comparative changes in the liver of female fischer-344 rats after short-term feeding of a...
TRANSCRIPT

http://tpx.sagepub.com/Toxicologic Pathology
http://tpx.sagepub.com/content/23/5/583The online version of this article can be found at:
DOI: 10.1177/019262339502300504
1995 23: 583Toxicol PatholKobayashi, Toshifumi Tsujiuchi, Ayumi Denda, Benito Lombardi and Yoichi Konishi
Dai Nakae, Yasushi Mizumoto, Nobuaki Andoh, Kazutoshi Tamura, Kohsuke Horiguchi, Takehiro Endoh, EisakuSemipurified or a Semisynthetic L-Amino Acid-Defined Choline-Deficient Diet
Comparative Changes in the Liver of Female Fischer-344 Rats after Short-Term Feeding of a
Published by:
http://www.sagepublications.com
On behalf of:
Society of Toxicologic Pathology
can be found at:Toxicologic PathologyAdditional services and information for
http://tpx.sagepub.com/cgi/alertsEmail Alerts:
http://tpx.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
http://tpx.sagepub.com/content/23/5/583.refs.htmlCitations:
by guest on July 13, 2011tpx.sagepub.comDownloaded from

583
Comparative Changes in the Liver of FemaleFischer-344 Rats after Short-Term Feeding of a
Semipurified or a Semisynthetic L-AminoAcid-Defined Choline-Deficient Diet*
DAI NAKAE,1 YASUSHI MIZUMOTO,1 NOBUAKI ANDOH,1 KAZUTOSHI TAMURA,1KOHSUKE HORIGUCHI,1 TAKEHIRO ENDOH,1 EISAKU KOBAYASHI,1 TOSHIFUMI TSUJIUCHI,1
AYUMI DENDA,1 BENITO LOMBARDI,2 AND YOICHI KONISHI2
1Department of Oncological Pathology, Cancer Center,Nara Medical University, Kashihara, Nara 634, Japan, and2Department of Pathology, University of Pittsburgh School of Medicine,
Pittsburgh, Pennsylvania 15261, USA
* Address correspondence to: Dr. Yoichi Konishi, Departmentof Oncological Pathology, Cancer Center, Nara Medical Uni-versity, 840 Shijo-cho, Kashihara, Nara 634, Japan.
ABSTRACT
Groups of female Fischer-344 rats were fed a semipurified choline-deficient (CD) diet, or a semisyntheticL-amino acid-defined choline-deficient (CDAA) diet, for up to 12 wk and effects of the 2 diets on the liverwere compared. Steatosis was diffuse and more severe throughout in rats fed the CDAA diet than in rats fedthe CD diet. Greater elevations in serum aspartate and alanine aminotransferase activities were also presentin the former rats, along with higher 2-bromodeoxyuridine labeling indices in the liver. Discrete amountsof 8-hydroxyguanine were detected in liver DNA, but were not significantly different in rats fed the 2 diets,or from those present in a group of control rats killed at 0 time. Glutathione S- transferase placental form-positive focal lesions were not observed in any of the rats. The results show that the CDAA diet causes moresevere degrees of steatosis and liver cell death and proliferation than the CD diet, raising the possibility thatit may, in contrast to the CD diet, result in the eventual induction of hepatocellular carcinomas in femaleFischer-344 rats.
Keywords. Dietary choline deficiency; hepatocarcinogenesis; histopathology; liver steatosis; liver cellnecrosis; regenerative cell proliferation; glutathione S-transferase placental form-positive hepatocyte focallesion; 8-hydroxyguanine
INTRODUCTION
It has been shown recently that sexually maturefemale Fischer-344 rats, in contrast to males of thesame strain, are largely resistant to the acute hepaticeffects of a semipurified choline-deficient (CD) diet,and do not develop hepatocellular carcinomas
(HCCs) when chronically fed the diet (13, 29, 30).In male Fischer-344 rats, development of HCC ispreceded by severe degrees of steatosis and of livercell death and regeneration, and a fibrosis that reach-es a stage of borderline cirrhosis after 10-12 mo (14,16). In sexually mature females of the same strain,however, only minor degrees of steatosis and oflivercell death and regeneration occur, and the lack of
HCC development in them was attributed to theattenuated forms of these acute effects of a CD diet
(13, 29, 30).In 1990, we introduced the use of a semisynthetic
L-amino acid-defined choline-deficient (CDAA) diethaving the same overall composition as the CD dietused in the above studies, but in which proteins arereplaced with an equivalent and corresponding mix-ture of L-amino acids (25). In male Fischer-344 rats,this diet proved to induce the same basic spectrumof acute and chronic morphological alterations inthe liver, but more rapidly and significantly moresevere than the alterations induced by the CD diet,as shown for example by the development of frankcirrhosis, more numerous and larger preneoplasticlesions, and a 100% incidence of HCC, all within a12-mo feeding period (12, 25, 26). In addition, theCDAA diet induced more rapid and marked oxi-dative damage of hepatocellular DNA, as revealed
by guest on July 13, 2011tpx.sagepub.comDownloaded from

584
by the formation of 8-hydroxyguanine (8-OHG),than did the CD diet (25).
