spatiotemporal cortical dynamics underlying abstract and concrete word reading
TRANSCRIPT

Spatiotemporal Cortical Dynamics Underlying Abstract andConcrete Word Reading
Rupali P. Dhond1, Thomas Witzel1, Anders M. Dale2, and Eric Halgren1,31 MGH/MIT/HMS Athinoula A. Martinos Center for Biomedical Imaging, Charlestown, MA 02129
2 Departments of Neurosciences and Radiology, University of California at San Diego, La Jolla, CA92037
3 INSERM E9926, Marseilles, France
AbstractIn the current study, we used whole head, anatomically constrained magnetoencephalography(aMEG) to spatiotemporally map brain responses while subjects made abstract/concrete judgmentson visually presented words. Both word types evoked a similar posterior to anterior sequence ofcortical recruitment involving occipital, temporal, parietal, and frontal areas from ~100–900ms post-stimulus. A prominent left temporo-frontal N400m was smaller to abstract words, while the righttemporal N400m was smaller to concrete words, suggesting that differences may exist in theirsemantic representation. The left temporo-frontal decrease seen here for abstract words is consistentwith EEG studies indicating a smaller N400 for abstract words based on a more extensive oraccessible lexicosemantic network. Furthermore, in the current study, the N400m peaked at ~420msand was followed by a large, right hemisphere, medial occipitoparietal as well as lateral parietalresponse to concrete words peaking at ~550ms, perhaps embodying imagistic processing. These datasuggest that words may be initially understood using a left lateralized fronto-temporal verbal-linguistic system that for concrete words is supplemented after a short delay by a right parietal andmedial occipital imagistic network.
Keywordssemantic; MEG; imagery; frontal; temporal; parietal
INTRODUCTIONAbstract words (e.g. “curiosity” and “justice”) are difficult to visualize and in general refer toideas or concepts. Concrete words, such as “building” and “ocean”, describe tangible entitiesand are easily perceived as mental images. Behavioral studies have demonstrated differencesin response times and accuracy with respect to abstract and concrete words (Day, 1977; Krolland Merves, 1986). Two models have been advanced to explain such findings. In the ‘dual-coding theory’ (Paivio, 1991), both concrete and abstract words are understood using a verbal-linguistic semantic system, while concrete words use an additional nonverbal-imagistic system.This dual-coding of concrete words results in faster reaction times. In the ‘context-availability’model (Schwanenflugel et al., 1992), both concrete and abstract words are processed withinthe same semantic system with the reaction time advantage for concrete words being explainedby stronger links to contextual information in semantic memory. Thus, the dual-coding model
Address correspondence to: Rupali P. Dhond, MGH/MIT/HMS Athinoula A. Martinos Center for Biomedical Imaging, Room 2301,Building 149, 13th Street, Charlestown, MA 02129, Phone: (617) 726-8790 ; Fax: (617) 726-7422, [email protected].
NIH Public AccessAuthor ManuscriptHum Brain Mapp. Author manuscript; available in PMC 2007 April 1.
Published in final edited form as:Hum Brain Mapp. 2007 April ; 28(4): 355–362.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

predicts qualitative differences in the distribution of semantic memory traces, while thecontext-availability model predicts only quantitative differences.
Using event related EEG averages, several studies have found that concrete and abstract wordsevoke different scalp topographies, and thus atleast partially distinct cortical generators(Kounios and Holcomb, 1994; Holcomb et al., 1999; Swaab et al., 2002). Two differences werefound. The first was a negativity peaking at ~400ms that was more right-lateralized and anteriorfor concrete (vs. abstract) words. This negativity was interpreted as the extensively studiedN400 component. The N400 is evoked by words (visual, auditory or signed) and pronounceablenon-words, occurring in isolation or in sentences; it is inversely proportional to the ease withwhich the stimulus may be integrated into the current context, being reduced by item repetition,semantic priming and elevated word frequency (for reviews see Kutas and Hillyard, 1984;Halgren, 1990). The finding of different N400 scalp topographies for concrete and abstractwords was taken as evidence for dual-coding theories, suggesting that at least partially distinctsemantic systems exist for the processing of these words (Holcomb et al., 1999). Secondly, alater component, termed the N700, was modulated for concrete word processing only; thiscomponent was interpreted as reflecting imagistic processing (West and Holcomb, 2000).Although these studies are important in demonstrating processing dissociations betweenconcrete vs. abstract words, ambiguities associated with the inverse problem prevented themfrom locating the anatomical source of these effects.
Like EEG, magnetoencephalography (MEG) provides millisecond resolution and has similarcortical generators (Dale and Halgren, 2001). Several recent studies have found MEG activitywith similar task correlates and latency to the N400 component (Simos et al., 1997; Heleniuset al., 1998; Dale et al., 2000; Halgren et al., 2002; Penney et al., 2003; Puregger et al., 2003;Pylkkanen and Marantz, 2003). This ‘N400m’ decreases with repetition priming, word order,or semantic context. It is evoked by both auditory and visual words, as well as word-stems,presented in isolation or in sentences, in a variety of tasks probing memory, morpho-syntax,and semantics. Studies utilizing whole-head MEG and a distributed noise-normalized sourcemodeling technique constrained by individual subject cortical anatomy have consistentlylocalized the N400m to fronto-temporal (left > right) areas (Dale et al., 2000; Dhond et al.,2001; Halgren et al., 2002; Marinkovic et al., 2003). These areas have previously been shownto generate N400-like activity in depth electrodes chronically implanted prior to surgicaltherapy (Halgren et al., 1994b; Halgren et al., 1994a).
