segmentation of short keying sequences does not spontaneously transfer to other sequences
TRANSCRIPT

Human Movement Science 28 (2009) 348–361
Contents lists available at ScienceDirect
Human Movement Science
journal homepage: www.elsevier .com/locate/humov
Segmentation of short keying sequencesdoes not spontaneously transfer to other sequences
Willem B. Verwey a,*, Elger L. Abrahamse a, Luis Jiménez b
a Cognitive Psychology and Ergonomics, Faculty of Behavioral Sciences, Universiteit Twente, Postbus 217,7500 AE Enschede, The Netherlandsb Facultad de Psicología, Universidad de Santiago de Compostela, Santiago de Compostela, Spain
a r t i c l e i n f o
Article history:Available online 8 January 2009
PsycINFO classification:23302340
Keywords:Motor skillsMovement sequencesKeying sequences
0167-9457/$ - see front matter � 2008 Elsevier B.Vdoi:10.1016/j.humov.2008.10.004
* Corresponding author. Tel.: +31 53 489 4764.E-mail address: [email protected] (W.B. V
a b s t r a c t
Previous research suggested that highly practiced discrete 6-keysequences are spontaneously segmented, sometimes even differ-ently for different persons. This suggests there is some limit inthe length of motor chunks that are assumed to underlie the seg-ments in the sequence. The present experiment examined whethera segmentation pattern induced in one 6-key sequence (the pre-structured sequence) determines segmentation in other 6-keysequences. The results are in line with segmentation, but showedneither transfer from the prestructured to a concurrently practicedunstructured sequence, nor to two new sequences that were car-ried out in a subsequent phase. Moreover, segmentation of thesetwo new sequences was mutually different. Hence, while segmen-tation seems a phenomenon affecting all 6-element keyingsequences, the exact segmentation pattern is not determined bythat of a familiar keying sequence. Another result of the presentresearch is that using different fingers of the same hand did slowexecution rate (thus indicating effector-specific sequence learn-ing), but the rate reduction was clearly smaller than in a previousstudy in which transfer to fingers of the other hand was assessed(Verwey & Wright, 2004). This is more in line with effector-specificlearning being a result of sequence learning in terms of a hand-based reference frame than learning to directly trigger particulareffectors (i.e., the fingers).
� 2008 Elsevier B.V. All rights reserved.
. All rights reserved.
erwey).

W.B. Verwey et al. / Human Movement Science 28 (2009) 348–361 349
1. Introduction
1.1. Controlling sequential action
Compared to most animals, humans excel in the ability to acquire sequential skills. We are less suc-cessful in understanding the mechanisms underlying these skills though we did develop some theo-ries about them. The classic notion is that the execution of movement sequences involves thecombining of elementary, sometimes innate, motor patterns or motor chunks (e.g., Book, 1908; Lash-ley, 1951; Miller, Galanter, & Pribram, 1960; Paillard, 1960). For example, Gallistel (1980) argued that‘‘individual movements that comprise the skill are first perfected to the point where they can be mademore rapidly and accurately with little variation. Then they become welded together into ‘chunks’ ”(p.367). Chunking is generally used to refer to a strategy to code multiple items in a relational structure,and goes beyond motor control in that the concept is used for central knowledge representations too.The benefit of chunking is that it allows circumventing limitations in human information processing asmore elaborate representations can be used (e.g., Anderson, 1996; Halford, Wilson, & Phillips, 1998).
We argue that motor chunks are the memory representations that allow familiar movement se-quences to be selected and executed as a single response (Verwey, 1999). Given the notion that move-ment control involves several hierarchically ordered levels (e.g., MacKay, 1982; Rosenbaum, Kenny, &Derr, 1983), we argue also that motor chunks are the lowest level of movement organization, straightabove the individual movements, controlling only relatively brief sequences (i.e., of up to about fiveelementary movements; Verwey, 2003a). Longer sequences would, in turn, be organized by usingmore abstract, higher level action representations (e.g., Engelkamp & Jahn, 2003; Hard, Lozano, & Tver-sky, 2006; Schack & Mechsner, 2006). According to the dual processor model, these representationsare being used by two independent, parallel processors (Verwey, 2001). The motor processor executesthe few more tightly coupled elements of a motor chunk. The cognitive processor either concatenatesthese chunks into longer sequences, or it may increase execution rate in parallel to the motor proces-sor by using explicit knowledge to prepare and executing the individual elements when there is noth-ing to concatenate (either because the sequence is short, or when the last chunk has been initiated; cf.Verwey & Eikelboom, 2003).
In the discrete sequence production (DSP) task participants usually execute two discrete, fixed ser-ies of three to six key presses, each of which is practiced for about 500 trials (see Rhodes, Bullock, Ver-wey, Averbeck, & Page, 2004, for an overview of among others the DSP task). This task is especiallysuited for studying the mechanisms underlying sequential motor skills because, as compared to se-quences requiring more complex movement elements, executing a key press takes so little time thatindividual interkey intervals (IKIs) are likely to reflect exclusively the action of the underlying controlmechanisms. Also, given the high execution rates in the DSP task, studying IKIs is likely to reveal prop-erties of the fastest mechanism at a specific level of practice because slower mechanisms (that mightbe active in parallel, like verbal ones, see Verwey, 2003b) are not likely to contribute much then.
1.2. Rhythm, segmentation, and motor chunks
It has been clear for many years that even with little practice certain properties, such as regularitiesin element order (like 12321), familiarity with certain parts, and temporal segmentation during prac-tice may cause sequences to be carried out as a few more closely integrated segments (e.g., Povel &Collard, 1982; Restle, 1970; Sakai, Hikosaka, & Nakamura, 2004; Sakai, Kitagucki, & Hikosaka, 2003;Verwey, 1996). A particularly interesting finding is that even without such regularities, longer keyingsequences appear to always include one or more relatively long response times (Kennerley, Sakai, &Rushwordth, 2004; Sakai et al., 2004; Verwey, 2003a; Verwey & Eikelboom, 2003; de Kleine & Verwey,2008). As the sequential positions of these slow responses were found to differ across participants,these sequences may have included individually different segmentation patterns. Due to the individ-ual differences, this had not been recognized before across groups of participants other than as a gen-eral effect on mean element execution time of sequence length (Verwey, 2003a; i.e., the rate effect,Sternberg, Monsell, Knoll, & Wright, 1978). Indications that sequences of over about 4-key presses

350 W.B. Verwey et al. / Human Movement Science 28 (2009) 348–361
are spontaneously segmented led to the notion that there is some neural limitation in the number ofelements represented in a single motor chunk (Verwey, Lammens, & van Honk, 2002).
