respiration and circulation in land crabs - oxford academic
TRANSCRIPT

AMER. ZOOL., 32:417-^27 (1992)
Respiration and Circulation in Land Crabs:Novel Variations on the Marine Design1
WARREN W. BURGGREN
Department of Biological Sciences, University of Nevada, Las Vegas, Nevada 89154-4004
SYNOPSIS. Both the "true" crabs (Brachyura) and hermit crabs (Ano-mura) include species that show numerous behavioral, morphological,and physiological specializations permitting terrestrial life. This paperexamines respiratory and circulatory adaptations for air breathing in theseland crabs. Respiratory specializations include modification of gas exchangestructures for air breathing (gills and elaborated branchial chamberlinings), ventilatory mechanisms permitting effective air pumping, an ele-vated hemolymph oxygen capacity, and a primarily CO2- rather than O2-sensitive ventilatory control system. The qualitative aspects of hemo-lymph oxygen transport and metabolic rate are apparently unchangedfrom that of marine crabs. While the basic cardiovascular morphology ofland crabs appears similar to that of marine forms, there is considerableelaboration of the vasculature of the branchial chamber lining, which insome species includes a unique double portal system. Cardiac output islower in land crabs (probably related to their higher hemolymph O2 capac-ity), but insufficient data on hemolymph pressures prevent comparisonswith marine forms. In general, land crabs have modified (sometimesextensively) existing structures and processes found in their marine rel-atives rather than evolving structures for terrestrial life de novo. Accord-ingly, land crabs present a useful model for the evolution of terrestriality,showing that even subtle anatomical changes can result in the large changesin physiological function necessary for the terrestrial invasion.
INTRODUCTION related elements of the cardiovascular sys-The evolutionary transition of animals t e m > i n particular, have become specialized
from water to land has been the subject of f o r a terrestrial existence. This paper dis-much study (and considerable speculation) cusses these specializations from the per-in the last decade (see monographs by spective of how the basic pattern of marineRandall et al, 1980; Little, 1983, 1990). brachyuran and anomuran crabs has beenLand crabs have been central in the study modified during the water-land transition,of the arthropod invasion of terrestrial hab-
invasion generally (see Vol. 8, American Morphology of gas exchange organsZoologist, 1968; Vol. 218, Journal of Exper- The morphology of the respiratory organsimental Zoology, 1981; Burggren and of most species of land crabs2 is extensivelyMcMahon, 1988a). Living on land requires modified from that of marine and intertidalnumerous specializations, in large part species. Modifications have occurred in bothbecause the properties of air and water are gill structure and in the branchial chamberso very different (Little, 1990; Burggren and within which the gills reside. However, it isRoberts, 1991). The respiratory system, and important to emphasize that in most cases
1 From the Symposium on The Compleat Crab pre- 2 Here a land crab is defined as a crab (brachyuransented at the Annual Meeting of the American Society or anomuran) that possesses behavioral, morphologi-of Zoologists, 27-30 December 1990, at San Antonio, cal, physiological and/or biochemical adaptation per-Texas. mitting extended activity out of water.
417
Dow
nloaded from https://academ
ic.oup.com/icb/article/32/3/417/1992302 by guest on 13 January 2022

418 WARREN W. BURGGREN
the respiratory adaptations to terrestrial liferepresent modification of respiratory struc-tures common to all crabs, rather than theappearance of structures that have arisen denovo.
The dominant feature of the gills of ter-restrial crabs is their reduced surface arearelative to marine forms. This results fromreduction in both number of gills and num-ber of lamellae on each gill (see McMahonand Burggren, 1988, for earlier literature).The reduced respiratory surface area of thegills of terrestrial crabs is thought to be anadaptation to reduce evaporative water loss.However, reduction of gill surface area alsopotentially reduces the available area acrosswhich the exchange of ions and respiratorygases can occur. In many species (e.g., thecoconut crab, Birgus lato), the posterior gills,which contain the preponderance of cellsresponsible for ion exchange with branchialchamber water, show less proportionatereduction than anterior gills, indicating thatthe gills may retain an important role inhydromineral regulation.
The basic pattern of gill structure inmarine crabs depends upon the buoyingnature of water to prevent the delicate gilllamellae from collapsing and adhering toeach other. When the gills of marine crabsare exposed to air, the collapse of the lamel-lae greatly reduces effective gill surface area.To prevent this, the gill lamellae of manyterrestrial crabs, both brachyuran and ano-muran, are sclerotized to provide greaterstiffness and may have small projectionsfrom their surfaces that act as spacers, keep-ing the lamellae vertically aligned and pre-venting them from collapsing on each other(for references see Burggren and McMahon,1988*).
Many terrestrial crabs exhibit major ana-tomical modifications of the branchialchambers. The branchial chambers arehighly enlarged in many terrestrial species(Cardisoma guanhumi, Gecarcinus later-alis, Dilocarcinus dentatus, Birgus latro)when compared with marine forms of sim-ilar body mass, and the ratio of gill to bran-chial chamber lining surface area appears todecrease progressively when examiningprogressively more terrestrial crabs (McMa-hon and Burggren, 1988). In some land
crabs, the branchial chamber lining remainsrelatively smooth, but in many terrestrialforms the inner surface of the branchialchamber is highly convoluted (e.g., Ocypodequadrata) to provide further enhancementof surface area (Farrelly and Greenaway,1987; Greenaway and Farrelly, 1990; seeMcMahon and Burggren, 1988, for earlierliterature). In Birgus latro, the inner surfaceof the laterally expanded branchial chamberforms numerous small projections ("tree-lets") that undoubtedly increase many-foldthe surface area of the branchial lining. Someof the most extensive branchial chambermodifications occur in Eudianella iturbi,Pseudothelphusa garmani, and Mictyrislongicarpus. The branchial chamber liningin these species is much thicker in manyother crabs but is highly perforated by tinypores (Innes et ai, 1987), giving a "sponge-like" microanatomical appearance superfi-cially resembling the parabronchi of birdlungs.
