interleukin 8: an autocrine growth factor for malignant mesothelioma
TRANSCRIPT

1999;59:367-371. Cancer Res Gabriella Galffy, Kamal A. Mohammed, Patrick A. Dowling, et al. MesotheliomaInterleukin 8: An Autocrine Growth Factor for Malignant
Updated version
http://cancerres.aacrjournals.org/content/59/2/367
Access the most recent version of this article at:
Cited Articles
http://cancerres.aacrjournals.org/content/59/2/367.full.html#ref-list-1
This article cites by 30 articles, 12 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/59/2/367.full.html#related-urls
This article has been cited by 7 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on January 13, 2014. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from
Research. on January 13, 2014. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from

[CANCER RESEARCH 59, 367–371, January 15, 1999]
Interleukin 8: An Autocrine Growth Factor for Malignant Mesothelioma
Gabriella Galffy, Kamal A. Mohammed, Patrick A. Dowling, Najmunnisa Nasreen, Melissa J. Ward, andVeena B. Antony1
Division of Pulmonary and Critical Care Medicine, Department of Medicine, Veterans’ Affairs Medical Center, Indiana University School of Medicine, Indianapolis, Indiana46202
ABSTRACT
Interleukin 8 (IL-8) is a potent chemokine that also has a directgrowth-potentiating effect on certain tumors. In the present study, wedetermined IL-8 levels in human malignant mesothelioma (MM) effusionsand congestive heart failure pleural fluids. We also investigated antigenicIL-8 production by different MM cell lines, and we describe the role ofIL-8 in the autocrine growth regulation of MMs. Mesothelial (CRL-9444 5 MC) and MM (CRL-2081 5 MM-1, CRL-5915 5 MM-2, andCRL-5820 5 MM-3) cell lines were grown using standard culture meth-ods. The bioactive IL-8 levels were measured in supernatants of culturedcells by ELISA, and the expression of cell-associated immunoreactive IL-8was observed by immunohistochemistry. The proliferative activity wasdetermined by thymidine ([3H]thymidine) incorporation and also by di-rect cell counts after incubation with varying concentrations of IL-8 in thepresence/absence of specific polyclonal IL-8 antibody. We found signifi-cantly higher levels of IL-8 in mesothelioma pleural fluids than congestiveheart failure and a time-dependent increase in IL-8 levels in MM-1 andMM-2 cell supernatants during 96 h of incubation. IL-8 levels were nearlyundetectable in MM-3 and MC cell line supernatants. In MM-1 and MM-2cells, IL-8 caused a dose-dependent increase of [3H]thymidine incorpora-tion to maximal levels of 46.36 3.6% and 12.3 6 1.6% (P < 0.001),respectively, when compared with serum-free medium as control. Neu-tralization of IL-8 significantly decreased proliferative activity of MM-1and MM-2. IL-8 did not induce proliferative activity in MM-3 and MCcells. We conclude that IL-8 had a direct growth-potentiating activity inMMs.
INTRODUCTION
MM,2 unlike other pulmonary tumors, is characterized by localtumor extension and invasion of surrounding tissue without distantmetastasis. It is a relatively rare tumor, but its incidence appears to beincreasing. The present annual incidence of MMs is;11.4 per onemillion in men and 2.8 per one million in women in the United States(1). Despite attempts to advance early diagnosis and use combinationtherapies, the prognosis of the patients with MM is poor. The averagesurvival time is about 7–9 months after diagnosis (1, 2). Unfortu-nately, there is no effective treatment for prolonging survival at thistime.
Several tumors produce specific autocrine growth factors that im-prove their growth capacity and ability for metastasis (3–5). IL-8, amember of the super gene family of C-X-C chemokines, is chemo-tactic for and activates neutrophils (6–9). By increasing expansion ofadhesion molecules (10), it also facilitates leukocyte-endothelial andleukocyte-mesothelial interactions, which are involved in the inflam-matory invasion of neutrophils. IL-8 has been described as an impor-tant angiogenic factor for the development of new capillariesin vivo(11–13). In empyema, IL-8 levels were reported to be severalfold
higher than mesothelioma and were comparable in parapneumoniceffusion; however, in tuberculosis, the IL-8 levels were very low (14).Several immune and nonimmune cells, including endothelial andmesothelial cells, have been found to produce IL-8 (15, 16). Inaddition, IL-8 has also been shown to be generated by a variety oftumors such as melanoma (3) and bronchogenic carcinoma (17). Itserves as an autocrine growth factor, in the case of melanoma,allowing for local tumor growth and invasion (18, 19).
The aim of the present study was to determine differences in IL-8levels from human MM effusions and CHF pleural fluids as control.It was also designed to investigate the autocrine IL-8 production bydifferent mesothelioma and mesothelial cells and describes the poten-tial role of IL-8 in the autocrine growth regulation of mesotheliomaand mesothelial cells. We found elevated levels of IL-8 in humanmesothelioma pleural fluids compared with CHF.In vitro, mesothe-lioma cells, but not mesothelial cells, produced high levels of IL-8.We verified that IL-8 is a direct growth-potentiating factor for me-sothelioma but not for mesothelial cells.
MATERIALS AND METHODS
Reagents.Recombinant human IL-8, monoclonal anti-IL-8 Ab (mouseIgG1; this Ab does not cross-react with other C-X-C chemokines), mouseIgG-isotype and specific polyclonal goat anti-IL-8 Ab (polyclonal IL-8 Ab;this Ab does not cross-react with MGSA, neutrophil activating protein-2, orany other chemokine) were purchased from R & D Systems (Minneapolis,MN). Purified nonspecific goat IgG was purchased from Sigma Chemical Co.(St. Louis, MO). [3H]Thymidine was obtained from Amersham Life Science(Arlington Heights, IL).
