the male copulatory system of european pea crabs (crustacea, brachyura, pinnotheridae)
TRANSCRIPT

The Male Copulatory System of European Pea Crabs(Crustacea, Brachyura, Pinnotheridae)
Carola Becker,1* Michael Turkay,1 and Dirk Brandis2
1Senckenberg Research Institute andNatural HistoryMuseum, Senckenberganlage 25, D-60325 Frankfurt/Main, Germany2Zoological Museum, Christian-Albrechts-University of Kiel, Hegewischstr. 3, D-24105 Kiel, Germany
ABSTRACT The male copulatory system of the Euro-pean pinnotherid species Pinnotheres pisum, Pinnotherespectunculi, and Nepinnotheres pinnotheres was investi-gated by gross morphology, scanning electron micros-copy, histological methods, and confocal laser scanningmicroscopy. The brachyuran copulatory system is consis-tently formed by paired penes and two pairs of abdomi-nal appendages, the gonopods, functioning in spermtransfer. In pinnotherids, the long first gonopods trans-fer the sperm mass into the female ducts. The first gono-pod has the ejaculatory canal inside that opens bothbasally and distally. The second gonopod is solid, short,and conical. During copulation, the penis and the secondgonopod are inserted into the basal lumen of the firstgonopod. While the penis injects the sperm mass, the sec-ond gonopod functions in the transport of spermatozoainside the ejaculatory canal toward its distal opening. Thesecond gonopod is adapted for the sealing of the tubularsystem in the first gonopod by its specific shape and theability to swell. Longitudinal cuticle foldings of the secondgonopod hook into structures inside the first gonopod. Thesecond gonopod can interact with the penis during copula-tion by a flexible flap separating the lumina in which thesecond gonopod and the penis are inserted. J. Morphol.273:1306–1318, 2012. � 2012 Wiley Periodicals, Inc.
KEY WORDS: Pinnotheres; morphology; gonopods;copulatory system; histology; confocal laser scanningmicroscopy
INTRODUCTION
Among true crabs (i.e., Brachyura Linnaeus,1758), internal fertilization has evolved. In ances-tral crabs (‘‘Podotremata’’ Guinot, 1977), the malesperm mass is transferred to simple invaginationsof the female’s sternum, while more complexstorage structures with an inner connection tothe oviduct, the receptacula semini have developedamong Eubrachyura Saint-Laurent, 1980 (in-cluding Heterotremata and Thoracotremata Gui-not, 1977). The male reproductive system co-evolved with the female genital ducts, in order tooptimize sperm transfer, probably driven by thetransition from external to internal fertilizationand by carcinization (Turkay, 1975; Hartnoll, 1979;Guinot and Quenette, 2005).
The internal male reproductive structures con-sist of paired gonads, the testes, and paired vasa
deferentia. Decapod spermatozoa are aflagellateand immotile. In the vas deferens, they becomeenclosed inside spermatophores and embedded intoa matrix of seminal plasma. Prior to copulation,the sperm mass is stored inside the long and coiledvasa deferentia. These open ventrally on theeighth thoracomere (Balss, 1944), the fifth and lastwalking leg segment, by ejaculatory ducts thatopen through penes.
In ‘‘Podotremata’’ and Heterotremata, penes withterminal gonopores are located on the coxae of thefifth pereiopods. In pinnotherids, as in all Thoraco-tremata, the gonopores open on the correspondingsternites. The elongated gonopores (‘‘penes’’) trans-fer the sperm mass to the actual external male cop-ulatory structures, i.e., gonopods. These gonopodsevolved from the first two pairs of abdominal limbs(pleopods), modified for spermatozoal transfer dur-ing copulation. The first gonopod (G1) and the sec-ond gonopod (G2) of each body half form, togetherwith the corresponding penis, a functional unitachieving the transfer of the sperm mass. By con-ducting experiments with excised gonopods, Ryan(1967) demonstrated their essential role in spermtransfer in Portunus sanguinolentus.
Brocchi (1875) established comparative gonopodmorphology with regard to decapod systematics.He included gonopods, penes, and inner reproduc-tive structures in his account. In recent taxonomicstudies, mostly G1s are described. Their diversityin form and fine structure is great among Brachyura(see Shen, 1932; Stephensen, 1946; Guinot, 1966–1971, 1976, 1979; Martin and Abele, 1986).
The gonopods of Brachyura and other decapodgroups, such as Astacidea Latreille, 1802, comprisea proximal short protopodite articulated to thepleon and an elongated endopodite (Beninger et al.,
*Correspondence to: Carola Becker, Senckenberg Forschungsinsti-tut und Naturmuseum, Senckenberganlage 25, D-60325 Frankfurt/Main, Germany. E-mail: [email protected]
Received 26 January 2011; Revised 12 June 2012;Accepted 21 June 2012
Published online 29 August 2012 inWiley Online Library (wileyonlinelibrary.com)DOI: 10.1002/jmor.20065
JOURNAL OF MORPHOLOGY 273:1306–1318 (2012)
� 2012 WILEY PERIODICALS, INC.

1991; Minagawa, 1993; Tsuchida and Fujikura,2000). The protopodite consists of coxa and basis(Balss, 1944). The presumed plesiomorphic charac-ter state is found in the Norway lobster Nephropsnorvegicus, whose endopodite of the G1 forms alongitudinal groove. In interaction, the G2 comple-ments the grooved G1 to form a tube that carriesthe sperm mass. An ‘‘appendix masculina’’ (sensuBalss, 1944) is present in the endopodite of the G2,which functions in sealing the tube (Guinot, 1979).
Among the Brachyura, the grooved G1 is rolledup longitudinally to form a tube, the ejaculatorycanal, by an overlapping of the (formerly) lateralmargins of the groove. A suture is therefore pres-ent along the length of the G1, which can be moreor less closed according to the degree of tubulation.A proximal and a distal opening are always pres-ent in the tubular G1. In contrast, the G2s areusually not tubular. During copulation, the G2 isinserted into the G1. Length and function of theG2s vary considerably among crabs. A long G2,which protrudes from the distal opening of theejaculatory canal inside the G1, can directly trans-fer the sperm mass into the female genital ducts.In contrast, a short G2 does not come in contactwith the female ducts during copulation and there-fore only functions inside the G1. Both pairs ofgonopods are essential for sperm transfer, though,either the G1 or the G2 serves as the actual copu-latory organ inserted into the female gonopores.For instance, in the primitive copulatory system ofthe Podotremata, the G1 is a barely closed tube(Hartnoll, 1975). The proximal opening is (still)wide, the distal opening narrow. In the spongecrab, Dromia personata, the G1 is shorter than theG2. The endopodite of G2 is long, thin, and flexibleand is directly involved in sperm transfer (Hart-noll, 1975). Conversely, in the frog crab, Raninaranina, the G2 is shorter than the G1 and, conse-quently, works inside it (Minagawa, 1993, Hart-noll, 1979, Guinot, 1979). Actually, the distributionof character states among podotreme crabs is sodiverse (see Guinot, 1979) that phylogenetic con-clusions based on gonopod morphology are impos-sible, which reflects the current paraphyletic sta-tus of ‘‘Podotremata’’ Guinot, 1977 (Spears et al.,1992; Schram, 2001; Ahyong et al., 2007; Scholtzand McLay, 2009).
In eubrachyuran copulatory systems, two evolu-tionary trends can be observed: (1) the increasingtubulation of the G1 and (2) the shortening of theG2 (Hartnoll, 1975). The G2s of HetereotremataGuinot, 1977 are variable in length and have spe-cific function, whereas in Thoracotremata Guinot,1977, G2s are always clearly shorter than G1s.While numerous heterotreme copulatory systemshave been studied (Spalding, 1942; Cronin, 1947;Ryan, 1967; Diesel 1989; Beninger et al., 1991;Neumann, 1996; Brandis et al., 1999), only mar-ginal data on thoracotreme copulatory systems are
represented in the literature (Lautenschlageret al., 2010). In particular, the short G2 of theThoracotremata and its specific function in spermtransfer is not entirely understood.
In this study, we investigate the morphology ofthe pinnotherid copulatory system using light mi-croscopy, scanning electron microscopy (SEM), andconfocal laser scanning microscopy (CLSM). Basedon the results, we infer possible functions of thecopulatory structures and evaluate their use forsystematics.
MATERIAL AND METHODSSampling of Specimens
Pinnotheres pisum was collected from a population of thehorse mussel, Modiolus modiolus, in the Helgoland Trench(GPS 54808.4190N-54808.5990N to 07850.9210E-07853.4310E) dur-ing cruises to the German Bight with R. V. Senckenberg in2004 and 2005 by hard bottom dredge and beam trawl, thedepth from 50 to 55 m.
