o en h O
?J v? V? en r^ ^
?? ̂ ^ en r^ |
^ 4 ^
vn * * O vo en odd -H -H 4H vn m oo r-H ?N CN
o5 vo en d i-? d
-H -H -H h- oo oo
i-? en en
'w' en CO "^^ S c^
^ i? fe* sr <n
^^ g ^ 0. CXh
ft X
a o
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions

MEGALOPA AND JUVENILE PACHYGRAPSUS 211
atlanticus Forest, 1957. Grapsid species with spines include riverine forms, in
which megalopae migrate from the sea back to the adult habitat, and marine
species, as is the case of Cyclograpsus integer H. Milne Edwards, 1837, the
single sesarmine presenting this feature. It is interesting to observe that there is
no described estuarine species with armed dactyli.
Plagusiines can be easily differentiated from grapsines, since the toothed dactyl condition extends to the 5th pereopod. Furthermore, in the Grapsinae the maxil
lule endopod is poorly setose, and in the case of Metopograpsus frontalis Miers,
1880, M. latifrons (White, 1847) and P. transversus, setae are absent.
This analysis does not intend to clearly separate each grapsid subfamily, but
attempts link the importance of some characteristics of the megalopal stage to
systematics; a suggestion that was previously raised by Rice (1981). Unfortu
nately, the available information concerning grapsid megalopal morphology is
not enough to allow a consistent phylogenetic analysis of the group at present. One of the most remarkable characteristics of the grapsine and, especially,
plagusiine megalopae is their size. The attainment of a big size at settlement,
besides its adaptive value mentioned above, may also allow the shortening of the
juvenile phase. Furthermore, Hines (unpubl. data) pointed out that larger recruits
may pass through fewer moults to attain the size at sexual maturity, which is an
alternative life history/growth adaptation to minimize this risky process.
The first crab stage in P. transversus is already very similar to the adults. The
density and distribution pattern of setae on the cephalic appendages and maxil
lipeds are typical of more advanced crab stages, such as those studied by Morita
(1974) in Eriocheir japonicus (De Haan, 1835). Furthermore, some behavioural
patterns like running, hiding under stones and feeding, clearly resemble the adult
life style. Sexes can be distinguished at the 2nd crab stage, and in subsequent instars the pleopods of both undergo profound and rapid differentiation. Captive
juvenile crabs from the 5th to 7th stage roughly correspond to the 6-7 mm carapace
length in wild individuals, which are usually about to achieve the puberty moult
(Flores & Negreiros-Fransozo, in prep.). Hence, it can be considered that this
species undergoes abbreviated juvenile development. In a similar way, the sexes
in the congener P. marmoratus can be distinguished in the 2nd or 3rd crab stage
and very small ovigerous females can be observed in the wild (Vernet-Cornubert,
1958). In the Plagusiinae, sexes can often be distinguished even earlier, and some
records on early postlarval stages revealed a surprisingly brief juvenile develop ment occurring in this group. Initial male gonopod differentiation at the 1st instar
was recorded by Hartnoll (1992) in Percnon abbreviatum (Dana, 1851) and by Muraoka (1963, 1965) in Plagusia dentipes (De Haan, 1835) and P. depressa
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions

218 A. A. V. FLORES ET AL.
(Fabricius, 1775) respectively. Based on the 1st crab and smaller adult crab sizes, Hartnoll (1992) also estimated that sexual maturity in Percnon abbreviatum and
P. guinotae (Crosnier, 1965) is attained in the 3rd and 4th stages respectively. Genital openings in the Varunine genus Eriocheir are visible at the 1st crab
stage (Hoestlandt, 1948; Lee et al., 1994), but in E. japonicus shrivelled non
functional pleopods remain until the 3rd instar and sexual pleopod differentiation
begins only at the 4th stage (Morita, 1974). In the Sesarmine species Sesarma
rectum Randall, 1840, remaining pleopods are present in the 1st and 2nd instars; at the 3rd stage, these structures disappear, and only at the 12th stage the defini
tive pleopods can be observed and the young crabs sexed (Fransozo, 1987). This extremely gradual process of pleopod differentiation throughout juvenile
development is likely to persist in other Sesarminae, as seems to be the case in
Armases rubripes (Rathbun, 1897). In this species, pleopod differentiation does
not take place at least until the 6th stage (Vieira & Rieger, pers. comm.).