In the present study we used anatomically constrained, whole-head MEG to determine thespatiotemporal dynamics of cortical response underlying abstract and concrete word reading.A concrete/abstract judgment task was used because previous studies indicated that theseproduce ERP differences between abstract and concrete words that are similar, but larger, thanthose produced by a more neutral task such as lexical decision (Kounios and Holcomb,1994).
METHODSSubjects, Task and Stimulus Presentation
Subjects consisted of 10 normal, right-handed, native English speaking males 18–30 years ofage (Edinburgh Inventory (Oldfield, 1971)). Subjects had no history of mental or physicalillness, head injuries, or of drug/alcohol dependence/abuse and were further screened for MEGartifacts due to dental work or excessive eye-blinks.
During the task, subjects were instructed to silently read individually presented words and toindicate whether they were “abstract” or “concrete” by lifting either left middle or index fingerrespectively. Subjects were shown 240 abstract words randomly intermixed with 240 concrete
Dhond et al. Page 2
Hum Brain Mapp. Author manuscript; available in PMC 2007 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

words. Words were roughly matched in their number of letters with abstract (mean ± std. error,6.81 ± .10) being slightly longer than concrete (5.61 ± .10), t(473) = 8.64, p < .001. Both wordtypes had high imageability ratings (used when available), but concrete words (576.21 ± 3.1)were on average higher than abstract (400.77 ± 5.9) words, t(359) = 27.78, p < .001 (Coltheart,1981). During task presentation, abstract and concrete words were both of medium frequency,55.01 ± 5.9 for abstract, and 33.79 ± 3.4 for concrete (Kucera, 1967), t(454) = 3.121, p < .002.To evaluate differences between abstract and concrete word processing, trials were re-averagedprior to subtraction so that the average word frequency within each category was 33 words permillion and only trials where subjects responded correctly were used. During the task, wordstimuli were presented in Geneva font as white letters on a black background using Mac Probesoftware (Hunt, 1994). Words were presented in the central 5% of visual angle for 700ms.Between word presentations subjects were asked to fixate on a central “+” sign. The intervalbetween onset of successive words was fixed at 2 seconds. The mapping analysis wasperformed from stimulus onset to 1000ms post-stimulus and a 500ms pre-stimulus baselinewas used.
MEG RecordingMEG was recorded using a Neuromag VectorView™ (Elekta, Stockholm, Sweden) with 204gradiometer channels covering the entire scalp. MEG recordings took place within amagnetically shielded room (IMEDCO, Hagendorf, Switzerland). Signals were sampled at601Hz after filtering from 0.1–200 Hz. Data were lowpass filtered at 20 Hz and separateaverages of each condition were constructed for all subjects. Trials were rejected from analysisbased on amplitude criteria supplemented by visual inspection for contamination by artifacts(identified as peak-to-peak amplitude >5000fT/cm in any channel), or eye-blinks (>200 μV inthe EOG electrode). Head movement was minimized using an individually molded bite bar(Marinkovic et al., In press).
Cortical Surface ReconstructionA geometrical representation of the cortical surface of each subject was obtained usingprocedures described previously (Dale et al., 1999; Fischl et al., 1999a). First, high-resolution3-D T1-weighted structural images were acquired for each subject using a 1.5T Picker Eclipse(Marconi Medical, Cleveland, OH). Then, the cortical white matter was segmented, and theestimated border between gray and white matter was tessellated, providing a topologicallycorrect representation of the surface with about 150,000 vertices per hemisphere. For theinverse computation, the cortical surface was decimated to approximately 3000 dipoles, whichis roughly equivalent to 1 dipole every 10 mm along the cortical surface. Finally the foldedsurface tessellation was “inflated”, in order to unfold cortical sulci, thereby providing aconvenient format for visualizing cortical response patterns (Dale et al., 1999; Fischl et al.,1999a). For purposes of intersubject averaging, the reconstructed surface for each subject wasmorphed into an average spherical representation, optimally aligning sulcal and gyral featuresacross subjects while minimizing metric distortions and shear (Fischl et al., 1999a) and MEGresponse power was mapped onto an average sulcal-gyral pattern. Compared to volumetricmorphing into Talairach (Collins et al., 1994) space, this method has been found to providebetter alignment across subjects of functional activation in a verbal task (Fischl et al., 1999b)and allows direct localization to regular gyri.
Forward solutionThe boundary element method (BEM) was used for calculating the signal expected at eachMEG sensor, for each dipole location (deMunck, 1992; Oostendorp and Van Oosterom,1992). The computation of the MEG forward solution has been shown to only require the innerskull boundary to achieve an accurate solution (Meijs et al., 1987; Meijs and Peters, 1987;
Dhond et al. Page 3
Hum Brain Mapp. Author manuscript; available in PMC 2007 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Hamalainen and Sarvas, 1989). The T1-weighted MRI described above was used forconstruction of the inner skull surface. The MEG sensor coordinate system was aligned withthe MRI coordinate system using four head position (HPI) coils, attached to cardinal locationson the scalp (Hämäläinen et al., 1993). The HPI coils generate weak magnetic signals, allowingthem to be directly localized by the MEG sensors before and after the recording session. Thepositions of the HPI coils with respect to the subject’s head (and thus MRI) are determined bymeasuring ~80 points (including the HPI coils) around the head using a Polhemus FastTrack3-D digitizer. These digitized points were later registered with the MRI image. Since the HPIcoils were thus localized with respect to both the MEG sensors and the structural MRI, acommon coordinate system was established allowing neural response to be estimated for eachcortical location.