In the Northern Adriatic Sea (Rovinj/Croatia), P. pisum andNepinnotheres pinnotheres were collected from different hostsin 2005 and 2007. Partly hand-collected by scuba- and skin-div-ing in depths from 1 to 35 m, partly by beam trawl on tripswith R. V. Burin from the Institute Rud-er Boskovic on differentsample sites ranging from 45802N–45807N to 13836E–13840E.
Pinnotheres pectunculi was collected from the host Glycymerisglycymeris around Roscoff (Brittany/France) in 2007 and 2008.Additional material of the species P. pisum was obtained fromthe commercially traded host Mytilus edulis bought on fishmarkets in Germany from 2007–2009. The research compliedwith the institutional guidelines of animal ethics and adheredto the local legal requirements.
Scanning Electron Microscopy
The SEM-study was conducted at the Research InstituteSenckenberg (Frankfurt, Germany). Six specimens of P. pisum,two of P. pectunculi, and four of N. pinnotheres were used forSEM-investigations. The material was fixed in 96% ethanol andcleaned in an ultrasonic bath for 30 s to 2 min. Samples weredried on a ‘‘Balzor’s CPD 030 critical point dryer’’ and sputter-coated with a gold/palladium-composite on an ‘‘Edwards S 150B’’ sputter coater for 3 min (equivalent to a coating of 20 nmthickness). Samples were examined with a CamScan (Elektrone-noptik GmbH) scanning electron microscope type. Photographswere taken with Orion1 software. The description of setal typesfollows the nomenclature established by Garm (2004).
Histology
The histological work was carried out at the Research Insti-tute Senckenberg and at the Morphisto GmbH (both Frankfurt,Germany). Three specimens of P. pisum and two specimens ofN. pinnotheres were used for histology. Fresh tissue was fixedin ‘‘Susa Heidenhain’’ (Romeis, 1989) and embedded inparaffine. Histological sectioning was done with a Leitz 1515microtome at 8–10 lm. For general tissue differentiation, thetrichromatic Masson-Goldner stainings with ‘‘aniline blue’’ and‘‘light green’’ were used (after Romeis, 1989).
Semithin Sections
Semithin and ultrathin sections were prepared in the EM-laboratory of Goethe-University (Frankfurt/Germany) and inthe Zoological Institute of the University of Heidelberg (Ger-many). Twelve specimens were used in total: six of P. pisumand six of N. pinnotheres. Fresh tissue was fixed in 4% glutar-
MALE COPULATORY SYSTEM PINNOTHERIDAE 1307
Journal of Morphology
aldehyde in cacodylate-buffer (pH 7.4). The tissue was post-fixed with 1% osmium tetroxide for 2 h. Cacodylate and male-ate buffer (pH 5.2) washing steps were followed by en-blocstaining with 1% uranyl acetate overnight. After dehydrationthrough a graded series of ethanol, the tissue was infiltratedand embedded in Araldite or Spurr’s resin. Semithin sections(1–2 lm) were made with glass knifes on a Reichert-Jungultramicrotome (by Reichert-Jung) and stained with‘‘Richardson’s blue’’ (after Richardson et al., 1960). Photo-graphs of semithin sections and paraffin sections were takenwith a Leica Diaplan light microscope with CamScan1 cameraand ProgRes software.
Confocal Laser Scanning Microscopy (CLSM)
CLSMwas conducted at the Department of Cell Biology and Com-parative Zoology in the Institute of Biology at University of Copen-hagen (Denmark). Six specimens (two of each species) were used.
Samples were infiltrated and embedded in glycerine. Analyses weredone using a CLSM on a Leica DM IRBE microscope. A scan serieswas performed through whole objects. Single scans were combinedin maximum projections of single photographs with the help of LeicaTCS NT software.
RESULTSOverview
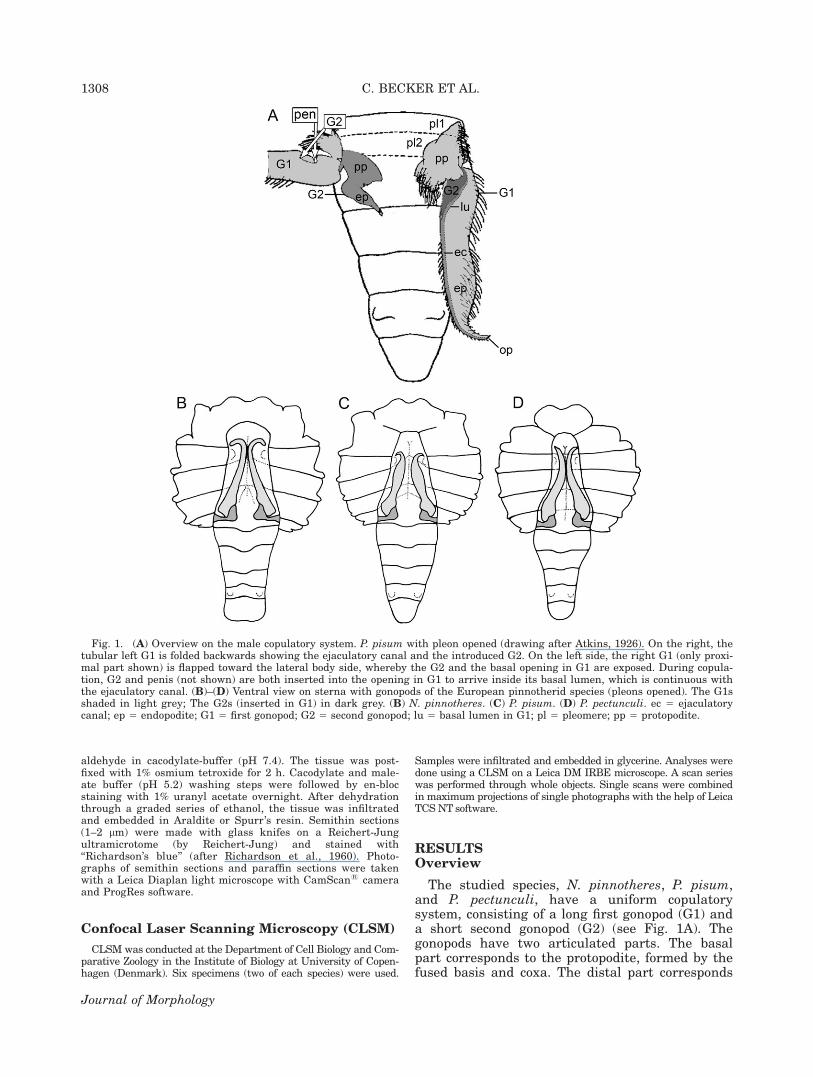
The studied species, N. pinnotheres, P. pisum,and P. pectunculi, have a uniform copulatorysystem, consisting of a long first gonopod (G1) anda short second gonopod (G2) (see Fig. 1A). Thegonopods have two articulated parts. The basalpart corresponds to the protopodite, formed by thefused basis and coxa. The distal part corresponds
Fig. 1. (A) Overview on the male copulatory system. P. pisum with pleon opened (drawing after Atkins, 1926). On the right, thetubular left G1 is folded backwards showing the ejaculatory canal and the introduced G2. On the left side, the right G1 (only proxi-mal part shown) is flapped toward the lateral body side, whereby the G2 and the basal opening in G1 are exposed. During copula-tion, G2 and penis (not shown) are both inserted into the opening in G1 to arrive inside its basal lumen, which is continuous withthe ejaculatory canal. (B)–(D) Ventral view on sterna with gonopods of the European pinnotherid species (pleons opened). The G1sshaded in light grey; The G2s (inserted in G1) in dark grey. (B) N. pinnotheres. (C) P. pisum. (D) P. pectunculi. ec 5 ejaculatorycanal; ep 5 endopodite; G1 5 first gonopod; G2 5 second gonopod; lu 5 basal lumen in G1; pl 5 pleomere; pp 5 protopodite.
1308 C. BECKER ET AL.
Journal of Morphology
to the endopodite. The endopodite of the G1 formsa tube with a wide basal opening and a narrowdistal opening (Fig. 1A). The endopodite of the G2is solid and coniform. The tube of the long firstgonopod (G1) transfers the sperm mass to thefemale ducts. The European species differ fromeach other in characters of the endopodite of G1and in the form of the pleon (Fig. 1B–D), which isnormally flexed into the sternal cavity to cover thegonopods.
