larval development of the eastern pacific mud crab acantholobulus mirafloresensis (abele and kim,...
TRANSCRIPT

*Corresponding author.
Invertebrate Reproduction and Development, 47:2 (2005) 133–145 133Balaban, Philadelphia/Rehovot0168-8170/05/$05.00 © 2005 Balaban
Larval development of the eastern Pacific mud crabAcantholobulus mirafloresensis (Abele and Kim, 1989)(Decapoda: Brachyura: Panopeidae) described from
laboratory-reared material
JOSÉ SALGADO-BARRAGÁN1* and MARCELA RUIZ-GUERRERO21Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México,
Código Postal 82000, Apartado Postal 811, Mazatlán, MéxicoTel. +52 (669) 985-2845; email: [email protected]
2Centro de Investigación en Alimentación y Desarrollo, A.C., Unidad Mazatlán,Código Postal 82010, Apartado Postal 711, Mazatlán, México
Received 29 March 2004; Accepted 19 May 2005
Summary
The complete larval development of the eastern Pacific mud crab Acantholobulus mira-floresensis is described and illustrated from laboratory-reared material. The larval developmentconsists of four zoeal stages and one megalopa. Comparison of A. mirafloresensis with descrip-tions of A. bermudensis and A. schmitti from the western Atlantic, shows that the first zoea of allthree species seems to be identical, whereas major differences appear between the species at thezoea II to megalopa stages, mostly related with setation of carapace, mouthparts and abdomen.
Key words: Acantholobulus mirafloresensis, Panopeidae, larval development, eastern Pacific
Introduction
The study of phylogenetic relationships amongxanthoid crabs traditionally included the adult mor-phology, but the evolutionary convergence of theexternal features in species and the scarcity of recog-nized criteria to group them, has complicated theirclassification and led to the construction of artificialtaxa (Martin and Abele, 1986). Therefore, the use of abroader set of characters should be used to elucidatethese taxonomic relationships such as the male firstpleopod morphology and the positioning of the genitalopenings with respect to the sternal plates (Guinot,1978; Martin and Abele, 1986; Hendrickx, 1998),
combined with analysis of DNA–rRNA sequences(Schubart et al., 2000) and the study of the larvalmorphology (Rice, 1981; Martin, 1984; Martin et al.,1985; Clark and Ng, 1998). Larval development is nowconsidered important to identify planktonic forms andthe reorganization of phylogenetic relationships amongdecapod crustaceans (Clark et al., 1998).
Based on a systematic revision of adult specimens(holotypes) and supported by larval morphology,Felder and Martin (2003) erected the genus Acantho-lobulus to contain A. schmitti (Rathbun, 1930), A. ber-mudensis (Benedict and Rathbun, 1891), A. pacificus(Edmondson, 1931) and A. mirafloresensis (Abele andKim, 1989), formerly assigned to Hexapanopeus and

J. Salgado-Barragán and M. Ruiz-Guerrero / IRD 47 (2005) 133–145134
Panopeus. Diagnostic character states of larvae ofA. bermudensis and A. schmitti include carapace ofzoeas without lateral spines, tapering antennal proto-pod, unarmed and slightly dilated at tip, reducedantennal exopod, telson forks with one dorsal spinelocated beyond the median point, and with no spines ormarginal setae beyond this structure.
A. mirafloresensis is a common mud crab intropical and temperate coastal lagoons from the Gulf ofCalifornia to the Panama Canal. The species is eury-haline and eurythermic, and frequently associated withmangrove prop roots and hard substrata, includingbivalves (Abele and Kim, 1989; Salgado-Barragán andHendrickx, 1996, 2002; Félix-Pico et al., 2003). Incoastal lagoons of the southeastern Gulf of California,it commonly coexists with the panopeids Eurypano-peus canalensis Abele and Kim, 1989, H. beebeiGarth, 1961, P. chilensis H. Milne Edwards and Lucas,1844, and P. purpureus Lockington, 1877 (Salgado-Barragán and Hendrickx, 2002).
In contrast to the western Atlantic panopeids, studyof the larval development of the eastern Pacific specieshas been restricted to a description of the first zoealstage of P. purpureus (Martin et al., 1998) and thecomplete larval development of E. canalensis (García-Guerrero et al., 2005). The purpose of this study is todescribe the complete larval development of A. mira-floresensis, from hatching to the megalopa stage, basedon laboratory-reared material, and compare larvalstages between this and two closely related species,A. bermudensis and A. schmitti.
Materials and Methods
Four ovigerous A. mirafloresensis were collectedtogether on March, 2003, from mangrove prop roots inthe upper portion of the Urías estuary, Mazatlán, SEGulf of California, Mexico. Females were maintainedin individual aquaria until larvae hatched. Each batchof larvae was reared in a 1-gal container at a salinity of36 and temperature 26–28EC, with constant aerationand a 12:12 light:dark photoperiod. For the first twodays zoeas were fed with a mixture of rotifers(Brachionus spp.) and nauplii of Artemia; later thelarvae were fed only with nauplii of Artemia until theappearance of megalopas.
