organic geochemical approaches to understanding early life
TRANSCRIPT

Organic geochemical approaches to understanding early life
The MIT Faculty has made this article openly available. Please share how this access benefits you. Your story matters.
As Published 10.1016/j.freeradbiomed.2019.03.005
Publisher Elsevier BV
Version Final published version
Citable link https://hdl.handle.net/1721.1/135118
Terms of Use Creative Commons Attribution-NonCommercial-NoDerivs License
Detailed Terms http://creativecommons.org/licenses/by-nc-nd/4.0/

Contents lists available at ScienceDirect
Free Radical Biology and Medicine
journal homepage: www.elsevier.com/locate/freeradbiomed
Organic geochemical approaches to understanding early lifeJulien Alleon, Roger E. Summons∗
Department of Earth, Atmospheric and Planetary Sciences, Massachusetts Institute of Technology, Cambridge, MA, USA
A B S T R A C T
Here we discuss the early geological record of preserved organic carbon and the criteria that must be applied to distinguish biological from non-biological origins.Sedimentary graphite, irrespective of its isotopic composition, does not constitute a reliable biosignature because the rocks in which it is found are generallymetamorphosed to the point where convincing signs of life have been erased. Rather, multiple lines of evidence, including sedimentary textures, microfossils, largeaccumulations of organic matter and isotopic data for co-existing carbon, nitrogen and sulfur are required before biological origin can be convincingly demonstrated.
1. Introduction
Multiple geological and inorganic geochemical lines of evidencesignal the permanent accumulation of dioxygen in Earth's surface en-vironments between ∼2.4 and ∼2.3 Ga, providing a minimum age forthe development of oxygenic photosynthesis (for details, see reviews byRefs. [1,2]). Opinions differ significantly on how to interpret this cri-tical event. On one hand, aspects of comparative biology, modernbiochemistry, and genomic and metagenomic data suggest that oxy-genic photosynthesis, as it exists in contemporary taxa, is a relativelylate (Paleoproterozoic) metabolic invention from the perspective ofEarth history (see review by Ref. [3]). In contrast, most recent mole-cular clock analyses [4,5] are suggesting the presence of oxygenicphotosynthesis as far back as Mesoarchean times. Advances in phylo-genomics, recognition of pivotal horizontal gene transfer events, andmolecular clocks calibrated to well established geochemical and pa-leontological transitions, offer a potential transformative understandingof the earliest biogeochemical cycles [5,6].
In parallel, carbonaceous matter from Archean sedimentary rocksshould record the key information needed to understanding the timingof the emergence and the evolution of life, including biological in-novations such as oxygenic photosynthesis [7]. Documenting the Ar-chean biosignature record remains highly challenging. Multiple exo-genous and endogenous processes could have sourced abiotic organicmatter on the early Earth including, for example, delivery on cometsand asteroids (e.g. Ref. [8]), photochemical reactions in the atmosphere(e.g. Ref. [9]), and organic synthesis in hydrothermal systems (e.g., Ref.[10]). Because all known Archean rocks are significantly altered bymetamorphic overprinting, it is sometimes very difficult to distinguishbiogenic objects from abiotic ones, and if biogenic, to attribute theirmorphological and/or geochemical characteristics to a particular or-ganism or metabolism. Recent reviews of the early life fossil record
have been provided by Bernard and Papineau [11]; Summons andHallman [12] and Knoll et al. [13]. Rather than giving a complete re-view of the Archean biosignature record, the present contribution aimsto illustrate, from examples selected in the literature, the problems thatneed to be resolved in order to reliably document the early life bio-signature record, and the analytical strategies that have been im-plemented for this purpose.
2. Eoarchean rocks: can graphite afford reliable evidence forpaleobiological activity?
Eoarchean rocks exposed within intensely metamorphosed geolo-gical formations from Canada (Nuvvuagittuq supracrustal belt, SaglekBlock) and southern Greenland (Isua supracrustal belt and Akilia is-land), may have recorded early life activity. Claims for ∼3.8 Ga lifewith possible photoautotrophic metabolism have been made on thebasis of the δ13C values of organic carbon present in rocks from Isua(e.g., Refs. [14,15]). The isotopic data in question, which are within therange expected for biological fractionation, were obtained from ana-lyses of bulk carbonaceous matter extracted from rock samples by de-mineralization with acids. Yet, in situ spatially resolved investigationsusing scanning electron microscopy (SEM) in rock samples from Isua,have shown the presence of organic matter produced by recent en-dolithic microorganisms [16]. Therefore, the antiquity of all the organiccarbon cannot be demonstrated for such old and altered rocks from thebulk analyses alone. Spatially-resolved tools such as scanning andtransmission electron microscopy (SEM and TEM, respectively), Ramanmicrospectroscopy, scanning transmission X-ray microscopy coupledwith X-ray absorption near edge structure spectroscopy (STXM-basedXANES spectroscopy), and ion micro- and nano-probe secondary ionmass spectrometry (SIMS and NanoSIMS, respectively), have thus beenextensively applied to address this problem. By allowing micrometer to
https://doi.org/10.1016/j.freeradbiomed.2019.03.005Received 3 December 2018; Received in revised form 2 March 2019; Accepted 5 March 2019
∗ Corresponding author.E-mail address: [email protected] (R.E. Summons).
Free Radical Biology and Medicine xxx (xxxx) xxx–xxx
0891-5849/ © 2019 The Authors. Published by Elsevier Inc. This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/BY-NC-ND/4.0/).
Please cite this article as: Julien Alleon and Roger E. Summons, Free Radical Biology and Medicine, https://doi.org/10.1016/j.freeradbiomed.2019.03.005

nanometer scale characterization of the elemental, molecular, struc-tural, and isotopic composition of carbonaceous materials, whilst pre-serving their textural relationships with the surrounding mineral phasesin the host rock, these techniques indeed offer key constraints on theantiquity and the origin of carbonaceous matter in old rocks.
As Eoarchean rocks have all experienced peak metamorphic con-ditions at least in the upper amphibolite facies (temperature of550–700 °C and pressure> 4 kbar; [17–22]), any potentially biogenicorganic matter initially present during the rock formation has now beendegraded and metamorphic carbon persists only in the form of crys-talline graphite. Mojzsis et al. [23] have reported in situ ion-microprobemeasurements of 13C-depleted graphite (δ13C values ranging from −49to −21‰) associated with apatite grains in a quartz–pyroxene rocksample from Akilia island. This 13C-depleted graphite has led theseauthors to interpret the graphite-apatite association as the earliest re-mains of life on Earth. There also, these conclusions about the existenceof Eoarchean life have since been challenged.
