spontaneous autocrine release of protons activates asic-mediated currents in hek293 cells
TRANSCRIPT

ORIGINAL ARTICLE 473
Spontaneous AutocrineRelease of Protons ActivatesASIC-Mediated Currents inHEK293 Cells
ULYANA LALO, YURI PANKRATOV, R. ALAN NORTH, AND ALEXEI VERKHRATSKY*Faculty of Life Sciences, The University of Manchester, Manchester, UK
When examining HEK293 cells by whole-cell patch-clamp electrophysiology we found spontaneous currents, present in almost all cells.These currents were carried by Naþ ions, were inhibited by amiloride and by cells exposure to acidic (pH 6.3) extracellular solutions.These properties (ion carrier, amiloride-sensitivity, and inactivation by constant lowering of extracellular pH) were similar to theproperties of proton-activated currents measured from the same cells. Spontaneous currents required intracellular ATP, werecompletely inhibited by intracellular Ca2þ buffering with BAPTA and were suppressed by intracellular administration ofvesicular HþATPase inhibitor bafilomycin. ATP-inducedCa2þ influx through P2X receptors inHEK293 cells stably transfectedwith P2X2,P2X2/3 or P2X4 purinoreceptor subunits transiently potentiated amplitude and frequency of spontaneous currents; this effect wasantagonized by bafilomycin. We concluded that spontaneous currents represent activation of acid-sensitive ion channels (ASICs) byautocrine vesicular release of protons from HEK cells.J. Cell. Physiol. 212: 473–480, 2007. � 2007 Wiley-Liss, Inc.
Contract grant sponsor: The Wellcome Trust.
*Correspondence to: Alexei Verkhratsky, Faculty of Life Sciences,The University of Manchester, 1.124 Stopford Building, OxfordRoad, Manchester M13 9PT, UK.E-mail: [email protected]
Received 5 November 2006; Accepted 4 January 2007
DOI: 10.1002/jcp.21043
Proton-activated conductances were initially found in spinal(Gruol et al., 1980) and in sensory (Krishtal and Pidoplichko,1980, 1981a,b) mammalian neurones. Subsequently Hþ-activated cationic currents were described in many types ofperipheral and central nerve cells (Vyklicky et al., 1990;Escoubas et al., 2000; Baron et al., 2002; Fukuda et al., 2006).Several types of acid-sensing ion channels (ASICs) were clonedand molecularly characterized; there are six members of theASIC family (Bassilana et al., 1997; Waldmann et al., 1997;Waldmann and Lazdunski, 1998; Reeh and Kress, 2001),represented by 4 main types (ASIC1–4) and 2 splice variants(ASIC1a, ASIC2a). The functional role of Hþ-activated ionchannels remains not fully understood, although theirinvolvement into pain sensation (Krishtal and Pidoplichko,1981b; Chen et al., 2002; Krishtal, 2003), integrative processesin the CNS (Wemmie et al., 2002), and neuronal responses toischemia (Allen and Attwell, 2002) were suggested.The HEK293 cell line, derived from human embryonic kidneycells immortalized with human adenovirus cells, are generallyused as one of the most versatile platforms for expression ofnumerous ion channels and ionotropic receptors (Thomas andSmart, 2005). Yet, the success of transfection approach is, to avery large extent, determined by the expression of endogenouschannels and signaling systems. The naive HEK293 cells werereported to possess numerous voltage- and ligand-gated ionchannels, including a7, a5, and d subunits of acetylcholinereceptors, GluR3 glutamate receptors,b3, g3, and e subunits ofGABA receptors and ASIC1a Hþ-activated channels(Gunthorpe et al., 2001; Thomas and Smart, 2005).In the present article we report a new phenomenon ofautocrine activation of Hþ-activated ion channels in ‘‘plain’’HEK293 cells and in HEK293 cells expressing several subunitsof P2X purinoreceptors. The auto-secretion of protonsresulted from Ca2þ-dependent exocytosis; ATP-inducedstimulation of Ca2þ entry in P2X receptor expressing cellstransiently increased the release of protons.
Materials and MethodsCell lines and electrophysiology
HEK293 cells were obtained from American Type Culture collection(ATCC, Manassas, Virginia), and were cultured in modified Eagles
� 2 0 0 7 W I L E Y - L I S S , I N C .
media supplementedwith 10% fetal calf serum (Kawashima et al., 1998).A stable cell lines expressing P2X2, P2X2/3 or P2X4 receptors wereprepared as described previously (Kawashima et al., 1998). Forelectrophysiological recordings cells were plated onto glass cover slipscoated with poly-D-lysine and used after 24–72 h in culture. Forrecordings, cover slips with cells were transferred into the chambermounted on the stage of an inverted microscope and incubated withextracellular solution containing (mM): NaCl 140; KCl 2.7; CaCl2 2.5;MgCl2 1; HEPES 15, NaH2PO4 1, pH 7.35.Whole-cell patch clamp recordings were performed by using the patchpipette (3–6 MV) filled with the intracellular solution (in mM): K-gluconate 110;NaCl 10;HEPES 10;MgATP 5;GTP0.5; EGTA10;CaCl21; pH7.35. In some experiments the ‘‘symmetricNaþ’’ solution (inmM:NaCl 140; KCl 2.7; HEPES 10; MgATP 5; GTP 0.5; EGTA 10; CaCl2 1;pH 7.35) was used. The series and input resistanceswere 8� 3MV and1,000–3,000 MV, respectively and varied by less than 20% in the cellsaccepted for analysis. Liquid junction potentials were measured withthe EPC-9 patch-clamp amplifier and PULSE software; all voltagesreported are corrected. All recordings were performed at holdingpotential�80 mV, unless stated otherwise. Currents were monitoredusing EPC-9 (HEKA, Lambrecht, Germany), filtered at 3.9 kHz anddigitized at 10 kHz. Experiments were controlled by PULSE/PULSEFITsoftware (HEKA) and data were analyzed by self-designed software.Agonists and antagonists were applied using RSC200 fast flow deliverysystem (Biologic Science Instruments, Grenoble, France).Alternatively, a modified ‘‘square-pulse’’ concentration jump method(Lalo et al., 2001; Pankratov et al., 2003) was used for rapid (solutionexchange time �20 msec) applications of solutions containing variousagents to single cells.
