herbivory by caribbean majid crabs: feeding ecology and plant susceptibility
TRANSCRIPT

•
J. Exp. Mar. Bioi. Ecol., 1988, Vol. 122. pp. 257-276Elsevier
JEM 01151
257
Herbivory by Caribbean majid crabs: feeding ecology andplant susceptibility
Loren D. Coen I
Department of Zoolog)'. Universit)' ofMaryland. College Park. Maryland. U.S.A.
(Received 2 May 1988; revision reccived II July 1988; acccpted 27 July 1988)
Abstract: The present study investigated plant susceptibility. plant quality and food preferences in two smallherbivorous mesograzers, the majid crabs Mithrax scukptus (Lamarck) and Mithrax coryphe (Herbst). Sevenplants were chosen based on characteristics hypothesized to influence grazer preferences. These includedrelative abundance. hypothesized food MqualityM (perccnt ash and water. caloric contents), previousobservations on plant susceptibility to herbivorous fishes and sea urchins and inherent structural andchemical traits.
In single preference (day and night) and multiple selection experiments. preferences for both crab specieswere similar among the seven plants tested and were concordant with those determined previously for fishand urchin grazers on other Caribbean reefs. Mithrax successfully manipulated and consumed both corti·cated (Laurencia. Acanthophora) and leathery (Padina. Lobophora ruffled form) macroalgae using theirspatulate chelae. except where plant secondary chemistry probably reduced consumption (Lobophoradecumbent form and St)'Popodium). Significant differences in plant calorific. perceni organic matter andwater values were found among the seven plants and correlations between these variables and feedingpreference ranks were negative and highly significant. Curiously. correlations between preference rank andcalorific value suggest that crabs were selecting plants with potentially lower net caloric returns. However,when feeding rates were adjusted on a per meal basis, observed grazing rates on "preferred" species maybe sufficiently high for crabs to potentially maximize net energy gain.
Key words: Belize; Coral reef; Crab; Feeding preference; Herbivory
INTRODUCfION
Tropical marine herbivore faunas are quite diverse but often fall into fairly discretefunctional groupings or guilds (Steneck, 1983; Carpenter, 1986; Menge et al., 1986).However, few field studies (see reviews by Borowitzka, 1981; Lubchenco & Gaines,1981; Steneck, 1983; Huston, 1985) have examined the possible impact of reefassociated herbivore species other than fishes and sea urchins, exceptions being thework of Carpenter (1986) and Coen (1988) for example.
Herbivores vary in the methods they employ to manipulate and consume plants inthe marine environment (e.g., Steneck & Watling, 1982; Steneck, 1983; Gaines, 1985;Lewis, 1985. 1986). In large measure, this results from constraints placed on these
Correspondence address: L D. Coen, Marine Environmental Sciences Consortium. Dauphin Island SeaLab, Box 369-370, Dauphin Island, AL 36528. U.S.A.
oo22-0981/88/S03.50 © 1988 Elsevier Science Publishers B.V. (Biomedical Division)
C.C.RE. CONTRIBUTION NO.SUPPORTED IN PART BY EXX~ CORP.
ot:,K,4 Z.'!,/

258 LD.COEN
•
grazers by their morphologies, and to some degree, on selection driven by inherent planttraits (Littler & Littler, 1980; Lubchenco & Cubit, 1980; Paul & Hay, 1986; Targettet al., 1986; Paul et al., 1987; Hay et al., 1988a). These deterrents are not absolute;rather their effectiveness is often dependent upon the herbivore's feeding apparatus (e.g.,Steneck, 1982, 1983; Steneck & Watling, 1982; Gaines, 1985; Padilla, 1985; Coen,1987). The ability of herbivores to overcome these structural and chemical traits mayvary considerably (e.g., Hay et al., 1987a, 1988b). Until recently (Coen, 1987; Hay et al.,1987a, 1988b), morphological and chemical traits as herbivore deterrents in tropicalmacrophytes have been investigated only with respect to macrograzers (e.g., Hay,1981a, 1984b; Littler et al., 1983a, b; Lewis,1985; Morrison, 1986; Paul & Hay, 1986;Targett et al., 1986). Few data are available to test the generality of plant susceptibilitypatterns for smaller tropical invertebrate herbivores (e.g., mesograzers, Brawley & Fei,1987; Hay et al., 1987a; Paul et al., 1987).
Herbivorous crustaceans, such as decapod crabs, have been shown to influencesemi-terrestrial (e.g., Beever et al., 1979) and marine (e.g., Sousa, 1979; Menge &Lubchenco, 1981; Robles & Cubit, 1981; Menge et al., 1986; Coen, 1988) plant communities in both temperate and tropical habitats. For example, Menge & Lubchenco(1981) and Menge et al. (1986) suggest that the scarcity offleshy algae in Pacific Panamais in large part a direct result of grazing crabs. Previous studies have shown that liveplants (as opposed to just plant detritus) often constitute a large proportion of the dietofmany temperate and tropical crabs (Crane, 1947; Hiatt, 1948; Herreid, 1963; Havens, .1974; Hagen, 1977; Warner, 1977; Beeveret 01.,1979; Hines, 1982; Klumpp & Nichols,1983; Leber, 1983; Choy, 1986; Coen, 1987), even in families not previously thoughtto have herbivorous representatives (e.g., Portunidae: Klumpp & Nichols, 1983; Choy,1986). In spite of this, plant grazing preferences of decapod crustaceans are poorlyknown (Coen, 1987), except from indirect work exploring decorating (or camouflaging)behavior of a few majid crabs (Getty & Hazlett, 1978; Mastro, 1981; Wicksten, 1983;Kitar & Lou, 1984, 1986).
Here I prc!lcnt results of a study, the objectives of which were: (1) to examine thecharacteristics ofseven species oftropical marine plants chosen for observed differencesin relative abundance, susceptibility to fish and sea urchin herbivores and previouslyidentified structural and chemical traits (for macrograzers); (2) to establish preferencehierarchies based on comparative grazing rates of two species ofMithrax on this groupof commonly encountered (by crabs) plants; and (3) to test whether crab preferencepatterns conform to those previously observed for macrograzers. Earlier experimentalwork in Belize on macrograzers provides a framework against which grazing by smallerinvertebrate herbivores, su,=h as decapod crabs can be compared.
STUDY SITE AND EXPERIMENTAL ORGANISMS
All collections and experiments were conducted at the Smithsonian Institution'sCame Bow Cay field station, Belize, Central America (16°48'N: 88°05'W) during
, .",
, :.- .. ~.

PLANT SUSCEPTIBILITY TO CRABS 259
October-November, 1984 and June-July, 1985. ROtzler & Macintyre (1982) summarize recent work on the geology, hydrology and biology of adjacent areas within theBelizean barrier reefcomplex. Noms & Bucher's (1982) floristic survey supplies detailedinformation on algae and seagrasses associated with habitats adjacent to Carrie BowCay.
HERBIVORES
The genus Mithrax is one of the most diverse and abundant crab genera in shallowwarm-temperate to tropical waters in the Gulf of Mexico, Caribbean Sea and easternPacific. Previous work (Warner, 1977; Coen, 1988) has demonstrated that CaribbeanMithrax spp. are primarily herbivorous and that they significantly depress algal coverwithin localized areas (Coen, 1988). Mithrax sculptus (Lamarck) and Mithrax coryphe(Herbst) are extremely common majid crabs on western Atlantic coral reefs fromFlorida to Brazil where they are found intertidally to 55 m in cavities in rocks, coralsand sponges (Rathbun, 1925; Powers, 1977; Warner, 1977).
In the present study, M. sculptus and M. coryphe were collected exclusively from smallindividual Porites porites (PalIas) (formafurcata) coral heads in the backreef northeastofCarrie Bow Cay in water < 1.5 m deep. However, both species were locally abundantwherever complex three-dimensional structure (e.g., branching coral colonies, loose andconsolidated rubble) was available. Mithrax coryphe individuals reached a maximumsize of 26 mm carapace width (cw) and a weight of 5.7 g; M. sculptus individualsattained a maximum size of 24.6 mm cw and a weight of 7.3 g (Coen, 1987).
ALGAL PREY SPECIES
Experimental plant species were chosen according to local relative abundance andprevious differences in herbivore susceptibility as derived from earlier fish and seaurchin studies (i.e., herbivore resistant traits, see Table I and Hay, 1981a, 1984a; Noms& Fenical, 1982; Littler et al., 1983a, b; Lewis, 1985, 1986; Taylor et al., 1986)...Distributions of all seven plants overlap those of both species of Mithrax. Seven marineplants were used in experiments to examine feeding rates and preferences of the twospecies of Mithrax.
Plants were divided into functional groupings (sensu Littler & Littler, 1980; Steneck& Watling, 1982) consisting of sheet-like, coarsely-branching, corticated, or leatheryplants. Experimental macrophytes (Table I) included: two red algae [Laurenciapapillosa(Forsskal) Greville and Acanthophora spicifera (Vahl) Boergesen], four brown algae[Padina boergesenii Allender & Kraft (designated P. gymnospora in Taylor 1960),Lobophora variegata (Lamouroux) Womersley ruffied form, Lobophora variegatadecumbent form and Stypopodium zonale (Lamouroux) Papenfuss] and a marine vascular plant, turtle grass (Thalassia testudinum Banks ex Koenig). An additional Lobophoraform (encrusting) was also examined. Wet and dry vouchers for all species weredeposited in the Algal Collection, National Herbarium, National Museum of NaturalHistory, Smithsonian Institution.