In this communication we present the results ofan initial short-term study, in which some of theacute effects of the CDAA or CD diet on the liverof female Fischer-344 rats were compared.
MATERIALS AND METHODS
Animals and Diets. Forty-five 5-wk-old femaleFischer-344 rats (Japan SLC Inc., Hamamatsu, Shi-zuoka, Japan) were housed in stainless-steel wirecages, in an air-conditioned atmosphere with con-stant temperature (25 ± 3°C) and humidity (55 ±8%), and under a 12-hr dark/light cycle. A labora-tory chow (Oriental MF diet, Oriental Yeast Co.,Ltd., Itabashi, Tokyo, Japan) and the CDAA (25,26) and CD (13, 29, 30) diets were used. The dietswere purchased from Dyets Inc. (Bethlehem, PA)and were immediately stored at 4°C upon arrival inthe laboratory; a single batch of each diet was usedto complete the study. The diets and laboratory chowwere in pellet form, and feed and water were offeredad libitum throughout.Experimental Protocols. After a 1-wk acclima-
tization on the Oriental MF diet, 5 rats were killedas a control group (group 1 ), and the others weredivided into 2 equal groups and placed on either the
~ CD or CDAA diet (groups 2 and 3, respectively).Subgroups of 5 rats on each diet were killed 2, 4, 8,and 12 wk, thereafter. Two hr before killing, 30 mg2-bromodeoxyuridine (BrdU) (Sigma Chemical Co.,St. Louis, MO) per kg body weight were injected ipinto all rats. Under light ether anesthesia, a bloodsample was taken from the abdominal aorta, andthe liver was removed and weighed. Blood sera wereimmediately prepared and used for determinationof aspartate aminotransferase (AST) and alanineaminotransferase (ALT) activities by routine clini-co-biochemical methods. Five-mm-thick slices fromthe left, median, and right lobes of the livers werefixed in an ice-cold mixture of dehydrated ethanol :glacial acetic acid ( 19:1, volume/volume) for 3 hr,left overnight in 99.5% ethanol at 4°C, and embed-ded in paraffin. Four-~m-thick serial sections of eachlobe were processed for hematoxylin and eosinstaining, Azan-Mallory staining, and immunope-roxidase staining (27) with a polyclonal anti-rat-glutathione S-transferase placental form (GST-P)antibody (Medical and Biological LaboratoriesCorp., Nagoya, Aichi, Japan) and a monoclonal anti-BrdU antibody (Becton Dickinson Immunocyto-metry Systems, San Jose, CA). BrdU labeling indices(BrdU-LIs) were determined by counting 5,000 he-patocytes in 5 randomly chosen fields, and scoringthe number of positive nuclei. The remaining livertissues were immediately frozen by liquid nitrogen
and stored at - 80°C. Two blocks of those liver tis-sues were used for determination of triacylglycerols(TGs) (approximately 0.5 g wet weight) (29, 30), andof 8-OHG in DNA (approximately 2.0 g wet weight)(23, 36). In the latter analyses, an authentic sampleof 8-hydroxydeoxyguanosine (8-OHdG) (Wako PureChemical Industries, Ltd., the City of Kyoto, Kyoto,Japan), and a control sample of deoxyguanosine (dG)(Sigma), were used. The results were expressed asU AST or ALT/L serum, mg TG/g wet liver, andnumber of 8-OHdG formed per 105 total dG nu-cleosides.
Statistics. Differences between means were eval-uated by the Student’s t-test, and were consideredsignificant when p < 0.05.
