reconstitution and pigment-binding properties of recombinant cp29
TRANSCRIPT

Eur. J. Biochem. 238, 112-120 (1996) 0 FEBS 1996
Reconstitution and pigment-binding properties of recombinant CP29 Elisabetta GIUFFRA, Daniela CUGINI, Roberta CROCE and Roberto BASSI
Universiti di Verona, Facolth di Scienze MM. FF. NN., Biotecnologie Vegetali, Verona, Italia
(Received 28 December 1995/26 February 1996) - EJB 95 2126/6
The minor light-harvesting chlorophyll-alb-binding protein CP29 (Lhcb4), overexpressed in Esche- richia coli, has been reconstituted in vitro with pigments. The recombinant pigment-protein complexes show biochemical and spectral properties identical to the native CP29 purified from maize thylakoids. The xanthophyll lutein is the only carotenoid necessary for reconstitution, a finding consistent with the structural role of two lutein moleculeslpolypeptide suggested by the crystallographic data for the homolo- gous protein light-harvesting chlorophyll-alb-binding protein of photosystem I1 (LHCII).
The CP29 protein scaffold can accommodate different chromophores. This conclusion was deduced by the observation that the pigment composition of the reconstituted protein depends on the pigments present in the reconstitution mixture. Thus, in addition to a recombinant CP29 identical to the native one, two additional forms of the complex could be obtained by increasing chlorophyll b content. This finding is typical of CP29 because the major LHCII complex shows an absolute selectivity for chromophore binding [Plumley, F. G. & Schmidt, G. W. (1987) Pruc. Natl Acad. Sci. USA 84, 146-150; Paulsen, H., Riimler, U. & Rudiger, W. (1990) Plunta (Heidelb.) 181, 204-2111, and it is consistent with the higher stability of CP29 during greening and in chlorophyll b mutants compared with LHCII.
Keywords: photosynthesis ; Lhcb4; xanthophyll proteins.
In the chloroplasts of higher plants, chlorophyll and carot- enoid molecules are non-covalently bound to specific transmem- brane proteins. These represent antenna complexes and are called light-harvesting chlorophyll-ah-binding proteins of pho- tosystem I and I1 (LHCI and LHCII, respectively). Light is har- vested by these two antenna complexes and excitation energy is delivered to the reaction centres of photosystem I (PSI) and photosystem I1 (PSII), where transmembrane electron transport occurs, generating a trans-thylakoid pH gradient, ATP synthesis and NADP' reduction.
Photosystem I1 light harvesting complex (LHC) is composed of four chlorophyll-alb-binding proteins : the major LHCII, bind- ing approximately 65 % of PSII chlorophyll, and the three minor complexes called CP24, CP26 and CP29 which, all together, bind about 15% of total PSII chlorophyll (Dainese and Bassi, 1991; Peter and Thornber, 1991 ; Jansson et al., 1992). Due to the low amount of bound pigments, it seems unlikely that the main function of the minor complexes is to harvest light. On the contrary, several lines of evidence indicate that they are involved in the regulation of the level of chlorophyll a excited states. Such regulation is required to prevent overexcitation and photo- inhibition of PS 11. At least 80% of the xanthophyll violaxanthin is located in minor complexes in maize (Bassi et al., 1993). This pigment is involved in the major photoprotection mechanism in plants, known as non-photochemical quenching (Demmig-
Correspondence to R. Bassi, Universiti di Verona, Facolti di Scienze MM. FF. NN., Strada Le Grazie, 1-37134 Verona, Italia
Abbreviations. (cHxN),C, dicychlohexylcarbodiimide; Deriphat, lauryl b-D-immhopropionidate ; LiDS, lithium dodecyl sulfate ; LHCII, light-harvesting chlorophyll-ah-binding protein of PSII; PS, photosys- tern; rCP29, recombinant CP29 reconstituted from the apoprotein deriva- tives overproduced in bacteria.
Dedication. Work dedicated to Prof. Francesco Ghiretti on the occa- sion of his 80th birthday.
Adams, 1992). Non-photochemical quenching operates through a xanthophyll cycle, that includes the deepoxidation of violaxan- thin to antheraxanthin and zeaxanthin (Bassi and Yamamoto, 1995). The involvement of minor complexes in regulation mech- anisms is also supported by the binding of the NPQ inhibitor dicychlohexylcarbodiimide [(cHxN),C] to CP26 and CP29 (Walters et al., 1994) and by the finding that photoinhibitory conditions lead to a conformational change of CP29 caused by phosphorylation. The phosphorylation takes place in cold-resis- tant, but not in cold-sensitive, maize plants (Bergantino et al., 1995). Finally, the location of the minor complexes between the reaction centre and the major LHCII complex (Bassi and Dainese, 1991) is well placed for regulating excitation-energy supply to, or diversion from, PSII.
The major antenna complex LHCII can be easily purified in bulk amounts, has been deeply studied by spectroscopic meth- ods, and its structure has been obtained from two-dimensional crystals (Kuhlbrandt et al., 1994). Conversely, the structural ba- sis for the regulative function of minor chlorophyll-proteins are mostly unknown due to the difficulties in the isolation of these proteins in sufficient amounts and in their native form. These problems have prevented not only their crystallisation but also a complete spectroscopic analysis.
To overcome this problem, we have attempted the reconstitu- tion of the CP29 protein from the apoprotein overexpressed in Escherichia coli. Here we report on the production of recombi- nant CP29 protein reconstituted with pigments, on its pigment- binding properties and on the procedure required in order to obtain a complex suitable for spectroscopic analysis.
MATERIALS AND METHODS Construction of a CP29(Lhcb4)-expression plasmid. To
overexpress plant CP29 in E. coli, the maize Lhcb4 cDNA (Ber-

Giuffra et al. ( E M J . Biochem. 238) 113
Fig.2. N-terminal sequences of the native CP29 protein and of the two recombinant proteins used in this study as deduced from DNA sequencing. The first residue of the mature protein is labelled with 1 , residues labelled with negative numbers either derive from the transit peptide or are introduced by the construction strategy.
gantino et al., 1995) was subcloned into an expression vector of the pDS series (Bujard et al., 1987). Two clones (pQBH11, pQBH22), differing in the number of amino acids of the transit peptide at the N-terminus, were obtained by PCR mutagenesis of Lhcb4 cDNA. The construct pQBH22 (Fig. 1) was obtained by using, for PCR mutagenesis, two primers [5' CATCGG- GATCCGGTTCGGGTT 3' (carrying the BamHI site), 5' GGGTTGATGCATCCCCAGGCT 3' (carrying the NsiI site)], allowing the amplification of a 900-bp fragment spanning the full coding region. After digestion with BarnHI and NsiI, the resulting fragment was subcloned into the pQE52 Qiagen ex- pression vector (pDS series; Bujard et al., 1987). Thus, the pQBH22 construct codes for a protein containing one additional Ile (which substitutes the first Ala of the transit peptide) and two amino acids contributed by the vector DNA sequence. The pQBHl1 construct allowed the expression of a protein contain- ing an NH,-terminal addition of 13 amino acids (11 amino acids of the transit peptide plus two amino acids of the vector se- quence (Fig. 2). The constructs were checked by DNA sequenc- ing.
Isolation of overexpressed CP29 apoprotein from bacte- ria. CP29 was isolated from the SGI 3009 strain transformed with one of the two CP29-constructs by means of published pro- tocols (Nagai and Thorgersen, 1987; Paulsen et al., 1990).
