transsynaptic retrograde transport of fragment c of tetanus toxin demonstrated by...
TRANSCRIPT

Brain Research, 380 (1986) 383-388 383 Elsevier
BRE 21725
Transsynaptic retrograde transport of fragment C of tetanus toxin demonstrated by immunohistochemical localization
CRAIG EVINGER and JONATHAN T. ERICHSEN
Department of Neurobiology and Behavior, SUNY at Stony Brook, Stony Brook, NY 11794 (U.S.A )
(Accepted May 6th, 1986)
Key words: transsynaptic transport - - fragment C of tetanus toxin - - immunohistochemical localization - - vestibulo-ocular pathway
Injection of the non-toxic fragment C of tetanus toxin into the superior oblique muscle of the eye results in strong direct retrograde labelling of the motoneurons in the contralateral trochlear nucleus and clear transsynaptic labelling of neurons in the ipsilateral and contralateral vestibular nuclei. Standard immunohistochemical procedures using a monoclonal antibody localize fragment C in the brain with high sensitivity and excellent resolution. An injection of fragment C into the superior oblique muscle labels the same pool of trochlear motoneurons as an HRP injection into the superior oblique muscle. A comparison of the fragment C labelling of trochlear motoneurons with intracellular injections of HRP into trochlear motoneurons suggests that fragment C stains not just the soma, but also the distal dendrites of motoneurons. Moreover, a fragment C injection into the superior oblique muscle labels transsynaptically more neurons in each vestibular nucleus than an injection of HRP into the trochlear nucleus labels directly.
A marke r that undergoes re t rograde t ranssynaptic
t ranspor t would be an impor tant tool for neuroana-
tomical research. Tetanus toxin exhibits this prop-
erty, but three p rob lems have p reven ted tetanus tox-
in from being commonly used. First , even in small
quanti t ies tetanus toxin is lethal, so long survival
t imes are impossible. Second, localizing tetanus toxin
or its fragments has requi red conjugat ion to 1251 and
au torad iography 2'4, a me thod that lacks the resolution
of techniques such as immunohis tochemis t ry as well
as being t ime-consuming. Third, the extent of retro-
grade transsynaptic t ranspor t of te tanus toxin frag-
ments has not been carefully tested in the central ner-
vous system. The recent deve lopment of non-toxic
fragments of te tanus toxin which show transsynaptic
t ranspor t overcomes the problem of toxicity 1-7'12'13.
The present demons t ra t ion that a monoclonal anti-
body can be used to localize the non-toxic f ragment C
of te tanus toxin supplants the need for autoradiogra-
phy. Finally, our compar ison of the re t rograde trans-
synaptic t ranspor t of f ragment C of te tanus toxin with
direct re t rograde t ranspor t of horseradish peroxidase
(HRP) in a vest ibulo-ocular pathway shows that loca-
l ization of f ragment C with a monoclonal ant ibody
provides a direct re t rograde transsynaptic labelling
method for de termining pathways in the central ner-
vous system.
Light microscopic examinat ion of brains tem sec-
tions, following inject ion of f ragment C of te tanus
toxin into the super ior obl ique muscle of the pigeon (Colurnba livia), reveals direct re t rograde t ranspor t
of f ragment C to the contra la tera l t rochlear nucleus
(Figs. 1B, 2A, B) and clear t ranssynaptic t ranspor t to
the vest ibular nuclei (Fig. 2C, D). We sought to com-
pare this pa t te rn of labelling with data obta ined using
wel l -developed, convent ional techniques.
Direct retrograde transport. Direct re t rograde la-
belling with H R P showed the locat ion and distribu-
tion of motoneurons innervat ing the superior oblique
muscle (Fig. 1A). Twenty-four hours after placing
the IVth nerve of two birds in a cuff containing 40%
HRP (in Tris buffer, pH 7.4), the birds were perfused
with avian saline followed by a fixative of 1% para-
formaldehyde and 1.25% glutara ldehyde in phos-
phate buffer (pH 7.4) (PB). Fifty ¢tm sections of the
brain were cut on a freezing micro tome, reacted with
the te t ramethyl benzidine method 11 modified for glu-
cose oxidase s and counters ta ined with neutral red.