Spontaneous currents analysis
The spontaneous currents (SC) were analyzed off-line. For thepreliminary detection spontaneous inward currents of amplitude
474 L A L O E T A L .
higher than 2 SD of baseline noise were selected. Each SC wassubsequently analyzed in the 140 msec-long time window (40 msecbefore and 100msec after peak of current). The SCpeak amplitudewasdetermined using a computer routine based on the fitting of eachcurrent trace by themodel curvewith single exponential rise and decayphases. Theminimal square root procedure was used to determine theamplitude of themodel curve, while the time constants and offset wereoptimized by the gradient method to minimize the mean square error.The parameters of eachmodel curvewere reasonably restricted by therange of 0.3–3 msec for rise and 4–40 msec for decay time constants.Usually mean square error of fit amounted 5–20% of SC amplitude,depending on baseline noise. If error of fit exceeded 25%, currentswere discarded from further analysis. For the accepted events theamplitude of the fitted curve was taken as SC amplitude. Asdemonstrated in (Pankratov and Krishtal, 2003) such a technique istolerant to noise and allows obtaining the noise-free amplitudedistribution of miniature synaptic currents.The set of 100–200 SCs was obtained for each protocol and used forstatistical analysis. The amplitude distributions were analyzed usingprobability density functions and likelihood maximization techniques,as described in (Stricker et al., 1996; Pankratov and Krishtal, 2003).The integral value of spontaneous current ‘‘Power’’ (P) was used forvisualization of time-dependent changes in both amplitude and
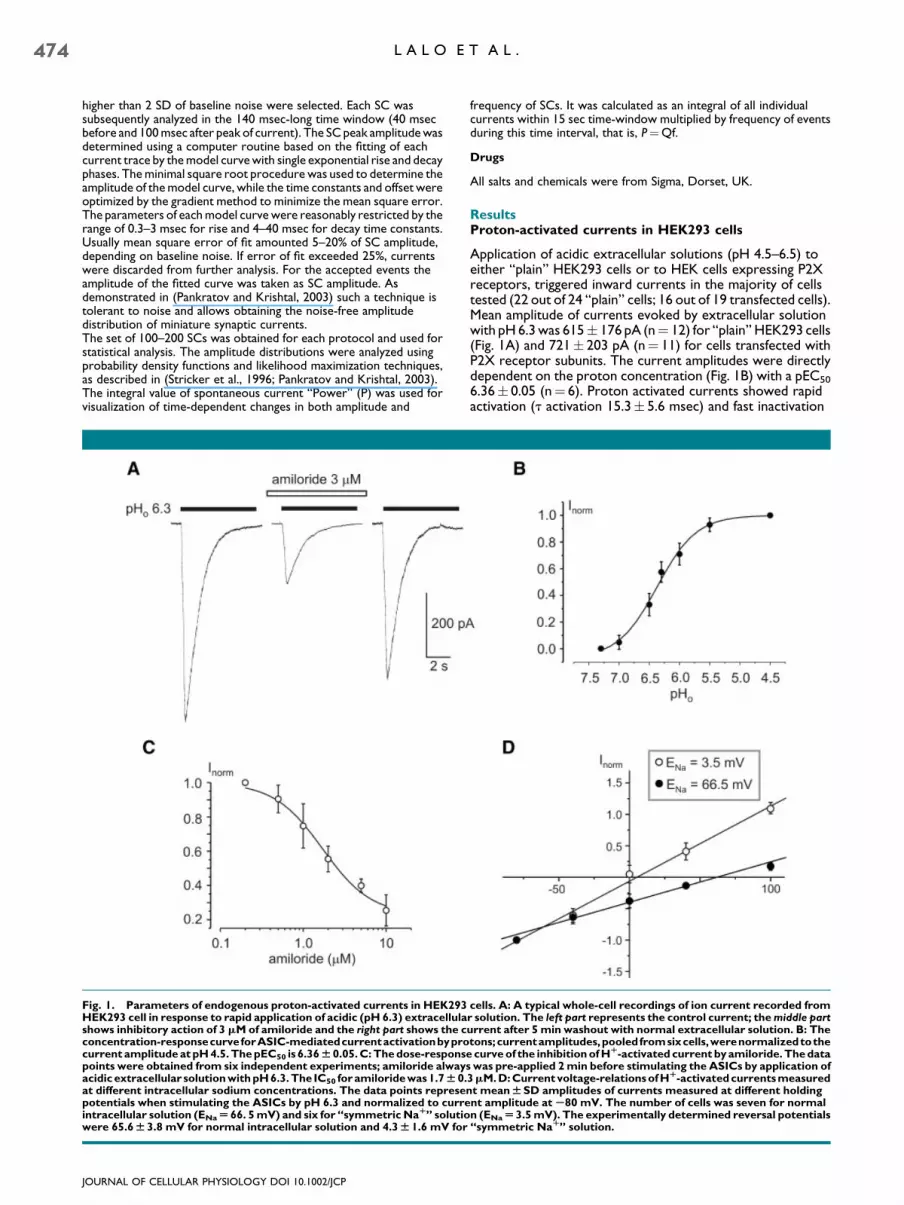
Fig. 1. Parameters of endogenous proton-activated currents in HEK293HEK293 cell in response to rapid application of acidic (pH 6.3) extracellulashows inhibitory action of 3 mM of amiloride and the right part shows the cconcentration-responsecurveforASIC-mediatedcurrentactivationbyprocurrent amplitude at pH 4.5. The pEC50 is 6.36W 0.05. C: The dose-responspoints were obtained from six independent experiments; amiloride alwayacidic extracellular solution with pH 6.3. The IC50 for amiloride was 1.7W 0.at different intracellular sodium concentrations. The data points represepotentials when stimulating the ASICs by pH 6.3 and normalized to curreintracellular solution (ENaU 66. 5 mV) and six for ‘‘symmetric NaR’’ solutiowere 65.6W 3.8 mV for normal intracellular solution and 4.3W 1.6 mV for
JOURNAL OF CELLULAR PHYSIOLOGY DOI 10.1002/JCP
frequency of SCs. It was calculated as an integral of all individualcurrents within 15 sec time-window multiplied by frequency of eventsduring this time interval, that is, P¼Qf.