Inverse solutionTo estimate the timecourses of cortical response, we used the noise-normalized, anatomicallyconstrained linear estimation approach described by Dale et al (2000). This approach is similarto the generalized least-squares or weighted minimum norm solution (Hamalainen andIlmoniemi, 1984), except that the modeled sources were constrained to lie in the cortical surface(as determined above) (Dale and Sereno, 1993), and the estimate was normalized for noisesensitivity such that source signal to noise ratio rather than current dipole moment was mapped(Dale et al., 2000). The noise normalization also has the effect of greatly reducing the variationin the point-spread function between locations (Liu et al., 2002). This approach providesstatistical parametric maps of cortical response, similar to the statistical maps typicallygenerated using fMRI, or PET data, but with a temporal resolution of 5ms or less. Note thatalthough the noise normalized values are not identical to the source current estimates, thesevalues are directly proportional to the current power estimated for a given site. This is aconsequence of the fact that the noise normalized value is calculated as the ratio between thetime variant signal power and the time invariant noise power. Since in the current study, no apriori assumptions were made about the local dipole orientation, three components werecalculated for each location. The noise normalized estimate of the source power (sum ofsquared source component strengths) at location i is given by
qi(t) =
∑j∈Gi
(w j ⋅ x(t))2
∑j∈Gi
w jCw jT ,
where Gi is the set of (three) dipole component indices for the ith location, and wi denotes theith row of the inverse operator W (Dale and Sereno, 1993; Liu et al., 1998; Dale et al., 2000).The noise covariance matrix C for all conditions was estimated from the baseline of the rawsignals after all filtering is applied. For the comparisons (i.e. Abstract vs. Concrete), thewaveforms of the individual conditions were subtracted prior to estimating the differentialsource activity pattern. Both non-subtracted and subtracted conditions were tested for the nullhypothesis that the signal was noise. Under the null hypothesis, qi (t) is F-distributed, withthree degrees of freedom for the numerator and a large number of degrees of freedom for thedenominator (about 160,000 time points per sensor were used to estimate the spatial sensornoise covariance). Noise-sensitivity normalized cortical surface constrained minimum norminverse solutions were calculated every 5 ms for every condition and every individual. Thesquare roots of these values were then averaged on the cortical surface across individuals afteraligning their sulcal-gyral patterns. The square root was used in order to de-emphasize outliersand ensure that the result is linearly proportional to the magnitude of the estimated sources(Dale et al., 2000; Dhond et al., 2001). The distribution under the null hypothesis of theaveraged estimates was estimated using monte carlo simulations in order to obtain significance
Dhond et al. Page 4
Hum Brain Mapp. Author manuscript; available in PMC 2007 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

thresholds. Figure 1 shows the posterior to anterior progression of responses to abstract andconcrete words, while Figure 2 demonstrates differences between these conditions. For Figure1, the same significance scales were used for both abstract and concrete words. For both Figure1 and 2 the minimum significance values were p<10−7 for threshold, p<10−10 indicated by fullred. The statistical values were chosen to maximize the dynamic range of the color scale, andthe statistical thresholds were chosen to conservatively correct for the number of comparisonsbeing performed. Figure 3 shows average timecourses for the indicated areas smoothed over10ms intervals.
It should also be noted that MEG gradiometer data in this study are unlikely to reveal responsesthat can be confidently assigned to deep, non-cortical structures. These structures lack thesynaptic/dendritic arrangements necessary to generate large MEG signals. Furthermore, theyare far from the sensors, and MEG gradiometer signals decline greatly with distance. Althoughsubcortical responses can sometimes be detected by MEG, this usually requires a large numberof averages, i.e. >1000 and a lack of response in overlying generators (such as may occur atvery short latencies prior to cortical involvement.) In contrast, the current study examinesresponse at relatively long latencies and averaged across relatively fewer trials. For thesereasons, deep non-cortical structures such as the basal ganglia and corpus callosum are notincluded as sources in the spatiotemporal maps of the medial and ventral surfaces. In general,it must be borne in mind that the estimation of electromagnetic sources from extracranial datais dependent on prior assumptions.
RESULTSBehavioral data
Average response times were longer for abstract (1164 ± 147 ms) than concrete (1045 ± 120ms) words with t(9) = 5.82, p < .000126. Subjects responded 76.47% correct for abstract wordsand 95.64% correct for concrete words.
Overall brain response pattern during abstract/concrete word readingBrain responses to abstract and concrete word presentations demonstrated similar posterior toanterior sequences of cortical recruitment as shown in Figure 1.
Response first occurred bilaterally at ~90 ms post-stimulus within the occipital pole andcalcarine sulcus then spread to anteroventral occipital cortex corresponding approximately toBrodmann area 19 (~BA19, left > right) by ~135ms. At ~200ms response was strongest withinleft temporo-parietal and occipitotemporal (ventral ~BA 37/19), followed by anterior andlateral temporal (~BA 38, 20, 21) regions. Response within the left ventral (~BA 11/47) andlateral (~BA 45, 44, 9, 46, 25) prefrontal cortices peaked at ~400ms. At this time, there wasalso strong bilateral recruitment of medial cortices, including the posterior cingulate area (alsoretrosplenial cortex), the precuneus (~BA 7/31), and medial occipital cortex (~BA 17/18). By~700ms activity had strongly decreased in many areas but remained prominent within anteriorfronto-temporal, as well as medial occipitoparietal cortical areas.
Differential response between abstract and concrete wordsDifferential response between abstract and concrete words is mapped in Figure 2. Locationswith significantly different response are displayed on the cortical surface at latencies of~330ms, ~465ms and ~550ms. Areas prominently involved included left prefrontal, bilateralanterior temporal, left lateral temporal, right lateral and medial parietal, and posterior cingulateareas (including retrosplenial cortex).