A unique adaptation for gas exchangeoccurs in the Scopimerinae, a group of ocyp-odid air breathing crabs (Maitland, 1986).The Australian sand bubbler crab Scopi-mara inflata, for example, shows little mod-ification to its gills or branchial chamber.However, single large oval patches on themeral segments of each walking leg showgreatly reduced cuticular and epidermal lay-ers (Fig. 1). Hemolymph spaces lay directlyunderneath these patches, which formrespiratory surfaces with an air-hemolymphdiffusion distance for respiratory gases ofless than 1 /im. Preventing gas exchangeacross these patches reduces total oxygenconsumption by up to 60% (Maitland, 1986).Dotilla, another member of this group, hassimilar respiratory patches located on theventral thoracic sternal plates rather thanon the walking legs (Maitland, 1986).
Ventilatory mechanismsThe ventilatory pumps propelling water
through the branchial chamber in marinecrabs are the paired scaphognathites, flat-tened "paddles" formed from the expoditesof the second maxillae (McMahon andWilkens, 1983). The complex, four-phasedbeat of the pump generates a flow of waterin through the Milne-Edwards openings at
Dow
nloaded from https://academ
ic.oup.com/icb/article/32/3/417/1992302 by guest on 13 January 2022

RESPIRATION AND CIRCULATION IN LAND CRABS 419
the base of the limbs, though the branchialchamber, past the scaphognathite, and outvia the exhalent canals near the base of thethird maxillipeds. Occasional periods ofreversed scaphognathite beating produce areversal of this water flow.
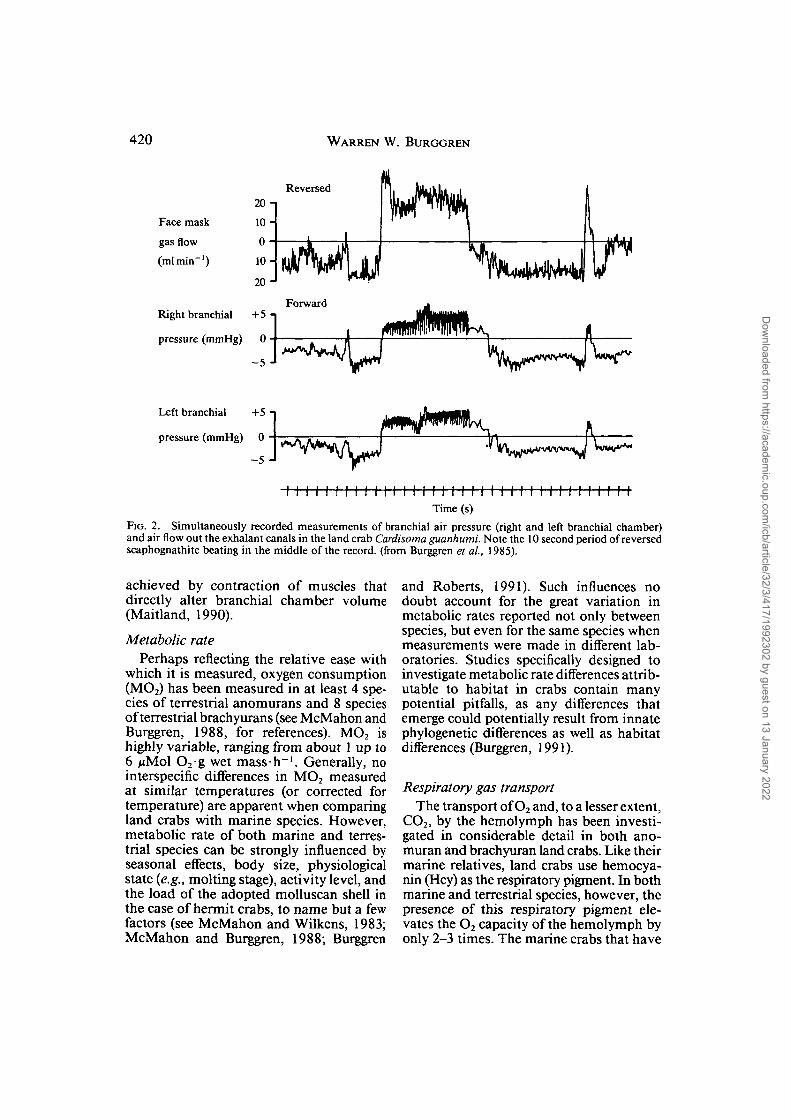
Both brachyuran and anomuran landcrabs continue to employ scaphognathitebeating as the primary ventilatory pump forpropelling air through the branchial cham-bers. The scaphognathites are highly effi-cient at air pumping (Cameron and Meck-lenberg, 1973; McMahon and Burggren,1979; Burggren et ah, 1985), and can sustaina continuous, relatively non-pulsatile pos-terior-to-anterior flow of air through thebranchial chamber (Fig. 2). Total air flowthrough the branchial chambers tends to beconsiderably lower than branchial water flowin marine species (see McMahon and Wil-kens, 1983; McMahon and Burggren, 1988),probably because of the higher O2 capaci-tance of air compared to water. Periods ofreversals occur during air pumping in landcrabs, just as during water pumping bymarine species. Periods of apnea are quitecommon in air breathing crabs, as they arein water breathing crabs. In short, thereappears to be little modification of the basicscaphognathite pumping mechanism whenair replaces water as the respiratory medium.