Drugs
All salts and chemicals were from Sigma, Dorset, UK.
ResultsProton-activated currents in HEK293 cells
Application of acidic extracellular solutions (pH 4.5–6.5) toeither ‘‘plain’’ HEK293 cells or to HEK cells expressing P2Xreceptors, triggered inward currents in the majority of cellstested (22 out of 24 ‘‘plain’’ cells; 16 out of 19 transfected cells).Mean amplitude of currents evoked by extracellular solutionwith pH6.3was 615� 176 pA (n¼ 12) for ‘‘plain’’ HEK293 cells(Fig. 1A) and 721� 203 pA (n¼ 11) for cells transfected withP2X receptor subunits. The current amplitudes were directlydependent on the proton concentration (Fig. 1B) with a pEC50
6.36� 0.05 (n¼ 6). Proton activated currents showed rapidactivation (t activation 15.3� 5.6 msec) and fast inactivation
cells. A: A typical whole-cell recordings of ion current recorded fromr solution. The left part represents the control current; the middle parturrent after 5 min washout with normal extracellular solution. B: Thetons;currentamplitudes,pooledfromsixcells,werenormalizedtothe
e curve of the inhibition of HR-activated current by amiloride. The datas was pre-applied 2 min before stimulating the ASICs by application of3mM. D: Current voltage-relations of HR-activatedcurrents measurednt meanWSD amplitudes of currents measured at different holdingnt amplitude at S80 mV. The number of cells was seven for normaln (ENaU3.5 mV). The experimentally determined reversal potentials‘‘symmetric NaR’’ solution.
A U T O C R I N E S T I M U L A T I O N O F A S I C s I N H E K 2 9 3 C E L L S 475
decay (t decay 1,156� 94 msec; n¼ 12; both parameters weredetermined for pH 6.3). The Hþ-activated currents wereinhibited by amiloride with an IC50 1.7� 0.3 mM (n¼ 6)(Fig. 1A,C). The reversal potential of the Hþ-activated currentsin HEK293 cells closely followed the equilibrium potentialfor Naþ ions (Fig. 1D). There was no difference in theproperties of Hþ-activated currents between normal HEK293cells and cells expressing P2X receptors. All in all biophysicalparameters of ASIC-mediated currents in HEK293 cells weresimilar to those obtained previously by Randall and colleagues(Gunthorpe et al., 2001).
ASICs mediate spontaneous currents in HEK cells
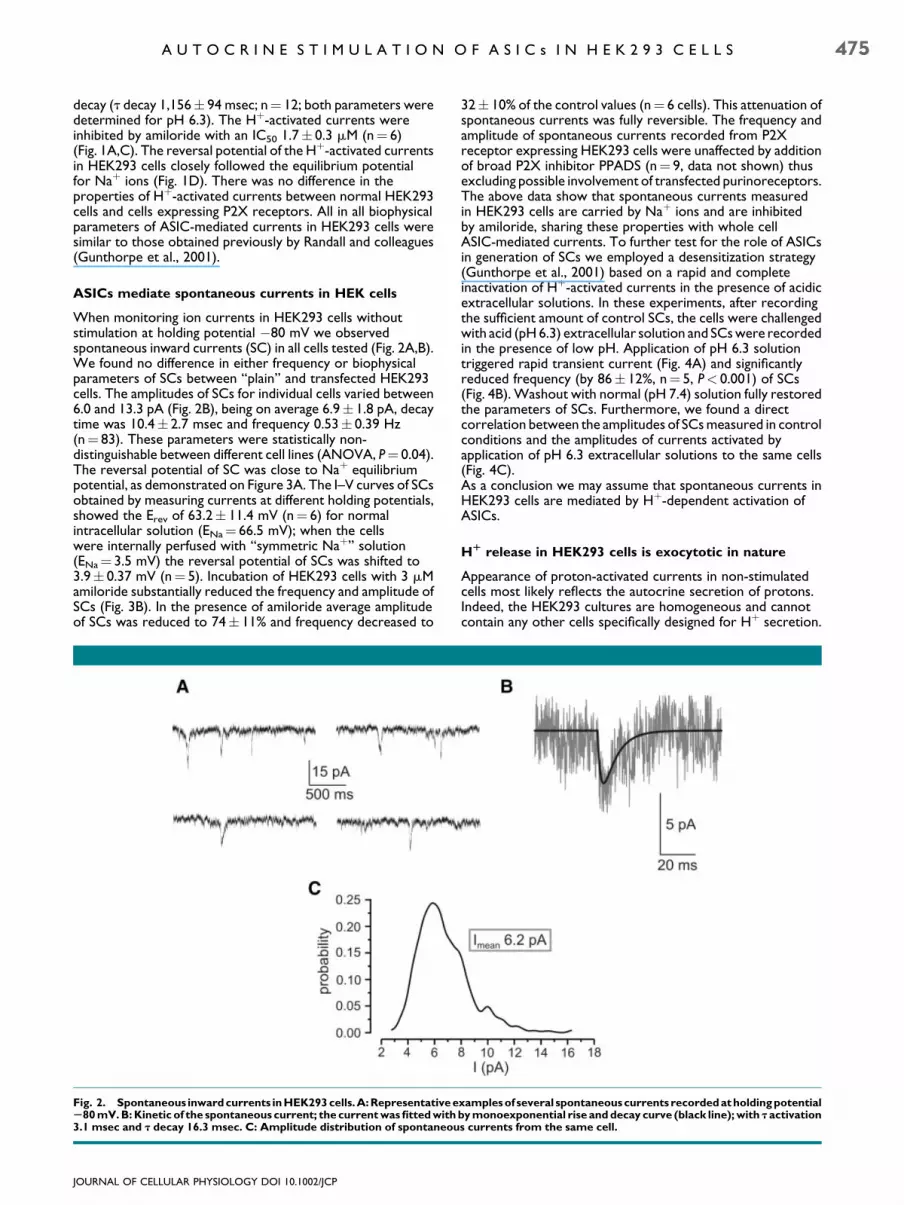
When monitoring ion currents in HEK293 cells withoutstimulation at holding potential �80 mV we observedspontaneous inward currents (SC) in all cells tested (Fig. 2A,B).We found no difference in either frequency or biophysicalparameters of SCs between ‘‘plain’’ and transfected HEK293cells. The amplitudes of SCs for individual cells varied between6.0 and 13.3 pA (Fig. 2B), being on average 6.9� 1.8 pA, decaytime was 10.4� 2.7 msec and frequency 0.53� 0.39 Hz(n¼ 83). These parameters were statistically non-distinguishable between different cell lines (ANOVA, P¼ 0.04).The reversal potential of SC was close to Naþ equilibriumpotential, as demonstrated on Figure 3A. The I–V curves of SCsobtained by measuring currents at different holding potentials,showed the Erev of 63.2� 11.4 mV (n¼ 6) for normalintracellular solution (ENa¼ 66.5 mV); when the cellswere internally perfused with ‘‘symmetric Naþ’’ solution(ENa¼ 3.5 mV) the reversal potential of SCs was shifted to3.9� 0.37 mV (n¼ 5). Incubation of HEK293 cells with 3 mMamiloride substantially reduced the frequency and amplitude ofSCs (Fig. 3B). In the presence of amiloride average amplitudeof SCs was reduced to 74� 11% and frequency decreased to
