tetanus toxin fragment c binds to a protein present in neuronal cell lines and motoneurons
TRANSCRIPT

Tetanus Toxin Fragment C Binds to a Protein Present inNeuronal Cell Lines and Motoneurons
Judit Herreros, Giovanna Lalli, *Cesare Montecucco, and Giampietro Schiavo
Molecular Neuropathobiology Laboratory, Imperial Cancer Research Fund, London, England; and*Centro CNR Biomembraneand Dipartimento di Scienze Biomediche, Universita` di Padova, Padova, Italy
Abstract: Tetanus neurotoxin is one of the most powerfulprotein toxins known, acting in vivo at femtomolar doses.Two main factors determine its high potency: a proteaseactivity restricted to a single intracellular substrate and itsabsolute neurospecificity. Whereas the enzymatic prop-erties of tetanus toxin have been thoroughly defined, thenature of its neuronal receptor(s) and their involvement inthe intracellular trafficking of tetanus toxin are poorlyunderstood. Using binding and crosslinking experiments,we report here on the characterisation of an N-glycosy-lated 15-kDa interacting protein, which behaves as anintegral membrane protein. This putative receptor specif-ically interacts with the binding domain (fragment C) oftetanus toxin and not with several related botulinum neu-rotoxins in spinal cord motoneurons and neuronal-likecell lines. Sialic acid-specific lectins antagonise the bind-ing of tetanus toxin to the cell surface and to the 15-kDaprotein, supporting the central role of sialic acid residuesin the recognition process. Altogether, these results indi-cate the existence of a neuronal protein receptor fortetanus toxin whose identification is likely to constitute akey step in the analysis of the molecular machinery in-volved in the toxin internalisation and retrograde trans-port. Key Words: Crosslinking—Motoneurons—PC12cells—Receptor—Tetanus neurotoxin.J. Neurochem. 74, 1941–1950 (2000).
Tetanus toxin (TeNT) is a bacterial protein toxin thatblocks irreversibly neurotransmitter release. Togetherwith the botulinum neurotoxins (BoNT/A–G), TeNT be-longs to the clostridial neurotoxin (CNT) family. CNTsshare a common structure composed of a heavy (H; 100kDa) and a light (L; 50 kDa) chain linked by a disulphidebond (Herreros et al., 1999). The H chain mediates thebinding and internalisation into neurons, whereas the Lchain is responsible for the intracellular metalloproteaseactivity that selectively cleaves synaptic proteins, conse-quently inhibiting neurotransmission (Niemann et al.,1994; Montecucco and Schiavo, 1995). All CNTs bind tothe neuromuscular junction (NMJ), but their intracellularactions take place at different levels of the nervoussystem. The proteolytic activity of TeNT occurs withininhibitory interneurons of the spinal cord, thereby caus-
ing the typical spastic paralysis of tetanus. In contrast,BoNTs act at peripheral cholinergic terminals, causingflaccid paralysis (Herreros et al., 1999).
CNTs are the most powerful toxins known (Gill,1982). Their potency is due to both their enzymaticactivity and their selective high-affinity binding to neu-rons. Early studies demonstrated the lack of competitionamong TeNT and several BoNT serotypes, suggestingthe existence of different cellular receptors for CNTs(Habermann and Dreyer, 1986). This could also explainthe differential sorting of these toxins at the NMJ, whichdetermines the recruitment of TeNT but not BoNTs inthe retrograde transport pathway (Bizzini et al., 1977;Schwab et al., 1977; Black and Dolly, 1986a,b).
Polysialogangliosides bind CNTs with aKD in thenanomolar range on the carboxy-terminal fragment ofthe H chain (fragment C or HC) (for review, see Halpernand Neale, 1995). The relatively low-affinity binding topolysialogangliosides suggests, however, that these gly-colipids are not the only CNT receptors. Protein recep-tor(s) may participate in CNT binding, as suggested bythe observation of protease-sensitive TeNT-binding sitesin different neuronal preparations (Pierce et al., 1986;Yavin and Nathan, 1986; Parton et al., 1988; Bakry et al.,1991b). Alternatively, a binary complex containing both
Received November 3, 1999; revised manuscript received December21, 1999; accepted January 3, 2000.
Address correspondence and reprint requests to Dr. G. Schiavo atMolecular Neuropathology Laboratory, Imperial Cancer ResearchFund, 44 Lincoln’s Inn Fields, London WC2A 3PX, U.K. E-mail:[email protected]
Abbreviations used:BoNT, botulinum neurotoxin; BSA, bovine serumalbumin; BSOCOES, bis[2(succinimidyloxycarbonyloxy)ethyl]sulphone;CNT, clostridial neurotoxin; DSS, disuccinimidyl suberate; DST, di-succinimidyl tartrate; H, heavy chain; HC, carboxy-terminal fragmentof H chain or fragment C; L, light chain; LFA,Limax flavusagglutinin;MA, Maackia amurensis;MES, 2-(N-morpholino)ethanesulphonicacid; MN, motoneuron; NeuNAc,N-acetylneuraminic acid; NGF,nerve growth factor; NMJ, neuromuscular junction; p15, 15-kDa pu-tative receptor protein for TeNT; PC12, rat pheochromocytoma; SDS-PAGE, sodium dodecyl sulphate–polyacrylamide gel electrophoresis;sulpho-EGS, ethylene glycolbis(sulphosuccinimidyl succinate); TeNT,tetanus neurotoxin; TV,Triticum vulgaris; TX-114, Triton X-114;VSV-G, vesicular stomatitis virus G protein.
1941
Journal of NeurochemistryLippincott Williams & Wilkins, Inc., Philadelphia© 2000 International Society for Neurochemistry

protein and polysialogangliosides could represent thehigh-affinity receptor (Montecucco, 1986). Lately, a fewcandidate proteins have been suggested as putative re-ceptors for CNTs. BoNT/A and TeNT were shown tobind to glycosylated proteins in brain synaptosomes(Schengrund et al., 1992). However, these proteins, iden-tified as synapsin-I and adducin (Schengrund et al.,1996), are unlikely to represent physiological receptorsbecause they lack extracellular domains. More recently,synaptotagmins I and II have been shown to bind toBoNT/A, B, and E in vitro (Nishiki et al., 1994, 1996a,b;Li and Singh, 1998). TeNT was suggested to interactwith the luminal domain of a synaptic vesicle protein incentral synapses (Matteoli et al., 1996). Crosslinkingexperiments performed on nerve growth factor (NGF)-differentiated PC12 cells with native TeNT suggested thepresence of a putative 20-kDa receptor protein for thisneurotoxin (Schiavo et al., 1991). However, no furthercharacterisation of this protein has yet been provided.
Here we used a recombinant version of TeNT HC as aprobe to study this putative protein receptor(s) for TeNT.HC is tagged at the amino-terminus with the vesicularstomatitis virus G protein (VSV-G) epitope and containsa phosphorylation site for protein kinase A radiolabel-ling. This avoids the direct chemical modification ofresidues within the HC and minimises loss of biologicalactivity. We performed binding and crosslinking exper-iments on a variety of neuronal cell types, includingNGF-differentiated PC12 cells and primary cultures ofspinal cord motoneurons (MNs), the key cell type forTeNT entry into the organism. We were able to demon-strate that recombinant TeNT HC binds to these cells andspecifically interacts with a 15- to 20-kDaN-glycosy-lated TeNT-binding protein (p15). p15 behaves as anintegral membrane protein and displays very similar fea-tures in neuronal-like cell lines and MNs, suggesting thatit may constitute a high-affinity protein receptor forTeNT.