For instance, Papineau et al. [24,25] have used SEM, TEM, as wellas Raman spectroscopy and STXM-based XANES spectroscopy, tocharacterize the graphite-apatite associations in rock samples from thesame locality. In contrast with previous claims, these authors did notreport any graphite occurring as inclusion in apatite crystals. Rather,graphite was shown to systematically occur as coatings on apatite
Glossary
Archean Eon The geologic eon that extends from ca. 4.0 Ga to theProterozoic 2.5 Ga. The Archean Eon is in the process ofbeing redefined chronometrically and subdivided intothe eras of Eoarchean (4.0–3.6 Ga), Paleoarchean(3.6–3.2 Ga), Mesoarchean (3.2–2.8 Ga), andNeoarchean (2.8–2.5 Ga). The InternationalCommission on Stratigraphy currently does not re-cognize the lower boundary of the Eoarchean
Biogenic Produced by once living organismsBiosignature A distinctive pattern, product, or characteristic by
which life can be identifiedBitumen Organic matter that is or was mobile and soluble in
organic solventsChert Rock almost exclusively made of silicaKerogen Macromolecular organic matter insoluble in organic
solventsMa/Ga Million/billion years before presentSyngenetic Formed at the same time.
Fig. 1. Schematic evolution of biological organic matter during geological burial of the host rocks. Biological organic matter is initially a mixture of diversebiomolecules. The fraction of these biomolecules that escape taphonomic alteration (i.e. degradation before fossilization) will be entrapped into sediments.Biodegradation and the increase of temperature and pressure during burial and diagenesis will then lead to the progressive loss of chemical elements other thancarbon [44]. As low temperature diagenetic reactions transition to metamorphic processes, the residual organic matter takes the form of a chemically inert, carbon-rich and hydrogen-poor material, structurally organized as small polyaromatic units randomly oriented (grey) and connected by short aliphatic chains. With theincreasing temperature and pressure associated with metamorphism, graphitization progresses and results in the lateral extension of the polyaromatic units and theirreorganization into stacked layers, and ultimately end as perfectly crystallized graphite at sufficiently elevated temperature and pressure [45]. The transformation oforganic matter during diagenesis and metamorphism therefore results in the progressive loss of geochemical information concerning any of the former livingorganisms. Organic molecules and graphite have been drawn using ChemDraw. Image of the green cell is from Wikipedia (https://ia.wikipedia.org/wiki/Cyanobacteria). Image of the inert graphitic carbon is the chemical model for Type II kerogen at the end of catagenesis, from Behar and Vandenbroucke [46].
J. Alleon and R.E. Summons Free Radical Biology and Medicine xxx (xxxx) xxx–xxx
2

grains, suggesting a secondary origin. Poorly crystalline graphite hasbeen reported to also occur as coatings on apatite crystals in Eoarcheanrocks from the Nuvvuagittuq supracrustal belt, with a disorderedstructure inconsistent with amphibolite facies metamorphism, implyingthat graphite formed after peak metamorphism [26]. Quartz-pyroxenerock samples from Akilia island have been reported to contain graphitewithin CO2-CH4-H2O-bearing fluid inclusions [27]. Graphite can indeedprecipitate directly from CH4- or CO2-bearing fluids associated withmetamorphic conditions typical of the upper amphibolite facies (e.g.Refs. [28,29]), or at temperatures and pressures as low as 400–500 °Cand 1–2 kbar, respectively [30]. Recent experimental work and ther-modynamic calculations have shown that solid carbonaceous mattercan also precipitate from the reduction of CO2 by H2 in hydrothermalfluids at even lower temperature (200–300 °C; [31,32]).
Recently, additional evidence for Eoarchean life has been claimedbased on the presence of 13C-depleted graphite particles within meta-sedimentary rocks from Isua [33] and from the Saglek block in northernLabrador, Canada [34]. Similarly, however, evidence for the antiquityof graphite is lacking. The geological history of these rocks is complexand poorly resolved. All known sequences from these locations haveexperienced at least two metamorphic events since their formation, themost recent reaching at least the upper amphibolite facies around2.8–2.5 Ga [17–22]. Graphite could thus have precipitated from hy-drothermal and metasomatic fluids at any time during post-Eoarcheanmetamorphic episodes and, at least partially, could be as young as2.8–2.5 Ga. Alternatively, thermal decomposition of siderite (iron car-bonate FeCO3) at 500–600 °C was proposed as an abiotic mechanism forthe formation of graphite found in metamorphosed rocks from Isua[35,36]. Adding further complexity, the sedimentary nature of theprotolith (i.e. the original, unmetamorphosed precursor) of theseEoarchean metamorphic rocks has been questioned, casting doubtabout the significance of the presence of carbonaceous matter, even iforiginally present in the protolith [19,20,37].
Last, but not least, even assuming that the reported carbonaceousparticles originate from the degradation of ≥3.8 Ga organic moleculestrapped in sediments, carbon isotopic compositions, alone, cannot beused to distinguish between abiotic and biogenic sources. 13C-depletedsignatures are indeed not unique to the chemistry of life (e.g. Refs.[38,39]), as demonstrated, for instance, by the experimental synthesisof similarly 13C-depleted abiotic hydrocarbons under hydrothermalconditions [40,41]. Consequently, even for better constrained geolo-gical settings, the interpretation of the origin of carbon based on itsisotopic composition is often controversial. For instance, 13C-depletedsignatures of methane found in fluid inclusions from hydrothermalcherts of the 3.5 Ga Dresser Formation were interpreted to be biogenicby some authors [42], while abiotic alternatives were considered to beequally plausible by others [43]. Additional information is thereforerequired in order to verify and augment the early life biosignature re-cord.
Apart from the problem of dating carbonaceous matter in Eoarcheanmetamorphic rocks, these examples thus emphasize the low paleobio-logical potential of graphite present in the oldest rocks on the Earth.Any organic matter present during the formation of Eoarchean rockshas, by now, been converted into crystalline graphite with the in-evitable loss of all chemical elements other than carbon. This precludesany complementary isotopic or diagnostic molecular data being ac-quired (Fig. 1).
3. The “disordered graphite” in a Hadean zircon
Despite the oldest known terrestrial rocks having Eoarchean ages,older zircon crystals have been preserved in younger sequences.Possibly the best-studied examples are the 3.0 Ga metamorphic rocksfrom Jack Hills, Western Australia [47]. Recently, and despite the manycrystals analyzed, a single 4.1 Ga zircon from Jack Hills was reported tocontain 13C-depleted carbonaceous micrograins, interpreted to be
derived from biological process that occurred more than 4.1 billionyears ago [48]. Yet, this carbonaceous matter is likely derived fromsecondary processes, possibly during subsequent metamorphism asproposed for other inclusions [49], or during polishing as it was thecase for putative Hadean diamonds in zircons from Jack Hills [50].Indeed, even assuming structural disorder induced by polishing arte-facts [51], the reported Raman spectral characteristics of this carbo-naceous matter, interpreted by the authors to be “disordered graphite”,indicate that it did not experienced peak temperatures above 300 °C[45], which is inconsistent with the crystallization history of the zircon,inferred from titanium concentrations to have occurred at temperaturesabove 600 °C [48].