Pigments. Total pigments were extracted from thylakoids of wild-type maize with 80% acetone, while chlorophyll a and ca- rotenoids were obtained from thylakoids of the chlorophyll-b- less mutant chlorina f2 (Simpson, 1979). Chlorophyll b and ca- rotenoids were purified by preparative HPLC using a reverse- phase C,, column bondclone (7.3 mmX300 mm) according to Gilmore and Yamainoto (1991). Reconstituted complexes were analysed for their pigment composition after 80% acetone ex- traction as previously described (Bassi et al., 1993). During all the procedures, care was taken to protect pigments from light and contact with oxygen. Concentrations of pigments were de- termined spectroscopically :determination of chlorophylls was performed as described by Porra et al. (1 989), whilst the absorp- tion coefficients given by Davies (1965) were used for xantho- phyll determinations. The concentration of carotenoid mixtures
was estimated on the basis of an average percentage absorption coefficient of 2500 at 444 nm (Davies, 1965).
Reconstitution of CP29-pigment complexes. The reconsti- tution procedure mimicked that described for LHCII (Plumley and Schmidt 1987; Paulsen et al., 1990). The reconstitution buffer contained 100 niM Tris/HCl, pH 9.0, the protease inhibi- tors 6-aminocaproic acid ( 5 mM) and benzamidine (1 mM), 12.5% sucrose and 2% lithium dodecylsulfate (LiDS). After ad- dition of 10 mM dithioerythritol, the pigments in ethanol solu- tion (7-10% final vol.) were added with vortex mixing, fol- lowed by bath sonication for 5 min. The CP29 apoprotein iso- lated from bacteria (350 pg/ml, previously stored in the reconsti- tution buffer) was heated to 100°C for 1 min and added to the mixture, which was again sonicated for 5 min. In the standard experiments, 0.4 mg/ml chlorophyll of total pigment extracted from thylakoid membranes and 0.35 mg/ml of total protein from inclusion bodies (Fig. 3A) were used. The carotenoid concentra- tion was kept at 0.15 mg/ml in all cases, and the chlorophyll a/ chlorophyll b ratio was adjusted to 1 .O, 3.0 or 8.0. Reconstitution was achieved by three subsequent cycles of freezing (1 h, -20°C) and thawing (15 min, 25°C). 1 % n-octyl p-D-ghCOpy- ranoside was then substituted for LiDS by precipitation of the potassium dodecylsulfate following addition of KC1 150 mM, incubation for 15 min on ice and centrifugation (10 min at 15000Xg). The mixture was then loaded on a 12-ml sucrose gradient (0.1 M to 1 M), containing 10 mM Hepes/KOH, pH 7.6 and 0.06 % dodecyl p-D-maltoside, and centrifuged overnight at 254000Xg in a Beckman SW41 rotor. The lower green band (at about 0.4 M sucrose) contained the reconstituted complex and was harvested with a syringe.
Removal of excess pigments from the reconstituted com- plex. Two alternative procedures were used with essentially the same results. The band from the sucrose gradient was run through a lauryl p-D-imminopropionidate (Deriphat)/PAGE (7 % acrylamide). After overnight migration the upper green band was excised and ground in a Potter homogeniser with 0.025% dodecyl P-D-maltoside, 10 mM Hepes/KOH, pH 7.6. After 1 h elution at 4 "C, the acrylamide was eliminated by filtration through a 20-pm nylon mesh and the complex loaded onto a glycerol gradient (15 % to 40 %) containing 0.06 % dodecyl p-D- maltoside and 10 mM Hepes-KOH, pH 7.6 The gradient was spun overnight in a SW 60 Beckman rotor at 450000 g yielding a faint upper band of free pigments and a lower band with the chlorophyll-protein complex which was frozen in liquid nitrogen and kept at -80°C until use. The DeriphatPAGE step could be substituted for by chromatography into a Fractogel EMD-DEAE 650 (s) column (10 mmXl50 mm) (Merck). The sucrose gradi- ent band was loaded onto the column, which was washed with 0.025 % dodecyl p-D-maltoside, 100 mM Tris/HCl, pH 7.4 for 60 min at 1.0 ml/min and 4". The chlorophyll-protein was then eluted by applying a 0 to 500mM NaCl gradient and loaded onto the glycerol gradient. The two procedures yielded identical results, however chromatography resulted in an higher yield.
Purification of reconstituted CP29. In some instances, such as stoichiometry determination (pigrnentslprotein ratio), it was necessary to obtain a fully purified complex which did not contain any residual contamination by bacterial proteins. This was obtained by preparative IEF of the reconstituted complex (Dainese et al., 1990) followed by DEAE chromatography (see above) in order to eliminate ampholytes and glycine which would interfere with ninhydrin reaction.
Protein and pigment concentration. The concentration of the CP29 apoprotein purified from E. coli inclusion bodies was determined by the bicinchoninic acid assay (Smith et al., 198.5). For stoichiometric (pigmentdprotein ratio) determination, the protein concentration was determined by the ninhidrin method
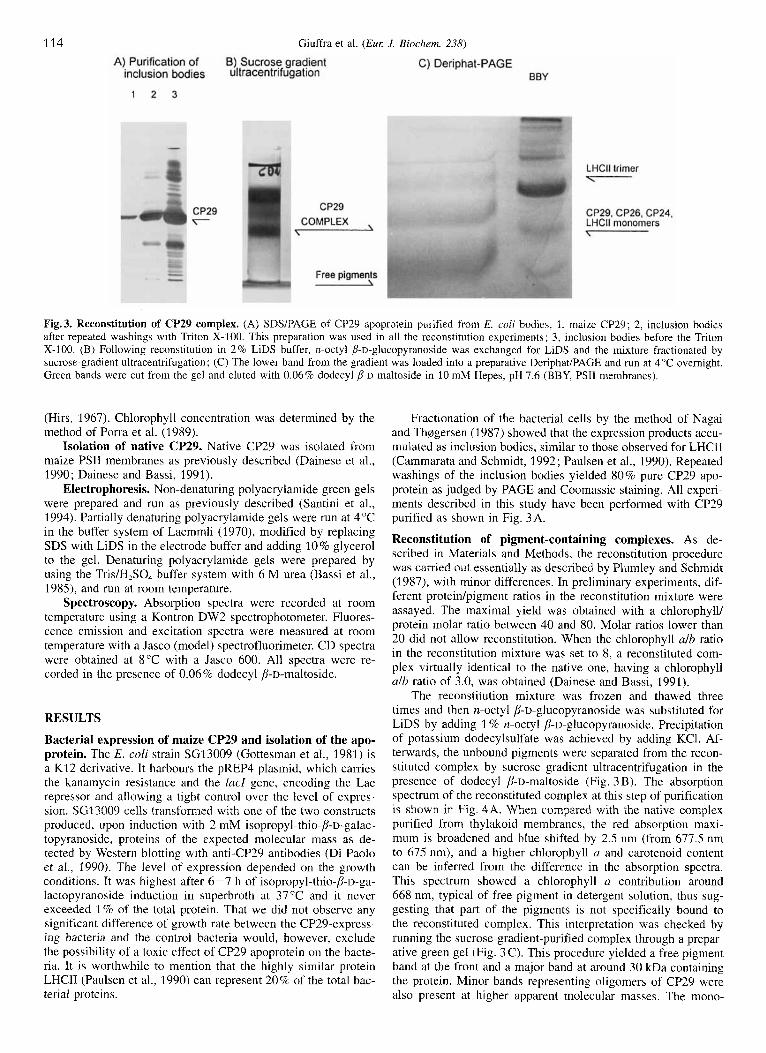
114 Giuffra et al. (Eul: J. Biochem. 238)
Fig.3. Reconstitution of CP29 complex. (A) SDSlPAGE of CF'29 apoprotein purified from E. coli bodies, 1, maize CP29; 2, inclusion bodies after repeated washings with Triton X-100. This preparation was used in all the reconstitution experiments; 3, inclusion bodies before the Triton X-100. (B) Following reconstitution in 2% LiDS buffer, n-octyl b-D-glucopyranoside was exchanged for LiDS and the mixture fractionated by sucrose-gradient ultracentrifugation; (C) The lower band from the gradient was loaded into a preparative Deriphat/PAGE and run at 4°C overnight. Green bands were cut from the gel and eluted with 0.06% dodecyl P-D-maltoside in 10 mM Hepes, pH 7.6 (BBY, PSII membranes).