RESULTS
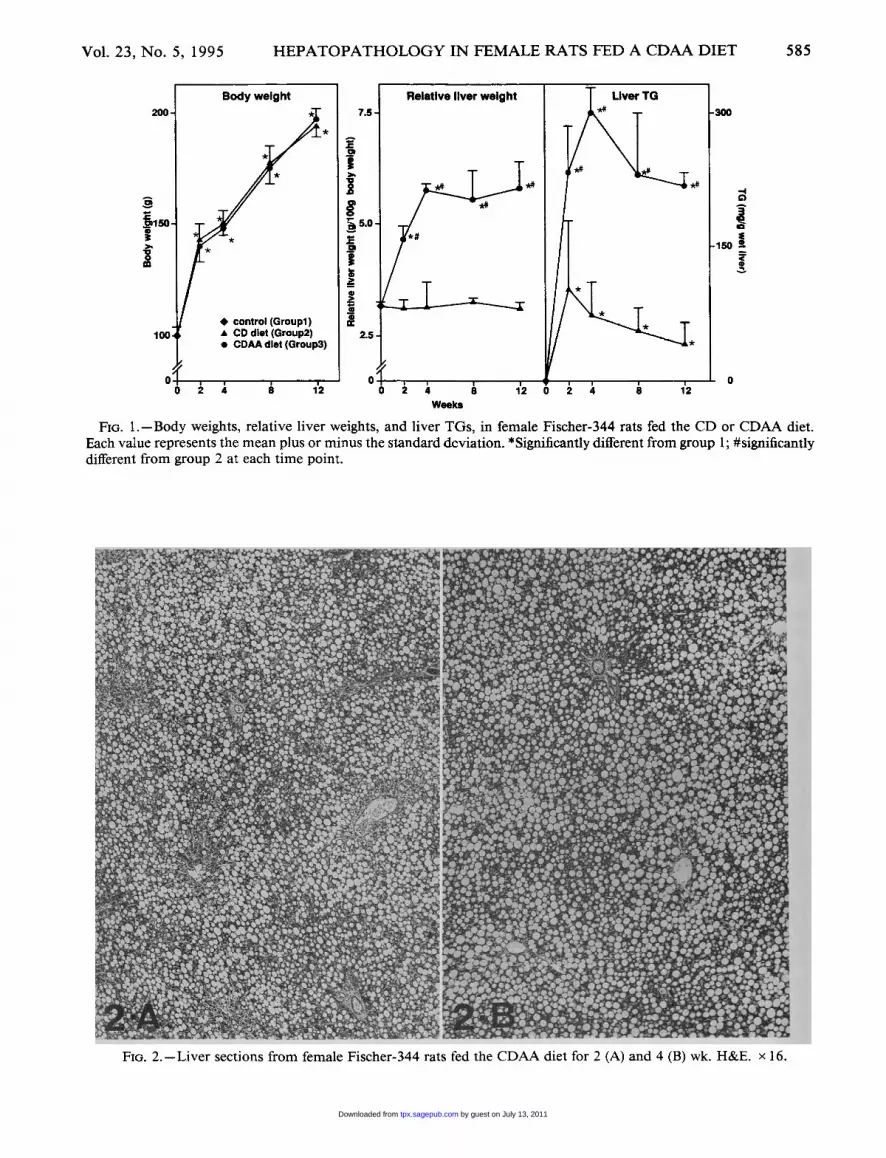
No significant differences were noted in the dietintake of the rats in groups 2 and 3 (data not shown),and no rat died during treatment. The initial averagebody weight of the rats is shown in Fig. 1, and ratsfed the CD or CDAA diet grew equally well. In group2, liver weights remained fairly constant throughoutthe 12-wk feeding period, but in group 3 liver weightsincreased up to 2.5-fold during the first 4 wk, andremained fairly constant thereafter (Fig. 1). Thesedifferences appeared to be largely due to the TGcontent of the livers. Indeed, the average TG contentwas -4 mg/g wet liver in group 1, and in group 2rose to a maximum of ~ 100 mg/g wet liver at 2 wk,and declined steadily thereafter (Fig. 1). On the otherhand, in group 3 TGs increased to a maximum of- 300 mg/g wet liver at 4 wk, and even though theydeclined thereafter, remained at high levels up tothe 12th wk ( ~ 214 mg/g wet liver) (Fig. 1). On grossexamination, all livers had a smooth surface. In
group 2, livers displayed patches of a slightly yel-lowish color in all lobes during the first 4 wk, but anormal color thereafter. In contrast, in group 3 thelivers were enlarged and diffusely yellowish at alltime points.On histopathological examination, a marked stea-
tosis was present in all rats of group 3. At week 2,small numbers of nonfatty hepatocytes could be seenaround the central veins and portal spaces (Fig. 2A),but the steatosis diffused throughout the liver byweek 4 (Fig. 2B), and remained extensive until theend of the experiment. A few small fatty cysts couldbe seen at week 4 (Fig. 2B), and were more numerousand larger thereafter. Single cell necrosis was de-tected at week 2, and gradually increased up to week12 (Fig. 3). In group 2, a modest degree of steatosispredominantly involving hepatocytes in zone 2 waspresent after 2 wk (Fig. 4A), persisted through week4, drastically declined by week 8 (Fig. 4B), and al-most completely disappeared by week 12. Single cell
by guest on July 13, 2011tpx.sagepub.comDownloaded from
585
Fm. 1.-Body weights, relative liver weights, and liver TGs, in female Fischer-344 rats fed the CD or CDAA diet.Each value represents the mean plus or minus the standard deviation. *Significantly different from group 1; #significantlydifferent from group 2 at each time point.