In land crabs such as Cardisoma, thebranchial chambers often contain somewater even when animals are breathing air.When Cardisoma stands in shallow (1 cm)water, they will sometimes show posturalmodifications (raising or lowering the car-apace to move the inhalant channels belowor above the surface of the water) that con-trol whether water or air is aspirated intothe branchial chamber at the limb bases(Burggren et al., 1985). Such posturalchanges are often associated with alternat-ing periods of forward and reversed sca-phognathite beating. While the physiologi-cal effects of this behavior are unknown, thesplashing and bubbling of branchial cham-ber water may moisten the superior aspectsof the gills and branchial chamber lining, aswell as enhance water and ion exchangebetween branchial chamber water andhemolymph. The semaphore crab, Heloe-cius cordiformus, similarly appears to splash
FIG. 1. The meral segments of the walking legs of theAustralian sand bubbler crab, Scopimera inflata, havea respiratory surface (indicated by star) produced bythinning of the cuticle and epidermis, (from Maitland,1986).
water within the branchial chamber and alsoalter the total amount of branchial waterduring air exposure (Maitland, 1990).
Ventilation of the branchial chamber insome land crabs appears to be aided by lat-eral movements of the abdominal mass,which acts like a piston to draw air in andout of the branchial chamber (e.g., Hol-thuisana transversa, Greenawayefa/., 1983).Birgus latro has muscles that elevate thebranchial chambers (see McMahon andBurggren, 1988, for details). Changes in vol-ume or conformation of the branchialchamber may well alter air flow through thebranchial chamber, and could even generatea gas flow (albeit quite transient) indepen-dent of the scaphognathites. Movement ofepimeral muscles appears to be responsiblefor sudden large pressure changes in thebranchial chambers of Coenobita clypeatus(McMahon and Burggren, 1979), and maysimilarly alter branchial air flow. Certainly,the movement of water (and presumablyair) through the branchial chamber of thesemaphore crab, Heloecius cordiformus, is
Dow
nloaded from https://academ
ic.oup.com/icb/article/32/3/417/1992302 by guest on 13 January 2022
420 WARREN W. BURGGREN
Face mask
gas flow
(mlmin"1)
Right branchial
pressure (mmHg)
Reversed
- 5 J
Left branchial +5 -\
pressure (mmHg) 0
- 5 J
I I I I I I I I I I I I I I I I I I I I 1 I I I I I I I I I I I I I I 1Time (s)
FIG. 2. Simultaneously recorded measurements of branchial air pressure (right and left branchial chamber)and air flow out the exhalant canals in the land crab Cardisoma guanhumi. Note the 10 second period of reversedscaphognathite beating in the middle of the record, (from Burggren et ai, 1985).
achieved by contraction of muscles thatdirectly alter branchial chamber volume(Maitland, 1990).
Metabolic ratePerhaps reflecting the relative ease with
which it is measured, oxygen consumption(MO2) has been measured in at least 4 spe-cies of terrestrial anomurans and 8 speciesof terrestrial brachyurans (see McMahon andBurggren, 1988, for references). MO2 ishighly variable, ranging from about 1 up to6 /JMO\ O 2 g wet mass-h"1. Generally, nointerspecific differences in MO2 measuredat similar temperatures (or corrected fortemperature) are apparent when comparingland crabs with marine species. However,metabolic rate of both marine and terres-trial species can be strongly influenced byseasonal effects, body size, physiologicalstate (e.g., molting stage), activity level, andthe load of the adopted molluscan shell inthe case of hermit crabs, to name but a fewfactors (see McMahon and Wilkens, 1983;McMahon and Burggren, 1988; Burggren
and Roberts, 1991). Such influences nodoubt account for the great variation inmetabolic rates reported not only betweenspecies, but even for the same species whenmeasurements were made in different lab-oratories. Studies specifically designed toinvestigate metabolic rate differences attrib-utable to habitat in crabs contain manypotential pitfalls, as any differences thatemerge could potentially result from innatephylogenetic differences as well as habitatdifferences (Burggren, 1991).
Respiratory gas transportThe transport of O2 and, to a lesser extent,
CO2, by the hemolymph has been investi-gated in considerable detail in both ano-muran and brachyuran land crabs. Like theirmarine relatives, land crabs use hemocya-nin (Hey) as the respiratory pigment. In bothmarine and terrestrial species, however, thepresence of this respiratory pigment ele-vates the O2 capacity of the hemolymph byonly 2-3 times. The marine crabs that have
Dow
nloaded from https://academ
ic.oup.com/icb/article/32/3/417/1992302 by guest on 13 January 2022

RESPIRATION AND CIRCULATION IN LAND CRABS 421
been examined carry a significant fractionof their total O2 in solution in the hemo-lymph rather than chemically bound to Hey(Mangum, 1980), though Hey remains theprimary transport vehicle in resting animals(for review see McMahon and Wilkens,1983; McMahon and Burggren, 1988). Interrestrial crabs, almost all O2 transport isdue to O2 bound to Hey, with dissolved O2in solution playing little or no role in O2transport (McMahon and Burggren, 1988).Does this mean that there are qualitativedifferences in O2 transport between terres-trial and marine forms? Probably not. Asbody temperature rises, the solubility of O2in hemolymph decreases even as O2 demandby the tissues increases, shifting the burdenof O2 transport to hemocyanin. Since almostall terrestrial crabs are tropical or sub-trop-ical (Wolcott, 1992), their body temperaturecan be as much as 20°C higher than tem-perate marine forms (e.g., Cancer magister),from which most data have been collected.Thus, the shift in O2 transport mechanismtowards O2 chemically bound to Hey andaway from O2 dissolved in solution inhemolymph is in all likelihood a direct resultof differences in body temperature ratherthan a physiological adaptation to terrestriallife per se.