The present comparative analysis shows that settlement and early postlarval
development in grapsid crabs tend to fall into two major patterns. Species that
recruit as larger benthic forms, mainly represented by the Grapsinae and the
Plagusiinae, present a shorter juvenile development and, therefore, a precocious sexual maturity. On the other hand, smaller recruits and long juvenile develop
ment, preceding delayed sexual maturity, seem to be characteristics of an alter
native strategy adopted chiefly by sesarmines. However, more work devoted to
the development of juvenile crabs is required before outlining more precise and
conclusive trends.
ACKNOWLEDGEMENTS
The authors are grateful to anonymous reviewers for providing valuable com
ments on earlier drafts of this manuscript. The present contribution was supported
by the "Funda?ao de Amparo ? Pesquisa do Estado de S?o Paulo" ? FAPESP
(proc. # 94/01688-3) as a Master Science fellowship to the first author.
Appendix I. Species and respective sources used in table II
SESARMINAE: Aratus pisonii: Warner (1968); Bresedium breviceps: Fielder & Greenwood
(1983b); Chasmagnathus convexus: Baba & Fukuda (1972); C. granulata: Boschi et al. (1967);
Cyclograpsus cinereus: Costlow & Fagetti (1967); C. integer. Gore & Scotto (1982); C. puncta
tus: Fagetti & Campodonico (1971); H?lice crassa: Wear & Fielder (1985); H. tridens tridens and H. t. wuana: Baba & Moryiama (1972); Helograpsus haswellianus: Fielder & Greenwood (1984);
Armases rubripes: Diaz & Ewald (1968); Sesarma catenata: Pereyra Lago (1987); S. {Holome
topus) dehaani: Baba & Myiata (1971); S. guttatum: Pereyra Lago (1993); S. meinerti: Pereyra
Lago (1989); S. rectum: Fransozo & Hebling (1986); S. reticulatum: Costlow & Bookhout, (1962); S. ricordi: Diaz & Ewald (1968).
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions

MEGALOPA AND JUVENILE PACHYGRAPSUS 219
Varuninae: Brachynotus atlanticus: Rodriguez et al. (1992); Cyrtograpsus altimanus: Scelzo
& Lichtschein de Bastida (1978); Eriocheir jap?nica {= E. japonicus): Morita (1974); E. rectus:
Shy & Yu (1992); Gaetice depressus: Muraoka (1971); Hemigrapsus nudus and H. oregonensis: Hart (1935); H. sanguineus: Muraoka (1971); Varuna litterata: Ryan & Choy (1990).
PLAGUSIINAE: Percnon abbreviatum: Hartnoll (1992); P. gibbesi: Paula & Hartnoll (1989);
P. guinotae and P. planissimum: Hartnoll (1992); Plagusia chabrus: Wear & Fielder (1985);
P. dentipes: Muraoka (1963); P. depressa: Muraoka (1965).
GRAPSINAE: Metopograpsus frontalis: Fielder & Greenwood (1983a); M. latifrons: Kakati
(1982); M. latifrons (as M. maculatus): Pasupathi & Kannupandi (1986); Pachygrapsus crassipes: Hiatt (1948); P. marmoratus: Vernet-Cornubert (1958) and Guerao et al. (in press); Planes cyaneus:
Muraoka (1973) and Wear & Fielder (1985); P. minutus: Lebour (1944).
