the decorin sequence syiriadtnit binds collagen type i
TRANSCRIPT

The Decorin Sequence SYIRIADTNIT Binds Collagen Type I*□S
Received for publication, January 3, 2007, and in revised form, April 3, 2007 Published, JBC Papers in Press, April 10, 2007, DOI 10.1074/jbc.M700073200
Sebastian Kalamajski‡, Anders Aspberg‡§, and Åke Oldberg‡1
From the ‡Department of Experimental Medical Science, University of Lund, SE-221 84 Lund, Sweden and the §Department ofMolecular Biology, University of Copenhagen, DK-2100 Copenhagen OE, Denmark
Decorin belongs to the small leucine-rich repeat proteoglycanfamily, interacts with fibrillar collagens, and regulates theassembly, structure, and biomechanical properties of connec-tive tissues. The decorin-collagen type I-binding region islocated in leucine-rich repeats 5–6. Site-directed mutagenesisof this 54-residue-long collagen-binding sequence identifiesArg-207 and Asp-210 in leucine-rich repeat 6 as crucial for thebinding to collagen. The synthetic peptide SYIRIADTNIT,which includes Arg-207 and Asp-210, inhibits the binding offull-length recombinant decorin to collagen in vitro. These col-lagen-binding amino acids are exposed on the exterior of the�-sheet-loop structure of the leucine-rich repeat. This resem-bles the location of interacting residues in other leucine-richrepeat proteins.
Decorin is an extracellular matrix proteoglycan involved incollagen type I fibril formation and collagenmatrix assembly ina wide range of connective tissues (1–3). Decorin belongs to thesmall leucine-rich repeat proteoglycan family (SLRPs)2 (4),which also includes, for example, biglycan (5), fibromodulin (6),lumican (7), and asporin (8, 9). The decorin core protein iscomposed of 12 tandem leucine-rich repeats (LRRs), each con-taining an average of 24 amino acid residues. The decorin three-dimensional structure, resolved by x-ray crystallography,reveals an arch-shaped molecule with LRRs composed of par-allel �-sheets on the concave surface and short �-strands, 310helices, and polyproline II helices on the convex face (10). Closeto the N terminus, decorin is substituted with a single chon-droitin or dermatan sulfate chain. Decorin also has three poten-tial consensus sites for N-glycosylation in the LRR domain (4).Several SLRPs are involved in the regulation of collagen fibril
formation and matrix assembly, as demonstrated in SLRP-de-ficient mice (11). Decorin deficiency causes skin fragility (12);biglycan-deficient mice suffer from reduced bone mass andosteoarthritis (13); lack of fibromodulin results in weaker ten-
dons and ligaments, causing osteoarthritis (14); and lumicanablation correlates with corneal opacity (15). In all these cases,ultrastructural imaging reveals abnormally developed collagenfibrils.Decorin presumably affects the lateral association of collagen
type I fibrils in vitro as it binds to collagenmonomers and delaystheir accretion to the growing fibrils (1). Collagen fibrils in theskin of decorin-deficient mice are thicker and irregularlyshaped, in contrast to smaller, uniform fibrils formed in wildtype mice (12). Decorin binds near the C terminus of collagen,close to an intermolecular cross-linking site (16), and seems tohave a tutelary function in collagen fibril assembly, which isimportant for the collagen cross-linking. In addition, decorinalso binds collagen type VI (17), transforming growth factor-�(18), fibronectin (19), and epidermal growth factor receptor(20).The precise decorin-binding site for collagen has not been
determined. An early study identified two collagen-bindingsites in decorin (21). A later report assigned a collagen-bindingsite to LRR 5–6 (22) and Glu-180 in decorin LRR 5 (Glu-180 inhuman decorin corresponds to Glu-181 in the bovinesequence) was proposed to be essential for the interaction (23).Decorin with an E180Q mutation was later demonstrated tobind more strongly to collagen type VI than wild type decorin,and amino acid residues located C-terminal of Gln-153 indecorin were implicated in the collagen binding (24). In addi-tion to the core protein, the dermatan sulfate chain has alsobeen reported to be involved in the collagen interaction (25).This glycosaminoglycan-collagen interaction is weaker thanthe core protein-collagen interactions (22–24).It is of interest to characterize this prototype decorin-colla-
gen interaction since other SLRPs, e.g. fibromodulin and lumi-can, also interact with fibrillar collagens and affect connectivetissue properties in similar ways. The concerted action of sev-eral SLRPs candetermine the structure and function of collagenmatrices. In this study, we used site-directed mutagenesis ofdecorin LRR 5–6 to clarify the location of the collagen-bindingsite.