MATERIALS AND METHODS
MaterialsTeNT, native TeNT HC, and BoNT/A were purified as
previously described (Schiavo and Montecucco, 1995). Recom-binant BoNT HC fragments were obtained as shown elsewhere(Lalli et al., 1999).Limax flavuslectin was from EY Labora-tories (U.S.A.),Triticum vulgaris (TV), Maackia amurensis(MA), andSambucus nigrawere from Sigma, and MA lectin II(MA-II) from Vector (U.S.A.).
Expression and radiolabelling of TeNT HCfragment
A recombinant TeNT HC (residues 855–1,314) tagged at theamino-terminus with the VSV-G epitope (Soldati and Perriard,1991) and containing the consensus sequence for protein kinaseA phosphorylation (Kaelin et al., 1992) was expressed inEsch-erichia coli as glutathioneS-transferase fusion protein andpurified on a HiTrap column (Amersham-Pharmacia Biotech,U.K.) (Lalli et al., 1999). For protein kinase A phosphorylation,5 mg of HC was incubated (5 min, 30°C) with 100mCi of
[g-32P]ATP (10 mCi/ml, 3,000 Ci/mmol; Amersham-Pharma-cia Biotech) in reaction buffer [50 mM 2-(N-morpholino)eth-anesulphonic acid (MES; pH 6.9), 10 mM MgCl2, 1 mMdithiothreitol, 0.5 mM EDTA, 1 mg/ml ovalbumin] togetherwith 0.4 mU of the catalytic subunit of protein kinase A (4mU/ml; Roche Boehringer-Mannheim, Germany) diluted in 5mM MES (pH 6.9), 0.5 mM dithiothreitol, 0.2 mM EDTA, and0.5 mg/ml ovalbumin. Free [g-32P]ATP was separated from thelabelled protein on PD-10 columns (Amersham-PharmaciaBiotech) equilibrated with 0.1% ovalbumin in phosphate-buff-ered saline. The fraction containing the maximal amount ofradiolabelled protein was stored at280°C. Specific activity ofthe radiolabelled HC was on average;2,000 Ci/mmol. Iodin-ated native HC was labelled with the Bolton and Hunter reagent(Amersham-Pharmacia Biotech) to a specific activity of 3–5mCi/mg as previously described (Schiavo et al., 1991).
Cell cultureRat pheochromocytoma (PC12) cells (obtained from Dr.
T. B. Rogers, University of Maryland, College Park, MD,U.S.A.) were cultured in Dulbecco’s modified Eagle’s mediumcontaining 7.5% foetal calf serum and 7.5% horse serum(Sigma). Cells were grown at 37°C in a humidified atmospherecontaining 10% CO2 on plastic dishes precoated with rat col-lagen. For crosslinking experiments, cells were seeded in 12-well plates (Costar-Corning, U.S.A.) at a density of 25,000cells/well. After 24 h, the medium was supplemented with 75ng/ml NGF (7S; Alexis, U.S.A.). Medium was changed every3–4 days, and cells were used after 7 days of NGF treatment.
Rat spinal cord MNs were purified from embryonic day 14rat embryos as previously described (Henderson et al., 1995).In brief, the ventral part of the spinal cords was dissected anddissociated with 0.025% trypsin (GibcoBRL, U.S.A.) for 10min at 37°C. After tissue trituration, cells were collected bycentrifugation through a bovine serum albumin (BSA) cushion.Immunopanning was performed by using dishes coated withIgG192 (Chandler et al., 1984). This antibody recognises thep75 low-affinity NGF receptor, which is specifically expressedby rat MNs at embryonic day 14 (Yan and Johnson, 1988).Purified MNs were seeded in 12-well plates or on glass cover-slips coated with polyornithine and laminin (Sigma) at a den-sity of 125,000 cells/well. Cultures were maintained in a hu-midified 7.5% CO2 incubator at 37°C in Neurobasal medium(Life Technologies GibcoBRL, U.S.A.) containing B27 supple-ment (Life Technologies GibcoBRL), 2% horse serum, 0.5 mML-glutamine, 25mM 2-mercaptoethanol (Fluka, U.K.), 10 ng/mlrat ciliary neurotrophic factor, and 100 pg/ml rat glial cellline-derived neurotrophic factor (both from R&D Systems,U.S.A.). L-Glutamate (25mM) was added to the medium onlyfor the first 4 days in culture (Arce et al., 1998).
Mouse spinal cord cells were isolated from 14-day foetalmice and cultured as described elsewhere (Williamson et al.,1996; Lalli et al., 1999). Experiments were performed after 2weeks in culture.
Human neuroblastoma cells from a primary explant wereobtained from Dr. R. Basso (Department of Paediatric Medi-cine, University of Padova, Padova, Italy) and cultured oncollagen in RPMI-1640 containing 2% sodium bicarbonate,10% foetal calf serum, and 50mg/ml gentamicin (GibcoBRL)at 37°C in a 5% CO2 atmosphere. Cells were seeded in 12-wellplates at a density of 75,000 cells/well. After 24 h, the mediumwas supplemented with 10mM retinoic acid (Sigma), and cellswere used after 7 days.
J. Neurochem., Vol. 74, No. 5, 2000
1942 J. HERREROS ET AL.

Binding and crosslinking experimentsBinding and internalisation of TeNT and its fragments
(40–80 nM) were performed as described elsewhere (Lalliet al., 1999). Toxin bound for 1 h at 4°C wasdetected withoutcell permeabilisation using a purified monoclonal antibodyagainst the VSV-G epitope (Soldati and Perriard, 1991) orrabbit polyclonal anti-TeNT antibody (Schiavo et al., 1991). Tomonitor internalisation, cells were incubated 1 h at37°C andthen treated with buffers containing 0.1% Triton X-100 (Lalliet al., 1999). Texas Red-coupled (Amersham-Pharmacia Bio-tech) or Alexa 488-coupled (Molecular Probes, U.S.A.) sec-ondary antibodies were then used following manufacturers’specifications.
For binding and crosslinking experiments, cells were cooledon ice and washed with ice-cold Hanks’ buffer (1.26 mM CaCl2,5.36 mM KCl, 0.44 mM KH2PO4, 0.81 mM MgSO4, 136 mMNaCl, 0.42 mM Na2HPO4, 6.1 mM glucose, 20 mM HEPES–NaOH, pH 7.4). Cells were incubated with32P-HC (200–300pM) in 0.2% BSA–Hanks’ for 2 h onice. In competitionassays,cells were preincubated for 20 min with 2mM cold toxins orwith the indicated amount of other substances, before addingthe32P-HC. After binding, cells were washed with Hanks’bufferand bound HC was crosslinked (10 min, 4°C) with 0.22 mMbis[2(succinimidyloxycarbonyloxy)ethyl]sulphone(BSOCOES;Pierce, U.S.A.) in Hanks’ buffer or with other crosslinkers atthe same concentration. The reaction was stopped by removingthe crosslinker and by adding 20 mM glycine in Hanks’ buffer.Cells were then solubilised in 4% octyl-b-D-glucopyranoside(Roche Boehringer-Mannheim) in Hanks’ buffer containing thefollowing protease inhibitors: 1 mM iodoacetamide, 100mMphenylmethanesulphonyl fluoride (Fluka, U.K.), 1 mM benza-midine (Fluka), 1mg/ml aprotinin, and 1mg/ml leupeptin(Sigma). After 5 min, detergent was diluted up to 0.8%. Solu-bilised samples were precipitated with 6.5% trichloroaceticacid and prepared for sodium dodecyl sulphate (SDS)–polyac-rylamide gel electrophoresis (SDS-PAGE). For the deglycosy-lation assay, after binding, crosslinking, and partial denatur-ation in 8% SDS (3 min, 100°C), selected samples were treatedwith 6.5 U of N-glycosidase F (Roche Boehringer-Mannheim)in 1% Nonidet P-40 (Sigma), 80 mM KH2PO4 (pH 8.6), 15mM EDTA, and 4 mM b-mercaptoethanol for 2 h at37°C (1%SDS final concentration). Proteins were analysed in 6–12%acrylamide gradient gels (Laemmli, 1970) and by autoradiog-raphy. Quantitative measurements were performed using thePhosphorImager system (Molecular Dynamics, U.S.A.), andbackground readings for each lane were subtracted to calculateboth the total 32P-HC binding and the amount of 65-kDacrosslinking band.