Verwey (1996) argued that the segmentation pattern used at the start of practice will eventuallyalso determine chunk boundaries. This notion was based on a study by Summers (1975), whichshowed that including pauses of 500 ms during practice at particular locations appeared to later causethese locations to be marked by longer response times when the pauses were no longer imposed (alsosee Summers, Sargent, & Hawkins, 1984). Summers (1975) argued that this was caused by a temporalpattern – a rhythm – that is closely associated with the sequence at hand. However, (Verwey 1996;Verwey & Dronkert, 1996) reported indications that, rather than a developing temporal pattern, thepauses in Summers (1975) practice phase may well have determined the chunk boundaries that de-velop with practice. So, when later the pauses were removed the IKIs at these positions remained rel-atively long because they involved a transition from one to the next motor chunk, rather than arhythm that was maintained. These chunks appeared so robust that when participants had carriedout two successive 2-key segments separated by a pause during practice, they stuck to this patterneven though a single 4-key segment would have been more efficient over the transfer phase (Verwey,1996, 2001).
One research issue in the present study concerns the possibility that training a 6-key sequence witha particular temporal pattern determines the way in which participants structure sequences that aretrained either simultaneously or successively. This idea was triggered by the unexpected observationin Verwey et al. (2002) that an unstructured 6-key sequence showed the same segmentation patternas that found in a simultaneously practiced sequence that consisted of a repeated 3-key segment. Inthe present study, participants practiced two 6-key sequences. One, the prestructured sequence, im-posed a temporal structure that, for one group (the 2-2-2 group) included pauses preceding the onsetof the third and of the fifth key press, and for another group (the 3-3 group) included a single pausepreceding the fourth key press. Simultaneously, participants practiced a second, unstructured sequence,that did not involve any pauses, and that may therefore be segmented in individually different ways(because it is assumed to be too long to be carried out as a single whole). The first research questionconcerned whether the temporal structure of the prestructured sequence would emerge in the simul-taneously practiced unstructured sequence as well as in two new successively executed sequences.
1.3. Effector-specific learning
In addition to investigating the temporal determinants of chunk learning, we were also interestedin the representational status of these chunks. A slowdown in execution rate has been found when the(same) keys of the sequence were pressed with different fingers than during practice (Verwey &Wright, 2004). This has been taken as evidence for effector-specific sequence learning, but it remainsunclear what this exactly means. Despite explicit search, no evidence could be obtained for effector(i.e., finger) specific triggering in response selection tasks (Proctor & Dutta, 1993). In contrast, thereare several indications with aiming tasks (Bock & Eckmiller, 1986; Darling & Gilchrist, 1991; Gordon,Ghilardi, & Ghez, 1994; Krakauer, Pine, Ghilardi, & Ghez, 2000) and response selection tasks (Cho &Proctor, 2002; Lippa, 1996) for the development of spatial hand- and forearm-based reference framesused for coding target locations. So, one may wonder whether a slowdown similar to that found byVerwey and Wright (2004) will be observed if other fingers of the same hand are assigned to eachkey. Motor learning in terms of automatic triggering of particular fingers predicts that performancewill reduce substantially because automatic response tendencies need to be suppressed and replacedby controlled selection of other fingers. Conversely, if a spatial hand-specific reference frame underliessequencing skill, little or no performance reduction is predicted of using adjacent fingers of the samehand because the existing hand-based reference frame is still applicable.
In short, the aim of the present study was twofold. First, it aimed at determining whether the tem-poral pattern imposed in prestructured sequence influences segmentation of a simultaneously prac-ticed, unstructured sequence, and of two new sequences that are practiced subsequently. Second,this study examined whether evidence for effector-specific sequence learning is observed also whenadjacent fingers of the same hand are used to distinguish between finger-specific sequence learningand hand-based spatial learning.

W.B. Verwey et al. / Human Movement Science 28 (2009) 348–361 351
2. Method
2.1. Participants
Forty-eight (24 female and 24 male) students from the University of Twente participated in ex-change for course credits. They were aged between 18 and 26 and signed informed consent beforethe start of the experiment.
2.2. Task
The computer screen displayed black outlines of six square placeholders on a white background inthe same spatial arrangement as the assigned keys on a QWERTY keyboard, i.e., D F G J K L. Stimuliconsisted of the area enclosed by a placeholder becoming homogeneously green. Participants re-sponded by depressing with three fingers from their left and three fingers from their right hand thespatially compatible key on a computer keyboard. Each participant was trained with two 6-key se-quences, resulting in a gradual development of keying skill. Immediately after depression of the cor-rect key the content of the square turned white again and the next placeholder was filled. As only thelatencies for depression of the keys were registered, it was acceptable to release a key after the ensu-ing one had been depressed. The last key press of each sequence was followed by blanking the entirescreen for 500 ms, then the placeholders were redrawn, and 1000 ms later the first key-specific cue ofthe next sequence was presented.
The visual angle was between about 1–2 � for each placeholder (at 60 cm face-display distance).The distance between adjacent placeholders was about 0.5 �, with the exception that the distance be-tween the third and fourth placeholder was about 1 � (corresponding to the larger distance betweenleft and right hand fingers). Errors resulted in the message ‘‘wrong key” (in Dutch) for 500 ms, and thetrial only advanced after the correct key had been pressed.
For all participants, one sequence in the practice phase did not contain a pause and was called theunstructured sequence. The second sequence in the practice phase involved one or two nonzero re-sponse stimulus intervals (RSIs) in order to impose a structure. For half the participants, this prestruc-tured sequence included a relatively long RSI between response three and four (3-3 group). For theremaining participants that sequence included two RSIs, one between the second and third key, andone between the fourth and fifth key (2-2-2 group). These RSIs consisted of non aging intervals of atleast 200 ms (starting at 200 ms, 5 ms was added in successive iterations, after each of which therewas a 1% chance that this interval lengthening process was halted). Non aging intervals imply thatshorter intervals have a higher probability than longer intervals. This reduces the predictability ofthe interval duration (see Gottsdanker, Perkins, & Aftab, 1986). In the test phase, none of the se-quences included RSIs exceeding 0 ms.
2.3. Procedure
The experiment started off with an instruction on the screen after which participants performedseven 148-trial blocks (each trial including a 6-key sequence) as a practice phase. The instruction in-cluded information on which fingers to use: half of the 2-2-2 and the 3-3 groups practiced with the leftlittle, ring, and middle fingers (thus excluding the left index finger), and the right index, middle, andring fingers (excluding the right little finger). The other half practiced with the left ring, middle, andindex finger (excluding the little finger), and the right middle, ring, and little fingers (excluding theindex finger). Halfway each block there was a 7 s break during which performance was presentedin terms of mean sequence execution time and error rate. At the end of each block, participants wereinformed also of their mean sequence execution time and error rate. Between successive blocks, par-ticipants rested for 7 min. Faulty key presses in the practice and test phases were immediatelyrepeated.
Block 8 was the test block and included four 40-trial subblocks. These were separated by 7 s breaksduring which, again, mean RT and error rates were presented. These four subblocks involved manip-

352 W.B. Verwey et al. / Human Movement Science 28 (2009) 348–361
ulation of sequence familiarity and of the fingers used. This implied that two new and the two familiarsequences were carried out once with the familiar finger setting (e.g., left little, ring, and middle fin-gers and right index, middle, and ring fingers), and once with unfamiliar finger settings (e.g., the leftring, middle, and index fingers, and the right middle, ring, and little fingers). Importantly, the unfamil-iar finger setting changed the mapping between fingers and the corresponding keys, but largely main-tained the hand-based reference frame, so that the keys assigned to each hand remained the same. Thecomputer screen each time instructed participants which fingers to use. In contrast to the practicephase, none of the test block sequences included a pause.