4. Paleoarchean record: structures with biological affinitiesrecord early life activity
The Pilbara Craton in Western Australia (e.g. Ref. [52]), and theKaapvaal Craton in South Africa (e.g. Ref. [53]), host some of the bestpreserved Paleoarchean sedimentary rocks. Primary features in theserocks have been less altered than those of Eoarchean age due to theirlower grade metamorphic history. Therefore, organic matter has notbeen converted into graphite, thereby potentially preserving moregeochemical information concerning the nature of biological precursors(Fig. 1).
4.1. Organic matter from Paleoarchean cherts: does it recordpaleobiological activity?
The solvent-soluble fraction of organic matter (i.e. bitumen) fromArchean rocks is dominated by information-poor aromatic and dia-mondoid hydrocarbons and is easily confounded by contaminants [54].In contrast, the solvent-insoluble fraction (i.e. kerogen) is considered tobe derived from syngenetic organic precursors [55,56]. In one study ofa kerogen isolated from a black chert vein in the 3.5 Ga Dresser For-mation, Pilbara Craton, catalytic pyrolysis in a stream of high-pressurehydrogen gas yielded abundant hydrocarbons with aliphatic chains upto 18 carbons in length with an abrupt step to only trace levels of longerchains [57]. In another study of a similarly aged kerogen from a dif-ferent locality in the Pilbara Craton, Curie-point pyrolysis afforded al-kene-alkane doublets with up to 25 carbon atoms and an odd over evencarbon number preference [58]. In both cases, these distributions wereclaimed to be evidence for Paleoarchean biological activity because ofmolecular characteristics similar to unambiguous biogenic organicmatter. Yet, the presence of such fragile, long-chain, aliphatic mole-cules in rocks having been exposed to low-grade metamorphism hasraised questions about their actual origins [56]. In addition, an alter-native scenario involving abiotic hydrothermal synthesis of organiccompounds has been proposed for the generation of carbonaceousmatter in these Paleoarchean black chert dykes and stratiform units[59]. Lastly, it should be noted that the elemental and isotopic com-positions measured on kerogens isolated from Paleoarchean rocks,namely the organic carbon content, the H/C ratio values, and the δ13Cvalues, can be impacted by late contamination, either during geologicalsecondary processes, or through the chemical isolation and concentra-tion of kerogen in the laboratory [60,61]. Thus, further work is neededto elevate these putative biosignatures beyond all doubt.
4.2. Macroscopic signatures of early life: stromatolites and associatedorganic matter
In both the Pilbara and Kaapvaal Cratons, 3.5–3.2 Ga sedimentaryrocks having sedimentary fabrics morphologically similar to those as-sociated with microbial activity in modern microbialites (i.e. stroma-tolites; microbially influenced sedimentary structures or MISS) havebeen widely interpreted as biosedimentary structures (e.g., Refs.[62–65]). Yet, their biogenicity has sometimes been questioned and
J. Alleon and R.E. Summons Free Radical Biology and Medicine xxx (xxxx) xxx–xxx
3

alternative mechanisms for an abiotic origin have been proposed[66,67]. Reassessment of the possible formation mechanisms of thePilbara stromatolites has largely focused on the Strelley Pool Formation(SPF, Pilbara Craton). There, trace element compositions of carbonatesand cherts indicate that they have been formed in shallow marine en-vironments [68,69], and the SPF stromatolites display distinct mor-photypes with macro- and micro-textural features more easily ex-plainable with the contribution of microbial activity, rather than bypurely abiotic physicochemical processes [70,71]. In addition to theseinorganic geochemical and morphological lines of evidence, organicmatter has been documented in the laminations of the Strelley Poolstromatolites [71–74]. Carbon isotopic compositions of this organicmatter measured using SIMS cover a large range of negative δ13C va-lues, interpreted as the result of multiple autotrophic pathways forcarbon fixation [74]. A similar line of reasoning applies to isotopic dataacquired with SIMS on organic matter present in microstructures pre-served in bedded cherts and hydrothermal chert veins of the DresserFormation. Although present mainly as disseminated clots and in finelaminae, the δ13C values in specific microstructural types range be-tween −33.6‰ and −25.7‰. Because the features are considered torepresent primary textures formed prior to host rock lithification, andthat no evidence was found for subsequent hydrocarbon migrationbeyond the micrometer scale, the carbonaceous structures were inter-preted to represent the remains of microbes that employed different C-fixation pathways [75].
NanoSIMS analyses of sulfur isotopic compositions of organicmatter preserved in the SPF stromatolites have revealed negative δ34Svalues interpreted to be the result of sulfur disproportionation by mi-croorganisms using organics, H2, or CH4 as electron donors to form H2S[72]. Organic matter present in black stromatolitic cherts has also beenreported to be closely associated with framboidal pyrite crystals, in-terpreted as the relict of microbial processes, possibly microbial sulfatereduction, even though the bulk sulfur isotopic composition of pyritedoes not provide information about a specific metabolism [73].
4.3. Microscopic signatures of early life: carbonaceous microstructures
Microfossils have not been reported in Paleoarchean stromatoliteswhere one, logically, would expect to find them. Nevertheless, mor-phologically diverse carbonaceous microstructures, reported inPaleoarchean cherts from both the Pilbara and the Kaapvaal Cratons,were originally interpreted as fossilized filamentous and coccoidal mi-croorganisms (e.g., Refs. [76–78]). Thereafter, both the age and bio-genicity of these objects have yet been questioned (e.g., Ref. [79]). Thechallenging nature of micropaleontological studies in Paleoarcheanrocks is well illustrated by the long-standing controversy about theorigin of microstructures described in black chert samples from the3.5 Ga Apex Basalt Formation (Pilbara Craton, Western Australia).There, Schopf and Packer [78] reported the occurrence of spherical andfilamentous microstructures. Based on morphological comparison withyounger fossil and extant microbial species, the authors interpretedthem as fossilized coccoidal cells and trichomes of prokaryotic, possiblyoxygen-producing, microorganisms. Schopf [80] subsequently de-scribed the morphological diversity of the Apex filamentous objects inmore detail where they were interpreted as trichomic cyanobacterium-like fossilized microorganisms, additionally suggesting that oxygenicphotosynthesis may have evolved as early as 3.5 Ga. Schopf et al. [81]applied Raman microspectroscopy to further document in situ thechemistry of the optically discernible microstructures, showing thatthey are composed of carbonaceous matter that the authors have in-terpreted to be kerogen, thereby providing further evidence for theirbiogenicity. As numerous authors have pointed out, however, the use ofRaman spectroscopy to assign an origin to any given carbonaceousmaterial is questionable because the technique probes the degree ofstructural organization of carbon but does not allow measurements thatprovide constraints concerning biogenicity. The elemental and
molecular compositional characteristics that typify the chemistry ofcomplex carbonaceous materials such as kerogens and other carbo-naceous materials can display similar Raman spectra, independent oftheir biogenic or abiotic origin [82]. Of note, as the Raman spectra ofcarbonaceous matter evolve systematically with metamorphic grade[83], it is possible to quantitatively estimate the peak temperature ex-perienced by carbonaceous matter during metamorphism [84,85].Raman spectroscopy has also been used to evaluate the degree of che-mical preservation of kerogens [86,87], although Raman spectralcharacteristics and elemental/molecular compositions of organicmatter can sometimes be unrelated in such complex disordered organicmaterials [82,85,88,89].