The presence of new zoeal stages or megalopas wasdeterminated by constant sampling. Larvae were fixedin 4% formaldehyde and preserved in 70% ethanol. Aminimum of five larvae for each zoeal stage and mega-lopa were used for measurements, setal counts anddrawings. Dissections were made under an OlympusSZ40 stereomicroscope, and figures were drawn with
a Leitz Wetzlar Ortholux II microscope, equipped witha camera lucida. All setal counts are from proximal todistal and from endopod to exopod according to Clarket al. (1998). The plumose natatory setae of the firstand second maxillipeds and the telson forks weredrawn in truncated form. Measurements include cara-pace length (CL) from the anterior margin of the orbitto posterior carapace margin, dorsal-spine length (DS)from base to tip, rostral spine length (RS) from base totip, and dorsal spine to rostral spine (DS–RS) from thetip of the dorsal spine to the tip of the rostral spine. Formegalopas, CL was measured from the rostrum tip tomiddle posterior carapace margin. Parental femalesand larval series were deposited in the crustaceanreference collection (EMU-6357) at the Unidad Mazat-lán, UNAM, Sinaloa, Mexico.
Results
No prezoeal stage was observed. Four zoeal stagesand a single megalopa were obtained. Larval develop-ment from hatching to megalopa lasted 14 days. Timeelapsed after hatching for each larval stage, and bodymeasurements are summarized in Table 1. The firstzoeal stage is described in detail; description of thesubsequent stages only includes the morphologicalchanges.
The larval description for A. mirafloresensis (Abeleand Kim, 1989) (Figs. 1–6) is given below.
Zoea I (Fig. 1)Carapace (Fig. 1A,B): dorsal spine long, distally
curved; rostral spine straight, tapering, slightly shorterthan antennal protopod, without spines; cephalothoraxsmooth, slightly globose without lateral spines butlateral protuberance; anterodorsal setae absent; a pairof posterodorsal setae present; ventral margin withoutsetae, ventrolateral margin, below eye, with acute pro-jection, not spiniform; eyes sessile.
Antennule (Fig. 1C): uniramous; endopod absent;exopod stout and conical, with 4 terminal aesthetascsof different lengths and thicknesses and 1 shortterminal seta.
Antenna (Fig. 1D): protopod length subequal torostrum, smooth, tapering distally, slightly dilated attip; endopod absent, exopod very reduced with 1 apicalseta.
Mandible: Palp absent.Maxillule (Fig. 1E): epipod seta absent; coxal
endite with 7 setae; basial endite with 4 plumodenti-culate setae and 1 sparsely plumose seta; endopod 2-segmented, proximal segment short with 1 seta; distal

J. Salgado-Barragán and M. Ruiz-Guerrero / IRD 47 (2005) 133–145 135
Fig. 1. Zoea I of Acantholobulus mirafloresensis (Abele and Kim, 1989). A, lateral view; B, ventral view (mouthparts,abdomen and appendages omitted); C, antennule; D, antenna; E, maxillule; F, maxilla; G, first maxilliped; H, secondmaxilliped; I, abdomen and telson; J, fork of telson in detail. All scale bars = 0.1 mm.
J. Salgado-Barragán and M. Ruiz-Guerrero / IRD 47 (2005) 133–145136
Table 1Time of the first appearance (in days) of each larval stage of Acantholobulus mirafloresensis and measurement range inmillimeters (± standard deviation) of larvae at different development stages
Stage Time elapsedafter hatching
CL DS RS RS to DS
Zoea I 0 0.37 ± 0.02 0.39 ± 0.03 0.45 ± 0.02 1.16 ± 0.03Zoea II 5 0.46 ± 0.03 0.47 ± 0.02 0.62 ± 0.04 1.4 ± 0.04Zoea III 8 0. 53 ± 0.03 0.6 ± 0.03 0.75 ± 0.03 1.9 ± 0.05Zoea IV 11 0.69 ± 0.05 0.72 ± 0.04 0.84 ± 0.05 2.22 ± 0.04Megalopa 14 0.79 ± 0.03 — — —
C, carapace length; DS, dorsal spine; RS, rostral spine.
segment longer with 2 subterminal and 4 terminalsetae; exopod seta absent.
Maxilla (Fig. 1F): coxal endite bilobed with 4 + 4setae; basial endite bilobed with 5 + 4 setae; endopodbilobed with 3 + 5 setae; scaphognathite with 4 plu-mose setae and plumose terminal process.