Two specific respiratory characteristics ofcrab hemolymph—Hey O2 affinity and HeyO2 capacity—have received considerableattention with respect to putative adapta-tions to the terrestrial habitat. Modulationof Hey O2 affinity by chemical factors is nowknown to involve numerous hemolymphfactors, including urate, lactate, and Ca++
(for reviews see Morris, 1990; Burnett,1992). However, the hemocynanin of ter-restrial species seems to be largely insensi-tive to organic modulators, and in any eventthe hemolymph concentration of thesemodulators can depend upon many of thesame considerations that affect metabolicrate (mentioned above). Not surprisingly,no clear relationship between Hey O2 affin-ity and the tendency towards terrestrialityhas been identified in brachyuran or ano-muran crabs.
What is clear, however, is that terrestrialcrabs generally have a higher total hemo-lymph O2 capacity than do temperate marine
forms due to higher concentrations ofhemolymph Hey (see McMahon and Burg-gren, 1988, for a summary of data and ref-erences). While the adaptive advantages (ifany) of this higher Hey concentration havenot been unequivocally demonstrated, thehigher O2 capacity would allow the samelevel,of tissue O2 transport for a reducedhemolymph flow through the respiratoryorgans. Since conserving water is a primaryconcern of land crabs (Greenaway, 1988;Wolcott, 1992), a reduced hemolymph flowthrough the gas exchange organs couldreduce evaporative water loss in terrestrialforms. Indeed, a decreased cardiac outputis characteristic of land crabs (see sectionbelow on cardiac output).
Carbon dioxide transport, a processinherently much more difficult to quantifythan O2 transport, has not been extensivelyinvestigated in terrestrial crabs. Partial invivo data (i.e., Pco2 in venous hemolymphor pH of arterial hemolymph) abound, asdo in vitro measurements of hemolymphcarriage of CO2 (e.g., Morris et al, 1988),but very few studies on land crabs have pro-vided sufficient data to provide the samequantitative approach to CO2 transport thathas been possible for O2 transport. One gen-eral finding is that hemolymph Pco2, rarelyhigher than 6 mmHg in aquatic crabs(McMahon and Wilkens, 1983), rangesbetween 5 and 15 mmHg in terrestrial crabs(see McMahon and Burggren, 1988). Thistendency for elevated CO2 (and thus lowerhemolymph pH) with increasing terrestri-ality is not unique to crabs, but has beenobserved in virtually every major animallineage showing either an evolutionary ordevelopmental transition from water to land.Because air has a higher O2 capacity thanwater (20-40 times higher, depending upontemperature), air breathing animals canhypoventilate relative to water breathinganimals. This hypoventilation, however,leads to an accumulation of CO2, which isreflected in the higher tissue and body fluidvalues of Pco2 in terrestrial animals. In thecase of land crabs, the higher hemolymphPco2 even at rest places these animals atgreater risk of passing into a respiratory aci-dosis during exercise, but at least in formswith access to water H+ ions can be elimi-
Dow
nloaded from https://academ
ic.oup.com/icb/article/32/3/417/1992302 by guest on 13 January 2022

422 WARREN W. BURGGREN
DS
PLEURAL
SINUSES
+
vs PLEURAL
SINUSES
1
BPVPV J
FIG. 3. A highly diagrammatic representation of thevenous circulation of the terrestrial crab, Ocypode cor-dimanus. DS, dorsal sinus; HS, hepatic sinus; VS, ven-tral sinus; IBS, infrabranchial sinus; AS, abdominalsinus; PS, pericardial sinus; BPV, branchiopericardialvein; PV, pulmonary vein, (from Greenaway and Far-relly, 1984).
nated, and most of the accumulating CO2is stored temporarily as bicarbonate.
Control of ventilationOne of the central dogmas in comparative
respiratory physiology is that, as animalsbecome more and more reliant upon airbreathing for gas exchange needs, ventila-tion of their gas exchange organs is stimu-lated increasingly by CO2 and decreasinglyby O2 (Burggren and Roberts, 1991). Thecontrol of ventilation has been thoroughlyinvestigated in marine crabs—ventilation isprimarily O2 driven, with CO2 playing onlya minor role in influencing the flow of waterpumped through the branchial chamber (seereview McMahon and Wilkens, 1983;McMahon and Burggren, 1988). Althoughrelatively few studies have investigated ven-tilatory control in land crabs, an enhancedrespiratory sensitivity to CO2 has beenreported for Birgus latro, Gecarcinus later-alis and Holthuisana transversa (for reviewsee McMahon and Burggren, 1988). Inter-estingly, the terrestrial hermit crab Coenob-ita clypeatus remains far more sensitive toO2 than CO2 (McMahon and Burggren,1979). This may reflect the fact that C. cly-peatus, although occupying a terrestrialmacrohabitat, probably makes importantand frequent use of its shell water for res-piration.
CIRCULATION: COMPARISON OFTERRESTRIAL AND MARINE FORMS
Cardiovascular morphology and thepattern of circulation
The basic cardiovascular pattern of deca-pod Crustacea is well known (see Burggrenand McMahon, 19886, McMahon and Bur-nett, 1990, and Greenaway and Farrelly,1990, for current reviews and earlier ref-erences). Oxygenated hemolymph ispumped from the heart into a series of pos-terior and anterior arteries. Hemolymphthen passes through the distributing vesselsand on to the exchange vessels, which havea diameter comparable to vertebrate cap-illaries (as small as 2-10 ^m in some spe-cies!). After perfusing the viscera, hemo-lymph flows into a ventral sinus, and thenflows on into the infrabranchial sinuseswhich, as the name implies, lie inferior tothe gills. The infrabranchial sinuses alsoreceive hemolymph returning from thelimbs. Hemolymph from the infrabranchialsinuses flows superiorly into the gills, whereit becomes oxygenated. After leaving thegills, the oxygenated hemolymph flows oninto the pericardial cavity, where it is aspi-rated into the heart through several pairs ofostia in the heart wall.
