prepubertal fischer 344 rats display stronger morphine-induced taste avoidance than prepubertal...
TRANSCRIPT

Prepubertal Fischer 344 RatsDisplay Stronger Morphine-Induced Taste Avoidance ThanPrepubertal Lewis Rats
ABSTRACT: The present report asked if the previously reported differences inmorphine-induced conditioned taste avoidance between adult F344 and LEWrats (F344> LEW) are also evident in prepubescence (early adolescence). Toassess this possibility, adult (Experiment 1) and prepubertal (Experiment 2)F344 and LEW rats were assessed for their ability to acquire morphine-inducedtaste avoidance (0, 3.2, 10, or 18mg/kg) in a modified taste avoidanceprocedure. In each experiment, rats of both strains were given repeated pairingsof saccharin and morphine followed by a final two-bottle avoidance test. Adultand prepubertal F344 subjects displayed a more rapid acquisition of theavoidance response as well as stronger suppression of consumption than theirLEW counterparts. These data suggest the strains differ in their sensitivity to theaversive effects of morphine and that this differential sensitivity is evident earlyin development and is developmentally stable. The basis for these straindifferences in morphine-induced avoidance was discussed. � 2013 WileyPeriodicals, Inc. Dev Psychobiol 56: 979–988, 2014.
Keywords: strain differences; prepubertal; early adolescence; adult; F344;LEW; morphine-induced taste avoidance; development
INTRODUCTION
Recent work on conditioned taste avoidance (CTA)
learning between the inbred F344 and LEW rat strains
has demonstrated strain differences in drug-induced
taste avoidance that vary depending on the drug of
abuse tested (for cocaine, see Davis & Riley, 2007;
Glowa, Shaw, & Riley, 1994; for nicotine, see Pesca-
tore, Glowa, & Riley, 2005; for ethanol, see Roma,
Flint, Higley, & Riley, 2006). One compound for which
such strain differences have been well characterized is
morphine. Specifically, Lancellotti, Bayer, Glowa,
Houghtling, and Riley (2001) reported that the F344
strain displayed dose-dependent morphine-induced taste
avoidance, while the LEW strain did not acquire
avoidance at any dose tested, even after repeated
conditioning trials (Davis, Cobuzzi, & Riley, 2012; see
also Gomez-Serrano, Kearns, & Riley, 2009; for a
review, see Riley, 2011). That the F344 animals show
stronger avoidance is interesting in that they also self-
administer less morphine (Ambrosio, Goldberg, &
Elmer, 1995; Garcıa-Lecumberri et al., 2011; Martın
et al., 1999, 2003) and form weaker place preferences
(Davis, Roma, Dominguez, & Riley, 2007; Grakalic,
Schindler, Baumann, Rice, & Riley, 2006) than the
LEW strain, suggesting that morphine intake in these
strains may be impacted by the relative balance of
these affective properties.
Although the differences between the F344 and
LEW strains with respect to morphine-induced taste
avoidance are consistently observed in adults (see
above), it remains unknown if such effects are evident
early in life and are developmentally stable or if they
Manuscript Received: 2 July 2013Manuscript Accepted: 30 September 2013The authors declared that they have no conflicts of interest.Correspondence to: Zachary E. HurwitzContract grant sponsor: Mellon FoundationContract grant sponsor: American University Artists and Scholars
FellowshipArticle first published online in Wiley Online Library
(wileyonlinelibrary.com): 28 October 2013DOI 10.1002/dev.21176 � � 2013 Wiley Periodicals, Inc.
Developmental Psychobiology
Zachary E. Hurwitz
Jennifer L. Cobuzzi
Andrew P. Merluzzi
Bradley Wetzell
Anthony L. Riley
Psychopharmacology LaboratoryDepartment of Psychology
American University4400 Mass. Ave., NW, Washington, DC
20016E-mail: [email protected]

are a function of specific developmental histories that
result in differential behavior in adulthood. Interesting-
ly, strain differences have been reported to vary over
development for a number of physiological and behav-
ioral endpoints in a variety of rodent strains. For
example, Wilking et al. (2012) reported that nicotine
preference varied as a function of age and dose in the
C3H/Ibg and C57bl/6j mouse strains. Specifically, there
were no differences between the strains across various
doses tested in late adolescence and adulthood. Howev-
er, in early adolescence C3H/Ibg subjects were more
sensitive than the C57bl/6j strain at 10mg concentration
while in middle adolescence C3H/Ibg subjects emitted
less behavior than C57bl/6j mice at 20 and 30mgconcentrations (for other examples of developmental
differences in strain comparisons, see Allam, 2012;
Fairless et al., 2012; Farid, Martinez, Geyer, &
Swerdlow, 2000; Moore, Forrest, & Boehm, 2013;
Moore, Linsenbardt, Melon, & Boehm, 2011; Paylor,
Baskall-Baldini, Yuva, & Wehner, 1996; Satinder,
1981; Sinaiko & Mirkin, 1974; Tonkiss, Shultz, &
Galler, 1992; Vogl, Atchley, & Xu, 1994).
Although age differences across a variety of strains
have been well characterized, little has been examined
with the F344 and LEW rat lines, especially studies
examining the ontogenesis of strain differences in drug
reactivity (see Gomez-Serrano, Sternberg, & Riley,
2002; Gomez-Serrano, Tonelli, Listwak, Sternberg, &
Riley, 2001). In the only developmental assessment of
behavioral differences between the two strains, Siviy,
Love, DeCicco, Giordano, and Seifert (2003) reported
that strain differences in play behavior (LEW>F344)
were present and comparable at all developmental
periods assessed. Although suggestive that these pheno-
typic differences are highly heritable and stable, no
developmental assessments have been made with the
F344 and LEW strains in relation to their relative
sensitivities to drugs of abuse yet alone their ability to
acquire drug-induced taste avoidance.