260 L.D.COEN
TABU! I
Detailed descriptions ofexperimental plant species (after Steneck & Watling, 1982; Littler et al., 1983a, b;Hay, 1984a; Lewis, 1985, 1986; Coen & Tanner, unpublished).
Division RHODOPHYTA (family Rhodomelaceae)Laurencia papillosa - corticated macrophyte, tough, eartilaginous, erect, densely clustered short determinatebranchlets, primary axes, 2-3 mm diameter; no unusual secondary chemistry, readily eaten by fishes andsea urchinsACQnthophora spicifera - corticated macrophyte, stiff, erect, coarsely branched, spinose, main axes 2-3 mmdiameter; no unusual secondary chemistry, readily eaten by fishes and sea urchins
Division PHAEOPHYTA (family Dictyotaceae)Padina boergesenii - lightly calcified, fan-shaped blades, 2-4 cells thick, 60-100 pm thick; no unusualchemistry, readily eaten by fishes and sea urchinsLobophora variegata (rumed form) - flat to rumed thallus at margins, usually found in clumps, golden brown,blades 100-180 pm thick; blades split easily; no apparent unusual chemistry, readily eaten by sea urchinsand fishes; collected where herbivorous fishes are uncommonLobophora variegata (decumbent form) - thin flat thallus, dark brown to orange brown, 105-130 pm thick;blades, while easy to tear do not split easily; chemistry under investigation, although previous analysisshowed none, rarely eaten by fishes or sea urchins; dominant alga on ridge(> 15 m) east ofCarry Bow Cay,but also found in shallow water in shaded microhabitats such as the undersides of patch reefsStypopodium zonale - leathery macrophyte, thick medulla, iridescent, blades fan shaped to irregularlybranched (caused by splitting ofthe blade), 165-480 pm thick; toxicity demonstrated, essentially untouchedby sea urchins and fishes
Division MAGNOLIOPHYTAThalassia testudinum - vascular plant (seagrass), cortex and epidermis, plades < 2 mm thick, 8-12 mm wide;high cellulose and lignin content; tannin-laden cells, grazed, though not extensively by sea urchins and fishes
MATERIALS AND METHODS
GENERAL METHODOLOGIES FOR FEEDING EXPERIMENTS
Two types of feeding experiments were conducted to determine whether M. sculptusand M. coryphe would mirror fish and sea urchin preference patterns. In the first seriesofexperiments (hence referred to as SPl and SP2), individual crabs were offered singleplants of known weights so that crab preferences could be ranked based on relativeamounts consumed. Both day (SP1) and night (SP2) experiments were conducted forM. sculptus but only day trials were run for M. coryphe. These experiments examinedboth palatability and ingestion rates for each plant separately (Vadas, 1977; Watanabe,1984).
In a second series of experiments (hereafter referred to as MP), all 7 plants (speciesand forms) were presented simultaneously to a group (three) of conspecific crabs(M. sculptus and M. coryphe separately), thereby allowing herbivores to discriminateamong plant species. Plants were then ranked based on relative consumption. Only daytrials were conducted. This method is often preferable to single-prey designs (e.g.,Vadas, 1977; Nicotri, 1980; Watanabe, 1984) because it requires herbivore discrimination, incorporating both tactile and chemosensory responses.

PL\NT Sl'SCEPTlBILlTY TO CRABS 261
Epibiont-free plant material was collected immediately prior to each trial. Plants ofequal weight were found to vary greatly in size (surface area) due to differences in blademorphology. To control for these potential differences in size, plants of similar area wereused. Individual plants were weighed to the nearest 0.01 g, after being shaken twice andpatted dry with paper towels. Concurrent controls were run with plants(11 = 5/species/trial) placed in similar containers without crabs to allow adjustments forany net weight changes due to growth or respiration. All experiments were conductedunder natural light.
At the conclusion of a trial, individual plants were dried as above and reweighed. Allfragments torn loose by crabs were carefully collected and weighed along with intactportions. For all experiments, net weight change (pre-weight minus post-weight) wascalculated rather than percent change as for example Hay (l984a) did, since the lattermethod in this case was judged to be sensitive to pre-weight differences within andamong plants employed in these experiments (Coen, 1987).
Preliminary experiments showed no size or male-female biases in preference or totalconsumption among individuals within either herbivore species. Thereafter, allindividuals were employed without regard to size or sex. Crabs were starved for amaximum of 24 h prior to a trial and used only once per experimental set. Crabsencountered no problems feeding on unattached plants based on extensive feedingobservations with both anchored and unanchored plants. Each crab species experimentwas analyzed individually using absolute plant tissue consumed. Comparisons betweenthe two Mithrax species were made using relative rankings rather than absolute planttissue consumed, since the focus of this study was to examine relative preferences andplant susceptibilities.
SINGLE-SPECIES FEEDING EXPERIMENTS (SPI, SP2)
In single-species experiments, individual crabs (M. sculptus and M. coryphe) fed oneach of the seven plants (Table I) singly in 300 ml plastic freezer containers(11 = 30/species) for 10 h (SPI, 0930-1930). Observations were made at the beginningand at various times throughout the course of each trial on crab manipulation andsubsequent feeding (Coen, 1987). Night experiments (SP2, M. sculptus, 2000-0600,11 = 10), were identical to day experiments except that feeding trials excluded Tha/assia.Concurrent controls were run for both experiments (see general methods). Running seawater was not added during trials; however, no potential dissolved O2 problems wereencountered and only 2 of 450 crabs failed to survive during the feeding trials. Differences in net weight change (for all algae by experiment and herbivore species) werethen analyzed by onc-way ANOVA after data were square-root transformed to meethomogeneity of variances assumption (F-max test, Sokal & Rohlf, 1969). Multiplecomparisons on significant ANOVA (P < 0.05) were performed using a StudentNewman-Keuls test (SNK experiment-wise error rate, P < 0.05). All statisticalanalyses used Statistical Analysis System's (SAS Institute, Version 82.3) GLMprocedure.

262 L.D.COEN
MULTIPLE-SPECIES PREFERENCE EXPERIMENTS
In this experiment, daytime trials (MP, 0930-1430) were conducted offering a crabgroup (three conspecifics) all seven plant species (Table I) simultaneously in an array.Experimental duration (5 h) was sufficient to allow crabs time to sample all algae, whilenot consuming 100% of anyone alga. As before, control thalli, with crabs excluded,were run concurrently. All seven plants were placed randomly (with respect to nearestspecies or form) in a circular array in large plastic trays (46 x 31 x 7 cm). No waterflowed into arenas during trials, but large surface-area and short trial durationsminimized potential dissolved O2 problems. Each group ofcrabs was placed within thecenter of the array and released to reduce positional biases. Observations were madefor the first 30 min to observe whether crabs sampled several species or tended to orientimmediately to particular species.
The quantity consumed for each plant was then determined after 5 h (n = 30 multipleruns). Mean consumption/intraspecific group/5 h for each crab species was comparedusing a Friedman non-parametric (Hollander & Wolfe, 1973) two-way test on rankeddata. This statistical method avoids the problems (E. Russek-Cohen, pers. comm.)normally encountered with multiple-ehoice preference experiments (cf. Hay et al.,1988b). Significant among-species differences (P < 0.01) were then calculated by handusing a Friedman multiple comparisons test (Hollander & Wolfe, 1973) at an experimentwise error rate of P < 0.05.
PLANT CALORIC ANALYSES
Samples ofall experimental plants were collected at Came Bow Cay during July 1985and split into subsamples for drying. Healthy plants free ofepibionts were rinsed in freshwater, patted dry and then weighed on a balance to the nearest 0.001 g for wet weight.They were then dried in an oven (at 60-80 cC) in the field for transport. Upon returnto the laboratory, subsamples were dried (::::::4 h) to constant weight at 80 cC andreweighed (% water) for calorific and ash-free dry weight analyses. Plants were nextfrozen with liquid N2 and pulverized to a fme powder. The resulting powder was pressedinto pellets (::::::0.20-0.26 g each) for calorimetry. An encrusting form of Lobophoravariegata (form c), collected from peat slopes beneath nearby Rhizophora mangle (L.)mangroves (Taylor et al., 1986) was also compared to the two forms of Lobophoraalready mentioned (Table I).
Calorific values (n = 5/macrophyte) were measured using a Parr AdiabaticCalorimeter (model 1243) with a semi-micro bomb (10 cm fuse) (model 1107) controlledby a Parr computer (model 1720). No corrections for endothermy or mineral oil forcombustion were deemed necessary based on preliminary runs. All calorific values arereported in cal/g (wet, dry or ash-free dry weights). Percent ash content (ash-free dryweight) was measured by heating samples (n = 5/macrophyte) to ignition in a mufflefurnace at 400 cC for 24 h. Calories per gram ash-free dry weight and calories per gramwet weight were then calculated using calories· g dry weight - 1, percent ash and water
PLANT SUSCEPTIBILITY TO CRABS 263
content for each plant. Calorific differences among species were analyzed by one-wayANOVA. Proportional data were arcsin-transformed. All data met the homogeneity ofvariances assumption (F-max test, Sokal & Rohlf, 1969). Multiple comparisons onsignificant ANOVA (P < 0.05) used a SNK test (P < 0.05).
