brachyuran crabs (decapoda, brachyura) associated with the green sponge amphimedon viridis...
TRANSCRIPT

Crustaceana 85 (4-5) 497-512
BRACHYURAN CRABS (DECAPODA, BRACHYURA) ASSOCIATED WITHTHE GREEN SPONGE AMPHIMEDON VIRIDIS (DEMOSPONGIAE) FROM
ITAGUÁ BEACH, SOUTH-EASTERN COAST OF BRAZIL
BY
DOUGLAS F. R. ALVES1,2,4), MARCELO C. R. CARVALHO3),SAMARA DE P. BARROS-ALVES1,2) and VALTER J. COBO2,3)
1) Postgraduate Course in Zoology, Instituto de Biociências de Botucatu,Universidade Estadual Paulista — UNESP
2) NEBECC — Group of Studies on Crustacean Biology, Ecology and Culture,Universidade Estadual Paulista — UNESP, Distrito de Rubião Junior,
CEP 18618-970, Botucatu, São Paulo, Brazil3) LabBMar (Marine Biology Laboratory), Universidade de Taubaté, UNITAU,
Av. Tiradentes, 500, Centro, Taubaté, CEP 12030-180, SP, Brazil
ABSTRACT
This study provides information on the community structure of brachyuran crabs associated withthe green sponge, Amphimedon viridis Duchassaing & Michelotti, 1864, from a beach on the south-eastern coast of Brazil. Monthly collections from May 2002 through August 2003 were carriedout on the rocky bottoms off Itaguá Beach (23°27′07′′S 45°02′49′′W). Crabs on the green spongebanks were hand-caught during snorkelling sessions, and 638 crabs belonging to 31 species, 19genera, and 10 families were obtained. Biological parameters, including sponge volume, number ofindividuals, density, richness, diversity, evenness, and dominance changed considerably over time.Species recruitment and foraging habit are some of the factors that drive the richness, dominance,and diversity patterns of this community. The green sponge banks are important sites for recruitment,refuge, and feeding of many species of brachyurans. Therefore, they are also important for theconservation of this group on the south-eastern coast of Brazil.
RESUMO
Este estudo providencia informações da estrutura da comunidade de caranguejos braquiúrosassociados com a esponja verde Amphimedon viridis Duchassaing & Michelotti, 1864 da praiado Itaguá, na costa sudeste do Brasil. Foram realizadas coletas mensais de maio de 2002 a agostode 2003 em fundos rochosos da Praia do Itaguá (23°27′07′′S 45°02′49′′W). Os caranguejos foramobtidos manualmente em bancos da esponja verde, durante sessões de mergulho livre. Foram obtidos638 caranguejos, pertencentes a 31 espécies, 19 gêneros e 10 famílias. Os parâmetros biológicos,incluindo o volume da esponja, número de indivíduos, densidade, riqueza, diversidade, equidade,
4) e-mail: [email protected]
© Koninklijke Brill NV, Leiden, 2012 DOI:10.1163/156854012X633385

498 DOUGLAS F. R. ALVES ET AL.
e dominância mudaram consideravelmente ao longo do tempo. O recrutamento das espécies e ohábito alimentar são alguns dos fatores que controlam a riqueza, dominância e diversidade destacomunidade. Os bancos da esponja verde representam sítios importantes para o recrutamento, refúgioe alimentação de muitas espécies de Brachyura. Portanto, eles também são importantes para aconservação deste grupo na costa sudeste do Brasil.
INTRODUCTION
Sessile invertebrates are common constructors of biogenic substrata, whichcan be exploited by a multitude of epibiont species. Decapods can be foundassociated with many other marine species belonging to several phyla, includingPorifera (cf. Duarte & Nalesso, 1996; Ribeiro et al., 2003; Bezerra & Coelho,2006), Cnidaria (cf. Pérez et al., 2005), Polychaeta (cf. Gore et al., 1978; Almaça,1990), Mollusca (cf. Boehs & Magalhães, 2004), Echinodermata (cf. Melo et al.,2008), and Bryozoa (cf. Mantelatto & Souza-Carey, 1998). Some of these decapodcrustaceans seem to be preferentially associated with specific biogenic substrata,such as some hermit crabs that live on sponges of the family Suberitidae (cf.Williams & McDermott, 2004); certain alpheid shrimps that also inhabit sponges;and brachyuran crabs of the genus Epialtus H. Milne Edwards, 1834 that occupybeds of Sargassum C. Agardh, 1820 on intertidal rocky shores.
Decapod crustaceans that use other living organisms as a habitat may find shelterand food; they may also spend their entire life in or on them, or alternatively mayuse these hosts for relatively short periods of time, for instance during specificstages in their ontogeny (Safriel & Eliahu-Ben, 1991; Porras et al., 1996). Exactknowledge of which species are involved in these symbiotic relationships couldprovide biological and ecological information concerning those decapods, andalso concerning the features of the supposed dependence-relationships betweenthe decapods and their biogenic substrata.
Sponges are a dominant component of many benthic communities in tropicaland temperate regions, and are common on both hard and soft substrata (Wenner etal., 1983). Sponges may be inhabited by a wide diversity of invertebrates, includingdecapod crustaceans, as reported by Klitgaard (1995), Duarte & Nalesso (1996),Caruso et al. (2005), and Bezerra & Coelho (2006).
Based on the role of sponges as biogenic substrata for brachyuran crabs, severalquestions arise: What species are most abundant in this association? Do certaingenera or species show a greater ability to exploit sponges? Do certain speciesuse sponges as a host only in a specific phase of their development? Whichspecies may use the green sponge Amphimedon viridis Duchassaing & Michelotti,1864 (Demospongiae) as a habitat during their entire post-larval life span? Theanswers to these questions will increase understanding interactions between crabs

BRACHYURA ASSOCIATED WITH AMPHIMEDON (DEMOSPONGIAE) 499
and sponges, and the potential benefits of these relationships for biodiversityconservation.
This study provides information on the community structure of crabs (Crus-tacea, Brachyura) associated with the green sponge, Amphimedon viridis fromItaguá Beach on the south-eastern coast of Brazil. The study was based on eco-logical indices, including richness, diversity, evenness, and dominance. Severalpopulation parameters of the crabs were also analysed: abundance, occurrence,size, and sex-ratio. In addition, the level of importance of the various species tothis community was evaluated by means of their abundance, relative occurrence,and equitability.