It is important to note in this context that prepubertal
(early adolescent) and adult outbred rats differ dramati-
cally in taste avoidance learning (Anderson, Agoglia,
Morales, Varlinskaya, & Spear, 2012; Anderson, Varlin-
skaya, & Spear, 2010; Cobuzzi et al., 2013; Hurwitz,
Merluzzi, & Riley, 2013; Infurna & Spear, 1979;
Schramm-Sapyta et al., 2007; Schramm-Sapyta, Morris,
& Kuhn, 2006; Shram, Funk, Li, & Le, 2006; Vetter-
O’Hagen, Varlinskaya, & Spear, 2009; Wilmouth &
Spear, 2004) with prepubertal rats exhibiting significant-
ly weaker taste avoidance relative to their adult counter-
parts. Such results suggest that prepubescent rats are less
sensitive to the aversive effects of such drugs than adults,
a differential sensitivity that may confer an increased
susceptibility to the subsequent use and abuse of these
compounds (see Doremus-Fitzwater, Varlinskaya, &
Spear, 2010; Misanin, Anderson, & Hinderliter, 2009;
Schramm-Sapyta et al., 2010; Spear, 2013).
Given that differences with other strains have been
shown to vary over development and that outbred
prepubertal and adult rats differ significantly in drug-
induced avoidance across a range of drugs of abuse, it
might be predicted that the patterns evident in F344 and
LEW may differ across development. If prepubertal
F344 and LEW rats do not differ in their avoidance
patterns (and as such differ from the pattern displayed by
adults), one might argue that some unique developmental
history interacting with the unique genetic backgrounds
of the two strains may mediate the results typically seen
in adults. Conversely, if the strain differences observed
in adults are also present in prepubescence, it would
suggest that these differences in avoidance learning are
highly heritable, substantiating the use of these strains as
genetic models for drug sensitivity, use and abuse
(Beitner-Johnson, Guitart, & Nestler, 1991; Guitart et al.,
1993; Kosten et al., 1997). To address this issue, pre-
pubescent rats of the F344 and LEW stains were given
access to a novel saccharin solution and injected with
varying doses of morphine (Experiment 2). The specific
procedure employed (see below) was modified from the
typical taste avoidance procedure to accommodate their
developmental window and allow for assessment of
avoidance learning under conditions that minimize the
effects of fluid deprivation (and concomitant weight loss;
see Hurwitz et al., 2013). This modification employs
deprivation procedures not generally used in work with
adults. Consequently, prior to this assessment, adult
F344 and LEW subjects were tested under similar
conditions to assure that the often-reported differences
between the two strains in adult animals were evident
under the modified procedure (Experiment 1).
METHOD
Subjects
One hundred thirty-nine experimentally naıve male
F344 and LEW rats (Harlan Laboratories, Indianapolis,
IN) arrived at the facility on PND 21, weighing
approximately 40 g. Experiments 1 and 2 were each run
in two replicates with subjects in all dose groups and
both strains represented in each replicate. Food and
water were available ad libitum unless stated otherwise.
Procedures recommended by the National Research
Council (1996) and the Committee on Guidelines for
the Care and Use of Animals in Neuroscience and
Behavioral Research (2003) were followed at all times.
The research was reviewed and approved by the
American University IACUC.
980 Hurwitz et al. Developmental Psychobiology

Apparatus
Upon arrival to the animal colony, subjects in each
strain were initially handled and then group housed (2–
4 rats per bin) in polycarbonate bins (23 cm� 44 cm
� 21 cm). All subjects were maintained on a 12:12
light–dark cycle (lights on at 0800 hr) and at an
ambient temperature of 23˚C. All conditioning and
testing occurred during the light phase of the light–dark
cycle at 0800 hr and in the same room in which the
animals were housed. During adaptation and condition-
ing, animals were transferred to individual hanging
wire-mesh test cages (24.3 cm� 19 cm� 18 cm) but
were returned to their group-housed bins after condi-
tioning trials (see details below).
Drugs
Morphine sulfate was generously supplied by the
National Institute on Drug Abuse. For behavioral
testing, morphine sulfate was dissolved in sterile
isotonic saline (.9%) at a concentration of 5mg/ml and
was subsequently filtered through a .2mm filter to
remove any contaminants before being administered
subcutaneously (SC) at a dose of 3.2, 10, or 18mg/kg.
Morphine administered at these doses and by this route
has been reported to induce dose-dependent avoidance
in outbred rats (Hurwitz et al., 2013) and in the F344
and LEW strains (Davis et al., 2012; Gomez-Serrano
et al., 2009; Lancellotti et al., 2001). Sterile isotonic
saline was also filtered before being administered to
vehicle controls and was given equivolume to the
highest dose of morphine administered (18mg/kg).
Volume of the injection was manipulated in favor of
concentration given the influence concentration has on
drug absorption and distribution.
Morphine-Induced Taste Avoidance
Experiment 1: Adults. Procedures were adapted from
Hurwitz et al. (2013). Specifically, F344 (n¼ 35) and
LEW subjects (n¼ 36) were brought into the laboratory
on PND 21 and maintained on ad libitum food and
water until PND 77 to permit the control of their
developmental environment (housing conditions, han-
dling, and light/dark cycle). Subjects were weighed and
handled and had their water consumption measured
from PND 77–83 at which point water adaptation began.
On PND 84, subjects in each strain underwent full fluid
deprivation at 0800 hr and on the following day (PND
85) they were removed from their group-housed bin at
0800 hr, weighed and placed into individual test cages
where they were given 45-min access to tap water in
graduated 50-ml Nalgene tubes beginning at 0830 hr.
This specific procedure was used to minimize the
potential stress associated with more severe deprivation
schedules, for example, extended 20-min daily access,
in adolescent subjects (see Anderson et al., 2010; Hur-
witz et al., 2013). Under this modified deprivation
procedure, fluid consumption is generally lower, neces-
sitating longer access to assure sufficient consumption.
After 45min, bottles were removed, consumption was
recorded and subjects remained in the hanging cages for
an additional 20min before being returned to their
group-housed bin and given ad libitum water until
0800 hr the following day (PND 86). This procedure
was repeated one additional time to adapt animals to
consuming fluid in the test cages prior to being given
access to saccharin during conditioning. On the day
prior to the first saccharin conditioning day, water access
was again fully restricted before subjects underwent
conditioning in the test cages. On this first conditioning
trial (PND 89), subjects were weighed and handled (as
previously described) and given 45-min access to a
novel saccharin solution (1 g/L) in the test cages after
which they remained for an additional 20min. At this
point, subjects in each strain (independent of their
group-housed bin) were assigned to one of four groups
such that saccharin intake was comparable among
groups. Based on these group assignments, subjects in
each strain were injected with either morphine (3.2, 10,
or 18mg/kg, SC) or vehicle and then returned to their
group-housed bins and given ad libitum water until
0800 hr the following day. This procedure yielded
Groups F0 (n¼ 9), L0 (n¼ 9), F3.2 (n¼ 8), L3.2
(n¼ 8), F10 (n¼ 8), L10 (n¼ 9), F18 (n¼ 10), and L18
(n¼ 10) where the letter refers to the strain of the
subject and the number indicates the dose of morphine
administered. On the next day, subjects in each bin had
their fluid consumption restricted before the next
saccharin conditioning trial. This 2-day procedure
(saccharin-water recovery-full deprivation) was repeated
four times. On the day after the fourth cycle (PND 97),
subjects were transferred to test cages where two 50-ml
Nalgene tubes (one containing tap water; the other
containing the .1% saccharin solution) were affixed to
the cage for 45min and consumption of both solutions
was recorded. Placement of the bottles was counter-
balanced across all subjects to prevent positioning
effects. After the 45min, the bottles were removed,
consumption was recorded and subjects were returned to
their home cages where water was available ad libitum.
Experiment 2: Prepubescents. The procedures de-
scribed above were identical for the prepubescent rats
with the following exceptions. F344 and LEW rats
(n¼ 34 per strain) were brought into the facility at
PND 21 and were weighed and handled and had their
water consumption measured until PND 29. Following
Developmental Psychobiology Prepubertal Strain Differences in Morphine CTA 981

water adaptation (PND 29–32; see above), conditioning
began. During conditioning (PND 33–40), subjects in
each strain were given four saccharin-morphine (or
vehicle) pairings until the two-bottle test, which was
administered on PND 41. The group designations were
similar to those in Experiment 1: Groups F0 (n¼ 9), L0
(n¼ 9), F3.2 (n¼ 8), L3.2 (n¼ 8), F10 (n¼ 8), L10
(n¼ 8), F18 (n¼ 9), and L18 (n¼ 9).
Statistical Analysis
Acquisition. A 2 (Replicate)� 2 (Strain)� 4 (Dose)� 4
(Trial) mixed model ANOVA on saccharin consumption
(ml) on the four conditioning trials was run for each
experiment to determine differences between the strains as
a function of dose and trial. One-way ANOVAs and
Tukey’s HSD posthost tests were employed where merited
by significant interactions to determine differences
between the strains and doses across trials. To determine if
there were differences in saccharin consumption between
Conditioning Trials 1 and 4, Bonferroni-corrected t-tests
were employed (a¼ .0125) for each group.
Two-Bottle Test. A 2 (Replicate)� 2 (Strain)� 4
(Dose) univariate ANOVA was run for each experiment
on the percent saccharin consumed on the two-bottle
test. One-way ANOVAs and Tukey’s HSD post hoc
tests were used where merited to evaluate differences
in the percent saccharin consumed between replicates
and strains at the different dose groups.
RESULTS
Experiment 1: Adults
Acquisition. Over conditioning, F344 subjects drank
significantly less saccharin than LEW subjects, indica-
tive of greater morphine-induced taste avoidance. The 2
(Replicate)� 2 (Strain)� 4 (Dose)� 4 (Trial) mixed-
model ANOVA yielded significant effects of Trial [F
(3, 156)¼ 11.611], Strain [F (1, 55)¼ 159.148], Dose
[F (3, 55)¼ 37.403] as well as significant Trial�Repli-
cate [F (3, 156)¼ 6.950], Trial�Strain [F (3,
156)¼ 23.220], Trial�Dose [F (9, 156)¼ 22.869],
Strain�Dose [F (3, 55)¼ 1.389], Replicate�Strain [F
(1, 55)¼ 7.536] and Trial� Strain�Dose [F (9,
156)¼ 6.098] interactions.
Although on Trial 1 F344 subjects did not differ in
saccharin consumption, on Trials 2–4 all drug-treated
F344 subjects drank less saccharin than vehicle-treated
controls with Group 18 drinking less saccharin than
Group 3.2. On Trial 1, LEW subjects also did not differ
in saccharin consumption. On Trials 2–4, Group 18
drank less than Group 0. On all trials, F344 subjects
consumed significantly less saccharin than LEW sub-
jects. Tukey’s HSD revealed that on Trial 1 Groups F0,
F3.2, and F18 drank significantly less saccharin than
Groups L0, L3.2, and L18, respectively, with no differ-
ences between Groups F10 and L10. On Trials 2–4,
Groups F0 and L0 did not differ but Groups F3.2, F10,
and F18 drank significantly less saccharin than Groups
L3.2, L10, and L18, respectively (see Fig. 1).
Over conditioning, Group F0 displayed a significant
increase in saccharin intake from Trials 1 to 4 [t
(8)¼ 5.812]. Group F3.2 [t (7)¼ 3.036] exhibited no
significant changes in saccharin consumption, while
Group F10 [t (7)¼ 10.583] and Group F18 [t
(7)¼ 11.239] significantly decreased saccharin intake
over trials. Group L0 [t (8)¼ 3.440] exhibited a
significant increase in saccharin consumption over trials,
while Groups L3.2 [t (7)¼ 1.843], L10 [t (8)¼ .000],
and L18 [t (9)¼ 1.467] did not display any significant
changes in saccharin intake from Trials 1 to 4.
Two-Bottle Test. On the two-bottle test, the percent
saccharin consumed by morphine-injected adult F344
A B
FIGURE 1 Mean (�SEM) saccharin consumption (ml) by F344 (A) and LEW (B) adults
during acquisition. #Significant differences between Group 3.2 and Group 18. �Significantdifferences between Group 0 and Groups 3.2, 10 and 18. $Significant differences between Group
0 and Group 18.
982 Hurwitz et al. Developmental Psychobiology
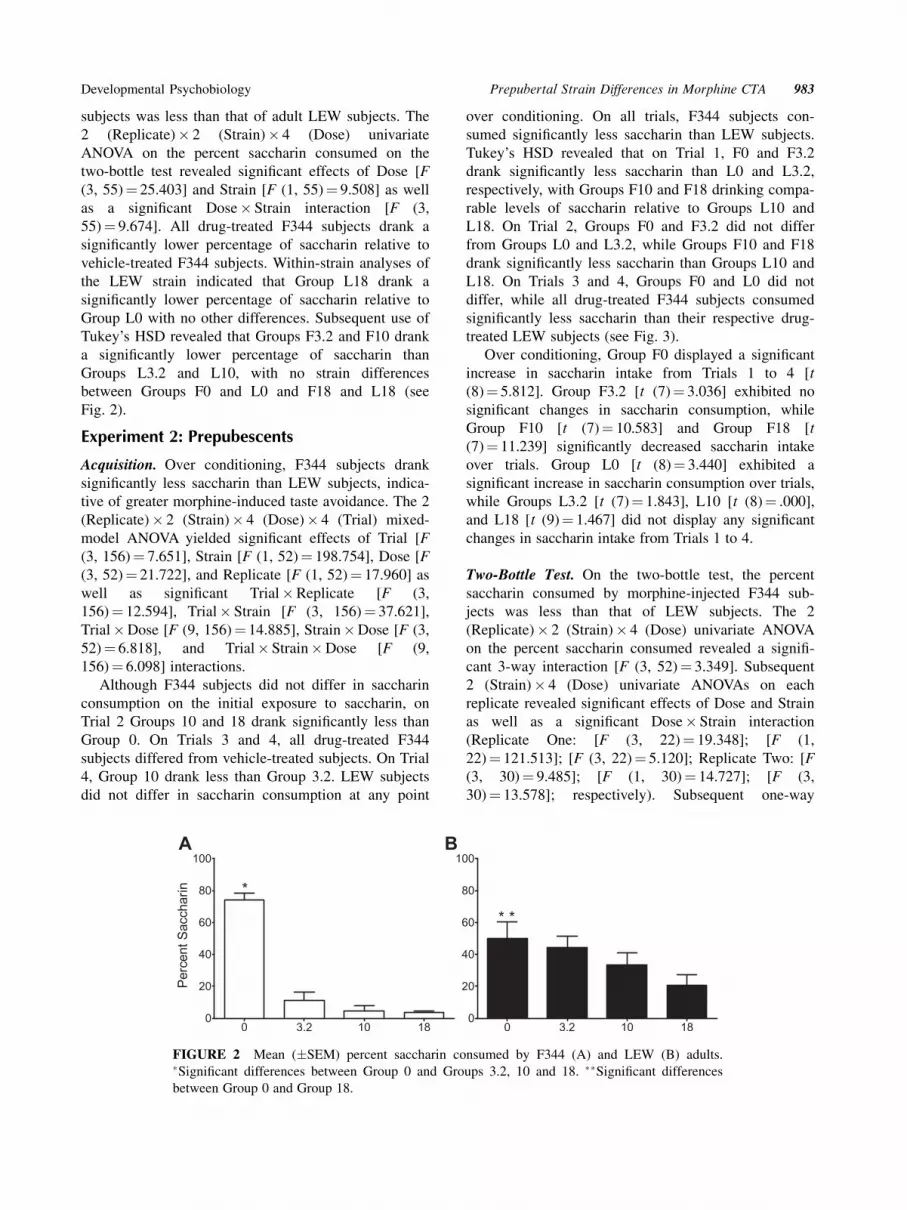
subjects was less than that of adult LEW subjects. The
2 (Replicate)� 2 (Strain)� 4 (Dose) univariate
ANOVA on the percent saccharin consumed on the
two-bottle test revealed significant effects of Dose [F
(3, 55)¼ 25.403] and Strain [F (1, 55)¼ 9.508] as well
as a significant Dose� Strain interaction [F (3,
55)¼ 9.674]. All drug-treated F344 subjects drank a
significantly lower percentage of saccharin relative to
vehicle-treated F344 subjects. Within-strain analyses of
the LEW strain indicated that Group L18 drank a
significantly lower percentage of saccharin relative to
Group L0 with no other differences. Subsequent use of
Tukey’s HSD revealed that Groups F3.2 and F10 drank
a significantly lower percentage of saccharin than
Groups L3.2 and L10, with no strain differences
between Groups F0 and L0 and F18 and L18 (see
Fig. 2).
Experiment 2: Prepubescents
Acquisition. Over conditioning, F344 subjects drank
significantly less saccharin than LEW subjects, indica-
tive of greater morphine-induced taste avoidance. The 2
(Replicate)� 2 (Strain)� 4 (Dose)� 4 (Trial) mixed-
model ANOVA yielded significant effects of Trial [F
(3, 156)¼ 7.651], Strain [F (1, 52)¼ 198.754], Dose [F
(3, 52)¼ 21.722], and Replicate [F (1, 52)¼ 17.960] as
well as significant Trial�Replicate [F (3,
156)¼ 12.594], Trial� Strain [F (3, 156)¼ 37.621],
Trial�Dose [F (9, 156)¼ 14.885], Strain�Dose [F (3,
52)¼ 6.818], and Trial� Strain�Dose [F (9,
156)¼ 6.098] interactions.
Although F344 subjects did not differ in saccharin
consumption on the initial exposure to saccharin, on
Trial 2 Groups 10 and 18 drank significantly less than
Group 0. On Trials 3 and 4, all drug-treated F344
subjects differed from vehicle-treated subjects. On Trial
4, Group 10 drank less than Group 3.2. LEW subjects
did not differ in saccharin consumption at any point
over conditioning. On all trials, F344 subjects con-
sumed significantly less saccharin than LEW subjects.
Tukey’s HSD revealed that on Trial 1, F0 and F3.2
drank significantly less saccharin than L0 and L3.2,
respectively, with Groups F10 and F18 drinking compa-
rable levels of saccharin relative to Groups L10 and
L18. On Trial 2, Groups F0 and F3.2 did not differ
from Groups L0 and L3.2, while Groups F10 and F18
drank significantly less saccharin than Groups L10 and
L18. On Trials 3 and 4, Groups F0 and L0 did not
differ, while all drug-treated F344 subjects consumed
significantly less saccharin than their respective drug-
treated LEW subjects (see Fig. 3).
Over conditioning, Group F0 displayed a significant
increase in saccharin intake from Trials 1 to 4 [t
(8)¼ 5.812]. Group F3.2 [t (7)¼ 3.036] exhibited no
significant changes in saccharin consumption, while
Group F10 [t (7)¼ 10.583] and Group F18 [t
(7)¼ 11.239] significantly decreased saccharin intake
over trials. Group L0 [t (8)¼ 3.440] exhibited a
significant increase in saccharin consumption over trials,
while Groups L3.2 [t (7)¼ 1.843], L10 [t (8)¼ .000],
and L18 [t (9)¼ 1.467] did not display any significant
changes in saccharin intake from Trials 1 to 4.
Two-Bottle Test. On the two-bottle test, the percent
saccharin consumed by morphine-injected F344 sub-
jects was less than that of LEW subjects. The 2
(Replicate)� 2 (Strain)� 4 (Dose) univariate ANOVA
on the percent saccharin consumed revealed a signifi-
cant 3-way interaction [F (3, 52)¼ 3.349]. Subsequent
2 (Strain)� 4 (Dose) univariate ANOVAs on each
replicate revealed significant effects of Dose and Strain
as well as a significant Dose� Strain interaction
(Replicate One: [F (3, 22)¼ 19.348]; [F (1,
22)¼ 121.513]; [F (3, 22)¼ 5.120]; Replicate Two: [F
(3, 30)¼ 9.485]; [F (1, 30)¼ 14.727]; [F (3,
30)¼ 13.578]; respectively). Subsequent one-way
A B
FIGURE 2 Mean (�SEM) percent saccharin consumed by F344 (A) and LEW (B) adults.�Significant differences between Group 0 and Groups 3.2, 10 and 18. ��Significant differencesbetween Group 0 and Group 18.
Developmental Psychobiology Prepubertal Strain Differences in Morphine CTA 983

ANOVAs on the percent saccharin consumed in Repli-
cate One indicated significant differences between
groups [F (7, 29)¼ 31.796]. Tukey’s HSD indicated
that Groups F0 and L0 did not differ while all drug-
treated F344 subjects drank a significantly lower
percentage of saccharin than drug-treated LEW sub-
jects. The one-way ANOVA on Replicate Two also
indicated significant differences between groups [F (7,
37)¼ 12.181]. Subsequent use of Tukey’s HSD on
Replicate Two indicated Groups F0 and L0, F3.2 and
L3.2, and F10 and L10 did not differ while F18
drank a significantly lower percentage of saccharin than
Group L18.
Given that for each replicate, the general effect was
similar (i.e., F344>LEW), data from each replicate
were pooled for presentation (see Fig. 4).
DISCUSSION
Given that strain differences have been reported to vary
over development for a number of rodent strains and
that robust differences have been observed in taste
avoidance learning induced by a variety of drugs of
abuse in outbred prepubertal and adult rats (see above),
Experiment 2 assessed if the often-reported differences
in morphine-induced avoidance learning in adult F344
and LEW rats are also evident in prepubescence. As
described, prepubescent F344 rats acquired the avoid-
ance at a faster rate and to a stronger degree than
animals in the LEW strain, a pattern that paralleled the
results of Experiment 1 with adults under the modified
deprivation procedure and those previously reported
with adults in these strains (Davis et al., 2012; Gomez-
Serrano et al., 2009; Lancellotti et al., 2001). The
present results demonstrate that differences in mor-
phine-induced taste avoidance are evident as early as
prepubescence and are developmentally stable, suggest-
ing that these differences are highly heritable (for other
developmental strain assessments, see Allam, 2012;
Fairless et al., 2012; Farid et al., 2000; Moore
et al., 2011, 2013; Paylor et al., 1996; Satinder, 1981;
Sinaiko & Mirkin, 1974; Tonkiss et al., 1992; Vogl
et al., 1994; Wilking et al., 2012).
A B
FIGURE 4 Mean (�SEM) percent saccharin consumed by F344 (A) and LEW (B)
prepubescents. �Significant differences between Group 0 and Groups 3.2, 10, and 18.
A B
FIGURE 3 Mean (�SEM) saccharin consumption (ml) by F344 (A) and LEW (B)
prepubescents during acquisition. #Significant differences between Group 0 and Groups 10 and
18. �Significant differences between Group 0 and Groups 3.2, 10 and 18. ^Significant differences
between Group 3.2 and Group 10.
984 Hurwitz et al. Developmental Psychobiology

Although strain differences are clearly evident in
morphine-induced taste avoidance as early as prepubes-
cence, the basis for these differences are unknown. One
obvious possibility is that the two strains may differ in
blood and/or brain levels of morphine. Interestingly, mor-
phine plasma levels do not differ between adult F344 and
LEW rats (Davis & Riley, 2007; Gosnell & Krahn, 1993;
Lancellotti et al., 2001), although one report has demon-
strated that F344 animals have higher brain morphine
levels relative to LEW animals 30min following mor-
phine injection (Gosnell & Krahn, 1993). It remains
unknown whether adolescents show a similar pattern of
peripheral and/or central distribution and how that may
relate to strain differences in avoidance learning.
Given that the F344 strain displays greater stress
reactivity then the LEW strain (Dhabhar, McEwen, &
Spencer, 1993; Dhabhar, Miller, McEwen, & Spencer,
1995; Sternberg et al., 1992; for a review see Kosten &
Ambrosio, 2002), the reported strain differences in
morphine-induced taste avoidance could be due to
potential stress associated with the specific procedures
utilized in conditioning taste avoidance in the present
experiments, for example, deprivation, handling, injec-
tion. Despite the differential stress reactivity in these
strains (F344>LEW), work assessing the effects of
stress on the acquisition and expression of taste avoid-
ance in outbred subjects is mixed, typically with the
results indicating no direct relationship between stress
and the degree of taste avoidance conditioned (Ander-
son, Hinderliter, & Misanin, 2006; Bourne, Calton,
Gustavson, & Schachtman, 1992; Bowers, Amit, &
Gringras, 1996; Misanin, Kaufhold, Paul, Hinderliter, &
Anderson, 2006; Revusky & Reilly, 1989). It is impor-
tant to note that the vast majority of this work has been
done in adults, precluding any conclusions as to how
stress might differentially impact prepubescent F344
and LEW subjects. Interestingly, in the one direct
assessment of the effects of different stressors (isolation
housing, restraint stress) on the acquisition of ethanol-
induced taste avoidance in outbred prepubertal and adult
rats stress had no significant effect on ethanol-induced
taste avoidance in either age group (Anderson
et al., 2010).
It is possible that differences in taste, learning and
memory processing mediate the differences between
the F344 and LEW strains. These are unlikely, howev-
er, given that the reported differences between the two
strains are highly dependent upon the specific drug
examined. For example, although F344 rats display
greater morphine-induced taste avoidance than LEW
rats, the difference is reversed for cocaine (i.e., LEW>F344; Davis & Riley, 2007; Glowa et al., 1994). For
other compounds such as LiCl (Foynes & Riley, 2004),
there are no strain differences. The most parsimonious
explanation for the present results is that morphine is
differentially aversive in these strains. What accounts
for this differential sensitivity, however, remains un-
known (for a discussion of the nature of avoidance
learning, see Verendeev and Riley, 2012), although
when c-fos activity in the brainstem is examined in the
two strains, cellular activity in avoidance-related areas
(Grabus, Glowa, & Riley, 2004) are differentially
activated following morphine and in ways that parallel
the ability of morphine to induce aversions in the two
strains, that is, morphine which induces greater avoid-
ance in the F344 rat induces significantly greater
activity in these areas than that seen in the LEW strain.
Such assessments are limited to a few drugs and in
adults, so it is again unclear if these results generalize
to other ages.
The present data are suggestive of highly heritable
behavioral differences between the two strains; howev-
er, they do not argue that such differences cannot be
impacted by a host of environmental challenges. In
fact, a number of behavioral differences between the
strains are impacted by stress (Grakalic et al., 2006;
Siviy et al., 2003), diurnal cycle (Gomez-Serrano et al.,
2009), and maternal rearing (Gomez-Serrano et al.,
2001, 2002; Gomez-Serrano & Riley, 2006; Riley,
2011; Roma, Davis, & Riley, 2007; Siviy et al., 2003).
Further, it is unknown to what degree prenatal experi-
ence impacts the behavioral differences reported here.
What is clear is that the differences in morphine-
induced taste avoidance learning in these two strains
are evident early in life and are developmentally stable.
Further assessments are needed to determine the point
at which these behavioral differences become evident
and the basis for this stability.
NOTES
This work was supported in part by grants from the Mellon
Foundation to ALR and ZEH and the American University
Artists and Scholars Fellowship to APM.
REFERENCES
Allam, A. A. (2012). The development of sensorimotor
reflexes in albino mice; albino rats and black-hooded rats.
International Journal of Developmental Neuroscience, 30
(7), 545–553.
Ambrosio, E., Goldberg, S., & Elmer, G. (1995). Behavior
genetic investigation of the relationship between spontane-
ous locomotor activity and the acquisition of morphine
self-administration behavior. Behavioural Pharmacology, 6
(3), 229–237.
Developmental Psychobiology Prepubertal Strain Differences in Morphine CTA 985

Anderson, M. J., Hinderliter, C. F., & Misanin, J. R.
(2006). The effects of chronic water deprivation on
metabolic rate and long-trace taste-aversion conditioning
in rats. Neurobiology of Learning and Memory, 85(3),
199–205.
Anderson, R. I., Agoglia, A. E., Morales, M., Varlinskaya, E.
I., & Spear, L. P. (2012). Stress, kappa manipulations, and
aversive effects of ethanol in adolescent and adult male
rats. Neuroscience, 249, 214–222.
Anderson, R. I., Varlinskaya, E. I., & Spear, L. P. (2010).
Ethanol-induced conditioned taste aversion in male
Sprague Dawley rats: Impact of age and stress. Alcohol-
ism: Clinical and Experimental Research, 34(12), 2106–
2115.
Beitner-Johnson, D., Guitart, X., & Nestler, E. J. (1991).
Dopaminergic brain reward regions of Lewis and Fischer
rats display different levels of tyrosine hydroxylase and
other morphine-and cocaine-regulated phosphoproteins.
Brain Research, 561(1), 147–150.
Bourne, M. J., Calton, J. L., Gustavson, K. K., & Schacht-
man, T. R. (1992). Effects of acute swim stress on LiCl-
induced conditioned taste aversions. Physiology & Behav-
ior, 51(6), 1227–1234.
Bowers, W., Amit, Z., & Gringras, M. (1996). Time-
dependent exacerbation of amphetamine-induced taste
aversions following exposure to footshock. Psychopharma-
cology, 125(1), 43–49.
Cobuzzi, J. L., Siletti, K. A., Hurwitz, Z. E., Wetzell, B.,
Baumann, M. H., & Riley, A. L. (2013). Age differences
in (�) 3,4-methylenedioxymethamphetamine (MDMA)-
induced conditioned taste aversions and monoaminergic
levels. Developmental Psychobiology, DOI: 10.1002/
dev.21132
Davis, C. M., Cobuzzi, J. L., & Riley, A. L. (2012).
Assessment of the aversive effects of peripheral mu opioid
receptor agonism in Fischer 344 and Lewis rats. Pharma-
cology, Biochemistry, and Behavior, 101(2), 181–186.
Davis, C. M., & Riley, A. L. (2007). The effects of cocaine
preexposure on cocaine-induced taste aversion learning in
Fischer and Lewis rat strains. Pharmacology, Biochemis-
try, and Behavior, 87(1), 198–202.
Davis, C. M., Roma, P. G., Dominguez, J. M., & Riley, A. L.
(2007). Morphine-induced place conditioning in Fischer
and Lewis rats: Acquisition and dose-response in a fully
biased procedure. Pharmacology, Biochemistry, and Be-
havior, 86(3), 516–523.
Dhabhar, F. S., McEwen, B. S., & Spencer, R. L. (1993).
Stress response, adrenal steroid receptor levels and corti-
costeroid-binding globulin levels—A comparison between
Sprague-Dawley, Fischer 344 and Lewis rats. Brain
Research, 616(1), 89–98.
Dhabhar, F. S., Miller, A. H., McEwen, B. S., & Spencer, R.
L. (1995). Effects of stress on immune cell distribution.
Dynamics and hormonal mechanisms. The Journal of
Immunology, 154(10), 5511–5527.
Doremus-Fitzwater, T. L., Varlinskaya, E. I., & Spear, L. P.
(2010). Motivational systems in adolescence: Possible
implications for age differences in substance abuse and
other risk-taking behaviors. Brain and Cognition, 72(1),
114–123.
Fairless, A. H., Katz, J. M., Vijayvargiya, N., Dow, H. C.,
Kreibich, A. S., Berrettini, W. H., … Brodkin, E. S.
(2012). Development of home cage social behaviors in
BALB/cJ vs. C57BL/6J mice. Behavioural Brain Research,
237, 338–347.
Farid, M., Martinez, Z. A., Geyer, M. A., & Swerdlow, N. R.
(2000). Regulation of sensorimotor gating of the startle
reflex by serotonin 2A receptors: Ontogeny and strain
differences. Neuropsychopharmacology, 23(6), 623–632.
Foynes, M., & Riley, A. (2004). Lithium-chloride-induced
conditioned taste aversions in the Lewis and Fischer 344
rat strains. Pharmacology, Biochemistry, and Behavior, 79,
303–308.
Garcıa-Lecumberri, C., Torres, I., Martın, S., Crespo, J. A.,
Miguens, M., Nicanor, C., … Ambrosio, E. (2011). Strain
differences in the dose–response relationship for morphine
self-administration and impulsive choice between Lewis
and Fischer 344 rats. Journal of Psychopharmacology, 25
(6), 783–791.
Glowa, J. R., Shaw, A. E., & Riley, A. L. (1994). Cocaine-
induced conditioned taste aversions: Comparisons between
effects in LEW/N and F344/N rat strains. Psychopharma-
cology, 114(2), 229–232.
Gomez-Serrano, M., & Riley, A. L. (2006). The effects of
cross-fostering on morphine-induced conditioned taste
aversions in Fischer and Lewis rats (in preparation).
Gomez-Serrano, M., Tonelli, L., Listwak, S., Sternberg, E., &
Riley, A. L. (2001). Effects of cross fostering on open-
field behavior, acoustic startle, lipopolysaccharide-induced
corticosterone release, and body weight in Lewis and
Fischer rats. Behavior Genetics, 31(5), 427–436.
Gomez-Serrano, M. A., Kearns, D. N., & Riley, A. L. (2009).
The effects of light cycle phase on morphine-induced
conditioned taste aversions in the Lewis, Fischer and
Sprague-Dawley rat strains. Behavioural Brain Research,
196(1), 116–122.
Gomez-Serrano, M. A., Sternberg, E. M., & Riley, A. L.
(2002). Maternal behavior in F344/N and LEW/N rats:
Effects on carrageenan-induced inflammatory reactivity
and body weight. Physiology & Behavior, 75(4), 493–505.
Gosnell, B. A., & Krahn, D. D. (1993). Morphine-induced
feeding: A comparison of the Lewis and Fischer 344
inbred rat strains. Pharmacology, Biochemistry, and Be-
havior, 44(4), 919–924.
Grabus, S. D., Glowa, J. R., & Riley, A. L. (2004). Morphine-
and cocaine-induced c-Fos levels in Lewis and Fischer rat
strains. Brain Research, 998(1), 20–28.
Grakalic, I., Schindler, C. W., Baumann, M. H., Rice, K. C.,
& Riley, A. L. (2006). Effects of stress modulation on
morphine-induced conditioned place preferences and plas-
ma corticosterone levels in Fischer, Lewis, and Sprague–
Dawley rat strains. Psychopharmacology, 189(3), 277–
286.
Guitart, X., Kogan, J. H., Berhow, M., Terwilliger, R. Z.,
Aghajanian, G. K., & Nestler, E. J. (1993). Lewis and
Fischer rat strains display differences in biochemical,
986 Hurwitz et al. Developmental Psychobiology

electrophysiological and behavioral parameters: Studies in
the nucleus accumbens and locus coeruleus of drug naive
and morphine-treated animals. Brain Research, 611(1), 7–
17.
Hurwitz, Z. E., Merluzzi, A. P., & Riley, A. L. (2013).
Age-dependent differences in morphine-induced taste
aversions. Developmental Psychobiology, 55(4), 415–
428.
Infurna, R. N., & Spear, L. P. (1979). Developmental changes
in amphetamine-induced taste aversions. Pharmacology,
Biochemistry, and Behavior, 11(1), 31–35.
Kosten, T. A., & Ambrosio, E. (2002). HPA axis function and
drug addictive behaviors: Insights from studies with Lewis
and Fischer 344 inbred rats. Psychoneuroendocrinology,
27(1), 35–69.
Kosten, T. A., Miserendino, M. J., Haile, C. N., DeCaprio, J.
L., Jatlow, P. I., & Nestler, E. J. (1997). Acquisition and
maintenance of intravenous cocaine self-administration in
Lewis and Fischer inbred rat strains. Brain Research, 778
(2), 418–429.
Lancellotti, D., Bayer, B. M., Glowa, J. R., Houghtling, R.
A., & Riley, A. L. (2001). Morphine-induced conditioned
taste aversions in the LEW/N and F344/N rat strains.
Pharmacology, Biochemistry, and Behavior, 68(3), 603–
610.
Martın, S., Lyupina, Y., Crespo, J. A., Gonzalez, B., Garcıa-
Lecumberri, C., & Ambrosio, E. (2003). Genetic differ-
ences in NMDA and D1 receptor levels, and operant
responding for food and morphine in Lewis and Fischer
344 rats. Brain Research, 973(2), 205–213.
Martın, S., Manzanares, J., Corchero, J., Garcıa-Lecumberri,
C., Crespo, J. A., Fuentes, J. A., & Ambrosio, E.
(1999). Differential basal proenkephalin gene expression
in dorsal striatum and nucleus accumbens, and vulnerabili-
ty to morphine self-administration in Fischer 344 and
Lewis rats. Brain Research, 821(2), 350–355.
Misanin, J. R., Anderson, M. J., & Hinderliter, C. F. (2009).
Conditioned taste aversion across the life span from
prenascence to senescence. In S. Reilly & T. R. Schacht-
man (Eds.), Conditioned taste aversion: Behavioral and
neural processes (pp. 281). New York, New York: Oxford
University Press.
Misanin, J. R., Kaufhold, S. E., Paul, R. L., Hinderliter, C. F.,
& Anderson, M. J. (2006). A time contraction effect of
acute tail-pinch stress on the associative learning of rats.
Behavioural Processes, 71(1), 16–20.
Moore, E. M., Forrest, R., & Boehm, S. (2013). Genotype
modulates age-related alterations in sensitivity to the
aversive effects of ethanol: An eight inbred strain analysis
of conditioned taste aversion. Genes, Brain and Behavior,
12(1), 70–77.
Moore, E. M., Linsenbardt, D. N., Melon, L. C., & Boehm, S.
L. (2011). Ontogenetic differences in adolescent and adult
C57BL/6J and DBA/2J mice: Anxiety-like, locomotor, and
consummatory behaviors. Developmental Psychobiology,
53(2), 141–156.
Paylor, R., Baskall-Baldini, L., Yuva, L., & Wehner, J. M.
(1996). Developmental differences in place-learning perfor-
mance between C57BL/6 and DBA/2 mice parallel the
ontogeny of hippocampal protein kinase C. Behavioral
Neuroscience, 110(6), 1415–1425.
Pescatore, K. A., Glowa, J. R., & Riley, A. L. (2005). Strain
differences in the acquisition of nicotine-induced condi-
tioned taste aversion. Pharmacology, Biochemistry, and
Behavior, 82(4), 751–757.
Revusky, S., & Reilly, S. (1989). Attenuation of conditioned
taste aversions by external stressors. Pharmacology, Bio-
chemistry, and Behavior, 33(1), 219–226.
Riley, A. L. (2011). The paradox of drug taking: The role of
the aversive effects of drugs. Physiology & Behavior, 103
(1), 69–78.
Roma, P. G., Davis, C. M., & Riley, A. L. (2007). Effects of
cross-fostering on cocaine-induced conditioned taste aver-
sions in Fischer and Lewis rats. Developmental Psychobi-
ology, 49(2), 172–179.
Roma, P. G., Flint, W. W., Higley, J. D., & Riley, A. L.
(2006). Assessment of the aversive and rewarding effects
of alcohol in Fischer and Lewis rats. Psychopharmacology,
189(2), 187–199.
Satinder, K. P. (1981). Ontogeny and interdependence of
genetically selected behaviors in rats: Avoidance response
and open field. Journal of Comparative and Physiological
Psychology, 95(1), 175–187.
Schramm-Sapyta, N. L., Cha, Y. M., Chaudhry, S., Wilson,
W. A., Swartzwelder, H. S., & Kuhn, C. M. (2007).
Differential anxiogenic, aversive, and locomotor effects of
THC in adolescent and adult rats. Psychopharmacology,
191(4), 867–877.
Schramm-Sapyta, N. L., DiFeliceantonio, A. G., Foscue, E.,
Glowacz, S., Haseeb, N., Wang, N., … Kuhn, C. M.
(2010). Aversive effects of ethanol in adolescent versus
adult rats: Potential causes and implication for future
drinking. Alcoholism: Clinical and Experimental Re-
search, 34(12), 2061–2069.
Schramm-Sapyta, N. L., Morris, R. W., & Kuhn, C. M.
(2006). Adolescent rats are protected from the conditioned
aversive properties of cocaine and lithium chloride.
Pharmacology, Biochemistry, and Behavior, 84(2), 344–
352.
Shram, M. J., Funk, D., Li, Z., & Le, A. D. (2006).
Periadolescent and adult rats respond differently in tests
measuring the rewarding and aversive effects of nicotine.
Psychopharmacology, 186(2), 201–208.
Sinaiko, A., & Mirkin, B. L. (1974). Ontogenesis of the
renin-angiotensin system in spontaneously hypertensive
and normal Wistar rats. Circulation Research, 34(5), 693–
696.
Siviy, S. M., Love, N. J., DeCicco, B. M., Giordano, S. B., &
Seifert, T. L. (2003). The relative playfulness of juvenile
Lewis and Fischer-344 rats. Physiology & Behavior, 80
(2), 385–394.
Spear, L. P. (2013). Adolescent neurodevelopment. Journal of
Adolescent Health, 52(2), S7–S13.
Sternberg, E. M., Glowa, J. R., Smith, M. A., Cologero, A.
E., Listwak, S. J., Aksentijevich, S., … Gold, P. W.
(1992). Corticotropin releasing hormone related behavioral
Developmental Psychobiology Prepubertal Strain Differences in Morphine CTA 987

and neuroendocrine responses to stress in Lewis and
Fischer rats. Brain Research, 570(1), 54–60.
Tonkiss, J., Shultz, P., & Galler, J. R. (1992). Long-Evans and
Sprague-Dawley rats differ in their spatial navigation
performance during ontogeny and at maturity. Develop-
mental Psychobiology, 25(8), 567–579.
Verendeev, A., & Riley, A. (2012). Conditioned taste aversion
and drugs of abuse: History and interpretation. Neurosci-
ence & Biobehavioral Reviews, 36(10), 2193–2205.
Vetter-O’Hagen, C., Varlinskaya, E., & Spear, L. (2009). Sex
differences in ethanol intake and sensitivity to aversive
effects during adolescence and adulthood. Alcohol and
Alcoholism, 44(6), 547–544.
Vogl, C., Atchley, W. R., & Xu, S. (1994). The ontogeny of
morphological differences in the mandible in two inbred
strains of mice. Journal of Craniofacial Genetics and
Developmental Biology, 14(2), 97–110.
Wilking, J. A., Hesterberg, K. G., Nguyen, V. H., Cyboron,
A. P., Hua, A. Y., & Stitzel, J. A. (2012). Comparison of
nicotine oral consumption and baseline anxiety measures
in adolescent and adult C57BL/6J and C3H/Ibg mice.
Behavioural Brain Research, 233(2), 280–287.
Wilmouth, C. E., & Spear, L. P. (2004). Adolescent and adult
rats’ aversion to flavors previously paired with nicotine.
Annals of the New York Academy of Sciences, 1021(1),
462–464.
988 Hurwitz et al. Developmental Psychobiology