EXPERIMENTAL PROCEDURES
Expression of GST-tagged bacDCN56 in Bacteria—Bovinedecorin cDNA (GenBankTM accession number NM_173906)was used as a template in PCR to amplify cDNA encoding LRR5–6 of decorin. The primers used for the PCRwere: 5�-AAAG-GATCCAACGAGATCACCAAAGTGCG-3� and 5�-AAAC-CCGGGTATATTTGTGTCAGCAATGCG-3�. The amplifiedcDNA was digested with the restriction enzymes BamHI andSmaI and ligated into the vector pGEX-4T-3 (Amersham Bio-sciences). The construct was sequenced to confirm the identity
* This work was supported by grants from the Swedish Research Council,Gustaf V:s 80-Anniversary Fund, Greta and Johan Kock’s Fund, and AlfredOsterlund’s Fund. The costs of publication of this article were defrayed inpart by the payment of page charges. This article must therefore be herebymarked “advertisement” in accordance with 18 U.S.C. Section 1734 solely toindicate this fact.
□S The on-line version of this article (available at http://www.jbc.org) containssupplemental Figs. 1 and 2.
1 To whom correspondence should be addressed: Lund University, Dept. ofExperimental Medical Science, BMC B12, SE-221 84 Lund, Sweden. Tel.:46-462228577; Fax: 46-462220855; E-mail: [email protected].
2 The abbreviations used are: SLRP, small leucine-rich repeat proteoglycan;LRR, leucine-rich repeat; GST, glutathione S-transferase; PBS, phosphate-buffered saline.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 282, NO. 22, pp. 16062–16067, June 1, 2007© 2007 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
16062 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 22 • JUNE 1, 2007
by guest on April 29, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on April 29, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on April 29, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on April 29, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on April 29, 2016
http://ww
w.jbc.org/
Dow
nloaded from

and transfected into BL21 Escherichia coli, and bacDCN56 pro-tein was expressed according to the manufacturer’s instruc-tions. bacDCN56 was purified under native conditions by glu-tathione affinity chromatography, as described previously (26),and dialyzed against PBS with 0.2% (v/v) Tween 20 prior to use.The protein concentration was determined with Coomassieprotein assay reagent (Pierce).Expression of His-tagged Fibromodulin in Bacteria—Bovine
fibromodulin cDNA (GenBank accession number X16485) wasused as a template in PCR to amplify fibromodulin cDNA. Theprimers used for the PCR were: 5�-CCGGATCCCCAATAT-GAGGAAGACTCTCAC-3� and 5�-CCCTCGAGGATCTC-GATGAGGCTAGCC-3�. The amplified cDNA was digestedwith XhoI and BamHI restriction enzymes and ligated into apET-27b(�) expression vector (Novagen), and the His-taggedprotein was expressed in Rosetta cells (Novagen). Fibromodu-lin was purified using nickel-nitrilotriacetic acid agarose (Qia-gen) according to the manufacturer’s instructions. Prior to use,the protein was dialyzed against PBS with 0.2% (v/v) Tween 20.Site-directed Mutagenesis—bacDCN56 cDNA and bovine