MATERIAL AND METHODS
Monthly collections were carried out on rocky bottoms off Itaguá Beach(23°27′07′′S 45°02′49′′W), Ubatuba, south-eastern coast of Brazil, from May 2002through August 2003. Sponges of the species Amphimedon viridis were hand-caught at a mean depth of 5 m, during snorkelling sessions of one hour permonth carried out by 4 divers, so totalling 4 hours of catch effort per month. Thesponges collected were stored in plastic bags during the dive, in order to avoid theloss of crabs. Samples were transferred to the Marine Biology Laboratory at theUniversidade de Taubaté — UNITAU, where the crabs were separated from thesponges.
Sponge samples were immersed in a graduated container filled with 10 L ofwater, and the sponge volume (L) was measured by water displacement, as readon the container scale. This measurement was used to assess the crab densityassociated with the sponges (individuals/m3). Water temperature (°C) and salinitywere recorded monthly at the sampling site, by means of a thermometer and opticalrefractometer, respectively.
In the laboratory, crabs sorted from the sponges were preserved with 70%ethanol, in labelled jars. For each crab, the greatest carapace width (CW) wasmeasured with Vernier callipers (nearest 0.1 mm). Sex and developmental stage(juvenile or adult) were determined from the external morphology of the abdomenand appendages, and classified as: adult female: abdomen large, covering almostthe entire surface of the thorax, and hairy pleopods; adult male: abdomen narrow,with 2 pairs of small gonopods; juvenile female and male: abdomen triangular,and 2 or 4 pairs of non-hairy pleopods, for males and females, respectively (seeHaefner, 1990).
The Brachyura were identified according to Melo (1996), and their nomencla-ture follows Ng et al. (2008). After analysis, the specimens were deposited in

500 DOUGLAS F. R. ALVES ET AL.
the Carcinology Collection of the Marine Biology Laboratory at the Universityof Taubaté (UNITAU).
The relative abundance (RAb) was calculated for each crab species, as RAb =n/N, where n = total absolute abundance recorded for a given species, and N =total of individuals recorded over the entire sampling programme. The relativeoccurrence (RO) was represented as O = p/P, where p = number of samplesin which the given species was recorded, and P = the total number of samples.Equitability (E) was also calculated for each species, represented as
E = Levinsjln(pj )
where
Levinsj = −N∑
i=1
RAbi,j · ln(RAbi,j )
pj = number of samples with occurrence of species “j”, and RAbi,j = relativeabundance of the species “j” in sample “i”. “E” corresponds to Pielou’s index, butit here refers to the equality of abundance of a given species recorded at everycollection point it occurred, as opposed to the traditional way. In order to estimatethe similarity of species present in the community, the species were groupedaccording to the Bray-Curtis similarity index, R-mode, followed by “UPGMA”clustering (Zar, 2010) on the indices (RAb, RO, and E).
Only the most important species, as shown by the grouping analyses, were usedfor the analyses of sex-ratio, frequency histograms, and median size of individuals.The sex-ratio was estimated, and tested by the chi-square test (p < 0.05) (Zar,2010).
Temporal changes in community structure were analysed through evaluationof crab density, and diversity indices (H′) (Shannon, 1948), evenness (J′) (Pielou,1966), and dominance (D) (Berger & Parker, 1970), for each of the 16 months.The diversity and evenness indices were calculated with the aid of the programKrebs/Win 0.9, and dominance with the software PAST, version 1.85.
The density, number of individuals, richness, diversity, evenness, and domi-nance were evaluated for correlation with each other by the Spearman correlationtest (α < 0.05). The associations between ecological parameters (number of indi-viduals, diversity, evenness, richness, dominance, and density) and environmentalfactors (temperature, salinity, and sponge volume) were evaluated by multiple lin-ear regressions (α < 0.05). Data were log-transformed prior to analysis, to satisfythe assumption of normality (Zar, 2010).

BRACHYURA ASSOCIATED WITH AMPHIMEDON (DEMOSPONGIAE) 501
RESULTS
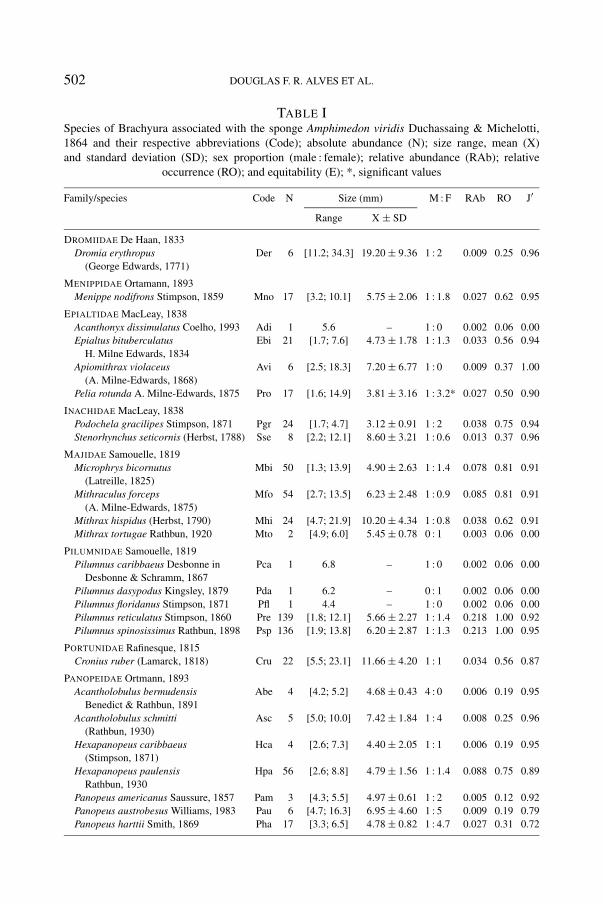
A total of 638 crabs belonging to 31 species, 19 genera, and 10 families, wascaught. The families Panopeidae and Pilumnidae were represented by the largestnumbers of species, 9 and 5, respectively. Pilumnus reticulatus Stimpson, 1860and Pilumnus spinosissimus Rathbun, 1898 comprised more than 40% of the totalnumber of individuals, and were also the only species recorded in all samples(table I).
