collagen reorganization in leech wound healing
TRANSCRIPT

Biol. Cell (2005) 97, 557–568 (Printed in Great Britain) Research article
Collagen reorganization in leechwound healingGianluca Tettamanti*, Annalisa Grimaldi*, Terenzio Congiu†, Gianpaolo Perletti‡, Mario Raspanti†,Roberto Valvassori* and Magda de Eguileor*1
*Department of Structural and Functional Biology, University of Insubria, Via J.H. Dunant 3, 21100 Varese, Italy, †Department of Human
Morphology, University of Insubria, Via Monte Generoso 71, 21052 Varese, Italy, and ‡Department of Structural and Functional Biology,
University of Insubria, Via A. da Giussano 12, 21052 Busto Arsizio, Italy
Background Information. Leeches respond to surgical lesions with the same sequence of events as that describedfor wound healing in vertebrates, where collagen is important for the development of tensions in healing wounds,functioning as an extracellular scaffold for accurate regeneration of the structures disrupted by surgical or traumaticactions.
Results. In surgically lesioned leeches, newly synthesized collagen is arranged in hierarchical structures. Fibrilscan be packed and shaped to form cords or tubular structures, thus acting as an extracellular scaffold that directsand organizes the outgrowth of new vessels and the migration of immune cells towards lesioned tissues. In theseanimals, the general architecture of collagen fibrils, generated during tissue regeneration, shows similarities to boththe structural pattern of collagen bundles and assembly processes observed in several vertebrate systems (fishscales, amphibian skin and human cornea).
Conclusions. The production of extracellular matrix during wound healing in leeches is a surprising example ofconservation of an extremely close relationship between the structure and function of molecular structures. It couldbe hypothesized that collagen structures, characterized not only by a striking structural complexity, but also bymultifunctional purposes, are anatomical systems highly conserved throughout evolution.
IntroductionCollagen is a generic term that covers a number ofproteins of diverse molecular mass and of hetero-geneous structure. In humans, about 26 geneticallydistinct types of collagen have been characterized(Babel and Glanville, 1984; Kadler et al., 1996).These molecules are synthesized and secreted by dif-ferent types of cells to form a complex, multifunc-tional ECM (extracellular matrix) (Chiquet et al.,1996; Frazier et al., 1996; Kletsas et al., 2000;Chiquet et al., 2003).
In fact, collagen represents not only a structuralcomponent of the ECM but also plays a major rolein the modulation of several cell functions, includ-ing adhesion, migration, growth and differentiation
1To whom correspondence should be addressed ([email protected]).Key words: atomic force microscopy (AFM), collagen, leech injury, scanningelectron microscopy (SEM), transmission electron microscopy (TEM).Abbreviations used: AFM, atomic force microscopy; ChABC, chondroitinaseABC; ECM, extracellular matrix; SEM, scanning electron microscopy; TEM,transmission electron microscopy; TMAFM, tapping-mode AFM.
(Hay, 1991; Birk and Zycband, 1994; Lim et al.,1994).
In vertebrates, the major types of fibrillar collagens,i.e. types I–III, V and X, are fundamental for ensuringoptimal mechanical properties of tissues such as ten-don, derma, cornea, cartilage and bone (Vilarta andDe Campos Vidal, 1989; Cheung et al., 1990; Birkand Zycband, 1994; Niederreither et al., 1994;Birk et al., 1996; Christiansen et al., 2000; Hirschet al., 2001; Ottani et al., 2001, 2002; Raspanti et al.,2002; Boote et al., 2003). These characteristics areachieved through optimal organization and inter-action between individual collagen fibrils as well asbetween fibres. In addition, fibrillar collagens are alsoinvolved in numerous processes, including stabiliz-ation of tissue shape and form during both verteb-rate development and tissue regeneration (Birk andTrelstad, 1984; Birk et al., 1989; Ingber, 1994;Niederreither et al., 1994; Frazier et al., 1996;Kletsas et al., 2000; Suzuma et al., 2000; Badylak,2002).
www.biolcell.org | Volume 97 (7) | Pages 557–568 557
G. Tettamanti and others
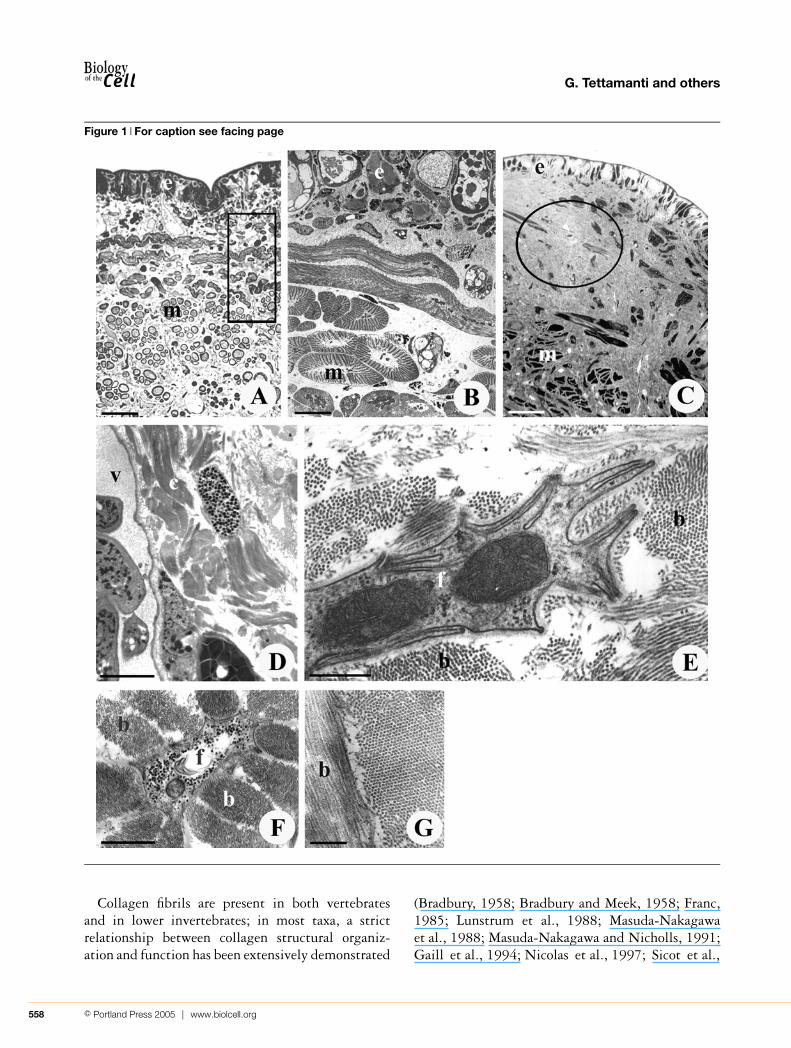
Figure 1 For caption see facing page
Collagen fibrils are present in both vertebratesand in lower invertebrates; in most taxa, a strictrelationship between collagen structural organiz-ation and function has been extensively demonstrated
(Bradbury, 1958; Bradbury and Meek, 1958; Franc,1985; Lunstrum et al., 1988; Masuda-Nakagawaet al., 1988; Masuda-Nakagawa and Nicholls, 1991;Gaill et al., 1994; Nicolas et al., 1997; Sicot et al.,
558 C© Portland Press 2005 | www.biolcell.org

Leech collagen reorganization Research article
Figure 1 Unlesioned and lesioned H. medicinalis: optical microscopy and TEMSemi-thin (A) and the respective (framed area) thin (B) cross-sections of the body of an unlesioned H. medicinalis. The body
wall of a control animal (A) is mainly composed of tightly packed helical muscle fibres (m) (easily recognizable due to their
peculiar cross-sections, showing a cytoplasmic core surrounded by a ring of contractile material) embedded in a scarce
and loose connective tissue, wrapped by a monolayered epithelium (e). Scale bars: (A) 25 µm and (B) 3 µm. (C, D) Semi-thin
(C) and thin (D) cross-sections of the body wall of an H. medicinalis subjected to surgical explant (C). This sample was processed