decorin cDNA (GenBank accession numberNM_173906) wereused for site-directedmutagenesis usingQuikChange II kit andpBluescript� II KS (�/�) cloning vector (both from Strat-agene) according to themanufacturer’s instructions.Mutationsof bacDCN56, used in bacterial protein expression, were thefollowing: E162Q, E181K, E181Q, T184V, E193Q, Y205F,R207S, D210N.Mutations of bovine decorin, used formamma-lian cell protein expression, were the following: E181K, R207S,and D210N. Primers used for mutagenesis are listed in supple-mental Fig. 1.Mammalian Expression of Wild Type Decorin and Mutated
Decorins—Wild type or mutated decorin cDNA was digestedwith the restriction enzymes HindIII and BamHI and ligatedinto the pCEP4 BM40-hisEK expression vector (27), attachingthe decorin cDNA to His tag DNA in the N terminus. Theconstructs were sequenced to confirm identity and transfectedinto human embryonic kidney 293 cells by electroporation, asdescribed (22). Cells were grown inDulbecco’smodified Eagle’smedium containing 10% (v/v) fetal calf serum (Invitrogen).After 24 h, hygromycin (InvivoGen) was added to a concentra-tion of 150 �g/ml. After 1 week, single clones were picked,incubated in medium containing [35S]sulfate, and analyzed forproteoglycan expression, as described (22). The proteoglycan-expressing clones were cultured in EX-Cell 325 PF Chinesehamster ovary protein-free medium (JRH Biosciences) for 12days with four medium changes every 3 days. The medium wasdialyzed against 300mMNaCl and 20mMNaH2PO4 buffer (pH8.0), and proteoglycanswere purified using nickel-nitrilotriace-tic acid agarose (Qiagen) and eluted with 250 mM imidazole in300 mM NaCl and 20 mM NaH2PO4 buffer (pH 8.0).Peptide Synthesis—Peptides SYIRIADTNIT and SYISIANT-
NIT were purchased from Schafer-N and dissolved in PBS with0.2% (v/v) Tween 20.Solution-phase Collagen Binding Assay—Collagen binding
assays were performed by the incubation of 15 �g of aceticacid-extractedmouse tail collagen in 100�l of PBS, pH7.4, withthe addition of different concentrations of radiolabeleddecorin, as described previously (17). Neutralization of acid sol-
ubilized collagen leads to the formation of collagen fibrils,which bind decorin. After incubation for 5 h at 37 °C, sampleswere centrifuged for 5 min at 13,000 � g to separate collagenfibrils with bound decorin in the pellet from non-bounddecorin in the supernatant. More than 90% of the collagen wasrecovered as fibrils in the pellet. The supernatant was removed,and the pellet was washed oncewith 1ml of PBS. The pellet andthe supernatant were then electrophoresed on a 10% SDS-PAGE, and the amount of radiolabeled proteoglycans wasdetermined in a FLA-3000 bioimaging analyzer (Fuji PhotoFilm Co.). The data were analyzed by non-linear regressionusing GraphPad Prism 3 software (GraphPad Software, Inc.).Solid-phase Collagen Binding Assay—Microtiter 96-well
Maxisorp plates (Nunc) were coated for 16 h using 10 �g/mlacetic acid-extracted mouse tail collagen in acetic acid. Thewells were washed with PBS and blocked for 1 h with 1 mg/mlbovine serum albumin in PBSwith 0.2% (v/v) Tween 20. After asingle wash using PBS with 0.5 mg/ml bovine serum albuminand 0.2% (v/v) Tween 20 (wash buffer), the GST-decorin fusionproteins were added, and after 3 h, the wells were washed fivetimes with the wash buffer. The amounts of GST fusion pro-teins bound to collagen were determined using 1-chloro-2,4-dinitrobenzene