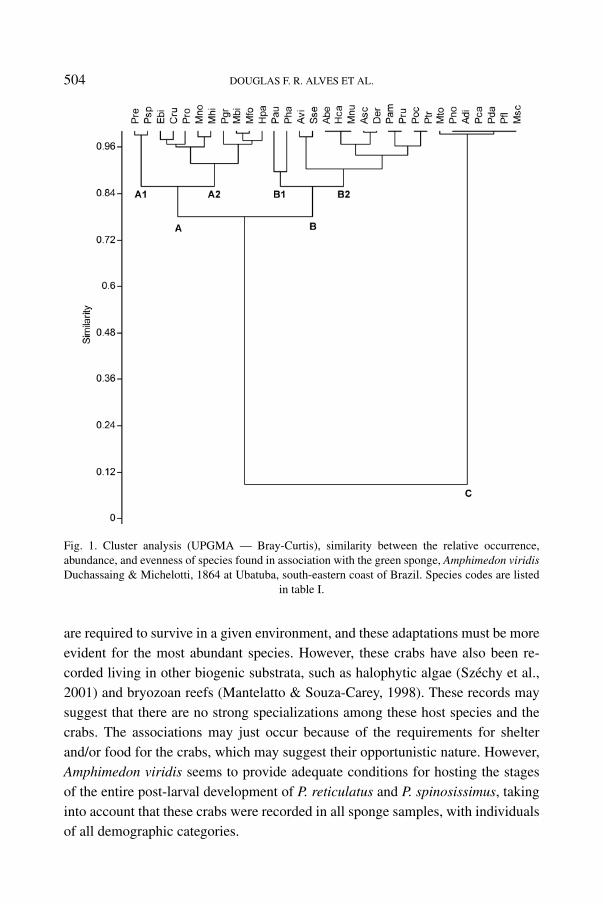
Species-importance ranking allowed the recognition of three groups (A, B,and C), composed of 11, 13, and 7 species, respectively. Group “A” was furthersubdivided into the subgroups “A1” and “A2” (fig. 1) with about 85% similarity,and this group was the most important with respect to RAb, RO, and E. The speciesbelonging to group “C” showed the smallest RAb, RO, and E, while group “B”was intermediate for these indices, and also subdivided into the subgroups “B1”and “B2”, with about 85% similarity. Subgroup “A1” was formed by 2 species,P. reticulatus and P. spinosissimus, the most important species in the community(fig. 1).
Among the species of the “A1” subgroup, P. reticulatus and P. spinosissimuswere recorded for entire demographic categories; whereas for the species ofthe “A2” subgroup, only Menippe nodifrons Stimpson, 1859, Mithrax hispidus(Herbst, 1790), and Cronius ruber (Lamarck, 1818) were not recorded in alldemographic classes, being represented only by juveniles (table I, fig. 2). Thespecies of group “A” showed the sex ratio expected according to Fisher’s theory(Fisher, 1930), that is, statistically equal to 1 : 1 (p > 0.05), except for Peliarotunda A. Milne-Edwards, 1875, which showed a bias towards females (1 : 3.2,χ2 = 4765; p = 0.029).
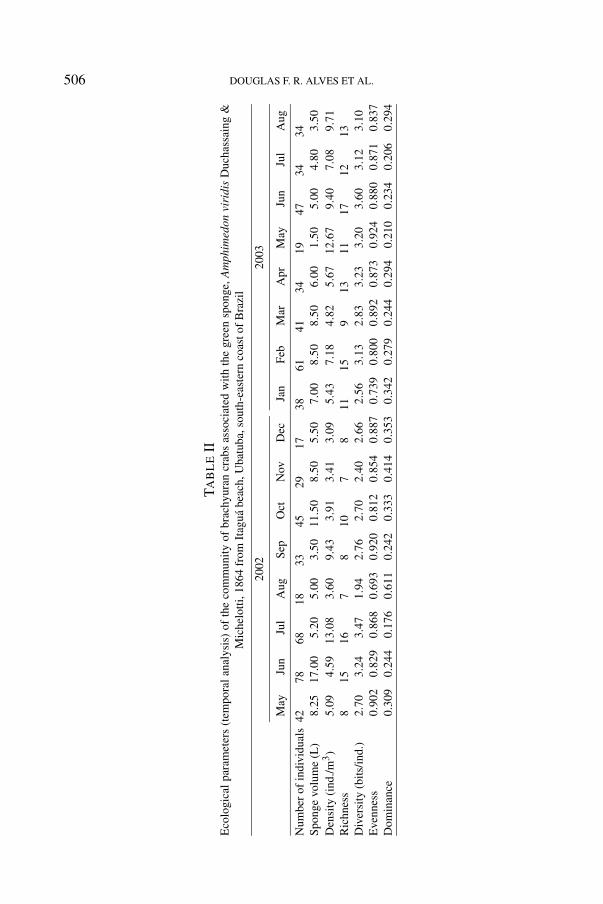
The mean number of individuals per sample was 39.9 ± 17.3, ranging from 17in December 2002 to 78 in June 2002. The largest volume of A. viridis, 17.0 L,was also recorded in June 2002 (table II).
The number of individuals was positively correlated with sponge volume,diversity, and richness (table III). The highest richness (R = 17) was recordedin June 2003, and the lowest (R = 7) in August and November 2002 (table II,fig. 3). Positive correlations were also found between richness and the number ofindividuals, density, and diversity (table III). The correlation between richness anddiversity showed the highest level of statistical significance (p < 0.001). Thisindicates the logarithmic base in which the index was calculated.
Crab diversity on the sponge banks ranged from 1.94 bits/individual in August2002 to 3.60 bits/individual in June 2003 (fig. 3a, b). The lowest evennessand highest dominance were also recorded in August 2002 (table II, fig. 3b).Positive correlations were found between diversity and the number of individuals,