(Hirs, 1967). Chlorophyll concentration was determined by the method of Porra et al. (1989).
Isolation of native CP29. Native CP29 was isolated from maize PSII membranes as previously described (Dainese et al., 1990; Dainese and Bassi, 1991).
Electrophoresis. Non-denaturing polyacrylamide green gels were prepared and run as previously described (Santini et al., 1994). Partially denaturing polyacrylamide gels were run at 4 "C in the buffer system of Laemmli (1970), modified by replacing SDS with LiDS in the electrode buffer and adding 10% glycerol to the gel. Denaturing polyacrylamide gels were prepared by using the Tris/H,SO, buffer system with 6 M urea (Bassi et al., 1985), and run at room temperature.
Spectroscopy. Absorption spectra were recorded at room temperature using a Kontron DW2 spectrophotometer. Fluores- cence emission and excitation spectra were measured at room temperature with a Jasco (model) spectrofluorimeter. CD spectra were obtained at 8°C with a Jasco 600. All spectra were re- corded in the presence of 0.06% dodecyl p-D-maltoside.
RESULTS
Bacterial expression of maize CP29 and isolation of the apo- protein. The E. coli strain SG13009 (Gottesman et al., 1981) is a K12 derivative. It harbours the pREP4 plasmid, which carries the kanamycin resistance and the l a d gene, encoding the Lac repressor and allowing a tight control over the level of expres- sion. SGI 3009 cells transformed with one of the two constructs produced, upon induction with 2 mM isopropyl-thio-p-D-galac- topyranoside, proteins of the expected molecular mass as de- tected by Western blotting with anti-CP29 antibodies (Di Paolo et al., 1990). The level of expression depended on the growth conditions. It was highest after 6-7 h of isopropyl-thio-p-D-ga- lactopyranoside induction in superbroth at 37°C and it never exceeded 1 % of the total protein. That we did not observe any significant difference of growth rate between the CP29-express- ing bacteria and the control bacteria would, however, exclude the possibility of a toxic effect of CP29 apoprotein on the bacte- ria. It is worthwhile to mention that the highly similar protein LHCII (Paulsen et al., 1990) can represent 20% of the total bac- terial proteins.
Fractionation of the bacterial cells by the method of Nagai and ThBgersen (1987) showed that the expression products accu- mulated as inclusion bodies, similar to those observed for LHCII (Cammarata and Schmidt, 1992; Paulsen et al., 1990). Repeated washings of the inclusion bodies yielded 80% pure CP29 apo- protein as judged by PAGE and Coomassie staining. All experi- ments described in this study have been performed with CP29 purified as shown in Fig. 3A.
Reconstitution of pigment-containing complexes. As de- scribed in Materials and Methods, the reconstitution procedure was carried out essentially as described by Plumley and Schmidt (1 987), with minor differences. In preliminary experiments, dif- ferent proteidpigment ratios in the reconstitution mixture were assayed. The maximal yield was obtained with a chlorophyll/ protein molar ratio between 40 and 80. Molar ratios lower than 20 did not allow reconstitution. When the chlorophyll alb ratio in the reconstitution mixture was set to 8, a reconstituted com- plex virtually identical to the native one, having a chlorophyll u/b ratio of 3.0, was obtained (Dainese and Bassi, 1991).
The reconstitution mixture was frozen and thawed three times and then n-octyl p-D-glucopyranoside was substituted for LiDS by adding 1 % n-octyl p-D-glucopyranoside. Precipitation of potassium dodecylsulfate was achieved by adding KCl. Af- terwards, the unbound pigments were separated from the recon- stituted complex by sucrose gradient ultracentrifugation in the presence of dodecyl p-D-maltoside (Fig. 3 B). The absorption spectrum of the reconstituted complex at this step of purification is shown in Fig. 4A. When compared with the native complex purified from thylakoid membranes, the red absorption maxi- mum is broadened and blue shifted by 2.5 nm (from 677.5 nm to 675 nm), and a higher chlorophyll a and carotenoid content can be inferred from the difference in the absorption spectra. This spectrum showed a chlorophyll a contribution around 668 nm, typical of free pigment in detergent solution, thus sug- gesting that part of the pigments is not specifically bound to the reconstituted complex. This interpretation was checked by running the sucrose-gradient-purified complex through a prepar- ative green gel (Fig. 3 C). This procedure yielded a free pigment band at the front and a major band at around 30 kDa containing the protein. Minor bands representing oligomers of CP29 were also present at higher apparent molecular masses. The mono-
Giuffra et al. ( E m
1,4 -
1 2 -
1 0 -
a,
( u -
$ 0 6 - . Q -
a04-
2 0 8 -
-F
0 2 -
0,o -
-0 2
J. Biochem. 238)
- A 438
~ rCP29 nCP29 rCP29-nCP29
495
I I I I I 300 400 500 600 700 800
115
0.8
8 . C 0.6 m e -
0,4 a
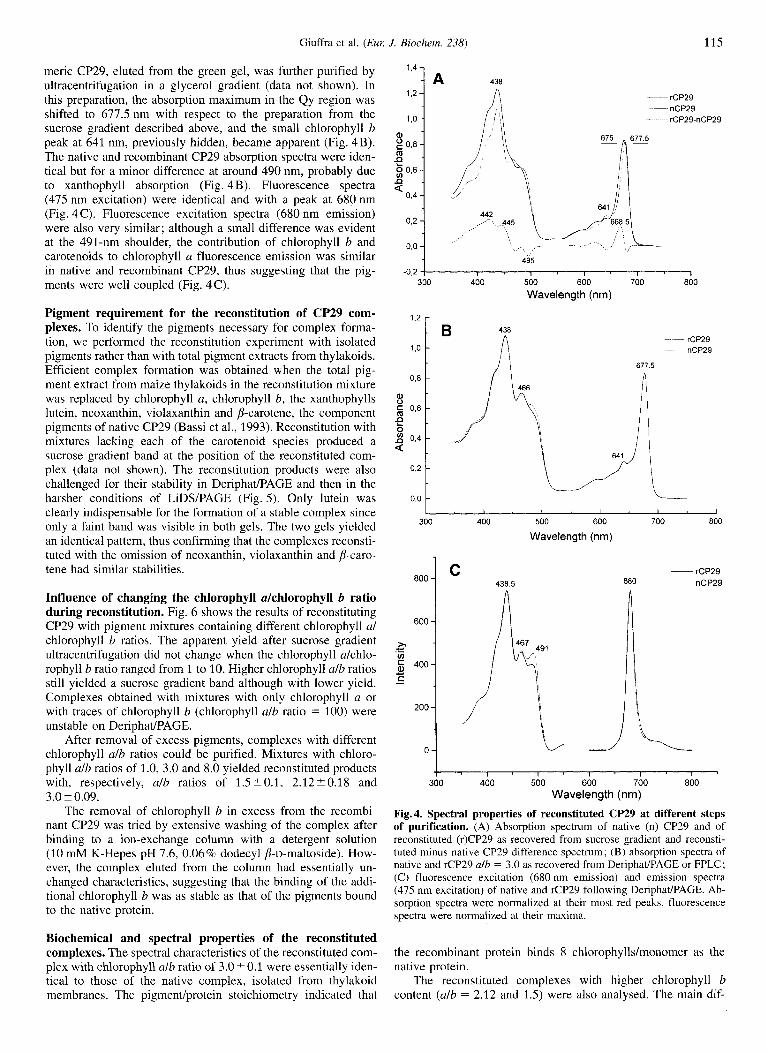
meric CP29, eluted from the green gel, was further purified by ultracentrifugation in a glycerol gradient (data not shown). In this preparation, the absorption maximum in the Qy region was shifted to 677.5 nm with respect to the preparation from the sucrose gradient described above, and the small chlorophyll b peak at 641 nm, previously hidden, became apparent (Fig. 4B). The native and recombinant CP29 absorption spectra were iden- tical but for a minor difference at around 490 nm, probably due to xanthophyll absorption (Fig. 4 B). Fluorescence spectra (475 nm excitation) were identical and with a peak at 680 nm (Fig. 4 C). Fluorescence excitation spectra (680 nm emission) were also very similar; although a small difference was evident at the 491-nm shoulder, the contribution of chlorophyll b and carotenoids to chlorophyll a fluorescence emission was similar in native and recombinant CP29, thus suggesting that the pig- ments were well coupled (Fig. 4C).