and reduced glutathione, as described previ-ously (28). A similar procedure was followed for assays withHis-tagged full-length decorin and decorin mutants, but thedetection was done with rabbit anti-His tag antibody ab9108(Abcam) diluted 1:250 in the wash buffer, swine anti-rabbitalkaline phosphatase-conjugated antibody (DAKO) diluted1:500 in the wash buffer, and detection with phosphatase sub-strate p-nitrophenyl phosphate (Sigma). For peptide inhibitionassays, the peptide in different concentrations was added todecorin or fibromodulin prior to incubation.Collagen Fibrillogenesis Assay—The assays were performed
as described previously (29). Pepsin-extracted acid-solubilizedcollagen (Vitrogen) was neutralized and diluted (167 pM) in 150mM NaCl, 20 mM HEPES, pH 7.4, with decorin or mutateddecorins at 4 °C. Solutions were degassed and then incubated at37 °C for 10 h in a spectrophotometer where the absorbancewas continuously recorded at 400 nm.
RESULTS
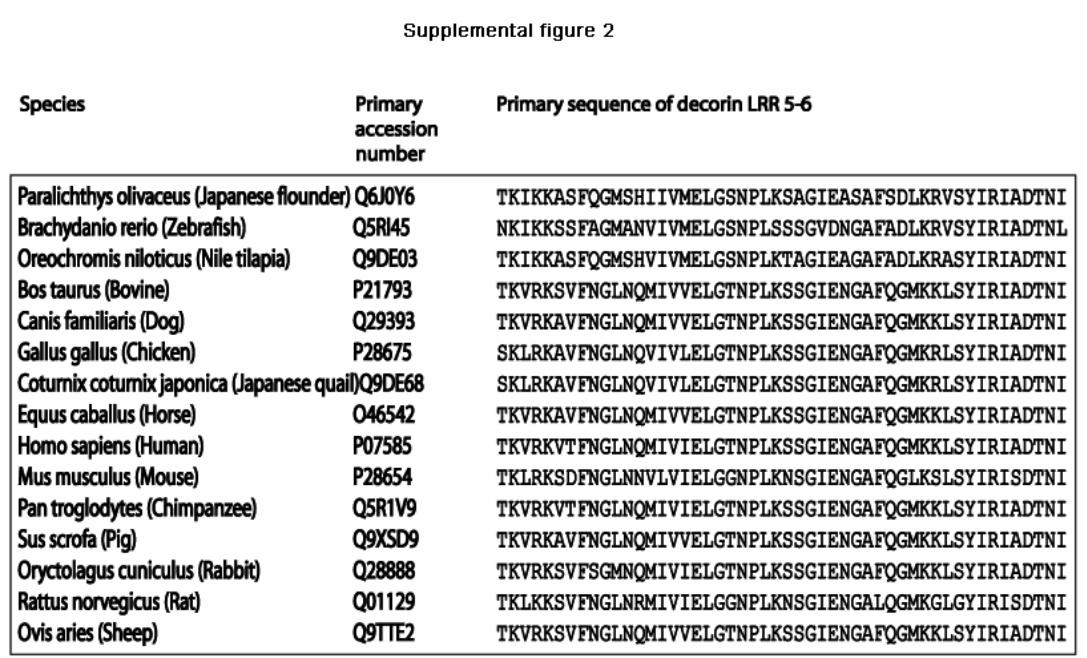
To study the interaction between decorin and collagen type I,we initially expressed decorin LRR 5–6, previously shown tobind collagen (22, 23), as a GST fusion protein in bacteria (bac-DCN56). This fusion protein inhibited the binding of mamma-lian-expressed full-length [35S]decorin to collagen in solution,indicating that they bind to the same site on collagen (Fig. 1).This confirms the collagen binding properties of bacDCN56,which was used in the further characterization of the decorin-collagen interaction. To identify the amino acid residuesinvolved in the interaction we performed site-directedmutagenesis. We selected residues in bacDCN56 that wereeither charged and conserved between species or polar andconserved between species, and also exposed on the exterior ofthe protein (Fig. 2). Thus, themutations E162Q, E181K, E181Q,T184V, E193Q, Y205F, R207S, and D210N were introducedinto bacDCN56 and expressed in E. coli. The binding of themutated decorin fragments was determined using a solid-phase
Decorin-Collagen Type I Interaction
JUNE 1, 2007 • VOLUME 282 • NUMBER 22 JOURNAL OF BIOLOGICAL CHEMISTRY 16063
by guest on April 29, 2016
http://ww
w.jbc.org/
Dow
nloaded from
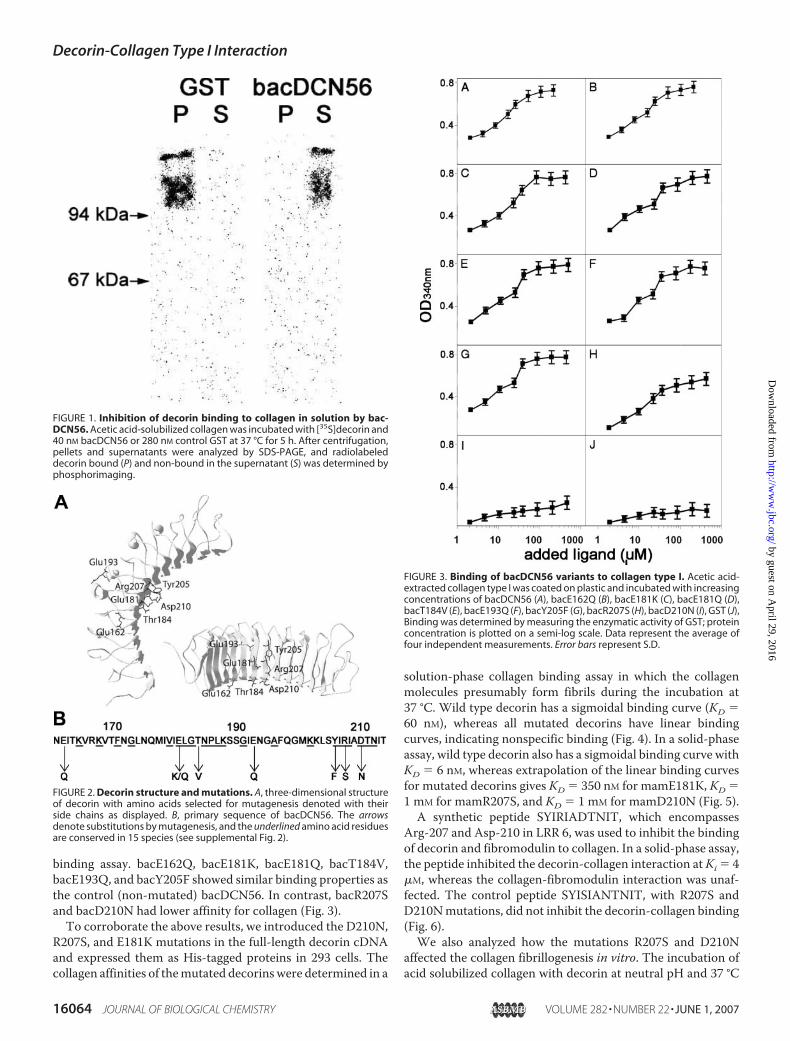
binding assay. bacE162Q, bacE181K, bacE181Q, bacT184V,bacE193Q, and bacY205F showed similar binding properties asthe control (non-mutated) bacDCN56. In contrast, bacR207Sand bacD210N had lower affinity for collagen (Fig. 3).To corroborate the above results, we introduced the D210N,
R207S, and E181K mutations in the full-length decorin cDNAand expressed them as His-tagged proteins in 293 cells. Thecollagen affinities of themutated decorinswere determined in a
solution-phase collagen binding assay in which the collagenmolecules presumably form fibrils during the incubation at37 °C. Wild type decorin has a sigmoidal binding curve (KD �60 nM), whereas all mutated decorins have linear bindingcurves, indicating nonspecific binding (Fig. 4). In a solid-phaseassay, wild type decorin also has a sigmoidal binding curve withKD � 6 nM, whereas extrapolation of the linear binding curvesfor mutated decorins gives KD � 350 nM for mamE181K, KD �1 mM for mamR207S, and KD � 1 mM for mamD210N (Fig. 5).