502 DOUGLAS F. R. ALVES ET AL.
TABLE ISpecies of Brachyura associated with the sponge Amphimedon viridis Duchassaing & Michelotti,1864 and their respective abbreviations (Code); absolute abundance (N); size range, mean (X)and standard deviation (SD); sex proportion (male : female); relative abundance (RAb); relative
occurrence (RO); and equitability (E); *, significant values
Family/species Code N Size (mm) M : F RAb RO J′
Range X ± SD
DROMIIDAE De Haan, 1833Dromia erythropus Der 6 [11.2; 34.3] 19.20 ± 9.36 1 : 2 0.009 0.25 0.96
(George Edwards, 1771)
MENIPPIDAE Ortamann, 1893Menippe nodifrons Stimpson, 1859 Mno 17 [3.2; 10.1] 5.75 ± 2.06 1 : 1.8 0.027 0.62 0.95
EPIALTIDAE MacLeay, 1838Acanthonyx dissimulatus Coelho, 1993 Adi 1 5.6 – 1 : 0 0.002 0.06 0.00Epialtus bituberculatus Ebi 21 [1.7; 7.6] 4.73 ± 1.78 1 : 1.3 0.033 0.56 0.94
H. Milne Edwards, 1834Apiomithrax violaceus Avi 6 [2.5; 18.3] 7.20 ± 6.77 1 : 0 0.009 0.37 1.00
(A. Milne-Edwards, 1868)Pelia rotunda A. Milne-Edwards, 1875 Pro 17 [1.6; 14.9] 3.81 ± 3.16 1 : 3.2* 0.027 0.50 0.90
INACHIDAE MacLeay, 1838Podochela gracilipes Stimpson, 1871 Pgr 24 [1.7; 4.7] 3.12 ± 0.91 1 : 2 0.038 0.75 0.94Stenorhynchus seticornis (Herbst, 1788) Sse 8 [2.2; 12.1] 8.60 ± 3.21 1 : 0.6 0.013 0.37 0.96
MAJIDAE Samouelle, 1819Microphrys bicornutus Mbi 50 [1.3; 13.9] 4.90 ± 2.63 1 : 1.4 0.078 0.81 0.91
(Latreille, 1825)Mithraculus forceps Mfo 54 [2.7; 13.5] 6.23 ± 2.48 1 : 0.9 0.085 0.81 0.91
(A. Milne-Edwards, 1875)Mithrax hispidus (Herbst, 1790) Mhi 24 [4.7; 21.9] 10.20 ± 4.34 1 : 0.8 0.038 0.62 0.91Mithrax tortugae Rathbun, 1920 Mto 2 [4.9; 6.0] 5.45 ± 0.78 0 : 1 0.003 0.06 0.00
PILUMNIDAE Samouelle, 1819Pilumnus caribbaeus Desbonne in Pca 1 6.8 – 1 : 0 0.002 0.06 0.00
Desbonne & Schramm, 1867Pilumnus dasypodus Kingsley, 1879 Pda 1 6.2 – 0 : 1 0.002 0.06 0.00Pilumnus floridanus Stimpson, 1871 Pfl 1 4.4 – 1 : 0 0.002 0.06 0.00Pilumnus reticulatus Stimpson, 1860 Pre 139 [1.8; 12.1] 5.66 ± 2.27 1 : 1.4 0.218 1.00 0.92Pilumnus spinosissimus Rathbun, 1898 Psp 136 [1.9; 13.8] 6.20 ± 2.87 1 : 1.3 0.213 1.00 0.95
PORTUNIDAE Rafinesque, 1815Cronius ruber (Lamarck, 1818) Cru 22 [5.5; 23.1] 11.66 ± 4.20 1 : 1 0.034 0.56 0.87
PANOPEIDAE Ortmann, 1893Acantholobulus bermudensis Abe 4 [4.2; 5.2] 4.68 ± 0.43 4 : 0 0.006 0.19 0.95
Benedict & Rathbun, 1891Acantholobulus schmitti Asc 5 [5.0; 10.0] 7.42 ± 1.84 1 : 4 0.008 0.25 0.96
(Rathbun, 1930)Hexapanopeus caribbaeus Hca 4 [2.6; 7.3] 4.40 ± 2.05 1 : 1 0.006 0.19 0.95
(Stimpson, 1871)Hexapanopeus paulensis Hpa 56 [2.6; 8.8] 4.79 ± 1.56 1 : 1.4 0.088 0.75 0.89
Rathbun, 1930Panopeus americanus Saussure, 1857 Pam 3 [4.3; 5.5] 4.97 ± 0.61 1 : 2 0.005 0.12 0.92Panopeus austrobesus Williams, 1983 Pau 6 [4.7; 16.3] 6.95 ± 4.60 1 : 5 0.009 0.19 0.79Panopeus harttii Smith, 1869 Pha 17 [3.3; 6.5] 4.78 ± 0.82 1 : 4.7 0.027 0.31 0.72

BRACHYURA ASSOCIATED WITH AMPHIMEDON (DEMOSPONGIAE) 503
TABLE I(Continued)
Family/species Code N Size (mm) M : F RAb RO J′
Range X ± SD
PANOPEIDAE Ortmann, 1893Panopeus occidentalis Saussure, 1857 Poc 2 [6.1; 8.6] 7.35 ± 1.77 0 : 2 0.003 0.12 1.00Panopeus rugosus Pru 3 [3.8; 6.2] 5.13 ± 1.22 1 : 0.5 0.005 0.12 0.92
A. Milne-Edwards, 1880
XANTHIDAE MacLeay, 1838Paractaea nodosa (Stimpson, 1860) Pno 1 14.9 – 0 : 1 0.002 0.06 0.00Micropanope nuttingi (Rathbun, 1898) Mnu 4 [2.2; 5.0] 3.53 ± 1.20 1 : 0 0.006 0.19 0.95Micropanope sculptipes Msc 1 2.6 – 1 : 0 0.002 0.06 0.00
Stimpson, 1871
GRAPSIDAE MacLeay, 1838Pachygrapsus transversus Ptr 2 [2.9; 8.5] 5.70 ± 3.96 1 : 1 0.003 0.12 1.00
(Gibbes, 1850)
density, richness, and dominance (table III). The correlation between diversityand dominance showed the highest level of statistical significance (p < 0.001)(table III).
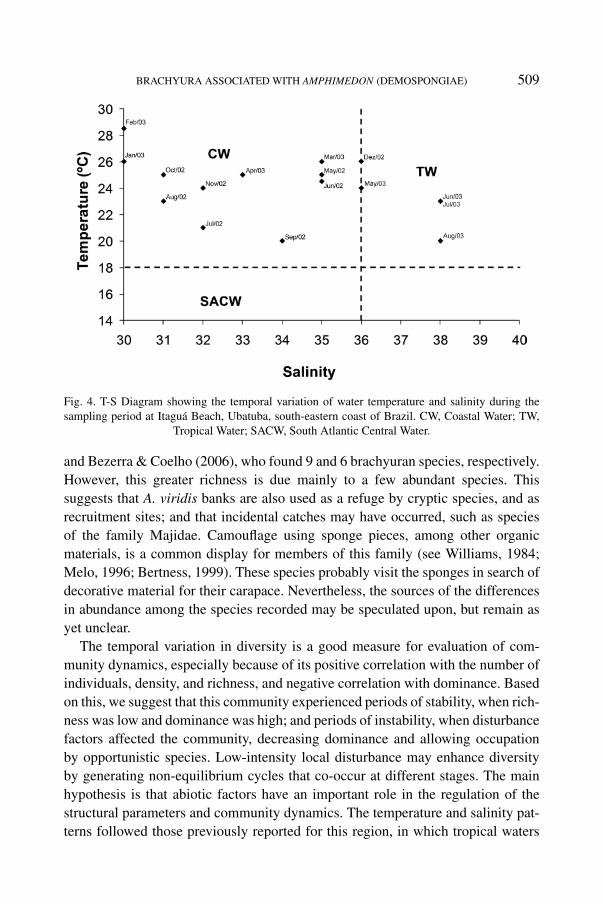
During the study period, the influences of two water-bodies were identifiedthrough the T-S diagram (Temperature and Salinity). Coastal Water (CW: T >
20°C, S < 36) generally predominated. Tropical Water (TW: T > 20°C, S > 36)only dominated from June through August 2003 (fig. 4).