First Gonopod (G1)
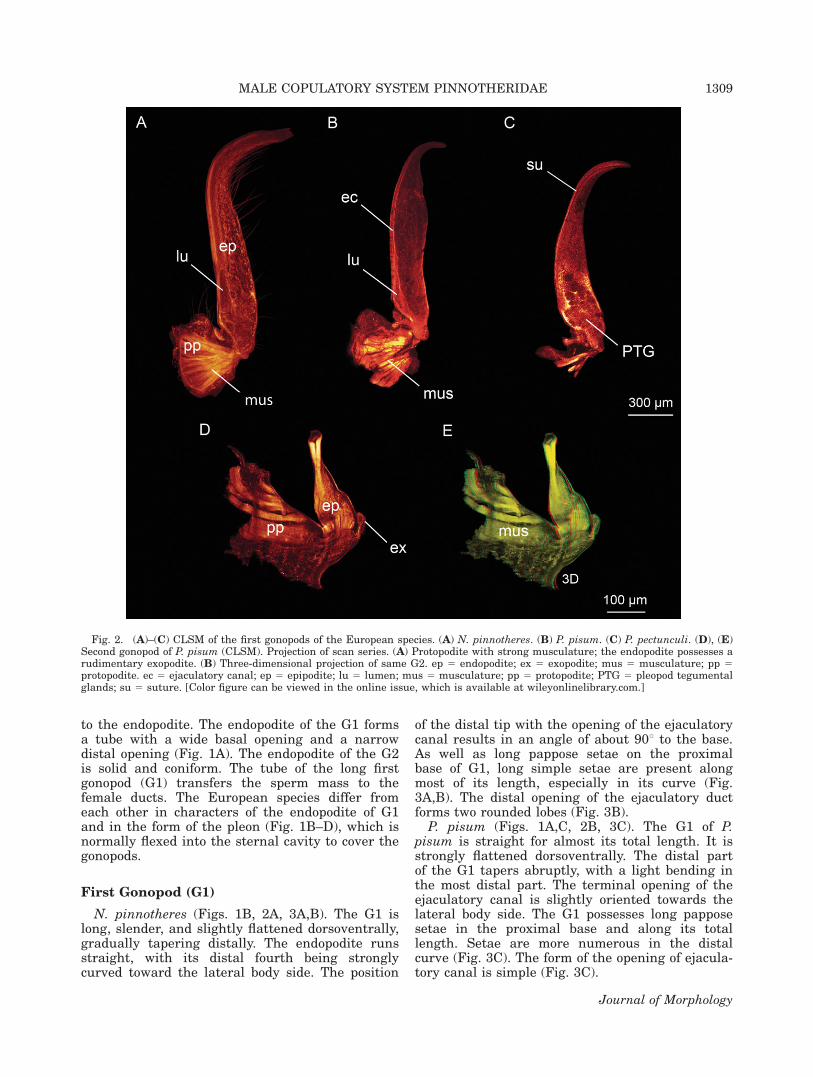
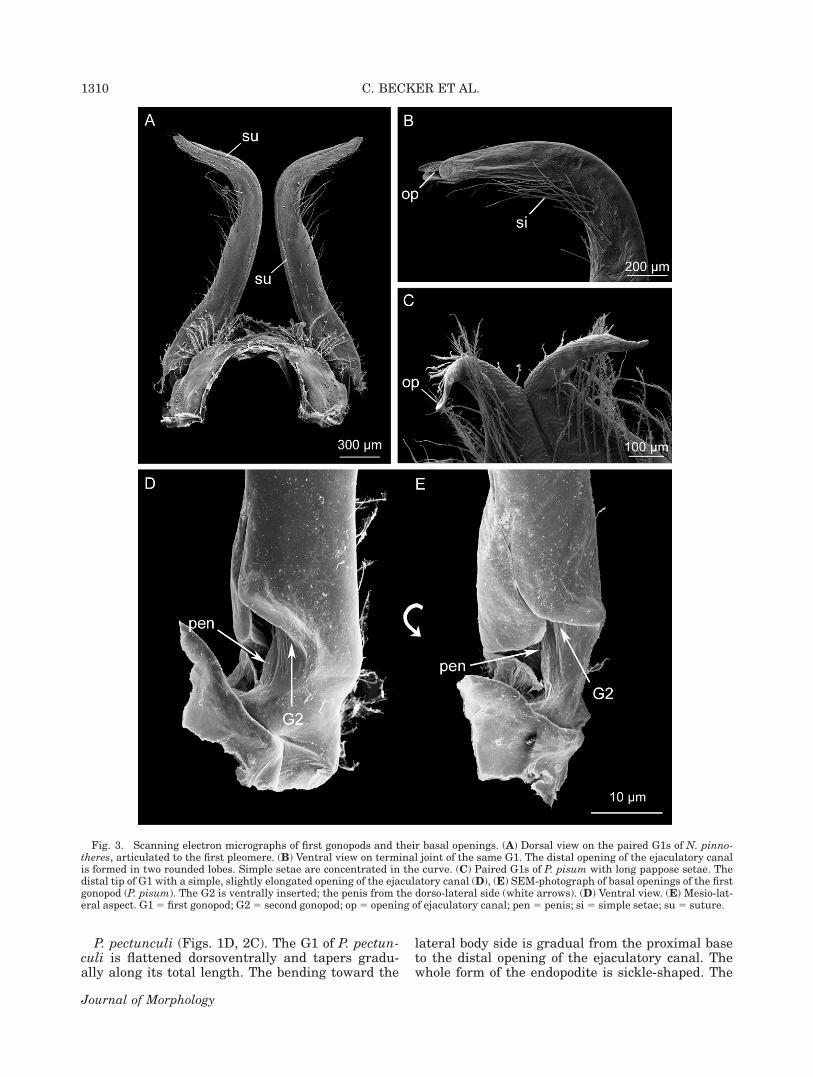
N. pinnotheres (Figs. 1B, 2A, 3A,B). The G1 islong, slender, and slightly flattened dorsoventrally,gradually tapering distally. The endopodite runsstraight, with its distal fourth being stronglycurved toward the lateral body side. The position
of the distal tip with the opening of the ejaculatorycanal results in an angle of about 908 to the base.As well as long pappose setae on the proximalbase of G1, long simple setae are present alongmost of its length, especially in its curve (Fig.3A,B). The distal opening of the ejaculatory ductforms two rounded lobes (Fig. 3B).
P. pisum (Figs. 1A,C, 2B, 3C). The G1 of P.pisum is straight for almost its total length. It isstrongly flattened dorsoventrally. The distal partof the G1 tapers abruptly, with a light bending inthe most distal part. The terminal opening of theejaculatory canal is slightly oriented towards thelateral body side. The G1 possesses long papposesetae in the proximal base and along its totallength. Setae are more numerous in the distalcurve (Fig. 3C). The form of the opening of ejacula-tory canal is simple (Fig. 3C).
Fig. 2. (A)–(C) CLSM of the first gonopods of the European species. (A) N. pinnotheres. (B) P. pisum. (C) P. pectunculi. (D), (E)Second gonopod of P. pisum (CLSM). Projection of scan series. (A) Protopodite with strong musculature; the endopodite possesses arudimentary exopodite. (B) Three-dimensional projection of same G2. ep 5 endopodite; ex 5 exopodite; mus 5 musculature; pp 5protopodite. ec 5 ejaculatory canal; ep 5 epipodite; lu 5 lumen; mus 5 musculature; pp 5 protopodite; PTG 5 pleopod tegumentalglands; su 5 suture. [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
MALE COPULATORY SYSTEM PINNOTHERIDAE 1309
Journal of Morphology
P. pectunculi (Figs. 1D, 2C). The G1 of P. pectun-culi is flattened dorsoventrally and tapers gradu-ally along its total length. The bending toward the
lateral body side is gradual from the proximal baseto the distal opening of the ejaculatory canal. Thewhole form of the endopodite is sickle-shaped. The
Fig. 3. Scanning electron micrographs of first gonopods and their basal openings. (A) Dorsal view on the paired G1s of N. pinno-theres, articulated to the first pleomere. (B) Ventral view on terminal joint of the same G1. The distal opening of the ejaculatory canalis formed in two rounded lobes. Simple setae are concentrated in the curve. (C) Paired G1s of P. pisum with long pappose setae. Thedistal tip of G1 with a simple, slightly elongated opening of the ejaculatory canal (D), (E) SEM-photograph of basal openings of the firstgonopod (P. pisum). The G2 is ventrally inserted; the penis from the dorso-lateral side (white arrows). (D) Ventral view. (E) Mesio-lat-eral aspect. G15 first gonopod; G25 second gonopod; op5 opening of ejaculatory canal; pen5 penis; si 5 simple setae; su5 suture.
1310 C. BECKER ET AL.
Journal of Morphology

pappose setae and the simple distal opening of theejaculatory duct are the same as in P. pisum (seeFig. 3C).