The morphology of the circulatory systemof a variety of marine crabs has also beenscrutinized, using a combination of dissec-tion of fresh tissues and examination of vas-cular corrosion casts of the hemolymph ves-sels (see Burggren and McMahon, 19886;Greenaway and Farrelly, 1990; Taylor,1990). Generally, with the exception of thecirculation of the branchial chamber lining,there are no qualitative differences in car-diovascular morphology between terrestrialcrabs and closely related marine forms.However, the specialization of the branchialchamber lining in many terrestrial crabs hasresulted in great elaboration of the vascu-lature to this tissue (see Greenaway and Far-relly, 1990). As evident in Figure 3, thebranchial chamber (often termed a "lung,"but see3) receives venous hemolymphdirectly from the dorsal sinus, hepatic sinus
3 The term "lung" is well ensconced in the literatureon air breathing crabs, and for convenience sake thisusage is retained. However, the current author opinesthat the term lung should be restricted to specialized,
Dow
nloaded from https://academ
ic.oup.com/icb/article/32/3/417/1992302 by guest on 13 January 2022

RESPIRATION AND CIRCULATION IN LAND CRABS 423
and ventral sinus, and thus indirectly fromvirtually every body tissue. Oxygenatedhemolymph draining the "lungs" returns tothe pericardial sinus where it mixes withoxygenated hemolymph from the gills, andis aspirated into the heart for redistributionto the tissues.
Corrosion casting of the branchial cham-ber vessels reveals an extensive network ofarborizing vessels draining into large "pul-monary" veins. One of the most fascinatingaspects of the circulation of the branchialchamber lining in many land crabs is thepattern of microcirculation. Essentially,there is a dual portal system, with hemo-lymph passing through three discrete seriesof vascular beds before finally draining intothe pulmonary vein (Greenaway and Far-relly, 1990). Whether there is any controlover the resistance of any of these vesselsis unknown. If such control exists, however,it would provide a mechanism by whichhemolymph could be preferentially distrib-uted between the gills and branchial cham-ber lining. Whatever the mechanism, redis-tribution of cardiac output from the gillstowards the branchial chamber lining occurswhen the amphibious crab Holthuisanatransversa emerges from water (Taylor andGreenaway, 1984). Whether this is a passiveresponse resulting from an increase in bran-chial resistance caused by collapse of thegills upon air exposure, or an active responseinvolving active muscular regulation of vas-cular resistance, remains equivocal. Mus-cular valves regulating flow through theperipheral hemolymph channels of the gilllamellae have been described in the marinecrabs Carcinus maenas and Ovalipes catha-rus (see Taylor, 1990), but whether similarstructures could alter branchial resistanceand thus redirect hemolymph flow to thebranchial chamber lining is as yet undeter-mined. Virtually all animals with two ormore sites for gas exchange have some wayof redistributing blood between these sites,as dictated by ambient conditions, inter-acting needs of respiration and hydromi-neral regulation, or needs for acid-base
distinctive organs that have evolved de now specifi-cally for gas exchange, rather than pre-existing struc-tures {e.g., branchial chambers) merely modified forrespiration.
adjustments (Farrell, 1991). Redistributionof cardiac output between anterior and pos-terior arteries, modulated by neurotrans-mitters, has now been demonstrated formarine crabs and lobsters (see McMahonand Burnett, 1990). Given all of these obser-vations, it would be surprising if terrestrialcrabs did not have a similarly sophisticatedability for altering perfusion between gillsand branchial chamber linings.
Cardiac outputCardiac output is one of the most difficult
of all physiological measurements to makein crabs. Nonetheless, data are available forboth marine (Cancer magister, C. produc-tus, Carcinus maenas) and terrestrial spe-cies {Cardisoma carnifex, Gecarcinus later-alis, Ocypode quadrata and Coenobitaclypeatus) (see Burggren and McMahon,19886; McMahon and Burnett, 1990, forreferences). Cardiac output is generally lowerin the terrestrial crabs. Low cardiac outputhas been correlated with a high blood oxy-gen capacity in both vertebrates and inver-tebrates (McMahon and Wilkens, 1983).This is because cardiac output can still meetoxygen demand if the blood O2 capacity iselevated. As mentioned above, the hemo-lymph O2 capacity of terrestrial crabs tendsto be greater than that in marine crabs, andwith similar metabolic rates, this suggeststhat cardiac output of terrestrial formsshould, indeed, be lower.
While these data suggest a correlationbetween cardiac output and habitat, choos-ing closely related, similarly sized speciesfrom different habitats is of critical impor-tance when attempting to delineate envi-ronmental adaptations from broader, phy-logenetic differences (see Burggren, 1991, fordiscussion and additional references). Com-parisons of the large, temperate, sub-tidalbrachyuran Cancer magister with the small,tropical anomuran Coenobita clypeatus maybe an inappropriate way to address the ques-tion of cardiac output and its relation tohabitat.