In order to be able to compare execution rate of key presses as a function of their location in thesequences, it is imperative that finger-specific effects are not associated with sequential positions(due to some fingers being faster; Coover, 1923; Kornblum, 1965; Lahy, 1924). This was accomplishedby counterbalancing across participants fingers over sequential positions. This involved rotating keysat each sequential position (D?F?G?J?K?L?D?F?G?. . .). For example, one participant executedKFGDJL and FKLJDG, a second participant carried out LGJFKD and GLDKFJ, and a third DJKGLF andJDFLGK. This yielded a set of six versions of each sequence. Each participant practiced one sequencewith one or two pauses (which was removed during testing). Across participants, each particular se-quence was practiced equally often with (one or two) pauses as without, while the order of presenta-tion of these sequences within blocks was random. Also, the two new sequences in test Block 8 weretaken from the same set of six sequences implying that across participants each sequence was as oftena practiced and a new sequence.
Following the experiment, participants filled out a paper questionnaire. It started by askingwhether participants could reproduce in writing the two sequences they had been trained with(‘reproduction’). Then they were to select their two sequences out of a set of 18 alternatives (‘forcedchoice’). Eventually they were asked to indicate how they had reproduced the sequences in the firstand second parts of the questionnaire: (a) by remembering the order of the letters on the keys, (b)by ticking the sequence with their fingers on the table, (c) by ticking the sequence in memory, (d)by remembering the order of the stimuli on the screen, and (e) in another way.
2.4. Apparatus and setting
Stimulus presentation, timing, and data collection was achieved using the E-prime� 1.1 experi-mental software package on a standard Pentium� IV class PC. Stimuli were presented on a 17 inchPhilips 107T5 display running at 1024 by 768 pixel resolution in 32 bit color, and refreshing at85 Hz. The viewing distance was approximately 60 cm, but not strictly controlled.
2.5. Dependent measures
The serial positions of the six responses in each sequence are denoted by their index: R1 throughR6. The interval between two successive key presses is denoted interkey interval (IKI), and the positionof a specific IKI is indicated by a T with an index denoting the response it precedes. So, T2–T6 indicatethe IKIs preceding R2–R6. As cue onset coincides with depression of the previous key, T2–T5 are alsoresponse times to their corresponding cues. T1 denotes the interval between the onset of the first cueand pressing the first key.
3. Results
3.1. Practice phase
Apart from providing participants with training on their two 6-key sequences, the practice phaseallowed also examining whether practicing the prestructured sequence affects segmentation of theunstructured sequence. This was investigated with a 2 (Group: 3-3 vs. 2-2-2) � 2 (Sequence: withvs. without pause) � 2 (Block 6 & 7) � 6 (Key) ANOVA on response times with group as between-sub-ject variable.
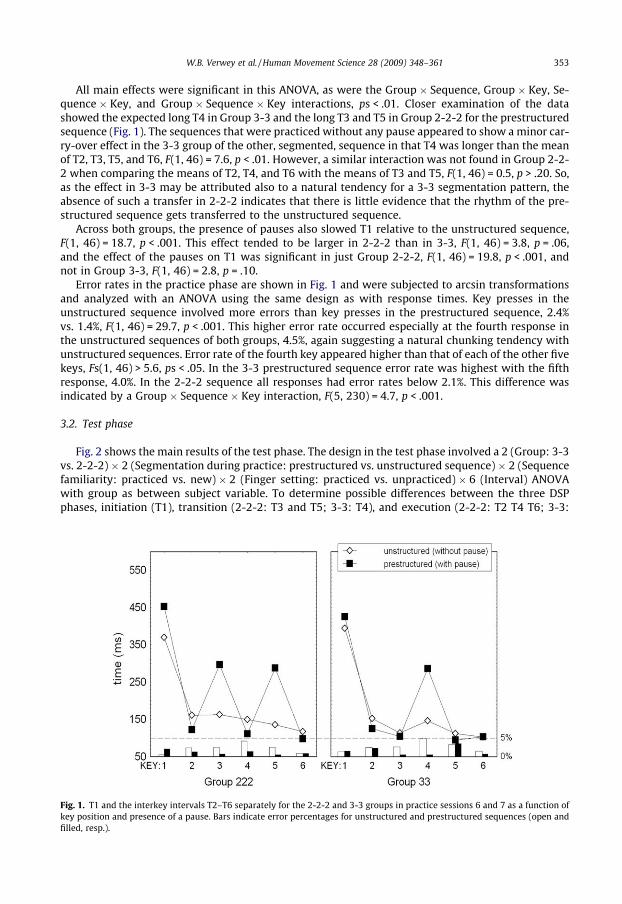
W.B. Verwey et al. / Human Movement Science 28 (2009) 348–361 353
All main effects were significant in this ANOVA, as were the Group � Sequence, Group � Key, Se-quence � Key, and Group � Sequence � Key interactions, ps < .01. Closer examination of the datashowed the expected long T4 in Group 3-3 and the long T3 and T5 in Group 2-2-2 for the prestructuredsequence (Fig. 1). The sequences that were practiced without any pause appeared to show a minor car-ry-over effect in the 3-3 group of the other, segmented, sequence in that T4 was longer than the meanof T2, T3, T5, and T6, F(1, 46) = 7.6, p < .01. However, a similar interaction was not found in Group 2-2-2 when comparing the means of T2, T4, and T6 with the means of T3 and T5, F(1, 46) = 0.5, p > .20. So,as the effect in 3-3 may be attributed also to a natural tendency for a 3-3 segmentation pattern, theabsence of such a transfer in 2-2-2 indicates that there is little evidence that the rhythm of the pre-structured sequence gets transferred to the unstructured sequence.
Across both groups, the presence of pauses also slowed T1 relative to the unstructured sequence,F(1, 46) = 18.7, p < .001. This effect tended to be larger in 2-2-2 than in 3-3, F(1, 46) = 3.8, p = .06,and the effect of the pauses on T1 was significant in just Group 2-2-2, F(1, 46) = 19.8, p < .001, andnot in Group 3-3, F(1, 46) = 2.8, p = .10.
Error rates in the practice phase are shown in Fig. 1 and were subjected to arcsin transformationsand analyzed with an ANOVA using the same design as with response times. Key presses in theunstructured sequence involved more errors than key presses in the prestructured sequence, 2.4%vs. 1.4%, F(1, 46) = 29.7, p < .001. This higher error rate occurred especially at the fourth response inthe unstructured sequences of both groups, 4.5%, again suggesting a natural chunking tendency withunstructured sequences. Error rate of the fourth key appeared higher than that of each of the other fivekeys, Fs(1, 46) > 5.6, ps < .05. In the 3-3 prestructured sequence error rate was highest with the fifthresponse, 4.0%. In the 2-2-2 sequence all responses had error rates below 2.1%. This difference wasindicated by a Group � Sequence � Key interaction, F(5, 230) = 4.7, p < .001.