The basal lumen in the endopodite of the G1splits into two openings (Fig. 3D,E). The G2 isinserted from ventrally into a wide opening. Thepenis is inserted dorsolaterally through a narrowslit-like opening between the articulation of proto-podite and endopodite (Fig. 3D,E).
Second Gonopod (G2)
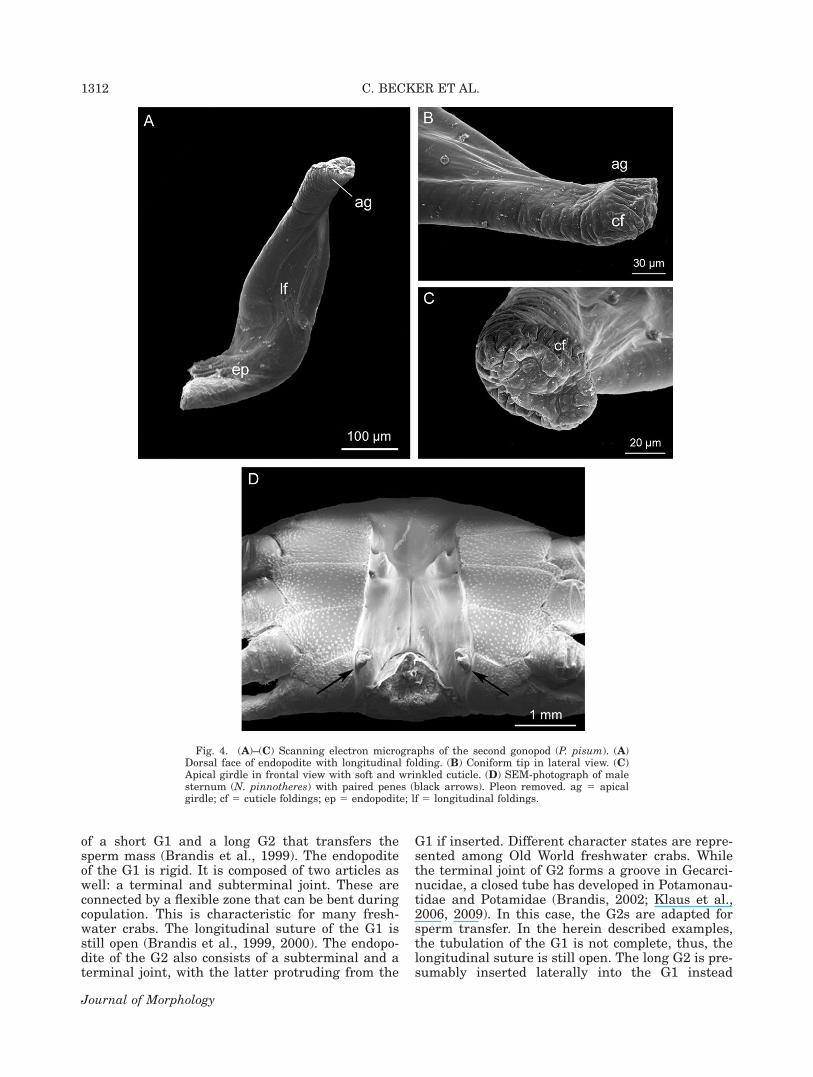
In the G2, protopodite and endopodite arealmost fused but still show a suture between them(Fig. 2D,E). The protopodite is robust and orientedmesially from the lateral edges of the second pleo-mere. The endopodite is turned anteriorly towardsthe basal opening in G1. A small finger-shapedexopodite is present at the ventromesial face of theG2 (Fig. 2D,E). The endopodite is solid and basallyslightly flattened. Distally, it has a coniform orstump-like form (Figs. 2D,E, 4A–C). The dorsaland ventral side of the G2 possess longitudinal cu-ticle foldings (Fig. 4A,B). The distal tip of the G2has a circular swelling, the ‘‘apical girdle’’ (sensuBeninger et al., 1991). The cuticle distal of the api-cal girdle is strongly folded and appears wrinkledin the SEM-photographs (Fig. 4A,C).
Ejaculatory Duct and Penis
In all Thoracotremata Guinot, 1977, the pinno-therids’ male gonopores open on the eighth thora-comere located on the slope of the sternal depres-sion (Fig. 4D).
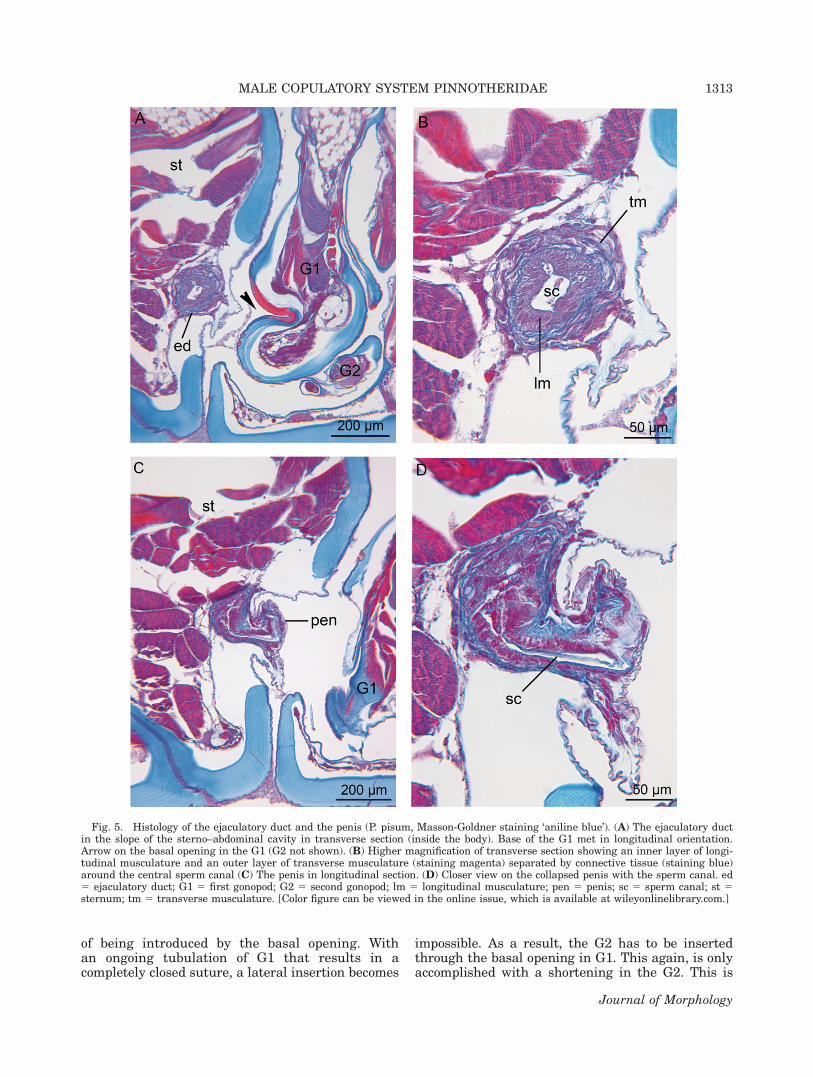
The distal part of the vas deferens is muscularand termed the ejaculatory duct. The penis can bedefined as the most distal part of the ejaculatoryduct that opens on the body’s outside and termi-nates by the gonopore. The paired penes areshown in Figure 4D. They are collapsed in SEM-observations (Fig. 4D) and in the histological sec-tions (Fig. 5C,D). The cuticle of the penis and thesurrounding integument is thin and wrinkled,indicating flexibility (Fig. 5A–D). The ejaculatoryduct (inside the body) is shown in transverse sec-tions (Fig. 5A,B), the penis (outside the body) inlongitudinal sections (Fig. 5C,D). An inner layer oflongitudinal musculature and outer layer of trans-verse musculature allow peristaltic actions of theejaculatory duct and penis (Fig. 5B). The collapsedpenis is short in fixed specimens and was neverobserved as being inserted into the G1. Duringcopulation, the penis is presumed to prolong toreach the opening in the G1.