Human Pleural Mesothelial and MM Cell Lines. One mesothelial cellline (CRL-9444) and three mesothelioma cell lines (CRL-2081, CRL-5915,and CRL-5820) were purchased from American Type Culture Collection(Rockville, MD). The mesothelial cells were resuspended in Ham’s 199 culturemedium (Life Technologies, Inc., Grand Island, NY) containing 10% fetalbovine serum (Harlan Bioproducts, Indianapolis, IN), penicillin (100 units/ml),and streptomycin (100 g/ml). Mesothelioma cells were cultured in RPMI 1640culture medium with 10% fetal bovine serum, 100 units/ml penicillin, and 100mg/ml streptomycin. The cells were plated in 75-cm2 tissue culture flasks(Corning Costar Corp., Cambridge, MA) and incubated at 37°C in 5% CO2 and95% air. The monolayer was confluent between 8 and 10 days. Mesothelialcells had a classic cobblestone morphology (20), absence of factor VIIIantigen, and presence of cytokeratin (21). All cells were used between the thirdand seventh passages.
Human Pleural Fluid. Twelve patients with pleural effusions secondary tomesothelioma (n5 6) and CHF (n5 6) were studied. Pleural fluids wereobtained after informed consent during diagnostic thoracocentesis, as de-scribed previously (16, 22). In all MM patients, the effusions were exudateswith culture negative for infective organisms. A malignant effusion secondaryto mesothelioma was defined by at least one of the following criteria: malig-nant mesothelioma cells in the pleural fluid on cytological examination, ormesothelioma on closed pleural biopsy. Biopsy tissue was examined bymicroscopy and a special panel of stains to diagnose MM. Three patients hada mixed-type mesothelioma: one was an epithelial type mesothelioma, and twowere sarcomatous in nature. Transudative pleural effusions from symptomaticpatients with a diagnosis of CHF served as control. The fluids were saved at270°C for IL-8 ELISA.
Cell Culture Supernatants for IL-8 Determination. To determine theconstitutive production of IL-8 without stimulation, CRL-2081, CRL-5915,
Received 3/12/98; accepted 11/9/98.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby markedadvertisementin accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1 To whom requests for reprints should be addressed, at Veterans’ Affairs MedicalCenter, 1481 West 10th Street, 111P, Indianapolis, IN 46202. Fax: (317) 554-0262.E-mail: [email protected].
2 The abbreviations used are: MM, malignant mesothelioma; IL, interleukin; CHF,congestive heart failure; Ab, antibody; MGSA, melanoma growth stimulatory activity;SFM, serum-free medium.
367
Research. on January 13, 2014. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from
CRL-5820 mesothelioma, and CRL-9444 mesothelial cells were plated in75-cm2 tissue culture flasks (13 106 cells/flask), grown to confluence,washed, and replaced with serum-free RPMI 1640 (8 ml/flask) for mesothe-lioma cells and Ham’s 199 for mesothelial cells. Conditioned media werecollected at 24, 48, 72, and 96 h, centrifuged at 9003 g for 10 min, and savedat 270°C for IL-8 analysis by ELISA.
IL-8 ELISA. Extracellular immunoreactive IL-8 levels were measured by“sandwich” enzyme immunoassay (R&D System, Minneapolis, MN) as de-scribed previously (16). The samples were added to 96-well microtiter plates,which were coated with murine monoclonal Ab to IL-8. The unbound proteinwas washed three times, and an enzyme-linked polyclonal antibody specific toIL-8 was added. The plates were again washed three times, and substratesolution was added to the wells. After 30 min of incubation, stop solution wasadded to each well. The amount of IL-8 was determined by absorbance of thesamples by comparing the standards at 450 nm using the ELISA reader.
Immunohistochemistry for IL-8. Unstimulated mesothelial and mesothe-lioma cells were immunostained with avidin-biotin conjugate and peroxidaseas described previously (7). The cells were cultured in Lab-Tek chamber slides(Nunc, Naperville, IL). The confluent cultures were fixed in absolute metha-nol, and endogenous peroxidase activity was quenched with 1% hydrogenperoxide. Nonspecific binding sites were blocked with 1% normal horse serumfor 50 min, permeabilized in 0.1% Triton X-100 for 10 min, and overlaid with1:1000 dilution of mouse monoclonal anti-human IL-8 Ab for 30 min. Thenegative controls were overlaid with mouse IgG isotype. The slides wererinsed with PBS and overlaid with avidin-biotin conjugated goat anti-mouseIgG (Vectastain ABC kit; Vector Laboratories, Burlingame, CA) for 60 min.The slides were washed and incubated for 5 min in peroxidase substratesolution (3,39-diaminobenzidine) washed in PBS and counterstained by May-er‘s hematoxylin (Sigma Chemical Co.). The cells were dehydrated in gradedalcohol and xylene and covered with glass.