RESULTS
SINGLE-SPECIES EXPERIMENTS
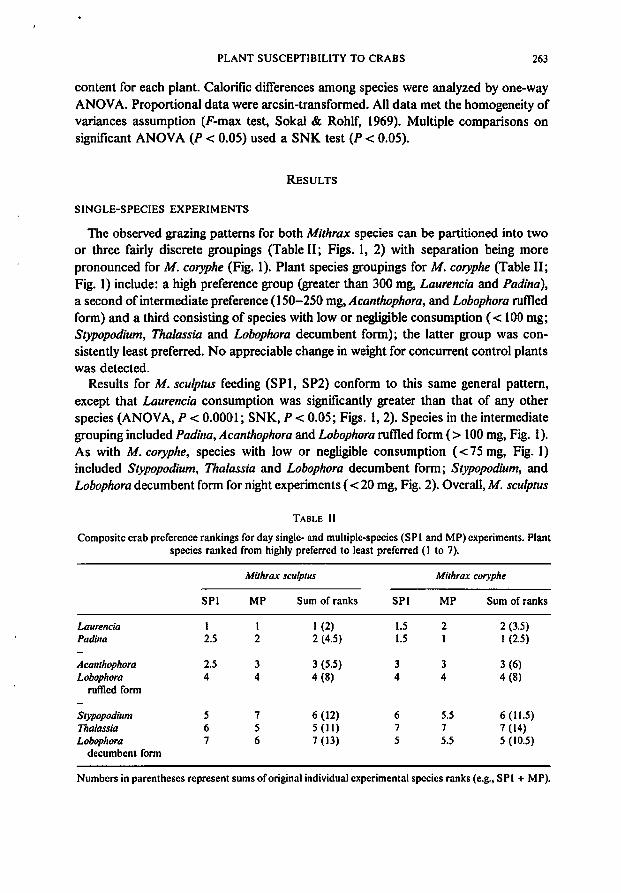
The observed grazing patterns for both Mithrax species can be partitioned into twoor three fairly discrete groupings (Table 11; Figs. I, 2) with separation being morepronounced for M. coryphe (Fig. 1). Plant species groupings for M. coryphe (Table 11;Fig. I) include: a high preference group (greater than 300 mg, Laurencia and Padina),a second ofintermediate preference (150-250 mg,Acanthophora, and Lobophora rumedform) and a third consisting of species with low or negligible consumption ( < 100 mg;Stypopodium, Tha/assia and Lobophora decumbent form); the latter group was consistently least preferred. No appreciable change in weight for concurrent control plantswas detected.
Results for M. scu/ptus feeding (SPI, SP2) conform to this same general pattern,except that Laurencia consumption was significantly greater than that of any otherspecies (ANOVA, P < 0.0001; SNK, P < 0.05; Figs. I, 2). Species in the intermediategrouping included Padina, Acanthophora and Lobophora rumed form (> 100 mg, Fig. I).As with M. coryphe, species with low or negligible consumption «75 mg, Fig. I)included Stypopodium, Thalassia and Lobophora decumbent form; Stypopodium, andLobophora decumbent form for night experiments ( < 20 mg, Fig. 2). Overall, M. scu/ptus
TABLE 11
Composite crab preference rankings for day single· and multiple·species (SP I and MP) experiments. Plantspecies ranked from highly preferred to least preferred (I to 7).
Mithrax sculptus Mithrax coryphe
SPI MP Sum of ranks SPI MP Sum of ranks
Laurencia I I 1(2) 1.5 2 2 (3.5)Padina 2.5 2 2 (4.5) I.S I I (2.5)
Acanthophora 2.5 3 3 (5.5) 3 3 3 (6)Lobophora 4 4 4 (8) 4 4 4 (8)
rumed form
Stypopodium 5 7 6 (12) 6 5.5 6 (11.5)Thalassia 6 5 5 (11) 7 7 7 (14)Lobophora 7 6 7 (13) 5 5.5 5 (10.5)
decumbent form
Numbers in parentheses represent sums oforiginal individual experimental species ranks (e.g., SPI + MP).
264 L.D. COEN
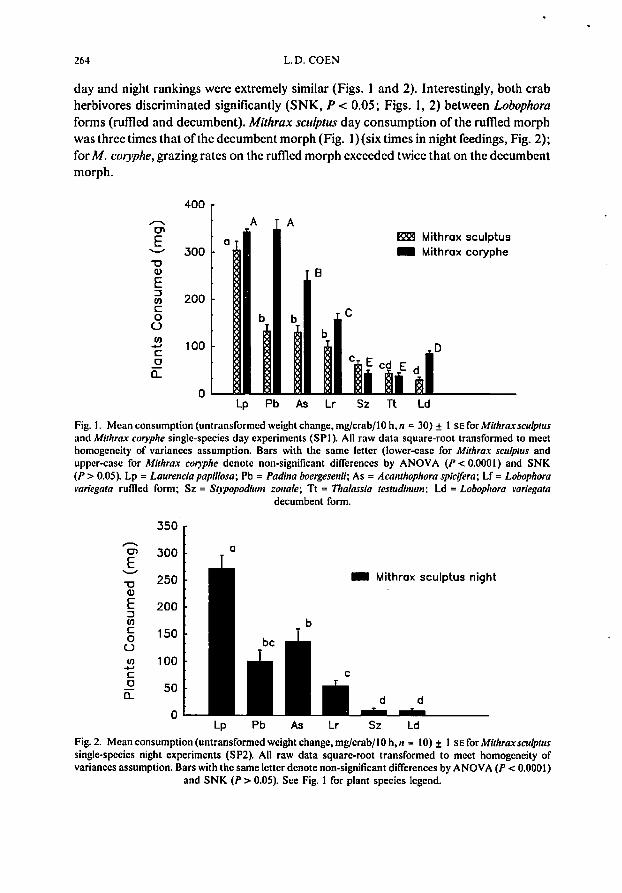
day and night rankings were extremely similar (Figs. 1 and 2). Interestingly, both crabherbivores discriminated significantly (SNK, P < 0.05; Figs. 1, 2) between Lobophoraforms (rumed and decumbent). Mithrax sculptus day consumption of the rumed morphwas three times that ofthe decumbent morph (Fig. 1) (six times in night fcedings, Fig. 2);for M. coryphe, grazing rates on the rumed morph exceeded twice that on the decumbentmorph.
400
........ A ACl
~ Mithrax sculptusE a'-" 300 -Mithrax coryphe"'0
Cl> 8E::l 200Cl)
c C0 b bU
bCl)
+J 100 0cc
a...0
Lp Pb As Lr 5z Tt Ld
Fig. I. Mean consumption (untransformed weight change, mgfcrab/IO h. n = 30) ± I SE for Mitllraxseulptusand Mitllrax eoryplle single-species day experiments (SPI). All raw data square-root transformed to meethomogcneity of varianecs assumption. Bars with the samc Icttcr (Iowcr-case for Mitllrax seu/plus andupper-case for Mitllrax eoryplle dcnote non-significant differences by ANOVA (P < 0.0001) and SNK(P> 0.05). Lp = Lallreneia papillosa; Pb = Padina boergesenii; As =Aeamhophora spieifera; Lf =Lobophoral'ariegata ruffled form; Sz = St.lpopodium zonafe; Tt = Tlla/assia tesllldinllm; Ld = Lobophora mriegma
decumbent form.
350........ cCl 300E
'-" _ Mithrax sCUlptU5 night"'C 250Q)
E 200::lCl)
c 1500U
Cl) 100+JCc 50
D...
0Lp Pb As Lr 5z Ld
Fig.2. Mean consumption (untransformed weight changc,mgfcrab/IO h,n = 10) ± 1 SF. for Mitllraxseulptussingle·speeies night experiments (SP2). All raw data squarc-root transformed to mect homogeneity ofvarianccs assumption. Bars with the same Ictter denote non-significant diffcrenecs by ANOVA (P < 0.0001)
and SNK (P> 0.05). Sce Fig. 1 for plant species Icgcnd.
PLANT SUSCEPTIBILITY TO CRABS 265
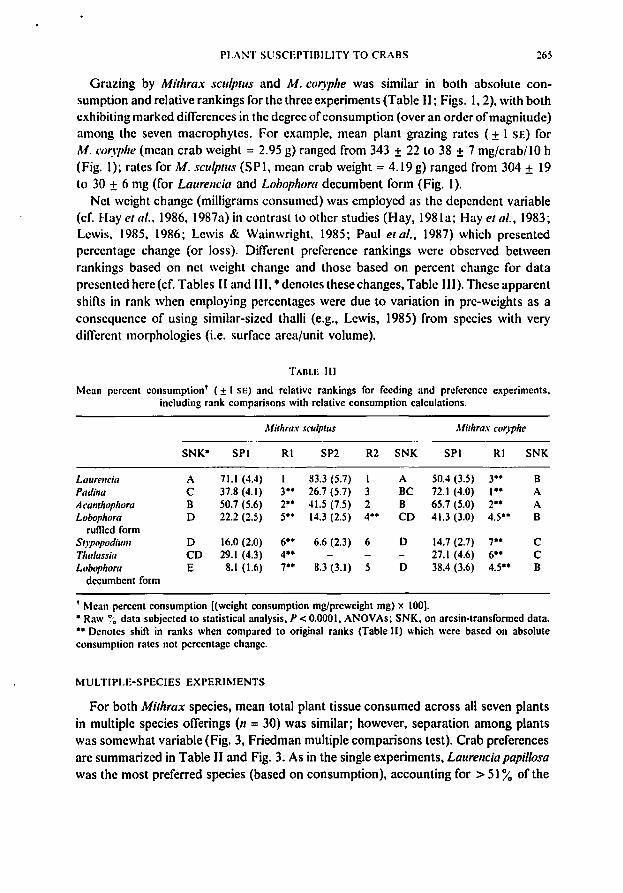
Grazing by Mithrax seulptus and M. eoryphe was similar in both absolute consumption and relative rankings for the three experiments (Table II; Figs. 1,2), with bothexhibiting marked differences in the degree ofconsumption (over an order ofmagnitude)among the seven macrophytes. For example, mean plant grazing rates (± I SE) forM. coryphe (mean crab weight = 2.95 g) ranged from 343 ± 22 to 38 ± 7 mg/crab/lO h(Fig. I); rates for M. selllplUs (SPI, mean crab weight = 4.19 g) ranged from 304 ± 19to 30 ± 6 mg (for Laurelle;a and LobopllOTll decumbent form (Fig. 1).