Interaction of Gonopods and Penis
The histology of the G1 and the G2 is shown inFigure 6 and 7. The G2 was always found to be
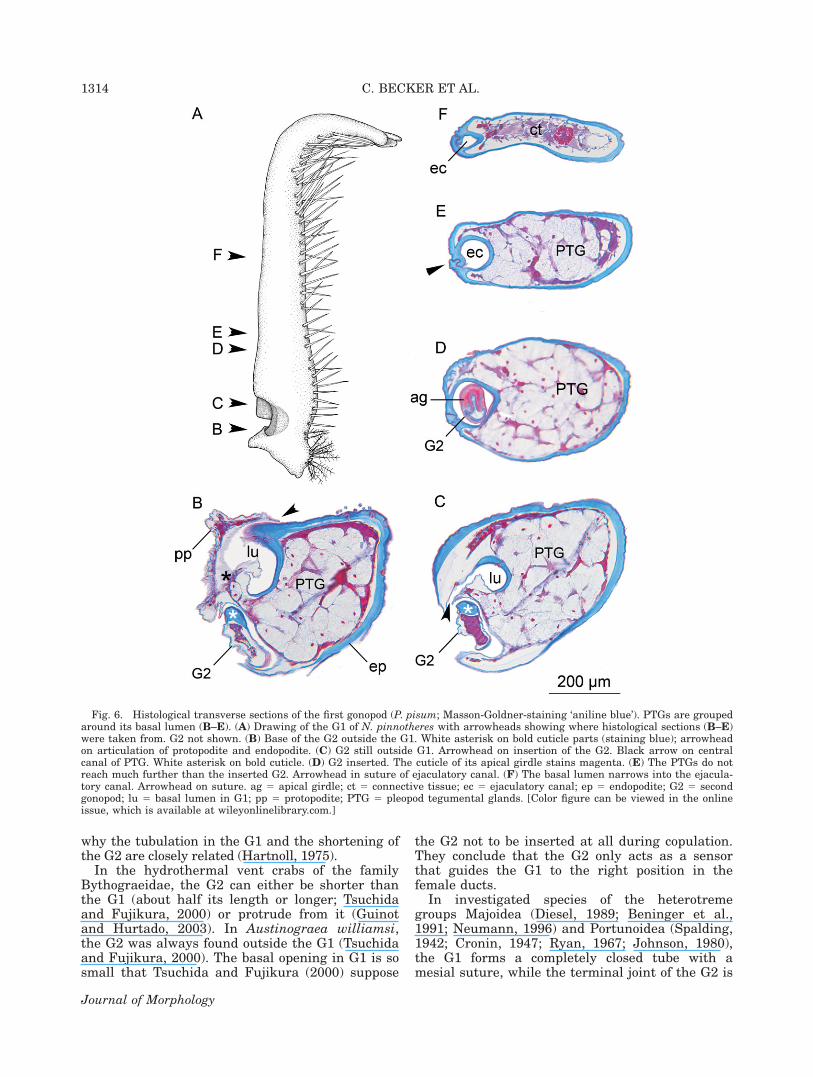
inserted in the G1 in living and fixed specimens.The base of the G1 is filled with rosette-shapedpleopod tegumental glands (PTG), which aregrouped around the basal lumen of the G1 (Figs.2C, 6B–D). Distally, they do not exceed furtherthan the inserted G2 (Figs. 2C, 6B–D). The coni-form tip of the G2 with the apical girdle is shownin Figure 6D. It dyes differently from the remain-ing cuticle in the Masson-Goldner staining. Thebasal lumen of the G1, which is continuous withthe ejaculatory canal, narrowing abruptly after theproximal third of the G1 (Figs. 2B, 6D, E).
The endopodite of the G2 is free of musculature.It possesses bold cuticle parts on the rounded lat-eral margins and thin cuticle on the ventral andespecially on the dorsal face (Figs. 6B–D, 7A–F).The form of the G2 follows the shape of the basallumen in the G1 precisely (Fig. 7B,C,E,F). Longi-tudinal cuticle foldings on the dorsal face of theG2 hook with the margin of the suture inside theG1 and, thus, arrest the G2 within the G1. Thelumen of the G1 where the G2 is introduced is sep-arated from the lumen, in which the penis isinserted, by a flexible flap with very thin cuticle.
DISCUSSIONReview on Brachyuran Gonopods
The copulatory system of the investigated pinno-therids, that is, a long first gonopod (G1) thattransfers the sperm mass and a reduced, solid sec-ond gonopod (G2), is characteristic for thoracot-reme crabs and is represented among a number ofhigher heterotremes as well.
In the ancestral heterotreme cancrids Cancerpagurus (Williamson, 1900) and Cancer gracilis(Orensanz et al., 1995), a long G2 acts in transfer-ring the sperm mass into the female gonopores.Their G2 is slender and flexible and its endopoditeconsists of two articles: a subterminal and a termi-nal joint (Williamson, 1900, Orensanz et al., 1995).The G1 is comparatively short and robust with anopen suture throughout most of its length. Whenthe G2 is inserted, its terminal joint protrudesfrom the G1. Williamson (1900) observed the rigidG1 to be held firmly during copulation, while theflexible G2 can freely move back and forwardinside the G1. By staining the sperm mass, Wil-liamson (1900) showed that the pumping of the G2inside the G1 forwards the sperm mass toward thedistal opening of the ejaculatory canal. As the G2is not grooved in the mentioned Cancer-species, itremains unclear how the elongated part of the G2is involved in sperm transfer and in the accurateplacement into the female duct. Therefore, theprocess of sperm transfer among Cancridae issupposed to be a rather unspecific kind of plugging(Elner et al., 1985, Williamson, 1900).
Heterotreme freshwater crabs of the genus Pota-mon also have a copulatory system, which consists
MALE COPULATORY SYSTEM PINNOTHERIDAE 1311
Journal of Morphology
of a short G1 and a long G2 that transfers thesperm mass (Brandis et al., 1999). The endopoditeof the G1 is rigid. It is composed of two articles aswell: a terminal and subterminal joint. These areconnected by a flexible zone that can be bent duringcopulation. This is characteristic for many fresh-water crabs. The longitudinal suture of the G1 isstill open (Brandis et al., 1999, 2000). The endopo-dite of the G2 also consists of a subterminal and aterminal joint, with the latter protruding from the
G1 if inserted. Different character states are repre-sented among Old World freshwater crabs. Whilethe terminal joint of G2 forms a groove in Gecarci-nucidae, a closed tube has developed in Potamonau-tidae and Potamidae (Brandis, 2002; Klaus et al.,2006, 2009). In this case, the G2s are adapted forsperm transfer. In the herein described examples,the tubulation of the G1 is not complete, thus, thelongitudinal suture is still open. The long G2 is pre-sumably inserted laterally into the G1 instead
Fig. 4. (A)–(C) Scanning electron micrographs of the second gonopod (P. pisum). (A)Dorsal face of endopodite with longitudinal folding. (B) Coniform tip in lateral view. (C)Apical girdle in frontal view with soft and wrinkled cuticle. (D) SEM-photograph of malesternum (N. pinnotheres) with paired penes (black arrows). Pleon removed. ag 5 apicalgirdle; cf 5 cuticle foldings; ep 5 endopodite; lf 5 longitudinal foldings.
1312 C. BECKER ET AL.
Journal of Morphology
of being introduced by the basal opening. Withan ongoing tubulation of G1 that results in acompletely closed suture, a lateral insertion becomes
impossible. As a result, the G2 has to be insertedthrough the basal opening in G1. This again, is onlyaccomplished with a shortening in the G2. This is
Fig. 5. Histology of the ejaculatory duct and the penis (P. pisum, Masson-Goldner staining ‘aniline blue’). (A) The ejaculatory ductin the slope of the sterno–abdominal cavity in transverse section (inside the body). Base of the G1 met in longitudinal orientation.Arrow on the basal opening in the G1 (G2 not shown). (B) Higher magnification of transverse section showing an inner layer of longi-tudinal musculature and an outer layer of transverse musculature (staining magenta) separated by connective tissue (staining blue)around the central sperm canal (C) The penis in longitudinal section. (D) Closer view on the collapsed penis with the sperm canal. ed5 ejaculatory duct; G1 5 first gonopod; G2 5 second gonopod; lm 5 longitudinal musculature; pen 5 penis; sc 5 sperm canal; st 5sternum; tm 5 transverse musculature. [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
MALE COPULATORY SYSTEM PINNOTHERIDAE 1313
Journal of Morphology
why the tubulation in the G1 and the shortening ofthe G2 are closely related (Hartnoll, 1975).
In the hydrothermal vent crabs of the familyBythograeidae, the G2 can either be shorter thanthe G1 (about half its length or longer; Tsuchidaand Fujikura, 2000) or protrude from it (Guinotand Hurtado, 2003). In Austinograea williamsi,the G2 was always found outside the G1 (Tsuchidaand Fujikura, 2000). The basal opening in G1 is sosmall that Tsuchida and Fujikura (2000) suppose
the G2 not to be inserted at all during copulation.They conclude that the G2 only acts as a sensorthat guides the G1 to the right position in thefemale ducts.