3.2. Test phase
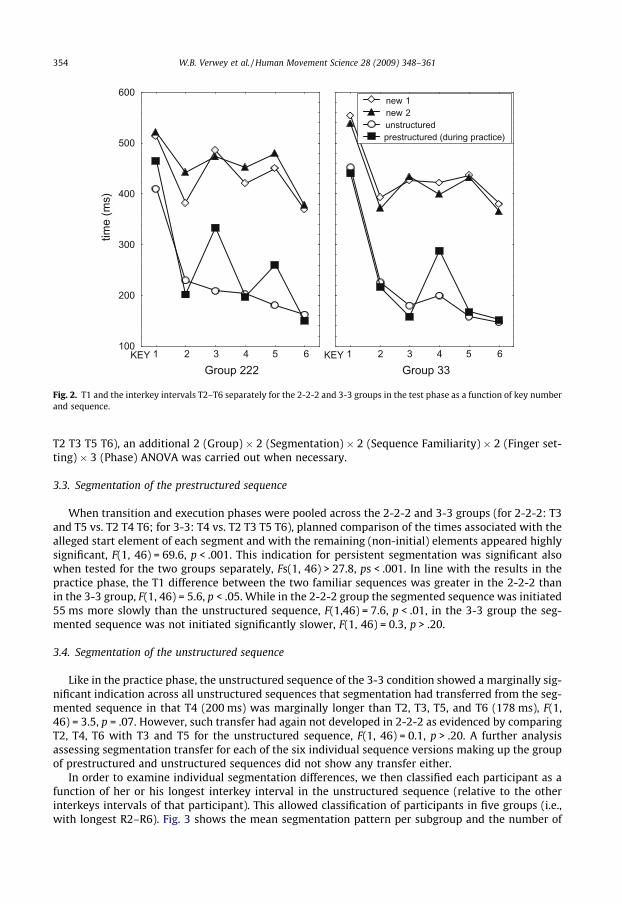
Fig. 2 shows the main results of the test phase. The design in the test phase involved a 2 (Group: 3-3vs. 2-2-2) � 2 (Segmentation during practice: prestructured vs. unstructured sequence) � 2 (Sequencefamiliarity: practiced vs. new) � 2 (Finger setting: practiced vs. unpracticed) � 6 (Interval) ANOVAwith group as between subject variable. To determine possible differences between the three DSPphases, initiation (T1), transition (2-2-2: T3 and T5; 3-3: T4), and execution (2-2-2: T2 T4 T6; 3-3:
Fig. 1. T1 and the interkey intervals T2–T6 separately for the 2-2-2 and 3-3 groups in practice sessions 6 and 7 as a function ofkey position and presence of a pause. Bars indicate error percentages for unstructured and prestructured sequences (open andfilled, resp.).
Group 222
tim
e (m
s)
100
200
300
400
500
600
KEY 1
Group 33
new 1new 2unstructuredprestructured (during practice)
2 3 4 5 6 KEY 1 2 3 4 5 6
Fig. 2. T1 and the interkey intervals T2–T6 separately for the 2-2-2 and 3-3 groups in the test phase as a function of key numberand sequence.
354 W.B. Verwey et al. / Human Movement Science 28 (2009) 348–361
T2 T3 T5 T6), an additional 2 (Group) � 2 (Segmentation) � 2 (Sequence Familiarity) � 2 (Finger set-ting) � 3 (Phase) ANOVA was carried out when necessary.
3.3. Segmentation of the prestructured sequence
When transition and execution phases were pooled across the 2-2-2 and 3-3 groups (for 2-2-2: T3and T5 vs. T2 T4 T6; for 3-3: T4 vs. T2 T3 T5 T6), planned comparison of the times associated with thealleged start element of each segment and with the remaining (non-initial) elements appeared highlysignificant, F(1, 46) = 69.6, p < .001. This indication for persistent segmentation was significant alsowhen tested for the two groups separately, Fs(1, 46) > 27.8, ps < .001. In line with the results in thepractice phase, the T1 difference between the two familiar sequences was greater in the 2-2-2 thanin the 3-3 group, F(1, 46) = 5.6, p < .05. While in the 2-2-2 group the segmented sequence was initiated55 ms more slowly than the unstructured sequence, F(1,46) = 7.6, p < .01, in the 3-3 group the seg-mented sequence was not initiated significantly slower, F(1, 46) = 0.3, p > .20.
3.4. Segmentation of the unstructured sequence
Like in the practice phase, the unstructured sequence of the 3-3 condition showed a marginally sig-nificant indication across all unstructured sequences that segmentation had transferred from the seg-mented sequence in that T4 (200 ms) was marginally longer than T2, T3, T5, and T6 (178 ms), F(1,46) = 3.5, p = .07. However, such transfer had again not developed in 2-2-2 as evidenced by comparingT2, T4, T6 with T3 and T5 for the unstructured sequence, F(1, 46) = 0.1, p > .20. A further analysisassessing segmentation transfer for each of the six individual sequence versions making up the groupof prestructured and unstructured sequences did not show any transfer either.
In order to examine individual segmentation differences, we then classified each participant as afunction of her or his longest interkey interval in the unstructured sequence (relative to the otherinterkeys intervals of that participant). This allowed classification of participants in five groups (i.e.,with longest R2–R6). Fig. 3 shows the mean segmentation pattern per subgroup and the number of
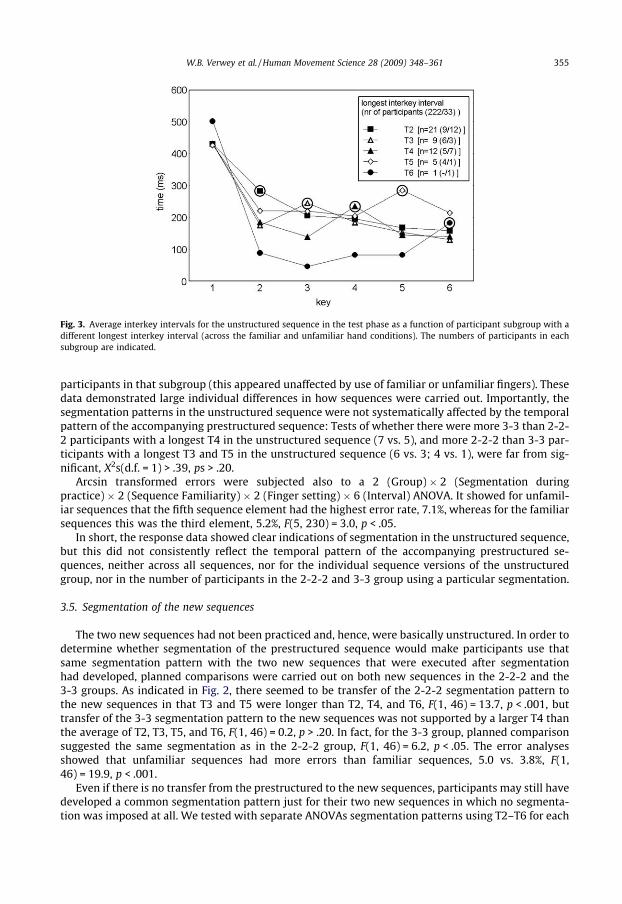
Fig. 3. Average interkey intervals for the unstructured sequence in the test phase as a function of participant subgroup with adifferent longest interkey interval (across the familiar and unfamiliar hand conditions). The numbers of participants in eachsubgroup are indicated.