-
-
-
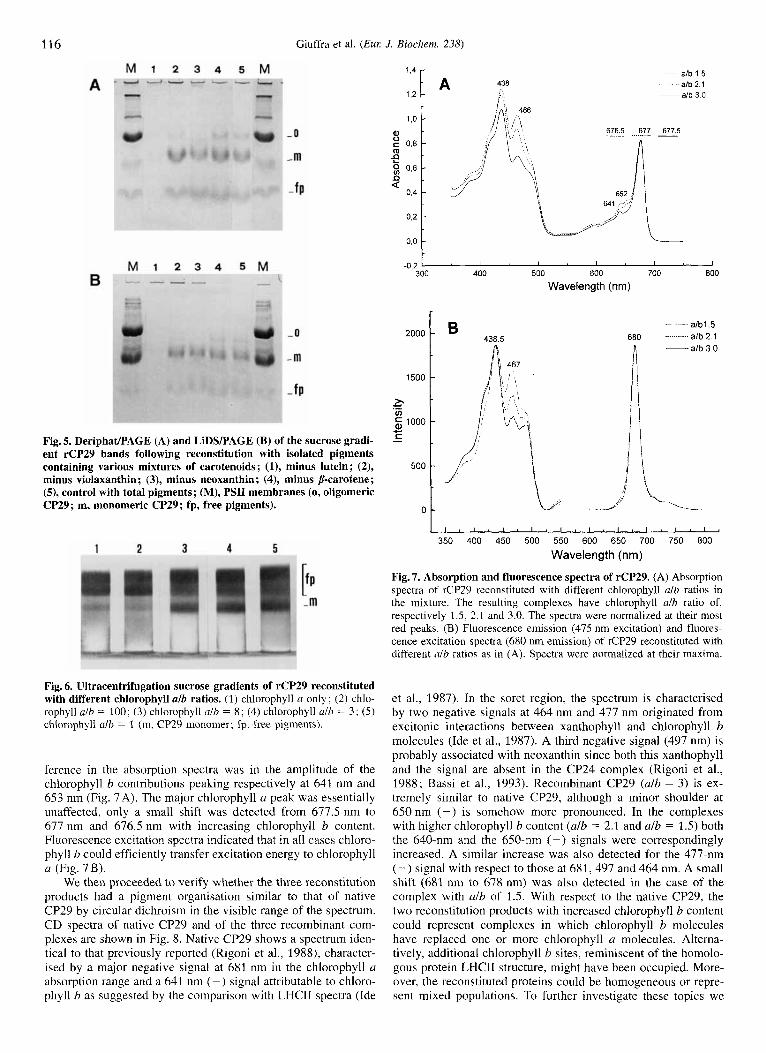
Pigment requirement for the reconstitution of CP29 com- plexes. To identify the pigments necessary for complex forma- tion, we performed the reconstitution experiment with isolated pigments rather than with total pigment extracts from thylakoids. Efficient complex formation was obtained when the total pig- ment extract from maize thylakoids in the reconstitution mixture was replaced by chlorophyll a, chlorophyll b, the xanthophylls lutein, neoxanthin, violaxanthin and p-carotene, the component pigments of native CP29 (Bassi et al., 1993). Reconstitution with mixtures lacking each of the carotenoid species produced a sucrose gradient band at the position of the reconstituted com- plex (data not shown). The reconstitution products were also challenged for their stability in Deriphat/PAGE and then in the harsher conditions of LiDSlPAGE (Fig. 5) . Only lutein was clearly indispensable for the formation of a stable complex since only a faint band was visible in both gels. The two gels yielded an identical pattern, thus confirming that the complexes reconsti- tuted with the omission of neoxanthin, violaxanthin and p-caro- tene had similar stabilities.
Influence of changing the chlorophyll alchlorophyll b ratio during reconstitution. Fig. 6 shows the results of reconstituting CP29 with pigment mixtures containing different chlorophyll a/ chlorophyll b ratios. The apparent yield after sucrose gradient ultracentrifugation did not change when the chlorophyll alchlo- rophyll b ratio ranged from 1 to 10. Higher chlorophyll alb ratios still yielded a sucrose gradient band although with lower yield. Complexes obtained with mixtures with only chlorophyll a or with traces of chlorophyll b (chlorophyll alb ratio = 100) were unstable on DeriphaVPAGE.
After removal of excess pigments, complexes with different chlorophyll alb ratios could be purified. Mixtures with chloro- phyll alb ratios of 1 .O, 3.0 and 8.0 yielded reconstituted products with, respectively, alb ratios of 1.5 kO.l. 2.1220.18 and 3 . O k 0.09.
The removal of chlorophyll b in excess from the recombi- nant CP29 was tried by extensive washing of the complex after binding to a ion-exchange column with a detergent solution (10 mM K-Hepes pH 7.6, 0.06% dodecyl p-D-rnaltoside). How- ever, the complex eluted from the column had essentially un- changed characteristics, suggesting that the binding of the addi- tional chlorophyll b was as stable as that of the pigments bound to the native protein.
Biochemical and spectral properties of the reconstituted complexes. The spectral characteristics of the reconstituted com- plex with chlorophyll alb ratio of 3.0 k 0.1 were essentially iden- tical to those of the native complex, isolated from thylakoid membranes. The pigment/protein stoichiometry indicated that
1.2
I , ;[" i": rCP29 nCP29
677 5
MI A 300 400 500 600 700 800
Wavelength (nm)
8oo] 438.5 ~ rCP29
680 nCP29
I ' I I I I
300 400 500 600 700 800 Wavelength (nm)
Fig. 4. Spectral properties of reconstituted CP29 at different steps of purification. (A) Absorption spectrum of native (n) CP29 and of reconstituted (r)CP29 as recovered from sucrose gradient and reconsti- tuted minus native CP29 difference spectrum; (B) absorption spectra of native and rCP29 alb = 3.0 as recovered from DeriphadPAGE or FPLC; (C) fluorescence excitation (680 nm emission) and emission spectra (475 nm excitation) of native and rCP29 following Deriphat/PAGE. Ab- sorption spectra were normalized at their most red peaks, fluorescence spectra were normalized at their maxima.
the recombinant protein binds 8 chlorophyllslmonomer as the native protein.