In investigated species of the heterotremegroups Majoidea (Diesel, 1989; Beninger et al.,1991; Neumann, 1996) and Portunoidea (Spalding,1942; Cronin, 1947; Ryan, 1967; Johnson, 1980),the G1 forms a completely closed tube with amesial suture, while the terminal joint of the G2 is
Fig. 6. Histological transverse sections of the first gonopod (P. pisum; Masson-Goldner-staining ‘aniline blue’). PTGs are groupedaround its basal lumen (B–E). (A) Drawing of the G1 of N. pinnotheres with arrowheads showing where histological sections (B–E)were taken from. G2 not shown. (B) Base of the G2 outside the G1. White asterisk on bold cuticle parts (staining blue); arrowheadon articulation of protopodite and endopodite. (C) G2 still outside G1. Arrowhead on insertion of the G2. Black arrow on centralcanal of PTG. White asterisk on bold cuticle. (D) G2 inserted. The cuticle of its apical girdle stains magenta. (E) The PTGs do notreach much further than the inserted G2. Arrowhead in suture of ejaculatory canal. (F) The basal lumen narrows into the ejacula-tory canal. Arrowhead on suture. ag 5 apical girdle; ct 5 connective tissue; ec 5 ejaculatory canal; ep 5 endopodite; G2 5 secondgonopod; lu 5 basal lumen in G1; pp 5 protopodite; PTG 5 pleopod tegumental glands. [Color figure can be viewed in the onlineissue, which is available at wileyonlinelibrary.com.]
1314 C. BECKER ET AL.
Journal of Morphology
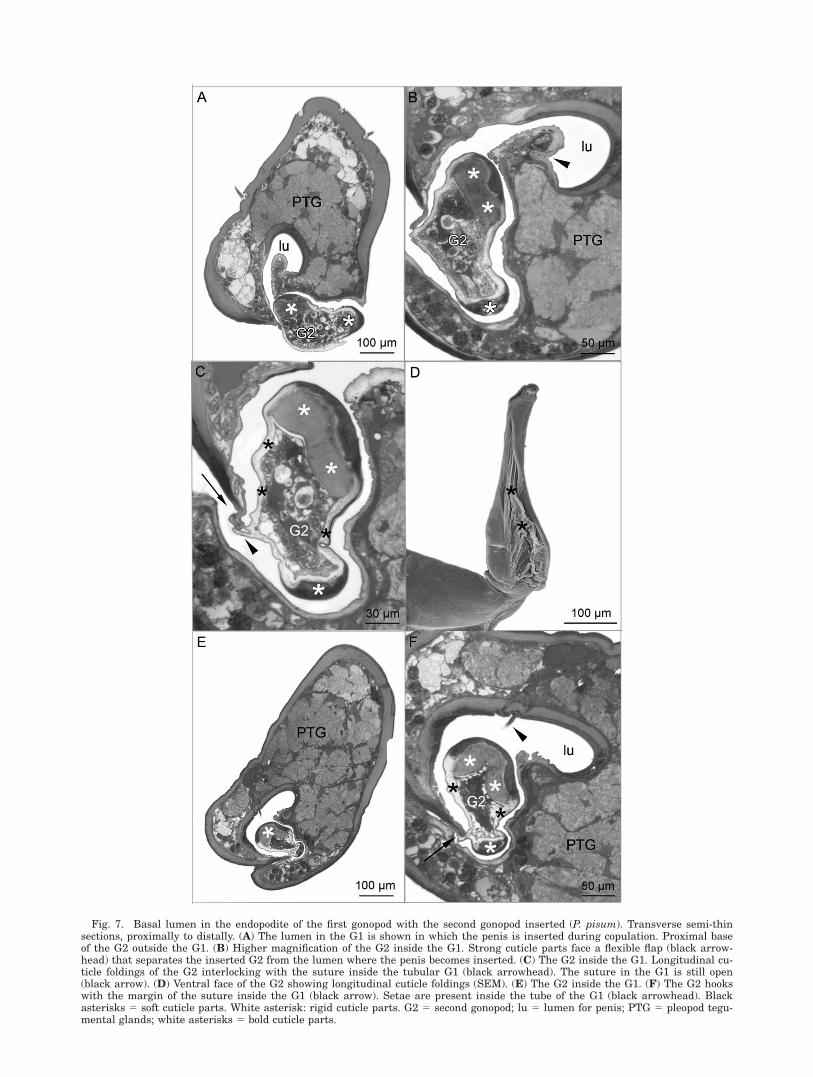
Fig. 7. Basal lumen in the endopodite of the first gonopod with the second gonopod inserted (P. pisum). Transverse semi-thinsections, proximally to distally. (A) The lumen in the G1 is shown in which the penis is inserted during copulation. Proximal baseof the G2 outside the G1. (B) Higher magnification of the G2 inside the G1. Strong cuticle parts face a flexible flap (black arrow-head) that separates the inserted G2 from the lumen where the penis becomes inserted. (C) The G2 inside the G1. Longitudinal cu-ticle foldings of the G2 interlocking with the suture inside the tubular G1 (black arrowhead). The suture in the G1 is still open(black arrow). (D) Ventral face of the G2 showing longitudinal cuticle foldings (SEM). (E) The G2 inside the G1. (F) The G2 hookswith the margin of the suture inside the G1 (black arrow). Setae are present inside the tube of the G1 (black arrowhead). Blackasterisks 5 soft cuticle parts. White asterisk: rigid cuticle parts. G2 5 second gonopod; lu 5 lumen for penis; PTG 5 pleopod tegu-mental glands; white asterisks 5 bold cuticle parts.

shortened. As in the shore crab Carcinus maenas,the length of the G2 is one-third of the G1(Spalding, 1942), while it is about one-sixth in thespider crabs Maja spp. (Neumann, 1996).
The role of a short G2 in sperm transfer is seenas a plunger or piston that accomplishes pumpingmovements inside the G1 and thereby drives thesperm mass within the ejaculatory canal to its dis-tal opening (Ryan, 1967; Bauer, 1986; Diesel, 1989;Beninger et al., 1991). The pumping movement isachieved by a flexing of the pleon during copula-tion (Watson, 1970, 1972; Diesel, 1989; Elner andBeninger, 1992). Even though the endopodite isfree of musculature, the G2 can also move alongits transverse axis by contracting the musculaturein its protopodite.
For the majoid, Chionoecetes opilio, severalstructures were described distally in the G2: the‘‘appendix masculina,’’ a ‘‘protuberance,’’ and an‘apical girdle’ (sensu Beninger et al., 1991). Neu-mann (1996) followed this terminology to describethe G2s of Maja spp. The term ‘‘appendix mascu-lina’’ is deduced from pleopod features in maleshrimps (see Balss, 1944). In our opinion, the useof the term ‘‘appendix masculina’’ on processes ofbrachyuran gonopods implies a homology of theseappendices, which is questionable.
The Pinnotherid Copulatory System
In the investigated pinnotherids, the G2 is reducedin length and appendices or protuberances are nolonger present. However, we found the coniform tipof the G2 of the studied pinnotherids to form an api-cal girdle that conforms to the description byBeninger et al. (1991). The apical girdle is character-ized by a circular cuticle swelling around the tip ofthe G2, distally followed by folded cuticle. The finestructure and histology of the endopodite of the G2and especially of its apical girdle suggest a certainability to swell. Due to the lack of musculature in theendopodite of the G2, a modification in form can onlyoccur by the build up of hemolymph pressure. Thewhole endopodite of the G2 possesses bold rigid cuti-cle parts laterally and flexible, folded cuticle on theslightly flattened ventral and dorsal face. With ahemolymph swelling, the G2 is supposed to enlargealong the cuticle foldings. The ability to swell in theendopodite with its apical girdle and the precise ad-aptation of the specific shape of the G2 to the tubeinside the G1 show that the G2 is really optimized totightly seal the basal lumen and, respectively, theejaculatory canal, inside the G1. Furthermore, theG2 seals the tubular system to the outside, whichcould minimize seawater influx and the loss of sper-matozoa. Observations on mating behavior of C. opi-lio revealed the sperm transfer process to be kind of‘‘leaky’’, which implies a partial loss of the spermmass (Watson 1970, 1972). Beninger et al. (1988)stated that the transmission process of spermatozoa
is sensitive in Brachyura because of the risk for sper-matophores to come in contact with sea water duringcopulation, which infiltrates through the proximalopenings in the G1 causing their dissolution. Fur-thermore, Beninger et al. (1991) supposed that asealing of the ejaculatory canal in the G1 by the G2is a precondition to enable the hydraulic pumping ofseminal fluids inside the ejaculatory canal. Beningeret al. (1988) also observed an asymmetry in cuticlethickness in the G2 of C. opilio and supposed that itfunctions in breaking the sealing of the ejaculatorycanal with every backwards movement due to theasymmetrical forces.