Proliferative Activity. Mesothelial and mesothelioma cell line prolifera-tion was determined by a [3H]thymidine uptake assay as described by Lauberet al. (23) and Hottet al. (24). Cells (23 104) were plated in 48-well platesand grown to confluence. Then the medium was replaced by RPMI 1640 orHam’s 199 and incubated overnight. Various concentrations of IL-8 (5, 25, 50,and 100 ng/ml), polyclonal goat anti-human IL-8 specific Ab (1, 5, 10 and 20mg/ml), or nonspecific purified goat IgG (1, 5, 10, and 20mg/ml) as controlwere added. After 24-h incubation at 37°C in 5% CO2 95% air, 0.5mCi of[3H]thymidine was added to each well, and the plates were reincubated for24 h. The cells were washed three times in complete HBSS and harvested fromthe plates with 0.5% EDTA, precipitated in 5% trichloroacetic acid, andcentrifuged at 3000 rpm for 10 min. They were resuspended in 0.1N NaOH,placed in scintillation fluid, and counted in ab-scintillation counter. Allexperiments were done in triplicate, and data are expressed as percentage ofstimulation (cpm produced by the test factors/cpm produced by SFM3 100).In addition, mesothelial and mesothelioma cell proliferation was measured bydirect cell counts (hemocytometer). After 48 h of seeding, the cell culturemedium was changed to SFM plus either 50 ng/ml IL-8 or 10mg/ml anti-IL-8Ab or SFM alone. Cell counts were obtained after 48 h of incubation. Theselected IL-8 concentrations had no effect on the mesothelial and mesotheli-oma cell viability as demonstrated by trypan blue exclusion.
Statistical Analysis. All values are expressed as mean6 SD. Data werecompared using Student’st test and ANOVA. Values were considered to bestatistically significant whenP was,0.05.
RESULTS
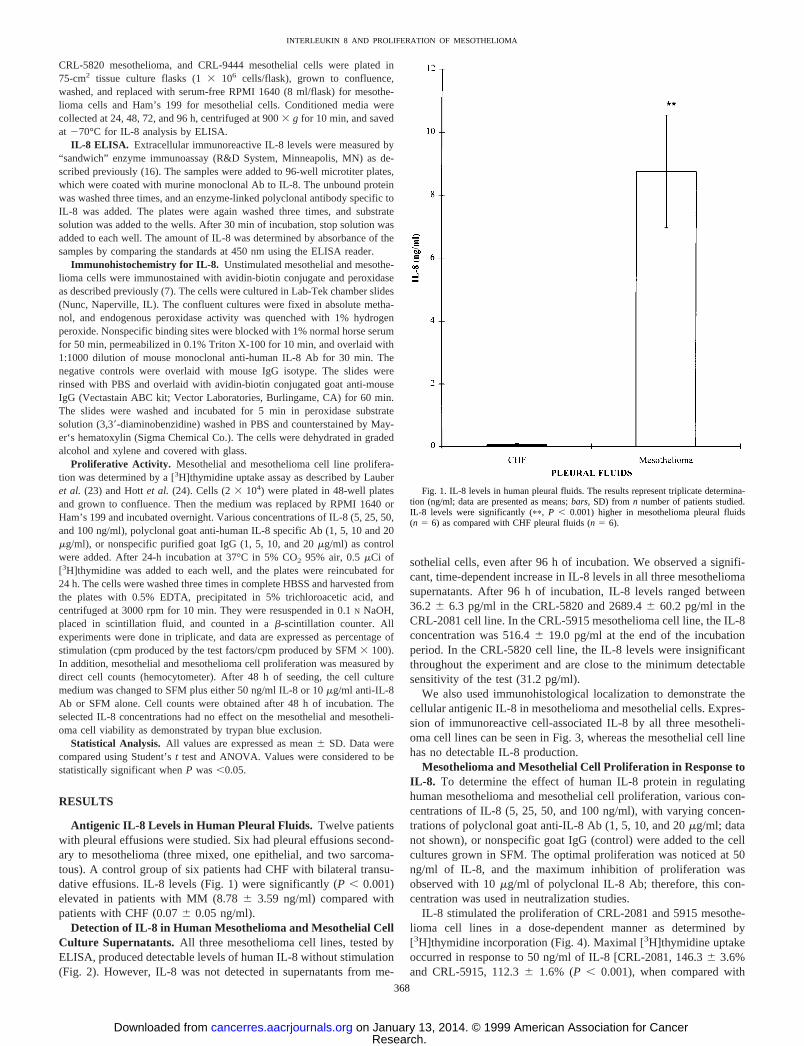
Antigenic IL-8 Levels in Human Pleural Fluids. Twelve patientswith pleural effusions were studied. Six had pleural effusions second-ary to mesothelioma (three mixed, one epithelial, and two sarcoma-tous). A control group of six patients had CHF with bilateral transu-dative effusions. IL-8 levels (Fig. 1) were significantly (P , 0.001)elevated in patients with MM (8.786 3.59 ng/ml) compared withpatients with CHF (0.076 0.05 ng/ml).
Detection of IL-8 in Human Mesothelioma and Mesothelial CellCulture Supernatants. All three mesothelioma cell lines, tested byELISA, produced detectable levels of human IL-8 without stimulation(Fig. 2). However, IL-8 was not detected in supernatants from me-
sothelial cells, even after 96 h of incubation. We observed a signifi-cant, time-dependent increase in IL-8 levels in all three mesotheliomasupernatants. After 96 h of incubation, IL-8 levels ranged between36.2 6 6.3 pg/ml in the CRL-5820 and 2689.46 60.2 pg/ml in theCRL-2081 cell line. In the CRL-5915 mesothelioma cell line, the IL-8concentration was 516.46 19.0 pg/ml at the end of the incubationperiod. In the CRL-5820 cell line, the IL-8 levels were insignificantthroughout the experiment and are close to the minimum detectablesensitivity of the test (31.2 pg/ml).