FIG. 2.-Liver sections from female Fischer-344 rats fed the CDAA diet for 2 (A) and 4 (B) wk. H&E. x 16.
by guest on July 13, 2011tpx.sagepub.comDownloaded from

586
FIG. 3.-Liver sections from female Fischer-344 ratsfed the CDAA diet for 12 wk. H&E. x 100. Arrows in-dicate single cell necroses.
necrosis was also detected in these rats, but to amuch lesser extent than in rats in group 3. In both
groups 2 and 3, the observed changes appeared toequally affect the 3 liver lobes examined. Focal, piece-meal, or bridging necrosis, and fibrotic or cirrhoticchanges were absent in all rats. Similarly, no foci ofGST-P-positive hepatocytes (more than 1 MM2 incross section) were seen, but single GST-P-positivecells, and small clusters of a few positive cells scat-tered throughout the parenchyma, were present atall time points in rats of both groups 2 and 3 (datanot shown).
Figure 5 shows that only minimal elevations inthe activities of serum AST and ALT were presentin rats of group 2. In rats of group 3, however, bothenzyme activities increased significantly during thefirst 2 wk, and remained at elevated though declin-ing levels throughout the experimental period. Cellproliferation estimated by means of BrdU-LIs in-creased steadily with time in both groups 2 and 3,but at all time points was significantly higher in ratsof group 3 than in those of group 2 (Fig. 5). Discrete
levels of 8-OHG were detected in liver DNA of therats in both groups 2 and 3 (Fig. 5), but were notsignificantly different from the level of 8-OHG inthe untreated rats of group 1.
DISCUSSION
The results obtained in the present study are con-sistent with previous findings, indicating that femaleFischer-344 rats are largely resistant to the acutesteatogenic and hepatonecrogenic effects of the CDdiet (13, 29, 30). However, no interlobar differenceswere observed in the degree of visible steatosis, aspreviously reported (29), a result that may be ac-counted for by the fact that the rats were probablysexually immature when placed on the diet at 6 wkof age (30). Indeed, TG accumulated moderately inthe liver of these rats during the ensuing 2 wk, butdeclined steadily thereafter.At the same time, the results show that the CDAA
diet is indeed more severely lipotrope-deficient (17)than the CD diet. This fact was clearly bom out byour previous studies (25, 26), in which the hepaticeffects of the 2 diets were compared in male Fischer-344 rats. As was the case in males, in the presentstudy the CDAA diet was found to cause degrees ofsteatosis and of liver cell death and proliferation(compensatory mitogenesis) in females that weresignificantly greater than those induced by the CDdiet. The reason for the greater lipotrope deficiencyof the CDAA diet is not known. However, at least2 factors may contribute to it: (a) smaller traces ofcholine (11, 12) in the CDAA diet than the CD diet(free choline: below detectable limits versus 630pmoles/kg diet; phosphatidylcholine: 11 versus 891 1pmoles/kg diet); and (b) although the 2 diets containthe same amount of methionine, the latter is offeredas proteins in the CD diet, and as a free amino acidin the CDAA diet; any easier and greater utilizationof free methionine by the intestinal flora could re-duce its absorption by the rats (32, 33).