Thus, both the geological context of the carbonaceous micro-structure-bearing cherts in the Apex Basalt Formation and the bio-genicity of the microstructures have been questioned. Previously in-terpreted as a bedded sedimentary deposit containing detrital clasts[80], the Apex chert has been reinterpreted as a hydrothermal brec-ciated vein resulting from the circulation of hot fluids (≅300 °C),therefore representing an environment that was adverse for the pre-sence of cyanobacterial activity but favorable for hyperthermophilebacteria or abiotic synthesis of hydrocarbons [90]. In addition, frompetrographic investigations on the same thin section studied by Schopfet al. [81]; as well as on additional samples re-collected from the samelocality, Brasier et al. [90] have reported more diverse and complexmorphologies, with filamentous microstructures displaying multipleand chaotic branching. This has led these authors to reinterpret themicrofossil-like objects as secondary artefacts formed during the cir-culation of hydrothermal fluids (see also [91]).
In parallel, Garcia-Ruiz et al. [92] have experimentally demon-strated that morphologically complex filamentous microstructures,with shapes and sizes similar to those from the Apex chert, can besynthesized by mixing a water-soluble barium salt with alkaline sodiumsilicate solutions at ambient temperature and pressure. When placedunder hydrothermal conditions in the presence of simple organic mo-lecules (formaldehyde-phenol mixtures), the inorganic filaments servedas templates for the adsorption and condensation of carbonaceousmatter. After having been heated at 500 °C for 5 h, the Raman spectralcharacteristics of the experimental carbonaceous products are reportedto be broadly similar to those of the Apex filaments [92]. Several otherstudies have also reported non-carbonaceous mineral [93,94] andcarbon-containing clay mineral [95,96] filamentous microstructuresfrom the Apex cherts, which have also been interpreted as abiotic ar-tefacts produced during hydrothermalism. Schopf and Kudryavtsev[97] and Schopf et al. [98] still question the significance of thesefindings and the debate continues because these newly described ob-jects are not from the originally documented Apex specimens. Schopfet al. [98] have reported carbon isotopic values collected in situ on theApex carbonaceous microstructures using SIMS. The measured 13C-de-pleted values are proposed by these authors to be morphotype-specific,thus reflecting taxon-specific values. Yet, the carbon isotopic compo-sitions of the different microstructures are not significantly differentfrom one another once one takes into account the variability in themeasurements with respect to the associated analytical uncertainties.
While the carbonaceous nature and the morphological character-istics of many Archean microstructures are not sufficiently informativeto unambiguously assess their origins, the isotopic compositions ofcarbonaceous matter, when associated with morphological character-istics of potential microfossils, might provide additional constraints forthis purpose. The complex multistage hydrothermal history of thebrecciated Apex chert [91,99] makes it challenging to interpret theprimary versus secondary origin of carbonaceous matter. In contrast,cherts resulting from sediment deposition in shallow marine environ-ments such as those from the Strelley Pool Formation clearly provide aless confused window for distinguishing Paleoarchean microfossils fromabiotic artefacts.
Wacey et al. [100] have used optical microscopy, SIMS, TEM, and
J. Alleon and R.E. Summons Free Radical Biology and Medicine xxx (xxxx) xxx–xxx
4

NanoSIMS, to document, down to the submicrometer scale, the mor-phological characteristics and the elemental and carbon isotopic com-positions of spheroidal, ellipsoidal and tubular microstructures in asandstone from the basal member of the 3.4 Ga Strelley Pool Formation.Spheroidal/ellipsoidal microstructures occur as clusters or chains anddisplay morphological features similar to those of well-accepted mi-crofossils from the 1.9 Ga Gunflint Formation (Canada) with an almostcontinuous wall containing carbon and nitrogen as evidenced by TEMand NanoSIMS analyses. These carbonaceous microstructures have 13C-depleted compositions as revealed by SIMS measurements [100] andare sometimes associated with micrometric pyrite crystals having sulfurisotopic compositions consistent with the products of sulfur metabo-lisms [101]. These observations have led Wacey et al. [100] to interpretthese carbonaceous microstructures as 3.4 Ga microfossils of sulfur-metabolizing cells.
Carbonaceous microstructures interpreted as microfossils have alsobeen reported in different chert horizons of the Strelley Pool Formation[102–108]. These objects are morphologically diverse, organized asfilm-like, spheroidal and lenticular forms. Spheroidal microstructuresoccur either as isolated individuals or as clusters while lenticular formsoccur as isolated individuals, and clusters of either aligned or randomlyoriented individuals, and chains [102–105].
SIMS measurements performed on individual microstructureswithin petrographic thin sections show that they are composed ofcarbonaceous matter displaying heterogeneous, texture-specific, 13C-depleted isotopic compositions and hydrogen-to-carbon ratio values,which have been interpreted as the marker of different carbon-fixationmetabolisms and/or the result of selective preservation of distinctbiomolecules during diagenesis [106,107].
Abiotic alternatives have also been proposed for some of the StrelleyPool Formation microstructures. Hydrothermally generated spheroidalmicrostructures morphologically similar to those from the Strelley Poolblack cherts have been reported in a Neoproterozoic (0.75 Ga) chertfrom the Callison Lake Formation, northwestern Canada [13]. Other-wise, Wacey et al. [109] have proposed that some of the spheroidal andlenticular microstructures previously described in the Strelley Poolsandstone (e.g. Ref. [100]), and cherts (e.g. Ref. [103]), could be abioticartefacts formed through the silicification of vesicular volcanic glassfragments coated by organic matter. As stated by Wacey et al. [109];various features of the Strelley Pool lenticular forms remain difficult toexplain with this abiotic mechanism. The carbonaceous lenticular ob-jects retain their structural integrity, morphological features (centralbody with a surrounding thin discoid flange of variable widths), andchain-like arrangements after they have been extracted from the rockby acid demineralization using HF and HCl, as demonstrated by SEMand TEM investigations [104]. In contrast, the disseminated carbo-naceous matter coating some of the volcanogenic vesicles reported byWacey et al. [109] does not display such morphological features andwould not preserve its structural integrity after acid demineralization.In addition, the morphological characteristics of the lenticular objectsco-vary with the sedimentary environment of their host cherts, whichhas been interpreted to reflect adaptive speciation of microorganisms[105].