Dhond et al. Page 5
Hum Brain Mapp. Author manuscript; available in PMC 2007 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

In Figure 3, the timecourses of the responses to concrete and abstract words are shown forselected regions. The spread of activity from the occipital pole was seen as a sharp rise frombaseline, occurring at progressively longer latencies in more anterior locations. This sharp riseended in a peak, also at increasingly longer latencies in anterior locations, in the order occipital,parietal, anterior temporal, and prefrontal. For example, the initial occipital response begins at~90ms and peaks at ~150, whereas the initial anterior temporal response begins at ~100ms andpeaks at ~210. These initial responses do not differentiate between abstract and concrete words.The differential response that occurred between word types was most prominent in left anteriortemporal, ventral and lateral prefrontal areas, peaking at ~400ms. This response was larger toconcrete words, but after ~620ms, response in the same areas became larger to abstract words.At a slightly longer latency (peak ~500ms) in the right anterior temporal lobe, the responsewas larger to abstract words, and did not reverse. A large, sustained differential response wasalso present in right medial and lateral parietal sites as well as the right retrosplenial and medialoccipital cortex. These did not peak until ~550ms.
DISCUSSIONGeneral spatiotemporal pattern underlying abstract and concrete word reading
Overall, the spatiotemporal cortical response patterns supporting abstract and concrete wordreading were similar and involved a common, distributed cortical network (Figure 1). Corticalregions were recruited in a posterior to anterior sequence, and following ~250ms post-stimulus,multiple areas within occipital, temporal, parietal and frontal cortex were simultaneouslyactive, and remained so for hundreds of milliseconds. This pattern of response has been seenpreviously within numerous aMEG studies investigating language processing (Dale et al.,2000;Dhond et al., 2001;Halgren et al., 2002;Marinkovic et al., 2003) and is also supported byintracranial EEG data (Halgren et al., 1994b;Halgren et al., 1994a;McCarthy et al., 1995;Nobreand McCarthy, 1995).
Following the initial bilateral response in the occipital pole, by ~135ms left-lateralized activitywas significant in anteroventral occipital areas implicated in form processing of words(Tarkiainen et al., 1999; Cohen et al., 2000; Dhond et al., 2001; Tarkiainen et al., 2002;Marinkovic et al., 2003). This activity continued to advance along the ventral visual pathwayand in addition, at ~200ms was also prominent in classical posterior language areas, includingtemporo-parietal regions implicated in phonological processing (Rumsey et al., 1997; Paulesuet al., 2000; Price, 2000). Later responses were predominantly in anterior temporal, leftprefrontal, and right parietal regions that also showed differential responses to abstract versusconcrete words, and are discussed below. The differential response occurred in areas that werealready active, consistent with it representing a network interaction (Dale et al., 2000).
Differential Responses to Abstract and Concrete Words: Modulation of Left HemisphereSemantic, and Right Hemisphere Imagery Related Areas
In this study, left fronto-temporal areas previously associated with the N400 in iEEG and aMEGstudies, responded more to concrete than to abstract words, beginning ~330ms, peaking shortlyafter 400ms. Conversely, right anterior temporal areas also previously associated with theN400m demonstrated an increased response to abstract words. Furthermore, peaking at slightlylonger latencies (~550ms), right medial occipitotemporal and lateral parietal areas, oftenassociated with imagery, demonstrated more significant response for concrete words.Previously, differences in the N400 scalp distribution evoked by concrete versus abstract wordshave been taken as evidence for the existence of multiple representations of meaning, imagisticand non-imagistic (Kounios and Holcomb, 1994; Holcomb et al., 1999; West and Holcomb,2000). The present data confirm these studies in finding different topographies for the MEGresponse to abstract versus concrete words, and further suggest that that these differences may
Dhond et al. Page 6
Hum Brain Mapp. Author manuscript; available in PMC 2007 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

be due to a stronger non-imagistic representation (left hemisphere) for abstract words and astronger imagistic representation (right hemisphere) for concrete words.
The N400 is triggered by potentially meaningful stimuli and is thought to embody processingwithin an associative semantic network, encompassing the integration/encoding of the currentevent with an ongoing context. A decreased N400 is observed when lexicosemantic integrationwith the current context is facilitated by contextual congruity, semantic and repetition priming,late sentence position, or elevated word frequency (Halgren, 1990; Kutas and Federmeier,2000). In the present task, abstract and concrete words were presented in random order so theircognitive context was equated. The average word frequency of the lists was also matched forassessment of spatiotemporal differences. Furthermore, all words were presented only onceand were unrelated so neither semantic nor repetition priming would be expected. Collectively,this suggests that the decreased left N400m to abstract words represents a more efficient orextensive representation for these words within frontotemporal networks. Analogously, it maybe inferred that the relatively decreased right N400m to concrete words represents a moreefficient or extensive representation within right hemisphere networks. Our findings ofincreased engagement of left frontotemporal areas for concrete words and of right temporalareas for abstract words is supported by hemodynamic studies. Specifically, greater activationof left temporal areas for concrete words has been found with PET (Noppeney and Price,2002) as well as fMRI (Fiebach and Friederici, 2004). Stronger activation of right temporalareas by abstract (vs. concrete) words has also been demonstrated (Mellet et al., 1998; Kiehlet al., 1999; Perani et al., 1999). Collectively, the current results are consistent with somespecialization of frontotemporal lexicosemantic networks with the left hemisphere areasrepresenting abstract words more efficiently, and right hemisphere networks representingconcrete words more efficiently.