Fig. 2. Spontaneous inward currents in HEK293 cells.A:Representative eS80 mV. B: Kinetic of the spontaneous current; the current was fitted with3.1 msec and t decay 16.3 msec. C: Amplitude distribution of spontaneou
JOURNAL OF CELLULAR PHYSIOLOGY DOI 10.1002/JCP
32� 10% of the control values (n¼ 6 cells). This attenuation ofspontaneous currents was fully reversible. The frequency andamplitude of spontaneous currents recorded from P2Xreceptor expressing HEK293 cells were unaffected by additionof broad P2X inhibitor PPADS (n¼ 9, data not shown) thusexcluding possible involvement of transfected purinoreceptors.The above data show that spontaneous currents measuredin HEK293 cells are carried by Naþ ions and are inhibitedby amiloride, sharing these properties with whole cellASIC-mediated currents. To further test for the role of ASICsin generation of SCs we employed a desensitization strategy(Gunthorpe et al., 2001) based on a rapid and completeinactivation of Hþ-activated currents in the presence of acidicextracellular solutions. In these experiments, after recordingthe sufficient amount of control SCs, the cells were challengedwith acid (pH6.3) extracellular solution and SCswere recordedin the presence of low pH. Application of pH 6.3 solutiontriggered rapid transient current (Fig. 4A) and significantlyreduced frequency (by 86� 12%, n¼ 5, P< 0.001) of SCs(Fig. 4B).Washout with normal (pH 7.4) solution fully restoredthe parameters of SCs. Furthermore, we found a directcorrelation between the amplitudes of SCsmeasured in controlconditions and the amplitudes of currents activated byapplication of pH 6.3 extracellular solutions to the same cells(Fig. 4C).As a conclusion we may assume that spontaneous currents inHEK293 cells are mediated by Hþ-dependent activation ofASICs.
HR release in HEK293 cells is exocytotic in nature
Appearance of proton-activated currents in non-stimulatedcells most likely reflects the autocrine secretion of protons.Indeed, the HEK293 cultures are homogeneous and cannotcontain any other cells specifically designed for Hþ secretion.
xamples of several spontaneouscurrents recorded atholding potentialby monoexponential rise and decay curve (black line); with t activations currents from the same cell.

Fig. 3. SpontaneouscurrentssharebiophysicalandpharmacologicalpropertieswithASIC-mediatedwhole-cellcurrents.A:Thecurrent-voltagedependence of SCs measured at different intracellular sodium concentrations. The middle part shows I–V relationships of SCs; the data pointsrepresent meanWSD amplitudes of currents recorded at different holding potentials and normalized to current amplitude atS80 mV. The rightand left parts show representative SC recordings at different holding potentials and different intracellular sodium concentrations as shown on thegraph. The number of cells was six for normal intracellular solution (ENaU 66. 5 mV) and five for ‘‘symmetric NaR’’ solution (ENaU 3.5 mV).B: Inhibitory action of amiloride on spontaneous currents. The graph shows amplitudes of the SCs taken from a single HEK293 cell over a period of800 sec. Administration of amiloride significantly decreased both the amplitudes and frequency of SCs.