STXM-based XANES spectroscopy investigations performed on in-dividual lenticular and film-like microstructures have revealed thatthey are composed of nitrogen- and oxygen-rich organic molecules,which are similar to those found in younger organic microfossils fromthe Gunflint Formation, Canada, and appear to be morphotype-specific[108]. Such a high degree of organic molecular preservation is re-markable given that the Strelley Pool microfossils have been exposed topeak temperatures of ≅300 °C at some time during the 3.4 Ga longgeological history of the Strelley Pool Formation, as indicated by theirRaman spectral features [108]. Burial temperatures below 150 °C areindeed generally sufficient to lead to intense chemical degradation[110,111]. The silica matrix surrounding the Strelley Pool organicmicrostructures is likely to have played a key role in the preservation
process, as previously proposed for organic microfossils from theyounger Gunflint cherts [112]. Organic microfossils reported from theStrelley Pool cherts were interpreted to have been initially entombedwithin amorphous opaline silica [103], a fossilization mechanism thatwas experimentally demonstrated to minimize the molecular degrada-tion of microorganisms when they are subsequently exposed to tem-peratures in the range 250–300 °C [113,114]. Of note, such molecularpreservation persists even if opaline silica ends up being completelyconverted into thermally stable quartz, suggesting that the protectionoffered by the mineral matrix may indeed persist beyond the laboratorytimescale [113].
Altogether, these observations strongly support the hypothesis thatthese organic microstructures found in cherts of the Strelley PoolFormation are authentic organic microfossils. Nevertheless, experi-mental investigations using advanced technologies should be conductedto continually re-assess the possibility for alternative abiotic mechan-isms including hydrothermal processes that might generate such mor-phologically and chemically complex organic microstructures.
5. Mesoarchean and Neoarchean organic geochemical records
Reports of bona-fide microfossil occurrences remain limited inMesoarchean and Neoarchean sedimentary rocks [115], which mayreflect a sampling bias towards the older Paleoarchean records.Nevertheless, spheroidal and lenticular microstructures interpreted asmicrofossils have been reported in Mesoarchean sedimentary rocksfrom both the Kaapvaal and Pilbara Cratons (e.g., Refs. [116–118]).Two studies, demonstrating the heightened use and value of in situmicrochemical analyses, have shown that carbon isotopic data acquiredby SIMS [107,119] on the large lenticular and spheroidal microfossils inthe 3.0 Ga Farrel Quartzite had a weighted mean δ13C value of−37.0‰ (±0.4‰), which differed significantly from the backgroundvalue of −32.7‰ (±0.6‰). The authors interpreted these data as bothevidence for biogenicity and an autotrophic lifestyle. A recent study,based on nitrogen isotopic data acquired in situ using NanoSIMS onfilm-like and lenticular microstructures isolated from the FarrelQuartzite through acid demineralization, suggested that the lenticularmicrostructures were oxygen-producing microorganisms [118]. Themorphological similarity of lenticular microfossils of the Farrel Quart-zite with those of the ca. 3.4 Ga Strelley Pool Formation and the 3.4 GaOnverwacht Group of South Africa was also interpreted as evidence forthese microbiota could be the oldest known morphologically-preservedremains with a long-lived (400Ma) fossil record.
Contrasting the paucity of studies reporting organic microfossils,organic matter has been reported to be relatively abundant inMesoarchean and Neoarchean stromatolitic carbonates [120–123].Organic globules closely associated with aragonite, similar to those thatoccur in modern settings, have been reported in the 2.7 Ga stromatolitesof the Tumbiana Formation [120,121]. Carbon isotopic data for non-marine kerogens of the Tumbiana Formation, are posited to support theconcept of a ‘Global Age of Methanotrophy’ at around 2.7 Ga[122,124–126] although not everyone is in agreement with this inter-pretation [127]. SIMS analyses of nanopyrites preserved within theorganic laminae of the Tumbiana stromatolites show an extremely largerange of δ34S values spanning from −33.7‰ to +50.4‰, interpretedto be the result of microbial sulfate reduction, with Rayleigh distilla-tion, within microbial mats during early diagenesis [128]. Data ac-quired at these very small spatial scales, in exceptionally well-preservedsediments for their age, strongly support the idea that microbial pro-cesses operated in similar ways to those of modern microbial matecosystems (see also [129,130]).
Large, global compilations of carbon, sulfur and nitrogen isotopicdata continue to be studied for clues about the evolution of the bio-geochemical cycles of these elements, particularly as they relate to theinception of relevant metabolic processes and their timing [131,132].Havig et al. [132] postulate that C- and S- isotopic trends signify major
J. Alleon and R.E. Summons Free Radical Biology and Medicine xxx (xxxx) xxx–xxx
5
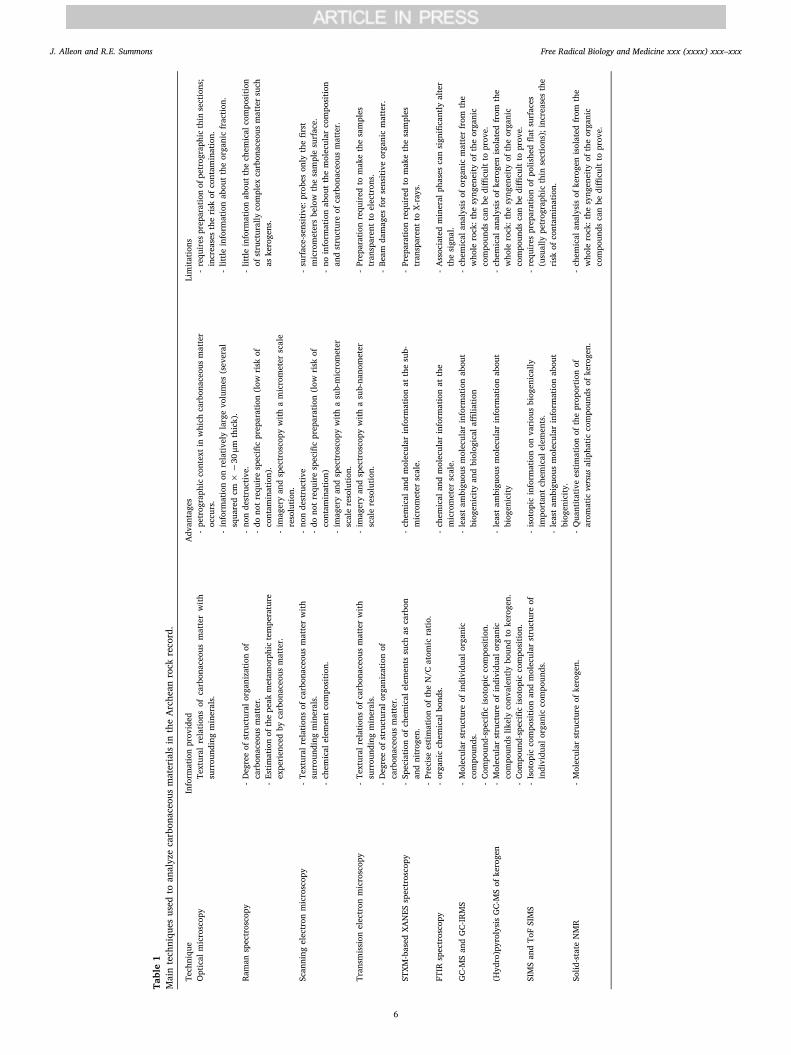
Table1
Maintechniques
used
toanalyzecarbonaceous
materialsin
theArchean
rock
record.