W.B. Verwey et al. / Human Movement Science 28 (2009) 348–361 355
participants in that subgroup (this appeared unaffected by use of familiar or unfamiliar fingers). Thesedata demonstrated large individual differences in how sequences were carried out. Importantly, thesegmentation patterns in the unstructured sequence were not systematically affected by the temporalpattern of the accompanying prestructured sequence: Tests of whether there were more 3-3 than 2-2-2 participants with a longest T4 in the unstructured sequence (7 vs. 5), and more 2-2-2 than 3-3 par-ticipants with a longest T3 and T5 in the unstructured sequence (6 vs. 3; 4 vs. 1), were far from sig-nificant, X2s(d.f. = 1) > .39, ps > .20.
Arcsin transformed errors were subjected also to a 2 (Group) � 2 (Segmentation duringpractice) � 2 (Sequence Familiarity) � 2 (Finger setting) � 6 (Interval) ANOVA. It showed for unfamil-iar sequences that the fifth sequence element had the highest error rate, 7.1%, whereas for the familiarsequences this was the third element, 5.2%, F(5, 230) = 3.0, p < .05.
In short, the response data showed clear indications of segmentation in the unstructured sequence,but this did not consistently reflect the temporal pattern of the accompanying prestructured se-quences, neither across all sequences, nor for the individual sequence versions of the unstructuredgroup, nor in the number of participants in the 2-2-2 and 3-3 group using a particular segmentation.
3.5. Segmentation of the new sequences
The two new sequences had not been practiced and, hence, were basically unstructured. In order todetermine whether segmentation of the prestructured sequence would make participants use thatsame segmentation pattern with the two new sequences that were executed after segmentationhad developed, planned comparisons were carried out on both new sequences in the 2-2-2 and the3-3 groups. As indicated in Fig. 2, there seemed to be transfer of the 2-2-2 segmentation pattern tothe new sequences in that T3 and T5 were longer than T2, T4, and T6, F(1, 46) = 13.7, p < .001, buttransfer of the 3-3 segmentation pattern to the new sequences was not supported by a larger T4 thanthe average of T2, T3, T5, and T6, F(1, 46) = 0.2, p > .20. In fact, for the 3-3 group, planned comparisonsuggested the same segmentation as in the 2-2-2 group, F(1, 46) = 6.2, p < .05. The error analysesshowed that unfamiliar sequences had more errors than familiar sequences, 5.0 vs. 3.8%, F(1,46) = 19.9, p < .001.
Even if there is no transfer from the prestructured to the new sequences, participants may still havedeveloped a common segmentation pattern just for their two new sequences in which no segmenta-tion was imposed at all. We tested with separate ANOVAs segmentation patterns using T2–T6 for each
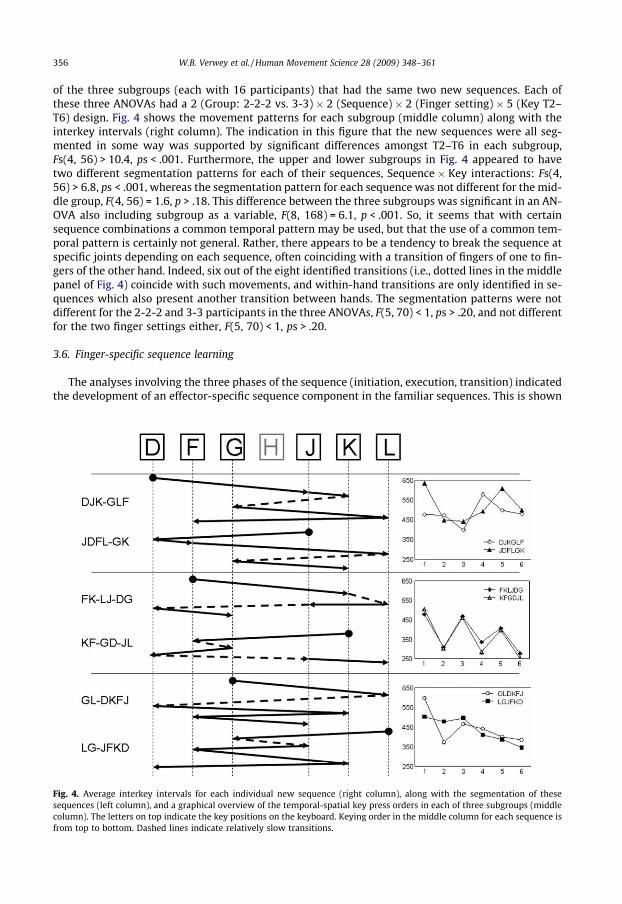
356 W.B. Verwey et al. / Human Movement Science 28 (2009) 348–361
of the three subgroups (each with 16 participants) that had the same two new sequences. Each ofthese three ANOVAs had a 2 (Group: 2-2-2 vs. 3-3) � 2 (Sequence) � 2 (Finger setting) � 5 (Key T2–T6) design. Fig. 4 shows the movement patterns for each subgroup (middle column) along with theinterkey intervals (right column). The indication in this figure that the new sequences were all seg-mented in some way was supported by significant differences amongst T2–T6 in each subgroup,Fs(4, 56) > 10.4, ps < .001. Furthermore, the upper and lower subgroups in Fig. 4 appeared to havetwo different segmentation patterns for each of their sequences, Sequence � Key interactions: Fs(4,56) > 6.8, ps < .001, whereas the segmentation pattern for each sequence was not different for the mid-dle group, F(4, 56) = 1.6, p > .18. This difference between the three subgroups was significant in an AN-OVA also including subgroup as a variable, F(8, 168) = 6.1, p < .001. So, it seems that with certainsequence combinations a common temporal pattern may be used, but that the use of a common tem-poral pattern is certainly not general. Rather, there appears to be a tendency to break the sequence atspecific joints depending on each sequence, often coinciding with a transition of fingers of one to fin-gers of the other hand. Indeed, six out of the eight identified transitions (i.e., dotted lines in the middlepanel of Fig. 4) coincide with such movements, and within-hand transitions are only identified in se-quences which also present another transition between hands. The segmentation patterns were notdifferent for the 2-2-2 and 3-3 participants in the three ANOVAs, F(5, 70) < 1, ps > .20, and not differentfor the two finger settings either, F(5, 70) < 1, ps > .20.