Recently, the resistance of female Fischer-344 ratstoward induction of a high degree of compensatorymitogenesis and eventual tumorigenesis was foundto be not absolute, but a function of the severity ofthe dietary lipotrope deficiency, because it was over-come by feeding a diet devoid of methionine, folicacid, and vitamin B12, as well as of choline (29).However, it is not known whether such an extremedegree of lipotrope deficiency is necessary, or wheth-er diets adequate in folic acid and vitamin B,2, butmore lipotrope-deficient than the CD diet, wouldalso result in such an outcome. The CDAA dietcertainly fits those requisites, as shown by the resultsof our present and previous studies (12, 19, 24-26,36). To test the possibility, therefore, experiments
by guest on July 13, 2011tpx.sagepub.comDownloaded from

587
FIG. 4.-Liver sections from female Fischer-344 rats fed the CD diet for 2 (A) and 8 (B) wk. H&E. x 16.
in female Fischer-344 rats fed the CDAA diet forintermediate and long-term periods of time havebeen initiated, and are in progress in this laboratory.We are waiting with anticipation the results of theseexperiments, because they may also provide the pos-sibility for evaluating other questions concerningthe pathogenesis of the HCCs induced by lipotrope-deficient diets, such as: (a) whether a protracted aswell as initial period of high liver cell proliferationis indeed required for tumorigenesis (16, 29), (b) thenature and role of foci of enzyme-altered hepato-cytes, and (c) the source and significance of 8-OHG.
Differences in methodological approaches pre-clude an exact comparison of the degree of cell pro-liferation induced by the CDAA diet in female rats(Fig. 5), and the degrees induced by the CD diet inmales (16), or by a severe lipotrope-deficient diet infemales (29). Nonetheless, the BrdU-LIs show thatthe CDAA diet caused an enhanced and high degreeof cell proliferation in females that increased steadi-ly for 12 wk, and might thus remain sufficiently highduring an intermediate period of feeding. Fibrosisbecomes active during such a period, and use ofBrdU-labeling will also allow evaluation of to what
extent the proliferation is sustained by hepatocytes,or by other cells, especially fibroblasts.Although the nature and role of foci of enzyme-
altered hepatocytes, as putative preneoplasic le-
sions, are widely accepted in the case of processesof chemical carcinogenesis in the rat (28), there ap-pears to be no consensus as yet about their natureand role in lipotrope-deficient diet models (9, 14,16, 31, 35). The major uncertainties stem from thefact that in the latter, enzyme-altered hepatocytesdevelop in small numbers, like those with which&dquo;spontaneous&dquo; foci have been observed to emergein colony or control groups of rats (31, 35). Foci ofGST-P-positive hepatocytes were not seen in therats treated for up to 12 wk in the present study,and it will be of interest, therefore, to see at whatpoint in time, and in what numbers, they may even-tually appear in females chronically fed the CDAAdiet.
Generation of abnormal amounts of 8-OHG inliver DNA is induced by both the CD (15, 2~) andCDAA (12, 19, 24, 25, 36) diets in male rats, withthe resulting level being greater in rats fed the latterdiet (25). Moreover, in a previous study in male rats
by guest on July 13, 2011tpx.sagepub.comDownloaded from

588
FIG. 5.-Serum AST and ALT activities, liver BrdU-LIs, and 8-OHG levels in liver DNA in female Fischer-344 ratsfed the CD or CDAA diet. Each value represents the mean plus or minus the standard deviation. *Significantly differentfrom group l; #significantly different from group 2 at each time point.