Hemolymph pressures and baroreceptorsThere is a paucity of data on hemolymph
pressures at any site in the circulation, inboth terrestrial and marine crabs. Thor-ough, systematic investigations of the
Dow
nloaded from https://academ
ic.oup.com/icb/article/32/3/417/1992302 by guest on 13 January 2022
424 WARREN W. BURGGREN
-5
-6
-7
-S
-9
-10
• 5 * -10%
Changes inIntracardiacPressure(mmHg)
Changes inHeart Rate(beats/min)
Changes in Hemolymph Volume(% of control volume)
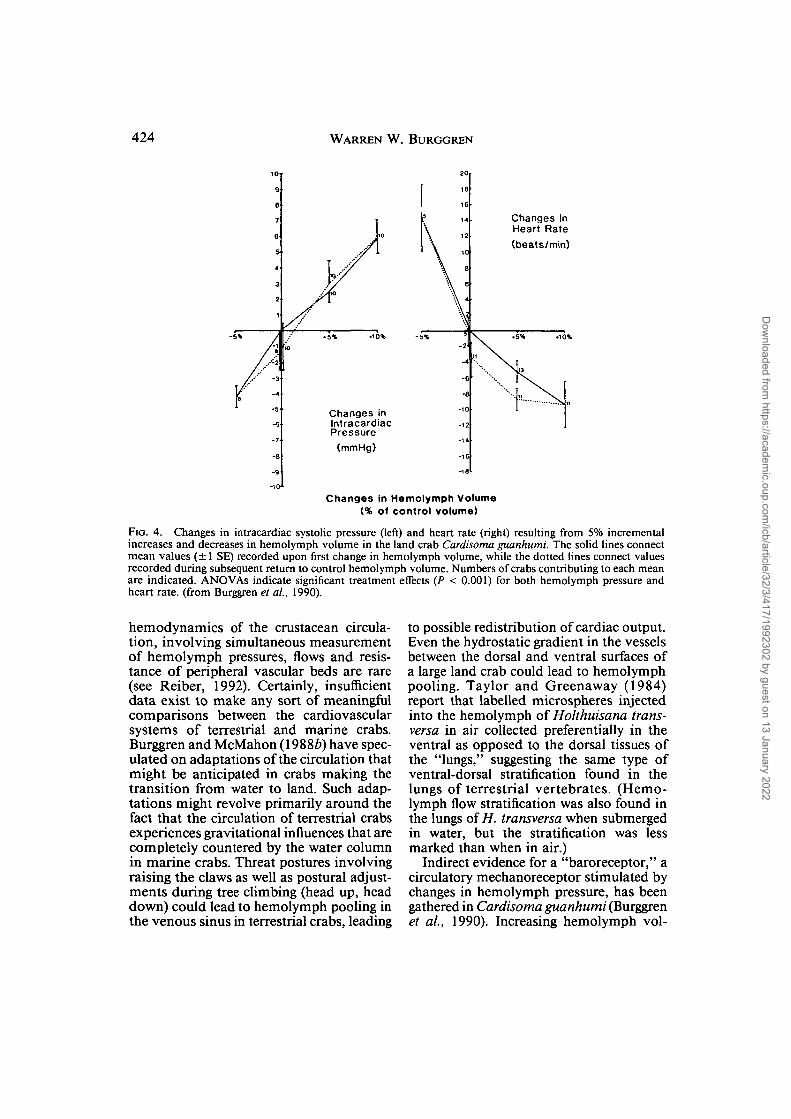
FIG. 4. Changes in intracardiac systolic pressure (left) and heart rate (right) resulting from 5% incrementalincreases and decreases in hemolymph volume in the land crab Cardisoma guanhumi. The solid lines connectmean values (± 1 SE) recorded upon first change in hemolymph volume, while the dotted lines connect valuesrecorded during subsequent return to control hemolymph volume. Numbers of crabs contributing to each meanare indicated. ANOVAs indicate significant treatment effects (P < 0.001) for both hemolymph pressure andheart rate, (from Burggren et ai, 1990).
hemodynamics of the crustacean circula-tion, involving simultaneous measurementof hemolymph pressures, flows and resis-tance of peripheral vascular beds are rare(see Reiber, 1992). Certainly, insufficientdata exist to make any sort of meaningfulcomparisons between the cardiovascularsystems of terrestrial and marine crabs.Burggren and McMahon (19886) have spec-ulated on adaptations of the circulation thatmight be anticipated in crabs making thetransition from water to land. Such adap-tations might revolve primarily around thefact that the circulation of terrestrial crabsexperiences gravitational influences that arecompletely countered by the water columnin marine crabs. Threat postures involvingraising the claws as well as postural adjust-ments during tree climbing (head up, headdown) could lead to hemolymph pooling inthe venous sinus in terrestrial crabs, leading
to possible redistribution of cardiac output.Even the hydrostatic gradient in the vesselsbetween the dorsal and ventral surfaces ofa large land crab could lead to hemolymphpooling. Taylor and Greenaway (1984)report that labelled microspheres injectedinto the hemolymph of Holthuisana trans-versa in air collected preferentially in theventral as opposed to the dorsal tissues ofthe "lungs," suggesting the same type ofventral-dorsal stratification found in thelungs of terrestrial vertebrates. (Hemo-lymph flow stratification was also found inthe lungs of//, transversa when submergedin water, but the stratification was lessmarked than when in air.)