REFERENCES
Baba, K. & Y. Fukuda, 1972. Larval development of Chasmagnathus convexus de Haan (Crus
tacea, Brachyura) reared under laboratory conditions. Mem. Fac. Educ. Kumamoto Uni v., 21
(1): 90-96.
Baba, K. & K. Miyata, 1971. Larval development of Sesarma {Holometopus) dehaani H. Milne
Edwards (Crustacea, Brachyura) reared in the laboratory. Mem. Fac. Educ. Kumamoto Uni v.,
19 (1): 49-68.
Baba, K. & M. Moriyama, 1972. Larval development of Heiice tridens wuana Rathbun and H. tri
dens De Haan (Crustacea, Brachyura) reared in the laboratory. Mem. Fac. Educ. Kumamoto
Univ., 20 (1): 49-68.
Boschi, E. E., M. A. Scelzo & B. Goldstein, 1967. Desarrollo larval de dos especies de
crust?ceos dec?podos en el laboratorio. Pachycheles haigae Rodrigues da Costa (Porcel
lanidae) y Chasmagnathus granulata Dana (Grapsidae). Bolm. Inst. Biol. mar., Mar del
Plata, 12: 1-46.
Brossi-Garcia, A. L. & M. D. RODRIGUES, 1993. Zoeal morphology of Pachygrapsus gracilis
(Saussure, 1858) (Decapoda, Grapsidae) reared in the laboratory. Invert. Reprod. Dev., 24
(3): 197-204.
Christiansen, M. E., 1973. The complete larval development of Hyas araneus (Linnaeus) and
Hyas coarctatus Leach (Decapoda, Brachyura, Majidae) reared in the laboratory. Norwegian Journ. Zool., 21: 63-89.
COSTLOW, J. D., Jr. & C. G. Bookhout, 1962. The larval development of Sesarma reticulatum
Say, reared in the laboratory. Crustaceana, 4: 281-294.
COSTLOW, J. D., JR. & E. FAGETTI, 1967. The larval development of the crab, Cyclograpsus cinereus Dana, under laboratory conditions. Pacific Sei., 21: 166-177.
CUESTA, J. A. & A. RODRIGUEZ, 1994. Early zoeal stages of Pachygrapsus marmoratus (Fabricius),
P. transversus (Gibbes) and P. maurus (Lucas) (Decapoda, Brachyura, Grapsidae) reared in
the laboratory. Scientia marina, 58 (4): 323-327.
Cuesta, J. A. & C. D. Schubart (in prep.). Morphological and molecular differentiation between
three allopatric populations of the grapsid crab Pachygrapsus transversus (Gibbes, 1850)
(Brachyura: Grapsidae: Grapsinae).
D?AZ, H. & J. J. EWALD, 1968. A comparison of the larval development of Metasesarma rubripes
(Rathbun) and Sesarma ricordi H. Milne Edwards (Brachyura, Grapsidae) reared under similar
laboratory conditions. Crustaceana, (suppl.) 2: 225-248.
FAGETTI, E. & I. Campodonico, 1971. The larval development of the crab Cyclograpsus punc tatus H. Milne Edwards, under laboratory conditions (Decapoda, Grapsidae, Sesarminae).
Crustaceana, 21 (2): 183-195.
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions

220 A. A. V. FLORES ET AL.
Felder, D. L., J. W. Martin & J. W. Goy, 1985. Patterns in early post-larval development of
decapods. In: A. M. Wenner (ed.), Factors of larval growth. Crustacean Issues, 3: 163-225.
Fielder, D. R. & J. G. Greenwood, 1983a. The complete larval development of Metopograpsus
frontalis Miers (Decapoda, Grapsidae), reared in the laboratory. Proc. R. Soc. Queensland, 94: 51-60.
-&-, 1983b. The zoeal stages and megalopa of Bresedium brevipes (De Man, 1899)
(Crustacea: Decapoda: Grapsidae), reared in the laboratory. Journ. Plankton Res., 5 (4):
585-598.