Subsequent to the N400 effects, and peaking at ~550ms, right medial occipitoparietal andlateral parietal areas demonstrated more significant response for concrete words.Hemodynamic studies have demonstrated imagery related activation within right parieto-temporal areas during the processing of concrete (Dehghani et al., 2004) or animal words(Lambert et al., 2002) in comparison with abstract words. The precuneus has also beenimplicated in mental imagery (Shallice et al., 1994 Grasby, 1993 #5420), though it is possiblethat this area is also involved in more general retrieval processes (Buckner et al., 1996; Krauseet al., 1999; Schmidt et al., 2002). Furthermore, there was significant differential responsewithin the right medial occipital cortex (anterior portions of the calcarine sulcus). Studies havesuggested that primary visual areas may participate mental imagery (for review Kosslyn et al.,2001). In the present study, the increased right medial occipitoparietal and lateral parietalresponses to concrete words (~400–700ms) suggests that their greater imageability allows themto engage additional encoding processes.
Strong abstract/concrete differences within right parietal areas occurred at longer latencies thanthose seen within left hemisphere frontotemporal areas, however, the duration of theseresponses are temporally overlapping and presumably support different underlying processesthat may interact. In particular, the early results of lexicosemantic processing in frontotemporalareas began to diverge for abstract/concrete words at ~300ms, this may have lead to theselective engagement of occipitoparietal and lateral parietal areas, possibly supportingimagery, for concrete words at ~400ms (when abstract/concrete waveforms began to divergethere). The differential response to concrete words in occipitoparietal and lateral parietal areaspeaks at ~550ms, subsequently at ~650ms the differential response in left frontotemporal areasreverses. One possibility is that the right occipitoparietal imagery-related processing maycontribute information to the frontotemporal N400m, thus leading to a faster termination ofthe N400m to concrete words, and ultimately a faster reaction time.
Dhond et al. Page 7
Hum Brain Mapp. Author manuscript; available in PMC 2007 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

CONCLUSIONWe found that all words evoked the typical sequence of cortical recruitment observed inprevious studies of word-reading thus, lending support for the existence of a common,distributed, intracortical network underlying word reading. However, the amplitude and time-course of the left fronto-temporal N400m was different between abstract and concrete words.In addition, later right medial occipitoparietal and lateral parietal processes were larger forconcrete words. These results were interpreted as indicating that although the sameanatomically extended fronto-temporal lexico-semantic network is used to encode concreteand abstract words, different parts of the network are relatively more specialized for eitherconcrete or abstract words. Furthermore, the results suggest that right parietal and occipitalimagistic processing contributes to the later parts of the lexico-semantic encoding for concretewords, allowing it to successfully conclude sooner than abstract words, resulting in theirreaction time advantage.
Acknowledgements
This work was funded by USPHS (NIH grants NS18741, NS44623), the Mental Illness and Neuroscience Discovery(MIND) Institute, and the National Center for Research Resources (P41RR14075). We thank Ksenija Marinkovic,Anthony Wagner, Dave Post, Sharelle Baldwin, Jeremy Jordin, Bruce Fischl, Arthur Liu, Jeffrey Lewine, Kim Paulsonand Bruce Rosen
ReferencesBuckner RL, Raichle ME, Miezin FM, Petersen SE. Functional anatomic studies of memory retrieval for
auditory words and visual pictures. J Neurosci 1996;16:6219–6235. [PubMed: 8815903]Cohen L, Dehaene S, Naccache L, Lehericy S, Dehaene-Lambertz G, Henaff MA, Michel F. The visual
word form area: spatial and temporal characterization of an initial stage of reading in normal subjectsand posterior split-brain patients [In Process Citation]. Brain 2000;123:291–307. [PubMed: 10648437]
Collins DL, Neelin P, Peters TM, Evans AC. Data in Standardized Talairach Space. Journal of ComputerAssisted Tomography 1994;18:292–205. [PubMed: 8126285]
Coltheart M. The MRC psycho-linguistic database. Quarterly Journal of Experimemtal Psychology1981;33A:497–505.
Dale AM, Sereno MI. Improved localization of cortical activity by combining EEG and MEG with MRIcortical surface reconstruction: A linear approach. Journal of Cognitive Neuroscience 1993;5:162–176.
Dale AM, Halgren E. Spatiotemporal mapping of brain activity by integration of multiple imagingmodalities. Curr Opin Neurobiol 2001;11:202–208. [PubMed: 11301240]
Dale AM, Fischl B, Sereno MI. Cortical surface-based analysis I: Segmentation and surfacereconstruction. NeuroImage 1999;9:179–194. [PubMed: 9931268]
Dale AM, Liu AK, Fischl BR, Buckner RL, Belliveau JW, Lewine JD, Halgren E. Dynamic statisticalparametric mapping: combining fMRI and MEG for high-resolution imaging of cortical activity.Neuron 2000;26:55–67. [PubMed: 10798392]
Day J. Right-hemisphere language processing in normal right-handers. J Exp Psychol Hum PerceptPerform 1977;3:518–528. [PubMed: 886282]
Dehghani N, Najafi AH, Kharazi HH. Effect of mental imagery duration on functional brain mapping:an fMRI study. Brain Cogn 2004;54:166–168. [PubMed: 15025055]
deMunck JC. A linear dicretization of the volume conductor boundary integral equation using analyticallyintegrated elements. IEEE Transactions on Biomedical Engineering 1992;39:986–990. [PubMed:1473829]
Dhond RP, Buckner RL, Dale AM, Marinkovic K, Halgren E. Spatiotemporal maps of brain activityunderlying word generation and their modification during repetition priming. J Neurosci2001;21:3564–3571. [PubMed: 11331385]
Fiebach CJ, Friederici AD. Processing concrete words: fMRI evidence against a specific right-hemisphereinvolvement. Neuropsychologia 2004;42:62–70. [PubMed: 14615076]
Dhond et al. Page 8
Hum Brain Mapp. Author manuscript; available in PMC 2007 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fischl B, Sereno MI, Dale AM. Cortical surface-based analysis II: Inflation, flattening, a surface-basedcoordinate system. NeuroImage 1999a;9:195–207. [PubMed: 9931269]
Fischl B, Sereno MI, Tootell RB, Dale AM. High-resolution intersubject averaging and a coordinatesystem for the cortical surface. Hum Brain Mapp 1999b;8:272–284. [PubMed: 10619420]
Halgren E (1990) Insights from evoked potentials into the neuropsychological mechanisms of reading.In: Neurobiology of Cognition (Scheibel A, Weschsler A, eds), pp 103–150. New York: Guilford.