The reconstituted complexes with higher chlorophyll b content (alb = 2.12 and 1.5) were also analysed. The main dif-
116
1500
h c v) 2 1000 -
Fig. 5. DeriphaUPAGE (A) and LiDSlPAGE (B) of the sucrose gradi- ent rCP29 bands following reconstitution with isolated pigments
minus violaxanthin ; (3), minus neoxanthin; (4), minus p-carotene; (9, control with total pigments; (M), PSI1 membranes (0, oligomeric CP29; m, monomeric CP29; fp, free pigments).
= containing various mixtures of carotenoids; (I), minus lutein; (2), 500
0 -
Giuffra et al. (Eur J. Biochem. 238)
__ a/b 3 0
'. -,
-
-
-
1.4
1,2
0,o o'2 I
A 438 a lb 1 5 d b 2 1
~ a/b 3 0
677 5 -
-0,z I I I I I
300 400 500 600 700 800 Wavelength (nrn)
2ooo c 4385 afbl 5
680 a/b 2 1
1 1 1 ~ ~ ~ ~ ~ 1 1 1 1 1 l I I I I ~ I I
350 400 450 500 550 600 650 700 750 800 Wavelength (nm)
Fig. 7. Absorption and fluorescence spectra of rCP29. (A) Absorption spectra of rCP29 reconstituted with different chlorophyll alb ratios in the mixture. The resulting complexes have chlorophyll alb ratio of, respectively 1.5, 2.1 and 3.0. The spectra were normalized at their most red peaks. (B) Fluorescence emission (475 nm excitation) and fluores- cence excitation spectra (680 nm emission) of rCP29 reconstituted with different alb ratios as in (A). Spectra were normalized at their maxima.
Fig. 6. Ultracentrifugation sucrose gradients of rCP29 reconstituted with different chlorophyll alb ratios. (1) chlorophyll a only; (2) chlo- rophyll alb = 100; (3) chlorophyll ulb = 8; (4) chlorophyll alb = 3; (5) chlorophyll alh = 1 (m, CP29 monomer; fp, free pigments).
ference in the absorption spectra was in the amplitude of the chlorophyll b contributions peaking respectively at 641 nm and 653 nm (Fig. 7A). The major chlorophyll u peak was essentially unaffected, only a small shift was detected from 677.5 nm to 677 nm and 676.5 nm with increasing chlorophyll b content. Fluorescence excitation spectra indicated that in a11 cases chloro- phyll b could efficiently transfer excitation energy to chlorophyll a (Fig. 7B).
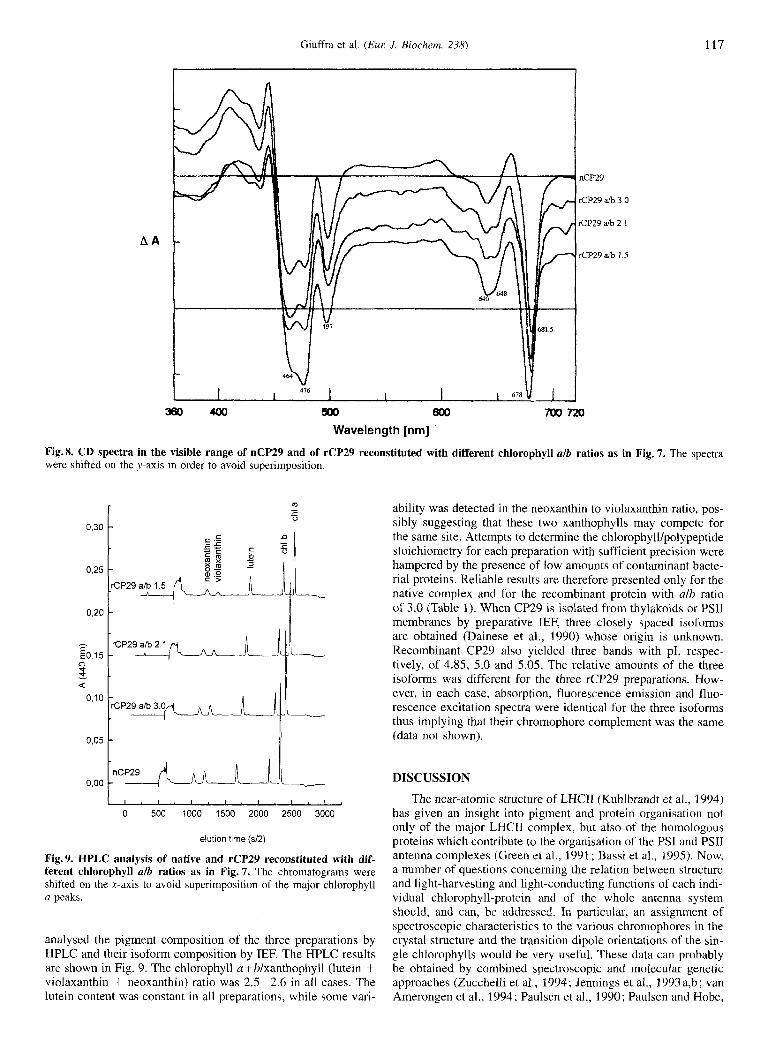
We then proceeded to verify whether the three reconstitution products had a pigment organisation similar to that of native CP29 by circular dichroism in the visible range of the spectrum. CD spectra of native CP29 and of the three recombinant com- plexes are shown in Fig. 8. Native CP29 shows a spectrum iden- tical to that previously reported (Rigoni et al., 1988), character- ised by a major negative signal at 681 nm in the chlorophyll u absorption range and a 641 nm (-) signal attributable to chloro-
et al., 1987). In the soret region, the spectrum is characterised by two negative signals at 464 nm and 477 nm originated from excitonic interactions between xanthophyll and chlorophyll b molecules (Ide et al., 1987). A third negative signal (497 nm) is probably associated with neoxanthin since both this xanthophyll and the signal are absent in the CP24 complex (Rigoni et al., 1988; Bassi et al., 1993). Recombinant CP29 (alb = 3) is ex- tremely similar to native CP29, although a minor shoulder at 650 nni (-) is somehow more pronounced. In the complexes with higher chlorophyll b content (alb = 2.1 and alb = 1.5) both the 640-nm and the 650-nm (-> signals were correspondingly increased. A similar increase was also detected for the 477-nm (-) signal with respect to those at 681, 497 and 464 nm. A small shift (681 nm to 678 nm) was also detected in the case of the complex with alb of 1.5. With respect to the native CP29, the two reconstitution products with increased chlorophyll b content could represent complexes in which chlorophyll b molecules have replaced one or more chlorophyll a molecules. Alterna- tively, additional chlorophyll b sites, reminiscent of the homolo- gous protein LHCII structure, might have been occupied. More- over, the reconstituted proteins could be homogeneous or repre-
phyll h as suggested by the comparison with LHCII spectra (Ide sent mixed populations. To further investigate these topics we
Giuffra et at. ( E m J. Biochem. 238) 117
Wavelength [nm] Fig.8. CD spectra in the visible range of nCP29 and of rCP29 reconstituted with different chlorophyll alb ratios as in Fig. 7. The spectra were shifted on the y-axis in order to avoid superimposition
0.30
0,25
0,20
- $0,15 0 v ‘0
L I . I . I . I . I . I . I ,
0 500 1000 1500 2000 2500 3000
elution time (s/2)
Fig.9. HPLC analysis of native and rCP29 reconstituted with dif- ferent chlorophyll alb ratios as in Fig. 7. The chromatograms were shifted on the x-axis to avoid superimposition of the major chlorophyll a peaks.
analysed the pigment composition of the three preparations by HPLC and their isoform composition by IEF. The HPLC results are shown in Fig. 9. The chlorophyll afblxanthophyll (lutein + violaxanthin + neoxanthin) ratio was 2.5-2.6 in all cases. The lutein content was constant in all preparations, while some vari-
ability was detected in the neoxanthin to violaxanthin ratio, pos- sibly suggesting that these two xanthophylls may compete for the same site. Attempts to determine the chlorophyll/polypeptide stoichiometry for each preparation with sufficient precision were hampered by the presence of low amounts of contaminant bacte- rial proteins. Reliable results are therefore presented only for the native complex and for the recombinant protein with d b ratio of 3.0 (Table 1). When CP29 is isolated from thylakoids or PSII membranes by preparative IEF, three closely spaced isoforms are obtained (Dainese et al., 1990) whose origin is unknown. Recombinant CP29 also yielded three bands with PI, respec- tively, of 4.85, 5.0 and 5.05. The relative amounts of the three isoforms was different for the three rCP29 preparations. How- ever, in each case, absorption, fluorescence emission and fluo- rescence excitation spectra were identical for the three isoforms thus implying that their chromophore complement was the same (data not shown).