Net weight change (milligrams consumed) was employed as the dependent variable(cf. Hay et al., 1986, 1987a) in contrast to other studies (Hay, 1981a; Hay et al., 1983;Lewis, 1985, 1986; Lewis & Wainwright. 1985; Paul et al., 1987) which presentedpercentage change (or loss). Different preference rankings were observed betweenrankings based on net weight change and those based on percent change for datapresented here (cf. Tables II and Ill, • denotes these changes, Table Ill). These apparentshifts in rank when employing percentages were due to variation in pre-wcights as aconsequence of using similar-sized thalli (e.g., Lewis, 1985) from species with verydifferent morphologies (i.e. surface area/unit volume).
TABLE III
Mean percent consumption' (± I SE) and relative rankings for feeding and preference experiments,including rank comparisons with relative consumption calculations.
Milhrax sCIIlplus Milhrax cor)'phe
SNKo SPI RI SP2 R2 SNK SPI RI SNK
Laurencia A 71.1 (4.4) I 83.3 (5.7) I A 50.4 (3.5) 3·· BPadina C 37.8 (4.\) 3·· 26.7 (5.7) 3 BC 72.1 (4.0) lOO AAcanthophora B 50.7 (5.6) 2·· 41.5 (7.5) 2 B 65.7 (5.0) 2.0 ALoboplwr" 0 22.2 (2.5) 5·· 14.3 (2.5) 4.0 CD 41.3 (3.0) 4.5·· B
rumcd formSI)'popodil/lII 0 16.0 (2.0) 6·· 6.6 (2.3) 6 0 14.7 (2.7) 7.0 CThalassia CD 29.1 (4.3) 4·· 27.1 (4.6) 600 CLobophora E 8.\ (1.6) 70 • 8.3 (3.1) 5 0 38.4 (3.6) 4.5·· B
decumbent form
, Mean percent consumption [(weight consumption mg/preweight mg) x lOO).o Raw ";~ data subjected to statistical analysis, P < 0.0001, ANOVAs; SNK, on arcsin·transformed data..0 Denotes shift in ranks when compared to original ranks (Table 11) which were based on absoluteconsumption rates not percentage change.
MULTIPLE-SPECIES EXPERIMENTS
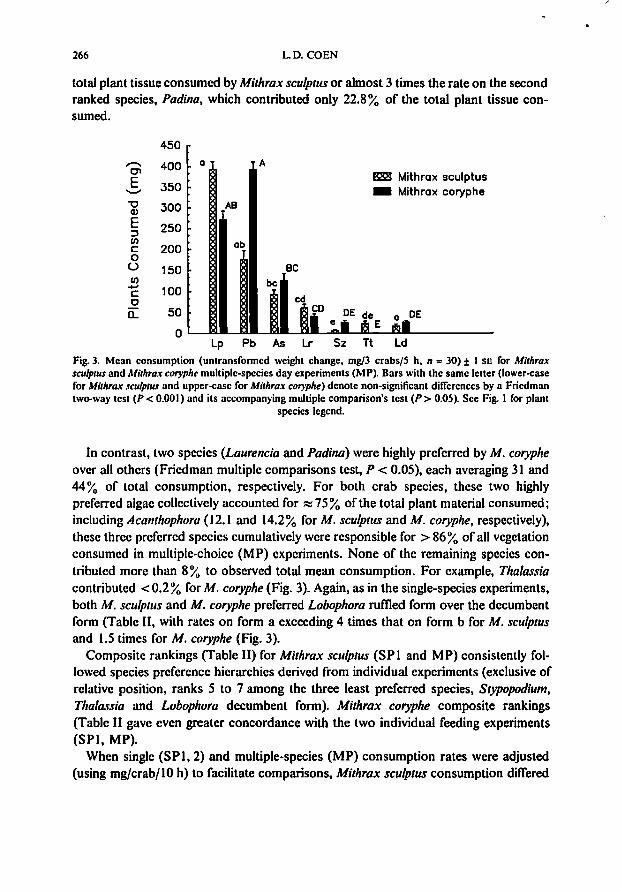
For both Mithrax species, mean total plant tissue consumed across all seven plantsin multiple species offerings (11 = 30) was similar; however, separation among plantswas somewhat variable (Fig. 3, Friedman multiple comparisons test). Crab preferencesare summarized in Table II and Fig. 3. As in the single experiments, Laurellc;a pap;/Iosawas the most preferred species (based on consumption). accounting for> 5I% of the
266 L.O. COEN
total plant tissue consumed by Mithrax sculprus or almost 3 times the rate on the secondranked species, Padina, which contributed only 22.8% of the total plant tissue consumed.
450......... 400 a A0'1
~ Mithrax sculptusE 350'-" _ Mithrax coryphe-0 300 ABQ)
E 250:lCl) abC 2000
U 150 BCCl) be.... 100c cd0
a.. 50
0Lp Pb As Lr Sz Tt Ld
Fig. 3. Mean consumption (untransformed weight change, mg/3 crabs/S h. n = 30) ± 1 SE for MilhraxSCU/PIUS and Milhra:c cor)'phe multiple-species day experiments (MP). Bars with the same leller (lower-casefor Milhrax SCU/pIUS and upper-case for Milhra:c coryphe) denote non-significant differences by a Friedmantwo-way test (P < 0.001) and its accompanying multiple comparison's test (P> 0.05). See Fig. 1 for plant
species legend.
In contrast, two species (Laurencia and Padina) were highly preferred by M. corypheover all others (Friedman mUltiple comparisons test, P < 0.05), each averaging 31 and44% of total consumption, respectively. For both crab species, these two highlypreferred algae collectively accounted for :::::75% of the total plant material consumed;including Acanthophora (12.1 and 14.2% for M. sculptus and M. coryphe, respectively),these three preferred species cumulatively were responsible for > 86%of all vegetationconsumed in multiple-choice (MP) experiments. None of the remaining species contributed more than 8% to observed total mean consumption. For example, Thalassiacontributed < 0.2%for M. coryphe (Fig. 3). Again, as in the single-species experiments,both M. sculptus and M. coryphe preferred Lobophora rumed form over the decumbentform (Table 11, with rates on form a exceeding 4 times that on form b for M. sculptusand 1.5 times for M. coryphe (Fig. 3).
Composite rankings (Table 11) for Mithrax sculprus (SPI and MP) consistently followed species preference hierarchies derived from individual experiments (exclusive ofrelative position, ranks 5 to 7 among the three least preferred species, Stypopodium,Thalassia and Lobophora decumbent form). Mithrax coryphe composite rankings(Table 11 gave even greater concordance with the two individual feeding experiments(SPI, MP).
When single (SPI, 2) and multiple-species (MP) consumption rates were adjusted(using mg/crab/IO h) to facilitate comparisons. Mithrax sculptus consumption differed
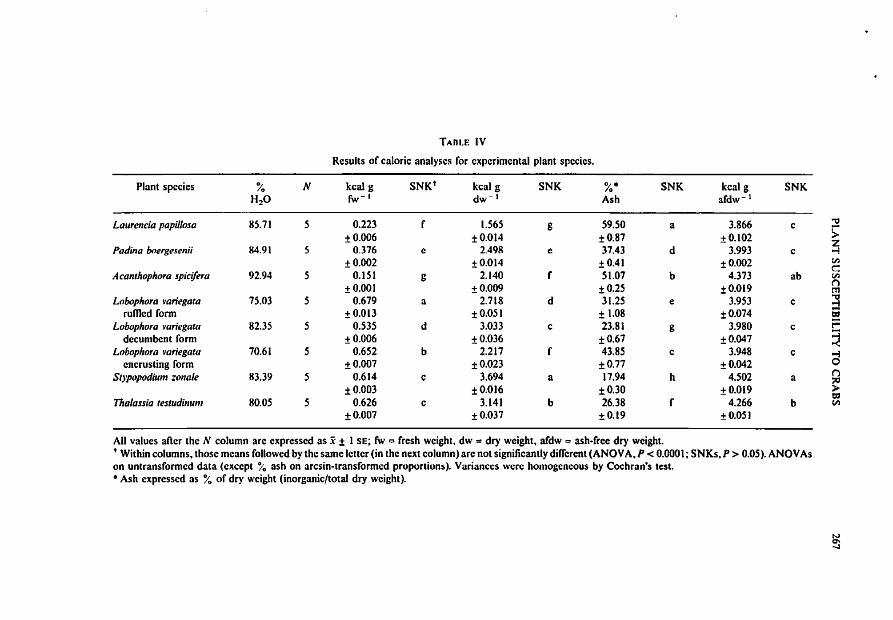
TABLE IV
Results of caloric analyses for experimental plant species.