First maxilliped (Fig. 1G): coxal segment with1 seta; basis with 10 ventral setae arranged in groupsof 2 or 3; endopod 5-segmented with 3, 2, 1, 2, 1 + 4setae respectively; exopod partially segmented, with 4terminal natatory setae.
Second maxilliped (Fig. 1H): coxa without setae;basis with 4 setae arranged 1, 1, 1, 1; endopod 3-segmented with 1, 1, 2 + 3 setae respectively; exopodpartially segmented, with 4 terminal natatory setae.
Third maxilliped: absent.Pereiopods: absent.Abdomen (Fig. 1I): 5 somites, somites 2 and 3 each
with 1 pair of dorsolateral processes, those on somite 2distinctly larger; somites 2–5 each with 1 pair ofminute posterodorsal setae; pleopod buds absent.
Telson (Fig. 1I–J): trapezoidal, with long slenderforks, recurved dorsally at tip, each fork with twosubparallel minute lateral spines; 1 large curved spineon inner margin of fork; posterior margin with 3 pairsof stout spinulate setae.
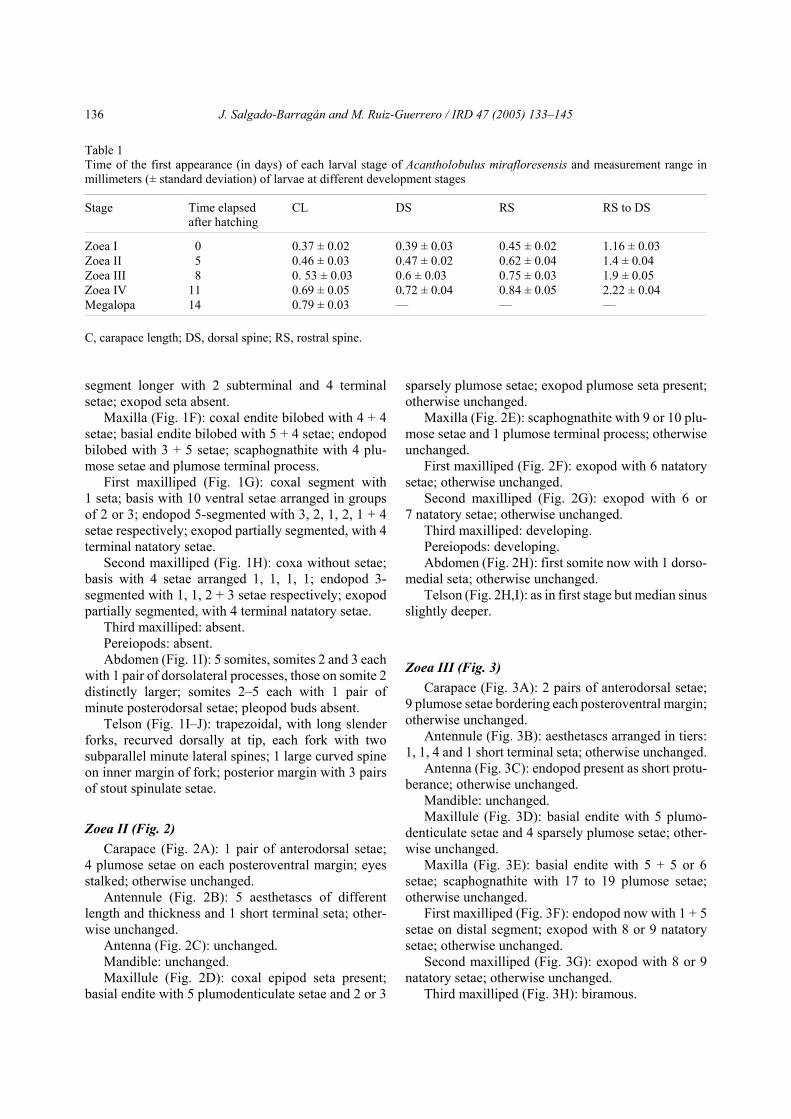
Zoea II (Fig. 2)Carapace (Fig. 2A): 1 pair of anterodorsal setae;
4 plumose setae on each posteroventral margin; eyesstalked; otherwise unchanged.
Antennule (Fig. 2B): 5 aesthetascs of differentlength and thickness and 1 short terminal seta; other-wise unchanged.
Antenna (Fig. 2C): unchanged.Mandible: unchanged.Maxillule (Fig. 2D): coxal epipod seta present;
basial endite with 5 plumodenticulate setae and 2 or 3
sparsely plumose setae; exopod plumose seta present;otherwise unchanged.
Maxilla (Fig. 2E): scaphognathite with 9 or 10 plu-mose setae and 1 plumose terminal process; otherwiseunchanged.
First maxilliped (Fig. 2F): exopod with 6 natatorysetae; otherwise unchanged.
Second maxilliped (Fig. 2G): exopod with 6 or7 natatory setae; otherwise unchanged.