DISCUSSION
The near-atomic structure of LHCII (Kuhlbrandt et al., 1994) has given an insight into pigment and protein organisation not only of the major LHCII complex, but also of the homologous proteins which contribute to the organisation of the PSI and PSII antenna complexes (Green et a]., 1991 : Bassi et al., 1995). Now, a number of questions concerning the relation between structure and light-harvesting and light-conducting functions of each indi- vidual chlorophyll-protein and of the whole antenna system should, and can, be addressed. In particular, an assignment of spectroscopic characteristics to the various chromophores in the crystal structure and the transition dipole orientations of the sin- gle chlorophylls would be very useful. These data can probably be obtained by combined spectroscopic and molecular genetic approaches (Zucchelli et al., 1994; Jennings et al., 1993a,b; van Amerongen et al., 1994; Paulsen et al., 1990; Paulsen and Hobe,

118 Giuffra et al. (Eul: J. Biochem. 238)
Table 1. Characteristics of native and recombinant CP29 pigment-proteins. (*), native complex purified from maize thylakoids ; (#), value determined ; (Q, value calculated assuming 3 xanthophyll molecules/monomer CP29 apoprotein.
~ _ _ _ _ _ _ ~
Chlorophyll ulb ratio Chlorophyll ulb ratio Absorption peak Chlorophyll/ Chlorophyll/ (reconstituted mixture) (complex) xantophyll ratio protein ratio
nm
(*I 3.0 C 0.08 617 5 2 6 8 +- 0.4 (#) 8 3.0 f 0.08 617.5 2.6 8 ? 0.4 (#) 3 2 12 C 0.18 611 2.5 8 f 0.4 (3) 1 1.5 f 0.1 676.5 2.5 8 20.6 (3)
1992) as previously shown in prokaryotic systems (Vermaas, 1993; Visschers et al., 1994). In the case of minor chlorophyll- proteins, besides the well-known functions of light-harvesting and energy transfer, a variety of regulatory mechanisms have been found (Bassi et al., 1993; Crofts and Yerkes, 1994; Walters et al., 1994; Bergantino et al., 1995) whose structural framework need to be elucidated by direct structural determination.
In this work we have reconstituted a CP29 complex by using pigments extracted from plant tissue and the apoprotein overex- pressed in E. coli. This method is a crucial step toward the crys- tallisation of the pigment protein. It will also allow the muta- tional analysis of CP29. In the case of LHCII (Plumley and Schmidt, 1987; Paulsen et al., 1990), the similarity of CD spectra was considered the best way to verify the correct organi- sation of the pigments. Nevertheless, we could verify that recon- stituted complexes, as obtained from the sucrose gradient, exhib- ited CD spectra identical to the fully purified complex, and yet they bound higher amounts of pigments/polypeptide compared with the native protein. The presence of not specifically bound pigments could be detected from absorption spectra. Native CP29 exhibits very typical red peaks at 677-678 nm (chloro- phyll a) and at 641 nm (chlorophyll b) while in the reconstituted complex the presence of not specifically bound pigments in- duced both a broadening and a blue shift in the red absorption peak and evidence for uncoupled pigments from fluorescence spectra (Fig. 3A, B), thus masking the underlying spectral fea- tures of the reconstituted protein.
The fully purified recombinant complexes are similar or identical to the native protein extracted from thylakoids. This conclusion follows a careful comparative analysis of the bio- chemical and spectral properties of the native and recombinant CP29. The purification procedure here described yields a CP29 holoprotein which exhibits the same pigment-protein interac- tions, as suggested by absorption and CD spectra, with respect to the native complex. This supports the view that pigment bind- ing and energy transfer is fully recovered in recombinant CP29. Moreover, native and recombinant CP29 have the same pigment/ protein ratio and pigment composition as revealed by HPLC.
Further support for the correctness of rCP29 structure comes from IEF which yields three isoforms as previously described in the case of native CP29 (Dainese et al., 1990). The reason for this heterogeneity is not understood : post-translational modifica- tion or expression of closely related genes could be hypothesised in the case of the thylakoid-derived protein but not in the case of overexpressed protein. Spectral properties appear to be iden- tical in the three isoforms thus suggesting that the pigment-bind- ing hydrophobic transmembrane region is probably not in- volved. Stable conformational states can more likely be formed by alternative folding in the large N-terminal domain.
Previous work with LHCII (Plumley and Schmidt, 1987; Paulsen et al., 1990) has shown that pigment binding during folding is independent of the composition of the reconstitution mixture. This is not the case for CP29: at least three stable forms
can be isolated differing, although within a limited extent, in chlorophyll a versus chlorophyll b binding. While the chloro- phyll alb ratio of 3.0 for the native CP29 (and the recombinant complex with alb ratio of 3.0) nicely fits the figure of 6 chloro- phyll a and 2 chlorophyll b molecules, the values of, respec- tively, 2.1 and 1.5, determined for the recombinant complexes with higher chlorophyll b content, call for heterogeneity either in the number of bound chromophores or in their ratio within individual molecules.
A possible explanation for this finding is that, depending on its concentration during folding, chlorophyll b competes for chlorophyll a sites. An alternative hypothesis can be cast on the basis of the similarity between CP29 and the LHCII complex which contains a higher number of chlorophyll b (5 versus 2) and of chlorophyll a (7 versus 6) chromophores/polypeptide. Within the three transmembrane helices and the short C-terminal amphiphilic helix containing the pigment-binding residues (Kuhlbrandt et al., 1994), the similarity between maize CP29 and LHCII, as deduced from the sequences of Lhcb4 (Bergan- tino et al., 1995) and Lhcbl (Matsuoka et al., 1987) cDNAs, is very high while all but one of the identified chlorophyll-binding residues are conserved. It is therefore likely that three chloro- phyll-b-binding sites are at least partially conserved, but not oc- cupied, in native CP29. They might be filled during folding in vitro, and this process might depend on the relative chromophore concentrations. Relevant to this question are the following : the chlorophyll/xanthophyll ratio in reconstituted complexes (Table 1) is constant; the three isoforms detected by IEF of each recon- stituted complex showed identical biochemical and spectro- scopic properties although identified chlorophyll binding resi- dues are mostly charged (Kuhlbrandt et al., 1994) and therefore IEF would seem the best way to detect heterogeneity in chromo- phore occupancy.
In conclusion, the above data suggest that chlorophyll b may compete for chlorophyll a binding sites. Thus, the complexes with chlorophyll alb ratios of 3.0 and 1.5 probably bind, respec- tively, 6 and 5 chlorophylls a, and 2 and 3 chlorophylls b. The complex with chlorophyll alb ratio of 2.1 probably represents a mixture of the 6: 2 and 5 : 3 populations. A spectroscopic analysis at higher resolution is currently in progress with the aim of elu- cidating this aspect. It is worthwhile to note that the major chlo- rophyll-b-binding protein in Proclzlorophyta is CP43', a well- known chlorophyll a protein in plants and cyanobacteria, thus supporting the view that chlorophyll b can functionally bind to chlorophyll a sites (LaRoche et al., 1995; van der Staay et al., 1995).