Plant species % N kcal g SNK' kcal g SNK %. SNK keal g SNKH2O fw- I dw- I Ash afdw- 1
Laurencia papillosa 85.71 5 0.223 1.565 g 59.50 a 3.866 c ""l:Ir-±0.OO6 ±0.014 ±O.87 ±O.102 >
ZPadina boergesenii 84.91 5 0.376 e 2.498 e 37.43 d 3.993 c o-:l
±0.OO2 ±0.OI4 ±O.41 ±O.OO2 V)
c:Acamhophoro spici/era 92.94 5 0.151 g 2.140 r 51.07 b 4.373 ab V)
±O.OOI ±0.OO9 ±0.2S ±O.019(")m
Lobophora ~Iariegata 75.03 5 0.679 a 2.718 d 31.25 e 3.953 c ~rumed form ±0.013 ±O.OSI ± 1.08 ±0.074 1:11
Lobophora ~Iariegata 82.35 5 0.535 d 3.033 c 23.81 g 3.980 c i=decumbent form ± 0.006 ±0.O36 ±O.67 ±0.047 ~
Lobophora var;egata 70.61 5 0.652 b 2.217 f 43.85 c 3.948 c """tencrusting form ±0.OO7 ±0.023 ±0.77 ±0.042 0
Slypopodium zonale 83.39 5 0.614 c 3.694 a 17.94 h 4.502 a(j::0
±0.OD3 ±0.OJ6 ±0.3D ±0.Ot9 >=Thalassia tesludinum 80.05 5 0.626 c 3.141 b 26.38 f 4.266 b (IJ
±O.OO7 ±O.D37 ±O.19 ±O.OS!
All values after the N column are expressed as x ± I SE; fw = fresh weight. dw = dry weight, afdw = ash-free dry weight., Within columns, those means followed by the same letter (in the next column) are not significantly different (ANOVA, P < 0.0001; SNKs,P > 0.05). ANOVAson untransformed data (except i'o ash on arcsin-transrormed proporlion5). Variances were homogeneous by Cochran's lest.• Ash expressed as % of dry weight (inorganic/total dry weight).
268 LD.COEN
'.
little between SPI, 2 and MP experiments. For example, adjusted mean grazing rateson Laurencia were 272 and 262 mg for single night (SP2, Fig. 2) and multiple experiments (MP) (Fig. 3). In contrast, M. coryphe's adjusted consumption for single-speciestrials always exceeded those from multiple-feeding experiments (for Laurencia, 343versus 18 \, respectively. Figs. I and 3).
CALORIC ANALYSES
Caloric values (for dry, ash-free and fresh weights) and percent ash and water contentfor each macrophyte are presented in Table IV. Significant differences (P < 0.0001,ANOVAs) were found among the eight plants for the variables fresh weight (fw), dryweight (dw), and ash-free dry weight (afdw) (kcal per g) and % ash (organics). Plantwater content ranged from 70.6% for Lobophora encrusting form to 93 % for the red algaAcamhophora spicifera. Mean percent ash ranged widely from 18% of dry weight forStypopodium zona/e to almost 60/0 for Laurencia papil/osa. A large proportion of theobserved variation among plant species resulted from interspecific differences in percentash and water content (see Table IV). Mean caloric values ranged from: (1) 0.151(Acam/lOp/lOra) to 0.679 kcal/g (Lobophora rumed form) for fresh weight; (2) 1.56(Laurencia) to 3.69 kcal/g (Stypopodium) for dry weight, and (3) 3.87 (Laurencia) to4.50 kcal/g (Stypopodium) ash-free dry weight.
Spearman rank correlations between caloric and ash data (% ash, kcal/g dw, kcal/gfw, kcal/g afdw) versus the three preference rankings (1-7, most to least preferred;single, multiple and composite) yielded highly significant correlations (Table V) for allbut one (Mithrax scu/ptus, SP I, versus kcal/g afdw) of the relationships. However, allcorrelations were positive (except for % ash) suggesting that crabs tended to preferspecies of lower caloric and organic contents such as Laurencia, Acam/lOphora andPadina. These highly susceptible species had moderate to high ash and water contents(Table IV) thereby shifting their position to the lower end of the caloric-%, ash species'distribution.
TARI.E V
Spearman rank sum correlation (r.) between preference rankings' (single (SPI) and multiple (MP) experi·ments) and mean caloric and ash values.
Experiment '10 Ash keal/g dw kcal/g fw keal/g afdw
M ..feu/plusSPI - 0.87· 0.84· 0.60· 0.22 NSl\IP - 0.95· 0.92· 0.58· 0.50·Composite ranks - 0.92· 0.86· 0.54· 0.35·M. eorypheSPI - 0.82· 0.88· 0.65· 0.50·MP - 0.77· 0.81· 0.61· 0.41·Composite ranks - 0.78· 0.84· 0.63· 0.48·
, Tics between ranks were averaged and used in correlation analyses (rankings 1-7,most to least preferred).• Significant r, correlation coefficients (SAS). P < 0.05.

PLANT SUSCEPTIBILITY TO CRABS 269
Howcvcr, comparisons of adjusted mean caloric intake (mg consumed x kcal valuefw, dw or afdw) per 10 h ofgrazing (SPI, SP2; Coen, 1987) suggest that the significantlygreater grazing rates of both Mitlrrax species on "preferred" plant species (e.g.,Laurencia, Padina, and to a lesser extent Acamlroplrora) may be sufficiently high tomaximize potentially available calories on a per meal basis, all other things being equal(e.g., palatability, handling times, availability). For all four measures (fresh weight, dryweight. percent organics and ash-free dry weight). Laurel1cia (e.g., SPI:mean ± I SE = 0.476 ± 0.030 kcal/dw), Padina (0.331 ± 0.035 keal/dw) and Acanthoplrora (0.278 ± 0.030 kcal/dw) were ranked ahead of all others (0.098 ± 0.018,0.228 ± 0.030, 0.139 ± 0.20 kcal/dw for Lobophora decumbent, Stypopodium andThalassia, respectively), with few exceptions (see Coen, 1987, for all experiments andspecies).
DISCUSSION
PREFERRED MACROPHYTE SPECIES
Crabs consistently discriminated among and even between forms within macrophyte(e.g., Loboplrora variegata) species (Table 11; Figs. 1-3). Marked preferences forLaurencia papillosa, Padina boergensenii and Acanthophora spicijera were exhibited insingle and multiple choice experiments. These species are among the fastest growing(Acanthophora, 2-4.5 mm/day, Kilar & Lou, 1986), abundant and most palatablemacrophyte prey on Caribbean reefs (e.g., Hay, 1981b,c; Littler et al., 1983b; Hay,1984a; Kilar& Lou, 1984, 1986; Lewis, 1985, 1986; Lewis & Wainwright, 1985; Paul& Hay, 1986; Taylor et al., 1986), being consistently preferred by crabs, fishes and seaurchins. All three species were grazed upon by Mitlrrax spp. The crabs either used theirspooned-ehelae to tear off portions of plants (e.g., Laurencia and Padina, Coen, 1987)or manipulated the thallus directly (e.g., Laurencia and Acamhoplrora) towards the oralregion with several of their eight pairs of oral appendages (primarily the 3rd maxillipedand mandibles).
LOW PREFERENCE SPECIES AND PLANT TRAITS
It is increasingly evident that parallels may be drawn between terrestrial and marineplant chemical defense systems (e.g., Norris & Fenical, 1982; Hay, 1984a; Steinberg,1985; Bakus et al., 1986; Paul & Fenical, 1986; Paul & Hay, 1986; Targett et al., 1986;Hay et al., 1987a, b; Paul & Van Alstyne, 1988). However, there has been an unfortunatetendency to equatc the presence of "unusual" or "unique" (terminology used in Hay,1984a; Paul & Hay, 1986) natural compounds as evidence ofchemical defense in manyreef-associated macrophytes (e.g., Hay, 1984a; Paul & Fenical, 1986; Paul & Hay,1986). Relatively few studies until recently (Paul & Fenical, 1983; Targett et al., 1986;Hay etal., 1987a,b, 1988a,b; Paul etal., 1987) experimentally manipulated plantextracts, thereby adequately testing a deterrence hypothesis.

270 L.D.COEN
The plants used in this study are only a small subset of those available to crabs onthe reef. However, it now appears that several of the species tested are consistently oflow preference to both vertebrate and invertebrate marine herbivores. Few speciesspecific plant specialists have been identified in marine plant-herbivore systems (e.g.,Jensen, 1980; Steneck, 1982; Vermeij, 1983). This apparent lack of specificity mayexplain observed generalized deterrent patterns seen for many chemically-defendedmarine plants such as Stypopodium, Caulerpa, Halimeda, and Lobophora (Norris &Fenical, 1982; Paul & Fenical, 1983; Hay, 1984a; Littler et al., 1986; Paul & Hay, 1986;Targett et al., 1986; Hay et al., 1988a; Coen & Tanner, unpublished), although recentwork suggests that there may be some differences (Hay et al., 1987a,b, 1988).Stypopodium and the decumbent form of Lobophora have been found previously to beamong the least preferred algal species based on field experiments with fishes (Hay,1981a, 1984a; Hay et al., 1983; Littler et al., 1986; Morrison, 1986; Paul & Hay, 1986;Coen & Tanner, unpublished) and sea urchins (Littler et al., 1983b).