A synthetic peptide SYIRIADTNIT, which encompassesArg-207 and Asp-210 in LRR 6, was used to inhibit the bindingof decorin and fibromodulin to collagen. In a solid-phase assay,the peptide inhibited the decorin-collagen interaction atKi � 4�M, whereas the collagen-fibromodulin interaction was unaf-fected. The control peptide SYISIANTNIT, with R207S andD210Nmutations, did not inhibit the decorin-collagen binding(Fig. 6).We also analyzed how the mutations R207S and D210N
affected the collagen fibrillogenesis in vitro. The incubation ofacid solubilized collagen with decorin at neutral pH and 37 °C
FIGURE 1. Inhibition of decorin binding to collagen in solution by bac-DCN56. Acetic acid-solubilized collagen was incubated with [35S]decorin and40 nM bacDCN56 or 280 nM control GST at 37 °C for 5 h. After centrifugation,pellets and supernatants were analyzed by SDS-PAGE, and radiolabeleddecorin bound (P) and non-bound in the supernatant (S) was determined byphosphorimaging.
FIGURE 2. Decorin structure and mutations. A, three-dimensional structureof decorin with amino acids selected for mutagenesis denoted with theirside chains as displayed. B, primary sequence of bacDCN56. The arrowsdenote substitutions by mutagenesis, and the underlined amino acid residuesare conserved in 15 species (see supplemental Fig. 2).
FIGURE 3. Binding of bacDCN56 variants to collagen type I. Acetic acid-extracted collagen type I was coated on plastic and incubated with increasingconcentrations of bacDCN56 (A), bacE162Q (B), bacE181K (C), bacE181Q (D),bacT184V (E), bacE193Q (F), bacY205F (G), bacR207S (H), bacD210N (I), GST (J),Binding was determined by measuring the enzymatic activity of GST; proteinconcentration is plotted on a semi-log scale. Data represent the average offour independent measurements. Error bars represent S.D.
Decorin-Collagen Type I Interaction
16064 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 22 • JUNE 1, 2007
by guest on April 29, 2016
http://ww
w.jbc.org/
Dow
nloaded from

leads to a slower collagen fibril formation and a lower finalturbidity (1). 2 �g/ml decorin completely inhibited the forma-tion of fibrils and 1 �g/ml inhibited the formation of fibrils by�50% (Fig. 7). The incubation of collagen in the presence ofmamD210N at 2 and 1 �g/ml affected the fibrillogenesis onlymarginally. mamR207S at 2 �g/ml resulted in �50% inhibitionof fibrillogenesis. Also, the peptide SYIRIADTNIT at 150�g/ml lowered the turbidity significantly, whereas the controlpeptide at the same concentration had no effect.
DISCUSSION
We show that decorin LRR 5–6 domain, encompassing Arg-207 and Asp-210, is of importance for collagen binding. Muta-tions of these 2 amino acid residues impaired binding to recon-stituted collagen fibrils in solution and monomeric collagenadsorbed on plastic surface. Glu-181 (Glu-180 in humandecorin corresponds to Glu-181 in the bovine sequence) waspreviously reported to be essential for the collagen interaction
(23) and, interestingly, the E181K and E181Q mutationsexpressed as GST fusion proteins in bacteria did not impairbinding to monomeric collagen-coated surfaces. However,mammalian full-length E181K decorin lost the affinity for col-lagen. This discrepancy may be due to structural changesinduced by the E181K and E181Qmutations rather than directinvolvement of Glu-181 in collagen binding. The crystal struc-ture of decorin indicates that the side chain of Glu-181 poten-tially formshydrogen bondswith the side chains ofArg-207 andHis-159 on the neighboring LRR domains. Glu-181 also forms ahydrogen bond with the peptide bond of the proximal �-sheet(10). Consequently, mutation of this glutamate residue maylead to misfolding of the protein core, and collagen-bindingsites could be sterically hindered from interaction in full-lengthdecorin.Weused the programCUPSAT (30) to predict changes
FIGURE 4. Binding of radiolabeled variants of full-length decorin to colla-gen in solution. Increasing concentrations of decorin (f), mamE181K (Œ),mamR207S (E), and mamD210N (ƒ) were incubated with acetic acid-ex-tracted collagen in PBS for 5 h at 37 °C. Collagen-bound and non-bounddecorin fractions were separated by centrifugation, electrophoresed on SDS-PAGE, and quantified by phosphorimaging. The concentration of addedligand is plotted on a semi-log scale. Data were collected in three independ-ent experiments.