The higher plasticity of CP29 in pigment binding (and possi- bly also of the other minor PS I1 chlorophyll ulb complexes) with respect to LHCII, might explain its higher stability in con- ditions of limited chlorophyll b supply, such as intermittent light growth (Marquardt and Bassi, 1993; Krol et al., 1995) or chloro- phyll-b-biosynthesis mutants (Simpson et al., 1985). It has been reported that CP29 is present in the Chlorina j2 barley mutant,

Giuffra et al. (Eul: J. Biochem. 238) 119
which completely lacks chlorophyll b (White and Green, 1988). Similarly, although it was rather unstable, we have isolated rCP29 after reconstitution with only chlorophyll a and carot- enoids. The report of White and Green (1988) would imply that a CP29 binding only chlorophyll a is stable in vivo, but not in vitro. Thus, it is possible that the contrasting reports on chloro- phyll b content of CP29 (Dainese and Bassi, 1991 ; Ruban et al., 1994; Irrgang et al., 1992; Camm and Green, 1989 ; Henrysson e t al., 1989) may not be due solely to differences in the isolation protocols, but also to different growth conditions leading to dif- ferences in chlorophyll b availability. The chlorophyll a l b ratio in maize thylakoids is around 3.4. In these conditions, a chloro- phyll alb ratio of about 2 would be expected for CP29. Thus chlorophyll b might be limiting during CP29 folding within thy- lakoids. Alternatively, pigment supply to folding chlorophyll- proteins in vivo might be mediated by buffer pigment-proteins (Paulsen, 1995; Funk et al., 1994).
Conclusions. This work describes the reconstitution of CP29 holoprotein from recombinant apoprotein overexpressed in E. coli and purified pigments. A purification procedure has been devised in order to produce proteins suitable for spectroscopic studies. The recombinant (r)CP29 is essentially identical t o the native protein extracted from maize membranes as assessed by absorption, fluorescence excitation, fluorescence emission and circular dichroism spectroscopy and by HPLC analysis of bound pigments. Optimal reconstitution was obtained with a chloro- phyll alb ratio of 8 in the reconstitution mixture, while condi- tions of higher chlorophyll b availability yielded products with lower chlorophyll a lb ratios with respect to the native complex. This is probably due to competition for chlorophyll-a-binding sites. In these conditions, however, the general organisation of pigments in the complexes is essentially conserved. Based on these results, it is now feasible to use molecular genetics t o study the structure and function of the minor chlorophyll proteins.
We would like to thank Dr Elisabetta Bergantino and Prof. Andrea Squartini (University of Padua, Italy), respectively, for the kind gift of Lhcb4 cDNA clone and help in planning the plasmid-construction strat- egy, and for providing the pQE plasmids. Prof. Harald Paulsen (Botany 111, Miinchen, Germany) is thanked for helpful suggestions and Prof. Evaristo Peggion (University of Padua) for the use of the Jasco 600. M. G. Testi and Paolo Pesaresi are gratefully acknowledged for thylakoid and pigment purification. Dorianna Sandona and Aldo Pagano are thanked for advice in DNA work. Prof. Angelo Spena is thanked for critically reading the manuscript. This study was supported by the grant no. 4.7240.90 from the Italian Ministry of Agriculture and Forestry to R. B.
REFERENCES Bassi, R. & Dainese, P. (1991) A supramolecular light harvesting com-
plex from the chloroplast photosystem I1 mernbranes, Eul: J. Bio- chem. 204, 317-326.
Bassi, R., Machold, 0. & Simpson, D. J. (1985) Chlorophyll-proteins of two Photosystem I preparations from maize, Carlsberg Res. Com- mun. 50, 145-162.
Bassi, R., Pineau, B., Dainese, P. & Marquardt, J. (1993) Carotenoid- binding proteins of photosystem 11, Eur: J. Biochem. 212, 297-303.
Bassi. R. & Samamoto, H. Y. (1995) Carotenoids: localization and func- tion in photosynthetic apparatus of higher plants, in Photosynthetic antenna systems (Yokum C., ed.) pp. 1-3.5, Springer-Verlag, Heidel- berg.
Bassi, R., Giuffra, E., Croce, R., Dainese, P. & Bergantino, E. (1995) Biochemistry and molecular biology of pigment binding proteins, in NATO Advanced Study Institute, International Course : Light as en- ergy Source and information carrier in plant photuphysiology, in the press.
Bergantino, E., Dainese, P., Cerovic, Z., Sechi, S. & Bassi, R. (1995) A post-translational modification of the photosystem I1 subunit CP29 protects maize from cold stress, J. Biol. Chem. 270, 8474-8481.
Bujard, H., Gentz, R., Lanzer, M., Stiiber, D., Miiller, M., Ibrahimi, I., Hauptle, M. T. & Dobberstein, B. (1987) A T5-promoter based tran- scription-translation system for the analysis of proteins in vitro and in vivo, Methods Enzymol. 155, 416-433.
Camm, E. L. & Green, B. R. (1989) The chlorophyll ab complex, CP29, is associated with the Photosystem I1 reaction centre core, Biochim.
Cammarata, K. V. & Schmidt, G. W. (1992) In vitro reconstitution of a light-harvesting gene product: deletion mutagenesis and analysis of pigment binding, Biochemistry 31, 2779-2789.
Dainese, P., Hoyer-Hansen, G. & Bassi, R. (1990) The resolution of chlorophyll d b binding proteins by a preparative method based on flat bed isoelectric focusing, Photochem. Photobiol. 51, 693 -703.
Dainese, P. & Bassi, R. (1991) Subunit stoichiometry of the chloroplast photosystem I1 antenna system and aggregation state of the compo- nent chlorophyll alb binding proteins, J. Biol. Chem. 266, 8136- 8142.
Davies, B. H. (1965) Carotenoids, in Chemistry and biochemistry of plant pigments (Goodwin, T. W., ed.) 2nd edn, pp. 38-155, Aca- demic Press, New York.
Demmig-Adams, B. (1992) Carotenoids and photoprotection in plants: a role for the xanthophyll zeaxanthin, Biochim. Biophys. Acta 1020,
Di Paolo, M. L., Peruffo dal Belin, A. & Bassi, R. (1990) Immunological studies on chlorophyll alb proteins and their location in chloroplast membrane domains, Planta (Heidelb.) 181, 27.5 -286.
Funk, C., Schroder, W. P., Green, B. R., Renger, G. & Anderson, B. (1994) The intrinsic 22 kDa protein is a chlorophyll-binding subunit of photosystem 11, FEBS Lett. 342, 261 -266.
Gilmore, A. M. & Samamoto, H. Y. (1991) Resolution of lutein and zeaxanthin using a non-endcapped, lightly carbon-loaded C,, high- performance liquid chromatograpic column, J. Chromatogr: 543,
Gottesman, S., Halpern, E. & Trisler, P. (1981) Role of sulA and sulB in filamentation by Lon mutants of Escherichia coli K-12, J. Bacte- rial. 148, 265-273.
Henrysson, T., Schroeder, W. P., Spangfort, M. & Akerlund, H. E. (1989) Isolation and characterization of the chlorophyll d6 protein complex CP29 from spinach, Biochim. Biophys. Acta 977, 301 -308.
Hirs, C. H. W. (1967) Detection of peptides by chemical methods, Meth- ods Enzymol. 11, 325-329.