fed the CDAA diet, we observed not only a pro-gressive increase of 8-OHG with time (36), but alsoa correlation between levels of 8-OHG and numberof developed foci of GST-P-positive hepatocytes (24,36). These findings led to the proposal (15, 24) that8-OHG may play an important role in the initiationstage of hepatocarcinogenesis by lipotrope-deficientdiets, given that this adduct is mutagenic (2, 10, 21,22), and that specific pathways exist for its removal(3, 7, 8, 18, 20), which could be overwhelmed ordeteriorated by the high rate of compensatory mi-togenesis induced by these diets in the liver of malerats (14, 16). It is indeed known that in regenerating
liver DNA is more susceptible to formation of8-OHG than in resting liver (1). In the present study,however, neither the CD nor the CDAA diet re-sulted in a significant accumulation of 8-OHG infemale rats (Fig. 5), even though cell proliferationwas significantly higher in rats fed the CDAA thanthe CD diet, and in both of these it was certainlyhigher than in the resting livers of the rats in group1. This result was unexpected, and is difficult toaccount for at the present time. It is not known, forexample, whether liver DNA of-female rats is lesssusceptible to formation of 8-OHG than that of malerats. It is also not known whether there is a threshold
by guest on July 13, 2011tpx.sagepub.comDownloaded from

589
of liver cell proliferation intensity, below which noaccumulation of 8-OHG occurs; despite the resultsobtained (Fig. 5), it could be that the degree of com-pensatory mitogenesis, induced even by the CDAAdiet, falls short of that resulting in male rats. 8-OHGis considered to be a reliable marker of DNA dam-
age induced by oxidative stress (10), but the originof such a stress in the liver of rats fed lipotrope-deficient diets is not clear as yet. Recently, addi-tional (4, 5) evidence was obtained that peroxidationof liver cell membrane lipids does not occur in ratsfed the CD diet (6). For this reason, the suggestionwas made that oxidative stress, and generation of8-OHG, may likely result from an inflammatoryresponse elicited in these livers by the need to re-move cell debris (34). Given the anatomical struc-ture of the liver, and unless a process of apoptosisis responsible for cell death in these livers, removalof the debris would have to be accomplished bymacrophages or other inflammatory cells. Were thisindeed the case, though, in the present study onewould have anticipated seeing levels of 8-OHG infemales fed the CDAA diet that were significantlygreater than in those fed the CD diet, reflecting thesignificant differences in the relative degrees of com-pensatory mitogenesis. Obviously, much remains tobe learned about these early responses of rat liverto lipotrope-deficient diets. At any rate, it will be ofinterest to see whether females are refractory to theinduction of 8-OHG, or whether significant amountsof it will appear during more protracted periods ofthe CDAA diet feeding, especially at the time ofemergence of the first foci of GST-P-positive he-patocytes.
ACKNOWLEDGMENTS
The authors would like to thank Megumi Inagaki,Elizabeth Jahnke-Spinnenweber, Yoshimi Kawai,Kumi Masumoto, Hiroko Megumi, and Yonglin Renfor valuable technical assistance. This work was sup-ported in part by a Grant-in-Aid for the Compre-hensive 10-Year Strategy for Cancer Control fromthe Ministry of Health and Welfare, Japan, and GrantCA23449 from the National Institute of Health.
REFERENCES
1. Adachi S, Kawamura K, and Takemoto K (1994).Increased susceptibility to oxidative DNA damage inregenerating liver. Carcinogenesis 15: 539-543.
2. Akiyama M, Horiuchi T, and Sekiguchi M (1987).Molecular cloning and nucleotide sequence of the mutT mutator of Escherichia coli that causes A:T to C:Gtransversion. Mol. Gen. Genet. 206: 9-16.
3. Akiyama M, Maki H, Sekiguchi M, and Horiuchi T(1989). A specific role of Mut T protein. To preventdG·dA mispairing in DNA replication. Proc. Natl.Acad. Sci. USA 86: 3949-3952.
4. Banni S, Day BW, Evans RW, Corongiu FP, andLombardi B (1994). Liquid chromatographic-massspectrophotometric analysis of conjugated diene fattyacids in a partially hydrogenated fat. J. Am. Oil Chem.Soc. 71: 1321-1325.
5. Banni S, Day BW, Evans RW, Corongiu FP, andLombardi B (1995). Detection of conjugated dieneisomers of linoleic acid in liver lipids of rats fed acholine-devoid diet indicates that the diet does notcause lipoperoxidation. J. Nutr. Biochem. 6: 281-289.
6. Banni S, Salgo MG, Evans RW, Corongiu FP, andLombardi B (1990). Conjugated diene and trans fattyacids in tissue lipids of rats fed an hepatocarcinogeniccholine-devoid diet. Carcinogenesis 11: 2053-2057.
7. Bessho T, Roy R, Yamamoto K, Kasai H, NishimuraS, Tano K, and Mitra S (1993). Repair of
8-hydroxyguanine in DNA by mammalian N- meth-ylpurine-DNA glycosylase. Proc. Natl. Acad. Sci. USA90: 8901-8904.