FIGURE 5. Solid phase collagen binding of mammalian-expressed full-length decorin variants. 96-well plastic plates were coated with acetic acid-extracted collagen type I and incubated with increasing concentrations ofwild type decorin (f), mamE181K (Œ), mamR207S (E), and mamD210N (ƒ).Binding was detected by anti-His antibody followed by alkaline phosphatase-conjugated antibody and phosphatase substrate. Background (withoutdecorins) was subtracted before plotting. Concentration of added ligand isplotted on a semi-log scale. Data represent the average of four independentmeasurements. Error bars represent S.D.
FIGURE 6. Inhibition of collagen-decorin interaction by synthetic pep-tides. 96-well plastic plates were coated with acid-extracted collagen type Iand incubated with mammalian-expressed decorin (A) or fibromodulin (B)and increasing concentrations of SYIRIADTNIT (Œ) and SYISIANTNIT (�).Bound decorin or fibromodulin was detected by anti-His antibody followedby alkaline phosphatase-conjugated antibody and phosphatase substrate.Background (without decorin or fibromodulin) was subtracted. Concentra-tion of added ligand is plotted on a semi-log scale. Data represent the averageof four independent measurements. Error bars represent S.D.
Decorin-Collagen Type I Interaction
JUNE 1, 2007 • VOLUME 282 • NUMBER 22 JOURNAL OF BIOLOGICAL CHEMISTRY 16065
by guest on April 29, 2016
http://ww
w.jbc.org/
Dow
nloaded from

in protein stability upon point mutations. The E181Kmutationhas a destabilized structure and an unfavorable torsion, withpredicted��G� �5.5 kcal/mol. Also, the R207Smutation hasan unfavorable ��G � �1.66 kcal/mol, which could indirectlyimpair collagen binding. In contrast, the D210N mutation has��G � 0.2 kcal/mol, not affecting the structure.The misfolding of decorin may also explain the reduction in
amount of E181K-mutated decorin synthesized by 293 cells asthe protein is presumably retained in the endoplasmic reticu-lum and not properly secreted. After screening of several clonesof 293 cells transformedwithmamE181K,we could not identifyclones expressing large amounts of mamE181K (data notshown). In bacE181K, only two LRRs are expressed, and thecollagen-binding site possibly remains exposed for interaction.The crystal structure of decorin indicates that the Asp-210 sidechain does not form intramolecular hydrogen bonds, has a highsolvent exposure (60%, according to CUPSAT analysis), and isnot involved in the putative dimer formation. On the otherhand, both Glu-181 and Arg-207 are less exposed to solvent (2and 22% respectively), buried within the putative dimer contactsurface, and have potential structure-stabilizing intramolec-ular hydrogen bonds. Our results indicate that Asp-210directly contributes to the collagen binding, whereas theArg-207 and Glu-181 may have structural functions. Indeed,sequence alignment of decorin LRR 5–6 sequences in 15species reveals conservation of these 3 amino acids, empha-sizing their structural role and involvement in collagen bind-ing (supplemental Fig. 2).Notably, amino acid residues in other LRR proteins involved
in interactions are positioned in the same region of the leucine-rich repeat as the decorin Asp-210; e.g.Asp-175 and Glu-128 ofglycoprotein Ib are critical for complex formation with vonWillebrand factor (31), andAsp-435 of ribonuclease inhibitor iscrucial for binding RNase A and angiogenin (32). In analogywith decorin, these charged amino acids are positioned in the�-sheet-loop region of the LRR domain, and the side chains areexposed to solvent. Similar location of binding sites can befound in other LRR proteins (33), implying conserved bindingpatterns.