For the relationships between ecological parameters and environmental factors,positive associations were obtained between salinity vs. evenness (t = 2.462;p = 0.028) and sponge volume vs. number of individuals (t = 2.850; p = 0.014).In contrast, salinity and dominance were negatively associated (t = −2.329;p = 0.038).
DISCUSSION
The composition of the crab community found suggests a complex structure,comprising species from different superfamilies that exploit the sponge banks.These observations agree with those of previous investigations along the Braziliancoast (see Ribeiro et al., 2003; Bezerra & Coelho, 2006). The superfamiliesMajoidea and Pilumnoidea are the most common groups that occur on spongebanks, which seems to reflect the affinity between these animals in symbiosis, asreported previously by Bezerra & Coelho (2006).
The crabs Pilumnus reticulatus and P. spinosissimus seem to be more special-ized to live in sponge banks, allowing them to exploit the association most suc-cessfully. Abele (1974) stated that morphological and physiological adaptations
504 DOUGLAS F. R. ALVES ET AL.
Fig. 1. Cluster analysis (UPGMA — Bray-Curtis), similarity between the relative occurrence,abundance, and evenness of species found in association with the green sponge, Amphimedon viridisDuchassaing & Michelotti, 1864 at Ubatuba, south-eastern coast of Brazil. Species codes are listed
in table I.
are required to survive in a given environment, and these adaptations must be moreevident for the most abundant species. However, these crabs have also been re-corded living in other biogenic substrata, such as halophytic algae (Széchy et al.,2001) and bryozoan reefs (Mantelatto & Souza-Carey, 1998). These records maysuggest that there are no strong specializations among these host species and thecrabs. The associations may just occur because of the requirements for shelterand/or food for the crabs, which may suggest their opportunistic nature. However,Amphimedon viridis seems to provide adequate conditions for hosting the stagesof the entire post-larval development of P. reticulatus and P. spinosissimus, takinginto account that these crabs were recorded in all sponge samples, with individualsof all demographic categories.
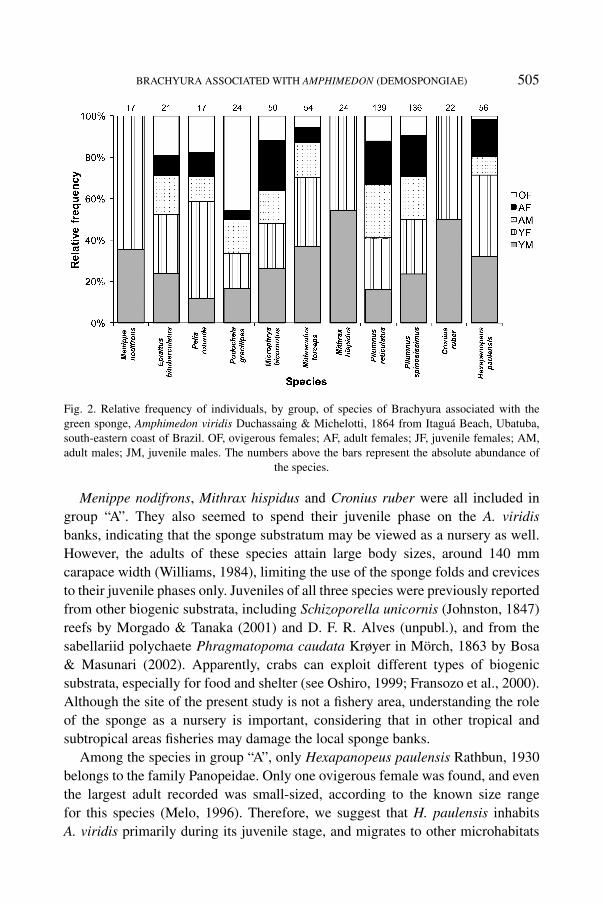
BRACHYURA ASSOCIATED WITH AMPHIMEDON (DEMOSPONGIAE) 505
Fig. 2. Relative frequency of individuals, by group, of species of Brachyura associated with thegreen sponge, Amphimedon viridis Duchassaing & Michelotti, 1864 from Itaguá Beach, Ubatuba,south-eastern coast of Brazil. OF, ovigerous females; AF, adult females; JF, juvenile females; AM,adult males; JM, juvenile males. The numbers above the bars represent the absolute abundance of
the species.
Menippe nodifrons, Mithrax hispidus and Cronius ruber were all included ingroup “A”. They also seemed to spend their juvenile phase on the A. viridisbanks, indicating that the sponge substratum may be viewed as a nursery as well.However, the adults of these species attain large body sizes, around 140 mmcarapace width (Williams, 1984), limiting the use of the sponge folds and crevicesto their juvenile phases only. Juveniles of all three species were previously reportedfrom other biogenic substrata, including Schizoporella unicornis (Johnston, 1847)reefs by Morgado & Tanaka (2001) and D. F. R. Alves (unpubl.), and from thesabellariid polychaete Phragmatopoma caudata Krøyer in Mörch, 1863 by Bosa& Masunari (2002). Apparently, crabs can exploit different types of biogenicsubstrata, especially for food and shelter (see Oshiro, 1999; Fransozo et al., 2000).Although the site of the present study is not a fishery area, understanding the roleof the sponge as a nursery is important, considering that in other tropical andsubtropical areas fisheries may damage the local sponge banks.
Among the species in group “A”, only Hexapanopeus paulensis Rathbun, 1930belongs to the family Panopeidae. Only one ovigerous female was found, and eventhe largest adult recorded was small-sized, according to the known size rangefor this species (Melo, 1996). Therefore, we suggest that H. paulensis inhabitsA. viridis primarily during its juvenile stage, and migrates to other microhabitats
506 DOUGLAS F. R. ALVES ET AL.
TA
BL
EII
Eco
logi
cal
para
met
ers
(tem
pora
lan
alys
is)
ofth
eco
mm
unity
ofbr
achy
uran
crab
sas
soci
ated
with
the
gree
nsp
onge
,Am
phim
edon
viri
dis
Duc
hass
aing
&M
iche
lotti
,186
4fr
omIt
aguá
beac
h,U
batu
ba,s
outh
-eas
tern
coas
tof
Bra
zil
2002
2003
May
Jun
Jul
Aug
Sep
Oct
Nov
Dec
Jan
Feb
Mar
Apr
May
Jun
Jul
Aug
Num
ber
ofin
divi
dual
s42
7868
1833
4529
1738
6141
3419
4734
34Sp
onge
volu
me
(L)
8.25
17.0
05.
205.
003.
5011
.50
8.50
5.50
7.00
8.50
8.50
6.00
1.50
5.00
4.80
3.50
Den
sity
(ind
./m3)
5.09
4.59
13.0
83.