-&-, 1984. The complete larval development of Helograpsus haswellianus (Whitelegge)
(Decapoda, Grapsidae), reared in the laboratory. Proc. R. Soc. Queensland, 95: 11-22.
Flores, A. A. V. & M. L. Negreiros-Fransozo (in prep.). Postlarval allometry of the secondary
sexual characters in the shore crab Pachygrapsus transversus (Gibbes, 1850) (Brachyura,
Grapsidae).
FRANSOZO, A., 1987. Desenvolvimento dos est?gios juvenis de Sesarma {Holometopus) rectum
Randall, 1840 (Decapoda, Grapsidae) obtidos em laboratorio. Naturalia, 11/12: 77-87.
FRANSOZO, A. & N. J. Hebling, 1986. Desenvolvimento larval de Sesarma {Holometopus) rectum
Randall, 1840 (Decapoda, Grapsidae), em laboratorio. Rev. Brasileira Biol., 46 (3): 353-364.
Fransozo, A. & M. L. Negreiros-Fransozo, 1987. Morfologia dos primeiros est?gios juvenis de
Eriphia gonagra (Fabricius, 1781) e Eurypanopeus abbreviatus (Stimpson, 1860) (Crustacea,
Decapoda, Xanthidae), obtidos em laboratorio. Pap?is Avulsos Zool., 36 (22): 257-277.
Fukuda, Y, 1981. Larval development of Trigonoplax unguiformis (De Haan) (Crustacea, Bra
chyura) reared in the laboratory. Zool. Mag., Tokyo, 90 (2): 163-173.
Gore, R. H. & L. E. Scotto, 1982. Cyclograpsus integer H. Milne Edwards, 1837 (Brachyura,
Grapsidae): The complete larval development in the laboratory, with notes on larvae of the
genus Cyclograpsus. Fish. Bull., U.S., 80 (3): 501-521.
Guerao, G., P. Abell? & J. A. Cuesta (in press). Morphology of the megalopa and first crab
stage of the mediolittoral crab Pachygrapsus marmoratus (Brachyura, Grapsidae, Grapsinae). Scientia marina.
Hart, J. F. L., 1935. Larval development of British Columbia Brachyura I. Xanthidae, Pinnotheri
dae (in part) and Grapsidae. Canadian. Journ. Res. Fish., 12: 411-432.
Hartnoll, R. G., 1992. Megalopae and early post-larval stages of East African Percnon (De
capoda: Brachyura: Grapsidae). Journ. Zool. London, 228: 51-67.
Hebling, N. J., A. Fransozo & M. L. Negreiros-Fransozo, 1982. Desenvolvimento dos
primeiros est?gios juvenis de Panopeus herbstii H. Milne Edwards, 1834 (Crustacea, De
capoda, Xanthidae), criados em laboratorio. Naturalia, 7: 177-188.
HlATT, R. W, 1948. The biology of the lined shore crab Pachygrapsus crassipes Randall. Pacific
Sei., 2: 135-213.
Hiyodo, C. M., A. Fransozo & M. L. Negreiros-Fransozo, 1994. Larval development of
the spider crab Acanthonyx petiverii H. Milne Edwards, 1834 (Decapoda, Majidae) in the
laboratory. Crustaceana, 66 (1): 53-66.
HOESTLANDT, H., 1948. Recherches sur la biologie de YEriocheir sinensis en France (Crustac?,
brachyoure). Ann. Inst, oc?anogr. Monaco, 24: 1-116.
Ingle, R. W., 1977. The larval and post-larval development of the scorpion spider crab, Inachus
dorsettensis (Pennant) (family: Majidae), reared in the laboratory. Bull. British Mus. (nat.
Hist), (Zool.) 30: 331-342.
-, 1987. The first zoea of three Pachygrapsus species and of Cataleptodius floridanus (Gibbes)
from Bermuda and Mediterranean (Crustacea: Decapoda: Brachyura). Bull. British. Mus.