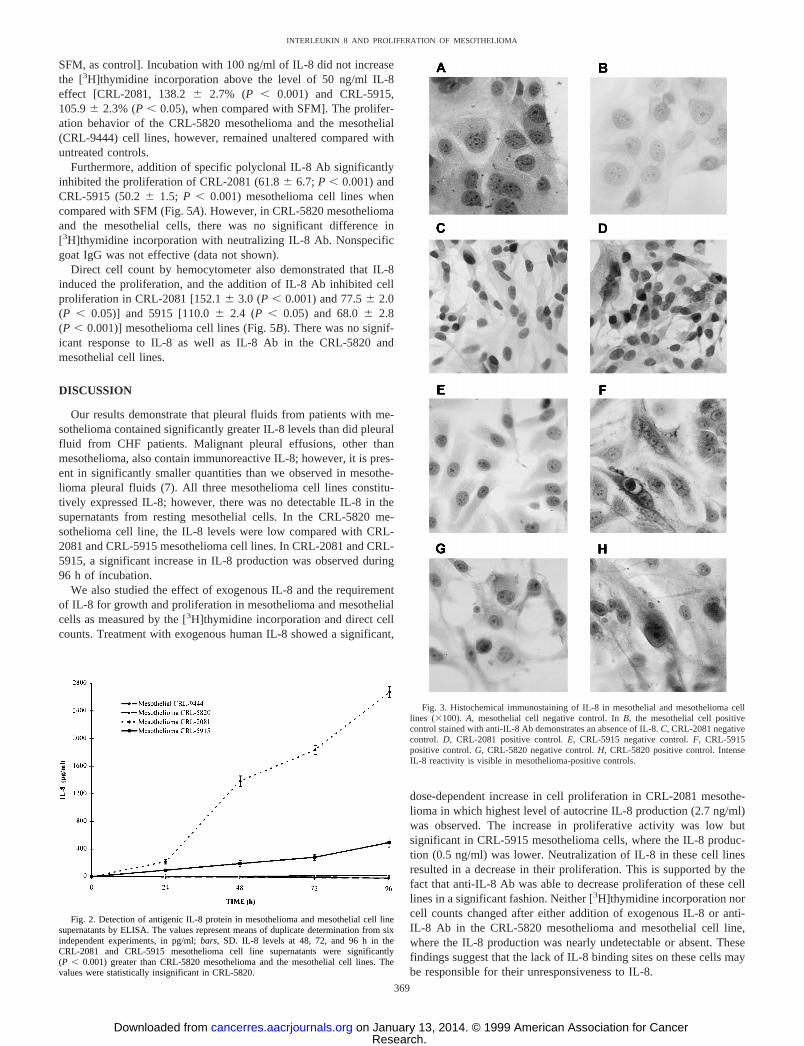
We also used immunohistological localization to demonstrate thecellular antigenic IL-8 in mesothelioma and mesothelial cells. Expres-sion of immunoreactive cell-associated IL-8 by all three mesotheli-oma cell lines can be seen in Fig. 3, whereas the mesothelial cell linehas no detectable IL-8 production.
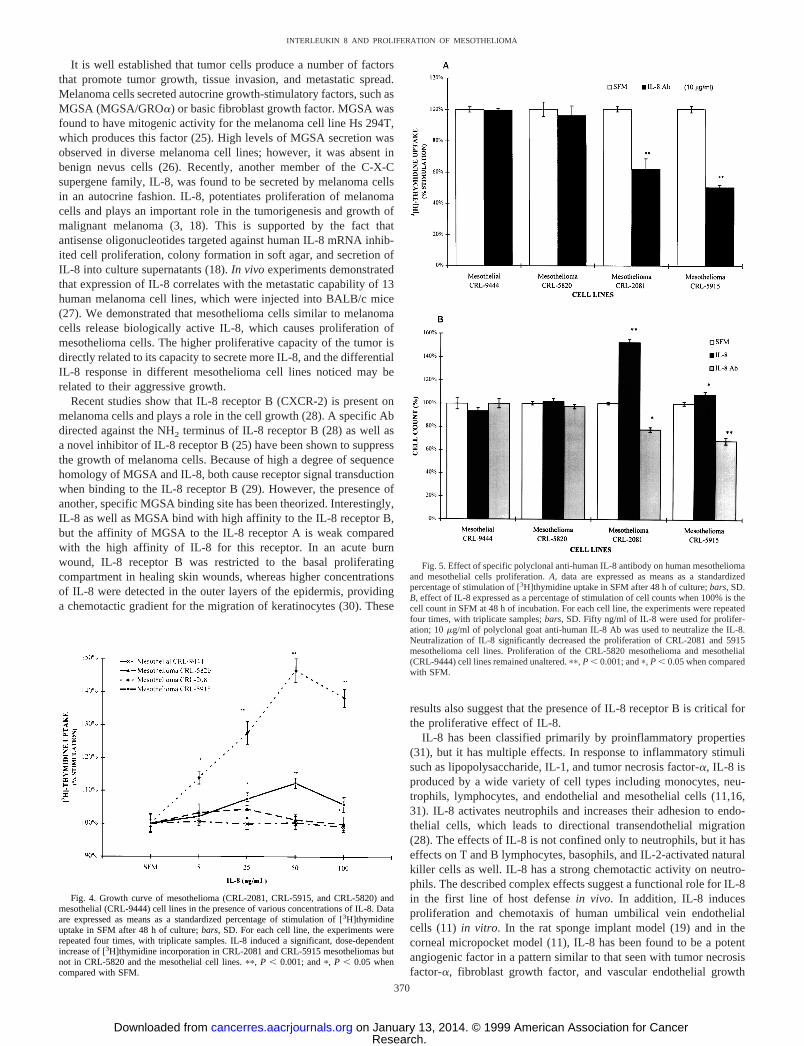
Mesothelioma and Mesothelial Cell Proliferation in Response toIL-8. To determine the effect of human IL-8 protein in regulatinghuman mesothelioma and mesothelial cell proliferation, various con-centrations of IL-8 (5, 25, 50, and 100 ng/ml), with varying concen-trations of polyclonal goat anti-IL-8 Ab (1, 5, 10, and 20mg/ml; datanot shown), or nonspecific goat IgG (control) were added to the cellcultures grown in SFM. The optimal proliferation was noticed at 50ng/ml of IL-8, and the maximum inhibition of proliferation wasobserved with 10mg/ml of polyclonal IL-8 Ab; therefore, this con-centration was used in neutralization studies.