Stypopodium zonale may be classified as a tough, leathery macrophyte (Littler et al.,1983b, 1986); however, this morphology does not deter crabs or fishes from tearing oreven shredding its uncalcified blades (Coen, pers. obs.). Rather. it appears that palatability is reduced directly by chemical deterrents, in the form of several secondarymetabolites (discussed in Gerwick & Fenical, 1981; Littler et al., 1986; Paul & Hay,1986; Hay et al., 1987b).
Similarly, Lobophora variegata provokes some interesting questions in regard toherbivore susceptibility. Lobophora (probably the mmed form) is readily eaten by fishes(Hay, 1984a; Lewis, 1985; Morrison, 1986; Coen & Tanner, unpublished), sea urchins(Morrison, 1986; Coen, pers. obs.) and crabs (Figs. 1-3). Elsewhere in the literatureLobophora (probably the decumbent form) shows few graze marks and little tissue lossbased on numerous field experiments (Hay, 1981a; Hay & Goertmiller, 1983; Morrison,1986; Paul & Hay, 1986; Coen & Tanner, unpublished). This finding was particularlyevident in areas with high fish grazing pressure in Belize (e.g.• the outer ridge where thedecumbent form dominates the algal community, Lewis & Wainwright, 1985). Bothforms are uncalcified and display no obvious morphologically resistant characters, yetboth Mithrax significantly preferred the rumed form to the decumbent form (up to 6times; Table 1I, Figs. 1-3). An hypothesis presently under investigation (L. Coen & N.Targett) is that some novel secondary chemistry in the decumbent form renders itunpalatable to a phylogenetically diverse group ofherbivores. Gerwick & Fenical (1982)have suggested a similar scenario for a related Pacific species. L. papenjussii. Thechemistry of several forms ofL. variegata is presently under study (L. Coen, N. Targett& C. Tanner, in prep.).
Thalassia testudinum has often been employed in field studies (e.g., Hay, 1981c; Hayetal., 1983; Lewis, 1985; Paul & Hay, 1986; Targett et al., 1986) to estimate localgrazing rates. Mithrax's low preference for Thalassia, as with Lobophora andStypopodium. may similarly be related to plant secondary metabolites. Zapata &McMillan (1979), McMillan et al. (1980), and Harrison & Durance (1985) have found

PLANT SUSCEPTIBILITY TO CRABS 271
relatively high concentrations of water-soluble phenolic acids in seagrasses. Others(Bjorndal, 1980; Dawes & Lawrence, 1980) have estimated that> 45 %of Thalassia'sdry weight is in the form of structural compounds such as cellulose, hemicellulose andlignin. When taken together, these characteristics may render epiphyte-free seagrassesa tough and low-nutritive food that is fairly undigestible to herbivores lacking cellulases(Bjorndal, 1980).
FOOD PREFERENCES AND PLANT QUALITY
Positive correlations between preferences and measured plant attributes (Table IV)initially suggested that crabs tended to choose species of lower organic and caloriccontents. Similar disparate patterns had been observed in earlier feeding studies (Paine& Vadas, 1969; Carefoot, 1973; Vadas, 1977; Larson etal., 1980; Nicotri, 1980;Watanabe, 1984). However, several of the least preferred species, with significantlygreater potential energy returns, also contained chemical feeding deterrents. Thus, these"defended" species may be unavailable to crabs. Alternatively, the plant attributesmeasured here may not be amongst those employed by crabs in selecting plants (e.g.,nitrogen, protein, structural and chemical defenses) (cf. Vadas, 1977; Montgomery &Gerking, 1980; Gaines, 1985; Steinberg, 1985; Hambrook & Sheath, 1987; Hay et al.,1988a). We are currently coUecting data on alternative nutritional cues used by Mithrax.In line with this, recent fmdings from terrestrial systems also suggest that herbivorechoice may be more related to minimizing secondary metabolites than maximizingnutritive gains (Crawley, 1983).
METHODOLOGY AND INTERPRETATION
Analyses based on comparisons of adjusted mean caloric values (see Results andCoen, 1987) suggest that potential available energy (based on calorific contents) fromhigh and low preference species may be equivalent over the course of a meal, due tosignificantly greater feeding rates on preferred plant species with significantly lowercaloric values (see Table IV). Few studies carry the analyses far enough to examine thispotential result (Paine & Vadas, 1969; Vadas, 1977). Most do not even calculatepotential caloric intake on a fresh weight basis. The remainder have tended not toexamine other variables (e.g., toxicity, plant morphology) germane to herbivore choice(but see Steneck & Watling, 1982).
An additional caution for future feeding studies involves the use of relative (orpercent) tissue loss among species (e.g., Lewis, 1985). As discussed earlier (see Resultsand Tables 11 and Ill), biases may be generated in feeding experiments employingpercentage weight change, when similar-sized plant thalli arc used, since natural variation (among species) in thalli weights per unit area is often encountered. Comparisonsof the two methods (% change and net weight change, Table Ill) suggest that rankingsand observed percentages may be quite different between the two methods when usinglarger plants. Low preference species are especially sensitive to these differences. For

272 L.D. COEN
example, suppose that a herbivore consumes 2 g of a particular plant. Grazing on aninitial 10 g segment would yield a 20% net weight change; however, if the initial weightwere decreased to only 4 g, the resulting percentage would be inflated to 50% (rangesare well within those used previously; e.g., 2.65-16.31 g, Hay et al., 1986). For thesingle-species experiments, over 65 % of the ordered rankings changed when the twomethods were compared. This included major changes in significant differences amongspecies means for SPI and SP2 (Table Ill, P< 0.0001, ANOVAs on arcsin-transformed data; P < O.D5, by SNK test). Based on these results it appears that net weightchange may be the preferred method to employ over percent change when using morethan one prey species and/or fonn in preference experiments (cf. Hay et al., 1986).
There has been a tendency in marine plant-animal interactions to ask questionsconcerning community structure by studying large herbivores and using coarsemeasures of macroherbivore impact such as standing crop, percent cover and speciesrichness across relatively large areas (e.g., Menge & Lubchenco, 1981; Menge et al.,1986). Hence, little is known of the effects of herbivores other than fishes, sea urchinsand some molluscs (Lubchenco & Gaines, 1981; Gaines & Lubchenco, 1982; Steneck,1983). The present study focussed on decapod crab grazing patterns and algal susceptibility in Belize. These results are among the first to demonstrate that these smallertropical herbivores are capable of feeding on a diverse group of macrophytes rangingin form from filaments to leathery thalli (see Table I and Littler et al., 1983a, b; Steneck& Watling, 1982). Previously (Coen, 1988), I demonstrated that Mithrax is able to keepcoral (Porites porites) heads free of fouling algal epibionts. This, in conjunction with thefact that the plant species employed in this study (see Table I) often constitute thedominant plant species on many Caribbean reefs (Hay, 1981a; Littler et al., 1983a, b;Kilar & Lou, 1984, 1986; Lewis, 1986), implies that the potential small-scale effects ofmesograzers, such as herbivorous crabs may be underestimated. Lastly, concordantresults of the single and multiple feeding experiments demonstrate that crab choicesreflect the feeding patterns observed previously for fish and sea urchin herbivores.
ACKNOWLEDGEMENTS
I gratefully acknowledge Drs. G. Venneij, M. Littler and J. Norris for supportthroughout all phases of this research. Dr. K. Ruetzler provided support for all Belizework. I. Castro, J. Dineen, C. von Dohlan, M. Hay, K. Heck, D. Littler, G. Naylor,R. Sims, C. Tanner, J. Valentine, and an anonymous reviewer provided assistance andadditional discussion. Support was generously given by the Department ofZoology, theGraduate School, the University Computing Center, the Smithsonian Institution,Lemer-Gray Foundation, and a Sigma-Xi Grant-in-Aid of Research. This paper iscontribution No. 249 to the Caribbean Coral Reef Ecosystems program, SmithsonianInstitution and is partly supported by a grant from the Exxon Corporation.

I'L·\l'T SUSCEPTIBILITY TO CRABS
REFERENCES
273
BAKUS. G.J., N.M. TARGErr & B. SCHULTE. 1986. Chemical ecology of marine organisms: an overview.J. Chem. Ecol., Vol. 12, pp. 951-987.
BEEVER, J. W., D. SIMBERI.OFF & L.L. KING, 1979. Herbivory, and predation by the mangrove tree crabArulllS pi.wlllii. Oecologia (Berlin). Vol. 43, pp. 317-328.
BJORN[)AL, K.A., 1980. Nutrition and grazing behavior of the green turtle Chelol/ia mydas. Mar. Bioi.,Vol. 56. pp. 147-154.