The peptide sequence SYIRIADTNIT is located in LRR 6 ofdecorin and includes the residues Arg-207 and Asp-210. Thispeptide inhibits the decorin-collagen type I interaction at Ki �4 �M. Although this makes the peptide a potent inhibitor ofdecorin-collagen interaction, the inhibition is not complete.This implies the existence of other, seeminglyweaker, collagen-binding sites in decorin and agrees with previous reports (21,22, 24). Also, binding of mutated decorins to collagen indicatesthe presence of multiple binding sites in decorin (Figs. 4 and 5).Themutated decorins have linear binding curves, as opposed tosigmoidal binding curve of wild type decorin, which indicatesweak or nonspecific binding. Since it is conceivable that theglycosaminoglycan chain could also contribute to collagenbinding, we assessed the binding of chondroitinase ABC-treated decorin in the solid-phase assay and compared it withdecorin proteoglycan. Both decorins had similar collagen bind-ing, suggesting minor contribution by the glycosaminoglycanchain in collagen binding (data not shown).We also investigated how mutated decorins and synthetic
peptides affect collagen fibrillogenesis. It has been demon-strated that decorin inhibits the rate of collagen fibril formationin vitro and reduces the turbidity of the formed fibrils (1). It isbelieved that decorin binds collagen and interferes with thelateral growth of fibrils. The reduction in turbidity is primarilydue to the formation of thinner collagen fibrils in the presenceof decorin. Our results show that decorin dramatically inhibitsfibrillogenesis at a collagen:decorin molar ratio of 1:8 (collagenmolecular mass 300 kDa and decorin molecular mass 100 kDa)(Fig. 7).mamR207S also significantly inhibits fibrillogenesis at amolar ratio of 1:8, although not as efficiently as decorin. Thismay be explained by the collagen bindingmediated by Asp-210in mamR207S. Also, mamD210N retains some inhibition offibril formation, which indicates additional collagen-bindingsites, not involving Asp-210 and Arg-207. Interestingly, thesynthetic peptide SYIRIADTNIT also showed inhibition of col-lagen fibril formation when present at high concentrations. Acollagen:peptide molar ratio of 1:360 (collagen molecular mass300 kDa and peptide molecular mass 1.3 kDa) resulted in 25%inhibition of fibril formation. The control peptide SYISIANT-NIT did not interfere with the fibril formation. Apparently, thesynthetic peptide SYIRIADTNIT has the capacity to bind col-lagen and interfere with the lateral growth of fibrils.The collagen-binding site of decorin is homologous to
sequences in asporin and biglycan, which are the closest rela-tives of decorin and belong to the same SLRP subfamily. Thedecorin sequence SYIRIADTNIT is partially conserved in theasporin sequence HIRIAEAKLT and in the biglycan sequenceNYLRISEAKLT. The sequences are located in LRR 6, and theircollagen binding properties are unknown.
REFERENCES1. Vogel, K. G., Paulsson, M., and Heinegard, D. (1984) Biochem. J. 223,
587–5972. Fleischmajer, R., Fisher, L. W., MacDonald, E. D., Jacobs, L., Perlish, J. S.,
and Termine, J. D. (1991) J. Struct. Biol. 106, 82–903. Krusius, T., and Ruoslahti, E. (1986) Proc. Natl. Acad. Sci. U. S. A. 83,
7683–76874. Iozzo, R. V. (1999) J. Biol. Chem. 274, 18843–188465. Fisher, L. W., Termine, J. D., and Young, M. F. (1989) J. Biol. Chem. 264,
FIGURE 7. Collagen fibrillogenesis assay. Pepsin-extracted acid solubilizedcollagen was neutralized and incubated at 37 °C in the presence of decorin(A), mamR207S (B), mamD210N (C), and synthetic peptides (D). Decorin,mamR207S, and mamD210N were present at 1 �g/ml (E) and 2 �g/ml (‚).Synthetic peptide SYIRIADTNIT was present at 10 �g/ml (F) and at 150 �g/ml(Œ). Control peptide SYISIANTNIT was present at 150 �g/ml (�). Control wasperformed with collagen alone (f). Turbidity at 400 nm was measured con-tinuously at 4-min intervals and plotted versus elapsed time.
Decorin-Collagen Type I Interaction
16066 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 22 • JUNE 1, 2007
by guest on April 29, 2016
http://ww
w.jbc.org/
Dow
nloaded from

4571–45766. Oldberg, Å., Antonsson, P., Lindblom, K., and Heinegard, D. (1989)
EMBO J. 8, 2601–26047. Blochberger, T., Vergnes, J.-P., Hempel, J., and Hassel, J. (1992) J. Biol.