Correspondence: C. Evinger, Dept. Neurobiology and Behavior, SUNY at Stony Brook, Stony Brook, NY 11794, U.S.A.
0006-8993/86/$03.50 © 1986 Elsevier Science Publishers B .V. (Biomedical Division)
384
r
A B m '
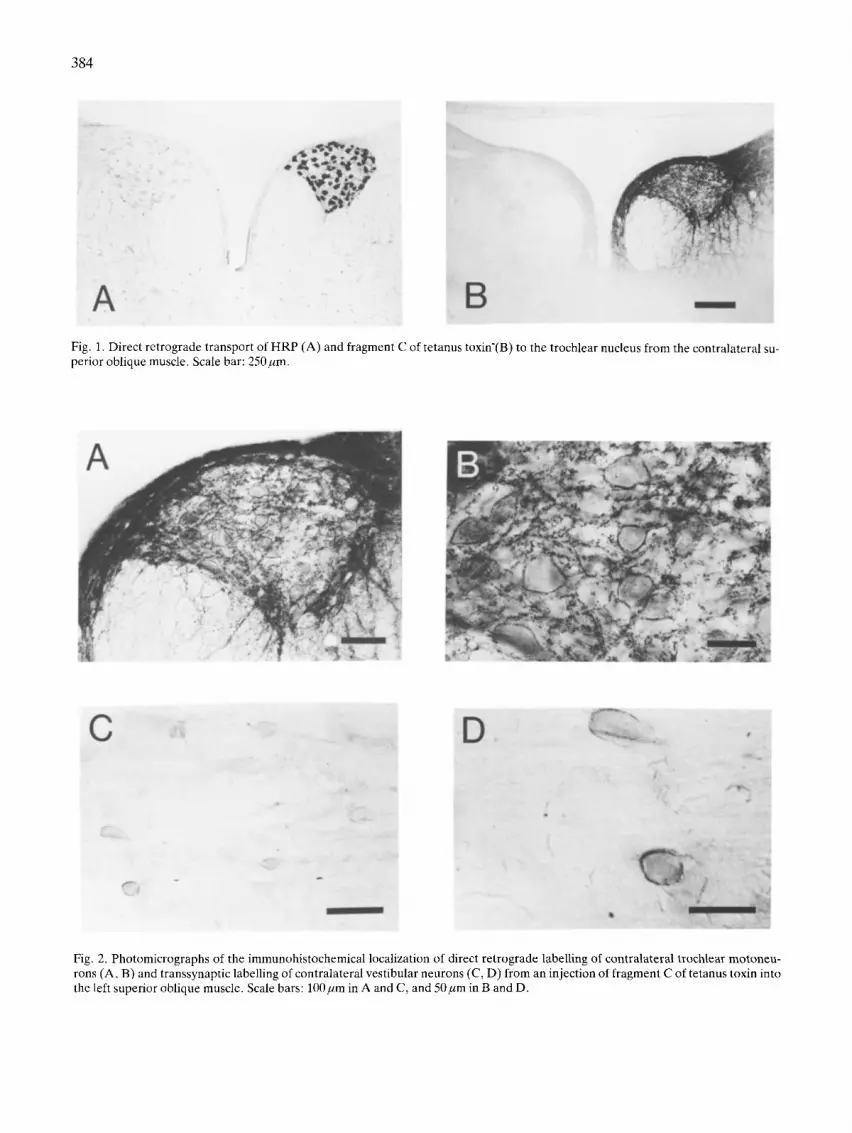
Fig. 1. Direct retrograde transport of HRP (A) and fragment C of tetanus toxin'(B) to the trochlear nucleus from the contralateral su- perior oblique muscle. Scale bar: 250 pm.