Technique
Inform
ationprovided
Advantages
Limitatio
nsOpticalmicroscopy
Textural
relatio
nsof
carbonaceous
matterwith
surroundingminerals.
-petrographiccontextinwhich
carbonaceous
matter
occurs.
-inform
ationon
relativ
elylargevolumes
(several
squaredcm
×∼30
μmthick).
-requirespreparationofpetrographicthinsections;
increasestherisk
ofcontam
ination.
-little
inform
ationaboutthe
organicfractio
n.
Raman
spectroscopy
-Degreeof
structuralorganizatio
nof
carbonaceous
matter.
-Estim
ationofthepeak
metam
orphictemperature
experiencedby
carbonaceous
matter.
-nondestructive.
-do
notrequirespecificpreparation(low
risk
ofcontam
ination).
-imageryandspectroscopy
with
amicrometer
scale
resolutio
n.
-little
inform
ationaboutthe
chem
icalcompositio
nofstructurallycomplex
carbonaceous
mattersuch
askerogens.
Scanning
electron
microscopy
-Texturalrelatio
nsof
carbonaceous
matterwith
surroundingminerals.
-chem
icalelem
entcompositio
n.
-nondestructive
-do
notrequirespecificpreparation(low
risk
ofcontam
ination)
-imageryandspectroscopy
with
asub-micrometer
scaleresolutio
n.
-surface-sensitive:p
robesonlythefirst
micrometersbelowthesamplesurface.
-no
inform
ationaboutthemolecular
compositio
nandstructureof
carbonaceous
matter.
Transm
ission
electron
microscopy
-Texturalrelatio
nsof
carbonaceous
matterwith
surroundingminerals.
-Degreeof
structuralorganizatio
nof
carbonaceous
matter.
-imageryandspectroscopy
with
asub-nanometer
scaleresolutio
n.-Preparationrequired
tomakethesamples
transparenttoelectrons.
-Beam
damages
forsensitive
organicmatter.
STXM
-based
XANES
spectroscopy
-Speciatio
nof
chem
icalelem
entssuch
ascarbon
andnitrogen.
-Preciseestim
ationof
theN/C
atom
icratio
.
-chem
icalandmolecular
inform
ationat
thesub-
micrometer
scale.
-Preparationrequired
tomakethesamples
transparenttoX-rays.
FTIR
spectroscopy
-organicchem
icalbonds.
-chem
icalandmolecular
inform
ationat
the
micrometer
scale.
-Associatedmineralphases
cansignificantlyalter
thesignal.
GC-MSandGC-IRMS
-Molecular
structureof
individualorganic
compounds.
-Co
mpound-specificisotopiccompositio
n.
-leastam
biguousmolecular
inform
ationabout
biogenicity
andbiologicalaffi
liatio
n-chem
icalanalysisof
organicmatterfrom
the
wholerock:the
syngeneity
oftheorganic
compounds
canbe
difficultto
prove.
(Hydro)pyrolysisGC-MSof
kerogen
-Molecular
structureof
individualorganic
compounds
likelyconvalently
boundto
kerogen.
-Co
mpound-specificisotopiccompositio
n.
-leastam
biguousmolecular
inform
ationabout
biogenicity
-chem
icalanalysisof
kerogenisolated
from
the
wholerock:the
syngeneity
oftheorganic
compounds
canbe
difficultto
prove.
SIMSandToFSIMS
-Isotopiccompositio
nandmolecular
structureof
individualorganiccompounds.
-isotopicinform
ationon
variousbiogenically
importantchem
icalelem
ents.
-leastam
biguousmolecular
inform
ationabout
biogenicity
.
-requires
preparationof
polishedflatsurfaces
(usuallypetrographicthin
sections);increasesthe
risk
ofcontam
ination.
Solid-stateNMR
-Molecular
structureof
kerogen.
-Quantitativ
eestim
ationof
theproportio
nof
arom
atic
vers
usaliphatic
compounds
ofkerogen.
-chem
icalanalysisof
kerogenisolated
from
the
wholerock:the
syngeneity
oftheorganic
compounds
canbe
difficultto
prove.
J. Alleon and R.E. Summons Free Radical Biology and Medicine xxx (xxxx) xxx–xxx
6

transitions in the operation of the carbon and sulfur cycles at 2.8, 2.45and 2.0 Ga after which there is a long record of apparent stasis in thePaleo- and Mesoproterozoic Eras, commonly referred to as the ‘boringbillion’ [133]. Yet, there is little consensus on whether these putativetransitions represent biological innovations or lags in responses of theentire Earth system. There is one transition marked by sulfur isotopegeochemistry that, after continued investigations, still retains its statusas a unidirectional and irreversible process and that is the Great Oxy-genation Event, signifying the permanent presence of molecular oxygenin the atmosphere at around 2.3 Ga [134–137]. The isotopic composi-tion of nitrogen bound into sedimentary kerogens deposited during thistime does indicate that there were concomitant changes in the cyclingof nitrogen in the oceans [138,139] as well as providing evidence forthe presence of Neoarchean alkaline lake systems [140].
One additional and notable feature that is observed in Mesoarcheanand Neoarchean sedimentary rocks is that they record some strati-graphically significant accumulations of total organic carbon (e.g. Ref.[141]), which is potentially sufficient for them to have served as ef-fective source rocks for petroleum generation [142,143]. Yet, modelingof isotopic records preserved in these sedimentary rocks do not showsignificant increase in the burial fraction of organic carbon until thePaleoproterozoic [141,144,145]. On the other hand, trends in organiccarbon isotopic data for Neoarchean organic-rich rocks from the PilbaraCraton strongly suggest that there was a transition from largely anae-robic microbial communities to a world where aerobiosis played anincreasingly important role [124,146].