Chem. 267, 347–3528. Lorenzo, P., Aspberg, A., Onnerfjord, P., Bayliss, M. T., Neame, P. J., and
Heinegard, D. (2001) J. Biol. Chem. 276, 12201–122119. Henry, S. P., Takanosu, M., Boyd, T. C., Mayne, P. M., Eberspaecher, H.,
Zhou, W., de Crombrugghe, B., Hook, M., and Mayne, R. (2001) J. Biol.Chem. 276, 12212–12221
10. Scott, P. G., McEwan, P. A., Dodd, C. M., Bergmann, E. M., Bishop, P. N.,and Bella, J. (2004) Proc. Natl. Acad. Sci. 101, 15633–15638
11. Ameye, L., and Young, M. F. (2002) Glycobiology 12, 107R–116R12. Danielson, K. G., Baribault, H., Holmes, D. F., Graham, H., Kadler, K. E.,
and Iozzo, R. V. (1997) J. Cell Biol. 136, 729–74313. Xu, T., Bianco, P., Fisher, L. W., Longenecker, G., Smith, E., Goldstein, S.,
Bonadio, J., Boskey, A., Heegaard, A. M., Sommer, B., Satomura, K.,Dominguez, P., Zhao, C., Kulkami, A. B., Robey, P. G., and Young, M. F.(1998) Nat. Genet. 20, 78–82
14. Svensson, L., Aszodi, A., Reinholt, F. P., Fassler, R., Heinegard, D., andOldberg, A. (1999) J. Biol. Chem. 274, 9636–9647
15. Chakravarti, S., Magnuson, T., Lass, J. H., Jepsen, K. J., LaMantia, C., andCarroll, H. (1998) J. Cell Biol. 141, 1277–1286
16. Keene, D. R., San Antonio, J. D., Mayne, R., McQuillan, D. J., Sarris, G.,Santoro, S. A., and Iozzo, R. V. (2000) J. Biol. Chem. 275, 21801–21804
17. Bidanset, D. J., Guidry, C., Rosenberg, L. C., Choi, H. U., Timpl, R., andHook, M. (1992) J. Biol. Chem. 267, 5250–5256
18. Hildebrand, A., Romaris, M., Rasmussen, L. M., Heinegard, D., Twardzik,
D. R., Border, W. A., and Ruoslahti, E. (1994) Biochem. J. 302, 527–53419. Schmidt, G., Hausser, H., and Kresse, H. (1991) Biochem. J. 280, 411–41420. Iozzo, R. V., Moscatello, D. K., McQuillan, D. J., and Eichstetter, I. (1999)
J. Biol. Chem. 274, 4489–449221. Schonherr, E., Hausser, H., Beavan, L., and Kresse, H. (1995) J. Biol. Chem.
270, 8877–888322. Svensson, L., Heinegard, D., and Oldberg, A. (1995) J. Biol. Chem. 270,
20712–2071623. Kresse, H., Liszio, C., Schonherr, E., and Fisher, L.W. (1997) J. Biol. Chem.
272, 18404–1841024. Nareyeck, G., Seidler, D. G., Troyer, D., Rauterberg, J., Kresse, H., and
Schonherr, E. (2004) Eur. J. Biochem. 271, 3389–339825. Liu, X., Yeh, M. I., Lewis, J. L., and Luo, Z. P. (2005) Biochem. Biophys. Res.
Commun. 338, 1342–134526. Frangioni, J. V., and Neel, B. G. (1993) Anal. Biochem. 210, 179–18727. Bengtsson, E., Aspberg, A., Heinegard, D., Sommarin, Y., and Spillmann,
D. (2000) J. Biol. Chem. 275, 40695–4070228. Habig, W. H., Pabst, M. J., and Jakoby, W. B. (1974) J. Biol. Chem. 249,
7130–713929. Kvist, A. J., Johnson, A. E., Morgelin, M., Gustafsson, E., Bengtsson, E.,
Lindblom, K., Aszodi, A., Fassler, R., Sasaki, T., Timpl, R., and Aspberg, A.(2006) J. Biol. Chem. 281, 33127–33139
30. Parthiban, V., Grohima, M. M., and Schomburg, D. (2006) Nucleic AcidsRes. 34,W239–42
31. Shimizu, A.,Matsushita, T., Kondo, T., Inden, Y., Kojima, T., Saito, H., andHirai, M. (2004) J. Biol. Chem. 279, 16285–16294
32. Chen, C. Z., and Shapiro, R. (1997) Proc. Natl. Acad. Sci. 94, 1761–176633. Kobe, B., and Kajava, A. V. (2001) Curr. Opin. Struct. Biol. 11, 725–732
Decorin-Collagen Type I Interaction
JUNE 1, 2007 • VOLUME 282 • NUMBER 22 JOURNAL OF BIOLOGICAL CHEMISTRY 16067
by guest on April 29, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Synthetic peptide SYIRIADTNIT at 10 µg/ml ( ) and 150 µg/ml ( ). Control peptide SYISIANTNIT at 150 µg/ml (X). Control with collagen alone ( ). Turbidity at 400 nm was measured continuously at 4 min intervals and plotted versus elapsed time.
SUPPLEMENTAL FIGURES
Supplemental Figure 1. Primers used for site-directed mutagenesis of bacDCN56 and bovine full-length decorin. Supplemental Figure 2. Sequence alignment of decorin LRR 5-6 in 15 species.
8


Sebastian Kalamajski, Anders Aspberg and Åke OldbergThe Decorin Sequence SYIRIADTNIT Binds Collagen Type I
doi: 10.1074/jbc.M700073200 originally published online April 10, 20072007, 282:16062-16067.J. Biol. Chem.
10.1074/jbc.M700073200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2007/04/11/M700073200.DC1.html
http://www.jbc.org/content/282/22/16062.full.html#ref-list-1
This article cites 33 references, 26 of which can be accessed free at
by guest on April 29, 2016
http://ww
w.jbc.org/
Dow
nloaded from