Third maxilliped: developing.Pereiopods: developing.Abdomen (Fig. 2H): first somite now with 1 dorso-
medial seta; otherwise unchanged.Telson (Fig. 2H,I): as in first stage but median sinus
slightly deeper.
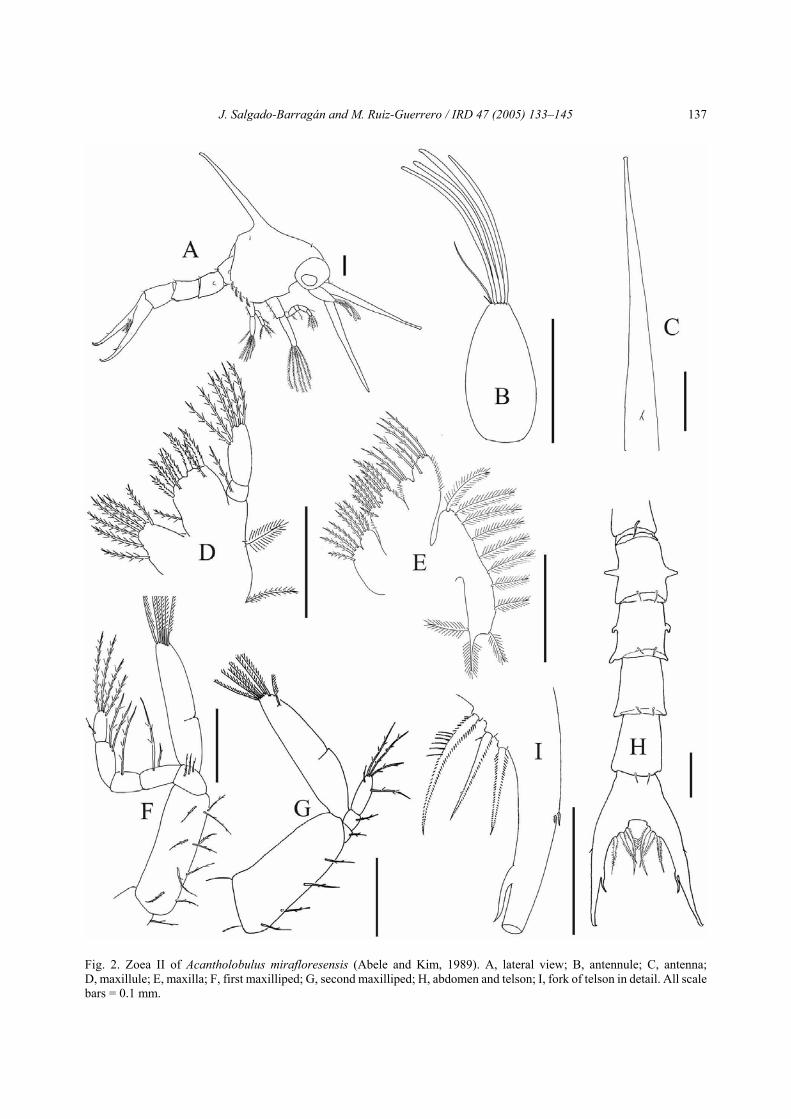
Zoea III (Fig. 3)Carapace (Fig. 3A): 2 pairs of anterodorsal setae;
9 plumose setae bordering each posteroventral margin;otherwise unchanged.
Antennule (Fig. 3B): aesthetascs arranged in tiers:1, 1, 4 and 1 short terminal seta; otherwise unchanged.
Antenna (Fig. 3C): endopod present as short protu-berance; otherwise unchanged.
Mandible: unchanged.Maxillule (Fig. 3D): basial endite with 5 plumo-
denticulate setae and 4 sparsely plumose setae; other-wise unchanged.
Maxilla (Fig. 3E): basial endite with 5 + 5 or 6setae; scaphognathite with 17 to 19 plumose setae;otherwise unchanged.
First maxilliped (Fig. 3F): endopod now with 1 + 5setae on distal segment; exopod with 8 or 9 natatorysetae; otherwise unchanged.
Second maxilliped (Fig. 3G): exopod with 8 or 9natatory setae; otherwise unchanged.
Third maxilliped (Fig. 3H): biramous.
J. Salgado-Barragán and M. Ruiz-Guerrero / IRD 47 (2005) 133–145 137
Fig. 2. Zoea II of Acantholobulus mirafloresensis (Abele and Kim, 1989). A, lateral view; B, antennule; C, antenna;D, maxillule; E, maxilla; F, first maxilliped; G, second maxilliped; H, abdomen and telson; I, fork of telson in detail. All scalebars = 0.1 mm.