C
r r
-, , ? , . ~o;, f.
Fig. 2. Photomicrographs of the immunohistochemical localization of direct retrograde labelling of contralatera| trochlear motoneu- rons (A, B) and transsynaptic labelling of contralateral vestibular neurons (C, D) from an injection of fragment C of tetanus toxin into the left superior oblique muscle. Scale bars: 100/~m in A and C, and 50pro in B and D.

Reconstruction of the trochlear nucleus demon- strated that all neurons innervated the contralateral superior oblique muscle and that no labelling oc- curred in the ipsilateral nucleus (Fig. 1A).
Injection of fragment C of tetanus toxin into the superior oblique muscle labelled the same population of motoneurons as that backfilled by an HRP injec- tion (Figs. 1B, 2A, B). Eight days after injection of 20 B1 of a 5% solution of fragment C (Calbiochem) into 3 pigeons, the birds were perfused with 6% dex- tran in PB followed by a fixative of 4% paraformal- dehyde in PB. Thirty-five gm sections of the brain were cut on a freezing microtome, incubated with a monoclonal antibody we raised against fragment C
(dilution 1:500), followed by incubation with biotinyl- ated anti-mouse IgG. The tissue was treated using the standard protocol for the Avidin-Biotin proce- dure (Vector Labs) and reacted with diaminobenzi- dine (DAB). The absence of labelling in the ipsilater- al trochlear nucleus (Fig. 1B) or in neurons other than those known to terminate on trochlear moto- neurons confirmed that non-specific binding did not O c c u r .
All contralateral trochlear motoneurons, but no motoneurons ipsilateral to the injected superior oblique muscle, contain reaction product following a fragment C injection (Fig. 1B, 2A, B). Nevertheless, the pattern of staining differs from that associated with HRP. First, fragment C appears to bind to the membrane rather than filling the inside of the cell (Fig. 2B). Indeed, the staining gives the impression that synaptic boutons surrounding the motoneurons contain label. Second, the staining associated with fragment C extends well outside the nucleus into the medial longitudinal fasciculus (MLF) and surround- ing reticular formation. To confirm that this is den- dritic staining, we determined the soma-dendrit ic organization of trochlear motoneurons with intracel- lular staining using HRP.
Four trochlear motoneurons in three pigeons were penetrated with micropipettes containing 10% HRP (in equal amounts of Tris buffer pH 7.4 and 0.5 M KC1), antidromically identified by stimulation of the IVth nerve, and tested for vestibular responses by electrical stimulation of the VIIIth nerves. All moto- neurons exhibited, at disynaptic latencies, an EPSP from stimulation of the contralateral VIIIth nerve and an IPSP from stimulation of the ipsilateral VIII th
385
nerve. HRP was then iontophoresed into the neurons
with steps of positive current ranging from 10 to 20 nA with a 75% duty cycle. One hour after the injec- tion, the bird was perfused with avian saline followed by the fixative used in the HRP protocol. One hundredj~m sections of the brain were cut on a freez- ing microtome, reacted with diaminobenzidine, and counterstained with cresyl violet. Reconstruction of each motoneuron revealed the same broad dendritic field extending into the MLF and stretching laterally into the surrounding reticular formation (Fig. 3). Thus, the extent of the dendritic arbor of intracellu- larly stained motoneurons corresponds to the pattern of labelling resulting from direct retrograde transport of fragment C to motoneurons. This implies that in addition to reaching the soma, fragment C also spreads throughout the dendritic tree of trochlear motoneurons.
Transsynaptic transport. The populations of vestib- ular neurons labelled transsynaptically by injections of fragment C into the superior oblique muscle (e.g. see Fig. 2C, D) are nearly the same as the vestibular neurons backfilled directly by a large HRP injection into the trochlear nucleus. Fragment C, however, la- bels more cells in each nucleus than does HRP (Fig. 4). In order to identify which vestibular nucleus neu- rons project to the trochlear nucleus, HRP was pressure-injected into the trochlear nucleus of two pigeons. Twenty-four hours later the birds were per- fused with avian saline and fixative. The brains were
/ ' /
/ -
~ j j ~ /
Fig. 3. Reconstruction of a trochlear motoneuron injected in- tracellularly with HRP. Abbreviations: A, axon; MLF, medial longitudinal fasciculus.