Membrane stripping and Triton X-114 (TX-114)partitioning
After binding and crosslinking, cells were resuspended inHanks’ buffer containing protease inhibitors and diluted withan equal volume of concentrated stripping buffer [0.2MNa2CO3 (pH 11.2; with NaHCO3), 2 M KCl, 4 mM EDTA,0.05% saponin]. Alternatively, cells were mixed with 4M KCladded under constant stirring up to 1M final concentration.Samples were incubated for 30 min at 4°C and centrifuged at100,000g for 30 min. In the TX-114 subfractionation experi-ment, cells were resuspended in 200ml of TX-114 buffer [0.1M potassium phosphate, pH 6.7, 5 mM MgCl2, 1% TX-114(previously precondensed) containing protease inhibitors].Samples were sequentially incubated on ice for 10 min, at 37°Cfor 3 min, and on ice for 10 min. Following centrifugation(3,000 g, 5 min, 4°C), the insoluble pellet was kept. The
supernatant was incubated at 37°C for 3 min and then underlaidwith 50 ml of cushion buffer (as the TX-114 buffer but con-taining 0.2M sucrose and 0.06% TX-114). After centrifugation(3,000g, 5 min, room temperature), the aqueous and detergentphases were collected. Proteins were trichloroacetic acid pre-cipitated, and pellets were washed with 80% cold acetone priorto SDS-PAGE analysis.
RESULTS
TeNT HC specifically interacts with a 15-kDaprotein in NGF-differentiated PC12 cells
To study the binding of TeNT in living cells, we firstinvestigated the binding of native or recombinant TeNTHC and TeNT holotoxin (80 nM) at 4°C to NGF-differ-entiated PC12 cells. Binding of the recombinant HC gavea punctate, discontinuous staining of the plasma mem-brane (Fig. 1A), indistinguishable from the immuno-staining patterns obtained either with TeNT holotoxin orwith the native HC fragment (Fig. 1C and D). Binding ofthe recombinant HC is specific as it was completelyabolished by pretreatment with an excess of native HC(Fig. 1B). Interaction with undifferentiated PC12 cellswas negligible (Fig. 1E) and similar to the control (Fig.1F), suggesting that the expression of TeNT acceptors inPC12 cells is dependent on NGF differentiation. Thisbinding to the PC12 cell surface is functional, because itis followed by the internalisation of TeNT HC into anendocytic compartment when the temperature is raised to37°C (Fig. 1G).
To assess the HC binding to putative protein receptors,we performed binding and chemical crosslinking of32P-labelled recombinant TeNT HC (200 pM) to intact NGF-differentiated PC12 cells. The addition of homo-bifunc-tional crosslinkers yielded an'65-kDa radioactive bandin addition to32P-HC (Fig. 2A), indicating the interactionof HC with a protein having an apparent molecular massof '15–20 kDa (p15). BSOCOES, a water-insoluble,membrane-permeable crosslinker that reacts with pri-mary amines, showed the highest efficiency in generatingthe 65-kDa product. Other homo-bifunctional crosslink-ers of similar arm length, solubility, and chemical spec-ificity, such as disuccinimidyl suberate (DSS) and disuc-cinimidyl tartrate (DST), were also capable of formingthe 65-kDa product, whereas their water-soluble forms[sulpho-BSOCOES and ethylene glycolbis(sulphosuc-cinimidyl succinate) (sulpho-EGS)] were less effective(Fig. 2A). BSOCOES was therefore used throughout,unless otherwise stated. A possible explanation for thehigher efficiency of apolar crosslinkers in the stabilisa-tion of the p15/32P-HC interaction might reside in theirability to access freely the interphase between the proteincomplex and the membrane bilayer, an area inaccessibleto charged crosslinkers. In line with our finding thatundifferentiated PC12 cells do not bind TeNT HC, theappearance of the 65-kDa crosslinking product waslargely reduced when crosslinking was performed inthese conditions (Fig. 2A, lane 1), indicating that theexpression of p15 is strongly induced by NGF differen-tiation.
J. Neurochem., Vol. 74, No. 5, 2000
1943TETANUS TOXIN RECEPTOR IN RAT MOTONEURONS

Additional crosslinking products migrating at'83and 90 kDa were also obtained (Fig. 2, asterisks). Gelfiltration experiments showed that isolated TeNT HC hasthe ability to self-associate and form homodimers (un-published results). The formation upon crosslinking ofradioactive bands with a molecular mass multiple of thatof TeNT HC is therefore expected. Both the 83- and the90-kDa radioactive bands were also observed upon
crosslinking in the absence of cells (data not shown).Based on these observations, the 65-kDa band appears tobe the unique product formed after interaction of HC witha putative receptor protein in NGF-differentiated PC12cells.
In similar conditions, HC obtained by limited proteol-ysis of TeNT generated a crosslinking product identicalto that observed with the recombinant HC (Fig. 2B). Thisresult confirms that the recombinant HC behaves as thenative fragment also in the crosslinking experiments andthat it is fully competent for binding to the cell surface.
Preincubation of the cells with an excess of unlabelledrecombinant, native HC, or TeNT resulted in a stronginhibition of the 32P-HC binding and a total disappear-ance of the 65-kDa band (Fig. 3). In the case of pretreat-ment with unlabelled recombinant TeNT HC, the IC50 ofthe competition is 1806 20 nM (data not shown). Incontrast, equal amounts of recombinant HC fragments ofdifferent BoNTs (serotypes A and B) or of an unrelatedprotein (ovalbumin) did not affect significantly HC bind-ing or the appearance of the 65-kDa crosslinking product(Fig. 3). In contrast, BoNT/E HC competed weakly boththe binding of TeNT HC to the cells and the formation ofthe 65-kDa band (see Discussion). These results suggestthat the 65-kDa band represents the interaction of the HCfragment with a putative protein receptor specific forTeNT. The effect of heparin on the binding was alsotested in an attempt to highlight possible similaritiesbetween p15 and the receptor for other bacterial proteintoxins. A heparin-binding epidermal growth factor-likeprecursor is involved in the binding of diphtheria toxin tosensitive cells (Naglich et al., 1992). In addition, a hep-
FIG. 1. Binding and internalisation of TeNT and its HC fragment in NGF-differentiated PC12 cells. Recombinant HC (80 nM ) wasimmunodetected with an antibody against the VSV-G tag (A, B, and E–H), whereas TeNT (C; 80 nM ) and its native HC (D; 80 nM ) wererevealed with a polyclonal antibody raised against the holotoxin. In B, the interaction of the recombinant HC is competed by an excess(3003) of native HC. Binding of HC to undifferentiated cells (E) is indistinguishable from the control (F) where HC was omitted.Internalisation of TeNT HC was achieved by incubating NGF-differentiated PC12 cells for 1 h at 37°C with 80 nM TeNT HC (G) or without(H), followed by staining with the same anti-VSV-G antibodies. Images in G and H were collected with a confocal laser scanningmicroscope and represent z sections obtained at 1.6 mm from the substrate. Bars 5 10 mm.