609.
433.
913.
413.
095.
437.
184.
825.
6712
.67
9.40
7.08
9.71
Ric
hnes
s8
1516
78
107
811
159
1311
1712
13D
iver
sity
(bits
/ind.
)2.
703.
243.
471.
942.
762.
702.
402.
662.
563.
132.
833.
233.
203.
603.
123.
10E
venn
ess
0.90
20.
829
0.86
80.
693
0.92
00.
812
0.85
40.
887
0.73
90.
800
0.89
20.
873
0.92
40.
880
0.87
10.
837
Dom
inan
ce0.
309
0.24
40.
176
0.61
10.
242
0.33
30.
414
0.35
30.
342
0.27
90.
244
0.29
40.
210
0.23
40.
206
0.29
4

BRACHYURA ASSOCIATED WITH AMPHIMEDON (DEMOSPONGIAE) 507
TABLE IIIThe “p” values of the Spearman correlation analysis among the ecological parameters. Vol., spongevolume; N° Ind., number of individuals; Dens., density; R, richness; H′, diversity; J′, evenness; D,
dominance; *, significant (p < 0.05)
Vol. N° Ind. Dens. R H′ J′ D
Vol. 0N° Ind. 0.040* 0Dens. 0.012* 0.337 0R 0.922 0.002* 0.011* 0H′ 0.612 0.018* 0.005* >0.001* 0J′ 0.140 0.373 0.279 0.639 0.344 0D 0.161 0.114 >0.001* 0.007 >0.001* 0.080 0
later, as reported by Fransozo et al. (1992) for sand bottoms, and by Melo (2008)for rocky bottoms, by Széchy et al. (2001) for halophytic algae, and by Mantelatto& Souza-Carey (1998) for bryozoan reefs.
Almost all group “A” species showed the expected 1 : 1 sex-ratio, accordingto Fisher’s theory (Fisher, 1930), suggesting that males and females exploit thesponge banks in a similar way. A departure from the 50% sex-ratio was observedonly for Pelia rotunda, which showed a higher proportion of females. This mightrelate to habitat partitioning for males and females during development, or toconstraints from differential birth and mortality rates between the sexes. However,the precise reason or reasons for this sex-ratio departure still need clarification.
The species of group “A2” might be visitors or residents that show some degreeof seasonality. The remaining species, belonging to groups “B” and “C”, werecharacterized as accidental or rare. Some species from groups “B” and “C” may beviewed as rare for the study site, e.g., Paractea nodosa (Stimpson, 1860), Pilumnusdasypodus Kingsley, 1879 and Pilumnus floridanus Stimpson, 1871; whereas otherspecies that were found in low abundance in this study are commonly recorded inother habitats in the same region: these include Stenorhynchus seticornis (Herbst,1788) and Acantholobulus schmitti (Rathbun, 1930), reported by Mantelatto et al.(2004) and Braga et al. (2005), respectively. This situation of minimal frequencymay have different biological meanings, such as cryptic behaviour, migration, ordispersal mechanisms; or the influence of adjacent habitats, which might reducethe frequency of individuals and thus the catch efficiency.
The richness of crabs recorded here (31 species) is considerably greater thanthat reported for other biogenic substrates on the south-eastern Brazilian coast,such as sand reefs of Phragmatopoma lapidosa Kinberg, 1867 (cf. Pinheiro et al.,1997) and the bryozoan Schizoporella unicornis (cf. Mantelatto & Souza-Carey,1998), where 5 and 20 crab species were found, respectively; as well as for othersponge banks along the Brazilian coast, as reported by Duarte & Nalesso (1996)
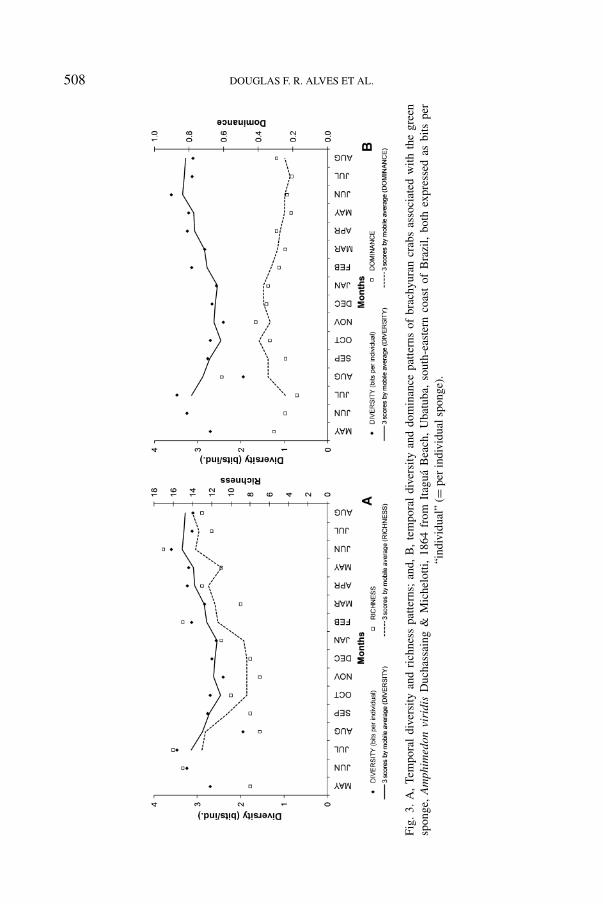
508 DOUGLAS F. R. ALVES ET AL.
Fig.
3.A
,Te
mpo
ral
dive
rsity
and
rich
ness
patte
rns;
and,
B,
tem
pora
ldi
vers
ityan
ddo
min
ance
patte
rns
ofbr
achy
uran
crab
sas
soci
ated
with
the
gree
nsp
onge
,A
mph
imed
onvi
ridi
sD
ucha
ssai
ng&
Mic
helo
tti,
1864
from
Itag
uáB
each
,U
batu
ba,
sout
h-ea
ster
nco
ast
ofB
razi
l,bo
thex
pres
sed
asbi
tspe
r“i
ndiv
idua
l”(=
per
indi
vidu
alsp
onge
).
BRACHYURA ASSOCIATED WITH AMPHIMEDON (DEMOSPONGIAE) 509
Fig. 4. T-S Diagram showing the temporal variation of water temperature and salinity during thesampling period at Itaguá Beach, Ubatuba, south-eastern coast of Brazil. CW, Coastal Water; TW,
Tropical Water; SACW, South Atlantic Central Water.
and Bezerra & Coelho (2006), who found 9 and 6 brachyuran species, respectively.However, this greater richness is due mainly to a few abundant species. Thissuggests that A. viridis banks are also used as a refuge by cryptic species, and asrecruitment sites; and that incidental catches may have occurred, such as speciesof the family Majidae. Camouflage using sponge pieces, among other organicmaterials, is a common display for members of this family (see Williams, 1984;Melo, 1996; Bertness, 1999). These species probably visit the sponges in search ofdecorative material for their carapace. Nevertheless, the sources of the differencesin abundance among the species recorded may be speculated upon, but remain asyet unclear.