J. Salgado-Barragán and M. Ruiz-Guerrero / IRD 47 (2005) 133–145138
Fig. 3. Zoea III of Acantholobulus mirafloresensis (Abele and Kim, 1989). A, lateral view; B, antennule; C, antenna;D, maxillule; E, maxilla; F, first maxilliped; G, second maxilliped; H, third maxilliped; I, pereipods; J, abdomen and telson;K, fork of telson in detail. All scale bars = 0.1 mm.

J. Salgado-Barragán and M. Ruiz-Guerrero / IRD 47 (2005) 133–145 139
Fig. 4. Zoea IV of Acantholobulus mirafloresensis (Abele and Kim, 1989). A, lateral view; B, antennule; C, antenna;D, maxillule; E, maxilla; F, first maxilliped; G, second maxilliped; H, third maxilliped; I, first and second pereiopods;J, abdomen and telson; K, fork of telson in detail. All scale bars = 0.1 mm.

J. Salgado-Barragán and M. Ruiz-Guerrero / IRD 47 (2005) 133–145140
Fig. 5. Megalopa of Acantholobulus mirafloresensis (Abele and Kim, 1989). A, carapace, dorsal view; B, antennule;C, antenna; D, mandible; E, maxillule; F, maxilla; G, first maxilliped; H, second maxilliped; I, third maxilliped. All scalebars = 0.1 mm.
Pereiopods (Fig. 3I): rudimentary, undifferentiatedinto segments; cheliped bilobed.
Abdomen (Fig. 3J): Six somites; first somite with 3dorsal setae, somites 2–5 with pairs of minute dorso-lateral setae; somite 6 length approximately half of that
of somites 4 and 5; pleopod buds now present onsomites 2–6.
Telson (Fig. 3J,K): 1 additional pair of inner shortsetae within medial sinus; otherwise unchanged.

J. Salgado-Barragán and M. Ruiz-Guerrero / IRD 47 (2005) 133–145 141
Zoea IV (Fig. 4)Carapace (Fig. 4A): 4 pairs of anterodorsal setae;
9 plumose setae bordering on each posteroventralmargin plus 3 longer plumose setae on anteroventralmargin.
Antennule (Fig. 4B): aesthetascs arranged in tiers:2, 5, 5; endopod bud present.
Antenna (Fig. 4C): endopod larger than in zoea III.Mandible: palp present as a small bud.Maxillule (Fig. 4D): coxal endite with 9 or10 setae;
basial endite with 6 plumodenticulate setae and 5sparsely plumose setae; otherwise unchanged.
Maxilla (Fig. 4E): coxal endite with 5 + 4 setae;basial endite with 6 + 7 setae; scaphognathite with 24to 27 plumose setae; otherwise unchanged.
First maxilliped (Fig. 4F): coxa with 2 setae;exopod with 9 natatory setae.
Second maxilliped (Fig. 4G): exopod with 10 or 11natatory setae.
Third maxilliped (Fig. 4H): epipod now present;arthrobranch gill present; endopod and exopoddeveloping.
Pereiopods (Fig. 4I): with incomplete segmentaldifferentiation.
Abdomen (Fig. 4J): pleopod buds larger than inzoea III; otherwise unchanged.
Telson (Fig. 4J,K): 1 additional pair of inner shortsetae within medial sinus; otherwise unchanged.
Megalopa (Figs. 5 and 6)Carapace (Fig. 5A): Longer than wide; frontal
region projected; rostrum blunt; sharp anterolateralinwardly curved horns; eyes stalked, with 4 or 5 setaeon peduncle; 3 or 4 pairs of setae lateral to rostrum;about 44 dorsal setae on anterior and posterior regionsas figured; 48 to 50 plumose setae bordering postero-lateral region.
Antennule (Fig. 5B): Peduncle 3-segmented with 3,1, 2 short + 3 long setae; endopod not segmented with1 short median and 4 long distal setae; exopod 4-segmented with 1, 0, 2, 2 setae respectively and 16closely packed aesthetascs on segments 2–4; segment2 with 4 aesthetascs, number of aesthetascs on seg-ments 3 and 4 could not be clearly determined.
Antenna (Fig. 5C): 11-segmented, with 2, 1, 1, 0, 0,3, 0, 4, 0, 4, 4 setae respectively.
Mandible (Fig. 5D): palp 2-segmented with 0, 8setae.
Maxillule (Fig. 5E): protopod with 2 long setae;coxal endite with 15 setae; basial endite with 2 longlateral setae and 19 plumodenticulate and scarcelysetulated setae; endopod 2-segmented, segment 1 with
2 long setae, segment 2 with 1 long setae and 2 shortterminal setae.
Maxilla (Fig. 5F): coxal endite with 7 + 5 setae;basial endite with 6 + 9 setae; endopod with 3 basalplumose setae and 1 long subterminal seta; scapho-gnathite with 43 setae and 3 spines on the surface.