3.6. Finger-specific sequence learning
The analyses involving the three phases of the sequence (initiation, execution, transition) indicatedthe development of an effector-specific sequence component in the familiar sequences. This is shown
Fig. 4. Average interkey intervals for each individual new sequence (right column), along with the segmentation of thesesequences (left column), and a graphical overview of the temporal-spatial key press orders in each of three subgroups (middlecolumn). The letters on top indicate the key positions on the keyboard. Keying order in the middle column for each sequence isfrom top to bottom. Dashed lines indicate relatively slow transitions.

W.B. Verwey et al. / Human Movement Science 28 (2009) 348–361 357
by the Finger Setting � Sequence Familiarity interaction, F(1, 46) = 5.9, p < .05. Planned comparisonsconfirmed that with the familiar sequences the unfamiliar finger setting yielded 18 ms slower re-sponses, F(1, 46) = 4.44, p < .05, whereas with the new sequences finger setting did not significantlyaffect response times, �8 ms, F(1, 46) = 1.4, p > .20.
A subsequent planned comparison, examining the three DSP phases separately, confirmed that theunfamiliar finger settings did not affect any of the phases of the new sequences, while it did affect thephases in the familiar sequences differently, F(1, 46) = 4.1, p < .05. A further planned comparison con-firmed for the familiar sequences that transition and execution intervals suffered more from using theunfamiliar finger settings (22 ms) than initiation (�2 ms), F(1, 46) = 4.4, p < .05.
When examining just the prestructured sequences, the use of other fingers than during practiceslowed execution by 23 ms, F(1, 46) = 5.7, p < .05, but not the transition intervals T3 and T5 in 2-2-2 and T4 in 3-3 (0 ms difference), F(1, 46) = 0.0, p > .20. This distinction between transition and execu-tion intervals was significant, F(1, 46) = 5.0, p < .05. So, changing fingers in prestructured sequencesslowed execution but not initiation and transition intervals.
This was different with unstructured sequences. On the assumption that the longest interkey inter-val (from T2–T6) in the unstructured sequence comprised a transition, we compared this longestinterval with the mean of the remaining intervals using a 2 (Group) � 2 (Peak vs. other intervals) � 2(Finger Setting) ANOVA. This did not show that effector-specific learning was less for the longest inter-val (24 ms) than for the remaining intervals (19 ms), F(1, 46) = 1, p > .20. Instead, the effector-specificeffect gradually increased with position (T1–T6: 0, 15, 20, 31, 44, 31 ms, resp.), F(5, 180) = 2.3, p < .05.So, in contrast to the prestructured sequences, the slowing observed when other fingers were usedthan during practice was not different for the longest and the remaining intervals of the unstructuredsequences, but increased with sequential position.
The error analysis showed a Sequence Familiarity � Finger Setting interaction indicating that famil-iar sequences carried out with the familiar finger setting involved less errors than when the unfamiliarsequence, the unfamiliar finger setting, or both were involved (3.2% vs. 4.4% and higher, F(1, 46) = 8.8,p < .01). Examination of the error rates at the various keys showed no clear patterns. Familiar unstruc-tured sequences carried out with the practiced finger settings still showed their highest error rates atthe fourth key (suggesting spontaneous segmentation), but this was no longer significantly higherthan the rest.
In short, using other fingers than during practice slowed execution of familiar 2-2-2 and 3-3 (pre-structured and unstructured) sequences, but this did not affect their initiation. In prestructured se-quences the finger change did not slow the relatively slow transition intervals either. Inunstructured sequences the effect of the finger change increased with position, while there was no dif-ference between the effect produced in the longest interkey interval and the effect produced in theremaining intervals. The new sequences never showed an effect of using unpracticed fingers, thus sup-porting the conclusion that the effector-specific effects were sequence specific, and not due to the factthat some fingers had become more skilled.
3.7. Awareness
The questionnaire results showed that the prestructured sequence was reproduced correctly by 42(88%) participants while the unstructured sequence was reproduced correctly by only 28 participants(58%), X2(d.f. = 1) = 10.3, p < .01. In the recognition phase, 44 (92%) participants recognized their pre-structured, and 45 (94%) their unstructured sequence. Thirty-six participants indicated that they hadreproduced the sequences in writing by ticking the sequence (either on the table or in memory; ofthese 36 participants, 10 indicated to have used just ticking on the table, and 2 used just ticking inmemory). Twenty-five participants indicated to have used remembering of stimulus order on thescreen as one of the reproduction cues (for 5 it was the only strategy). The amount of explicit sequenceknowledge (reproduction and recognition performance) in familiar sequences appeared not to be sig-nificantly correlated to execution rate, rs < �.40. Together these findings suggest that the sequenceswere represented primarily in a code that is tailored for executing these keying sequences, and thatthe need to write down (or recognize) a sequence involved strategies to derive explicit from implicit

358 W.B. Verwey et al. / Human Movement Science 28 (2009) 348–361
sequence knowledge (cf. Heuer & Schmidtke, 1996). With the unstructured sequence this strategyproved harder than with prestructured sequences.
4. Discussion
4.1. No transfer of segmentation
Data confirmed that inclusion of pauses during practice in a structured sequence results in a con-tinuation of that segmentation pattern after pause removal (cf. Verwey, 1996). The present studyshowed that this segmentation pattern is sequence specific in that there was no clear transfer to othersequences in various tests: First, the transfer of the segmentation pattern from the prestructured tothe unstructured sequence, observed in the 3-3 group, was not found in the 2-2-2 group. This indicatesthat the apparent transfer in 3-3, found in the practice and test phases, was probably due to a naturalsegmentation tendency and is independent of the accompanying prestructured sequence. A naturalsegmentation tendency in 3-key chunks was corroborated by the error data of the unstructured se-quences in both groups. So, the impetus for the present study, namely the observation that an unstruc-tured 6-key sequence showed the same segmentation pattern as in the simultaneously practiced 2 � 3sequence (Verwey et al., 2002), seemed to have been caused by a tendency to segment 6-key se-quences in threes, rather than that the 2 � 3 segmentation pattern in that study transferred to the1 � 6 sequence.
Second, the unstructured sequence in the test phase showed some systematic differences between2-2-2 and 3-3 groups that could have indicated transfer from the prestructured sequence, but theassociated segmentation patterns appeared to differ across the various versions of the unstructuredsequence, and did not reflect the accompanying 2-2-2 or 3-3 segmentation patterns either.
Third, classification of participants as a function of the position of longest interkey interval inunstructured sequences showed large individual differences as to how the unstructured sequenceswere segmented in the test phase (Fig. 3). Importantly, the groups with a long T3 or T5 did not includesignificantly more 2-2-2 participants, and the group with a longest T4 did not include significantlymore 3-3 participants. Inspection of segmentation for each particular sequence version did not pro-vide any indication that segmentation had been influenced by the use of specific fingers in a particularsequential position (e.g., because the little finger was slower).