FIG. 2. TeNT HC interacts with a 15-kDa protein in NGF-differ-entiated PC12 cells. A: Binding and crosslinking of 32P-recom-binant HC (200 pM ) to undifferentiated (lane 1) and to NGF-differentiated (lanes 2–7) PC12 cells in the presence or absenceof different crosslinkers (see text; sBSOCOES and sEGS, sul-pho-BSOCOES and sulpho-EGS, respectively). B: Binding andcrosslinking of 125I-native HC (300 pM ) to NGF-differentiatedPC12 cells. With either recombinant (lane 1) or native (lane 2) HC,the formation of an '65-kDa radioactive product is observedafter crosslinking (arrows). Asterisks mark the 83- and 90-kDabands (see text).
J. Neurochem., Vol. 74, No. 5, 2000
1944 J. HERREROS ET AL.
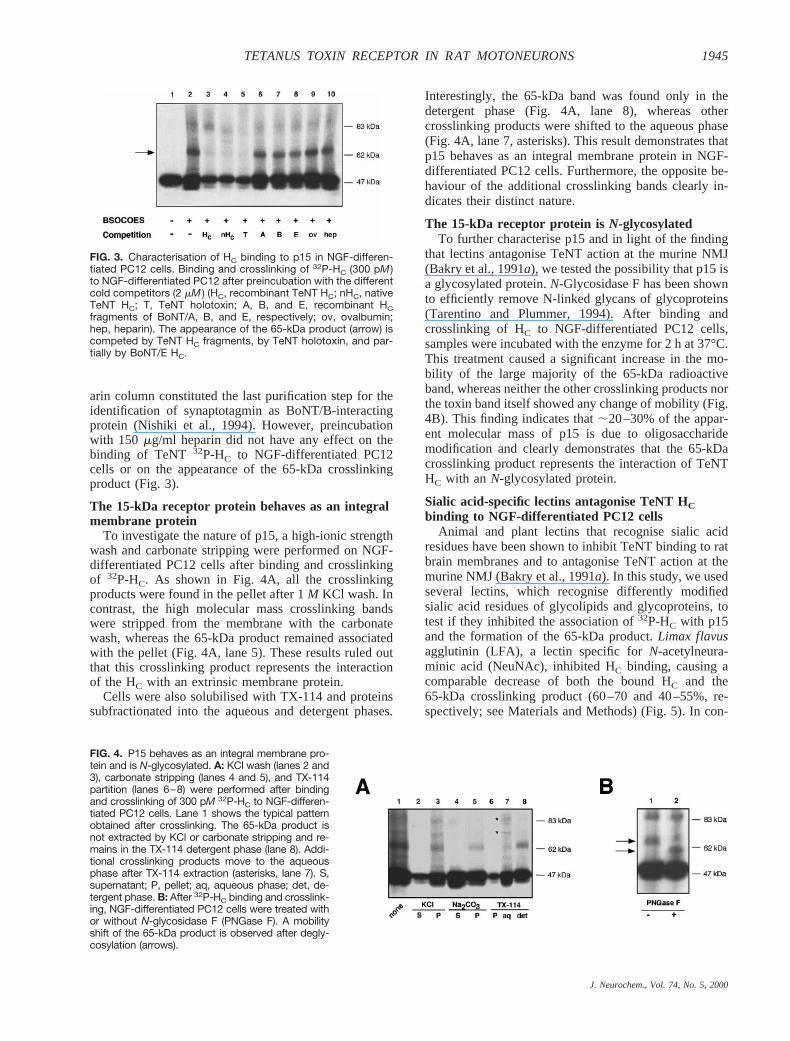
arin column constituted the last purification step for theidentification of synaptotagmin as BoNT/B-interactingprotein (Nishiki et al., 1994). However, preincubationwith 150 mg/ml heparin did not have any effect on thebinding of TeNT 32P-HC to NGF-differentiated PC12cells or on the appearance of the 65-kDa crosslinkingproduct (Fig. 3).
The 15-kDa receptor protein behaves as an integralmembrane protein
To investigate the nature of p15, a high-ionic strengthwash and carbonate stripping were performed on NGF-differentiated PC12 cells after binding and crosslinkingof 32P-HC. As shown in Fig. 4A,all the crosslinkingproducts were found in the pellet after 1M KCl wash. Incontrast, the high molecular mass crosslinking bandswere stripped from the membrane with the carbonatewash, whereas the 65-kDa product remained associatedwith the pellet (Fig. 4A, lane 5). These results ruled outthat this crosslinking product represents the interactionof the HC with an extrinsic membrane protein.
Cells were also solubilised with TX-114 and proteinssubfractionated into the aqueous and detergent phases.
Interestingly, the 65-kDa band was found only in thedetergent phase (Fig. 4A, lane 8), whereas othercrosslinking products were shifted to the aqueous phase(Fig. 4A, lane 7, asterisks). This result demonstrates thatp15 behaves as an integral membrane protein in NGF-differentiated PC12 cells. Furthermore, the opposite be-haviour of the additional crosslinking bands clearly in-dicates their distinct nature.
The 15-kDa receptor protein isN-glycosylatedTo further characterise p15 and in light of the finding
that lectins antagonise TeNT action at the murine NMJ(Bakry et al., 1991a), we tested the possibility that p15 isa glycosylated protein.N-Glycosidase F has been shownto efficiently remove N-linked glycans of glycoproteins(Tarentino and Plummer, 1994). After binding andcrosslinking of HC to NGF-differentiated PC12 cells,samples were incubated with the enzyme for 2 h at37°C.This treatment caused a significant increase in the mo-bility of the large majority of the 65-kDa radioactiveband, whereas neither the other crosslinking products northe toxin band itself showed any change of mobility (Fig.4B). This finding indicates that;20–30% of the appar-ent molecular mass of p15 is due to oligosaccharidemodification and clearly demonstrates that the 65-kDacrosslinking product represents the interaction of TeNTHC with an N-glycosylated protein.
Sialic acid-specific lectins antagonise TeNT HCbinding to NGF-differentiated PC12 cells
Animal and plant lectins that recognise sialic acidresidues have been shown to inhibit TeNT binding to ratbrain membranes and to antagonise TeNT action at themurine NMJ (Bakry et al., 1991a). In this study, we usedseveral lectins, which recognise differently modifiedsialic acid residues of glycolipids and glycoproteins, totest if they inhibited the association of32P-HC with p15and the formation of the 65-kDa product.Limax flavusagglutinin (LFA), a lectin specific forN-acetylneura-minic acid (NeuNAc), inhibited HC binding, causing acomparable decrease of both the bound HC and the65-kDa crosslinking product (60–70 and 40–55%, re-spectively; see Materials and Methods) (Fig. 5). In con-
FIG. 3. Characterisation of HC binding to p15 in NGF-differen-tiated PC12 cells. Binding and crosslinking of 32P-HC (300 pM )to NGF-differentiated PC12 after preincubation with the differentcold competitors (2 mM ) (HC, recombinant TeNT HC; nHC, nativeTeNT HC; T, TeNT holotoxin; A, B, and E, recombinant HCfragments of BoNT/A, B, and E, respectively; ov, ovalbumin;hep, heparin). The appearance of the 65-kDa product (arrow) iscompeted by TeNT HC fragments, by TeNT holotoxin, and par-tially by BoNT/E HC.
FIG. 4. P15 behaves as an integral membrane pro-tein and is N-glycosylated. A: KCl wash (lanes 2 and3), carbonate stripping (lanes 4 and 5), and TX-114partition (lanes 6–8) were performed after bindingand crosslinking of 300 pM 32P-HC to NGF-differen-tiated PC12 cells. Lane 1 shows the typical patternobtained after crosslinking. The 65-kDa product isnot extracted by KCl or carbonate stripping and re-mains in the TX-114 detergent phase (lane 8). Addi-tional crosslinking products move to the aqueousphase after TX-114 extraction (asterisks, lane 7). S,supernatant; P, pellet; aq, aqueous phase; det, de-tergent phase. B: After 32P-HC binding and crosslink-ing, NGF-differentiated PC12 cells were treated withor without N-glycosidase F (PNGase F). A mobilityshift of the 65-kDa product is observed after degly-cosylation (arrows).