386
C
BRECT RETROGRADE
TRAN~SYNAPTIC ( ~ 2 mm
HRP /
TEC
~-I~ P o.s
P 0.7
f v~ ~ l~, ~>
',MLF/ .....
C HRP

387
treated as described for the extracellular H R P exper- iments. In both cases the H R P injection filled the
trochlear nucleus, as well as extending rostrally into the inferior rectus subdivision of the oculomotor nu-
cleus, and spreading dorsally into the reticular for- mation (Fig. 4, right side). The illustrated injection
retrogradely labelled neurons bilaterally in the nu- cleus tangentialis (P 0.3-0.7) l°, the lateral vestibular
nucleus (P 0.1-0.5) , neurons contralaterally in the de-
scending vestibular nucleus (P 0.1-0.7) , the medial
vestibular nucleus (P 0.3) and neurons in the ipsilat-
eral superior vestibular nucleus (A 0.1) which extend into the immediately caudal dorsal lateral vestibular
nucleus (P 0.1). The fragment C muscle injection
transsynaptically labels similar populations of vestib-
ular neurons as does the H R P injection into the trochlear nucleus, except for the rostrally located ip- silateral superior vestibular and lateral vestibular nu-
clei (Fig. 4, compare A 0.1 and P 0.1 in the left and right panels). Fragment C, however, surpasses the H R P injection in the number of labelled neurons in
each of the nuclei. The absence of fragment C-
labelled neurons in the ipsilateral superior vestibular nucleus may suggest that fragment C does not move transsynaptically across this synapse. Alternatively,
the large H R P injection may have labelled afferents
terminating outside the trochlear nucleus or fibers of
passage. Immunohistochemical localization of transsynapti-
cally transported fragment C offers important advan-
tages over H R P techniques for identifying disynaptic pathways. First, our demonstration of the transport
of fragment C to premotor afferent neurons shows that fragment C labels a larger number of neurons
than does direct retrograde HRP. This enhanced staining probably results from retrograde transport
of fragment C to not only the soma of the motoneu-
ron, but also to its distal dendrites, potentially expos- ing all afferent synapses to the tetanus toxin frag-
ment. An H R P injection confined to the cytoarchi- tectonic boundaries of a nucleus (as defined by soma
distribution) would miss afferent inputs on dendrites
extending outside of the nucleus. Second, discrete HRP injections into nuclei containing motoneurons innervating more than one muscle (e.g. oculomotor
nucleus, facial nucleus) are virtually impossible. In-
jection of fragment C directly into a muscle with sub- sequent transsynaptic transport obviates this prob- lem. Third, H R P injections invariably label fibers
passing through the injection site. Because tetanus
toxin appears to enter neurons only at synaptic junc-
tions 12'13 (unpublished observations), fragment C should not label fibers of passage. Fourth, unlike the transsynaptic transport of W G A - H R P , which must
be facilitated by electrical stimulation of afferent ax- ons 9, transsynaptic labelling with fragment C re-
quires no special treatment. Thus, when localized with a monoclonal antibody, tetanus toxin fragment
C is a reliable and powerful transsynaptic retrograde
marker for neuroanatomical research.
This work was supported by NIH Grants EY04829 (C.E.), EY04587 (J.T.E.) and NSF Grant
BNS84818752 (C.E.). C.E. was an Alfred P. Sloan Research Fellow. We would like to thank H. Karten
for his aid and encouragement, A. Gero and D. An- thony for their technical assistance, and K.A. Man-
ning and J. Hamos for helpful comments on an earlier version of this manuscript.