6. Concluding remarks and future prospects
Technological advances have afforded series of complementaryanalytical techniques to investigate the Archean biosignature record atall spatial scales (see Table 1). The most ancient metasedimentary rockson the Earth, namely the >3.8 Ga rocks from the Greenland and Ca-nadian Cratons, might have recorded traces of early life processes. Yet,distinguishing traces of paleobiological activity from abiotic artefacts inthese heavily altered rocks remains highly challenging and renders theexisting data dubious, at best. Graphitization processes have inevitablydegraded any original organic molecules, leaving only graphite parti-cles which possess little relevant geochemical information for dis-cerning their age and origin (see Fig. 1 and discussion about Eoarcheanrocks). Younger and much less altered rocks preserved in the Paleo-,Meso- and Neo-archean geological formations have, in contrast, re-corded multiple lines of evidence for early life activity. Even the mostancient Archean sedimentary cherts, whilst having a long and complexgeological history so far believed to be incompatible with the chemicalpreservation of organic matter, may have instead a great potential forpreserving molecular clues about the biochemical nature of the earliestbiosphere. Not only the isotopic information has been preserved butalso molecular information. Despite contamination issues that limit theuse of bulk analyses such as GC-MS or NMR to reveal the molecularstructures of organic compounds composing organic microfossils pre-served in Archean cherts, such an analytical approach would be wellsuited for searching for molecular biosignatures in Martian rocks thatwill be collected for a future sample return mission. Along with otherkey factors of the fossilization process, such as the temperature, pres-sure and redox conditions, it is clear that mineralogy plays a criticalrole in the morphological and molecular preservation of fossilized mi-croorganisms, as demonstrated by recent experimental studies[113,114,147–150]. Much experimental work remains to be done toprecisely assess the taphonomic and diagenetic impact on organicmatter preservation of mineral phases relevant for sedimentary andhydrothermal settings on the early Earth and, potentially, on Mars.
Acknowledgments
The authors gratefully acknowledge the Simons Foundation via the
Simons Collaboration on the Origins of Life (award #290361) forsupport during the preparation of this review, as well as the twoanonymous reviewers for their suggestions that improved the manu-script, and Woodward W. Fischer for editorial handling.
Appendix A. Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.freeradbiomed.2019.03.005.
References
[1] J. Farquhar, A.L. Zerkle, A. Bekker, Geological constraints on the origin of oxy-genic photosynthesis, Photosynth. Res. 107 (2011) 11–36.
[2] T.W. Lyons, C.T. Reinhard, N.J. Planavsky, The rise of oxygen in Earth's earlyocean and atmosphere, Nature 506 (2014) 307.
[3] W.W. Fischer, J. Hemp, J.E. Johnson, Evolution of oxygenic photosynthesis, Annu.Rev. Earth Planet Sci. 44 (2016) 647–683.
[4] D.A. Gold, A. Caron, G.P. Fournier, R.E. Summons, Paleoproterozoic sterol bio-synthesis and the rise of oxygen, Nature 543 (2017) 420.
[5] C. Magnabosco, K. Moore, J. Wolfe, G. Fournier, Dating phototrophic microbiallineages with reticulate gene histories, Geobiology 16 (2018) 179–189.
[6] G.P. Fournier, J. Huang, J.P. Gogarten, Horizontal gene transfer from extinct andextant lineages: biological innovation and the coral of life, Philos. Trans. R. Soc.Lond. B Biol. Sci. 364 (2009) 2229–2239.
[7] R.E. Summons, Precambrian organic matter, in: W. White (Ed.), Encyclopedia ofGeochemistry. Encyclopedia of Earth Sciences Series, Springer, 2018.
[8] C. Chyba, C. Sagan, Endogenous production, exogenous delivery and impact-shocksynthesis of organic molecules: an inventory for the origins of life, Nature 355(1992) 125.
[9] E.T. Wolf, O.B. Toon, Fractal organic hazes provided an ultraviolet shield for earlyEarth, Science 328 (2010) 1266–1268.
[10] T.M. McCollom, Miller-Urey and beyond: what have we learned about prebioticorganic synthesis reactions in the past 60 years? Annu. Rev. Earth Planet Sci. 41(2013) 207–229.
[11] S. Bernard, D. Papineau, Graphitic carbons and biosignatures, Elements 10 (2014)435–440.
[12] R. Summons, C. Hallman, Organic geochemical signatures of early life on Earth,Treatise on Geochemistry (2014) 33–46.
[13] A.H. Knoll, K.D. Bergmann, J.V. Strauss, Life: the first two billion years,Philosophical Transactions of the Royal Society B 371 (2016) 20150493.
[14] M. Schidlowski, A 3,800-million-year isotopic record of life from carbon in sedi-mentary rocks, Nature 333 (1988) 313–318.
[15] M.T. Rosing, 13C-depleted carbon microparticles in >3700-Ma sea-floor sedi-mentary rocks from West Greenland, Science 283 (1999) 674–676.
[16] F. Westall, R.L. Folk, Exogenous carbonaceous microstructures in Early Archaeancherts and BIFs from the Isua Greenstone Belt: implications for the search for lifein ancient rocks, Precambrian Res. 126 (2003) 313–330.
[17] J.L. Boak, R.F. Dymek, Metamorphism of the ca. 3800 Ma supracrustal rocks atIsua, West Greenland: implications for early Archaean crustal evolution, EarthPlanet. Sci. Lett. 59 (1982) 155–176.
[18] A.P. Nutman, V.R. McGregor, C.R. Friend, V.C. Bennett, P.D. Kinny, The Itsaqgneiss complex of southern West Greenland; the world's most extensive record ofearly crustal evolution (3900-3600 Ma), Precambrian Res. 78 (1996) 1–39.
[19] C.M. Fedo, M.J. Whitehouse, Metasomatic origin of quartz-pyroxene rock, Akilia,Greenland, and implications for Earth's earliest life, Science 296 (2002)1448–1452.
[20] M.J. Whitehouse, J.S. Myers, C.M. Fedo, The Akilia Controversy: field, structuraland geochronological evidence questions interpretations of> 3.8 Ga life in SWGreenland, J. Geol. Soc. 166 (2009) 335–348.
[21] J. O'Neil, R.W. Carlson, J.-L. Paquette, D. Francis, Formation age and metamorphichistory of the Nuvvuagittuq greenstone belt, Precambrian Res. 220 (2012) 23–44.
[22] T. Komiya, S. Yamamoto, S. Aoki, Y. Sawaki, A. Ishikawa, T. Tashiro, K. Koshida,M. Shimojo, K. Aoki, K.D. Collerson, Geology of the Eoarchean, >3.95 Ga, Nulliaksupracrustal rocks in the Saglek Block, northern Labrador, Canada: the oldestgeological evidence for plate tectonics, Tectonophysics 662 (2015) 40–66.
[23] S. Mojzsis, G. Arrhenius, K. McKeegan, T. Harrison, A. Nutman, C. Friend,Evidence for life on earth before 3,800 million years ago, Nature 384 (1996)55–59.
[24] D. Papineau, B.T. De Gregorio, G.D. Cody, M.D. Fries, S.J. Mojzsis, A. Steele,R.M. Stroud, M.L. Fogel, Ancient graphite in the Eoarchean quartz–pyroxene rocksfrom Akilia in southern West Greenland I: petrographic and spectroscopic char-acterization, Geochem. Cosmochim. Acta 74 (2010) 5862–5883.