J. Neurochem., Vol. 74, No. 5, 2000
1945TETANUS TOXIN RECEPTOR IN RAT MOTONEURONS
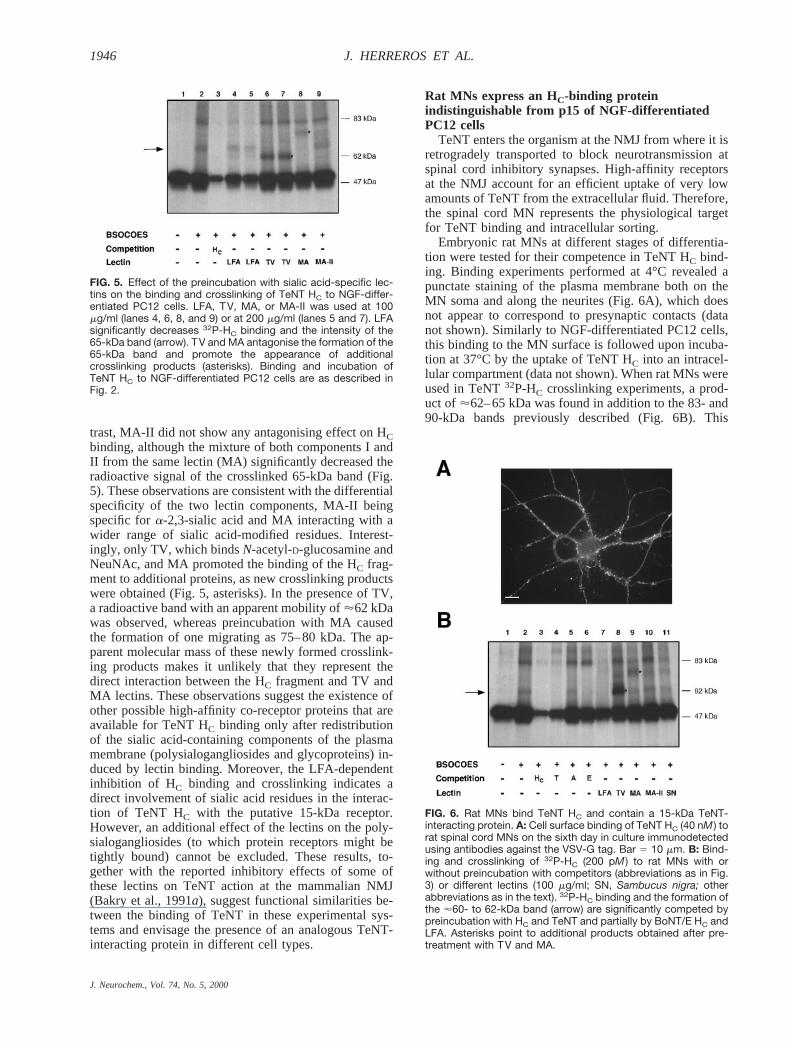
trast, MA-II did not show any antagonising effect on HCbinding, although the mixture of both components I andII from the same lectin (MA) significantly decreased theradioactive signal of the crosslinked 65-kDa band (Fig.5). These observations are consistent with the differentialspecificity of the two lectin components, MA-II beingspecific fora-2,3-sialic acid and MA interacting with awider range of sialic acid-modified residues. Interest-ingly, only TV, which bindsN-acetyl-D-glucosamine andNeuNAc, and MA promoted the binding of the HC frag-ment to additional proteins, as new crosslinking productswere obtained (Fig. 5, asterisks). In the presence of TV,a radioactive band with an apparent mobility of'62 kDawas observed, whereas preincubation with MA causedthe formation of one migrating as 75–80 kDa. The ap-parent molecular mass of these newly formed crosslink-ing products makes it unlikely that they represent thedirect interaction between the HC fragment and TV andMA lectins. These observations suggest the existence ofother possible high-affinity co-receptor proteins that areavailable for TeNT HC binding only after redistributionof the sialic acid-containing components of the plasmamembrane (polysialogangliosides and glycoproteins) in-duced by lectin binding. Moreover, the LFA-dependentinhibition of HC binding and crosslinking indicates adirect involvement of sialic acid residues in the interac-tion of TeNT HC with the putative 15-kDa receptor.However, an additional effect of the lectins on the poly-sialogangliosides (to which protein receptors might betightly bound) cannot be excluded. These results, to-gether with the reported inhibitory effects of some ofthese lectins on TeNT action at the mammalian NMJ(Bakry et al., 1991a), suggest functional similarities be-tween the binding of TeNT in these experimental sys-tems and envisage the presence of an analogous TeNT-interacting protein in different cell types.
Rat MNs express an HC-binding proteinindistinguishable from p15 of NGF-differentiatedPC12 cells
TeNT enters the organism at the NMJ from where it isretrogradely transported to block neurotransmission atspinal cord inhibitory synapses. High-affinity receptorsat the NMJ account for an efficient uptake of very lowamounts of TeNT from the extracellular fluid. Therefore,the spinal cord MN represents the physiological targetfor TeNT binding and intracellular sorting.
Embryonic rat MNs at different stages of differentia-tion were tested for their competence in TeNT HC bind-ing. Binding experiments performed at 4°C revealed apunctate staining of the plasma membrane both on theMN soma and along the neurites (Fig. 6A), which doesnot appear to correspond to presynaptic contacts (datanot shown). Similarly to NGF-differentiated PC12 cells,this binding to the MN surface is followed upon incuba-tion at 37°C by the uptake of TeNT HC into an intracel-lular compartment (data not shown). When rat MNs wereused in TeNT32P-HC crosslinking experiments, a prod-uct of '62–65 kDa was found in addition to the 83- and90-kDa bands previously described (Fig. 6B). This
FIG. 5. Effect of the preincubation with sialic acid-specific lec-tins on the binding and crosslinking of TeNT HC to NGF-differ-entiated PC12 cells. LFA, TV, MA, or MA-II was used at 100mg/ml (lanes 4, 6, 8, and 9) or at 200 mg/ml (lanes 5 and 7). LFAsignificantly decreases 32P-HC binding and the intensity of the65-kDa band (arrow). TV and MA antagonise the formation of the65-kDa band and promote the appearance of additionalcrosslinking products (asterisks). Binding and incubation ofTeNT HC to NGF-differentiated PC12 cells are as described inFig. 2.
FIG. 6. Rat MNs bind TeNT HC and contain a 15-kDa TeNT-interacting protein. A: Cell surface binding of TeNT HC (40 nM ) torat spinal cord MNs on the sixth day in culture immunodetectedusing antibodies against the VSV-G tag. Bar 5 10 mm. B: Bind-ing and crosslinking of 32P-HC (200 pM ) to rat MNs with orwithout preincubation with competitors (abbreviations as in Fig.3) or different lectins (100 mg/ml; SN, Sambucus nigra; otherabbreviations as in the text). 32P-HC binding and the formation ofthe '60- to 62-kDa band (arrow) are significantly competed bypreincubation with HC and TeNT and partially by BoNT/E HC andLFA. Asterisks point to additional products obtained after pre-treatment with TV and MA.
J. Neurochem., Vol. 74, No. 5, 2000
1946 J. HERREROS ET AL.

crosslinking product was observed from the second tothird day in culture and coincided with a remarkableaxonal and dendritic growth and an increase of32P-HCbinding (data not shown). The 62-kDa band was com-peted for by addition of cold HC and displayed manyfeatures of the crosslinking product observed in NGF-differentiated PC12 cells, providing indirect evidencethat TeNT binding is similar in the two cellular systems.Accordingly, pretreatment of MNs with lectins showedthe same specificity of inhibition on the HC binding asseen in PC12 cells, including the appearance of theadditional crosslinking products with TV and MA (Fig.6B).