The G2 of the investigated pinnotherids appearsto be adapted for functioning in hydraulic trans-port. We suggest that the G2 swells along its lat-eral faces and in the apical girdle with upwardmovements to tightly seal the ejaculatory canaland, respectively, the basal lumen (see Fig. 7).Thus, hydraulic pressure is built up, which drivesthe sperm mass distally. With backwards move-ments, the hydraulic pressure is released by theG2’s endopodite collapsing in its flexible parts. Inaddition to this, the G2 of the studied pinnotheridswas observed to interlock with the internal sutureof the G1. This clearly improves the sealing of thebasal opening and the internal suture of the G1 to-ward the outside. Again, the interlocking mightalso function in keeping the G2 in position whilemoving, by acting like a guide rail.
Furthermore, the G2 is supposed to interactwith the penis and with the lumen where thepenis becomes inserted. The basal lumen in the G1is split into two cavities for the G2 and the penis.The lumina where G2 and penis are inserted dur-ing copulation are only separated by a flexible flap.If the G2 moves inside the G1, it presumablypresses with its bold cuticle face against that flapwhereby the lumen wherein the penis lies becomesconstricted. This might additionally contribute tothe sealing of the hydraulic system, but it is alsopossible that the flexible flap directly interacts withthe penis by pressing against it. Williamson (1900)observed such an interaction of the G2 and thepenis in Cancer pagurus: the G2 pressed againstthe penis with every movement whereby the spermmass is conveyed into the tube of the G1.
Pleopod Tegumental Glands
Rosette-shaped ‘‘pleopod tegumental glands’’ (PTG)have so far been found inside the first gonopods(G1s) of all investigated brachyuran crabs(Spalding, 1942; Diesel, 1989; Beninger and Laroc-que, 1998; Brandis et al., 1999). The PTGs areultrastructurally similar to functionally diversetegumental glands of other decapod crustaceans(Johnson and Talbot, 1987; Schmidt et al., 2006).They are generally composed of a central cell,numerous secretory cells arranged in a rosette,and one canal cell that leads through cuticular
1316 C. BECKER ET AL.
Journal of Morphology

pores (Talbot and Demers, 1993). The PTGs havebeen demonstrated to communicate with the ejacu-latory canal via ducts passing through cuticularpores and concluded that their secretions areinvolved in the process of sperm transfer (Spalding,1942; Beninger and Larocque, 1998). As in the stud-ied pinnotherids, the PTGs are generally groupedaround the basal lumen of the G1 and extend dis-tally as far as the inserted G2 (Spalding, 1942; Die-sel, 1989; Beninger and Larocque, 1998; Brandiset al., 1999). The function of the PTGs in the G1 ofthe Brachyura is still discussed. Spalding (1942)suggested that their secretions form the spermato-phores, which is presently clearly disproved,because completed spermatophores are alreadyfound in the vas deferens (Adiyodi and Anilkumar,1988; Beninger et al., 1988; Diesel, 1989). A preva-lent idea is that secretions of the PTGs contributeto the formation of the so-called ‘‘sperm plug,’’astructure of hardened secretions often found to plugthe female ducts after copulation (Williamson, 1900;Spalding, 1942; Cronin, 1947; Ryan, 1967; Bawaband El-Sherief, 1989). The sperm plug was sup-posed to prevent the loss of spermatozoa or theentry of sea water in impregnated females (Wil-liamson, 1900), but it also plays a role in spermcompetition by closing the female’s genital openingsto inhibit subsequent copulations. In the spidercrab Inachus phalangium, the sperm received fromseveral matings are separated by layers of spermplugs inside the spermatheca, which suggests thatmales from consecutive copulations seal off rivalsperm masses (Diesel, 1990; 1991).
However, in the investigated pinnotherid spe-cies, sperm plugs were not present in the femalegenital ducts (Becker et al., 2011), which is alsonot the case in a number of other crabs that stillhave the PTGs in their G1s. Johnson and Talbot(1987) identified at least two different types of se-cretory cells in the PTGs. Beninger and Larocque(1998) showed by biochemical assays that the com-position of the substances secreted by the PTGsvaries between species. They concluded that onlysome might contribute to the sperm plug whileothers function in the protection of spermato-phores from opportunistic microbes. Furthermore,secretions may function directly in the transport ofthe sperm inside the G1. Beninger and Larocque(1998) proposed that they might act as a lubricantto reduce mechanical wear of the ejaculatory canalby G2 or by reducing the viscosity of the ejaculateas it enters the narrow ejaculatory canal. Thesecretions of the PTGs may also help in buildingup pressure inside the tubular system of G1,which is necessary for the transport of spermato-zoa. They may also just contribute to the seminalplasma or somehow function inside the femalerecepataculum seminis. But the spermatophoresare already embedded in seminal plasma producedby the vas deferens and the PTGs secrete directly
into the ejaculatory canal and the basal lumen inthe G2. That is why we rather suppose its functionin the process of transmitting sperm.
Our results show that the G2 of pinnotherids isreduced in size indeed, but still essential for thefunction of the copulatory system in sperm trans-fer, because the G2 is precisely adapted to the ba-sal lumen of the G1, it specifically interacts with.In the future, G2s should get more attention, notonly in terms of comparative morphology but alsoconsidering the functional aspects and the spe-cific role of the G2 during copulation and spermtransfer.
ACKNOWLEDGMENTS
We thank Michael Gudo (Morphisto GmbH,Frankfurt, Germany) for his expertise in histology.We also acknowledge Claudia Kempendorf (Uni-versity Heidelberg, Germany), Volker Knauff (Mor-phisto, Frankfurt, Germany), Marion Stohr, andSusann Munzner (University Frankfurt, Germany)for their kind help in preparing techniques for his-tology and transmission electron microscopy. Wealso thank Sebastian Klaus and Chris Tudge forreviewing the manuscript. For the CLSM-study,the authors acknowledge Synthesys (grant DK-TAF-4264) and Jens T. Hoeg and Andreas Wan-ninger (University of Copenhagen, Denmark).
LITERATURE CITED
Adiyodi KG, Anilkumar G. 1988. Arthropoda-Crustacea. In:Adiyodi KG, Adiyodi RG, editors. Reproductive Biology ofInvertebrates: accessory sex glands, Vol. 3. Chichester, Eng-land: Wiley. pp 261–318.
Ahyong ST, Lai JCY, Sharkey D, Colgan DJ, Ng PKL. 2007.Phylogenetics of the brachyuran crabs (Crustacea: Decapoda):The status of Podotremata based on small subunit nuclearribosomal RNA. Mol Phylogenet Evol 45:576–586.
Atkins D. 1926. The moulting stages of the pea-crab (Pinno-theres pisum). J Mar Biol Assoc UK. 14:475–492.
Balss H. 1944. Morphologischer teil. In: Schellenberg A,Grunder HE, editors. Dr HG Bronns Klassen und Ordnungendes Tierreichs: Decapoda. Leipzig, Germany: AkademischeVerlagsgesellschaft Geest & Portig KG. pp 22–667.
Bauer RT. 1986. Phylogenetic trends in sperm transfer andstorage complexity in decapod crustaceans. J Crustacean Biol6:313–325.
Bawab FM, El-Sherief SS. 1989. Contributions to the study ofthe origin, nature and formation of the plug in the sperma-theca of the female crab Portunus pelagicus (Linnaeus, 1766)(Decapoda, Brachyura). Crustaceana 57:9–24.
Becker C, Brandis D, Storch V. 2011. Morphology of the femalereproductive system of European pea crabs. (Crustacea: Bra-chyura: Pinnotheridae). J Morphol 272:12–26.
Beninger PG, Larocque R. 1998. Gonopod tegumental glands: Anew accessory sex gland in the Brachyura. Mar Biol 132:435–444.
Beninger PG, Elner RW, Foyle TP, Odense PH. 1988. Functionalanatomy of the male reproductive system and the femalespermatheca in the snow crab Chionoecetes opilio (O. Fabri-cius) (Decapoda: Majidae) and a hypothesis for fertilization. JCrustacean Biol 8:322–332.
Beninger PG, Elner RW, Poussart Y. 1991. Gonopods of themajid crab Chionoecetes opilio (O. Fabricius). J CrustaceanBiol 11:217–228.
MALE COPULATORY SYSTEM PINNOTHERIDAE 1317
Journal of Morphology

Brandis D, Storch V, Turkay M. 1999. Morphology and functionof the copulatory system in freshwater crabs of the GenusPotamon. J Morphol 239:157–166.
Brandis D, Storch V, Turkay M. 2000. Taxonomy and zoogeogra-phy of the freshwater crabs of Europe, North Africa, and theMiddle East (Crustacea, Decapoda, Potamidae). Sencken-bergiana Biol 80:5–56.