4 weeks after surgery. The regenerating tissue is characterized by a large amount of newly synthesized collagen (C, c; encircled
area) located under the epithelium (e) and among muscle fibres (m), forming a scaffold (D) to drive and direct the growth of new
blood vessels (v). Scale bars: (C) 50 µm and (D) 3 µm. (E, F) Thin sections of leech granulation tissue. Longitudinally sectioned
(E) and cross-sectioned (F) activated fibroblasts (f), showing cytoplasmic laminae lining extracellular microenvironments involved
in the organization of fibril bundles (b). Scale bars: (E) 0.5 µm and (F) 1 µm. (G) Thin section of newly synthesized collagen bundles
(b) orthogonally orientated. This general architecture of leech collagen bundles is identical with the spatial organization of collagen
bundles in vertebrates. Scale bar, 0.3 µm.
1997; Cluzel et al., 2000; Trotter et al., 2000;Ottani et al., 2002; Koch et al., 2003). In this con-text, it is important to emphasize that basic studies onnon-mammalian models are extensively performed tounderstand better the wound healing process and, inparticular, the granulation tissue stage, characterizedby a large synthesis of collagen.
We have previously demonstrated that Hirudo medi-cinalis (Annelida, Hirudinea) could be considered as agood animal model for two major reasons (de Eguileoret al., 2001, 2004). First of all, tissue repair in leechesshows a high degree of similarity to wound healingprocesses occurring in vertebrates, both under thestructural–functional and the biochemical points ofview. Secondly, the body of the leech shows a simpleorganization. Under the epithelium, the musclelayers are made of grouped fibres separated by scarceECM. The muscular sac contains several organs andcharacteristic tissues, i.e. botryoidal and vasofibroustissues (Sawyer, 1986), embedded in a loose connect-ive tissue characterized by a few widely spread fibro-blasts and scarce collagen fibres (Sawyer, 1986;de Eguileor et al., 1999; Tettamanti et al., 2004).
The wound healing process in leeches can be di-vided, as in vertebrates, into three different stages:(i) inflammation, (ii) granulation tissue and (iii)scar tissue remodelling. In leeches, the granulationtissue stage is characterized by re-epithelialization,angiogenesis and fibroplasia. During these steps,the formation of new epithelium occurs, followedby the formation of a network of blood vessels span-ning the entire body wall and then massive produc-tion of connective tissues that can be used as a scaf-fold not only for tissue reconstruction but also formigration of immune cells and for proper orientation
of growth of new vessels (de Eguileor et al., 2004;Tettamanti et al., 2004).
In the present study, different approaches [light mi-croscopy, AFM (atomic force microscopy), SEM (scan-ning electron microscopy), TEM (transmission elec-tron microscopy), immunocytochemical studies andselective enzymatic digestions] were used not onlyto characterize the synthesis of new collagens in re-sponse to surgical lesions in the leech animal model(Tettamanti et al., 2003), but also to evaluate howa newly formed collagen reaches its peculiar spatialreorganization.
Our results contribute to the characterization of thefunctional role of this specialized ECM; the strikingsimilarities between leech collagen matrix, assembledduring tissue regeneration, and the structure of thevertebrate collagen matrix (in different tissues) leadsus to hypothesize the recruitment of structures ac-quired along the evolutionary scale for specializedtissue structures and functions in higher taxa.
ResultsRole of the collagen matrix in the body wall ofunlesioned and lesioned leechesThe body wall of unlesioned H. medicinalis is charac-terized by a monolayered epithelium surrounding athick muscular wall made of helical fibres tightlypacked, embedded in scarce and loose connective tis-sues characterized by a few widely spread fibroblasts(Sawyer, 1986; de Eguileor et al., 1999; Tettamantiet al., 2004). In control leeches, the quantity of fibril-lar collagen is small and a few collagen fibres are vis-ible among muscle fibres, under the epithelium andin the space surrounding the gut (Figures 1A and 1B).
www.biolcell.org | Volume 97 (7) | Pages 557–568 559

G. Tettamanti and others
When leeches were subjected to an experimentalsurgical lesion, the gross organization of the bodywall markedly changed. In fact, the tissue regener-ation process was characterized by the proliferationof fibroblasts, major producers of the extracellularmatrix (Tettamanti et al., 2004). During the firstdays of the healing process, maturation of new col-lagen fibrils, easily detectable due to their supra-molecular organization, led to the formation of a ro-bust, solid extracellular network (Figure 1C). Thisnew collagenic scaffold, ‘infiltrated’ among musclefibres, directed and regulated cell migration and newvessels crawling from the gut towards the lesionedarea (Figure 1D).
After a surgical lesion, the morphology of leech-activated fibroblasts resembled that of the secret-ing fibroblasts of vertebrates. Secreting fibroblastsof H. medicinalis are characterized by projections ofcytoplasmic laminae stretching towards the extracel-lular space (Tettamanti et al., 2004). The cytoplasmicfoldings not only formed the microenvironment inwhich fibrillogenesis occurred but were also involvedin the spatial organization of collagen bundles (Fig-ures 1E and 1F), governing the orientation of the col-lagen fibrils with respect to the cell axis (Figures 1E,1F and 1G).
Spatial organization of collagen fibrilsIn samples of lesioned leeches, examined 24 h aftersurgery, newly synthesized and secreted collagen wasorganized into tightly packed, hexagonally arrangedfibrils of approx. 28 nm in diameter; these fibrilswere interconnected to one another by transverse pro-teoglycan bridges of approx. 14 nm, forming a latticeperpendicular to the fibril axis (Figures 2 and 3).
In each fibril, the collagen molecules lying around atranslucent central core (Figure 3) formed an annulusof approx. 7 nm thickness.
During collagen maturation, the diameter offibrils increased from 20 to 28 nm: this effect couldbe easily detected in each microenvironment lined bylaminae, by comparing fibrils lying in the extracel-lular juxtamembrane area of fibroblasts with fibrilslocated at a higher distance from the cell membrane(Figure 3).
Spatial organization of collagen bundlesIn surgically lesioned leeches, newly synthesizedand secreted collagen fibrils were regularly arranged
Figure 2 Drawing to explain the spatial organization ofcollagen fibrilsSchematic representation to describe, in lesioned leech, the
spatial organization of collagen fibrils interconnected by pro-
teoglycan bridges.
Figure 3 Spatial organization of fibrils(A) Thin sections of a newly synthesized collagen bundle.
Collagen epitaxial fibrils are hexagonally packed and inter-
connected by proteoglycan bridges (arrowhead) that form a
lattice perpendicular to the fibril axis (see Figure 2). Fibrils
grow up to a final diameter of 28 nm from the pericellular area
of fibroblasts (f). Scale bar, 0.15 µm.
in parallel, forming bundles. As observed by AFM(Figure 4A), the surface of each ‘rope’ showed a pre-cise three-dimensional alignment: fibrils were dis-posed in parallel and bundled up by a few associatedfibrils disposed at different orientations (Figure 4A).
560 C© Portland Press 2005 | www.biolcell.org

Leech collagen reorganization Research article
Figure 4 Three-dimensional organization of fibrils and bundles: optical, AFM and SEM(A) AFM image of the surface of a collagenic bundle (b) in the wounded area of a surgically lesioned leech. The sample was
processed 24 h after surgery. Collagen fibrils forming this bundle run parallel to each other. A few fibrils (arrowheads) are
positioned perpendicular to the main axis of the bundle (arrow). Scale bar, 0.1 µm. (B, C) SEM micrographs of a collagen
‘scaffold’ made of fibril bundles (b), showing rope-like (B) or tube-like (C) shapes. The collagenic tubes (C) are made of several
bundles (b). Scale bars: (B) 3 µm and (C) 1.5 µm. (D) Semi-thin tangential section of the regenerating area of surgically lesioned
H. medicinalis. Newly synthesized collagen bundles are orientated, forming layers at approx. 50◦ with respect to the main axis
of the leech body. Scale bar, 20 µm.
When the newly synthesized stroma was observedunder the transmission electron microscope, adjacentbundles were orientated orthogonally to one another(Figure 1G). When observed under the scanning elec-tron microscope, this collagen scaffold was charac-terized by bundles of fibrils assembled in differentways, i.e. forming either a compact cluster of colla-
gen bundles (Figure 4B) or a cylinder (Figure 4C).The collagenic cylinder or ‘tube’ was the mostcomplex spatial organization observed in the newlyforming collagen matrix (Figure 4C), because it wasthe result of a simultaneous synthesis of several (5–10)collagenic bundles involving several microenviron-ments all around the secreting fibroblast (Figure 1F).
www.biolcell.org | Volume 97 (7) | Pages 557–568 561
G. Tettamanti and others
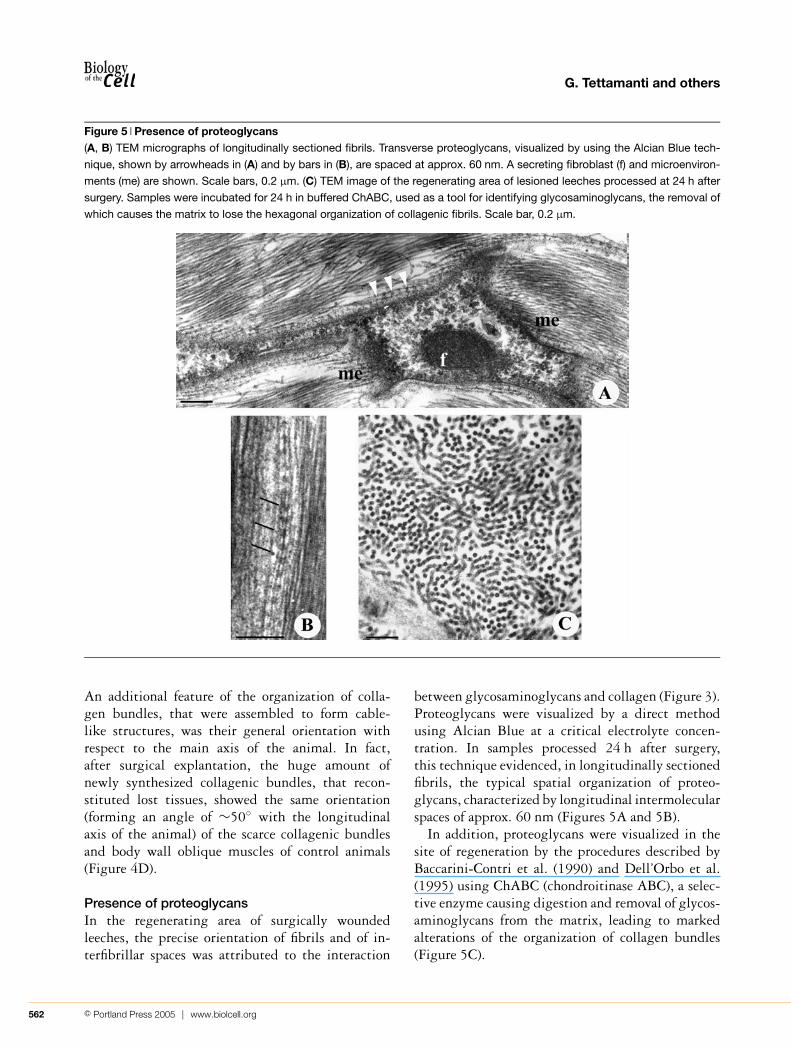
Figure 5 Presence of proteoglycans(A, B) TEM micrographs of longitudinally sectioned fibrils. Transverse proteoglycans, visualized by using the Alcian Blue tech-
nique, shown by arrowheads in (A) and by bars in (B), are spaced at approx. 60 nm. A secreting fibroblast (f) and microenviron-
ments (me) are shown. Scale bars, 0.2 µm. (C) TEM image of the regenerating area of lesioned leeches processed at 24 h after
surgery. Samples were incubated for 24 h in buffered ChABC, used as a tool for identifying glycosaminoglycans, the removal of
which causes the matrix to lose the hexagonal organization of collagenic fibrils. Scale bar, 0.2 µm.
An additional feature of the organization of colla-gen bundles, that were assembled to form cable-like structures, was their general orientation withrespect to the main axis of the animal. In fact,after surgical explantation, the huge amount ofnewly synthesized collagenic bundles, that recon-stituted lost tissues, showed the same orientation(forming an angle of ∼50◦ with the longitudinalaxis of the animal) of the scarce collagenic bundlesand body wall oblique muscles of control animals(Figure 4D).
Presence of proteoglycansIn the regenerating area of surgically woundedleeches, the precise orientation of fibrils and of in-terfibrillar spaces was attributed to the interaction
between glycosaminoglycans and collagen (Figure 3).Proteoglycans were visualized by a direct methodusing Alcian Blue at a critical electrolyte concen-tration. In samples processed 24 h after surgery,this technique evidenced, in longitudinally sectionedfibrils, the typical spatial organization of proteo-glycans, characterized by longitudinal intermolecularspaces of approx. 60 nm (Figures 5A and 5B).
In addition, proteoglycans were visualized in thesite of regeneration by the procedures described byBaccarini-Contri et al. (1990) and Dell’Orbo et al.(1995) using ChABC (chondroitinase ABC), a selec-tive enzyme causing digestion and removal of glycos-aminoglycans from the matrix, leading to markedalterations of the organization of collagen bundles(Figure 5C).
562 C© Portland Press 2005 | www.biolcell.org
Leech collagen reorganization Research article
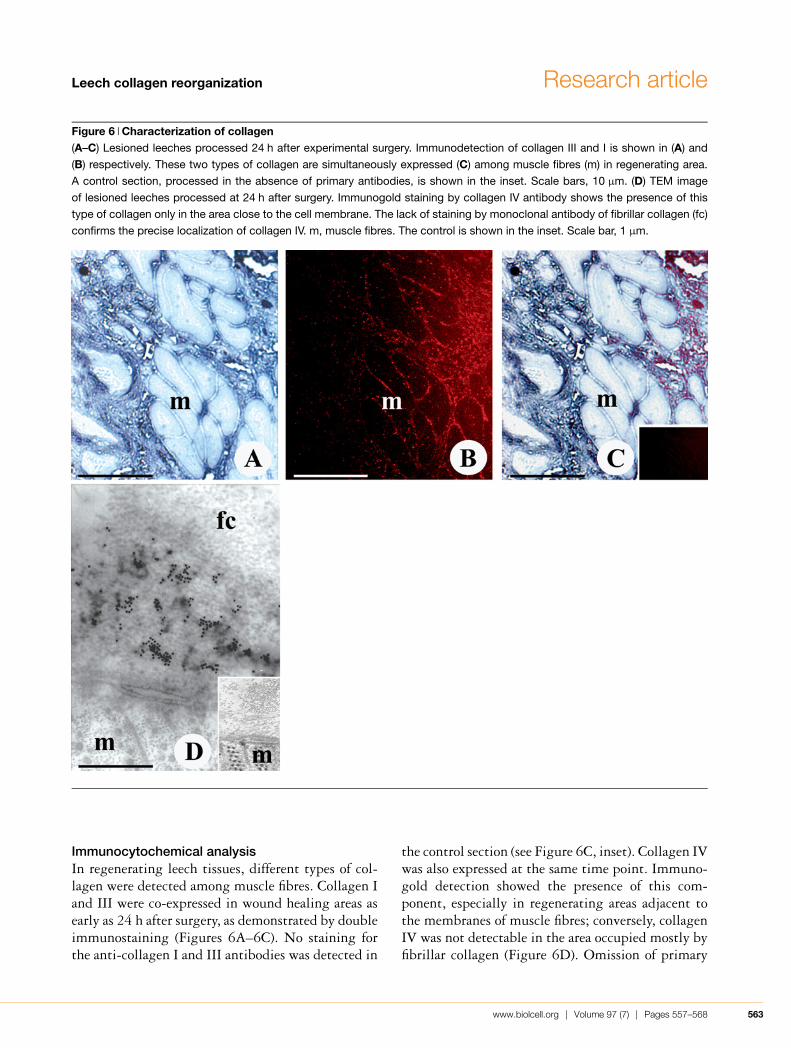
Figure 6 Characterization of collagen(A–C) Lesioned leeches processed 24 h after experimental surgery. Immunodetection of collagen III and I is shown in (A) and
(B) respectively. These two types of collagen are simultaneously expressed (C) among muscle fibres (m) in regenerating area.
A control section, processed in the absence of primary antibodies, is shown in the inset. Scale bars, 10 µm. (D) TEM image
of lesioned leeches processed at 24 h after surgery. Immunogold staining by collagen IV antibody shows the presence of this
type of collagen only in the area close to the cell membrane. The lack of staining by monoclonal antibody of fibrillar collagen (fc)
confirms the precise localization of collagen IV. m, muscle fibres. The control is shown in the inset. Scale bar, 1 µm.
Immunocytochemical analysisIn regenerating leech tissues, different types of col-lagen were detected among muscle fibres. Collagen Iand III were co-expressed in wound healing areas asearly as 24 h after surgery, as demonstrated by doubleimmunostaining (Figures 6A–6C). No staining forthe anti-collagen I and III antibodies was detected in
the control section (see Figure 6C, inset). Collagen IVwas also expressed at the same time point. Immuno-gold detection showed the presence of this com-ponent, especially in regenerating areas adjacent tothe membranes of muscle fibres; conversely, collagenIV was not detectable in the area occupied mostly byfibrillar collagen (Figure 6D). Omission of primary
www.biolcell.org | Volume 97 (7) | Pages 557–568 563

G. Tettamanti and others
antibody revealed no gold-labelled structures (Fig-ure 6D, inset).
DiscussionLeeches respond to surgical lesions with the samesequence of wound healing and tissue regenerationevents as that described for vertebrates (Tettamantiet al., 2003). The goals of the present study were todescribe and characterize the collagen matrix synthes-ized during tissue repair events after experimentalsurgery and to investigate the relationship betweenthe structure and properties of collagen in regenerat-ing leech tissues.
Although collagen IV is highly expressed in regen-erating leech tissues, fibrillar collagens (types I andIII) are the main components of the connective tis-sues in wound healing sites. It may be hypothesizedthat the simultaneous presence of these two types ofcollagen is due to the generation of heterotypic fibrils(containing type I and III collagens). Alternatively,the presence of a large amount of type III collagenin adult regenerating tissues could be related to theongoing wound healing process as shown by Barneset al. (1976) in guinea-pig dermal skin.
Fibrillar collagen, in leeches as well as in ver-tebrates, is essential for the preservation of shape inhealing wounds, because it functions as an extracel-lular scaffold for the accurate regeneration of struc-tures disrupted by surgery or traumatic events. Fromour results, it appears that H. medicinalis collagenfibrillogenesis shares striking similarities with thesame process occurring in vertebrates (Trelstad andHayashi, 1979; Birk and Trelstad, 1984; Scottand Bosworth, 1990). This process takes place in anextracellular compartment defined by fibroblastmembrane expansions (Tettamanti et al., 2004).Mature fibrils, grown up to their final diameter of ap-prox. 28 nm, are connected by proteoglycan bridgesof approx. 14 nm length, thus forming a lattice per-pendicular to the fibril axis, as shown by the AlcianBlue technique (Figures 5A and 5B). This latticehas the function of maintaining the permanent axialalignment as well as the regular lateral separation offibrils (Figure 2).
It is interesting to note that the general archi-tecture of the collagen components of regeneratingleech tissues is identical with the structural patternof collagen bundles observed in several vertebrate
tissues such as petromyzon skin, fish scale, amphi-bian skin and human cornea (Weiss and Ferris, 1956;Hirsch et al., 2001; Ottani et al., 2002; Ikoma et al.,2003). It is remarkable that the structural organiz-ation of the leech collagen is the same as that ofhuman cornea; in both cases, collagen is assembledusing a strikingly similar process, leading to identicalfibril organization and bundle orientation (Miyagawaet al., 2000; Boote et al., 2003). In the fibro-blasts of H. medicinalis (Tettamanti et al., 2004) aswell as in mammalian corneal fibroblasts (Birk andTrelstad, 1984), cell-surface foldings exert an im-portant role by positioning and packaging collagenfibrils within a defined, regular spatial organiz-ation, to build flexible bundles of parallel elementswithin the extracellular microenvironment. Inlesioned leeches, newly synthesized collagen is ar-ranged in hierarchical structures: fibrils (the func-tional units of the collagen scaffold) of constant dia-meter are tightly packed in hexagonal patterns andsubsequently cross-linked to one another. Thus fibrilsform a sort of strong multithreaded cable (surface andthree-dimensional view of which are visible respect-ively under an AFM and SEM) that can be furtherassembled in different ways after being packed andshaped to form a cluster of collagen bundles or a hol-low cylinder, the collagen ‘tube’ (the entire processis schematically represented in Figure 7). The tuberepresents the highest organization level of collagenfibres, and functions as a strong support structure. It isused as a scaffold to direct and organize the outgrowthof new vessels and the migration of immune cells to-wards lesioned areas of the animal body. We specu-late that collagen tubes, displaying a complexorganization, are characterized by peculiar mechan-ical properties responsible for the construction of alight and solid structure. The wall of each tube con-sists of several bundles of fibrils. The subdivisionof a collagen tube into multiple parallel bundles ofgrouped threads of equivalent cross-sectional sur-face does not influence its final strength, but shouldprovide several advantages according to mechanicalproperties (Parry and Craig, 1984; Gordon, 1988;Fratzl et al., 1998; Ottani et al., 2001; Gutsmannet al., 2003): (i) the subdivision of the tube into dis-tinct, easy-to-replace units increases structural resist-ance and safety; (ii) tubular structures are character-ized by high flexibility, which is directly proportionalto the thread number but inversely proportional to
564 C© Portland Press 2005 | www.biolcell.org

Leech collagen reorganization Research article
Figure 7 Sketch to explain the collagen fibrils disposition in control and explanted leechesDiagram representing the sequence of events that leads to the formation and spatial organization of fibrillar collagen in
non-explanted (control) and explanted H. medicinalis. Left panel: in control leech, the fibroblast (green) secretes a few col-
lagenic fibrils. In a cross-section of the body wall, the scarce connective tissue is loosely distributed among epithelium and
muscle fibres. Right panel: in surgically explanted leech, the activating fibroblast (green) synthesizes a large amount of collagenic
fibrils geometrically organized in bundles. Bundles form a solid cord or tube structures filling the explant zone.
the radius of each thread; and (iii) the tube structureis lighter and metabolically ‘cheaper’ than a solidcylinder.
As far as the inner architecture of each collagenbundle is concerned, it is possible to observe thatleech epitaxial fibrils, characterized by collagen mol-ecules laid upon a central electron-lucent core, areuniform in diameter and show a constant and uniforminterfibrillar space, ensured by proteoglycan bridges.Such an organization enables the mechanical coupl-ing of fibrils, which can resist compressive forcesand distribute the mechanical stress throughout thewhole tissue. Moreover, the above-described spatialdistribution enables the leech tissue to resist not only
compressive forces but also, because they were shapedas a ‘twisted strand’ (like a cable or rope), longitud-inal traction. As a consequence of these features, col-lagen tubes are optimally suited to withstand tensilestresses during wound healing and repair processes. Inaddition, in wounded areas of leeches, newly synthes-ized collagen tubes, which were found to be moreabundant when compared with unlesioned leeches,were orientated at approx. 50◦ with respect to themain axis of the animal (this angle is the same asthat of oblique muscles in the unlesioned body wall)(Lanzavecchia et al., 1977). Therefore a new col-lagen structure transiently replaces, for at least4 months, the entire muscle wall, lost during surgical
www.biolcell.org | Volume 97 (7) | Pages 557–568 565

G. Tettamanti and others
explantation, thus efficiently and optimally provid-ing an adequate hydrostatic pressure to ensure move-ment, tissular resistance and containment of internalorgans.
If it is assumed that “structures are always thetargets of selection according to their function”(Langman, 1989), then the question is why the samefinal complex organization of the collagen is foundfrom wound healing leeches to human cornea; infact, the general architecture of leech collagen compo-nents (i.e. fibril organization and bundle orientation)is identical with the structural pattern of collagenbundles observed in vertebrate cornea, that are as-sembled using a strikingly similar process (Birk andTrelstad, 1984).
Many interesting examples of conservation, alongthe zoological scale, of various anatomical structuresand related functions can be used to support the ob-servation that “Nature has followed economic andconservative strategies based on the conservation ofa lot of molecules and related functions” (Ottavianiet al., 2001). In particular, collagen components arestill present in older metazoa to demonstrate thatthose animals possess and express the genes regu-lating the synthesis and ordered aggregation of allmatrix components. Probably, evolution preservedthese primitive models because of their excellentfunctional utility, redeploying them for greater ef-fectiveness.
It is possible that the selection of anatomical sys-tems, characterized not only by a striking structuralcomplexity but also by multifunctional purposes, canbe viewed as invaluable advantages that should bemaintained along the zoological evolutionary scale.
Materials and methodsAnimals and treatmentsLeeches (H. medicinalis, Annelida, Hirudinea, obtained fromRicarimpex, Eysines, France) measuring 10 × 1.00 cm were keptin water at 22–23◦C in aerated tanks. Animals were fed monthlywith calf blood and starved for 4 weeks before each experiment.The animals were randomly divided into separate experimentalgroups (ten animals/group) according to the different methodsand treatments used.
Before each experiment, leeches were anaesthetized with a sat-urated solution of mephenesin (3-o-toloxy-1,2-propanediol). Toobtain a wound healing response, leeches were subjected to sur-gical lesion affecting the entire thickness of the body wall (i.e.an explant of 2 mm × 2 mm × 2 mm) at the hind part of theanimal (80th dorsal, superficial metamere). Leeches were killedand dissected, 24 h and 1 month after surgery, corresponding
respectively to the maximum synthesis and accumulation ofthe collagen. Tissues undergoing regeneration and tissues ad-jacent to surgical wounds were processed according to differentprocedures.
Light microscopy, TEM, SEM and AFMLeech samples were prepared as described by Tettamanti et al.(2004).
In addition, specimens from control and explanted leecheswere fixed in 0.25% glutaraldehyde and 0.25% paraformalde-hyde in 0.1 M sodium cacodylate buffer [0.1 M Na(CH3)2AsO2in distilled water] for 20 min at room temperature (18◦C), andthen rinsed in cold/cacodylate buffer and dehydrated in gradedethanol and hexamethyldisilazane. Samples were observed intapping-mode AFM (TMAFM) on a Digital Instruments Nano-scope III multimode microscope, fitted with Nanosensors SSS-NCH silicon probes (force constant, 30–60 N/m and resonancefrequency, 300–350 kHz). All measurements were performed inair at a scan rate of approx. 2 Hz.
To enhance the resolution of TMAFM images without com-promising their accuracy and linearity, TMAFM files wererendered in three-dimensions by photorealistic ray-tracing ren-dering software (POV-RAY, version 3.0).
HistochemistryTo visualize proteoglycans, leech samples were treated as de-scribed previously (Tettamanti et al., 2004).
Enzyme digestionChABC was used for removing glycosaminoglycans from ECMas described previously (Baccarini-Contri et al., 1990; Dell’Orboet al., 1995).
Regenerating tissues from surgically lesioned leeches were in-cubated for 24 h at 37◦C in 0.1 M Tris/HCl buffer (pH 8.3) con-taining 0.5 unit/ml ChABC (proteinase-free; Seikagaku, Tokyo,Japan).
After surgical treatment, samples were fixed in 0.25% glutar-aldehyde and 0.25% paraformaldehyde in 0.1 M sodium caco-dylate buffer (pH 7.2) for 20 min at room temperature, and thendehydrated and processed for electron microscopy as describedabove.
ImmunocytochemistryLeeches were dissected and fixed in 4% (w/v) paraformal-dehyde solution in 0.2 M PBS (pH 7.2) containing 0.1% glutar-aldehyde for 2 h at 4◦C. The specimens were then washedwith the same buffer and dehydrated. After a standard stepof serial ethanol dehydration, tissue samples were embeddedin Epon–Araldite, as described above, and sectioned with aReichert Ultracut S ultratome. After etching with 3% NaOHin 100% ethanol (Causton, 1984), 0.7 µm sections were in-cubated for 30 min with PBS containing 2% (w/v) BSA.Double stainings were performed by, first, incubating thesample, for 1 h in a dark moist chamber at 37◦C, with anti-human collagen I polyclonal antibody (Santa Cruz Biotech-nology, Santa Cruz, CA, U.S.A.) (working dilution, 1:40).Specimens were washed and then incubated with TRITC(tetramethylrhodamine β-isothiocyanate)-conjugated secondaryantibody (Jackson ImmunoResearch Laboratories, West Grove,PA, U.S.A.). Subsequently, the same specimens were incubated
566 C© Portland Press 2005 | www.biolcell.org

Leech collagen reorganization Research article
with anti-human collagen III polyclonal antibody (working di-lution, 1:40; Santa Cruz Biotechnology) followed by alkalinephosphatase-conjugated antiserum (Jackson ImmunoResearchLaboratories). Immunoreactivity was visualized using BCIP(5-bromo-4-chloroindol-3-yl phosphate)/Nitro Blue Tetrazo-lium (Sigma, St. Louis, MO, U.S.A.).
Mounted slides were examined with an Olympus BH2 mi-croscope. Control sections were incubated in PBS/BSA in theabsence of the primary antibody.
Thin sections were collected on gold grids of high meshnumber to improve adherence during incubation. Grids, sec-tion face down, were placed, for 10 min, on a 25 µl droplet ofPBS, 0.1% BSA buffer and then transferred on to the surfaceof a 25 µl droplet of anti-human collagen IV monoclonal anti-body (1:20 dilution; Santa Cruz Biotechnology) and incubatedfor 4 h. Washed grids were incubated for 1 h in a 25 µl dropletof the appropriate secondary antibody-gold conjugate (particlesize, 10 nm; dilution 1:100; Sigma).
Control sections were incubated in PBS/BSA buffer withoutthe primary antibody. After wash steps in a series of 50 µldroplets of distilled water, the sections were stained in uranylacetate.
The sections were examined under Jeol 1010 EX electronmicroscope (Jeol, Tokyo, Japan).
AcknowledgmentsWe are grateful to M.L. Guidali for the excellent tech-nical assistance, to Dr R. Ferrarese for assistance inpreparing the Figures and to K. Histon for correctingthe language of this paper. This work was partiallysupported by University of Insubria ‘Progetto di ec-cellenza per la ricerca di Ateneo’ (2003–4).
ReferencesBabel, W. and Glanville, R.W. (1984) Structure of human basement
membrane (type IV) collagen. Complete amino-acid sequence of a914-residue-long pepsin fragment from the alpha-I (IV) chain.Eur. J. Biochem. 143, 545–556
Baccarini-Contri, M., Vincenzi, D., Cicchetti, F., Mori, G. andPasquali-Ronchetti, I. (1990) Immunocytochemical localization ofproteoglycans within normal elastin fibers. Eur. J. Cell Biol. 53,305–312
Badylak, S.F. (2002) The extracellular matrix as a scaffold for tissuereconstruction. Semin. Cell Dev. Biol. 13, 377–383
Barnes, M.J., Morton, L.F., Bennett, R.C., Bailey, A.J. and Sims, T.J.(1976) Presence of type III collagen in guinea-pig dermal scar.Biochem. J. 157, 263–266
Birk, D.E. and Trelstad, R.L. (1984) Extracellular compartments inmatrix morphogenesis: collagen fibril, bundle, and lamellarformation by corneal fibroblasts. J. Cell Biol. 99, 2024–2033
Birk, D.E. and Zycband, E. (1994) Assembly of the tendonextracellular matrix during development. J. Anat. 184, 457–463
Birk, D.E., Southern, J.F., Zycband, E., Fallon, J.T. and Trelstad, R.L.(1989) Collagen fibril bundles: a branching assembly unit in tendonmorphogenesis. Development 107, 437–443
Birk, D.E., Hahn, R.A., Linsemayer, C.Y. and Zycband, E. (1996)Characterization of collagen fibril segments from chicken embryocornea, dermis and tendon. Matrix Biol. 15, 111–118
Boote, C., Dennis, S., Newton, R.H., Puri, H. and Meek, K.M. (2003)Collagen fibrils appear more closely packed in the prepupillarycornea: optical and biomedical implications. Invest. Ophthal.Vis. Sci. 44, 2941–2948
Bradbury, S. (1958) A cytological and histochemical study of theconnective-tissue fibres of the leech, Hirudo medicinalis.Q. J. Microsc. Sci. 99, 131–142
Bradbury, S. and Meek, G.A. (1958) A study of fibrogenesis in theleech, Hirudo medicinalis. Q. J. Microsc. Sci. 99, 143–148
Causton, B.E. (1984) The choice of resins for electronimmunocytochemistry. In Immunolabelling for Electron Microscopy(Polack, J.M. and Varndell I.M., eds.), pp. 17–28, Elsevier,Amsterdam
Cheung, D.T., Benya, P.D., Perelman, N., Dicesare, P.E. and Nimni,M.E. (1990) A highly specific and quantitative method fordetermining Type III–I collagen ratios in tissues. Matrix 10, 164–171
Chiquet, M., Matthisson, M., Koch, M., Tannheimer, M. andChiquet-Ehrismann, R. (1996) Regulation of extracellular matrixsynthesis by mechanical stress. Biochem. Cell Biol. 74, 737–744
Chiquet, M., Renedo, A.S., Huber, F. and Fluck, M. (2003) How dofibroblasts translate mechanical signal into changes in extracellularmatrix production? Matrix Biol. 22, 73–80
Christiansen, D.L., Huang, E.K. and Silver, F.H. (2000) Assembly oftype I collagen: fusion of fibril subunits and the influence of fibrildiameter on mechanical properties. Matrix Biol. 19, 409–420
Cluzel, C., Lethias, C., Garrone, R. and Exposito, J.H. (2000) Seaurchin fibrillar collagen 2α chain participates in heterotrimericmolecules of (1α)2 2α stoichiometry. Matrix Biol. 19, 545–547
de Eguileor, M., Grimaldi, A., Boselli, A., Tettamanti, G., Lurati, S.,Valvassori, R. and Lanzavecchia, G. (1999) Possible roles ofextracellular matrix and cytoskeleton in leech body wall muscle.J. Microsc. 196, 6–18
de Eguileor, M., Grimaldi, A., Tettamanti, G., Ferrarese, R., Congiu, T.,Protasoni, M., Perletti, G., Valvassori, R. and Lanzavecchia, G.(2001) Hirudo medicinalis: a new model system for testingactivators and inhibitors of angiogenesis. Angiogenesis 4, 299–313
de Eguileor, M., Tettamanti, G., Grimaldi, A., Perletti, G., Congiu, T.,Rinaldi, L. and Valvassori, R. (2004) Hirudo medicinalis: avasculartissues for clear-cut angiogenesis studies? Curr. Pharm. Des. 10,1979–1988
Dell’Orbo, C., De Luca, G., Giglio, L., Quacci, D. and Soldi, C. (1995)The role of proteoglycans in maintaining collagen fibrilsmorphology. Histol. Histopathol. 10, 583–588
Franc, S. (1985) Collagen of Coelenterates. In Biology ofInvertebrates and Lower Vertebrates Collagens (Bairati, A. andGarrone, R., eds.), pp. 197–210, Plenum, New York
Fratzl, P., Misof, K., Zizak, I., Rapp, G., Amenitsch, H. andBernstorff, S. (1998) Fibrillar structure and mechanical propertiesof collagen. J. Struct. Biol. 122, 119–122
Frazier, K., Williams, S., Kothapalli, D., Klapper, H. and Grotendorst,G.R. (1996) Stimulation of fibroblast cell growth, matrix productionand granulation tissue formation by connective tissue growthfactor. J. Invest. Dermatol. 107, 404–411
Gaill, F., Hamraoui, L., Sicot, F.X. and Timpl, R. (1994) Immunologicalproperties and tissue localization of two different collagen types inannelid and vestimentifera species. Eur. J. Cell Biol. 65, 392–401
Gordon, J.E. (1988) The Science and Structure of Materials,Freeman, New York
Gutsmann, T., Fantner, G.E., Venturoni, M., Ekani-Nkodo, A.,Thompson, J.B., Kindt, J.H., Morse, D.E., Kuchnir Fygenson, D.and Hansma, P.K. (2003) Evidence that collagen fibrils in tendonsare inhomogeneously structured in a tubelike manner. Biophys. J.84, 2593–2598
Hay, E. (1991) Collagen and other matrix glycoproteins inembryogenesis. In Cell Biology of Extracellular Matrix (Hay, E., ed.),pp. 419–462, Plenum Publishing, New York
www.biolcell.org | Volume 97 (7) | Pages 557–568 567

G. Tettamanti and others
Hirsch, M., Prenant, G. and Renard, G. (2001) Three-dimensionalsupramolecular organization of the extracellular matrix in humanand rabbit corneal stroma, as revealed by ultrarapid-freezing anddeep-etching methods. Exp. Eye Res. 72, 123–135
Ingber, D. (1994) Cellular tensegrety: exploring how mechanicalchanges in the cytoskeleton regulate cell growth, migration andtissue pattern during morphogenesis. Int. Rev. Cytol. 150, 173–224
Ikoma, T., Kobayashi, H., Tanaka, J., Walsh, D. and Mann, S. (2003)Microstructure, mechanical and biomimetic properties of fishscales from Pagrus major. J. Struct. Biol. 142, 327–333
Kadler, K.E., Holmes, D.F., Trotter, J. and Chapman, J.A. (1996)Collagen fibril formation. Biochem. J. 316, 1–11
Kletsas, D., Pratsinis, H., Zervolea, I., Handris, P., Sevaslidou, E.,Ottaviani, E. and Statakos, D. (2000) Fibroblast responses toexogenous and autocrine growth factors relevant to tissue repair.The effect of aging. Ann. N.Y. Acad. Sci. 908, 155–166
Koch, M., Laub, F., Zhou, P., Hahn, R.A., Tanaka, S., Burgeson, R.E.,Gerecke, D.R., Ramirez, F. and Gordon, M.K. (2003) Collagen XXIV,a vertebrate fibrillar collagen with structural features of invertebratecollagens: selective expression in developing cornea and bone.J. Biochem. Chem. 278, 43236–43244
Langman, R.E. (1989) The Immune System. Evolutionary PrinciplesGuide our Understanding of this Complex Biological DefenceSystem, Academic Press, London, U.K.
Lanzavecchia, G., de Eguileor, M., Vailati, G. and Valvassori, R. (1977)Studies on the helical and paramiosinic muscles. VI.Submicroscopic organization and function of body wall musclefibers in some leeches. Boll. Zool. 44, 311–326
Lim, A., Greenspan, D.S. and Smith, B.D. (1994) Expression of α2type I collagen in W8 cells increases cell adhesion and decreasescolony formation in soft agar. Matrix Biol. 14, 21–30
Lunstrum, G.P., Bechinger, H.-P., Fessler, L., Duncan, K.G.,Nelson, R. and Fessler, J.H. (1988) Drosophila basementmembrane procollagen IV. J. Biol. Chem. 263, 18318–18327
Masuda-Nakagawa, L.M. and Nicholls, J.G. (1991) Extracellularmatrix molecules in development and regeneration of the leechCNS. Philos. Trans. R. Soc. Lond. 331, 323–335
Masuda-Nakagawa, L.M., Beck, K. and Chiquet, M. (1988)Identification of molecules in leech extracellular matrix thatpromote neurite outgrowth. Proc. R. Soc. Lond. 235, 247–257
Miyagawa, A., Kobayashi, M., Fujita, Y., Nakamura, M., Hirano, K.,Kobayashi, K. and Miyake, Y. (2000) Surface topology of collagenfibrils associated with proteoglycan in mouse cornea and sclera.Jpn. J. Ophthalmol. 44, 591–595
Nicolas, G., Gaill, F. and Zylberberg, L. (1997) In situ localization oftwo fibrillar collagens in two compact connective tissues byimmunoelectron microscopy after cryotechnical processing.J. Histochem. Cytochem. 45, 119–128
Niederreither, K., D’Souza, R., Metsaranta, M., Eberspaecher, H.,Toman, P.D., Vuorio, E. and De Crombrugghe, B. (1994)Co-ordinate patterns of expression of type I and III collagen duringmouse development. Matrix Biol. 14, 705–713
Ottani, V., Raspanti, M. and Ruggeri, A. (2001) Collagen structure andfunctional implications. Micron 32, 251–260
Ottani, V., Martini, D., Franchi, M., Ruggeri, A. and Raspanti, M. (2002)Hierarchical structures in fibrillar collagens. Micron 33, 587–596
Ottaviani, E., Franchini, A. and Kletsas, D. (2001) Platelet-derivedgrowth factor and transforming growth factor-β in invertebrateimmune and neuroendocrine interactions: another sign ofconservation in evolution. Comp. Biochem. Physiol. 129, 295–306
Parry, D.A.D. and Craig, A.S. (1984) Growth and development ofcollagen fibrils in connective tissue. In Ultrastructure of theConnective Tissue Matrix (Ruggeri, A. and Motta, P.M., eds.),pp. 34–64, Martinus Nijhoff, The Hague
Raspanti, M., Congiu, T. and Guizzardi, S. (2002) Structural aspectsof the extracellular matrix of tendon: an atomic force and scanningelectron microscopy study. Arch. Histol. Cytol. 65, 37–43
Sawyer, R.T. (1986) Leech Biology and Behaviour, 1: Anatomy,Physiology and Behaviour, Oxford Science Publications,Oxford, U.K.
Scott, E.S. and Bosworth, T.R. (1990) A comparative biochemical andultrastructural study of proteoglycan-collagen interactions incorneal stroma. Functional and metabolic implications.Biochem. J. 270, 491–497
Sicot, F.X., Exposito, J.Y., Masselot, M., Garrone, R., Deutsch, J.and Gaill, F. (1997) Cloning of an annelid fibrillar-collagen gene andphylogenetic analysis of vertebrate and invertebrate collagen.Eur. J. Biochem. 246, 50–58
Suzuma, K., Naruse, K., Suzuma, I., Takahara, N., Ueki, K., Aiello, L.P.and King, G. (2000) Vascular endothelial growth factor inducesexpression of connective tissue growth factor via KDR, Flt1, andphosphatidylinositol 3-kinase-Akt-dependent pathways in retinalvascular cells. J. Biol. Chem. 52, 40725–40731
Tettamanti, G., Grimaldi, A., Ferrarese, R., Palazzi, M., Perletti, G.,Valvassori, R., Cooper, E.L., Lanzavecchia, G. and de Eguileor, M.(2003) Leech responses to tissue transplantation. Tissue Cell 35,199–212
Tettamanti, G., Grimaldi, A., Rinaldi, L., Arnaboldi, F., Congiu, T.,Valvassori, R. and de Eguileor, M. (2004) The multifunctional role offibroblasts during wound healing in Hirudo medicinalis (Annelida,Hirudinea). Biol. Cell 96, 443–455
Trelstad, R.L. and Hayashi, K. (1979) Tendon collagen fibrillogenesis:intracellular subassemblies and cell surface changes associatedwith fibril growth. Dev. Biol. 71, 228–242
Trotter, J.A., Kadler, K.E. and Holmes, D.F. (2000) Echinodermcollagen fibrils grow by surface-nucleation-propagation from bothcentres and ends. J. Mol. Biol. 300, 531–540
Vilarta, R. and De Campos Vidal, B. (1989) Anisotropic andbiochemical properties of tendons modified by exercise anddenervation: aggregation and macromolecular order in collagenbundles. Matrix 9, 55–61
Weiss, P. and Ferris, W. (1956) The basement lamella of amphibianskin. Its reconstruction after wounding. J. Biophys.Biochem. Cytol. 2, 275–281
Received 26 July 2004/2 November 2004; accepted 9 November 2004
Published as Immediate Publication 9 May 2005, DOI 10.1042/BC20040085
568 C© Portland Press 2005 | www.biolcell.org