Indirect evidence for a "baroreceptor," acirculatory mechanoreceptor stimulated bychanges in hemolymph pressure, has beengathered in Cardisoma guanhumi (Burggrenet ai, 1990). Increasing hemolymph vol-
Dow
nloaded from https://academ
ic.oup.com/icb/article/32/3/417/1992302 by guest on 13 January 2022

RESPIRATION AND CIRCULATION IN LAND CRABS 425
ume in increments equivalent to 5% totalhemolymph volume by infusing filtered seawater into an infrabranchial sinus causedcorresponding increases in intracardiachemolymph systolic pressure (Fig. 4). Ashemolymph pressure increased, heart ratedecreased. When hemolymph was removed,hemolymph pressure decreased and heartrate increased. These responses are quali-tatively similar to those occurring in a ver-tebrate circulation in which a baroreflexmaintains blood pressure. In fact, the rateof heart rate change in Cardisoma, about 2beats minmmHg-1, is within the rangereported for vertebrate baroreceptor reflexes,conforming to the contemporary view of thedecapod circulation as a relatively complex,efficient system (McMahon and Burnett,1990). Possession of a sensitive barorecep-tor reflex could allow Cardisoma to makeadjustments in hemolymph pressure (andthus in hemolymph flow) during transientperturbations caused by gravitational effectson hemolymph flow and distribution (Burg-gren et al., 1990). Recently, Rajashekhar andWilkens (1991) have manipulated hemo-lymph pressure in the branchiostegal sinusof the shore crab Carcinus maenas. Thisreflexly alters the action of the dorsal ventralmuscles that modify branchiostegal sinusvolume, leading these researchers to con-clude similarly that a reflex involving theregulation of hemolymph pressure residesin this intertidal crab. Independently, Tay-lor and Taylor (1991) elucidated the role ofthe branchiostegal muscles in modifyinghemolymph pressure in Carcinus maenas,and concluded as had Burggren et al. (1990)and Rajashekhar and Wilkens (1991) that abaroreceptor reflex was operating in brachy-urans. It would appear that there may be atleast two reflexes operant—one regulatinghemolymph pressure through heart rate andone through the action of the dorsal ventralmuscles on hemolymph-filled sinuses.Whether the baroreceptor reflexes of Car-disoma and Carcinus are primitive char-acters shared by all brachyurans, or derivedcharacters that have evolved in response toselection pressures associated with exposureto air that is occasional {Carcinus) or fre-quent (Cardisoma), is currently unknown—
other species both marine and terrestrialobviously must be examined in this respect.
CONCLUSIONS
The terrestrial habitat is in many respectsa potentially hostile environment that inmany animals has required substantialpreadaptation for a successful transitionfrom a marine environment (Randall et al.,1980; Little, 1983, 1990; Burggren andRoberts, 1991). Does this need for extensivepreadaptation similarly apply to brachy-uran decapods? Although the proportion ofbrachyuran and anomuran crabs that liveon land is small indeed, certain genera(especially Cardisoma, Coenobita, Gecar-cinus, Geosarma, Ocypode, Sesarma andUca) have shown varying degrees of successin invading the terrestrial habitat. In manyinstances, these crabs have made the tran-sition to air breathing without the evolutionof major new cardio-respiratory structures.Gills are still retained and still achieve somegas exchange in most species, and even thespectacular respiratory surfaces formed bythe branchial chamber lining in Birgus andEudianella represent primarily quantitativemodifications (albeit of great degree) toexisting structures rather than evolution ofqualitatively different structures with sep-arate embryonic origins. Cardiovascularadaptations to the terrestrial habitat are evenmore equivocal, except for the great elab-oration of the vasculature of the branchialchamber lining, most land crabs have a car-diovascular system similar in gross mor-phology to that of marine forms. Thus,major morphological adaptations do notseem necessary for terrestriality in brachy-uran decapods.
If one's goal is to study the evolution ofterrestriality, are not land crabs, with theirsubstantial morphology resemblance to theirmarine relatives (i.e., lack of overt mor-phological adaptation to land), somewhatof a disappointment? To the contrary, theland crabs offer important lessons about theevolution of terrestriality. Perhaps the mostfundamental of these is that major adjust-ments in organ and systems level mor-phology are not an absolute necessity for a
Dow
nloaded from https://academ
ic.oup.com/icb/article/32/3/417/1992302 by guest on 13 January 2022

426 WARREN W. BURGGREN
successful invasion of land by decapods,particularly when combined with adjust-ments in behavior that allow exploitationof microhabitats with some marine features.Generally speaking, the evolution of small,even subtle, alterations in morphologicalcharacteristics can result in much more pro-found evolutionary alterations in physio-logical function—i.e., anatomy and physi-ology are not as tightly linked as someevolutionary biologists would advocate (seeBurggren and Bemis, 1990, for discussion).The terrestrial crabs abound in examplesillustrating this point, but perhaps no moregraphic is the example of Scopimera, inwhich a thinning of the cuticle (a moderatemorphological change) converts the leg fromstrictly an organ of locomotion to an organof respiration!
In summary, land crabs hold a fascinationfor zoologists focusing on changes in respi-ratory and circulatory function during theevolutionary transition from water to air—as much for what these crabs have managedto do with novel variations on the basicmarine theme as for any fundamentally newadaptations that may have arisen.
ACKNOWLEDGMENTS
The author thanks Andrew Gannon andCarl Reiber for providing very useful crit-ical comments on this manuscript. Theauthor was supported by NSF operatinggrant (DCB8916938) during the prepara-tion of this manuscript.
REFERENCES
Burggren, W. W. 1991. Does comparative respiratoryphysiology have a role in evolutionary biology (andvice versa)? In A. J. Woakes, M. K. Grieshaber,and C. R. Bridges (eds.), Comparative insights intostrategies for gas exchanges and metabolism, pp.1-13. Cambridge University Press, Cambridge.
Burggren, W. W. and W. E. Bemis. 1990. Studyingphysiological evolution: Paradigms and pitfalls. InM. H. Nltecki (ed.), Evolutionary innovations: Pat-terns and processes, pp. 191-228. Oxford Univer-sity Press, Oxford.
Burggren, W. W., A. W. Pinder, B. R. McMahon, M.Doyle, and M. G. Wheatly. 1990. Heart rate andhemolymph pressure responses to hemolymphvolume changes in the land crab Cardisoma guan-humi: Evidence for "baroreflex" function. Physiol.Zool. 63:167-181.
Burggren, W. W. and B. R. McMahon (eds.) 1988a.
Biology of the land crabs. Cambridge UniversityPress, New York.
Burggren, W. W. and B. R. McMahon. 19886. Cir-culation. In W. W. Burggren and B. R. McMahon(eds.), Biology of the land crabs, pp. 298-332.Cambridge University Press, New York.