IL-8 stimulated the proliferation of CRL-2081 and 5915 mesothe-lioma cell lines in a dose-dependent manner as determined by[3H]thymidine incorporation (Fig. 4). Maximal [3H]thymidine uptakeoccurred in response to 50 ng/ml of IL-8 [CRL-2081, 146.36 3.6%and CRL-5915, 112.36 1.6% (P , 0.001), when compared with
Fig. 1. IL-8 levels in human pleural fluids. The results represent triplicate determina-tion (ng/ml; data are presented as means;bars, SD) from n number of patients studied.IL-8 levels were significantly (pp, P , 0.001) higher in mesothelioma pleural fluids(n 5 6) as compared with CHF pleural fluids (n 5 6).
368
INTERLEUKIN 8 AND PROLIFERATION OF MESOTHELIOMA
Research. on January 13, 2014. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from
SFM, as control]. Incubation with 100 ng/ml of IL-8 did not increasethe [3H]thymidine incorporation above the level of 50 ng/ml IL-8effect [CRL-2081, 138.26 2.7% (P , 0.001) and CRL-5915,105.96 2.3% (P, 0.05), when compared with SFM]. The prolifer-ation behavior of the CRL-5820 mesothelioma and the mesothelial(CRL-9444) cell lines, however, remained unaltered compared withuntreated controls.
Furthermore, addition of specific polyclonal IL-8 Ab significantlyinhibited the proliferation of CRL-2081 (61.86 6.7; P , 0.001) andCRL-5915 (50.26 1.5; P , 0.001) mesothelioma cell lines whencompared with SFM (Fig. 5A). However, in CRL-5820 mesotheliomaand the mesothelial cells, there was no significant difference in[3H]thymidine incorporation with neutralizing IL-8 Ab. Nonspecificgoat IgG was not effective (data not shown).
Direct cell count by hemocytometer also demonstrated that IL-8induced the proliferation, and the addition of IL-8 Ab inhibited cellproliferation in CRL-2081 [152.16 3.0 (P, 0.001) and 77.56 2.0(P , 0.05)] and 5915 [110.06 2.4 (P , 0.05) and 68.06 2.8(P , 0.001)] mesothelioma cell lines (Fig. 5B). There was no signif-icant response to IL-8 as well as IL-8 Ab in the CRL-5820 andmesothelial cell lines.
DISCUSSION
Our results demonstrate that pleural fluids from patients with me-sothelioma contained significantly greater IL-8 levels than did pleuralfluid from CHF patients. Malignant pleural effusions, other thanmesothelioma, also contain immunoreactive IL-8; however, it is pres-ent in significantly smaller quantities than we observed in mesothe-lioma pleural fluids (7). All three mesothelioma cell lines constitu-tively expressed IL-8; however, there was no detectable IL-8 in thesupernatants from resting mesothelial cells. In the CRL-5820 me-sothelioma cell line, the IL-8 levels were low compared with CRL-2081 and CRL-5915 mesothelioma cell lines. In CRL-2081 and CRL-5915, a significant increase in IL-8 production was observed during96 h of incubation.
We also studied the effect of exogenous IL-8 and the requirementof IL-8 for growth and proliferation in mesothelioma and mesothelialcells as measured by the [3H]thymidine incorporation and direct cellcounts. Treatment with exogenous human IL-8 showed a significant,
dose-dependent increase in cell proliferation in CRL-2081 mesothe-lioma in which highest level of autocrine IL-8 production (2.7 ng/ml)was observed. The increase in proliferative activity was low butsignificant in CRL-5915 mesothelioma cells, where the IL-8 produc-tion (0.5 ng/ml) was lower. Neutralization of IL-8 in these cell linesresulted in a decrease in their proliferation. This is supported by thefact that anti-IL-8 Ab was able to decrease proliferation of these celllines in a significant fashion. Neither [3H]thymidine incorporation norcell counts changed after either addition of exogenous IL-8 or anti-IL-8 Ab in the CRL-5820 mesothelioma and mesothelial cell line,where the IL-8 production was nearly undetectable or absent. Thesefindings suggest that the lack of IL-8 binding sites on these cells maybe responsible for their unresponsiveness to IL-8.
Fig. 2. Detection of antigenic IL-8 protein in mesothelioma and mesothelial cell linesupernatants by ELISA. The values represent means of duplicate determination from sixindependent experiments, in pg/ml;bars, SD. IL-8 levels at 48, 72, and 96 h in theCRL-2081 and CRL-5915 mesothelioma cell line supernatants were significantly(P , 0.001) greater than CRL-5820 mesothelioma and the mesothelial cell lines. Thevalues were statistically insignificant in CRL-5820.