476 L A L O E T A L .
Furthermore, the extracellular solution is heavily buffered(10 mM HEPES) which make intercellular diffusion of protonsimpossible. The discrete nature of SCs measured from HEKcells implies the similarly discrete release of protons, whichmaybe liberated in packets associated with exocytosis of Hþ
containing vesicles.We therefore tested the possibility of exocytotic Hþ release byemploying several complementary protocols. First, we testedfor intracellular Ca2þ dependence of spontaneous currents.When the cells were perfusedwith solution supplementedwith10 mM of fast Ca2þ buffer BAPTA, spontaneous currents
JOURNAL OF CELLULAR PHYSIOLOGY DOI 10.1002/JCP
became undetectable within less than 1 min of whole-cellrecording (n¼ 10; Fig. 5A), thus demonstrating the [Ca2þ]i-dependence of Hþ release.Second, we tested for the release dependence on intracellularATP level. When the cells were perfused with intracellularsolutions with no ATP added, the progressive and relativelyrapid run-down of SCs was observed in all nine cells tested(Fig. 5B); frequency of SCs in these cellswas very low even at thebeginning of experiment, mean value being 0.087� 0.025 Hz(n¼ 9). On average, 10 min of intracellular perfusion withATP-free intracellular solutions reduced the frequency of SCs
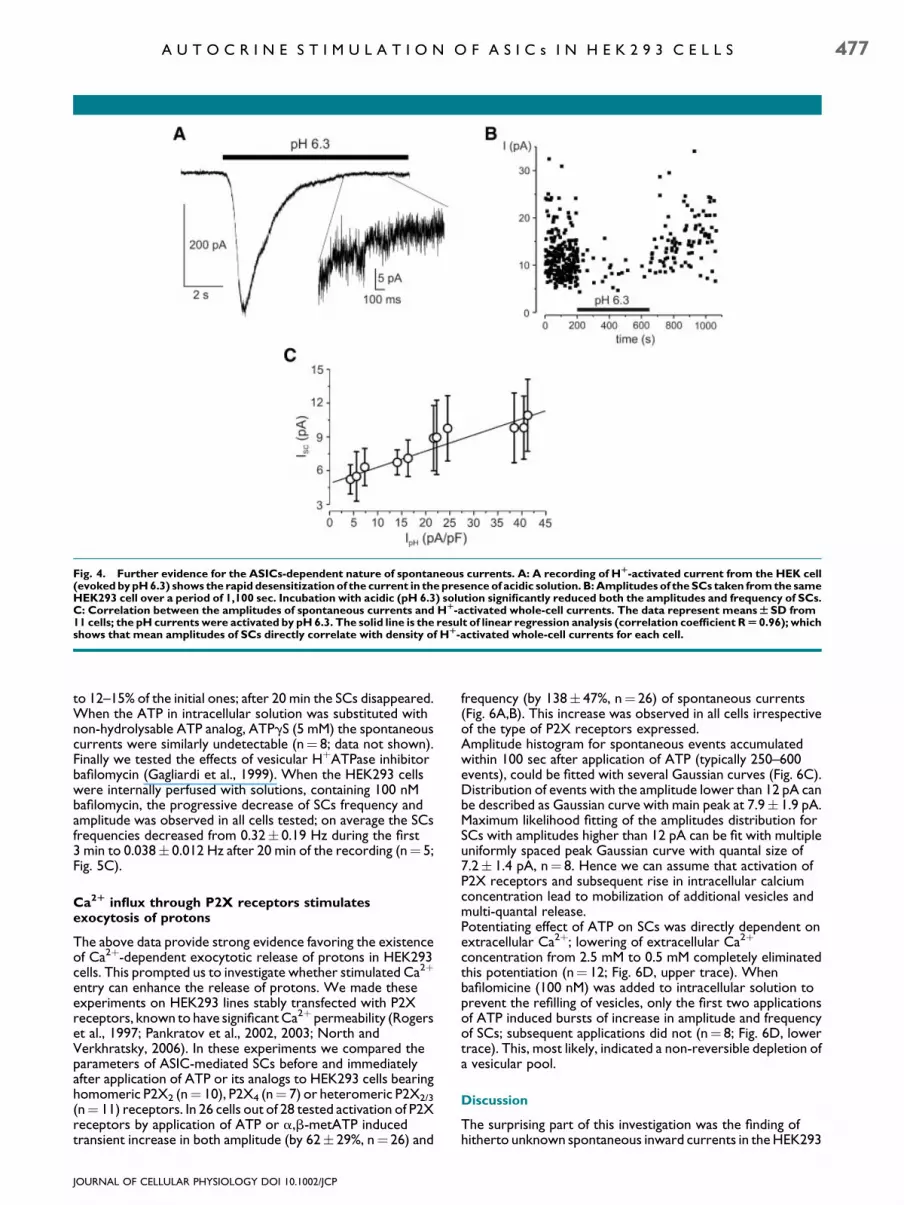
Fig. 4. Further evidence for the ASICs-dependent nature of spontaneous currents. A: A recording of HR-activated current from the HEK cell(evoked by pH 6.3) shows the rapid desensitization of the current in the presence of acidic solution. B: Amplitudes of the SCs taken from the sameHEK293 cell over a period of 1,100 sec. Incubation with acidic (pH 6.3) solution significantly reduced both the amplitudes and frequency of SCs.C: Correlation between the amplitudes of spontaneous currents and HR-activated whole-cell currents. The data represent meansWSD from11 cells; the pH currents were activated by pH 6.3. The solid line is the result of linear regression analysis (correlation coefficient RU 0.96); whichshows that mean amplitudes of SCs directly correlate with density of HR-activated whole-cell currents for each cell.
A U T O C R I N E S T I M U L A T I O N O F A S I C s I N H E K 2 9 3 C E L L S 477
to 12–15% of the initial ones; after 20 min the SCs disappeared.When the ATP in intracellular solution was substituted withnon-hydrolysable ATP analog, ATPgS (5 mM) the spontaneouscurrents were similarly undetectable (n¼ 8; data not shown).Finally we tested the effects of vesicular HþATPase inhibitorbafilomycin (Gagliardi et al., 1999). When the HEK293 cellswere internally perfused with solutions, containing 100 nMbafilomycin, the progressive decrease of SCs frequency andamplitude was observed in all cells tested; on average the SCsfrequencies decreased from 0.32� 0.19 Hz during the first3 min to 0.038� 0.012 Hz after 20 min of the recording (n¼ 5;Fig. 5C).