(nat. Hist.), (Zool.) 52: 31-41.
KAKATI, V. S., 1982. Larval development of the Indian grapsid crab, Metopograpsus latifrons
H. Milne-Edwards in vitro. Indian Journ. mar. Sei., 11: 311-316.
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions

MEGALOPA AND JUVENILE PACHYGRAPSUS 221
KONISHI, K. & M. MlNAGAWA, 1990. The first zoeal larva of the gulfweed crab Planes cyaneus
Dana, 1851 (Crustacea: Brachyura: Grapsidae). Proc. Japan Soc. syst. Zool., 42: 14-20.
Kurata, H., 1969. Larvae of decapod Brachyura of Arasaki, Sagami Bay. IV. Majidae. Bull. Tokai
reg. Fish. Res. Lab., 57: 81-127.
Lebour, M. V., 1944. Larval crabs from Bermuda. Zool?gica, New York, 29: 113-128.
LEE, T. H., M. Yamauchi & F. Yamazaki, 1994. Sex differentiation in the crab Eriocheir japonicus
(Decapoda, Grapsidae). Invert. Reprod. Dev., 25 (2): 123-138.
MORITA, T., 1974. Morphological observation on the development of larva of Eriocheir jap?nica
De Haan. Zool. Mag., Tokyo, 83: 24-81.
Muraoka, K., 1963. On the secondary sexual characters of the post-larval stages of a shore crab,
Plagusia dentipes De Haan. Res. on Crust., 1: 54-65.
-, 1965. On the post-larval stage of Plagusia depressa. Res. on Crust., 2: 83-90.
-, 1971. On the post-larval characters of the two species of shore crabs. Res. on Crust., 4/5:
225-234.
-, 1973. On the characters of the larva and the seasonal change of occurrence of the Sargassum
crab, Planes cyaneus Dana. Bull. Kanagawa Pref. Mus., 6: 45-53.
Negreiros-Fransozo, M. L. & A. Fransozo, 1991. Larval stages of Epialtus brasiliensis Dana,
1852 (Decapoda, Brachyura, Majidae) reared in the laboratory, with notes on characters of
the majid subfamilies. Crustaceana, 60 (2): 201-212.
Pasupathi, K. & T. Kannupandi, 1986. Laboratory reared larval stages of the mangrove grapsid
crab, Metopograpsus maculatus H. Milne Edwards. Mahasagar, 19 (4): 233-244.
PAULA, J., 1985. The first zoeal stages of Polybius henslowi Leach. Maja squinado (Herbst),
Pachygrapsus marmoratus (Fabricius), and Uca tangeri (Eydoux) (Crustacea, Decapoda,
Brachyura). Arq. Mus. Bocage, (BU) 17: 137-147.
Paula, J. & R. G. HARTNOLL, 1989. The larval and post-larval development of Percnon gibbesi
(Crustacea, Brachyura, Grapsidae) and the identity of the larval genus Pluteocaris. Journ.
Zool. London, 218: 17-37.
PEREYRA-LAGO, R., 1987. Larval development of Sesarma catenata Ortmann (Brachyura, Grap
sidae, Sesarminae) reared in the laboratory. South African Tydskr. Dierk., 22 (3): 200-212.
-, 1989. The larval development of the red mangrove crab Sesarma meinerti de Man
(Brachyura: Grapsidae) reared in the laboratory. South African Journ. Zool., 24 (3): 199-211.
-, 1993. Larval development of Sesarma guttatum A. Milne Edwards (Decapoda: Brachyura:
Grapsidae) reared in the laboratory, with comments on larval generic and familial characters.
Journ. Crustacean Biol., 13 (4): 745-762.
Pohle, G. & M. Telford, 1981. Morphology and classification of decapod crustacean larval
setae: a scanning electron microscopy study of Dissodactylus crinitichelis Moreira, 1901
(Brachyura: Pinnotheridae). Bull. mar. Sei., Uni v. Miami, 31 (3): 736-752.