Halgren E, Baudena P, Heit G, Clarke JM, Marinkovic K, Clarke M. Spatio-temporal stages in face andword processing. 1. Depth-recorded potentials in the human occipital, temporal and parietal lobes.Journal of Physiology (Paris) 1994a;88:1–50.
Halgren E, Baudena P, Heit G, Clarke JM, Marinkovic K, Chauvel P, Clarke M. Spatio-temporal stagesin face and word processing. 2. Depth-recorded potentials in the human frontal and Rolandic cortices.Journal of Physiology (Paris) 1994b;88:51–80.
Halgren E, Dhond RP, Christensen N, Van Petten C, Marinkovic K, Lewine JD, Dale AM. N400-likemagnetoencephalography responses modulated by semantic context, word frequency, and lexicalclass in sentences. Neuroimage 2002;17:1101–1116. [PubMed: 12414253]
Hamalainen MS, Ilmoniemi RJ (1984) Interpreting measured magnetic fields of the brain: Estimates ofcurrent distribution. Helsinki: University of Technology, Dept. of Technical Physics Report TKK-F-A559.
Hamalainen MS, Sarvas J. Realistic conductivity geometry model of the human head for interpretationof neuromagnetic data. IEEE Trans Biomed Eng 1989;36:165–171. [PubMed: 2917762]
Hämäläinen MS, Hari R, Ilmoniemi RJ, Knuutila J, Lounasmaa OV. Magnetoencephalography - theory,instrumentation, and applications to noninvasive studies of the working human brain. Reviews ofModern Physics 1993;65:413–497.
Helenius P, Salmelin R, Service E, Connolly JF. Distinct time courses of word and context comprehensionin the left temporal cortex. Brain 1998;121:1133–1142. [PubMed: 9648548]
Holcomb PJ, Kounios J, Anderson JE, West WC. Dual-coding, context-availability, and concretenesseffects in sentence comprehension: an electrophysiological investigation. J Exp Psychol Learn MemCogn 1999;25:721–742. [PubMed: 10368929]
Hunt SMJ. MacProbe: A Macintosh-based experimenter’s workstation for the cognitive sciences.Behavioral Research Methods,Instrumentation and Computing 1994;26:345–351.
Kiehl KA, Liddle PF, Smith AM, Mendrek A, Forster BB, Hare RD. Neural pathways involved in theprocessing of concrete and abstract words. Hum Brain Mapp 1999;7:225–233. [PubMed: 10408766]
Kosslyn SM, Ganis G, Thompson WL. Neural foundations of imagery. Nat Rev Neurosci 2001;2:635–642. [PubMed: 11533731]
Kounios J, Holcomb PJ. Concreteness effects in semantic processing: ERP evidence supporting dual-coding theory. J Exp Psychol Learn Mem Cogn 1994;20:804–823. [PubMed: 8064248]
Krause BJ, Schmidt D, Mottaghy FM, Taylor J, Halsband U, Herzog H, Tellmann L, Muller-GartnerHW. Episodic retrieval activates the precuneus irrespective of the imagery content of word pairassociates. A PET study. Brain 1999;122 ( Pt 2):255–263. [PubMed: 10071054]
Kroll JF, Merves JS. Lexical access for concrete and abstract words. J Exp Psychol Learn Mem Cogn1986;12:92–107.
Kucera H, & Francis, W. N. (1967). .Providence, RI: Brown University Press. (1967) Computationalanalysis of present-day American English. Providence, RI: Brown University Press.
Kutas M, Hillyard SA. Brain potentials during reading reflect word expectancy and semantic association.Nature 1984;307:161–163. [PubMed: 6690995]
Kutas M, Federmeier KD. Electrophysiology reveals semantic memory use in language comprehension.Trends Cogn Sci 2000;4:463–470. [PubMed: 11115760]
Lambert S, Sampaio E, Scheiber C, Mauss Y. Neural substrates of animal mental imagery: calcarinesulcus and dorsal pathway involvement--an fMRI study. Brain Res 2002;924:176–183. [PubMed:11750903]
Liu AK, Belliveau JW, Dale AM. Spatiotemporal imaging of human brain activity using fMRIconstrained MEG data: Monte Carlo simulations. Proceedings of the National Academy of Sciencesof the United States of America 1998;95:8945–8950. [PubMed: 9671784]
Dhond et al. Page 9
Hum Brain Mapp. Author manuscript; available in PMC 2007 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Liu AK, Dale AM, Belliveau JW. Monte Carlo simulation studies of EEG and MEG localization accuracy.Human Brain Mapping 2002;16:47–62. [PubMed: 11870926]
Marinkovic K, Cox B, Reid K, Halgren E (In press) Head position in the MEG helmet affects thesensitivity to anterior sources. Neurology and Clinical Neurophysiology.