BORowlTzKA, M.A., 1981. Algae and grazing in coral reef ecosystems. Endem·ollr. Vol. 5, pp. 99-106.BRAWI.EY, S. H. & X. G. FEI. 1987. Studies ofmesoherbivory in aquaria and in an unbarricaded mariculture
f;mu on the Chinese coast. J. Phycol.• Vol. 23, pp. 614-623.CARU"o I, T.II.. 1913. Fccdin~, food prcfcrcncc and uptakc offood cncrgy by thc supralillor,.1 i,opod Li/:ia
I'alli.\ii. ,\{ar. mol.. VIlI. IS. pp. 22S-236.C\RI'ENTER, R.C, IlJS6. Partitioning herbivory and its eflccts on coral reef algal communities. I::colog,
MO/wgr., Vol. 56, pp. 345-363.CIIOY, S.C, 1986. Natural diet and feeding habits of the crabs Liocarcillllspuber and L holsallls (Deeapoda,
Brachyura, Portunidae). Mur. £COol. Prog. Ser.• Vol. 31, pp. 87-99.COEN. L. D., 1987. Plant-animal interactions: ecology and comparative functional morphology of plant
grazing decapod (Brachyuran) crustaceans. Ph.D. Dissertation, University of Maryland, College Park.I\ID. 241 pp.
Con.:. L D.• 1988. Herbivol')' by crabs and the control ofalgal epibionts on Caribbean host corals. Oecologiu(Berlin), Vol. 75, pp. 198-203.
CRANE. J.• 1947. E"stern Pacific expeditions of the New York Zoological Society. XXXVIII. Intertid,,1brachygnathous crabs from the wcst coast of tropic,,1 America, \\;th special reference to ecology.Zoologim. Vol. 32, pp. 69-95.
CRAWLEY, M. J., 1983. lIerbimry: Ihe dynamics ofanimal-planl illleraClions. Studies in Ecology. Volume 10.University of California Press, Berkcley, CA. 437 pp.
DAWES, CJ. & J.M. LAWRENCE, 1980. Seasonal changes in the proximate constituents of the seagrassesrhalas.fia lesllldim/m, lIulodllle II'righlii. and SyrillgodiwlI filifonlle. Aqllal. Bol., Vol. 8. pp. 371-380.
GAINES, S. D., 1985. Ilcrbivory and between-habitat diversity: the differential effectiveness of defenses ina marine pl"nt. Ecolo1:,I', Vol. 66. pp. 473-485.
GAINES. S. D. & J. LtHlclIENCO, 1982. A unified approach to marine plant-herbivore interaclions. ".Biogeography. All/Ill. RCI·. Ecol. S,I'SI., Vol. 13, pp. 111-138.
GERWICK. W.B. & W. FENICAL, 1981. lehthyotoxic and cytotoxic metabolites of the Iropical brown algaSlypopodilllll zOIwle (Lamouroux) Papenfuss. J. Org. Chem., Vol. 46, pp. 22-27.
GERWICK. W.H. & W. FENICAL. 1982. Phenolic lipids from related marine algae of the order Dictyotales.PhylOcllemislr.I', Vol. 21, pp. 633-637.
GETTY, T. & B.A. HAZI.ETT. 1978. Decoration behavior in Microphrys bicomullLl (Latreille, 1825)(Decapoda. Braehyura). Cruslacealla, Vol. 34, pp. 105-108.
HAGEN. H. O. VON, 1977. The tree-climbing crabs ofTrinidad. SllId. Fauna Cllracuo Curibb. Islands, Vol. 54.pp. 25-59.
HAMBROOK, J.A. & R.G. SIIEATlI, 1987. Grazing of freshwater Rhodophyta. J. Phycol., Vol. 23,pp. 656-662.
HARRISON, P. G. & C. D. DURANcE, 1985. Reductions in photosynthetic carbon uptake in epiphytic diatomsby water-soluble extracts of leaves of ZOSlera marina. Mar. Bioi.• Vol. 90. pp. 117-119.
HAVENS, A. D.• 1974. The ecology ofeight species ofintertidal crabs ofthe family Xanthidae in the MarshallIslands. Ph.D. Dissertation, University of Southern California. 424 pp.
HAY. M. E., 1981a. Spatial patterns of grazing intensity on a Caribbean barrier reef: herbivory and algaedistribution. Aql/lll. BOI.. Vol. 11. pp. 97-109.
H,,,". M. E.• 1911lb. The funclion,,1 morphology of turf forming seaweeds: persistence in stressful marinehabitats. Ecology, Vol. 62, pp. 739-750.
HAY. M. E., 198\c. Herbivory, algal dislribution and the maintenance ofbetween-habilat diversity upon atropical fringing reef. Am. Nal.• Vol. 118. pp. 520-540.
HAY. M. E.. 1984a. Prcdiclable spatial escapes from herbivory: how do these affect the evolution ofherbivore resistance in tropical marine communities? Oecologia (Berlill). Vol. 64. pp. 396-407.

274 LD.COEN
HAY. M. E.• 1984b. Patterns of fish and urchin grazing on Caribbean coral reefs: arc previous patternstypical? Ecology. Vol. 65. pp. 446-454.
HAY. M. E. & T. GOERTEMILLER. 1983. Between·habitat differences in herbivore impact on Caribbean coralreefs. In. The Ecology ofDeep and Shallow Coral Reefs. Symposia Seriesfor Undersea Research. Vol. I.editedby M. L Reaka. Office of Undersea Research, NOAA, MD. pp. 97-102.
HAY. M. E.• T. COLBURN & D. DOWNING, 1983. Spatial and temporal patterns of herbivory on a Caribbeanfringing reef: the effects on plant distributions. Oecologia (Berlin). Vol. 58, pp. 299-308.
HAY. M. E.• R.R. LEE, JR. & M.M. BENNETI', 1986. Food preference and chemotaxis in the sea urchinArbacia punctulata (Lamarck) Philippi. J. Exp. Mar. Bioi. Ecol., Vol. 96, pp. 147-153.
HAY. M. E.• J. E. DUFFY. C.A. PFISTER & W. FENICAL, 1987a. Chemical defense against different marineherbivores: are amphipods insect equivalents? Ecology. Vol. 68, pp. 1567-1580.
HAY, M. E., W. FENICAL & K. GUSTAFSON. 1987b. Chemical defense against diverse coral·reefherbivores.Ecology. 68, pp. 1581-1591.
HAY. M.E., V.J. PAUL, S.M. LEWIS, K. GUSTAFSON. J. TUCKER & R.N. TRINDELL. 1988a. Can tropicalseaweeds reduce herbivory by growing at night?: dic! patterns of growth, nitrogen content. herbivory,chemical versus morphological defenses. Oecologio (Berlin), Vol. 75, pp. 233-245.
HAY, M.E., R.E. RENAUD & W. FENICAL, 1988b. Large mobile versus sedentary herbivores and theirresistancc to seaweed chemical defenses. Oecologia (Berlin), Vol. 75, pp. 246-252.
HERREID, C. F.• 11, 1963. Observations on the feeding behaviour of Cardisoma guanhumi (Latreille) insouthern Florida. Crustaceana, Vol. 5. pp. 176-180.
HIATT. R. W.• 1948. The biology of the lined shore crab, Pachygrapsus crassipes Randall. Pac. Sci., Vol. 2.pp. 135-213.
HINES, A. H., 1982. Coexistence in a kelp forest: size. population dynamics. and resource partitioning ina guild of spider crabs (Brachyura, Majidae). Ecol. Monogr., Vol. 52, pp. 179-198.
HOI.l.ANDER, M. & D.A. WOLFE, 1973. Nonparametric statistical methods. Witey and Sons, New York.503 pp.
HUSTON. M.A., 1985. Patterns of species diversity on coral reefs. Annu. Rev. Ecol. Syst•• Vol. 16.pp. 149-177.
JENSEN, K. R., 1980. A review ofsacoglossan diets. with comparative notes on radular and bueeal anatomy.Malacol. Rev., Vol. 13, pp. 55-77.
KILAR, J.A., & R.M. Lou, 1984. Ecological and behavioral studies of the decorator crab, Microphrysbicomutus Latreille (Decapoda: Brachyura): a test of optimal foraging theory. J. Exp. Mar. Bioi. Ecol.,Vol. 74, pp. 157-167.
KILAR, J. A. & R. M. Lou, 1986. The subtleties ofcamouflage and dietary preference ofthe decorator crab,Microphrys bicomutus Latreille (Decapoda: Brachyura). J. Exp. Mar. Bioi. Ecol.• Vol. 101, pp. 143-160.
KLUMPP. D. W. & P.D. NICHOLS. 1983. Utilisation of the seagrass Pasidonia australis as food by the rockcrab Nectocareinus integriji-ons (Latreille) (Crustacea: Decapoda: Portunidae). Mar. Bioi. Lett•• Vol. 4.pp. 331-339.
LARSON, B.R.• R.L VADAS & M. KESER. 1980. Feeding and nutritional ecology of the sea urchinStrongylocentrotus drobachiensis in Maine. USA. Mar. Bioi.• Vol. 59, pp. 49-62.
LEBER, K. M.• 1983. Feeding ecology of decapod crustaceans and the influence of vegetation on foragingsuccess in a subtropical seagrass meadow. Ph.D. Dissertation. Florida State University, Tallabassce. FL,166 pp.
LEWIS. S. M.• 1985. Herbivory on coral reefs: algal susceptibility to herbivorous fishes. Oecologio (Berlin),Vol. 65. pp. 370-375.
LEWIS, S. M., 1986. The role ofherbivorous fishes in the organization ofa Caribbean reefcommunity. Ecol.Monogr., Vol. 56. pp. 183-200.