Burggren, W., A. Pinder, B. R. McMahon, M. Wheatly,and M.Doyle. 1985. Ventilation, circulation andtheir interactions in the land crab, Cardisomaguanhumi. J. Exp. Biol. 117:133-154.
Burggren, W. W. and J. L. Roberts. 1991. Respirationand Metabolism. In C. L. Prosser (ed.), Compar-ative animal physiology: Environmental and met-abolic animal physiology, pp. 353-435. Wiley, NewYork.
Burnett, L. E. 1992. Integrated function of the respi-ratory pigment hemocyanin in crabs. Amer. Zool.32:438-446.
Cameron, J. N. and T. A. Mecklenberg. 1973. Aerialgas exchange in the coconut crab, Birgus latro withsome notes on Gecarcoidea lalandii. Respir. Phys-iol. 19:245-261.
Farrell.A.P. 1991. Circulation. In C. L. Prosser (ed.),Comparative animal physiology: Environmentaland metabolic animal physiology, pp. 509-558.Wiley, New York.
Farrelly, C. A. and P. Greenaway. 1987. The mor-phology and vasculature of the lungs and gills ofthe soldier crab, Mictyris longicarpus Latreille. J.Morphol. 193:285-304.
Greenaway, P. 1988. Ion and water balance. In W.W. Burggren and B. R. McMahon (eds.), Biologyof the land crabs, pp. 211-248. Cambridge Uni-versity Press, New York.
Greenaway, P., H. H. Taylor, and J. Bonaventura.1983. Aquatic gas exchange in the freshwater/land crab, Holthuisana transversa. J. Exp. Biol.103:225-236.
Greenaway, P. and C. A. Farrelly. 1984. The venoussystem of the terrestrial crab Ocypode cordimanus(Desmarest 1825) with particular reference to thevasculature of the lungs. J. Morphol. 181:133-142.
Greenaway, P. and C. A. Farrelly. 1990. Vasculatureof the gas-exchange organs in air-breathingbrachyurans. Physiol. Zool. 63:117-139.
Innes, A. J., E. W. Taylor, and A. J. El Haj. 1987.Air-breathing in the Trinidad mountain crab: Aquantum leap in the evolution of the invertebratelung?Comp. Biochem. Physiol. 87A:l-8.
Little, C. 1983. The colonisation of land. CambridgeUniversity Press, Cambridge.
Little, C. 1990. The terrestrial invasion. CambridgeUniversity Press, Cambridge.
Maitland, D. P. 1986. Crabs that breathe air withtheir legs—Scopimera and Dotilla. Nature 319:493-495.
Maitland, D. P. 1990. Carapace and branchial watercirculation; and water-related behaviors in thesemaphore crab Heloecius cordiformus (Decap-oda; Brachyura: Ocypodidae). Mar. Biol. 105:275—286.
Mangum, C. P. 1980. Respiratory function of thehaemocyanins. Amer. Zool. 20:19-38.
Dow
nloaded from https://academ
ic.oup.com/icb/article/32/3/417/1992302 by guest on 13 January 2022

RESPIRATION AND QRCULATION IN LAND CRABS 427
McMahon, B. R. and W. W. Burggren. 1979. Res-piration and adaptation to the terrestrial habitatin the terrestrial hermit crab, Coenobita clypeatus.J. Exp. Biol. 79:265-281.
McMahon, B. R. and W. W. Burggren. 1988. Res-piration. In W. W. Burggren and B. R. McMahon(eds.), Biology of the land crabs, pp. 249-297.Cambridge University Press, New York.
McMahon, B. R. and L. E. Burnett. 1990. The crus-tacean open circulatory system: A reexamination.Physiol. Zool. 63:35-71.
McMahon, B. R. and J. L. Wilkens. 1983. Ventila-tion, perfusion and oxygen uptake. In D. Bliss andL. H. Mantel (eds.), Physiology of the Crustacea,pp. 289-371. Academic Press, New York.
Morris, S. 1990. Organic ions as modulators of respi-ratory pigment function during stress. Physiol.Zool. 63:253-287.
Morris, S., P. Greenaway, and B. R. McMahon. 1988.Adaptations to a terrestrial existence by the robbercrab Birgus latro. I. An in vitro investigation ofblood gas transport. J. Exp. Biol. 140:477-491.
Rajashekhar, K. P. and J. L. Wilkens. 1991. Controlof "pulmonary" pressure and coordination withgill ventilation in the shore crab Carcinus maenas.J. Exp. Biol. 155:147-164.
Randall, D. J., W. W. Burggren, M. S. Haswell, andA. P. Farrell. 1980. The Evolution of air breath-ing in vertebrates. Cambridge University Press,Cambridge.
Reiber, C. 1992. The hemodynamics of the crusta-cean open circulatory systems: Hemolymph flowin the crayfish (Procambarus clarkii) and the lob-ster (Homarus americanus). Ph.D. diss., Univer-sity of Massachusetts, Amherst.
Taylor, H. H. 1990. Pressure-flow characteristics ofcrab gills: Implications for regulation of hemo-lymph pressure. Physiol. Zool. 63:72-89.
Taylor, H. H. and P. Greenaway. 1984. The role ofthe gills and branchiostegites in gas exchange in abimodally breathing crab, Holthuisana transversa:Evidence for a facultative change in the distribu-tion of the respiratory circulation. J. Exp. Biol.111:103-122.
Taylor, H. H. and E. W. Taylor. 1991. The dorso-ventral muscles of Carcinus maenas: Evidence forhydrostatic pressure control in a crab. Physiol. Zool.(In press)
Wolcott, T. G. 1992. Water and solute balance in thetransition to land. Amer. Zool. 32:428—437.
Dow
nloaded from https://academ
ic.oup.com/icb/article/32/3/417/1992302 by guest on 13 January 2022