Fourth, in new sequences a 2-2-2 segmentation pattern seemed dominant across both 2-2-2 and 3-3groups (Fig. 2), again refuting transfer from the prestructured sequences. Separate analyses on each ofthree subgroups carrying out a particular set of new sequences in the test phase showed that segmen-tation of new sequences was not necessarily the same within the two new sequences either. One group(middle group in Fig. 4) had an almost identical 2-2-2 segmentation pattern for both new sequences(even though half the participants had practiced in the 3-3 group). For another group (lower groupin Fig. 4) the interkey interval patterns were quite different though they may still reflect a similar seg-mentation pattern for the two new sequences. However, the upper group in Fig. 4 showed segmentationpatterns that were clearly different for both new sequences. Notice that the results in Fig. 4 indicate alsothat the relatively slow interkey intervals were not caused so much by a slow finger (e.g., the little fin-ger), but rather by a combination of segment length, a switch between hands, and perhaps position inthe sequence of the index finger. As explained below, the finding that the first key press following aninter-hand transition was relatively often quite slow, is in line with hand-based sequence coding.
In short, while confirming that unstructured sequences are spontaneously segmented (Verwey,2003a; Verwey & Eikelboom, 2003), the data did not show indications that the spontaneously devel-oped segmentation patterns in the unstructured and the new sequences are influenced by the imposedsegmentation of the accompanying prestructured sequence. Likewise, the new sequences do not nec-essarily involve the same segmentation pattern.
4.2. Transfer to new finger settings
The data also demonstrated an effector-specific sequence learning component in that using otherfingers of the same hand caused execution rates to reduce. Nevertheless, the size of this component

W.B. Verwey et al. / Human Movement Science 28 (2009) 348–361 359
was quite small (18 ms) compared to the (65 ms) effect size found with transfer between fingers ofone and two hands, and vice versa (Verwey & Wright, 2004). This small rate reduction seems hardto reconcile with the notion in the introduction that changing fingers implies that automatic triggeringof individual fingers is suppressed and replaced by controlled triggering of other fingers. The relativelysmall performance reduction when other fingers of the same hands are used is more in line with thenotion that effector-specific learning is based on the development and use of a spatial component witha hand-based reference frame. At least for the serial RT task, recent results confirm that implicit aswell as explicit sequence learning is based on spatial learning (Liu, Lungu, Waechter, Willingham, &Ashe, 2007). However, that study did not involve the extensive practice of the present experiment,and can therefore not be taken as immediate evidence that spatial learning is dominant with extensivepractice too as practice may well involve development of another type of coding (Hikosaka et al.,1999). Still, the present results are in line with the notion that effector-specific sequence learning isspatial. In terms of the dual processor model, these spatial components might all be a function ofthe motor processor. Hand-based sequence learning can explain also why in the present data seg-ments often seem to start following an inter-hand transition in that another reference frame needsto be activated after such a transition (Fig. 4). Obviously, the notion that effector-specific sequencelearning is actually based on a spatial, hand-based frame of reference has to be tested more directly.
4.3. Phases of sequence execution
Though not directly anticipated, the present data confirmed the theoretical distinction betweenexecution of a motor chunk and the transition from one to the next motor chunk within the prestruc-tured sequence as postulated by the hierarchically operating dual processor model (Verwey, 2001).That is, using adjacent fingers lengthened intervals that were within the predefined segments (i.e., in-tra-chunk intervals), but not those that started a segment (i.e., inter-chunk intervals). This extends re-cent findings supporting the distinction between execution of chunks and transition between chunksin that chunk execution was slowed when the hand position was changed relative to the body whileinitiation and transition phases were not (de Kleine & Verwey, 2008). This endorses the distinction be-tween a cognitive processor responsible for concatenating motor chunks, and a motor processor car-rying out motor chunks that represent individual key presses in terms of various spatial referenceframes.
The observation in the unstructured sequences that the effector-specific effect seemed to increasewith sequential position may be explained by the dual processor model too (Verwey, 2001). As thebuffer used to prepare short keying sequences (sometimes denoted motor buffer) is assumed to havea limited capacity (Rosenbaum, 1990; Sternberg et al., 1978), one could argue that the increasingeffector-dependence with position in the unstructured sequence can be attributed to a simultaneouscontribution of the (effector-independent) cognitive processor to the (effector-dependent) motor pro-cessor solely at the start of the sequence (Verwey, 2003b).
5. Conclusions
The main findings are (a) that the temporal pattern found with keying sequences is sequence-dependent and does not influence other sequences, and (b) that execution slows when other fingersof the same hand are being used. This slowing seems smaller than when fingers of the other handare used (as in Verwey & Wright, 2004), which can be explained by the notion that effector-specificsequence learning involves the development of a hand-based reference frame. Additional results are(c) a confirmation that different processes are involved in concatenating and executing motor chunks,and that (d) spontaneous segmentation is highly variable within and between participants. (e) The dif-ficulty some participants have in explicitly reproducing discrete sequences (here and in earlier stud-ies) confirms that even though participants are aware they are producing two fixed 6-key sequencestheir motor chunks are not directly accessible to processes used to reproduce the sequences in anothermode (like verbal expression), and that in order to verbally reproduce the sequence they may revert toplaying back their motor chunks.

360 W.B. Verwey et al. / Human Movement Science 28 (2009) 348–361
Acknowledgment
We would like to thank Malou Bolscher, Janine den Hertog, Matthias Kampermann, Milou Kievik,Malto Risto, Judith Wagner, Mike Walpuski, and Noshik Zeko for running the experiment. This re-search was supported in part by a visiting researcher grant to the first author from the Max PlanckInstitute for Human Cognitive and Brain Sciences in Leipzig, Germany, and by Grant No. 461-04-620 from the Netherlands Organization for Scientific Research NWO. Jiménez was supported by grantsSEJ2005 25754-E and SEJ2006 27564-E from the Spanish Ministerio de Educación y Ciencia.