The HC of BoNT/E (but not of BoNT/A) partiallycompeted with the binding of TeNT32P-HC to rat MNs,while strongly inhibiting the formation of the 65-kDaband (Fig. 6B). This result is in agreement with thereported TeNT HC antagonism of BoNT/E action at themurine phrenic nerve–hemidiaphragm (Simpson, 1984)and suggests that BoNT/E may interact with the sameprotein receptor of TeNT in these experimental systems.Consistently, a crosslinking product equivalent to the oneformed by TeNT32P-HC was obtained after binding ofBoNT/E 32P-HC to MNs that had been maintained inculture for at least 10 days (data not shown).
To extend the validity of our observations, other neu-ronal cultures were screened with our binding andcrosslinking assay. Crosslinking products with similarmolecular masses (56–62 kDa) were obtained after in-cubation of retinoic acid-differentiated human neuroblas-toma cells (Fig. 7A) and of a mixed population of mousespinal cord cells in culture (Fig. 7B) with TeNT32P-HC.This last preparation has been shown to display func-tional receptors for TeNT and has been extensively usedto test the intracellular activity of CNTs (Williamsonet al., 1996; Lalli et al., 1999). Thus, p15s from differentcell types display slight differences in their apparentmobility in SDS-PAGE. This result could be explained
by different cell- and species-specific glycosylation pat-terns, which could be altered in certain cancerous celllines (Rosenberg, 1995) such as the human neuroblas-toma, and is supported by the finding that the mobility ofthe TeNT HC–p15 complex is altered byN-glycosidaseactivity.
DISCUSSION
Natural toxins have been widely exploited as specifictools with which to study the molecular mechanisms ofseveral physiological processes. CNTs have been instru-mental in the functional analysis of neurotransmitterrelease and in the definition of the central role played bysynaptic SNAREs (syntaxin, SNAP-25, and VAMP/syn-aptobrevin) (Niemann et al., 1994; Montecucco andSchiavo, 1995). In addition, the binding fragment ofTeNT has been used as a marker for retrograde transport(Schwab et al., 1977; Bizzini et al., 1981; Fishman andCarrigan, 1987; Manning et al., 1990) and for the target-ing and delivery of exogenous proteins into the CNS(Beaude et al., 1990; Dobrenis et al., 1992; Coen et al.,1997; Figueiredo et al., 1997). Despite these applica-tions, no precise information is available on the molec-ular mechanism used by TeNT to interact with motor,sensory, and adrenergic neurons (Sto¨ckel et al., 1975).This specific recognition is essential for the routing ofTeNT to the retrograde transport pathway and the con-sequent dendritic targeting and transsynaptic transfer(Schwab et al., 1979).
A key element in this investigation is the characteri-sation of the TeNT-neurospecific receptor(s). Polysialo-gangliosides of the G1b series have been implicated inTeNT binding and uptake (Habermann and Dreyer,1986; Halpern and Neale, 1995; Williamson et al., 1999).Although TeNT binding to these lipids is undisputed,concerns arise about their role as the unique toxin recep-tors at the synapse. Their ubiquitous distribution andtheir relatively low binding affinity suggest that otheracceptors may be involved. In addition, BoNTs andTeNT bind to polysialogangliosides with similar speci-ficities and affinities (Critchley et al., 1986; Habermannand Dreyer, 1986; Ochanda et al., 1986; Schengrundet al., 1991), although their intracellular trafficking isremarkably different (Herreros et al., 1999). A possibleexplanation for this is the existence of additional co-receptors for different CNTs that confer specialised sort-ing and, in the case of BoNTs, serotype specificity.Despite several attempts to identify them, the nature ofthese receptors remains elusive.
Preliminary experiments using TeNT holotoxin shedlight on the presence of a putative receptor protein of15–20 kDa on NGF-differentiated PC12 cells (Schiavoet al., 1991). In this study, we characterise this putativeprotein receptor of 15 kDa specific for TeNT and possi-bly BoNT/E by using a binding and crosslinking ap-proach. This protein displays similar features in differentneuronal cell types including NGF-differentiated PC12cells and rat spinal cord MNs. NGF-differentiated PC12
FIG. 7. TeNT HC interacts with a p15-like protein in humandifferentiated neuroblastoma cells and mouse spinal cord cells.Binding and crosslinking of 200 pM 32P-HC to retinoic acid-differentiated human neuroblastoma cells (A) and to mouse spi-nal cord cells in culture (B) are shown. Bands of 58 (A) and 62 (B)kDa appear only after treatment with BSOCOES (lanes 2, ar-rows). The formation of the crosslinking products is abolished bycompetition with an excess of cold HC fragment (lanes 3).
J. Neurochem., Vol. 74, No. 5, 2000
1947TETANUS TOXIN RECEPTOR IN RAT MOTONEURONS

cells represent a widely used neuronal-like model sys-tem. After differentiation, they develop extended neu-rites and an increased number of synaptic-like vesicles(Tao-Cheng et al., 1995). In our investigation, we used aPC12 subclone that has been shown to bind TeNT and todisplay high-affinity receptors with pharmacological fea-tures similar to those found in mammalian brain (Sand-berg et al., 1989). This subclone expresses complexpolysialogangliosides before NGF differentiation (Wal-ton et al., 1988). In contrast, TeNT-induced blockade ofacetylcholine release is dependent upon NGF treatment(Figliomeni and Grasso, 1985; Sandberg et al., 1989).Our identification of p15 as a protein interacting withTeNT can now be explained by the NGF-dependentexpression of specific receptor(s) for TeNT in certainPC12 subclones. In addition, PC12 cells are known toshow a variable degree of autodifferentiation if main-tained at high density in culture, conditions in which lowamounts of the crosslinking band can be observed (Fig.2A). Nonconfluent cells, such as those used in immuno-fluorescence experiments, are lacking this band, whichstarts to appear after a few days of NGF treatment (datanot shown).
Solubilisation of cellular proteins with TX-114showed that the 65-kDa crosslinking product partitionsinto the detergent phase, whereas part of the radiola-belled HC co-fractionates with this phase. This findingmay reflect the existence of an HC subpool stronglyassociated with the plasma membrane. However, analy-sis of bound32P-HC in noncrosslinked cells revealed thatsuch a hydrophobic interaction is totally dependent onthe modification of the HC by the crosslinker (data notshown).
TeNT and several BoNT serotypes do not competewith each other for binding to rat brain preparations(Habermann and Dreyer, 1986). As expected, the bindingof TeNT HC to the cells and to p15 is abolished by TeNTor the native HC fragment but is not altered by the HCdomains of BoNT/A and B. This observation indicatesthat binding to polysialogangliosides, a feature shared byall CNTs, is not sufficient to compete with the interactionof TeNT with neurons and highlights the importance ofp15 as specific receptor. In contrast, the HC fragment ofBoNT/E decreased the binding of TeNT HC to p15,particularly in the MNs. This finding is in agreementwith the observed TeNT HC antagonism of BoNT/Eaction at the murine phrenic nerve–hemidiaphragm(Simpson, 1984). Preliminary crosslinking experimentsindicate that the HC fragment of BoNT/E interacts with aprotein with mobility similar to that of p15 (data notshown). Thus, these two CNTs may share the samecellular receptor. The less efficient formation of theBoNT/E–p15 product suggests, however, a lower affinityof BoNT/E compared with TeNT for p15.