Ide, J. P., Klug, D. R., Kuhlbrandt, W., Giorgi, L. B. & Porter, G. (1987) The state of detergent solubilised chlorophyll-alb protein complex as monitored by picosecond time-resolved fluorescence and circular dichroism, Biochim. Biophys. Acta 893, 349-364.
Irrgang, K. D., Renger, G. & Vater, J. (1991) Isolation, purification and partial characterization of a 30-kDa chlorophyll-&-binding protein from spinach, Eur: J. Biochem. 201, 515-522.
Jansson, S., Pichersky, E., Bassi, R., Green, B. R., Ikeuki, M., Melis, A., Simpson, D. J., Spangforth, M., Staehelin, L. A. & Thoi-nber, J. I? (1992) A nomenclature for the genes encoding the chlorophyll d b binding proteins of higher plants, Plant Mol. Bid. Rep. 10, 242- 253.
Jennings, R. C., Bassi, R., Garlaschi, F. M., Dainese, P. & Zucchelli, G. (1 993 a) Distribution of the chlorophyll spectral forms in the chloro- phyll-protein complexes of photosystem I1 antenna, Biochemistry 32,
Jennings, R. C., Garlaschi, F. M., Bassi, R., Zucchelli, G., Vianelli, A. & Dainese, P. (1993b) A study of photosystem I1 fluorescence emission in terms of the antenna chlorophyll-protein complexes, Biochim. Bio- phys. Acta 1183, 194-200.
Kr61, M., Spangfort, M. D., Huner, N. P. A,, Oquist, G., Gustaffson, P. & Jansson, S. (1995) Chlorophyll ulb-binding proteins, pigment conversions, and early light-induced proteins in a chlorophyll b-less barley mutant, Plant Physiol. (Brthesda) 107, 873-883.
Kiihlbrandt W., Wang, D. N. & Fujiyoshi, Y. (1994) Atomic model of plant light-harvesting complex by electron crystallography, Nature 367, 614-621.
LaRoche, J., Partensky, F. & Falkowsky, P. (1995) The major Light har- vesting chlorophyll binding protein of Prochlorococcus marinus is
Biophys. Acts 974, 180- 184.
1-24.
137- 145.
3203-3210.

120 Giuffra et al. (Eur: J. Biochem. 238)
similar to CP43’, a chl binding protein induced by iron depletion in cyanobacteria, in Photosynthesis : from light to biosphere (Mathis, P., ed.) vol. 1, pp. 171-174.
Marquardt, J. & Bassi, R. (1993) Chlorophyll-proteins from maize seed- lings grown under intermittent light conditions. Their stoichiometry and pigment content, Planta (Heidelb.) 191, 265-273.
Nagai, K. & Thpgersen, H. C. (1987) Synthesis and sequence-specific proteolysis of hybrid proteins produced in Escherichia coli, Mefhods Enzymol. 153, 461 -481.
Paulsen, H. (1995) Chlorophyll db-binding proteins, Photochem. Pho- tobiol. 62, 367-382.
Paulsen, H. & Hobe, S. (1992) Pigment-binding properties of mutant light-harvesting chlorophyll alb-binding protein, Eua J . Biochem. 205, 71 -76.
Paulsen, H., Riimler, U. & Rudiger, W. (1990) Reconstitution of pig- ment-containing complexes from light-harvesting chlorophyll ulb- binding protein overexpressed in Escherichia coli. Planta (Heidelb.)
Peter, G. F. & Thornber, J. P. (1991) Biochemical composition and or- ganisation of higher plant photosystem I1 light harvesting pigment- proteins, J . Bid. Chem. 266, 16745 - 16 754.
Plumley, F. G. & Schmidt, G. W. (1987) Reconstitution of chlorophyll alb light harvesting complexes: xanthophyll-dependent assembly and energy transfer, Proc. Natl Acad. Sci. USA 84, 146-150.
P o ~ T ~ , R. J., Thompson, W. A. & Kriedermann, P. E. (1989) Determina- tion of accurate extinction coefficients and simultaneous equations for assaying chlorophyll a and b extracted with four different solvents : verification of the concentration of chlorophyll standards by atomic absorption spectroscopy, Biochim. Biophys. Acta 975,
Rigoni, F., Bassi, R. & Giacometti, G. M. (1988) Spectroscopic charac- terisation of purified chlorophyll d b proteins CP29, CP26 CP24 from maize antenna complex, in Techniques and new developments in photosynthesis research (Barber, J. & Malkin, R., eds) pp. 141 - 144, Plenum Press, London, New York.
Ruban, A. V., Walters, R. G. & Horton, P. (1994) The molecular mecha- nism of the control of excitation energy dissipation in chloroplast membranes. Inhibition of ApH-dependent quenching of chlorophyll fluorescence by dicyclohexylcarbodiimide, FEBS Left. 309, 175 ~
179. Santini, C., Tidu, V., Tognon, G., Ghiretti Magaldi, A. & Bassi, R. (1994)
Three-dimensional structure of the higher-plant photosystem I1 reac-
181, 204-21 1.
384-394.
tion centre and evidence for its dimeric organization in vivo, Eur: J . Biochern. 221, 307-315.
Simpson, D. (1979) Freeze-fracture studies on barley plastid membranes 111. Location of the chlorophyll a-protein 1, Carlsberg Rex Commun.
Simpson, D. J., Mahold, O., Hoyer-Hansen, G. & von Wettstein, D. (1985) Chlorina mutants of barley (Hordeum vulgare L.), Carlsberg Res. Commun. 50, 223-238.
Smith, P. K., Krohn, R. I., Hermanson, G. T., Mallia, A. K., Gartner, F. H., Provenzano, M. D., Fujimoto, E. K., Goeke, N. M., Olson, B. J. & Klenk, D. C. (1985) Measurement of protein using bicinchoni- nic acid, Anal. Biochem. 150, 76-85.
Van Amerongen, H., van Bolthuis, B. M., Betts, S., Mei, R., van Gron- delle, R., Yokum, C. F. & Dekker, J. P. (1994) Spectroscopic charac- terisation of CP26, a chlorophyll alb binding protein of the higher plant photosystem I1 complex, Biochim. Biophys. Actu 1188, 227- 234.
Van der Staay, G. W. M., Ducret, A., Aebersold. R., Li, R., Golden, S. S., Hiller, R. G., Wrench, P. M., Larkum, A. W. I). & Green, B. R. (1995) The chlorophyll alb antenna from Prochlorophytes is related to the iron-stress induced chlorophyll a antenna (IsiA) from Cyano- bacteria, in Photosynthesis; from light fo biosphere (Mathis, P., ed.) VOI. 1, pp. 175-178.
Vermaas, W. (1 993) Molecular-biological approaches to analyze pho- tosystem-I1 structure and function, Annu. Rev. Plant Physiol. 44, 457-481.
Visscher, R. W., Crielaard, W., Fowler, G. J. S., Hunter, C. N. & van Grondelle, R. (1994) Probing the B800 bacteriochlorophyll binding site of the accessory light-harvesting complex from Rhodobacter sphperoides using site-directed mutants. 2. A low temperature spectroscopy study of stuctural aspects of the pigment-protein con- formation, Biochim. Biophys. Acta 1183, 483 -490.
Walters, R. G., Ruban, A. V. & Horton, P. (1994) Higher plant light- harvesting complexes LHCIIa and LHCIIc are bound by dicyclohex- ylcarbodiimide during inhibition of energy dissipation, Eur: J. Bio- chem. 226, 1063-1069.
White, M. J. & Green, B. R. (1988) Intermittent-light chloroplasts are not developmentally equivalent to chlorinu $2 chloroplasts in barley, Photosynth. Res. 15, 195-203.
44, 305-336.