References
Anderson, J. R. (1996). A simple theory of complex cognition. American Psychologist, 51, 355–365.Bock, O., & Eckmiller, R. (1986). Goal-directed arm movements in sequential pointing tasks. Experimental Brain Research, 62,
451–458.Book, W. F. (1908). The psychology of skill. Missoula: Montana Press.Cho, Y. S., & Proctor, R. W. (2002). Influences of hand posture and hand position on compatibility effects for up–down stimuli
mapped to left–right responses: Evidence for a hand referent hypothesis. Perception and Psychophysics, 64, 1301–1315.Coover, J. E. (1923). A method of teaching typewriting based on a psychological analysis of expert typing. National Education
Association Addresses and Proceedings, 61, 561–567.Darling, W. G., & Gilchrist, L. (1991). Is there a preferred coordinate system for perception of hand orientation in three-
dimensional space? Experimental Brain Research, 85, 405–416.Engelkamp, J., & Jahn, P. (2003). Lexical, conceptual and motor information in memory for action phrases: A multi-system
account. Acta Psychologica, 113, 147–165.Gallistel, C. R. (1980). The organization of action: A new synthesis. Hillsdale, NJ: Erlbaum.Gordon, J., Ghilardi, M. F., & Ghez, C. (1994). Accuracy of planar reaching movements. I. Independence of direct and extent
variability. Experimental Brain Research, 99, 97–111.Gottsdanker, R., Perkins, T., & Aftab, J. (1986). Studying reaction time with nonaging intervals: An effective procedure. Behavior
Research Methods, Instruments, and Computers, 18, 287–292.Halford, G. S., Wilson, W. H., & Phillips, S. (1998). Processing capacity defined by relational complexity: Implications for
comparative, developmental, and cognitive psychology. Behavioral and Brain Sciences, 21, 803–864.Hard, B. M., Lozano, S. C., & Tversky, B. (2006). Hierarchical encoding of behavior: Translating perception into action. Journal of
Experimental Psychology: General, 135, 588–608.Heuer, H., & Schmidtke, V. (1996). Secondary-task effects on sequence learning. Psychological Research, 59, 119–133.Hikosaka, O., Nakahara, H., Rand, M. K., Sakai, K., Lu, X., Nakamura, K., et al. (1999). Parallel neural networks for learning
sequential procedures. Trends in Neuroscience, 22, 465–471.Kennerley, S. W., Sakai, K., & Rushwordth, M. F. S. (2004). Organization of action sequences and the role of pre-SMA. Journal of
Neurophysiology, 91, 978–993.de Kleine, E., & Verwey, W. B. (2008). Representations underlying skill in the discrete sequence production task: Effect of
hand-based and hand position on the discrete sequence production task. Psychological Research, doi:10.1007/s00426-008-0174-2.
Kornblum, S. (1965). Response competition and/or inhibition in two-choice reaction time. Psychonomic Science, 2, 55–56.Krakauer, J. W., Pine, Z. M., Ghilardi, M.-F., & Ghez, C. (2000). Learnng of visuomotor transformations for vectorial planning of
trajectories. Journal of Neuroscience, 20, 8916–8924.Lahy, J. M. (1924). Motion study in typewriting. In Studies and reports: Series J (Educational), 3. Geneva: International Labour
Office.Lashley, K. S. (1951). The problem of serial order in behavior. In L. A. Jeffress (Ed.), Cerebral mechanisms in behavior
(pp. 112–136). New York: Wiley.Lippa, Y. (1996). A referential coding explanation for compatibility effects of physically orthogonal stimulus and response
dimensions. Quarterly Journal of Experimental Psychology, 49, 950–971.Liu, T., Lungu, O. V., Waechter, T., Willingham, D. T., & Ashe, J. (2007). Frames of reference during implicit and explicit learning.
Experimental Brain Research, 180, 273–280.MacKay, D. G. (1982). The problems of flexibility, fluency, and speed-accuracy trade-off in skilled behavior. Psychological Review,
89, 483–506.Miller, G. A., Galanter, E., & Pribram, K. H. (1960). Plans and the structure of behavior. New York: Holt.Paillard, J. (1960). Neurophysiology. In H. W. Magoun (Ed.), Handbook of physiology. A critical comprehensive presentation of
physiological knowledge and concepts (Section 1, Neurophysiology, Vol. III, Chapter 67, The pattern of skilled movements(pp. 1679–1708). Washington, DC: American Physiological Society.
Povel, D. J., & Collard, R. (1982). Structural factors in patterned finger tapping. Acta Psychologica, 52, 107–123.Proctor, R. W., & Dutta, A. (1993). Do the same stimulus-response relations influence choice reactions initially and after
practice? Journal of Experimental Psychology: Learning, Memory and Cognition, 19, 922–930.Restle, F. (1970). Theory of serial pattern learning: Structural trees. Psychological Review, 77, 481–495.Rhodes, B. J., Bullock, D., Verwey, W. B., Averbeck, B. B., & Page, M. P. A. (2004). Learning and production of movement
sequences: Behavioral, neurophysiological, and modelling perspectives. Human Movement Science, 23, 699–746.Rosenbaum, D. A. (1990). On choosing between movement sequences: Comments on Rose (1988). Journal of Experimental
Psychology: Human Perception and Performance, 16, 439–444.Rosenbaum, D. A., Kenny, S. B., & Derr, M. A. (1983). Hierarchical control of rapid movement sequences. Journal of Experimental
Psychology: Human Perception and Performance, 9, 86–102.

W.B. Verwey et al. / Human Movement Science 28 (2009) 348–361 361
Sakai, K., Hikosaka, O., & Nakamura, K. (2004). Emergence of rhythm during motor learning. Trends in Cognitive Sciences, 8,547–553.
Sakai, K., Kitagucki, K., & Hikosaka, O. (2003). Chunking during human visuomotor sequence learning. Experimental BrainResearch, 152, 229–242.
Schack, T., & Mechsner, F. (2006). Representation of motor skills in human long-term memory. Neuroscience Letters, 391, 77–81.Sternberg, S., Monsell, S., Knoll, R. L., & Wright, C. E. (1978). The latency and duration of rapid movement sequences:
Comparisons of speech and typewriting. In G. E. Stelmach (Ed.), Information processing in motor control and learning(pp. 117–152). New York: Academic Press.
Summers, J. J., Sargent, G. I., & Hawkins, S. R. (1984). Rhythm and the timing of movement sequences. Psychological Research, 46,107–119.
Summers, J. J. (1975). The role of timing in motor program representation. Journal of Motor Behavior, 7, 229–241.Verwey, W. B. (1996). Buffer loading and chunking in sequential keypressing. Journal of Experimental Psychology: Human
Perception and Performance, 22, 544–562.Verwey, W. B., & Dronkert, Y. (1996). Practicing a structured continuous keypressing task: Motor chunking or rhythm
consolidation? Journal of Motor Behavior, 28, 71–79.Verwey, W. B. (1999). Evidence for a multi-stage model of practice in a sequential movement task. Journal of Experimental
Psychology: Human Perception and Performance, 25, 1693–1708.Verwey, W. B. (2001). Concatenating familiar movement sequences: The versatile cognitive processor. Acta Psychologica, 106,
69–95.Verwey, W. B., Lammens, R., & van Honk, J. (2002). On the role of the SMA in the discrete sequence production task. A TMS study.
Neuropsychologia, 40, 1268–1276.Verwey, W. B. (2003a). Effect of sequence length on executing familiar keying sequences: Lasting segmentation and
preparation? Journal of Motor Behavior, 35, 343–354.Verwey, W. B. (2003b). Processing modes and parallel processors in producing familiar keying sequences. Psychological Research,
67, 106–122.Verwey, W. B., & Eikelboom, T. (2003). Evidence for lasting sequence segmentation in the discrete sequence-production task.
Journal of Motor Behavior, 35, 171–181.Verwey, W. B., & Wright, D. (2004). Effector-independent and effector-dependent learning in the discrete sequence production
task. Psychological Research, 68, 64–70.