Brandis D. 2002. Taxonomy and biogeography of the family Iso-lapotamidae. J Nat Hist 36:1291–1339.
Brocchi M. 1875. Recherches sur les organes genitaux malesdes Crustaces Decapodes. Ann Sci Nat (Zool) Ser 6:1–131.
Cronin LE. 1947. Anatomy and physiology of the male repro-ductive system of Callinectes sapidus Rathbun. J Morphol81:209–239.
Diesel R. 1989. Structure and function of the reproductive sys-tem of the symbiotic spider crab Inachus phalangium (Decap-oda: Majidae): Observations on sperm transfer, sperm storage,and spawning. J Crustacean Biol 9:266–277.
Diesel R. 1990. Sperm competition and reproductive success inthe decapod Inachus phalangium (Majidae): A male ghost spi-der crab that seals off rivals’ sperm. J Zool (Lond) 220:213–223.
Diesel R. 1991. Sperm competition and the evolution of matingbehaviour in Brachyura, with special reference to spidercrabs (Decapoda, Majidae). In: Bauer RT, Martin JW, editors.Crustacean Sexual Biology. New York: Columbia UniversityPress. pp 145–163.
Elner RW, Beninger PG. 1992. The reproductive biology of snowcrab, Chionoecetes opilio: A synthesis of recent contributions.Am Zool 32:524–533.
Elner RW, Gass CA, Campbell A. 1985. Mating behaviour of thejonah crab, Cancer borealis Stimpson (Decapoda, Brachyura).Crustaceana 14:34–39.
Garm A 2004. Revising the definition of the crustacean setaand setal classification systems based on examinations of themouthpart setae of seven species of decapods. Zool J Linn Soc142:233–252.
Guinot D. 1966–1971. Recherches preliminaires sur les groupesnaturels chez les Crustaces Decapodes Brachyoures. I–VIII.Bull Mus Natl Hist Nat Ser 2 38:744–1090.
Guinot D. 1976. Constitution de quelques groupes naturels chezles Crustaces Decapodes Brachyoures. I. La superfamille desBellioidea et trois sous-familles de Xanthidae (PolydectinidaeDana, Trichiinae de Haan, Actaeinae Alcock). Mem Mus NatlHist Nat Ser A 97:1–308.
Guinot D. 1979. Donnee nouvelle sur la morphologie, la phylogen-ese et la taxonomie des Crustaces Decapodes Brachyoures: VIIPleopodes sexuels males. Mus Natl Hist Nat Ser A 112:1–354.
Guinot D, Hurtado LA. 2003. Two new species of hydrothermalvent crabs of the genus Bythograea from the southern EastPacific Rise and from the Galapagos Rift (Crustacea Decap-oda Brachyura Bythograeidae). C R Biol 326:423–439.
Guinot D, Quenette G. 2005. The spermatheca in podotremecrabs (Crustacea, Decapoda, Brachyura, Podotremata) and itsphylogenetic implications. Zoosystema 27:267–342.
Hartnoll RG. 1969. Mating in the Brachyura. Crustaceana16:160–181.
Hartnoll RG. 1975. Copulatory structure and function in theDromiacea, and their bearing on the evolution of Brachyura.Pubbl Staz Zool Napoli 39Suppl:657–676.
Hartnoll RG. 1979. The phyletic implications of spermathecalstructure in the Raninidae (Decapoda: Brachyura). J Zool(Lond) 187:75–83.
Johnson PT. 1980. Histology of the blue crab, Callinectes sap-idus. A Model for the Decapoda: The Reproductive System,Chapter 15. New York: Praeger Publishers. pp 327–367.
Johnson B, Talbot P. 1987. Ultrastructural analysis of the pleo-pod tegumental glands in male and female lobsters, Homarusamericanus. J Crustacean Biol 7:288–301.
Klaus S, Schubart CD, Brandis D. 2006. Phylogeny, biogeogra-phy and a new taxonomy for the Gecarcinucoidea Rathbun,1904 (Decapoda: Brachyura). Organ Divers Evol 6:199–217.
Klaus S, Brandis D, Ng PKL, Yeo DCJ, Schubart CD. 2009.Phylogeny and biogeography of Asian freshwater crabs of thefamily Gecarcinucidae (Brachyura: Potamoidea). In: MartinJW, Crandall KA, Felder DL, editors. Crust Issues 18:Decapod Crustacean Phylogenetics. Boca Raton, Florida:Taylor & Francis/CRC Press. pp 509–531.
Lautenschlager A, Brandis D, Storch V. 2010. Morphology andfunction of the reproductive system of representatives of thegenus Uca. J Morphol 271:1281–1299.
Minagawa M. 1993. Gonopods of the red frog crab Ranina raninaLinnaeus (Decapoda: Raninidae). Crustacean Res 22:45–54.
Martin JW, Abele LG. 1986. Notes on male pleopod morphologyin the brachyuran crab family Panopeidae Ortmann, 1893,sensu Guinot (1978) (Decapoda). Crustaceana 50:182–198.
NeumannV. 1996. Comparative gonopodmorphology of the Europeanspider crabs of the genus Maja Lamarck 1801 (Crustacea: Decap-oda: Brachyura:Majidae). Senckenbergiana biol 75:143–157.
Orensanz JM, Parma AM, Armstrong DA, Armstrong J,Wardrup P. 1995. Feeding ecology of Cancer gracilis (Crus-tacea: Decapoda: Cancridae) and the mating systems ofcancrid crabs. J Zool 235:411–437.
Richardson KC, Jarett L, Finke EH. 1960. Embedding in epoxyresins for ultrathin sectioning in electron microscopy. StainTechnol 35:313–323.
Romeis B. 1989. Mikroskopische Technik. 17th edition. Munchen:Urban & Schwarzenberg.697 p.
Ryan EP. 1967. Structure and function of the reproductive sys-tem of the crab Portunus sanguinolentus (Herbst) (Brachyura:Portunidae). 1. The male system. In: Proceedings of the Sym-posium on Crustacea, 12–15 January, 1965, Ernakulam,India. Marine Biological Association of India, SymposiumSeries, Bangalore Press, Bangalore, India, 2:506–521.
Schmidt M, Chien H, Tadesse T, Johns ME, Derby CD. 2006.Rosette-type tegumental glands associated with aesthetascsensilla in the olfactory organ of the Caribbean spiny lobster,Palinurus argus. Cell Tissue Res 325:369–395.
Scholtz G, McLay CL. 2009. Is the Brachyura Podotremata amonophyletic group? In: Martin JW, Crandall KA, Felder DL,editors. Crust Issues 18: Decapod Crustacean Phylogenetics.Boca Raton, Florida: Taylor & Francis/CRC Press. pp 417–437.
Schram FR. 2001. Phylogeny of decapods: Moving towards aconsensus. In: Flores AAV, Fransen CHJM, editors. Nether-lands: Kluwer Academic Publishers. Hydrobiol 449:1–20.
Shen CJ. 1932. The brachyuran Crustacea of north China. ZoolSinica Ser A 9:84–92.
Spalding JF. 1942. The nature and formation of the spermato-phore and the sperm plug in Carcinus maenas. Q J MicroscSci. 83:399–422.
Spears T, Abele LG, Kim W. 1992. The monophyly of bra-chyuran crabs: A phylogenetic study based on 18S rRNA.Syst Biol 41:446–461.
Stephensen K. 1946. The Brachyura of the Iranian Gulf. Withan appendix: The male pleopoda of the Brachyura. DanishScient Investig Iran 4:58–237.
Talbot P, Demers D. 1993. Tegumental glands of Crustacea. In:Horst MN, Freeeman JA, editors. The Crustacean Integu-ment: Morphology and Biochemistry. Boca Raton, USA: CRCPress, Taylor & Francis Group. pp 151–191.
Tsuchida S, Fujikura K. 2000. Heterochely, relative growth, andgonopod morphology in the bythograeid crab, Austinograeawilliamsi (Decapoda, Brachyura). J Crustacean Biol 20:199–206.
Turkay M. 1975. Statement: Die Bedeutung des Gonopodenauf-baus fur die Aufklarung von Verwandtschaftsverhaltnissenbei dekapoden Crustaceen. Aufs Red Senckenb Naturf Ges27:114–115.
Watson J. 1970. Maturity, mating and egg laying in the spidercrab, Chionoecetes opilio. J Fish Res Bd Canada 27:1607–1616.
Watson J. 1972. Mating behaviour in the spider crab, Chionoe-cetes opilio. J Fish Res Bd Canada 29:447–449.
Williamson HC. 1900. Contributions to the life-history of the ed-ible crab (Cancer pagurus, Linn). Rep Fish Board Scotl18:77–142.
1318 C. BECKER ET AL.
Journal of Morphology