First maxilliped (Fig. 5G): epipod with long 5naked setae; coxal endite with 7 setae; basial enditewith 19 setae; endopod unsegmented with 4 shortsetae; exopod 2-segmented, with 2, 5 setae.
Second maxilliped (Fig. 5H): epipod with 2 ter-minal setae; coxa and basis not differentiated, with2 setae; endopod 4-segmented with 2, 1, 6 and 9 setaescarcely setulated and plumodenticulate setae respect-ively; exopod 2-segmented, proximal segment with1 medial seta, distal segment with 5 plumose setae.
Third maxilliped (Fig. 5I); epipod with 5 plumo-denticulate setae plus 14 long naked setae; coxa with7 setae; basis with 8 setae; endopod 5-segmented with16, 11, 5, 9, 9 setae; exopod 2-segmented with 0, 1 + 5setae.
Cheliped (Fig. 6A): covered with short setae asfigured; ischium with recurved hook; propod with 4 or5 irregular teeth, inner margin of dactyl sinuous.
Pereiopods (Fig. 6B–F): Covered with dispersedshort setae as shown in Fig. 6B; dactyl of pereiopods2–4 (Fig. 6C–E) with 3 strong serrate ventral spinesand 1 subterminal spine; dactyl of fifth pereiopod(Fig. 6F) with 1 dorsal and 1 subterminal serratespines, and 1 subterminal long seta.
Pleopods (Fig. 6G): present in abdominal segments2–6, the anterior the longer; endopod with 3 hookedsetae; pleopod 1 with 14 natatory setae.
Abdomen (Fig. 6H): Six somites plus telson,covered with sparse dorsal minute setae symmetricallyarranged as figured.
Telson (Fig. 6H): subquadrate with posterior anglesrounded; 4 dorsal and 4–6 marginal setae arrangedsymmetrically.
Discussion
The information obtained from larval descriptionsof brachyuran crabs could be very useful to clarifytaxonomic relationships among species and highertaxonomic levels. As more descriptions of larvaldevelopment are published in the same group, morpho-logical patterns can be detected which can help todistinguish associations between brachyuran species(see Rice, 1981; Martin, 1984; Martin et al., 1985).According to Clark and Ng (1998), the carapace spinearmature, antennal shape and the spinulation of thetelson forks can be useful information to distinguishtaxa below family level.

J. Salgado-Barragán and M. Ruiz-Guerrero / IRD 47 (2005) 133–145142
Fig. 6. Megalopa of Acantholobulus mirafloresensis (Abele and Kim, 1989). A, left cheliped; B, second pereiopod; C, dactylusof second pereiopod; D, dactylus of third pereiopod; E, dactylus of fourth pereiopod; F, dactylus of fifth pereiopod 5; G, firstpleopod; H, abdomen and telson in dorsal view; All scale bars = 0.1 mm.

J. Salgado-Barragán and M. Ruiz-Guerrero / IRD 47 (2005) 133–145 143
Table 2Comparison of morphological characters and setation formulae of zoeas II, III and IV of Acantholobulus bermudensisaccording to Martin et al. (1985), A. schmitti according to Bakker et al. (1989) and A. mirafloresensis (this work)
A. bermudensis A. schmitti A. mirafloresensisZOEA IDorsal sp to rostral sp length (mm) 1.30 1.71 1.16 ± 0.03 ZOEA IICarapace ventral margin (s) 0 5 4Basial endite of maxillule (s) 10–11 8 8Scaphognathite (s) 10 11 10First abdominal somite (s) 0 0 1ZOEA IIICarapace ventral margin (s) 6 6 9Basial endite of maxillule (s) 7–10 8 9Scaphognathite (s) 15–16 20 19Mxllip 1, distal segment of endopod (s) 1 + 4 1 + 5 1 + 5First abdominal somite (s) 0 0 3ZOEA IVCarapace ventral margin (s) 8–10 10 – 12 9 + 3 longAntennule (s) (a) 2, 5 1 – 2, 4, 3 2, 5, 5Coxal endite of maxillule (s) 9–10Basial endite of maxillule (s) 10–12 10 11Coxal endite of maxilla (s) 4 + 4–5 4 + 6 5 + 4Basial endite of maxilla (s) 5 + 7 5 + 6 6 + 7Scaphognathite (s) 21–23 30 24–27Mxllip 1, distal segment of endopod (s) 1 + 4 1+5 1+5First abdominal somite (s) 0 0 3MEGALOPAAntennal first segment (s) 2 3 2Scaphognathite (s), (sp) 40–44, 0 47, 11 43, 3Mxllip 1 coxal endite (s) 9–10 13 7Mxllip 1 epipod (s) 5–6 7 5Mxllip 2 endopod (s) 1–2, 1, 6, 8–9 3, 1, 7, 9 2, 1, 6, 9Mxllip 2 exopod (s) 2, 5–6 0, 5 1, 5Mxllip 3 endopod (s) 16–18, 8, 4–6, 8, 7–8 21, 9, 8, 8, 6 16, 11, 5, 9, 9Mxllip 3 epipod (s) 4 + 12–13 nd 5 + 14Mxllip 3 exopod (s) 2, 5–6 2, 7 0, 6Telson, dorsal, marginal (s) 4, 0 4, 0 4, 4–6
a, aesthetascs; s, setae; seg., segment; sp, spines; nd, no data; mxllip, maxilliped.