Ca2R influx through P2X receptors stimulatesexocytosis of protons
The above data provide strong evidence favoring the existenceof Ca2þ-dependent exocytotic release of protons in HEK293cells. This prompted us to investigate whether stimulated Ca2þ
entry can enhance the release of protons. We made theseexperiments on HEK293 lines stably transfected with P2Xreceptors, known to have significantCa2þ permeability (Rogerset al., 1997; Pankratov et al., 2002, 2003; North andVerkhratsky, 2006). In these experiments we compared theparameters of ASIC-mediated SCs before and immediatelyafter application of ATP or its analogs to HEK293 cells bearinghomomeric P2X2 (n¼ 10), P2X4 (n¼ 7) or heteromeric P2X2/3
(n¼ 11) receptors. In 26 cells out of 28 tested activation of P2Xreceptors by application of ATP or a,b-metATP inducedtransient increase in both amplitude (by 62� 29%, n¼ 26) and
JOURNAL OF CELLULAR PHYSIOLOGY DOI 10.1002/JCP
frequency (by 138� 47%, n¼ 26) of spontaneous currents(Fig. 6A,B). This increase was observed in all cells irrespectiveof the type of P2X receptors expressed.Amplitude histogram for spontaneous events accumulatedwithin 100 sec after application of ATP (typically 250–600events), could be fitted with several Gaussian curves (Fig. 6C).Distribution of events with the amplitude lower than 12 pA canbe described as Gaussian curve with main peak at 7.9� 1.9 pA.Maximum likelihood fitting of the amplitudes distribution forSCs with amplitudes higher than 12 pA can be fit with multipleuniformly spaced peak Gaussian curve with quantal size of7.2� 1.4 pA, n¼ 8. Hence we can assume that activation ofP2X receptors and subsequent rise in intracellular calciumconcentration lead to mobilization of additional vesicles andmulti-quantal release.Potentiating effect of ATP on SCs was directly dependent onextracellular Ca2þ; lowering of extracellular Ca2þ
concentration from 2.5 mM to 0.5 mM completely eliminatedthis potentiation (n¼ 12; Fig. 6D, upper trace). Whenbafilomicine (100 nM) was added to intracellular solution toprevent the refilling of vesicles, only the first two applicationsof ATP induced bursts of increase in amplitude and frequencyof SCs; subsequent applications did not (n¼ 8; Fig. 6D, lowertrace). This, most likely, indicated a non-reversible depletion ofa vesicular pool.
Discussion
The surprising part of this investigation was the finding ofhitherto unknown spontaneous inward currents in theHEK293

Fig. 5. Sensitivity of spontaneous currents to intracellular ATP and bafilomycin. A:Middle part shows amplitudes (ISC) and power (P) of the SCstaken from HEK293 cell over a period of 600 sec. The cell was perfused with intracellular solution supplemented with 10 mM BAPTA. Note almostimmediate disappearance of SCs after the beginning of recording. The representative current traces taken at the beginning and at the end of theexperiment(as indicatedonthetimegraph)areshownonthe leftand rightparts, respectively.B:Middlepartshowsamplitudes(ISC)andpower(P)ofthe SCs taken from HEK293 cell over a period of 1,000 sec. The cell was perfused with intracellular solution without any ATP added. Noteprogressive decrease of SCs amplitude and frequency. The representative current recordings taken at the beginning and at the end of theexperiment(asindicatedonthetimegraph)areshownonthe leftandrightparts, respectively.C:Middlepartshowsamplitudes(ISC)andpower(P)oftheSCstaken fromHEK293celloveraperiodof1,000sec.Thecellwasperfusedwith intracellular solutionsupplementedwith100nMbafilomycin,whichcausedprogressivedecrease inSCsamplitudeandfrequency.Therepresentativecurrentrecordingstakenatthebeginningandattheendofthe experiment (as indicated on the time graph) are shown on the left and right parts, respectively.
478 L A L O E T A L .
cells. These spontaneous currents were observed in all cellsstudied; the biophysical parameters of SCs were exactly thesame in ‘‘plain’’ HEK293 cells and in cell lines expressing P2Xreceptors. We can conclude therefore that the mechanismunderlying the generation of SC is intrinsic to the HEK293 cells.Investigation of the channel identity of SCs demonstratedthe role for Hþ-activated ASIC channels. As a matter of fact,HEK293 cells constitutively express ASIC1a channels(Gunthorpe et al., 2001), which, when activated by lowextracellular pH, generate large inward currents, carried
JOURNAL OF CELLULAR PHYSIOLOGY DOI 10.1002/JCP
predominantly by Naþ ions and sensitive to amiloride((Gunthorpe et al., 2001) and Fig. 1). We found that biophysicaland pharmacological profile of spontaneous currents wasidentical to that of ASIC-mediated whole-cell currents. Indeed,the SCs demonstrated the very same Naþ-dependent reversalpotential and can be effectively inhibited by amiloride (cf. Figs. 1and 3). When ASIC channels were forcefully desensitized bycells incubation in acidic extracellular solutions, thespontaneous currents disappeared; washout with normalsolution restored them (Fig. 4). Furthermore we found direct
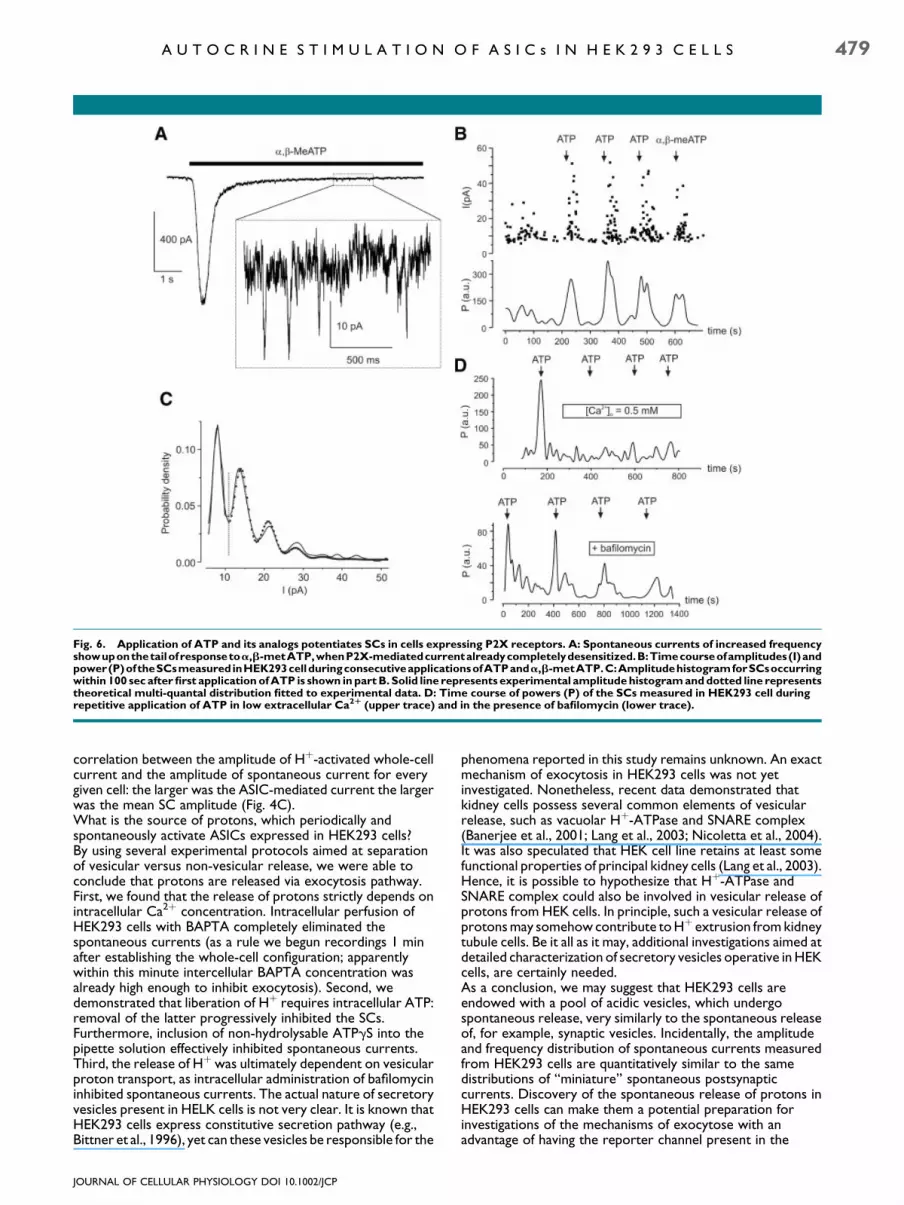
Fig. 6. Application of ATP and its analogs potentiates SCs in cells expressing P2X receptors. A: Spontaneous currents of increased frequencyshowuponthetailofresponsetoa,b-metATP,whenP2X-mediatedcurrentalreadycompletelydesensitized.B:Timecourseofamplitudes(I)andpower(P)oftheSCsmeasured inHEK293cellduringconsecutiveapplicationsofATPanda,b-metATP.C:AmplitudehistogramforSCsoccurringwithin 100 sec after first application of ATP is shown in part B. Solid line represents experimental amplitude histogram and dotted line representstheoretical multi-quantal distribution fitted to experimental data. D: Time course of powers (P) of the SCs measured in HEK293 cell duringrepetitive application of ATP in low extracellular Ca2R (upper trace) and in the presence of bafilomycin (lower trace).
A U T O C R I N E S T I M U L A T I O N O F A S I C s I N H E K 2 9 3 C E L L S 479
correlation between the amplitude of Hþ-activated whole-cellcurrent and the amplitude of spontaneous current for everygiven cell: the larger was the ASIC-mediated current the largerwas the mean SC amplitude (Fig. 4C).What is the source of protons, which periodically andspontaneously activate ASICs expressed in HEK293 cells?By using several experimental protocols aimed at separationof vesicular versus non-vesicular release, we were able toconclude that protons are released via exocytosis pathway.First, we found that the release of protons strictly depends onintracellular Ca2þ concentration. Intracellular perfusion ofHEK293 cells with BAPTA completely eliminated thespontaneous currents (as a rule we begun recordings 1 minafter establishing the whole-cell configuration; apparentlywithin this minute intercellular BAPTA concentration wasalready high enough to inhibit exocytosis). Second, wedemonstrated that liberation of Hþ requires intracellular ATP:removal of the latter progressively inhibited the SCs.Furthermore, inclusion of non-hydrolysable ATPgS into thepipette solution effectively inhibited spontaneous currents.Third, the release of Hþwas ultimately dependent on vesicularproton transport, as intracellular administration of bafilomycininhibited spontaneous currents. The actual nature of secretoryvesicles present in HELK cells is not very clear. It is known thatHEK293 cells express constitutive secretion pathway (e.g.,Bittner et al., 1996), yet can these vesicles be responsible for the
JOURNAL OF CELLULAR PHYSIOLOGY DOI 10.1002/JCP
phenomena reported in this study remains unknown. An exactmechanism of exocytosis in HEK293 cells was not yetinvestigated. Nonetheless, recent data demonstrated thatkidney cells possess several common elements of vesicularrelease, such as vacuolar Hþ-ATPase and SNARE complex(Banerjee et al., 2001; Lang et al., 2003; Nicoletta et al., 2004).It was also speculated that HEK cell line retains at least somefunctional properties of principal kidney cells (Lang et al., 2003).Hence, it is possible to hypothesize that Hþ-ATPase andSNARE complex could also be involved in vesicular release ofprotons from HEK cells. In principle, such a vesicular release ofprotonsmay somehowcontribute toHþ extrusion fromkidneytubule cells. Be it all as it may, additional investigations aimed atdetailed characterization of secretory vesicles operative inHEKcells, are certainly needed.As a conclusion, we may suggest that HEK293 cells areendowed with a pool of acidic vesicles, which undergospontaneous release, very similarly to the spontaneous releaseof, for example, synaptic vesicles. Incidentally, the amplitudeand frequency distribution of spontaneous currents measuredfrom HEK293 cells are quantitatively similar to the samedistributions of ‘‘miniature’’ spontaneous postsynapticcurrents. Discovery of the spontaneous release of protons inHEK293 cells can make them a potential preparation forinvestigations of the mechanisms of exocytose with anadvantage of having the reporter channel present in the

480 L A L O E T A L .
secreting cell. The great advantage of this preparation is the‘‘classical’’ quantal behavior. At the same time, many factorswhich potentially may hamper investigations of quantalrelease in central synapses, such as filtering by dendrites ornon-uniformity in quantal size in various synapses, simply do notexist in HEK preparation.
Literature Cited
Allen NJ, Attwell D. 2002. Modulation of ASIC channels in rat cerebellar Purkinje neurons byischaemia-related signals. J Physiol 543:521–529.
Banerjee A, Li G, Alexander EA, Schwartz JH. 2001. Role of SNAP-23 in trafficking ofHþ-ATPase in cultured inner medullary collecting duct cells. Am J Physiol Cell Physiol280:C775–C781.
Baron A, Waldmann R, Lazdunski M. 2002. ASIC-like, proton-activated currents in rathippocampal neurons. J Physiol 539:485–494.
Bassilana F, Champigny G,Waldmann R, deWeille JR, Heurteaux C, Lazdunski M. 1997. Theacid-sensitive ionic channel subunit ASIC and the mammalian degenerin MDEG form aheteromultimeric Hþ-gated Naþ channel with novel properties. J Biol Chem 272:28819–28822.
Bittner MA, Bennett MK, Holz RW. 1996. Evidence that syntaxin 1A is involved in storage inthe secretory pathway. J Biol Chem 271:11214–11221.
Chen CC, Zimmer A, Sun WH, Hall J, Brownstein MJ. 2002. A role for ASIC3 in themodulation of high-intensity pain stimuli. Proc Natl Acad Sci USA 99:8992–8997.
Escoubas P,DeWeille JR, LecoqA,Diochot S,WaldmannR,ChampignyG,MoinierD,MenezA, Lazdunski M. 2000. Isolation of a tarantula toxin specific for a class of proton-gatedNaþ
channels. J Biol Chem 275:25116–25121.Fukuda T, Ichikawa H, Terayama R, Yamaai T, Kuboki T, Sugimoto T. 2006. ASIC3-immunoreactive neurons in the rat vagal and glossopharyngeal sensory ganglia. Brain Res1081:150–155.
Gagliardi S, Rees M, Farina C. 1999. Chemistry and structure activity relationships ofbafilomycin A1, a potent and selective inhibitor of the vacuolar Hþ-ATPase. Curr MedChem 6:1197–1212.
Gruol DL, Barker JL, Huang LY, MacDonald JF, Smith TG, Jr. 1980. Hydrogen ions havemultiple effects on the excitability of culturedmammalian neurons. Brain Res 183:247–252.
Gunthorpe MJ, Smith GD, Davis JB, Randall AD. 2001. Characterisation of a human acid-sensing ion channel (hASIC1a) endogenously expressed in HEK293 cells. Pflugers Arch442:668–674.
Kawashima E, Estoppey D, Virginio C, Fahmi D, Rees S, Surprenant A, North RA. 1998.A novel and efficient method for the stable expression of heteromeric ion channels inmammalian cells. Receptors Channels 5:53–60.
JOURNAL OF CELLULAR PHYSIOLOGY DOI 10.1002/JCP
Krishtal O. 2003. The ASICs: Signaling molecules? Modulators? Trends Neurosci 26:477–483.
Krishtal OA, Pidoplichko VI. 1980. A receptor for protons in the nerve cell membrane.Neuroscience 5:2325–2327.
Krishtal OA, Pidoplichko VI. 1981a. Receptor for protons in the membrane of sensoryneurons. Brain Res 214:150–154.
Krishtal OA, Pidoplichko VI. 1981b. A ‘‘receptor’’ for protons in small neurons of trigeminalganglia: Possible role in nociception. Neurosci Lett 24:243–246.
Lalo UV, Pankratov YV, Arndts D, Krishtal OA. 2001. Omega-conotoxin GVIA potentlyinhibits the currents mediated by P2X receptors in rat DRG neurons. Brain Res Bull54:507–512.
Lang K, Wagner C, Haddad G, Burnekova O, Geibel J. 2003. Intracellular pH activatesmembrane-bound Naþ/Hþ exchanger and vacuolar Hþ-ATPase in human embryonickidney (HEK) cells. Cell Physiol Biochem 13:257–262.
Nicoletta JA, Ross JJ, Li G, Cheng Q, Schwartz J, Alexander EA, Schwartz JH. 2004. Munc-18-2regulates exocytosis of Hþ-ATPase in rat inner medullary collecting duct cells. Am JPhysiol Cell Physiol 287:C1366–C1374.
North RA, Verkhratsky A. 2006. Purinergic transmission in the central nervous system.Pflugers Arch. 452:479–485.
Pankratov YV, Krishtal OA. 2003. Distinct quantal features of AMPA and NMDA synapticcurrents in hippocampal neurons: Implication of glutamate spillover and receptorsaturation. Biophys J 85:3375–3387.
Pankratov Y, Lalo U, Krishtal O, Verkhratsky A. 2002. Ionotropic P2X purinoreceptorsmediate synaptic transmission in rat pyramidal neurones of layer II/III of somato-sensorycortex. J Physiol 542:529–536.
Pankratov Y, Lalo U, Krishtal O, Verkhratsky A. 2003. P2X receptor-mediated excitatorysynaptic currents in somatosensory cortex. Mol Cell Neurosci 24:842–849.
Reeh PW, Kress M. 2001. Molecular physiology of proton transduction in nociceptors. CurrOpin Pharmacol 1:45–51.
Rogers M, Colquhoun LM, Patrick JW, Dani JA. 1997. Calcium flux through predominantlyindependent purinergic ATP and nicotinic acetylcholine receptors. J Neurophysiol77:1407–1417.
Stricker C, Field AC, Redman SJ. 1996. Statistical analysis of amplitude fluctuations in EPSCsevoked in rat CA1 pyramidal neurones in vitro. J Physiol 490:419–441.
Thomas P, Smart TG. 2005. HEK293 cell line: A vehicle for the expression of recombinantproteins. J Pharmacol Toxicol Methods 51:187–200.
Vyklicky L, Jr., Vlachova V, Krusek J. 1990. The effect of external pH changes on responses toexcitatory amino acids in mouse hippocampal neurones. J Physiol 430:497–517.
Waldmann R, Lazdunski M. 1998. H(þ)-gated cation channels: Neuronal acid sensors in theNaC/DEG family of ion channels. Curr Opin Neurobiol 8:418–424.
Waldmann R, Champigny G, Bassilana F, Heurteaux C, Lazdunski M. 1997. A proton-gatedcation channel involved in acid-sensing. Nature 386:173–177.
Wemmie JA, Chen J, Askwith CC, Hruska-Hageman AM, Price MP, Nolan BC, Yoder PG,Lamani E, Hoshi T, Freeman JH, Jr., Welsh MJ. 2002. The acid-activated ion channel ASICcontributes to synaptic plasticity, learning, and memory. Neuron 34:463–477.