Marinkovic K, Dhond RP, Dale AM, Glessner M, Carr V, Halgren E. Spatiotemporal dynamics ofmodality-specific and supramodal word processing. Neuron 2003;38:487–497. [PubMed: 12741994]
McCarthy G, Nobre AC, Bentin S, Spencer DD. Language-related field potentials in the anterior-medialtemporal lobe: I. Intracranial distribution and neural generators. Journal of Neuroscience1995;15:1080–1089. [PubMed: 7869084]
Meijs JWH, Peters MJ. The EEG and MEG, Using a model of eccentric spheres to describe the head.IEEE Transactions on Biomedical Engineering 1987;34:913–920. [PubMed: 3692512]
Meijs JWH, Bosch FGC, Peters MJ, Lopes da Silva FH. On the magnetic field distribution generated bya dipolar current source situated in a realistically shaped compartment model of the head.Electroencephalography and Clinical Neurophysiology 1987;66:286–298. [PubMed: 2434313]
Mellet E, Tzourio N, Denis M, Mazoyer B. Cortical anatomy of mental imagery of concrete nouns basedon their dictionary definition. Neuroreport 1998;9:803–808. [PubMed: 9579669]
Nobre AC, McCarthy G. Language-related field potentials in the anterior-medial temporal lobe: II. Effectsof word type and semantic priming. J Neurosci 1995;15:1090–1098. [PubMed: 7869085]
Noppeney U, Price CJ. Retrieval of visual, auditory, and abstract semantics. Neuroimage 2002;15:917–926. [PubMed: 11906232]
Oldfield. The assesment and analysis of handedness: the Edinburgh inventory. Neuropsychologia1971;9:97–113. [PubMed: 5146491]
Oostendorp TF, Van Oosterom A (1992) Source parameter estimation using realistic geometry inbioelectricity and biomagnetism. In: Biomagnetic Localization and 3D Modeling (Nenonen J, RajalaHM, Katila T, eds). Helsinki: Helsinky Univ. of Technology, Rport TKK-F-A689.
Paivio A. Dual Coding Theory: Retrospect and Current Status. Canadian Journal of Psychology1991;45:255–287.
Paulesu E, McCrory E, Fazio F, Menoncello L, Brunswick N, Cappa SF, Cotelli M, Cossu G, Corte F,Lorusso M, Pesenti S, Gallagher A, Perani D, Price C, Frith CD, Frith U. A cultural effect on brainfunction. Nat Neurosci 2000;3:91–96. [PubMed: 10607401]
Penney TB, Maess B, Busch N, Derrfuss J, Mecklinger A. Cortical activity reduction with stimulusrepetition: a whole-head MEG analysis. Brain Res Cogn Brain Res 2003;16:226–231. [PubMed:12668231]
Perani D, Schnur T, Tettamanti M, Gorno-Tempini M, Cappa SF, Fazio F. Word and picture matching:a PET study of semantic category effects. Neuropsychologia 1999;37:293–306. [PubMed: 10199643]
Price CJ. The anatomy of language: contributions from functional neuroimaging. Journal of Anatomy2000;197:335–359. [PubMed: 11117622]
Puregger E, Walla P, Deecke L, Dal-Bianco P. Magnetoencephalographic--features related to mildcognitive impairment. Neuroimage 2003;20:2235–2244. [PubMed: 14683725]
Pylkkanen L, Marantz A. Tracking the time course of word recognition with MEG. Trends Cogn Sci2003;7:187–189. [PubMed: 12757816]
Rumsey JM, Horwitz B, Donohue BC, Nace K, Maisog JM, Andreason P. Phonological and orthographiccomponents of word recognition. A PET- rCBF study. Brain 1997;120:739–759. [PubMed: 9183247]
Schmidt D, Krause BJ, Mottaghy FM, Halsband U, Herzog H, Tellmann L, Muller-Gartner HW. Brainsystems engaged in encoding and retrieval of word-pair associates independent of their imagerycontent or presentation modalities. Neuropsychologia 2002;40:457–470. [PubMed: 11684178]
Schwanenflugel PJ, Akin C, Luh WM. Context availability and the recall of abstract and concrete words.Mem Cognit 1992;20:96–104.
Shallice T, Fletcher P, Frith CD, Grasby P, Frackowiak RS, Dolan RJ. Brain regions associated withacquisition and retrieval of verbal episodic memory. Nature 1994;368:633–635. [PubMed: 8145849]
Simos PG, Basile LF, Papanicolaou AC. Source localization of the N400 response in a sentence-readingparadigm using evoked magnetic fields and magnetic resonance imaging. Brain Res 1997;762:29–39. [PubMed: 9262155]
Dhond et al. Page 10
Hum Brain Mapp. Author manuscript; available in PMC 2007 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Swaab TY, Baynes K, Knight RT. Separable effects of priming and imageability on word processing: anERP study. Brain Res Cogn Brain Res 2002;15:99–103. [PubMed: 12433385]
Tarkiainen A, Cornelissen PL, Salmelin R. Dynamics of visual feature analysis and object-levelprocessing in face versus letter-string perception. Brain 2002;125:1125–1136. [PubMed: 11960901]
Tarkiainen A, Helenius P, Hansen PC, Cornelissen PL, Salmelin R. Dynamics of letter string perceptionin the human occipitotemporal cortex. Brain 1999;122 ( Pt 11):2119–2132. [PubMed: 10545397]
West WC, Holcomb PJ. Imaginal, semantic, and surface-level processing of concrete and abstract words:an electrophysiological investigation. J Cogn Neurosci 2000;12:1024–1037. [PubMed: 11177422]
Dhond et al. Page 11
Hum Brain Mapp. Author manuscript; available in PMC 2007 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 1. Main Sequence of Cortical Response During Abstract and Concrete Word ReadingBoth word conditions demonstrated similar cortical response patterns beginning with thebilateral visual response at ~90ms within occipital cortex (yellow arrow). By ~135ms responsemoved anteriorly to occipitotemporal regions (left > right, green arrow), and at ~200ms wasstrongest within posterior and lateral temporal cortex (purple arrow). At ~400ms, response wasstrong within ventral and lateral prefrontal regions (left > right) as well as anterolateral temporalcortex (blue arrows) similar to that seen for the N400m (Halgren et al., 2002). During this timethere was also strong bilateral response within anterior and posterior cingulate areas. Activityis also prominent in bilateral medial occipital cortex. At ~700ms activity remained prominentwithin anterior fronto-temporal and medial occipitoparietal (orange arrows) cortical areas. In
Dhond et al. Page 12
Hum Brain Mapp. Author manuscript; available in PMC 2007 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

the current figure, threshold values are the same for both abstract and concrete words with aminimum significance for response of p < 10−7 and full red indicating a minimum of p <10−9.