The presence of tapering antennas with a veryreduced exopod, a carapace without lateral spines anda telson with a minute pair of lateral spines, as well asone dorsal spine located posterior to the median pointof forks, are observed in all zoeal stages of A. mira-floresensis and in the first zoeal stage of A. ber-mudensis and A. schmitti (P.F. Clark, unpublisheddata). This feature clearly distinguishes this group ofthree species from other previously described xanthidlarvae. In addition, comparison between zoeas of A.mirafloresensis and other panopeid species (see Luppiet al., 2003) shows that main differences are related tothe arrangement and number of setae in mouthpartsand maxillipeds.
Within the species of Acantholobulus, morpho-logical differences among larvae increases throughouttheir development and major differences are observedin late zoeas and megalopa. Zoea I of A. miraflo-resensis, A. bermudensis and A. schmitti are similar,and the only relevant difference between them is thesize and the arrangement of the lateral spines on telson(unpublished data provided by P.F. Clark for zoea I ofA. bermudensis and A. schmitti). From zoea II to zoeaIV, differences in morphology between A. bermudensis(cf. Martin et al., 1985), A. schmitti (cf. Bakker et al.,1989) and A. mirafloresensis (present work) are mostlyrelated to the setal arrangement, such as setation on theventral side of the carapace, setation on the coxal

J. Salgado-Barragán and M. Ruiz-Guerrero / IRD 47 (2005) 133–145144
endite of the maxilla, setation of the scaphognathite,and dorsal setae on the first abdominal somite(Table 2).
In addition, the posterodorsal setae of the carapaceobserved in all zoeal stages of A. mirafloresensis werenot mentioned in the original descriptions of larvae ofA. bermudensis and A. schmitti. However, the descrip-tions by Clark (unpublished data) of the first zoealstage of these species suggest that these setae wereoverlooked in the original works. Thus, carapace setaethat appear in later zoeal stages were probably alsooverlooked by the authors and we cannot assume thatthis feature was different among the species.
The comparison of A. mirafloresensis megalopawith those of the Panopeid species, revised by Luppi etal. (2003), confirm that all those species are phylo-genetically related. Morphological features present inA. mirafloresensis megalopa (e.g., anterolateral hornsin carapace, ischium hook on the first pereiopod andsetae formulas in mouthparts and maxillipeds) are alsoobserved in the megalopa of those species and maindifferences are observed in scaphognatithe and maxil-liped setation and in the arrangement of carapace setae(Table 2).
The larval development of A. mirafloresensisagrees with the previous Acantholobulus zoeal descrip-tion provided by Felder and Martin (2003), based onthe original description of the larvae of A. bermudensisand A. schmitti, and in the later examination of A.bermudensis larvae. Our description agrees with thestatement of these authors that the larvae of A. mira-floresensis “should resemble those of the two otherspecies of Acantholobulus” and supports the newcombination of P. mirafloresensis to this new genus, assuggested by adult morphology. Moreover, morpho-logical differences of Acantholobulus with the rest ofPanopeidae support the idea that this genus probablydiverged from the main group of Panopeidae, as sug-gested by Schubart et al. (2000) for A. bermudensis, onthe basis of molecular phylogeny.
Acknowledgements
We thank Ana Puello Cruz (CIAD, UnidadMazatlán) for laboratory assistance. Gabriela Velasco(CIAD, Unidad Mazatlán) and Domenico Voltolina(Universidad Autónoma de Sinaloa) provided live foodfor rearing the larvae. Arturo Nuñez Pastén (ICMyL,Unidad Mazatlán) assisted in the sampling of parentalfemales of A. mirafloresensis. Samuel Gómez, GermánRamirez Reséndiz and Sandra Castañeda gave tech-nical advice. Special thanks to Darryl L. Felder, PaulF. Clark, and an anonymous reviewer for their criticalsuggestions to the manuscript and to Marcelo García
Guerrero and Albert van der Heiden for their valuablecomments to the early version of this manuscript.
ReferencesAbele, L.G. and Kim, W., The decapod crustaceans of the
Panama Canal. Smith. Contrib. Zool., 482 (1989) 1–50.Bakker, C., de, Montú, M., Anger, K. and Loureiro
Fernandes, L., Larval development of Hexapanopeusschmitti Rathbun, 1930 (Decapoda, Brachyura, Xan-thidae) reared in the laboratory. Nerítica, 4 (1989) 137–164.