Fig. 3. Histochemical immunostaining of IL-8 in mesothelial and mesothelioma celllines (3100).A, mesothelial cell negative control. InB, the mesothelial cell positivecontrol stained with anti-IL-8 Ab demonstrates an absence of IL-8.C, CRL-2081 negativecontrol. D, CRL-2081 positive control.E, CRL-5915 negative control.F, CRL-5915positive control.G, CRL-5820 negative control.H, CRL-5820 positive control. IntenseIL-8 reactivity is visible in mesothelioma-positive controls.
369
INTERLEUKIN 8 AND PROLIFERATION OF MESOTHELIOMA
Research. on January 13, 2014. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from
It is well established that tumor cells produce a number of factorsthat promote tumor growth, tissue invasion, and metastatic spread.Melanoma cells secreted autocrine growth-stimulatory factors, such asMGSA (MGSA/GROa) or basic fibroblast growth factor. MGSA wasfound to have mitogenic activity for the melanoma cell line Hs 294T,which produces this factor (25). High levels of MGSA secretion wasobserved in diverse melanoma cell lines; however, it was absent inbenign nevus cells (26). Recently, another member of the C-X-Csupergene family, IL-8, was found to be secreted by melanoma cellsin an autocrine fashion. IL-8, potentiates proliferation of melanomacells and plays an important role in the tumorigenesis and growth ofmalignant melanoma (3, 18). This is supported by the fact thatantisense oligonucleotides targeted against human IL-8 mRNA inhib-ited cell proliferation, colony formation in soft agar, and secretion ofIL-8 into culture supernatants (18).In vivo experiments demonstratedthat expression of IL-8 correlates with the metastatic capability of 13human melanoma cell lines, which were injected into BALB/c mice(27). We demonstrated that mesothelioma cells similar to melanomacells release biologically active IL-8, which causes proliferation ofmesothelioma cells. The higher proliferative capacity of the tumor isdirectly related to its capacity to secrete more IL-8, and the differentialIL-8 response in different mesothelioma cell lines noticed may berelated to their aggressive growth.
Recent studies show that IL-8 receptor B (CXCR-2) is present onmelanoma cells and plays a role in the cell growth (28). A specific Abdirected against the NH2 terminus of IL-8 receptor B (28) as well asa novel inhibitor of IL-8 receptor B (25) have been shown to suppressthe growth of melanoma cells. Because of high a degree of sequencehomology of MGSA and IL-8, both cause receptor signal transductionwhen binding to the IL-8 receptor B (29). However, the presence ofanother, specific MGSA binding site has been theorized. Interestingly,IL-8 as well as MGSA bind with high affinity to the IL-8 receptor B,but the affinity of MGSA to the IL-8 receptor A is weak comparedwith the high affinity of IL-8 for this receptor. In an acute burnwound, IL-8 receptor B was restricted to the basal proliferatingcompartment in healing skin wounds, whereas higher concentrationsof IL-8 were detected in the outer layers of the epidermis, providinga chemotactic gradient for the migration of keratinocytes (30). These
results also suggest that the presence of IL-8 receptor B is critical forthe proliferative effect of IL-8.
IL-8 has been classified primarily by proinflammatory properties(31), but it has multiple effects. In response to inflammatory stimulisuch as lipopolysaccharide, IL-1, and tumor necrosis factor-a, IL-8 isproduced by a wide variety of cell types including monocytes, neu-trophils, lymphocytes, and endothelial and mesothelial cells (11,16,31). IL-8 activates neutrophils and increases their adhesion to endo-thelial cells, which leads to directional transendothelial migration(28). The effects of IL-8 is not confined only to neutrophils, but it haseffects on T and B lymphocytes, basophils, and IL-2-activated naturalkiller cells as well. IL-8 has a strong chemotactic activity on neutro-phils. The described complex effects suggest a functional role for IL-8in the first line of host defensein vivo. In addition, IL-8 inducesproliferation and chemotaxis of human umbilical vein endothelialcells (11) in vitro. In the rat sponge implant model (19) and in thecorneal micropocket model (11), IL-8 has been found to be a potentangiogenic factor in a pattern similar to that seen with tumor necrosisfactor-a, fibroblast growth factor, and vascular endothelial growth
Fig. 4. Growth curve of mesothelioma (CRL-2081, CRL-5915, and CRL-5820) andmesothelial (CRL-9444) cell lines in the presence of various concentrations of IL-8. Dataare expressed as means as a standardized percentage of stimulation of [3H]thymidineuptake in SFM after 48 h of culture;bars,SD. For each cell line, the experiments wererepeated four times, with triplicate samples. IL-8 induced a significant, dose-dependentincrease of [3H]thymidine incorporation in CRL-2081 and CRL-5915 mesotheliomas butnot in CRL-5820 and the mesothelial cell lines.pp, P , 0.001; andp, P , 0.05 whencompared with SFM.
Fig. 5. Effect of specific polyclonal anti-human IL-8 antibody on human mesotheliomaand mesothelial cells proliferation.A, data are expressed as means as a standardizedpercentage of stimulation of [3H]thymidine uptake in SFM after 48 h of culture;bars,SD.B, effect of IL-8 expressed as a percentage of stimulation of cell counts when 100% is thecell count in SFM at 48 h of incubation. For each cell line, the experiments were repeatedfour times, with triplicate samples;bars,SD. Fifty ng/ml of IL-8 were used for prolifer-ation; 10mg/ml of polyclonal goat anti-human IL-8 Ab was used to neutralize the IL-8.Neutralization of IL-8 significantly decreased the proliferation of CRL-2081 and 5915mesothelioma cell lines. Proliferation of the CRL-5820 mesothelioma and mesothelial(CRL-9444) cell lines remained unaltered.pp, P , 0.001; andp, P , 0.05 when comparedwith SFM.