LEWIS, S. M. & P. C. WAINWRIGHT. 1985. Herbivore abundance and grazing intensity on a Caribbean coralreer. J. Exp. Mar. Bioi. Ecol., Vol. 87. pp. 215-228.
LITTLER, M. & D. LITTLER, 1980. The evolution of thallus form and survival strategies in benthic marinemacroalgae: field and laboratory tests of a functional fonn model. Am. Nat., Vol. 116, pp. 25-44.
Ll'rrLER. M.M.• D.S. LITTLER & P.R. TAYLOR. 1983a. Evolutionary strategies in a tropical barrier reefsystem: functional·form groups of marine macroalgac. J. Phycol., Vol. 19. pp. 229-237.
LITTLER. M. M.,P.R. TAYLOR & D.S. LITTLER, 1983b. Algal resistancc to herbivory on a Caribbean barrierreer. Coral Reefs, Vol. 2, pp. 111-118.

, .
PLANT SUSCEPTIBILITY TO CRABS 275
LITIl.ER, M. M., P. R. TAYLOR & D. S. LITILER, 1986. Plant defense associations in the marine environment.Coral Reefs, Vol. 5, pp. 63-71.
LUBClIENCO. J. & J. CuBIT, 1980. Heteromorphic life histories of certain marine algae as adaptations tovariations in herbivory. Eeology. Vol. 61, pp. 676-687.
LUBCIIENCO. J. & S.D. GAINES. 1981. A unified approach to marine plant-herbivore interactions. I.Populations and communities. Annu. Rev. Eeol. Syst.• Vol. 12, pp. 405-437.
McMII.LAN. c.. O. ZAPATA & L. ESCODAR. 1980. Sulphated phenolic compounds in seagrasses. Aq. Bor.•Vol. 8, pp. 267-278.
MAsTRo, E.• 198I. Algal preferences for decoration by the Californian kelp crab, Pugeltia produeta (Randatl)(Deeapoda, Majidae). Crustaeeana, Vol. 41. pp. 64-70.
MIlNGE, B.A. & J. LUBClIENCO, 1981. Community organization in temperate and tropical rocky intertidalhabitat: prey refuges in relation to consumer pressure gradients. Eeol. Monogr., Vol. 5 I, pp. 429-450.
MIlNGE. B. A.• J. LUBCHENCO, L. R. ASIIKENAS & F. RAMSEY, 1986. Experimental separation ofeffects ofconsumers on sessile prey in the low zone of a rocky shore in the Bay of Panama: direct and indirectconsequences of food web complexity. J. Exp. Mar. Bioi. Eeol.• Vol. lOO, pp. 225-269.
MOJl,TGOMERY. W. L. & S. D. GERKING. 1980. Marine maeroalgae as foods for fishes: an evaluation ofpotential food quality. Env. Bioi. Fishes. Vol. 5. pp. 143-153.
MORRISON. D.E., 1986. Algal-herbivore interactions on a Jamaican coral reef. Ph.D. dissertation.University of Georgia. Athens. 127 pp.
NICOTRI. M. E.• 1980. Factors involved in herbivore food preference. J. Exp. Mar. Bioi. Eeol.• Vol. 42.pp. 13-26.
NORRIS. J. N. & K. E. BUCIIER. 1982. Marine algae and seagrasses from Carrie Bow Cay. Belize. In. TheAtlantic Barrier Reef Ecosystem at Carrie BolV Cay. Belize, edited by K. ROtzler & I.G. Maeintyre.SmithsOllian Colltrib. Mar. Sci., Vol. 12, pp. 167-238.
NORRIS. J. N. & W. FENICAL. 1982. Chemical defense in tropical marine algae. In. The Atlantic Bamer Reefecosystem at Came Boil' Cay. Belize, edited by K. Riltzler & I. G. Macintyre. Smilhsonian Contrib. Mar.Sci., Vol. 12. pp. 417-431.
PAOILLA. D. K., 1985. Struclural resistance of algae to herbivores. Mar. Bioi., Vol. 90, pp. 103-109.PAINE, R. T. & R. L. VAOAS. 1969. Calorific values of marine benthic algae and their postulated relation
to invertebrate food preference. Mar. Bi"I., Vol. 4. pp. 79-86.PAUL. V.J. & W. FENICAL. 1983. Isolation ofhalimedatrial: chemical defense adaptation in the calcareous
reef building alga Halimeda. Science, Vol. 221. pp. 747-749.PAUL. V.J. & W. FENICAL, 1986. Chemical defense in tropical green algae, Caulerpales. Mar. Eeol. Prog.
Ser.• Vol. 34. pp. 157-169.PAUl., V.J. & M.E. HAY. 1986. Seaweed susceptibility to herbivory: chemical and morphological correlates.
Mar. Eeol. Prog. Ser.• Vol. 33, pp. 255-264.PAUl., V.J. & K.L. VAN ALSTYNE. 1988. Chemical defense and chemical variation in some tropical Pacific
species of Halimeda (Halimedaceae: Chlorophyta). Coral Reeft.• Vol. 6. pp. 263-269.PAUl., V.J., M.E. HAY.J. E. DUFFY. W. FENICAL & K. GUSTAFSON. 1987. Chemicaldefense in the seaweed
Oetodes seeundiramea (Montagne) Howe (Rhodophyta): effects of its monoterpenoid components upondiverse coral-reef herbivores. J. Exp. Mar. Bi"l. Eeol.• Vol. 114. pp. 249-260.
POWERS, L. W., 1977. A catalogue and bibliography to the crabs of the Gulf of Mexico. Vni\'. Texas Mar.Sci. Inst. Comrib. Mar. Sci. Suppl.• Vol. 20. pp. 1-190.
RATIIBUN. M.J.• 1925. The spider crabs of America. Bull. V.S. National Museum. Vol. 129, pp. 1-613.ROBI.ES. C. D. & J.D. CUBIT. 1981. Influence of biotic factors in an upper intertidal community: effects of
grazing diptera larvae on algae. Ecology. Vol. 62, pp. 1536-1547.ROTZLER K. & LG. MACtNTYRE (lOOS.). 1982. The Atlantic Barrier Reef Ecosystem at Carrie Bow Cay,
Belize. Smirhsonian Contrib. Mar. Sci., Vol. 12. 539 pp.SOKAL, R. R. & F.J. ROHLF. 1969. Biometry. Freeman & Sons, San Francisco. 776 pp.SOUSA. W. P.• 1979. Experimental investigations of disturbance and ecological succession in a rocky
intertidal algal community. Eeol. Monogr., Vol. 49, pp. 227-254.STmNBERG, P.D.• 1985. Feeding preferences of Tegulafunebralis and chemical defenses off marine brown
algae. Eeol. MOllogr., Vol. 55. pp. 333-349,STENECK, R.S., 1982. A Iimpet-coratline alga association: adaptations and defenses between a selective
herbivore and its prey. Eeology. Vol. 63. pp. 507-522.

276 L.D.COEN
Sn'NI'CK. RS.• 1983. Escal:lling hcrbivory and rcsulling adaptivc trcnds in c:llcarcolls :l1g'll crusts. PaleobMtlg)·. Vol. 9. pp. 44-61.
STJ:NI'CK. R.S. & L. WATLlNG. 1982. Fccding capabilities and Iimilations of herbivorous molluscs: AfUllctional group approach. Mtlr. Bioi•• Vol. 68. pp. 299-319.
TAllmrn. N.M .. T.E. TARGETT. N.B. VltOI.IJK & I.C. OGDEN, 1986. Effeel of m:lcrophyte secondaryIllctabolites on feeding preferences ofthe herbivorous parrotfish Sparisoma rat/iatl.r. Mtlr. Ecol. Prog. Ser.,Vol. 92. pp. 141-148.
TAYI.OIl, I>.R., M.M. LITTI.ER & D.S. Lrrl'l.l'R, 1986. Escapes from hcrbivory in rcl:llion to the structureof m:lIIgrove island macroulgal communities, Deeologia (Berlill), Vo!. 69. pp. 481-490.
TAYI.OR, \V,lt, 1960. Maril/e tllgae oftire ea.rtem tmt/ tropical tl/ulsllbtropical coasts ofthe Americas. Universityof Michigan I'rcss, Ann Arbor. Michigun. 870 pp.
VA1)AS, R. L., 1977. Preferential feeding: :111 optimizution strategy in sea urchins. Erol. /II01I0gr•• Vo!. 47,pp. 337-371.
VI:RMHIJ. G.I .• 1983. Intimate associations und coevolution in the sea. In. CtlI:l'tllllliOlI, edited by D.J.Futllyma & M. Slatkin, Sinauer Press. Sunderland. MA. pp. 31 1-327.
WARNI:R. G. F.• 1977. Ti,e biololt)' ofcmbs. Van Nostrand. New York. 202 pp.WATAN,\OH. I.M.• 1984. Food preference. food quality, and diels of three herbivorous gastropods
(Trochidae: Tegltla) in a temperate kelp forest. Deeologia (Berlill), Vo!. 62, pp. 47-52.WICKSTIJN, M. K.. 1983. Camouflage in marine invertebrates. DceatlOgr. Mar. Hi"I. AIIIIIl. Rei'•• Vo!. 21.
pp. 177-193.ZAI'ATt\, O. & C. McMIU.AN. 1979. I'henolic acids in scagrasses. Aqua', Btll., Vo!. 7. pp. 307-317.
"