The temporal variation in diversity is a good measure for evaluation of com-munity dynamics, especially because of its positive correlation with the number ofindividuals, density, and richness, and negative correlation with dominance. Basedon this, we suggest that this community experienced periods of stability, when rich-ness was low and dominance was high; and periods of instability, when disturbancefactors affected the community, decreasing dominance and allowing occupationby opportunistic species. Low-intensity local disturbance may enhance diversityby generating non-equilibrium cycles that co-occur at different stages. The mainhypothesis is that abiotic factors have an important role in the regulation of thestructural parameters and community dynamics. The temperature and salinity pat-terns followed those previously reported for this region, in which tropical waters

510 DOUGLAS F. R. ALVES ET AL.
predominated in autumn and winter (see Castro-Filho et al., 1987; Pires-Vanin etal., 1993; Mantelatto & Fransozo, 1999). Temperature seemed to act seasonally oncrab density on the A. viridis banks. Indeed, the macrobenthos structure on the con-tinental shelf off Ubatuba is related to hydrodynamic patterns (Pires-Vanin, 1993).
The occurrence of opportunistic species seems to be related to recruitmentevents and foraging behaviour, which are influenced by the richness, dominance,and diversity dynamics of the brachyuran community associated with the A. viridisbanks. The green sponge A. viridis provides a highly heterogeneous substrate atthe millimetre level, offering shelter for marine invertebrates of many sizes. Theseshelters, their morphology, and the substrate features seem to play an importantrole in recruitment success for many species (Shulman, 1984).
The influence of disturbance factors on the benthos depends on their intensity,length, and frequency, but all these factors affect recruitment and life-history traits(Posey et al., 1996). The banks of Amphimedon viridis are an important biogenicsubstrate, providing sites for recruitment, shelter, and foraging, and contributing tothe maintenance of the brachyuran fauna of the south-eastern coast of Brazil.
ACKNOWLEDGEMENTS
The authors are indebted to Dr. Gustavo Augusto Schmidt de Melo for thecareful examination of specimens. Dr. Janet W. Reid kindly revised the English.Thanks are also due to colleagues of the LabBMar/UNITAU, who assisted in thedive sessions. All sampling was conducted in accordance with applicable state andfederal laws.
REFERENCES
ABELE, L. G., 1974. Species diversity of decapod crustaceans in marine habitats. Ecology, 55:156-161.
ALMAÇA, C., 1990. Structure and interactions in the crab community inhabiting sabellariid wormcolonies at Praia de Ribeira d’Ilhas (Ericeira, Portugal). Arq. Mus. Bocage, 37: 505-519.
BERGER, W. H. & F. L. PARKER, 1970. Diversity of planktonic Foraminifera in deep-sea sediments.Science, New York, 168: 1345-1347.
BERTNESS, M. D., 1999. The ecology of Atlantic shorelines: 1-417. (Sinauer, Sunderland).BEZERRA, L. E. A. & P. A. COELHO, 2006. Crustáceos decápodos associados a esponjas no litoral
do Estado do Ceará, Brasil. Rev. Brasileira Zool., 23: 699-702.BOEHS, G. & A. R. M. MAGALHÃES, 2004. Simbiontes associados com Anomalocardia brasiliana
(Gmelin) (Mollusca, Bivalvia, Veneridae) na Ilha de Santa Catarina e região continentaladjacente, Santa Catarina, Brasil. Rev. Brasileira Zool., 21: 865-869.
BOSA, C. R. & S. MASUNARI, 2002. Crustáceos decápodos associados aos bancos de Phrag-matopoma caudata (Kröyer) (Polychaeta, Sabellariidae) na Praia de Caiobá, Matinhos, Paraná.Rev. Brasileira Zool., 19: 117-133.

BRACHYURA ASSOCIATED WITH AMPHIMEDON (DEMOSPONGIAE) 511
BRAGA, A. A., A. FRANSOZO, G. BERTINI & P. B. FUMIS, 2005. Composição e abundânciados caranguejos (Decapoda, Brachyura) nas regiões de Ubatuba e Caraguatatuba, litoral nortepaulista, Brasil. Biota Neotropica, 5(2): 1-34.
CARUSO, T., L. FALCIAI & V. ZUPO, 2005. Do hermit crabs like living in sponges? Paguristeseremita and Suberites domuncula: biometric data from the southern Mediterranean Sea. Journ.mar. biol. Ass. U. K., 85: 1353-1357.
CASTRO-FILHO, B. M., L. B. MIRANDA & S. Y. MYAO, 1987. Condições hidrográficas naplataforma continental ao largo de Ubatuba: variações sazonais e em média escala. Bol. Inst.Oceanogr. São Paulo, 35: 135-151.
DUARTE, L. F. L. & R. C. NALESSO, 1996. The sponge Zygomycale parishii (Bowerbank) and itsendobiotic fauna. Est. coast. Shelf Sci., 42: 139-151.
FISHER, R. A., 1930. The genetical theory of natural selection: 1-291. (Dover, New York).FRANSOZO, A., G. BERTINI & M. O. D. CORRÊA, 2000. Population biology and habitat utilization
of the stone crab Menippe nodifrons Stimpson, 1859 (Decapoda, Xanthidae) in Ubatuba region,Brazil. In: J. C. VON VAUPEL KLEIN & F. R. SCHRAM (eds.), The biodiversity crisis andCrustacea: 275-281. (Crustacean Issues, 12, A.A. Balkema, Rotterdam).
FRANSOZO, A., M. L. NEGREIROS-FRANSOZO, F. L. M. MANTELATTO & S. SANTOS, 1992.Composição e distribuição dos Brachyura (Crustacea, Decapoda) do sublitoral não consolidadona Enseada da Fortaleza, Ubatuba (SP). Rev. Brasileira Zool., 52(4): 667-675.
GORE, R. H., L. E. SCOTTO & L. J. BECKER, 1978. Community composition, stability, and trophicpartitioning in decapod crustaceans inhabiting some subtropical sabellariid worm reefs. Bull.mar. Sci., 28: 221-248.
HAEFNER, P. A., JR., 1990. Morphometry and size at maturity of Callinectes ornatus (Brachyura,Portunidae) in Bermuda. Bull. mar. Sci., 46: 274-286.