370
INTERLEUKIN 8 AND PROLIFERATION OF MESOTHELIOMA
Research. on January 13, 2014. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from

factor. Tumor growth beyond 1–2 mm3 was dependent on angiogen-esis (32). The dysregulation of the balance between angiogenic andangiostatic factors is one of the mechanisms maintaining tumorgrowth by neovascularization. IL-8 is expressed in and secreted by avariety of transformed neoplastic cells (3, 33). Significantly elevatedIL-8 levels were found in human non-small cell lung cancer (adeno-carcinoma and squamous cell carcinoma), which were four timesgreater than normal lung tissue (17). These observations suggest thatthe more aggressive course of adenocarcinomas could be related totheir capacity to generate IL-8. Inhibition of IL-8, by addition ofneutralizing antisera, attenuated both corneal (17) and neoplastic (33)neovascularization. In non-small cell lung cancer, IL-8 did not act asan autocrine growth factor for proliferation; however, treatment withneutralizing antibody to IL-8 resulted in a decline of tumor-associatedvascular density, a reduction in primary tumor size, and reduction ofthe rate of metastasis (33). This well-described angiogenic effect ofIL-8 could also play an important role in the spread and growth ofmalignant mesotheliomas.
The findings described in this report demonstrate for the first timethat IL-8, which is produced by mesothelioma cells, is a importantfactor in growth regulation in a subgroup of human MMs. Likewise,the progression of MM may correlate with the level of autocrine IL-8production. However, it remains to be determined whether IL-8 me-diates its proliferative effects through IL-8 receptor B or through arelated molecule of the same receptor family, as in melanoma cells(25). The mechanism responsible for these findings needs to bestudied further.
REFERENCES
1. Connelly, R. R., Spirtas, R., Myers, M. H., Percy, C. L., and Fraumeni, J. F., Jr.Demographic patterns for mesothelioma in the United States. J. Natl. Cancer Inst.,78:1053–1060, 1987.
2. Branscheid, D., Krysa, S., Bauer, E., Bulzebruck, H., and Schirren, J. Diagnostic andtherapeutic strategy in malignant pleural mesothelioma. Eur. J. Cardio-Thorac. Surg.,5: 466–472, 1991.
3. Forster, E., Kirnbauer, R., Urbanski, A., Kock, A., Luger, T. A., and Schwarz, T.Human melanoma cells produce interleukin-8, which function as an autocrine growthfactor. Arch. Dermatol. Res.,283: 26, 1991.
4. Relf, M., LeJeune, S., Scott, P. A., Fox, S., Smith, K., Leek, R., Moghaddam, A.,Whitehouse, R., Bicknell, R., and Harris, A. L. Expression of the angiogenic factorsvascular endothelial cell growth factor, acidic and basic fibroblast growth factor,tumor growth factorb-1, platelet-derived endothelial cell growth factor, placentagrowth factor, and pleiotrophin in human primary breast cancer and its relation toangiogenesis. Cancer Res.,57: 963–969, 1997.
5. Halaban, R., Kwon, B. S., Ghosh, S., Delli Bovi, P., and Baird, A. bFGF as anautocrine growth factor for human melanomas. Oncogene Res.,3: 177–186, 1988.
6. Nanney, L. B., Mueller, S. G., Bueno, R., Peiper, S. C., and Richmond, A. Distri-butions of melanoma growth stimulatory activity of growth-regulated gene and theinterleukin-8 receptor B in human wound repair. Am. J. Pathol.,147: 1248–1260,1995.
7. Antony, V. B., Godbey, S. W., Kunkel, S. L., Hott, J. W., Hartman, D. L., Burdick,M. D., and Strieter, R. M. Recruitment of inflammatory cells to the pleural space.J. Immunol.,151: 7216–7223, 1993.
8. Strieter, R. M., Koch, A. E., Antony, V. B., Fick, R. B., Jr., Standiford, T. J., andKunkel, S. L. The immunopathology of chemotactic cytokines: the role of interleu-kin-8 and monocyte chemoattractant protein-1. J. Lab. Clin. Med.,123: 183–197,1994.
9. Peveri, P., Walz, A., Dewald, B., and Baggiolini, M. A novel neutrophil-activatingfactor produced by human mononuclear phagocytes. J. Exp. Med.,167: 1547–1559,1988.
10. Paccaud, J. P., Schifferli, J. A., and Baggiolini, M. NAP-1/IL-8 induces up-regulationof CR1 receptors in human neutrophil leukocytes. Biochem. Biophys. Res. Commun.,166: 187–192, 1990.
11. Koch, A. E., Polverini, P. J., Kunkel, S. L., Harlow, L. A., DiPietro, L. A., Elner,V. M., Elner, S. G., and Strieter, R. M. Interleukin-8 as a macrophage-derivedmediator of angiogenesis. Science (Washington DC),258: 1798–1801, 1992.
12. Strieter, R. M., Kunkel, S. L., Elner, V. M., Martonyi, C. L., Koch, A. E., Polverini,P. J., and Elner, S. G. Interleukin-8: a corneal factor that induces neovascularization.Am. J. Pathol.,141: 1279–1284, 1992.