Sialic acid-specific lectins have been proven to an-tagonise TeNT intoxication at the murine phrenic nerve–hemidiaphragm (Bakry et al., 1991a) with a specificitysimilar to that observed in the inhibition of TeNT HCbinding to p15 and to the neuronal surface. Sialic acid-
containing oligosaccharides are therefore essential forthe functional interaction of TeNT with its receptor(s).However, lectins may bind to the sialic acid residues ofpolysialogangliosides and/or to oligosaccharides presenton a putative protein receptor (Middlebrook, 1989).Here, we provided direct evidence that p15 is a glyco-protein, its mobility being dramatically increased afterdeglycosylation. To our knowledge, this is the first directdemonstration of the involvement of a plasma membraneglycoprotein in TeNT binding.
Additional evidence supporting the neurospecific ex-pression of p15 was gathered by screening for TeNTbinding and crosslinking a variety of neuronal cells,including rat spinal cord MNs and mouse spinal cordcells, and also nonneuronal cell lines such as teratomasand neuroepitheliomas, which were found negative inthese assays (unpublished results). We were able to dem-onstrate that rat spinal cord MNs express a protein thatbehaves similarly to the p15 observed in NGF-differen-tiated PC12 cells. This finding is particularly importantbecause MNs are directly involved in TeNT uptake invivo. However, we were unable to confirm the identity ofp15 by microsequencing because of the low abundanceof the crosslinking product in our system and the rich-ness of p15 in oligosaccharides. Similarly, an affinityapproach has also proven ineffective, most likely due tothe instability of the double lipid and protein receptorcomplex in the presence of detergents. An alternativeapproach based on expression cloning and using thecrosslinking strategy as screening assay is now underwayto identify p15 at the molecular level.
In conclusion, our results demonstrate the presence ofa glycoprotein specifically interacting with TeNT andBoNT/E in a variety of neuronal cell types, including thecellular target of TeNT. Its identification will constitutean important step in unravelling the mechanism of TeNTinternalisation and retrograde transport in neurons.
Acknowledgment: We thank Dr. T. B. Rogers (Universityof Maryland, College Park, MD, U.S.A.) for the PC12 sub-clone, Dr. R. Basso (University of Padova, Padova, Italy) forthe human primary neuroblastoma, Drs. C. E. Henderson andV. Arce (CNRS Marseille, Marseille, France) for advice withthe motoneuron culture, and Dr. T. Iglesias for valuable un-published data. We are thankful to Drs. T. Iglesias, C. Reis eSousa, G. Stenbeck, S. Tooze, and G. Warren for criticalreading of the manuscript. J.H. is a fellow of the HumanFrontier Science Program. This work was supported by theImperial Cancer Research Fund and Telethon-Italia grant 1062(C.M.).
REFERENCES
Arce V., Pollock R. A., Philippe J., Pennica D., Henderson C. E., anddeLaperiere O. (1998) Synergistic effects of Schwann- and mus-cle-derived factors on motoneuron survival involve GDNF andcardiotrophin-1 (CT-1).J. Neurosci.18, 1440–1448.
Bakry N., Kamata Y., and Simpson L. L. (1991a) Lectins fromTriti-cum vulgarisandLimax flavusare universal antagonists of botu-linum neurotoxin and tetanus toxin.J. Pharmacol. Exp. Ther.258,830–836.
J. Neurochem., Vol. 74, No. 5, 2000
1948 J. HERREROS ET AL.

Bakry N., Kamata Y., Sorensen R., and Simpson L. L. (1991b) Tetanustoxin and neuronal membranes: the relationship between bindingand toxicity.J. Pharmacol. Exp. Ther.258,613–619.
Beaude P., Delacour A., Bizzini B., Domuado D., and Remy M. H.(1990) Retrograde axonal transport of an exogenous enzyme co-valently linked to B-IIb fragment of tetanus toxin.Biochem. J.271,87–91.
Bizzini B., Stoeckel K., and Schwab M. (1977) An antigenic polypep-tide fragment isolated from tetanus toxin: chemical characteriza-tion, binding to gangliosides, and retrograde axonal transport invarious neuron systems.J. Neurochem.28, 529–542.
Bizzini B., Grob P., and Akert K. (1981) Papain-derived fragment IIcof tetanus toxin: its binding to isolated synaptic membranes andretrograde axonal transport.Brain Res.210,291–299.
Black J. D. and Dolly J. O. (1986a) Interaction of125I-labeled botuli-num neurotoxins with nerve terminals. II. Autoradiographic evi-dence for its uptake into motor nerves by acceptor-mediatedendocytosis.J. Cell Biol. 103,535–544.
Black J. D. and Dolly J. O. (1986b) Interaction of125I-labeled botuli-num neurotoxins with nerve terminals. I. Ultrastructural autora-diographic localization and quantitation of distinct membraneacceptors for types A and B on motor nerves.J. Cell Biol. 103,521–534.
Chandler C. E., Parsons L. M., Hosang M., and Shooter E. M. (1984)A monoclonal antibody modulates the interaction of nerve growthfactor with PC12 cells.J. Biol. Chem.259,6882–6889.
Coen L., Osta R., Maury M., and Brulet P. (1997) Construction ofhybrid proteins that migrate retrogradely and transsynapticallyinto the central nervous system.Proc. Natl. Acad. Sci. USA94,9400–9405.
Critchley D. R., Habig W. H., and Fishman P. H. (1986) Reevaluationof the role of gangliosides as receptors for tetanus toxin.J. Neu-rochem.47, 213–222.
Dobrenis K., Joseph A., and Rattazzi M. C. (1992) Neuronal lysosomalenzyme replacement using fragment C of tetanus toxin.Proc.Natl. Acad. Sci. USA89, 2297–2301.
Figliomeni B. and Grasso A. (1985) Tetanus toxin affects the K1-stimulated release of catecholamines from nerve growth factor-treated PC12 cells.Biochem. Biophys. Res. Commun.128, 249–256.
Figueiredo D. M., Hallewell R. A., Chen L. L., Fairweather N. F.,Dougan G., Savitt J. M., Parks D. A., and Fishman P. S. (1997)Delivery of recombinant tetanus-superoxide dismutase proteins tocentral nervous system neurons by retrograde axonal transport.Exp. Neurol.145,546–554.
Fishman P. S. and Carrigan D. R. (1987) Retrograde transneuronaltransfer of the C-fragment of tetanus toxin.Brain Res.406,275–279.
Gill D. M. (1982) Bacterial toxins: a table of lethal amounts.Microbiol.Rev.46, 86–94.
Habermann E. and Dreyer F. (1986) Clostridial neurotoxins: handlingand action at the cellular and molecular level.Curr. Top. Micro-biol. Immunol.129,93–179.
Halpern J. L. and Neale E. A. (1995) Neurospecific binding, internal-ization, and retrograde axonal transport.Curr. Top. Microbiol.Immunol.195,221–241.
Henderson C. E., Bloch-Gallego E., and Camu W. (1995) Purificationand culture of embryonic spinal motoneurons, inNerve CellCulture: A Practical Approach(Cohen J. and Wilkin G. P., eds),pp. 69–81. Oxford University Press, London.
Herreros J., Lalli G., Montecucco C., and Schiavo G. (1999) Patho-physiological properties of clostridial neurotoxins, inThe Com-prehensive Sourcebook of Bacterial Protein Toxins(Freer J. H.and Alouf J. E., eds), pp. 202–228. Academic Press, London.
Kaelin W. G. Jr., Krek W., Sellers W. R., DeCaprio J. A., AjchenbaumF., Fuchs C. S., Chittenden T., Li Y., Farnham P. J., and BlanarM. A. (1992) Expression cloning of a cDNA encoding a retino-blastoma-binding protein with E2F-like properties.Cell 70, 351–364.
Laemmli U. K. (1970) Cleavage of structural proteins during theassembly of the head of bacteriophage T4.Nature227,680–685.