Dhond et al. Page 13
Hum Brain Mapp. Author manuscript; available in PMC 2007 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
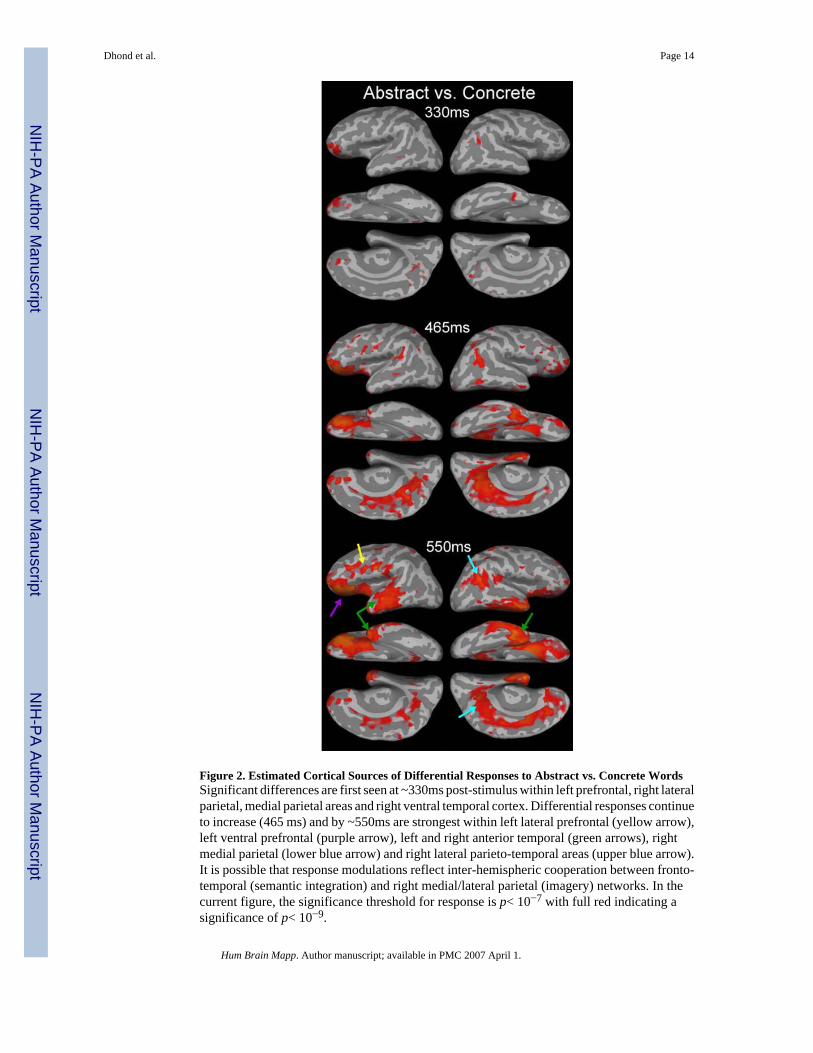
Figure 2. Estimated Cortical Sources of Differential Responses to Abstract vs. Concrete WordsSignificant differences are first seen at ~330ms post-stimulus within left prefrontal, right lateralparietal, medial parietal areas and right ventral temporal cortex. Differential responses continueto increase (465 ms) and by ~550ms are strongest within left lateral prefrontal (yellow arrow),left ventral prefrontal (purple arrow), left and right anterior temporal (green arrows), rightmedial parietal (lower blue arrow) and right lateral parieto-temporal areas (upper blue arrow).It is possible that response modulations reflect inter-hemispheric cooperation between fronto-temporal (semantic integration) and right medial/lateral parietal (imagery) networks. In thecurrent figure, the significance threshold for response is p< 10−7 with full red indicating asignificance of p< 10−9.
Dhond et al. Page 14
Hum Brain Mapp. Author manuscript; available in PMC 2007 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 3. Timecourses for Selected Locations, Estimated for Concrete (red) and Abstract (black)WordsThe initial response peak at ~135ms in lateral occipital, ~180ms in parietal, ~210ms intemporal, and ~230ms in frontal cortices (purple arrows), is equal to concrete and abstractwords, and is followed by sustained distributed procession in all brain areas. Four phases ofdifferential response are then observed: (1) left frontotemporal peak at ~400ms, larger toconcrete (red arrows, a typical N400m); (2) right anterior temporal, slightly later and withopposite modulation (i.e., larger to abstract words, orange arrow); (3) right parietal, both medialand lateral, as well as medial occipital, later and stronger to concrete words (blue arrows); and
Dhond et al. Page 15
Hum Brain Mapp. Author manuscript; available in PMC 2007 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

(4) a reversed modulation after ~650ms of the left frontotemporal N400m (green arrows). Thescale bars indicate a significance level of p < 10−9.
Dhond et al. Page 16
Hum Brain Mapp. Author manuscript; available in PMC 2007 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript