pulmonary structural and extracellular matrix alterations in fischer 344 rats following subchronic...
TRANSCRIPT

FUNDAMENTAL AND APPLIED TOXICOLOGY 37 , 5 4 - 6 3 (1997)
ARTICLE NO FA972298
Pulmonary Structural and Extracellular Matrix Alterations in Fischer 344Rats Following Subchronic Phosgene Exposure12
Urmila P. Kodavanti, Daniel L. Costa, Shri N. Giri,* Barry Starcher.f and Gary E. Hatch
Pulmonary Toxicology Branch, Experimental Toxicology Division, National Health and Environmental Effects Research Laboratory,U.S. Environmental Protection Agency, Research Triangle Park, North Carolina 27711; 'Department of Veterinary Molecular Biosciences,
School of Veterinary Medicine, University of California at Davis, Davis, California 95616; and ^Department of Biochemistry,The University of Texas Health Center at Tyler, Tyler, Texas 75710
Received August 20, 1996; accepted February 10, 1997
Pulmonary Structural and Extracellular Matrix Alterations inFischer 344 Rats Following Subchronic Phosgene Exposure. Koda-vanti, U. P., Costa, D. L., Giri, S. N., Starcher, B., and Hatch,G. E. (1997). Fundam. Appl. Toxicol. 37, 54-63.
Phosgene, an acylating agent, is a very potent inducer of pulmo-nary edema. Subchronic effects of phosgene in laboratory animalsare not well characterized. The purpose of the study was to eluci-date potential long-term effects on collagen and elastin metabolismduring pulmonary injury/recovery and obtain information aboutthe concentration X time (C X T) behavior of low levels of phos-gene. Male Fischer 344 rats (60 days old) were exposed either toclean air or phosgene, 6 hr/day: 0.1 ppm (5 days/week), 0.2 ppm(5 days/week), 0.5 ppm (2 days/week), and 1.0 ppm (1 day/week),for 4 or 12 weeks. A group of rats was allowed clean air recoveryfor 4 weeks after 12 weeks of phosgene exposure. This exposurescenario was designed to provide equal C x T product for allconcentrations at one particular time point except for 0.1 ppm(50% C X T). Phosgene exposure for 4 or 12 weeks increasedlung to body weight ratio and lung displacement volume in aconcentration-dependent manner. The increase in lung displace-ment volume was significant even at 0.1 ppm phosgene at 4 weeks.Light microscopic level histopathology examination of lung wasconducted at 0.0, 0.1, 0.2, and 1.0 ppm phosgene following 4 and12 and 16 weeks (recovery). Small but clearly apparent terminalbronchiolar thickening and inflammation were evident with 0.1ppm phosgene at both 4 and 12 weeks. At 0.2 ppm phosgene,terminal bronchiolar thickening and inflammation appeared to bemore prominent when compared to the 0.1 ppm group and changesin alveolar parenchyma were minimal. At 1.0 ppm, extensive in-flammation and thickening of terminal bronchioles as well as alve-olar walls were evident. Concentration rather than C X T seems
to drive pathology response. Trichrome staining for collagen atthe terminal bronchiolar sites indicated a slight increase at 4 weeksand marked increase at 12 weeks in both 0.2 and 1.0 ppm groups(0.5 ppm was not examined), 1.0 ppm being more intense. Whole-lung prolyl hydroxylase activity and hydroxyproline, taken as anindex of collagen synthesis, were increased following 1.0 ppm phos-gene exposure at 4 as well as 12 weeks, respectively. Desmosinelevels, taken as an index of changes in elastin, were increased inthe lung after 4 or 12 weeks in the 1.0 ppm phosgene group.Following 4 weeks of air recovery, lung hydroxyproline was furtherincreased in 0.5 and 1.0 ppm phosgene groups. Lung weight alsoremained significantly higher than the controls; however, desmo-sine and lung displacement volume in phosgene-exposed animalswere similar to controls. In summary, terminal bronchiolar andlung volume displacement changes occurred at very low phosgeneconcentrations (0.1 ppm). Phosgene concentration, rather than CX T product appeared to drive toxic responses. The changes in-duced by phosgene (except of collagen) following 4 weeks werenot further amplified at 12 weeks despite continued exposure.Phosgene-induced alterations of matrix were only partially revers-ible after 4 weeks of clean air exposure, c 1997 sod«y or To
1 Preliminary results have been presented at the 1994 American ThoracicSociety Meetings in Boston, MA, and published as an abstract in Am. J.Respir. Crit. Care Med. 149(4, Part 2), A159 (1994).
2 The research described in this article has been reviewed by the NationalHealth and Environmental Effects Research Laboratory, U.S. Environmen-tal Protection Agency, and approved for publication. Approval does notsignify that the contents necessarily reflect the views and the policies of theAgency nor does mention of trade names or commercial products constituteendorsement or recommendation for use.
0272-0590/97 $25.00 54Copyright © 1997 by the Society of Toxicology.All rights of reproduction in any form reserved.
Phosgene, a widely used intermediate in the synthesis ofchemicals and plastics, is a highly reactive acylating agent(chemical formula: COC12). It is known to induce noncardio-genic pulmonary edema upon acute inhalation exposure(Frosolono and Pawlowski, 1977; Pawlowski and Frosolono,1977; Diller et al., 1985). Although environmental levelsare relatively low, reaching to about 60 parts per trillion inthe urban areas such as Los Angeles, occupational hazardsresulting from accidental exposure are well known (Galdstonet al., 1947; Gerritsen and Buschmann, 1960; Sjogren et al.,1991). Likewise, emissions from industrial facilities maypose episodic risks to the public living nearby (Davis et al.,1989). The acute toxicity of phosgene has been reported to,involve oxidative tissue damage and pulmonary edema dueto increased vascular permeability (Franch and Hatch, 1986;Kennedy et al., 1989), which may involve arachidonic acidmetabolites (Guo et al., 1990). Exposure to phosgene is alsoknown to induce pulmonary inflammation in rats (Currie
by guest on July 15, 2011toxsci.oxfordjournals.org
Dow
nloaded from

SUBCHRONIC PULMONARY EFFECTS OF PHOSGENE 55
et al., 1987). Impaired alveolar macrophage function andclearance of microorganisms from the lung as a result ofsubchronic phosgene exposure have also been reported (Sel-grade et al., 1995). At lower concentrations, phosgene ap-pears to preferentially deposit in and affect terminal bronchi-oles, whereas at higher concentrations phosgene damageclearly extends to include alveolar epithelial cells (Grosset al., 1965; Pawlowski and Frosolono, 1977). It has beensuggested that the mechanism of action of phosgene is simi-lar to that of ozone and other oxidants (Kennedy et al., 1989;Ghio et al., 1991).
Although residual chronic lung injury has been noted inhumans accidentally exposed to phosgene (Cucinell and Ar-senal, 1974), long-term, low-level chronic effects are largelyunknown. Indeed, systematic chronic toxicity studies in ro-dents using low levels of this common fugitive emissionhave not been conducted. Pulmonary emphysema and oblit-erative bronchitis have been reported in dogs following high-level phosgene exposure (Clay and Rossing, 1964). Wehypothesized that while high-level phosgene exposures areassociated with destructive emphysema in humans and ani-mals, low-level, longer-term phosgene exposures will affectthe structural integrity and induce remodeling of the rat lungconnective tissues.
Because phosgene is known to be emitted by industrialactivities, both protracted low-level and incidental high-levelreleases into the air are expected; therefore, there is a regula-tory need to assess the relative importance of these potentialexposure scenarios. It has been proposed that physiologicaland toxic effects of phosgene exposure are proportional tothe concentration X time (C X 7) product (Rinehart andHatch, 1964). A primary objective of this study was to inves-tigate the relationship between toxicity and the C X T prod-uct in a subchronic format to ascertain the relative impor-tance of peak concentration, frequency of exposure, and totaldose to the lesion observed. We report here that pulmonarychanges were observed at phosgene concentrations as lowas 0.1 ppm, which is the current TLV. Also, it appears thatthe phosgene-induced changes were primarily determinedby concentration rather than C XT product. Moreover, theselesions were only partially reversible, suggesting that phos-gene exposures even at low levels may be deleterious to thelung.
METHODS
Animals. Specific pathogen-free male Fischer-344 (F344) rats,CDF*(F344)/CiiBr (260-300 g, 60 days old), were purchased from CharlesRiver Breeding Laboratories (Kingston, NY). The animals were housed intemperature- and humidity-controlled (72 ± 2°F and 50 ± 5% relativehumidity), AAALAC-approved animal facilities with a 12-hr light/darkcycle and were fed certified rat chow (Agway Inc. Country Foods Division,Syracuse, New York) ad libitum. A group of rats was tested for presenceof specific pathogen at the time of arrival, at the start of exposure, andperiodically thereafter until the end of experiment, and they were found tobe specific pathogen free.
Phosgene exposure. Rats were housed in large 2.0-m3 Hazelton 2000(0.0, 0.1, 0.2, and 0.5 ppm groups) and small 0.3-m3 Rochester inhalationchambers (0.0 and 1.0 ppm groups) (approved by AAALAC) during phos-gene exposure. Animals housed in Hazelton chambers remained in thechamber during nonexposure periods; however, animals housed in Roches-ter inhalation chambers were relocated to the animal colony. No significantdifferences in body weight gain (large chambers vs small chambers; 4weeks, 251.5 ± 17.9 vs 244.7 ± 10.5; 12 weeks, 319.2 ± 11.9 vs 309.7± 13.3; 16 weeks, 355.3 ±23.5 vs 347.1 ± 14.5) or in any other parametersoccurred between two different control groups housed in two differentsettings. Following 12 weeks of phosgene exposure all recovery groupanimals were housed in animal colony for 4 weeks of recovery period.Animals were randomly assigned to different groups using a computer-based weight-matched randomization program. A mass-flow controller (Ty-lan, Torrance, CA) metered phosgene (300 ppm in nitrogen; Matheson GasProducts, Morrow, GA), with an airflow of 1.0 m3/min for Hazelton cham-bers and 0.32 mVmin for Rochester inhalation chambers. The gas mixturepassed downward through the cages holding the test animals and was ex-hausted at the bottom into a water scrubber. Temperature and humidity inthe chambers as well as in the animal colony were (mean ± SD) 23.0 ± 4°Cand 50 ± 10%, respectively. Control animals were placed simultaneously inan identical chamber and allowed to breath filtered air.
Phosgene concentrations in the chambers were determined with Hewlett-Packard 5880 and 5840 gas chromatographs. Methane (5%) in argon (P-5) was used as the carrier phase. For Model 5880, a fused silica capillarycolumn, SPB-1, was used (Supelco Inc., Bellefonte, PA) with a flow rateof 30.0 ml/min. The column temperature was maintained at 35°C. ForModel 5840, a 10% SP-2100, 80/100 Supelcoport SS 1/8" column (Supelco,Bellefonte, PA) was used with a flow rate of 30 ml/min. Column temperaturewas maintained at 40°C during operation. Prior to the start of phosgeneexposure, an appropriate representative sampling port was determined byanalyzing samples collected at various locations within the chamber. Duringexposure, phosgene concentrations were monitored once every 20 min froma representative sampling port. The 12-week average chamber phosgeneconcentrations were (mean ± SD (range)) for 0.1 ppm = 0.101 ± 0.002(0.098-0.113), 0.2 ppm = 0.201 ± 0.002 (0.196-0.207), 0.5 ppm = 0.505± 0.008 (0.495-0.536), and 1.0 ppm = 0.976 ± 0.03 (0.912-1.009).
Rats were exposed to either air or phosgene, 6 hr/day at 0.1 (5 days/week), 0.2 (5 days/week), 0.5 (2 days/week), or 1.0 ppm (I day/week) inorder to derive equal C X T product for one particular time point exceptin the 0.1 ppm group (50% C X T). The experimental protocol and numberof animals used in each group are depicted in Table 1. At the end of 4 or12 weeks of phosgene exposure, or 4 weeks of clean air recovery after 12weeks of phosgene exposure, rats were killed and the lungs analyzed.
Lung fixation and displacement volume measurements. Rats were an-esthetized with urediane (1.0 g/ml/kg body wt), the abdomen was opened,blood was removed by exsanguination of the dorsal aorta, and the tracheawas cannulated. The left lung was used for histopathology and the rightlung was used for biochemical analysis. The right lung was tied, and thenthe lung lobes were clipped off and weighed. Following lung weight mea-surements, the lobes were quickly frozen in liquid nitrogen and stored at-80°C until analyzed. The trachea with the left lung attached was suspendedin a petri dish containing distilled water and inflated to 25 cm water transpul-monary pressure with 2.5% glutaraldehyde/0.16 M cacodylate buffer. Thirtymin postinflation, the lung was removed and the displacement volume wasobtained. The fixed tissues were then stored in 10% buffered formalin untilfurther processing.
Histopathology. Midsagittal sections (4 fim) of the lung tissue wereprepared from paraffin blocks and mounted on glass microscope slides.Slides were stained with Masson's Trichrome stain for identification ofcollagen fibers and counter stained with hematoxylin and eosin. Lung sec-tions were examined under a light microscope. Histopathological evaluationof these lung tissues was done in a blinded manner for exposure group designa-tion (Experimental Pathology Laboratory, Research Triangle Park, NQ.
by guest on July 15, 2011toxsci.oxfordjournals.org
Dow
nloaded from

56 KODAVANTI ET AL.
TABLE 1Phosgene Exposure Protocol and Concentration X Time Product
C x T product (ppm • hr)
Exposure 4 Weeks 12 Weeks
16 Weeks(12 weeks exposure +
4 weeks recovery)
Air control, large chambers'0.1 ppm, 6 hr/day, 5 days/week0.2 ppm, 6 hr/day, 5 days/week0.5 ppm, 6 hr/day, 2 days/week1.0 ppm, 6 hr/day, 1 day/weekAir control, small chambers
0 (/i = 8)"12 (n = 8)24 (n = 8)24 (n = 8)24 (n = 8)0 (/i = 4)
0 (n = 8)36 (n = 8)72 (n = 8)72 (n = 8)72 (n = 8)
0 (n = 4)
0 (n = 8)36 (n = 8)72 (n = 8)72 (n = 8)72 (n = 8)
0 (n = 4)
" Since large and small chambers were used for exposure to phosgene, two separate air control groups were run simultaneously.6 Number of animals used per group.
Biochemical assays. The right caudal lobe was used for hydroxyprolineand desmosine analysis and the rest of the lobes were used for prolylhydroxylase assay. The right lung lobes (except a caudal lobe) were thawedand homogenized in cold 20 mAf Tris—HC1 buffer, pH 7.6. Prolyl hydroxy-lase activity was measured as the enzyme-catalyzed release of tritiatedwater from L-[3,4-3H]proline-labeled procollagen (Gin el ah, 1983). Forhydroxyproline and desmosine analysis the caudal lobes were lyophilizedand then hydrolyzed in 6 M hydrochloric acid at 110°C for 16 hr. Thehydrolyzate was filtered through 0.45-^m Teflon filters. A portion of thefiltrate was then neutralized using 6 M sodium hydroxide to a pH of 7-7.5and the samples were diluted for calorimetric detection of hydroxyprolineas described (Bergman and Loxley, 1961). An aliquot of filtrate was driedunder nitrogen. The residue was resuspended in water and dried in orderto completely eliminate hydrochloric acid from the sample. The dried sam-ples were then suspended in water and analyzed for desmosine by radioim-munoassay (Starcher, 1982).
Statistics. A multivariate analysis of variance (MANOVA) (SAS 516;SAS Institute, Cary, NC) was used. According to the study protocol, ifsignificant (p as 0.1) multivanate time, exposure, or exposure by timeinteraction effects were found, further univariate analysis using the sameanalysis model as that used in the MANOVA were performed. Correctedpairwise comparisons were used as subtests to evaluate differences betweenexposure groups. The Type I error rate was set at p = 0 05 for significance.Control values (air chambers) for large and small chambers were pooledbecause no statistical differences were noted between the animals exposedin two chambers in any of the parameters measured at any time. Datarepresents the mean ± SE of 10-12 controls or 8 exposed rats.
RESULTS
Lung Weight and Displacement Volume Changes
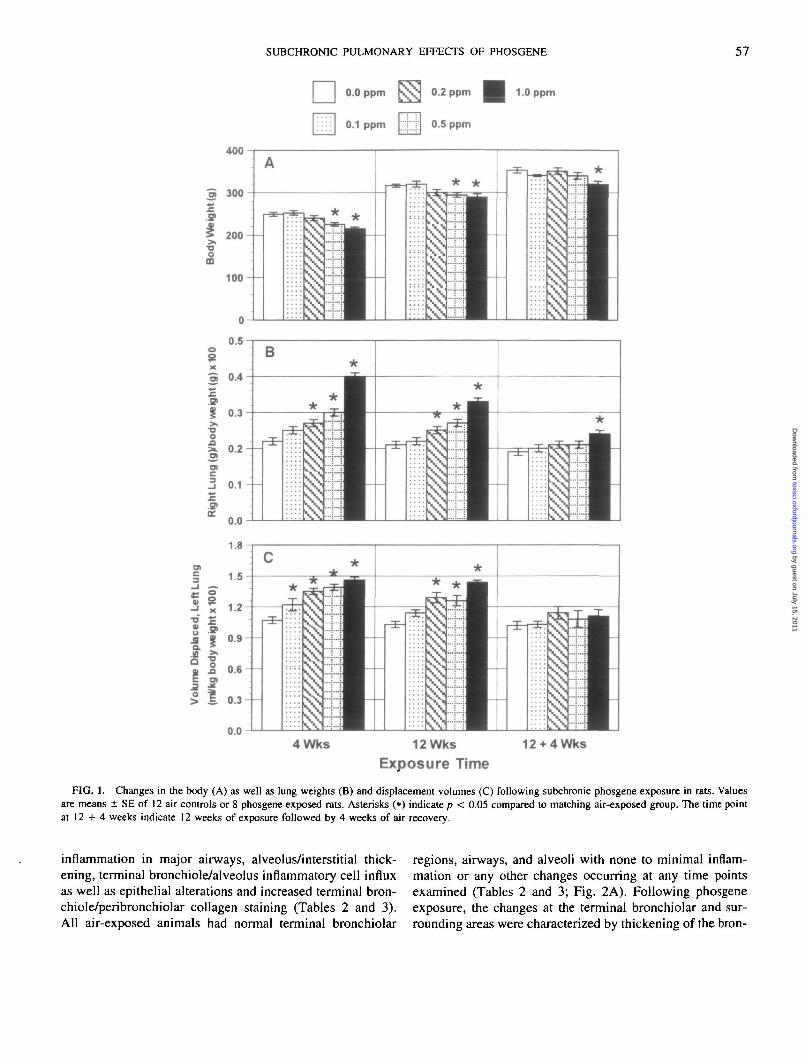
There was a small but significant decrease in weight gainin rats exposed to 0.5 and 1.0 ppm phosgene at 4 and 12weeks (Fig. 1 A); therefore, lung to body weight ratios weretaken into account in determining the effect on the lungweight. Lung to body weight ratios were increased in aconcentration-dependent manner at 4 and 12 weeks of phos-gene exposure (Fig. IB). The phosgene concentration at ei-ther time point seemed to drive this response rather than theC X T product. The effect appeared to be more severe at
the end of 4 weeks than after 12 weeks of exposure. After 4weeks of clean air recovery following 12 weeks of phosgeneexposure, the lung to body weight ratio remained slightly,but significantly, higher in the 1.0 ppm phosgene-exposedanimals. Phosgene also increased the lung displacement vol-ume (measured for the left lung and normalized to bodyweight) in a concentration-dependent manner (Fig. 1C).Measurement of displacement volume is taken as an indexof change in the total lung volume (Scherle, 1970). Theeffect was significant even at 0.1 ppm at 4 weeks. The in-crease in displacement volume at 4 weeks was not furtheramplified following 12 weeks despite continued exposure.Following 4 weeks of clean air recovery, the displacementvolumes returned to control levels regardless of phosgeneconcentration.
Histopathology and Collagen Changes
Light microscopic examination was done only at 0.0, 0.1,0.2, and 1.0 ppm phosgene following 4 and 12 weeks ofexposure and recovery (the 0.5 ppm samples were inadver-tently lost). It was possible to compare pathology of twogroups with equal C X T (0.2 and 1.0 ppm) instead of three.Pulmonary histopathological changes were associated withphosgene exposure and were concentration dependent (Ta-bles 2 and 3). Phosgene appeared to affect all animals in aconsistent manner within a group. The changes occurredprimarily at the terminal bronchiolar regions at low phosgeneconcentrations (0.1 and 0.2 ppm). The more peripheral areaswere affected at 1.0 ppm. The severity of the pathologicalchanges in the lung seemed to depend upon phosgene con-centrations rather than the C X T product based on subjectivescreening of the degree of injury (Tables 2 and 3).
Four-week phosgene exposure. Controls for both cham-bers as well as phosgene-exposed animals were scored forseverity of lesions such as epithelial alterations including
by guest on July 15, 2011toxsci.oxfordjournals.org
Dow
nloaded from
SUBCHRONIC PULMONARY EFFECTS OF PHOSGENE 57
0.0 ppm hoOJ 0.2 ppm 1.0 ppm
0.1 ppm ;;jHj 0.5 ppm
400
4Wks 12Wks
Exposure Time12 + 4Wks
FIG. 1. Changes in the body (A) as well as lung weights (B) and displacement volumes (C) following subchronic phosgene exposure in rats. Valuesare means ± SE of 12 air controls or 8 phosgene exposed rats. Asterisks (•) indicate p < 0.05 compared to matching air-exposed group. The time pointat 12 + 4 weeks indicate 12 weeks of exposure followed by 4 weeks of air recovery.
inflammation in major airways, alveolus/interstitial thick-ening, terminal bronchiole/alveolus inflammatory cell influxas well as epithelial alterations and increased terminal bron-chiole/peribronchiolar collagen staining (Tables 2 and 3).All air-exposed animals had normal terminal bronchiolar
regions, airways, and alveoli with none to minimal inflam-mation or any other changes occurring at any time pointsexamined (Tables 2 and 3; Fig. 2A). Following phosgeneexposure, the changes at the terminal bronchiolar and sur-rounding areas were characterized by thickening of the bron-
by guest on July 15, 2011toxsci.oxfordjournals.org
Dow
nloaded from

58 KODAVANTI ET AL.
TABLE 2Histopathology Incidence Table Indicating the Number of Animals Affected Following Phosgene Exposure"
Phosgene concentration (ppm):
Number of animals examined
0.0
(12)
4
0.1
(8)
Weeks
0.2
(8)
1.0
(6)
0.0
(12)
12
0.1
(8)
Weeks
0.2
(8)
1.0
(8)
0.0
(9)
16
0.1
(6)
Weeks
0.2
(7)
1.0
(5)
Alveolar effusionAlveolus, Interstitial thickeningBronchus, epithelial alterationBronchus, inflammationTerminal bronchiole/alveolus
Inflammatory cell influxTerminal bronchiole, peribronchiolarAlveolus, epithelial alterationTerminal bronchiole/peribronchiolar
Increased collagen staining
0001
2
2
1
0212
3
4
1
0522
8
5
8
2623
6
6
6
0000
1
0
2
0200
3
1
2
0410
8
7
8
1811
8
8
8
2000
0
0
2
0000
0
2
1
0000
1
3
7
0000
1
2
5
° Number in each column indicate number of animals affected of total numbers used in analysis.
chioles as well as alveolar walls with infiltration of inflam-matory cells (Figs. 2B-2D). At 0.1 ppm phosgene, a smallbut readily apparent terminal bronchiolar thickening was no-ticed at the light microscopic level with an increased numberof inflammatory cells (Fig. 2B). Pathology severity scoringand incidence data indicated that changes seen at 0.1 ppmcould be distinguished from control groups (Tables 2 and3). These changes were much more apparent at 0.2 ppm,and seemed to be uniformly distributed in all terminal bron-chioles within the section. At 1.0 ppm phosgene, more severeinflammation and thickening were noticeable at the terminalbronchiolar regions and alveolar walls (Fig. 2D). There wasan increase in collagen stain within thickened terminal bron-chioles at 0.2 and 1.0 ppm (0.5 ppm animals were not exam-ined) following 4 weeks of exposure (Figs. 2C and 2D).The staining was much more intense at 1.0 ppm phosgene.Although there was an increase in collagen staining at termi-nal bronchiolar regions and surrounding alveolar sites, espe-cially at 1.0 ppm, fibrotic patches were less apparent withinthe alveolar parenchyma.
Twelve-week phosgene exposure. Despite continual ex-posure, the inflammatory cell influx at the terminal bronchio-lar area and surrounding pleura was not further increasedfollowing 12 weeks of exposure when compared to 4 weeksfor any phosgene concentrations examined (Tables 2 and 3;Figs. 2E and 2F). Slight terminal bronchiolar thickening wasnoted at 0.1 ppm with very few inflammatory cells (notshown). The intensity of collagen stain increased followingexposure for 12 weeks at both 0.2 and 1.0 ppm in the termi-nal bronchiolar region; Figs. 2E and 2F). At 1.0 ppm phos-gene, in addition to the terminal bronchiolar changes, therewas marked alveolar wall thickening and inflammation, giv-ing an appearance of rounded alveolar spaces (Fig. 2G).Fibrotic lesions involving multialveolar units were minimal
in rats exposed to 0.1 or 0.2 ppm and only slightly apparentat 1.0 ppm phosgene (not shown).
Twelve-week phosgene exposure plus 4-week air recovery.Histopathology following 4 weeks of clean air recovery (12+ 4 weeks groups) indicate almost complete resolution ofinflammation with residual inflammatory cells and epithelialcell alterations (Tables 2 and 3). The 0.1 ppm phosgenegroup appeared to be similar to air-exposed controls (notshown). Terminal bronchiolar thickening and inflammationin the 0.2 ppm phosgene-exposed animals appeared to haveresolved considerably (Fig. 2H). Collagen staining remainedat the same level of intensity as seen in 12-week groups of0.2 and 1.0 ppm (Tables 2 and 3).
Prolyl hydroxyla.se, hydroxyproline, and desmosine fol-lowing phosgene exposure. Hydroxyproline and prolyl hy-droxylase activity were measured in a lung lobe as a measureof increased collagen synthesis following phosgene expo-sure. Prolyl hydroxylase activity was significantly increasedover air controls at 1.0 ppm phosgene following either 4 or12 weeks of exposure. The increase was of similar magnitudeat either time point (Fig. 3A). Prolyl hydroxylase activityreturned to control levels by the end of the recovery period.Total hydroxyproline, measured as an index of collagen de-position, was significantly elevated at the 1.0 ppm phosgeneconcentration following 12 weeks of exposure (Fig. 3B).At 4 weeks (1.0 ppm phosgene), the hydroxyproline levelappeared to be higher than that in the air-exposed controls;however, the difference was Jiot statistically significant.Right lung hydroxyproline levels appeared to have furtherincreased following 4 weeks of air recovery (Fig. 3B). Theincrease at this time was significant at 0.5 and 1.0 ppmphosgene. Analysis of variance also indicated a significanttime related increases in hydroxyproline in the air exposed
by guest on July 15, 2011toxsci.oxfordjournals.org
Dow
nloaded from

SUBCHRONIC PULMONARY EFFECTS OF PHOSGENE 59
TABLE 3Pulmonary Histopathology Severity Score in Rats Following Subchronic Phosgene Exposure"
Phosgene concentration (ppm):
Number of animals examinedAlveolar effusionAlveolus, Interstitial thickeningBronchus, epithelial alterationBronchus, inflammationTerminal bronchiole/alveolus
Inflammatory cell influxTerminal bronchiole, peribronchiolar
Alveolus, epithelial alterationTerminal bronchiole/penbronchiolar
Increased collagen staining
0.0
(12)0000.08
0.17
0.17
0.08
4
0.1
(8)00.250.130.25
0.38
0.5
0.13
Weeks
0.2
(8)00.630.500.4
1.00
0.63
1.00
1.0
(6)0.331.830.330.83
3.00
2.50
1.00
0.0
(12)0000
0.08
0
0.17
12
0.1
(8)00.2500
0.38
0.13
0.25
Weeks
0.2
(8)00.50.130
1.13
0.88
1.0
1.0
(8)0.132.130.250.13
2.13
2.38
2.0
0.0
(9)0.4000
0
0
0.2
16
0.1
(6)0000
0
0.33
0.17
Weeks
0.2
(7)0000
0.14
0.43
1.0
1.0
(5)0000
0.16
0.60
1.0
"Severity score given to individual animal from a complete pathological examination are 0, not remarkable; 1, minimal; 2, slight/mild; 3, moderate;4, moderately severe; and 5, severe, based upon relative evaluation of lesions. Based on severity scoring, a maximum of score 3 was assigned for someof the lesion types at highest phosgene concentrations. Severity scores for each animal within a group were added and an average score per animal wascalculated which is shown in the table.
animals during the 16 weeks of the experiment as would beexpected based on the lung growth with age.
Lung desmosine concentrations were measured as an in-dex of elastin content of the lung tissue. Desmosine wassignificantly increased at the 1.0 ppm phosgene concentra-tion following 4 and 12 weeks of exposure. After 4 weeksof recovery, although the desmosine level appeared to behigher than that in the air-exposed controls, the differencewas not statistically significant (Fig. 3C). There was a sig-nificant time-dependent increase in lung desmosine levels inair-exposed controls. This effect appeared to be similar tothat of hydroxyproline and reflects age-related lung growth.
DISCUSSION
Acute pulmonary effects of phosgene exposures at highconcentrations are associated with edema and bronchiolitisin animals (Frosolono and Pawlowski, 1977; Pawlowski andFrosolono, 1977; Guo et al., 1990). Here we report that very-low-level subchronic phosgene exposures (0.1 ppm, 6 hr/day, 5 days/week) for a period of 4 weeks in rats are associ-ated with pathological abnormalities in the lung. Small mi-croscopical changes at the terminal bronchiolar regions andalso small but significant increases in the lung volume dis-placement occurred at phosgene concentrations as low as0.1 ppm. This suggests that changes in the displacementvolume can be detected at the level at which very subtlehistopathological abnormalities occur and may represent avery sensitive indicator of lung structural changes followingphosgene exposure. This concentration of phosgene (0.1ppm) is the current TLV for 8 hr/day, 5 days per week
exposure over a life time and thus is regarded as safe forhumans.
Our results indicate that in subchronic phosgene exposurescenarios at low concentrations, the effects (i.e., thickeningand inflammation) are primarily associated with the terminalbronchiolar areas and are not as pronounced in the alveolarregions. It was also apparent that at the highest phosgeneconcentration (1.0 ppm) alveolar parenchyma, in addition tothe terminal bronchioles, was thickened and inflamed despitea C X T product that was identical to that of the 0.2 ppmexposure group, suggesting that phosgene concentration isthe primary determinant of the injury and the degree ofpathological damage within the lung. This dominant effect ofconcentration (over time of exposure) has been demonstratedwith other endpoints, including pulmonary function tests,lavage fluid protein (unpublished observations), and hostresistance to bacterial infections (Selgrade et al., 1995).Acute exposure studies using high phosgene concentrations,which demonstrate development of emphysema and damageto the alveolar walls (Clay and Rossing, 1964; Pawlowskiand Frosolono, 1977), support the present findings of periph-eral alveolar damage at higher phosgene concentrations.Changes in the alveolar parenchyma at higher phosgene con-centrations would be consistent with a proportionatelygreater dose reaching the alveolar units, resulting in a moresevere alveolar injury. A similar pattern of dose and injuryhas been previously noted after ozone exposure in animals(Pinkerton et al., 1995). Also, the pathological changes thatwe observed at the terminal bronchiolar sites following phos-gene exposure were similar to those reported followingozone and nitrogen dioxide exposures in animals (Freeman
by guest on July 15, 2011toxsci.oxfordjournals.org
Dow
nloaded from

60 KODAVANTl ET AL.
by guest on July 15, 2011toxsci.oxfordjournals.org
Dow
nloaded from

SUBCHRONIC PULMONARY EFFECTS OF PHOSGENE 61
et al., 1974), suggesting that perhaps the mechanism of lunginjury following phosgene exposure may be similar to thatof other oxidant air pollutants (Kennedy et al., 1989; Ghioet al., 1991).
Evidence of increased hydroxyproline concentrations fol-lowing a subchronic phosgene exposure of 0.25 ppm hasbeen reported previously (Franch and Hatch, 1986). Ourstudies also indicate that subchronic phosgene exposure isassociated with an increase in whole-lung hydroxyproline,but only at 1.0 ppm. Although quantitative analysis of wholelung hydroxyproline revealed an increase only after 1.0 ppmphosgene exposure, trichrome staining of lung sections indi-cated that collagen deposition occurred in terminal bronchi-oles even at 0.2 ppm phosgene. Thus, as we have previouslyreported with ozone in guinea pigs, whole-tissue analysis ofhydroxyproline may have diluted the more localized effectsseen on collagen deposition (Kodavanti et al., 1995). Analy-sis of lung tissue hydroxyproline has been used as an indexof fibrosis. However, the pattern of collagen deposition inthe tissue and its relation with the pathological appearance offibrotic foci should be carefully evaluated since considerablecollagen deposition at the thickened terminal bronchiolarregions can occur without evidence of multialveolar fibroticlesions. At 1.0 ppm, the increases in lung hydroxyproline inrats probably reflects collagen accumulation at both thick-ened terminal bronchioles and alveolar walls which appearto show remodeling of the parenchyma.
Increased collagen deposition may be attributable to stim-ulated collagen synthesis since prolyl hydroxylase activitywas also increased following phosgene exposure. The in-crease in prolyl hydroxylase activity in the whole lung tissuehomogenate coincided with an increase in hydroxyprolineat 4 and 12 weeks of 1.0 ppm phosgene exposure. Althoughprolyl hydroxylase activity returned to control levels duringrecovery, lung hydroxyproline continued to accumulate, per-haps as a result of slower collagen degradation in additionto continued synthesis. Induction of prolyl hydroxylase ac-tivity in the lung has been reported following extended ozoneexposures in animals (Hussain et al., 1976; Hesterberg andLast, 1981; Bhatnagar et al., 1982; Kodavanti et al., 1995).Since we observed an increase in collagen at the terminalbronchiolar regions with concentrations as low as 0.2 ppm
phosgene, it is likely that prolyl hydroxylase was also in-duced in a site specific manner in those terminal bronchiolarregions.
The development of lung emphysema can be correlatedwith a loss of elastin in the alveolar walls. Since emphysemahas been previously reported in animals following phosgeneexposure (Clay and Rossing, 1964), we hypothesized thatemphysema might result from our phosgene exposure sce-narios, with or without a concurrent increase in collagen.However, analysis of lung desmosine following phosgeneexposure indicated an increase in elastin rather than a de-crease. One possible explanation is that the rounding of alve-olar spaces, such as that seen at 1.0 ppm phosgene, is likelyassociated with an increase in tissue matrix production inthe alveolar walls. The increase in lung desmosine may beindicative of tissue remodeling and perhaps is a reflectionof the terminal bronchiolar and alveolar wall thickening thatwas observed histologically. Concurrent increases in colla-gen and elastin have been reported for animal models offibrotic lung diseases (Costa et al., 1983).
In the past, C X T products have been used in calculatingcumulative dose/effects of a toxic substances (Rinehart andHatch, 1964; Cucinell and Arsenal, 1974) and are currentlyused in computing reference concentrations (Jarabek, 1994).However, in most of the acute experimental scenarios, con-centration alone appears to drive toxic response rather thancumulative dose. In the exposure scenarios examined in thepresent study, concentration also appeared to drive all indica-tors of the toxicity. For example, pathology, changes in lungweight and volumes, and changes in hydroxyproline anddesmosine that occurred at 0.2 ppm (6 hr/day, 5 days/weekfor 4 or 12 weeks) were much less evident than those ob-served at 1.0 ppm (6 hr/day, 1 day/week for 4 or 12 weeks),despite the fact that the C X T products were similar atthese phosgene concentrations at one given time. In addition,changes observed after 12 weeks were not three times greaterthan those of 4 weeks, despite the C X T product being threetimes higher at 12 weeks.
In summary, low-level subchronic phosgene exposure (0.1ppm, current TLV) resulted in histological changes at theterminal bronchioles and changes in lung displacement vol-umes. Hydroxyproline increases in the lung following phos-
F1G. 2. Histopathological changes in the lung following subchronic phosgene exposure. The tissue sections were stained with trichrome stain forcollagen fibers and counter stained with hematoxylin and eosin. (A) Control rat lung after 4 weeks of air exposure showing normal alveolar and terminalbronchiolar structure (10X). (B) Lung section from a 0.1 ppm phosgene exposed rat (4 weeks) showing slight terminal bronchiolar thickening andinflammation around the terminal bronchiole (lOx). (C) Lung section from a 0.2 ppm phosgene exposed rat (4 weeks) showing marked terminalbronchiolar thickening and inflammation but less severe alveolar changes (lOx). (D) Lung section from a 1.0 ppm phosgene-exposed rat (4 weeks)showing intense terminal bronchiolar thickening and inflammation (20x). (E) Lung section from a 0.2 ppm phosgene exposed rat (12 weeks) showinginflammation and increased collagen staining at the terminal bronchiolar site (20x). (F) Lung section from a 1.0 ppm phosgene exposed rat (12 weeks)showing marked increase in collagen staining at the terminal bronchiolar and alveolar region (20x). (G) Lung section from a 1.0 ppm phosgene exposedrat (12 weeks) showing alveolar area with rounded air spaces and thickened alveolar walls (20x). (H) Lung section from a rat exposed to phosgene (0.2ppm) for 12 weeks and allowed to recover in clean air for 4 weeks (20X). Notice that the terminal bronchiolar thickening and inflammation have largelydisappeared; however, marked collagen staining is still evident.
by guest on July 15, 2011toxsci.oxfordjournals.org
Dow
nloaded from

62 KODAVANTI ET AL.
I 0.0 ppm KVH 0.2 ppm 1.0 ppm
:: 0.1 ppm ;j;;;|; 0.5 ppm
150
4Wks 12Wks
Exposure Time
12 + 4Wks
FIG. 3. Increases in whole lung prolyl hydroxylase activity (A), hydroxyproline (B), and desmosine (C) following subchronic phosgene exposure inrats. Values are means ± SE of 12 air controls or 8 phosgene exposed rats. Asterisks (•) indicate P < 0.05 compared to matching air exposed group.
gene exposure were reflective of increased collagen primar-ily at terminal bronchiolar regions. Enlargement of alveolar the C X T product,spaces may occur at high phosgene concentrations. How-ever, this may not be consistent with frank emphysema, sincesignificant alveolar wall and terminal bronchiolar remodel-ing may reflect paracintrial disease and not overt interstitialdegeneration. Phosgene-induced changes in collagen werenot reversible within 4 weeks of recovery period. Finally,
the severity of lesions was dependent on concentration, not
ACKNOWLEDGMENTS
The authors thank following collaborators: Mr. Mark Higuchi, Mr. Has-sell Hilliard, Miss Margaret Beamon, and Miss Charlie Micor for conduct-ing phosgene exposure; Mr. Joe Sandy for animal care during exposure;
by guest on July 15, 2011toxsci.oxfordjournals.org
Dow
nloaded from

SUBCHRONIC PULMONARY EFFECTS OF PHOSGENE 63
Mr. John McGee for system development; Mr. David Davies for overallsupervision of phosgene exposures; Mr. Donald Doerfler for performingstatistical analysis; and Dr. John Vandenberg and Dr. Andrew Ghio forcritically reviewing the manuscript.
REFERENCES
Bergman, 1., and Loxley, R. (1961). Two improved and simplified methodsfor the spectrophotometric determination of hydroxyproline. Anal. Chem.35, 1961-1965.
Bhatnagar, R. S., Hussain, M. Z., Sorenson, K., Mustafa, M. G., von Doh-len, F. M., and Lee, S. D. (1982). Effect of ozone on lung collagenbiosynthesis. In Advances in Modern Environmental Toxicology, Vol. V,International Symposium on the Biochemical Effects of Ozone RelatedPhotochemical Oxidants (S. D. Lee, M. G. Mustafa, and M. A. Mehlman,Eds.), pp. 311-321. Princeton Sci Pub., Princeton, NJ.
Clay, J. R., and Rossing, R. G. (1964). Histopathology of exposure to phos-gene. An attempt to produce pulmonary emphysema experimentally.Arch. Pathol. 78, 544-551.
Costa, D. L., Lehmann, J. R., Slatkin, D. N., Popenoe, E. A., and Drew,R. T. (1983). Chronic airway obstruction and bronchiectasis in the ratafter intratracheal bleomycin. Lung 161, 287-300.
Cucinell, S. A., and Arsenal, E. (1974). Review of the toxicity of long-termphosgene exposure. Arch. Environ. Health 28, 272-275.
Currie, W. D., Hatch, G. E., and Frosolono, M. F. (1987). Pulmonary alter-ations in rats due to acute phosgene inhalation. Fundam. Appl. Toxicol.8, 107-114.
Davis, D. S., DeWolf, G. B., Penrod, S. F., and Quass, J. D. (1989). ProjectSummary: Prevention Reference Manual: Chemical Specific, Vol. 14,Control of Accidental Releases of Phosgene. Air and Energy EngineeringResearch Laboratory, U.S. EPA, EPA/600/S8-87/034n.
Diller, W. F., Bruch, J., and Dehnen, W. (1985). Pulmonary changes in therat following low phosgene exposure. Arch. Toxicol 57, 184-190.
Franch, S., and Hatch, G. E. (1986). Pulmonary biochemical effects ofinhaled phosgene in rats. J. Toxicol. Environ. Health 19, 413-423.
Freeman, G.t Juhos, L. T., Furiosi, N. J., Mussenden, R., Stephens, R. J.,and Evans, M. J. (1974). Pathology of pulmonary disease from exposureto interdependent ambient gases (nitrogen dioxide and ozone). Arch.Environ. Health 29, 203-210.
Frosolono, M. F., and Pawlowski, R. (1977). Effect of phosgene on ratlungs after single high-level exposure. 1. Biochemical alterations. Arch.Environ. Health 32, 27'1 -277.
Galdston, M., Luetscher, Jr., J. A., Longcope, W. T., and Ballich, N. L.(1947). A study of the residual effects of phosgene poisoning in humansubjects. II. After chronic exposure. J. Clin. Invest. 26, 169-181.
Gerritsen, W. B., and Buschmann, C. H. (1960). Phosgene poisoning causedby the use of chemical paint removers containing methylene chloride inill-ventilated rooms heated by kerosene stoves. Br. J. Ind. Med. 17, 187-189.
Ghio, A. J., Kennedy, T. P., Hatch, G. E., and Tepper, J. S. (1991) Induc-tion of neutrophil influx diminishes lung injury and mortality followingphosgene inhalation. J. Appl. Physiol. 71, 657-665.
Giri, S. N., Misra, H. P., Chander, D. B., and Chan, Z. (1983). Increasein lung prolyl hydroxylase and superoxide dismutase activities duringbleomycin-induced lung fibrosis in hamsters. Exp. Mol. Pathol. 39, 317-326.
Gross, P., Rinehart, W. E., and Hatch, T. (1965). Chronic pneumonitiscaused by phosgene. Arch. Environ. Health 10, 768-775.
Guo, Y.-L., Kennedy, T. P., Michael, J. R., Sciuto, A. M., Ghio, A. J., Ad-kinson, F. A., Jr., and Gunner, G. H. (1990). Mechanism of phosgene-induced lung toxicity: Role of arachidonate mediators. J. Appl. Physiol.69, 1615-1622
Hesterberg, T. W., and Last, J. A. (1981). Ozone-induced acute pulmonaryfibrosis in rats: Prevention of increased areas of collagen synthesis bymethylprednisolone. Am. Rev. Respir. Dis. 123, 47-52.
Hussain, M. Z., Cross, C. E., Mustafa, M. G., and Bhatnagar, R. S. (1976).Hydroxyproline contents and prolyl hydroxylase activities in lung of ratsexposed to low levels of ozone. Life Sci. 18, 897-904.
Jarabek, A. M. (1994). Inhalation RfC methodology: Dosimetric adjust-ments and dose-response estimation of non-cancer toxicity in the upperrespiratory tract, lnhal. Toxicol. 6(Suppl.), 301-325.
Kennedy, T. P., Michael, J. R., Hoidal, J. R., Hasty, D., Sciuto, A. M.,Hopkins, C , Lazar, R., Bysani, G. K., Tolley, E., and Gurtner, G. (1989).Dibutyryl cATP, aminophylhne, and /3-adrenergic agonists protectagainst pulmonary edema caused by phosgene. J. Appl. Physiol. 67,2542-2552.
Kodavanti, U. P., Hatch, G. E., Starcher, B., Gin, S. N., Winsett, D., andCosta, D. L. (1995). Ozone-induced pulmonary functional, pathological,and biochemical changes in normal and vitamin C-deficient guinea pigs.Fundam. Appl. Toxicol. 24, 154—164.
Pawlowski, R., and Frosolono, M. F. (1977). Effect of phosgene on ratlungs after single high-level exposure. II. Ultrastructural alterations. Arch.Environ. Health 32, 278-283.
Pinkerton, K. E., Menache, M. G., and Plopper, C. G. (1995). Changes in theterminal bronchiolar epithelium, pulmonary acinus, and lung antioxidantenzyme activity. In Health Effects Institute Research Report Number 65,Parts VIII and IX, pp. 41-98, Capital City Press, Montpelier, VT.
Rinehart, W. E., and Hatch, T. (1964). Concentration-time product (CT) asan expression of dose in sublethal exposures to phosgene. Am. Ind, Hyg.Assoc. J. 25, 545-553.
Rossing, R. G. (1964). Physiologic effects of chronic exposure to phosgenein dogs. Am. J. Physiol. 207, 265-272.
Scherle, W. A. (1970). A simple method for volumetry of organs in quantita-tive stereology. Mikroskopie 26, 57-60.
Selgrade, M. J. K., Gilmour, M. I., Yang, Y. G., Burlson, G. R., and Hatch,G. E. (1995). Pulmonary host defenses and resistance to infection follow-ing subchronic exposure to phosgene. Ink Toxicol. 7, 1257-1268.
Sjogren, B., Plato, N., Alexandersson, R., Eklund, A., and Falkenberg,C. (1991). Pulmonary reactions caused by welding-induced decomposedtrichloroethylene. Chest 99, 237-238.
Starcher, B. C. (1982). Antibodies to desmosine. In Methods in Enzymology:Structural and Contractile Proteins. Part A. Extracellular Matrix (F. W.Cunningham and D. W. Fredericksen, Eds.), Vol. 82, pp. 759-762. Aca-demic Press, New York.
by guest on July 15, 2011toxsci.oxfordjournals.org
Dow
nloaded from