Lalli G., Herreros J., Osborne S. L., Montecucco C., Rossetto O., andSchiavo G. (1999) Functional characterisation of tetanus andbotulinum neurotoxins binding domains.J. Cell Sci.112, 2715–2724.
Li L. and Singh B. R. (1998) Isolation of synaptotagmin as a receptorfor type A and type E botulinum neurotoxin and analysis of theircomparative binding using a new microtiter plate assay.J. Nat.Toxins7, 215–226.
Manning K. A., Erichsen J. T., and Evinger C. (1990) Retrogradetransneuronal transport properties of fragment C of tetanus toxin.Neuroscience34, 251–263.
Matteoli M., Verderio C., Rossetto O., Iezzi N., Coco S., Schiavo G.,and Montecucco C. (1996) Synaptic vesicle endocytosis mediatesthe entry of tetanus neurotoxin into hippocampal neurons.Proc.Natl. Acad. Sci. USA93, 13310–13315.
Middlebrook J. L. (1989) Cell surface receptors for protein toxins, inBotulinum Neurotoxin and Tetanus Toxin(Simpson L. L., ed), pp.95–119. Academic Press, San Diego.
Montecucco C. (1986) How do tetanus and botulinum toxins bind toneuronal membranes?Trends Biochem. Sci.11, 315–317.
Montecucco C. and Schiavo G. (1995) Structure and function of tetanusand botulinum neurotoxins.Q. Rev. Biophys.28, 423–472.
Naglich J. G., Metherall J. E., Russell D. W., and Eidels L. (1992)Expression cloning of a diphtheria toxin receptor: identity with aheparin-binding EGF-like growth factor precursor.Cell 69,1051–1061.
Niemann H., Blasi J., and Jahn R. (1994) Clostridial neurotoxins: newtools for dissecting exocytosis.Trends Cell Biol.4, 179–185.
Nishiki T., Kamata Y., Nemoto Y., Omori A., Ito T., Takahashi M., andKozaki S. (1994) Identification of protein receptor forClostridiumbotulinum type B neurotoxin in rat brain synaptosomes.J. Biol.Chem.269,10498–10503.
Nishiki T., Tokuyama Y., Kamata Y., Nemoto Y., Yoshida A., Sato K.,Sekiguchi M., Takahashi M., and Kozaki S. (1996a) The high-affinity binding of Clostridium botulinumtype B neurotoxin tosynaptotagmin II associated with gangliosides GT1b/GD1a.FEBSLett. 378,253–257.
Nishiki T., Tokuyama Y., Kamata Y., Nemoto Y., Yoshida A., Sekigu-chi M., Takahashi M., and Kozaki S. (1996b) Binding of botuli-num type B neurotoxin to Chinese hamster ovary cells transfectedwith rat synaptotagmin II cDNA.Neurosci. Lett.208,105–108.
Ochanda J. O., Syuto B., Ohishi I., Naiki M., and Kubo S. (1986)Binding of Clostridium botulinumneurotoxin to gangliosides.J. Biochem. (Tokyo)100,27–33.
Parton R. G., Ockleford C. D., and Critchley D. R. (1988) Tetanustoxin binding to mouse spinal cord cells: an evaluation of the roleof gangliosides in toxin internalization.Brain Res.475,118–127.
Pierce E. J., Davison M. D., Parton R. G., Habig W. H., and CritchleyD. R. (1986) Characterization of tetanus toxin binding to rat brainmembranes. Evidence for a high-affinity proteinase-sensitive re-ceptor.Biochem. J.236,845–852.
Rosenberg A. (1995)Biology of Sialic Acids,pp. 168–196. PlenumPress, New York.
Sandberg K., Berry C. J., and Rogers T. B. (1989) Studies on theintoxication pathway of tetanus toxin in the rat pheochromocy-toma (PC12) cell line. Binding, internalization, and inhibition ofacetylcholine release.J. Biol. Chem.264,5679–5686.
Schengrund C.-L., DasGupta B. R., and Ringler N. J. (1991) Binding ofbotulinum and tetanus neurotoxins to ganglioside GT1b and de-rivatives thereof.J. Neurochem.57, 1024–1032.
Schengrund C. L., Ringler N. J., and Dasgupta B. R. (1992) Adherenceof botulinum and tetanus neurotoxins to synaptosomal proteins.Brain Res. Bull.29, 917–924.
Schengrund C. L., DasGupta B. R., Hughes C. A., and Ringler N. J.(1996) Ganglioside-induced adherence of botulinum and tetanusneurotoxins to adducin.J. Neurochem.66, 2556–2561.
Schiavo G. and Montecucco C. (1995) Tetanus and botulism neuro-toxins: isolation and assay.Methods Enzymol.248,643–652.
Schiavo G., Ferrari G., Rossetto O., and Montecucco C. (1991) Specificcross-linking of tetanus toxin to a protein of NGF-differentiatedPC12 cells.FEBS Lett.290,227–230.
J. Neurochem., Vol. 74, No. 5, 2000
1949TETANUS TOXIN RECEPTOR IN RAT MOTONEURONS

Schwab M., Agid Y., Glowinski J., and Thoenen H. (1977) Retrogradeaxonal transport of125I-tetanus toxin as a tool for tracing fiberconnections in the central nervous system; connections of therostral part of the rat neostriatum.Brain Res.126,211–224.
Schwab M. E., Suda K., and Thoenen H. (1979) Selective retrogradetranssynaptic transfer of a protein, tetanus toxin, subsequent to itsretrograde axonal transport.J. Cell Biol. 82, 798–810.
Simpson L. L. (1984) The binding fragment from tetanus toxin antag-onizes the neuromuscular blocking actions of botulinum toxin.J. Pharmacol. Exp. Ther.229,182–187.
Soldati T. and Perriard J.-C. (1991) Intracompartmental sorting ofessential myosin light chains: molecular dissection and in vivomonitoring by epitope tagging.Cell 66, 277–289.
Stockel K., Schwab M., and Thoenen H. (1975) Comparison betweenthe retrograde axonal transport of nerve growth factor and tetanustoxin in motor, sensory and adrenergic neurons.Brain Res.99,1–16.
Tao-Cheng J. H., Dosemeci A., Bressler J. P., Brightman M. W., andSimpson D. L. (1995) Characterisation of synaptic vesicles andrelated neuronal features in nerve growth factor and ras oncogenedifferentiated PC12 cells.J. Neurosci. Res.42, 323–334.
Tarentino A. L. and Plummer T. H. (1994) Enzymatic deglycosylationof asparagine-linked glycans: purification, properties, and speci-ficity of oligosaccharide-cleaving enzymes fromFlavobacteriummeningosepticum. Methods Enzymol.230,44–57.
Walton K. M., Sandberg K., Rogers T. B., and Schnaar R. L. (1988)Complex ganglioside expression and tetanus toxin binding byPC12 pheochromocytoma cells.J. Biol. Chem.263,2055–2063.
Williamson L. C., Halpern J. L., Montecucco C., Brown J. E., andNeale E. A. (1996) Clostridial neurotoxins and substrate proteol-ysis in intact neurons: botulinum neurotoxin C acts on synapto-somal-associated protein of 25 kDa.J. Biol. Chem.271, 7694–7699.
Williamson L. C., Bateman K. E., Clifford J. C. M., and Neale E. A.(1999) Neuronal sensitivity to tetanus toxin requires gangliosides.J. Biol. Chem.274,25173–25180.
Yan Q. and Johnson E. J. (1988) An immunohistochemical study of thenerve growth factor receptor in developing rats.J. Neurosci.8,3481–3498.
Yavin E. and Nathan A. (1986) Tetanus toxin receptors on nerve cellscontain a trypsin-sensitive component.Eur. J. Biochem.154,403–407.
J. Neurochem., Vol. 74, No. 5, 2000
1950 J. HERREROS ET AL.