13. Strieter, R. M., Polverini, P. J., Arenberg, D. A., Walz, A., Opdenakker, G., VanDamme, J., and Kunkel, S. L. Role of C-X-C chemokines as regulators of angiogen-esis in lung cancer. J. Leukocyte Biol.,57: 752–762, 1995.
14. Broaddus, V. C., Hebert, C. A., Vitangcol, R. V., Hoeffel, J. M., Bernstein, M. S., andBoylan, A. M. Interleukin-8 is a major neutrophil chemotactic factor in pleural liquidof patients with empyema. Am. Rev. Respir. Dis.,146: 825–830, 1992.
15. Boylan, A. M., Ruegg, C., Kim, K. J., Hebert, C. A., Hoeffel, J. M., Pytela, R.,Sheppard, D., Goldstein, I. M., and Broaddus, V. C. Evidence of a role for mesothelialcell-derived interleukin 8 in the pathogenesis of asbestos-induced pleurisy in rabbits.J. Clin. Invest.,89: 1257–1267, 1992.
16. Antony, V. B., Hott, J. W., Kunkel, S. L., Godbey, S. W., Burdick, M. D., andStrieter, R. M. Pleural mesothelial cell expression of C-C (monocyte chemotacticpeptide) and C-X-C (interleukin 8) chemokines. Am. J. Resp. Cell Mol. Biol.,12:581–588, 1995.
17. Smith, D. R., Polverini, P. J., Kunkel, S. L., Orringer, M. B., Whyte, R. I., Burdick,M. D., Wilke, C. A., and Strieter, R. M. Inhibition of interleukin 8 attenuatesangiogenesis in bronchogenic carcinoma. J. Exp. Med.,179: 1409–1415, 1994.
18. Schadendorf, D., Moller, A., Algermissen, B., Worm, M., Sticherling, M., andCzarnetzki, B. M. IL-8 produced by human malignant melanoma cellsin vitro is anessential autocrine growth factor. J. Immunol.,151: 2667–2675, 1993.
19. Hu, D. E., Hori, Y., and Fan, T. P. Interleukin-8 stimulates angiogenesis in rats.Inflammation,17: 135–143, 1993.
20. Andrews, P. M., and Porter, K. R. The ultrastructural morphology and possiblefunctional significance of mesothelial microvilli. Anat. Record,177: 409 – 426,1973.
21. Connell, N. D., and Rheinwald, J. G. Regulation of cytoskeleton in mesothelial cells:reversible loss of keratin and increase in vimentin during rapid growth in culture. Cell,34: 245–253, 1983.
22. Goodman, R. B., Wood, R. G., Martin, T. R., Hanson-Painton, O., and Kinasewitz,G. T. Cytokine stimulated human mesothelial cells produce chemotactic activity forneutrophils including NAP-1/IL-8. J. Immunol.,148: 457–465, 1992.
23. Lauber, B., Leuthold, M., Schmitter, D., Cano-Santos, J., Waibel, R., and Stahel,R. A. An autocrine mitogenic activity produced by a pleural human mesothelioma cellline. Int. J. Cancer,50: 943–950, 1992.
24. Hott, J. W., Sparks, J. A., Godbey, S. W., and Antony, V. B. Mesothelial cell responseto pleural injury: thrombin-induced proliferation and chemotaxis of rat pleural me-sothelial cells. Am. J. Respir. Cell Mol. Biol.,6: 421–425, 1992.
25. Hayashi, S., Kurdowska, A., Cohen, A. B., Stevens, M. D., Fujisawa, N., and Miller,E. J. A synthetic peptide inhibitor for alpha-chemokines inhibits the growth ofmelanoma cell lines. J. Clin. Invest.,99: 2581–2587, 1997.
26. Richmond, A., and Thomas, H. G. Melanoma growth stimulatory activity: isolationfrom human melanoma tumors and characterization of tissue distribution. J. Cell.Biochem.,36: 185–198, 1988.
27. Singh, R. K., Gutman, M., Radinsky, R., Bucana, C. D., and Fidler, I. J. Expressionof interleukin 8 correlates with the metastatic potential of human melanoma cells innude mice. Cancer Res.,54: 3242–3247, 1994.
28. Norgauer, J., Metzner, B., and Schraufstatter, I. Expression and growth-promotingfunction of the IL-8 receptor b in human melanoma cells. J. Immunol.,156: 1132–1137, 1996.
29. Arenberg, D. A., Kunkel, S. L., Polverini, P. J., Glass, M., Burdick, M. D., andStrieter, R. M. Inhibition of interleukin-8 reduces tumorigenesis of human non-smallcell lung cancer in SCID mice. J. Clin. Invest.,97: 2792–2802, 1996.
30. Lee, J., Horuk, R., Rice, G. C., Bennett, G. L., Camerato, T., and Wood, W. I.Characterization of two high affinity human interleukin-8 receptors. J. Biol. Chem.,267: 16283–16287, 1992.
31. Harada, A., Mukaida, N., and Matsushima, K. Interleukin-8 as a novel target forintervention therapy in acute inflammatory disease. Mol. Med. Today,2: 482–489,1996.
32. Folkman, J. Tumor angiogenesis. Adv. Cancer Res.,43: 175–203, 1985.33. Hebert, C. A., and Baker, J. B. Interleukin-8: a review. Cancer Invest.,11: 743–750,
1993.
371
INTERLEUKIN 8 AND PROLIFERATION OF MESOTHELIOMA
Research. on January 13, 2014. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from