Clark, P.F., Calazans, D.K. and Pohle, G.W., Accuracy andstandardization of brachyuran larval descriptions. Invert.Reprod. Develop., 33 (1998) 127–144.
Clark, P.F. and Ng, P.K.L., The larval development of thepoisonous mosaic crab, Lophozozymus pictor (Fabricius,1798) (Crustacea, Decapoda, Brachyura, Xanthidae,Zosiminae), with comments on familial characters forfirst stage zoeas. Zoosistema, 20 (1998) 201–220.
Félix-Pico, E.F., Holguin Quiñones, O.E., Campos, E. andSalgado-Barragán, J., Cangrejos (Decapoda: Brachyura)de los sistemas lagunares con mangles de la costa orientalde Baja California Sur. In: Contributions to the study ofthe East Pacific Crustaceans [Contribuciones al estudiode los Crustáceos del Pacífico Este], Vol. 2, M.E.Hendrickx (ed.), Instituto de Ciencias del Mar y Limno-logía, UNAM, México, 2003, pp. 191–203.
Felder, D.L. and Martin, J.W., Establishment of a new genusfor Panopeus bermudensis Benedict and Rathbun, 1891and several other xanthoid crabs from the Atlantic andPacific oceans (Crustacea: Decapoda: Xanthoidea). Proc.Biol. Soc. Wash., 116 (2003) 438–452.
García-Guerrero, M.U., Rodríguez, A., Cuesta, J.A. andHendrickx, M.E., The complete larval development ofEurypanopeus canalensis Abele and Kim, 1989 (Crus-tacea: Brachyura: Panopeidae) described from laboratoryreared material. Sci. Mar., 69 (2005) in press.
Guinot, D., Principes d’une classification évolutive desCrustacés Décapodes Brachyoures. Bull. biol. Fr. Belg.,112 (1978) 211–279.
Hendrickx, M.E., A new genus and species of “goneplacid-like” brachyuran crab (Crustacea: Decapoda) from theGulf of California, Mexico, and a proposal for the use ofthe family Pseudorhombiliidae Alcock, 1900. Proc. Biol.Soc. Wash., 111 (1998) 634–644.
Luppi, T.A., Rodríguez, A. and Spivak, E.D., Larval mor-phology of the southwestern Atlantic mud crab Panopeusmeridionalis (Decapoda: Brachyura: Panopeidae)described from laboratory-reared material. J. Crust. Biol.,23 (2003) 920–935.
Martin, J.W., Notes and bibliography on the larvae ofxanthid crabs, with a key to known xanthid zoeas of thewestern Atlantic and Gulf of Mexico. Bull. Mar. Sci., 34(1984) 220–239.
Martin, J.W. and Abele, L.G., Notes on the male pleopodmorphology in the brachyuran crab family PanopeidaeOrtmann, 1893, sensu Guinot (1978) (Decapoda). Crus-taceana, 50 (1986) 182–198.

J. Salgado-Barragán and M. Ruiz-Guerrero / IRD 47 (2005) 133–145 145
Martin, J.W., Rodríguez, A. and Zimmerman, T.L., Mor-phological description of the first zoeal stage of thetropical xanthid crab Panopeus purpureus Lockington,1877 (Crustacea, Decapoda, Brachyura, Xanthidae).Cienc. Mar., 24 (1998) 225–232.
Martin, J.W., Truesdale, F.M. and Felder, D.L., Larvaldevelopment of Panopeus bermudensis Benedict andRathbun, 1891 (Brachyura, Xanthidae) with notes onzoeal characters in xanthid crabs. J. Crust. Biol., 5 (1985)84–105.
Rice, A.L., Crab zoeae and brachyuran classification: a re-appraisal. Bull. Br. Mus. Nat. Hist. (Zool.), 40 (1981)287–296.
Salgado-Barragán, J. and Hendrickx, M.E., Decapod
crustaceans from the Pacific coast of Mexico, includingnew records, taxonomic remarks and ecological data.Rev. Biol. Trop., 44/45 (1996) 680–683.
Salgado-Barragán, J. and Hendrickx, M.E., Panopeid crabs(Crustacea: Brachyura: Panopeidae) associated to proproots of Rhizophora mangle L. in a tropical coastallagoon of the SE Gulf of California, Mexico. In: ModernApproaches to the Study of Crustacea, E. Escobar-Briones and F. Alvarez (eds.), Academic Press, NewYork, 2002, pp. 163–169.
Schubart, C.D., Neigel, J.E. and Felder, D.L., Molecularphylogeny of mud crabs (Brachyura: Panopeidae) fromthe northwestern Atlantic and the role of morphologicalstasis and convergence. Mar. Biol., 137 (2000) 11–18.