1 Bizzini, B., Tetanus toxin, Microbiol. Rev.. 43 (1979) 224-240.
2 Bizzini, B., Grob, P. and Akert, K., Papain-derived frag- ment II c of tetanus toxin: its binding to isolated synaptic membranes and retrograde axonal transport, Brain Re- search, 210 (1981) 291-299.
3 Bizzini, B., Stoeckel, K. and Schwab, M., An antigenic
polypeptide fragment isolated from tetanus toxin: chemical characterization, binding to gangliosides and retrograde axonal transport in various neuron systems, J. Neurochem.. 28 (1977) 529-542.
4 Buttner-Ennever, J.A., Grob, P., Akert, K. and Bizzini, B., Transsynaptic retrograde labelling in the oculomotor system of the monkey with [125I]tetanus toxin BII b frag-
Fig. 4. Labelled neurons in the vestibular complex from an HRP injection into the trochlear nucleus (right panel) and transsynaptic la- belling from injection of fragment C into the superior obliqtae muscle (left panel). Sections are at levels determined from the Karten and Hodos atlas. Each dot is a single labelled neuron. Abbreviations: DV, descending vestibular nucleus: IO, isthmo-optic nucleus; LVD, lateral vestibular nucleus dorsal; LVV, lateral vestibular nucleus ventral; MLF, medial longitudinal fasciculus; MV, medial ves- tibular nucleus; PL, nucleus pontis lateralis; SL, nucleus semiluminaris; SV, superior vestibular nucleus; TA, nucleus tangentialis; TEC, tectum; TRO, trochlear nucleus; V, ventricle; VN, vestibular nuclei; IVn, IVth nerve; VI, abducens nucleus; VIac, accessory abducens nucleus; VIII, VIIIth nerve.

388
ment, Neurosci. Lett., 26 (1981) 233-238. 5 Dumas, M., Schwab, M.E., Baumann, R. and Thoenen,
H., Retrograde transport of tetanus toxin through a chain of two neurons, Brain Research, 165 (1979) 354-357.
6 Helting, T.B. and Zwisler, O., Structure of tetanus toxin. I. Breakdown of the toxin molecule and discrimination be- tween polypeptide fragments, J. Biol. Chem., 252 (1977) 187-193.
7 Helting, T.B., Zwisler, O. and Wiegandt, H., Structure of tetanus toxin. II. Toxin binding to ganglioside, J. Biol. Chem., 252 (1977) 194-198.
8 Itoh, K., Konishi, A., Nomura, S., Mizuno, N., Nakamura, Y. and Sugimoto, T., Application of coupled oxidation reaction to electron microscopic demonstration of horse- radish peroxidase: cobalt-glucose oxidase method, Brain Research, 175 (1979) 341-346.
9 Jankowska, E., Further indications for enhancement of ret- rograde transneuronal transport of WGA-HRP by synaptic
activity, Brain Research, 341 (1985) 403-408. 10 Karten, H.J. and Hodos, W., A Stereotaxic Atlas of the
Brain o f the Pigeon (Columba livia), Johns Hopkins Press, Baltimore, 1967.
11 Mesulam, M.-M., Tetramethyl benzidine for horseradish peroxidase neurohistochemistry: a non-carcinogenic blue reaction-product with superior sensitivity for visualizing af- ferents and efferents, J. Histochem. Cytochem., 26 (1978) 106-117.
12 Schwab, M.E. and Thoenen, H., Electron microscope evi- dence for a transsynaptic migration of tetanus toxin on spi- nal cord motoneurons: an autoradiographic and morpho- metric study, Brain Research, 105 (1976) 213-227.
13 Schwab, M.E., Suda, K. and Thoenen, H., Selective retro- grade transsynaptic transfer of a protein, tetanus toxin, subsequent to its retrograde axonal transport, J. Cell Biol., 82 (1979) 798-810.
Endoscopic Retrograde Cholangiography for Biliary Anastomotic Strictures After Liver Transplantation
Anomalous information reception by research mediums demonstrated using a novel triple-blind protocol