8. Bessho T, Tano K, Kasai H, Ohtsuka E, and Nishi-mura S (1993). Evidence for two DNA repair en-zymes for 8-hydroxyguanine (7,8-dihydro-8-oxo-guanine) in human cells. J. Biol. Chem. 268: 19416-19421.
9. Chandar N and Lombardi B (1988). Liver cell pro-liferation and incidence of hepatocellular carcinomasin rats fed consecutively a choline-devoid and cho-line-supplemented diet. Carcinogenesis 9: 259-263.
10. Cheng KC, Cahill DS, Kasai H, Nishimura S, andLoeb LA (1992). 8-Hydroxyguanine, an abundantform of oxidative DNA damage, causes G → T andA → C substitutions. J. Biol. Chem. 267: 166-172.
11. DaCosta K-A, Cochary EF, Blusztajn JK, Garner SF,and Zeisel SH (1993). Accumulation of 1,2-sn-dira-dylglycerol with increased membrane-associated pro-tein kinase-C may be the mechanism for spontaneoushepatocarcinogenesis in choline deficient rats. J. Biol.Chem. 268: 2100-2105.
12. Denda A, Tang Q, Endoh T, Tsujiuchi T, HoriguchiK, Noguchi O, Mizumoto Y, Nakae D, and KonishiY (1994). Prevention by acetylsalicylic acid of livercirrhosis and carcinogenesis as well as generation of8-hydroxydeoxyguanosine and thiobarbituric acid-reactive substances caused by a choline-deficient, L-amino acid-defined diet in rats. Carcinogenesis 15:1279-1283.
13. Ghazarian DM, Stoick D, Farber E, and Ghoshal AK(1990). The protective role of ovary in the liver dam-age induced by a choline deficient diet. Proc. Am.Assoc. Cancer Res. 31: 143A.
14. Ghoshal AK and Farber E (1993). Choline deficiency,lipotrope deficiency and the development of liver dis-ease including liver cancer. A new perspective. Lab.Invest. 68: 255-260.
15. Hinrichsen LI, Floyd RA, and Sudilovsky O (1990).Is 8-hydroxydeoxyguanosine a mediator of carcino-genesis by a choline-devoid diet in the rat liver? Car-cinogenesis 11: 1879-1881.
16. Lombardi B and Smith ML (1994). Tumorigenesis,
by guest on July 13, 2011tpx.sagepub.comDownloaded from

590
protooncogene activation, and other gene abnormal-ities in methyl deficiency. J. Nutr. Biochem. 5: 2-9.
17. Lucas CC and Ridout JJ (1967). Fatty livers and lip-otropic phenomena. Prog. Chem. Fats Other Lipids10: 1-150.
18. Michaels ML, Cruz C, Grollman AP, and Miller JH(1992). Evidence that Mut Y and Mut M combine toprevent mutations by an oxidatively damaged formof guanine in DNA. Proc. Natl. Acad. Sci. USA 89:7022-7025.
19. Mizumoto Y, Nakae D, Yoshiji H, Andoh N, Hor-iguchi K, Endoh T, Kobayashi E, Tsujiuchi T, Shi-moji N, Denda A, Tsujii T, Nagao M, WakabayashiK, and Konishi Y (1994). Inhibitory effects of 2-O-octadecylascorbic acid and other vitamin C and Ederivatives on the induction of enzyme-altered pu-tative preneoplastic lesions in the livers of rats fed acholine deficient L-amino acid defined diet. Carci-
nogenesis 15: 241-246.20. Mo YY, Maki H, and Sekiguchi M (1992). Hydrolytic
elimination of a mutagenic nucleotide, 8-oxodGTP,by human 18-kilodalton protein. Sanitization of nu-cleotide pool. Proc. Natl. Acad. Sci. USA 89: 11021-11025.
21. Moriya M (1993). Single-stranded shuttle plasmid formutagenesis studies in mammalian cells. 8-Oxogua-nine in DNA induces targeted G·C → T·A trans-
versions in simian kidney cells. Proc. Natl. Acad. Sci.USA 90: 1122-1126.
22. Moriya M, Ou C, Bodepudi V, Johnson F, TakeshitaM, and Grollman AP (1991). Site-specific mutagen-esis using a gapped duplex vector. A study of tran-slesion synthesis past 8-oxodeoxyguanosine in E. coli.Mutat. Res. 254: 281-288.
23. Nakae D, Andoh N, Mizumoto Y, Endoh T, ShimojiN, Horiguchi K, Shiraiwa K, Tamura K, Denda A,and Konishi Y (1994). Selective 8-hydroxyguanineformation in pancreatic DNA due to a single intra-venous administration of 4-hydroxyaminoquinoline1-oxide in rats. Cancer Lett. 83: 97-103.
24. Nakae D, Mizumoto Y, Yoshiji H, Andoh N, Hor-iguchi K, Shiraiwa K, Kobayashi E, Endoh T, ShimojiN, Tamura K, Tsujiuchi T, Denda A, and KonishiY (1994). Different roles of 8-hydroxyguanine for-mation and 2-thiobarbituric acid-reacting substancegeneration in the early phase of liver carcinogenesisinduced by a choline-deficient, L-amino acid-defineddiet in rats. Jpn. J. Cancer Res. 85: 499-505.
25. Nakae D, Yoshiji H, Maruyama H, Kinugasa T, Den-da A, and Konishi Y (1990). Production of both8-hydroxydeoxyguanosine in liver DNA and γ-gluta-myl-transferase-positive hepatocellular lesions in ratsgiven a choline-deficient, L-amino acid defined diet.Jpn. J. Cancer Res. 81: 1081-1084.
26. Nakae D, Yoshiji H, Mizumoto Y, Horiguchi K, Shi-raiwa K, Tamura K, Denda A, and Konishi Y (1992).
High incidence of hepatocellular carcinomas inducedby a choline deficient L-amino acid defined diet inrats. Cancer Res. 52: 5042-5045.
27. Obara T, Makino T, Ura H, Yokose Y, Kinugasa T,Moore MA, Sato K, and Konishi Y (1986). Com-parative histochemical investigation of the gluta-thione S-transferase placental form and γ-glutamyl-transpeptidase during N-nitrosobis(2-hydroxypro-pyl)amine-induced pancreatic carcinogenesis in ham-ster. Carcinogenesis 7: 801-805.
28. Pitot HC (1990). AHF: Their role in murine hepa-tocarcinogenesis. Annu. Rev. Pharmacol. Toxicol. 30:465-500.
29. Saito R, Jahnke-Spinnenweber E, Shinozuka H, andLombardi B (1994). On the role of compensatorymitogenesis in the hepatocarcinogenicity of cholineand multiple-lipotrope devoid diets. Carcinogenesis15: 1413-1419.
30. Saito R, Palomba L, Rao KN, and Lombardi B (1991).Resistance of female Fischer-344 rats to the hepa-tonecrogenic and hepatocarcinogenic actions of acholine-devoid diet. Carcinogenesis 12: 1451-1457.
31. Sawada N, Poirier L, Moron S, XuY-H, and PitotHC (1990). The effect of choline and methionine de-ficiencies on the number and volume percentage ofaltered hepatic foci in the presence or absence of die-thylnitrosamine initiation in rat liver. Carcinogenesis11:273-281.
32. Silk DB, Fairclough PD, Clark ML, Hegarty JE, MarrsTC, Addison JM, Burston D, Clegg KM, and Mat-thews DM (1980). Use of a peptide rather than freeamino acid nitrogen source in chemically defined"elemental" diets. J. Parenter. Enter. Nutr. 4: 548-553.
33. Silk DB, Perrett D, and Clark M (1972). Absorptionof amino acids and peptides in man. Gut 13: 854-855.
34. Smith ML, Yeleswarapu L, Scalamogna P, Locker J,and Lombardi B (1993). p53 Mutations in hepato-cellular carcinomas induced by a choline-devoid dietin male Fischer-344 rats. Carcinogenesis 14: 503-510.
35. Yokota K, Singh U, and Shinozuka H (1990). Effectsof a choline-deficient diet and a hypolipidemic agenton single glutathione S-transferase placental form-positive hepatocytes in rat liver. Jpn. J. Cancer Res.81:129-134.
36. Yoshiji H , Nakae D, Mizumoto Y, Horiguchi K,Tamura K, Denda A, Tsujii T, and Konishi Y (1992).Inhibitory effect of dietary iron deficiency on induc-tions of putative preneoplstic lesions as well as
8-hydroxydeoxyguanosine in DNA and lipid perox-idation in the livers of rats caused by exposure to acholine-deficient L-amino acid defined diet. Carci-
nogenesis 13: 1227-1233.
by guest on July 13, 2011tpx.sagepub.comDownloaded from