Rice, A. L., 1981. The megalopa stage in brachyuran crabs. The Podotremata Guinot. Journ. nat.
Hist. London, 15: 1003-1011.
RlEGER, P. J., 1986. Desenvolvimento larval e juvenil de Hepatus pudibundus (Herbst, 1785)
(Decapoda, Calappidae), em laboratorio: 1-76. (M. Se. Thesis, S?o Paulo, Brasil).
Rodriguez, A., J. I. Gonzalez-Gordillo & J. A. Cuesta, 1992. Larval stages of Brachynotus atlanticus Forest, 1957 (Crustacea: Decapoda: Grapsidae) reared under laboratory conditions.
Journ. Plankton Res., 14 (6): 867-883.
ROSSIGNOL, M., 1957. IL Crustac?s D?capodes Marines de la R?gion de Pointe-Noire. In: J. Col
LIGNON, M. ROSSIGNOL & Ch. Roux (eds.), Mollusques, crustac?s, poissons marines des
c?tes d'A.E.F. en collection au Centre d'Oc?anographie de l'Institut d'?tudes Centraficaines
de Pointe-Noire: 71-136. (Minist?re de la France d'Outre-Mer, Paris).
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions

222 A. A. V. FLORES ET AL.
Ryan, P. A. & S. C. Choy, 1990. Observations on the mass upstream migration of Varuna litterata
(Fabricius) megalopae (Decapoda, Brachyura, Grapsidae) in Fiji. Crustaceana, 58 (3): 237
249.
Scelzo, M. A. & V. Lichtschein de Bastida, 1978. Desarrollo larval y metamorfosis del can
grejo Cyrtograpsus altimanus Rathbun, 1914 (Brachyura, Grapsidae) en laboratorio, con
observaciones sobre la ecologia de la especie. Physis, (A) 38 (94): 103-126.
SCHLOTTERBECK, R. E., 1976. The larval development of the lined shore crab Pachygrapsus cras
sipes Randall, 1840 (Crustacea, Brachyura, Grapsidae) reared in the laboratory. Crustaceana,
30: 184-200.
Shen, C. J., 1935. An investigation of the post-larval development of the shore crab Carcinus
maenas, with special reference to the external secondary sexual characters. Proc. Zool. Soc.
London, 1935 (1): 1-33.
Shy, J. Y & H. P. Yu, 1992. Complete larval development of the mitten crab Eriocheir rectus
Stimpson, 1858 (Decapoda, Brachyura, Grapsidae) reared in the laboratory. Crustaceana, 63
(3): 277-290.
VERNET-CORNUBERT, G., 1958. Recherches sur la sexualit? du crabe Pachygrapsus marmoratus
(Fabricius). Arch. Zool. exp. g?n., 96: 104-274.
Warner, G. F., 1968. The larval development of the mangrove tree crab, Aratus pisonii (H. Milne
Edwards) reared in the laboratory (Brachyura, Grapsidae). Studies on decapod larval devel
opment. Crustaceana, (suppl.) 2: 249-258.
Wear, R. G. & D. R. Fielder, 1985. The marine fauna of New Zealand: larvae of the Brachyura
(Crustacea, Decapoda). New Zealand oceanogr. Inst. Mem., 92: 1-90.
Wilson, K. A., 1980. Studies on Decapod Crustacea from the Indian River Region of Florida. IV.
The larval development under laboratory conditions of Euchirograpsus americanus A. Milne
Edwards, 1880 (Crustacea, Decapoda, Grapsidae) with notes on grapsid subfamilial larval
characters. Bull. mar. Sei. Univ. Miami, 30: 756-775.
Received for publication 10 December 1996.
This content downloaded from 186.217.7.233 on Thu, 5 Dec 2013 07:58:28 AMAll use subject to JSTOR Terms and Conditions