KLITGAARD, A. B., 1995. The fauna associated with outer shelf and upper slope sponges (Porifera,Demospongiae) at the Faroe Islands, northeastern Atlantic. Sarsia, 80: 1-22.
MANTELATTO, F. L. M., F. C. R. FARIA, R. BIAGI & G. A. S. MELO, 2004. Majoid crabscommunity (Crustacea: Decapoda) from infralittoral rocky/sandy bottom of Anchieta Island,Ubatuba. Brazilian Arch. Biol. Techn., 47: 273-279.
MANTELATTO, F. L. M. & A. FRANSOZO, 1999. Characterization of the physical and chemicalparameters of Ubatuba Bay, northern coast of São Paulo State, Brazil. Rev. Brasileira Biol.,59(1): 23-31.
MANTELATTO, F. L. M. & M. M. SOUZA-CAREY, 1998. Brachyura (Crustacea, Decapoda)associated to Schizoporella unicornis (Bryozoa, Gymnolaemata) in Ubatuba Bay (SP), Brazil.Brazilian Arch. Biol. Techn., 41: 212-217.
MELO, G. A. S., 1996. Manual de identificação dos Brachyura (caranguejos e siris) do litoralbrasileiro: 1-604. (Editora Plêiade, São Paulo).
— —, 2008. The Brachyura (Decapoda) of Ilha Grande Bay, Rio de Janeiro, Brazil. Nauplius, 16(1):1-22.
MELO, J. T. A., A. E. SILVA, M. A. SILVA & L. L. G. SOUZA, 2008. Estudo comparativoda ocorrência de Mellita quinquiesperforata (Leske, 1778) (Echinodermata, Echinoidea) nasPraias de Cotovelo e Genipabu, no Estado do Rio Grande do Norte, Brasil. Rev. Publ.Ca., 4:47-57.
MORGADO, E. H. & M. O. TANAKA, 2001. The macrofauna associated with the bryozoanSchizoporella errata (Walters) in southeastern Brazil. Sci. mar., 65(3): 173-181.
NG, P. K. L., D. GUINOT & P. J. F. DAVIE, 2008. Systema Brachyurorum: Part I. An annotatedchecklist of extant brachyuran crabs of the world. Raffles Bull. Zool., (Suppl.) 17: 1-208.
OSHIRO, L. M. Y., 1999. Aspectos reprodutivos do caranguejo guaiá, Menippe nodifrons Stimpson(Crustacea, Decapoda, Xanthidae) da Baía de Septiba, Rio de Janeiro, Brasil. Rev. BrasileiraZool., 16(3): 827-834.

512 DOUGLAS F. R. ALVES ET AL.
PÉREZ, C. D., D. A. VILA-NOVA & A. M. SANTOS, 2005. Associated community with thezoanthid Palythoa caribaeorum (Duchassaing & Michelotti, 1860) (Cnidaria, Anthozoa) fromlitoral of Pernambuco, Brasil. Hydrobiologia, 548: 207-215.
PIELOU, E. C., 1966. The measurement of diversity in different types of biological collections.Journ. theor. Biol., 13: 131-144.
PINHEIRO, M. A. A., G. BERTINI, L. C. FERNANDES-GÓES & A. FRANSOZO, 1997. Decapodcrustaceans associated to sands reefs of Phragmatopoma lapidosa Kinberg, 1867 (Polychaeta,Sabellariidae) at Praia Grande, Ubatuba, SP, Brazil. Nauplius, 5: 77-83.
PIRES-VANIN, A. M. S., 1993. A macrofauna bêntica da plataforma continental ao largo deUbatuba, São Paulo, Brasil. Publção esp. Inst. Oceanogr., 10: 137-158.
PIRES-VANIN, A. M. S., C. L. D. B. ROSSI-WONGTSCHOWSKI, E. AIDAR, H. S. L. MESQUITA,L. S. H. SOARES, M. KATSURAGAWA & Y. MATSUURA, 1993. Estrutura e função doecossistema de plataforma continental do Atlântico Sul brasileiro: síntese dos resultados.Publção esp. Inst. Oceanogr., 10: 217-231.
PORRAS, R., J. V. BATALLER, E. MURGUI & M. T. TORREGROSA, 1996. Trophic-structure andcommunity composition of polychaetes inhabiting some Sabellaria alveolata (L.) reefs alongthe Valencia Gulf coast, Western Mediterranean. Mar. Ecol., 17(4): 583-602.
POSEY, M., W. LINDBERG, T. ALPHIN & F. VOSE, 1996. Influence of storm disturbance on anoffshore benthic community. Bull. mar. Sci., 59: 523-529.
RIBEIRO, S. M., E. P. OMENA & G. MURICY, 2003. Macrofauna associated to Mycale microsig-matosa (Porifera, Demospongiae) in Rio de Janeiro State, SE Brazil. Est. coast. Shelf Sci., 57:951-959.
SAFRIEL, U. N. & M. N. BEN-ELIAHU, 1991. The influence of habitat structure and environmentalstability on the species diversity of polychaetes in vermetid reefs. In: S. S. BELL, E. D.MCCOY & H. R. MUSHINSKY (eds.), Habitat structure — the physical arrangement of objectsin space: 349-369. (Chapman and Hall, London).
SHANNON, C. E., 1948. A mathematical theory of communication. Bell System Technical Journ.,27: 379-423.
SHULMAN, M. J., 1984. Resource limitation and recruitment patterns in a coral reef fish assemblage.Journ. exp. mar. Biol. Ecol., 74: 85-109.
SZÉCHY, M. T. M., V. G. VELOSO & E. J. PAULA, 2001. Brachyura (Decapoda, Crustacea) ofphytobenthic communities of the sublittoral region of rocky shores of Rio de Janeiro and SãoPaulo, Brazil. Tropical Ecology, 42(2): 231-242.
WENNER, E. L., D. M. KNOTT, R. F. VAN DOLAH & V. G. BURRELL, 1983. Invertebratecommunities associated with hard bottom habitats in the South Atlantic Bight. Estuar. coast.mar. Sci., 17: 143-158.
WILLIAMS, A. B., 1984. Shrimps, lobsters, and crabs of the Atlantic coast of the eastern UnitedStates, Maine to Florida: 1-550. (Smithsonian Institution Press, Washington).
WILLIAMS, J. D. & J. J. MCDERMOTT, 2004. Hermit crab biocoenoses: a worldwide review of thediversity and natural history of hermit crab associates. Journ. exp. mar. Biol. Ecol., 305: 1-128.
ZAR, J. H., 2010. Biostatistical analysis (5th ed.): 1-944. (Prentice-Hall, Upper Saddle River, NewJersey).
First received 2 August 2011.Final version accepted 22 December 2011.