[25] D. Papineau, B.T. De Gregorio, R.M. Stroud, A. Steele, E. Pecoits, K. Konhauser,J. Wang, M.L. Fogel, Ancient graphite in the Eoarchean quartz-pyroxene rocksfrom Akilia in southern West Greenland II: isotopic and chemical compositions andcomparison with Paleoproterozoic banded iron formations, Geochem. Cosmochim.Acta 74 (2010) 5884–5905.
[26] D. Papineau, B. De Gregorio, G. Cody, J. O'Neil, A. Steele, R. Stroud, M. Fogel,Young poorly crystalline graphite in the> 3.8-Gyr-old Nuvvuagittuq banded ironformation, Nat. Geosci. 4 (2011) 376.
[27] A. Lepland, M. Van Zuilen, P. Philippot, Fluid‐deposited graphite and its
J. Alleon and R.E. Summons Free Radical Biology and Medicine xxx (xxxx) xxx–xxx
7

geobiological implications in early Archean gneiss from Akilia, Greenland,Geobiology 9 (2011) 2–9.
[28] D. Rumble, T.C. Hoering, Carbon isotope geochemistry of graphite vein depositsfrom New Hampshire, USA, Geochem. Cosmochim. Acta 50 (1986) 1239–1247.
[29] F. Luque, E. Crespo-Feo, J. Barrenechea, L. Ortega, Carbon isotopes of graphite:implications on fluid history, Geoscience Frontiers 3 (2012) 197–207.
[30] L. Ortega, D. Millward, F. Luque, J. Barrenechea, O. Beyssac, J.-M. Huizenga,M. Rodas, S. Clarke, The graphite deposit at Borrowdale (UK): a catastrophicmineralizing event associated with Ordovician magmatism, Geochem.Cosmochim. Acta 74 (2010) 2429–2449.
[31] V. Milesi, F. Guyot, F. Brunet, L. Richard, N. Recham, M. Benedetti, J. Dairou,A. Prinzhofer, Formation of CO2, H2 and condensed carbon from siderite dis-solution in the 200–300 °C range and at 50 MPa, Geochem. Cosmochim. Acta 154(2015) 201–211.
[32] V. Milesi, T.M. McCollom, F. Guyot, Thermodynamic constraints on the formationof condensed carbon from serpentinization fluids, Geochem. Cosmochim. Acta 189(2016) 391–403.
[33] T. Hassenkam, M. Andersson, K. Dalby, D. Mackenzie, M. Rosing, Elements ofEoarchean life trapped in mineral inclusions, Nature 548 (2017) 78–81.
[34] T. Tashiro, A. Ishida, M. Hori, M. Igisu, M. Koike, P. Méjean, N. Takahata, Y. Sano,T. Komiya, Early trace of life from 3.95 Ga sedimentary rocks in Labrador, Canada,Nature 549 (2017) 516–518.
[35] M.A. Van Zuilen, A. Lepland, G. Arrhenius, Reassessing the evidence for the ear-liest traces of life, Nature 418 (2002) 627–630.
[36] M.A. van Zuilen, A. Lepland, J. Teranes, J. Finarelli, M. Wahlen, G. Arrhenius,Graphite and carbonates in the 3.8 Ga old Isua supracrustal belt, southern WestGreenland, Precambrian Res. 126 (2003) 331–348.
[37] M.J. Whitehouse, R. Schoenberg, C. Fedo, B. Kamber, Does a heavy Fe-isotopecomposition of Akilia quartz-amphibole-pyroxene rocks necessitate a BIF origin?Astrobiology 15 (2015) 816–824.
[38] Y. Bottinga, Calculated fractionation factors for carbon and hydrogen isotope ex-change in the system calcite-carbon dioxide-graphite-methane-hydrogen-watervapor, Geochem. Cosmochim. Acta 33 (1969) 49–64.
[39] E. Galimov, Isotope organic geochemistry, Org. Geochem. 37 (2006) 1200–1262.[40] J. Horita, M.E. Berndt, Abiogenic methane formation and isotopic fractionation
under hydrothermal conditions, Science 285 (1999) 1055–1057.[41] T.M. McCollom, J.S. Seewald, Carbon isotope composition of organic compounds
produced by abiotic synthesis under hydrothermal conditions, Earth Planet. Sci.Lett. 243 (2006) 74–84.
[42] Y. Ueno, K. Yamada, N. Yoshida, S. Maruyama, Y. Isozaki, Evidence from fluidinclusions for microbial methanogenesis in the early Archaean era, Nature 440(2006) 516.
[43] B. Sherwood-Lollar, T. McCollom, Biosignatures and abiotic constraints on earlylife, Nature 444 (2006) E18.
[44] M. Vandenbroucke, C. Largeau, Kerogen origin, evolution and structure, Org.Geochem. 38 (2007) 719–833.
[45] P.R. Buseck, O. Beyssac, From organic matter to graphite: Graphitization,Elements 10 (2014) 421–426.
[46] F. Behar, M. Vandenbroucke, Chemical modelling of kerogens, Org. Geochem. 11(1987) 15–24.
[47] W. Compston, R. Pidgeon, Jack Hills, evidence of more very old detrital zircons inWestern Australia, Nature 321 (1986) 766–769.
[48] E.A. Bell, P. Boehnke, T.M. Harrison, W.L. Mao, Potentially biogenic carbon pre-served in a 4.1 billion-year-old zircon, Proc. Natl. Acad. Sci. Unit. States Am. 112(2015) 14518–14521.
[49] B. Rasmussen, I.R. Fletcher, J.R. Muhling, C.J. Gregory, S.A. Wilde, Metamorphicreplacement of mineral inclusions in detrital zircon from Jack Hills, Australia:implications for the hadean earth, Geology 39 (2011) 1143–1146.
[50] L. Dobrzhinetskaya, R. Wirth, H. Green, Diamonds in Earthʼs oldest zircons fromJack Hills conglomerate, Australia, are contamination, Earth Planet. Sci. Lett. 387(2014) 212–218.
[51] O. Beyssac, B. Goffé, J.-P. Petitet, E. Froigneux, M. Moreau, J.-N. Rouzaud, On thecharacterization of disordered and heterogeneous carbonaceous materials byRaman spectroscopy, Spectrochim. Acta Mol. Biomol. Spectrosc. 59 (2003)2267–2276.
[52] M.J. Van Kranendonk, Volcanic degassing, hydrothermal circulation and theflourishing of early life on Earth: a review of the evidence from c. 3490-3240 Marocks of the Pilbara Supergroup, Pilbara Craton, Western Australia, Earth Sci. Rev.74 (2006) 197–240.
[53] D.R. Lowe, G.R. Byerly, Stratigraphy of the West-Central Part of the BarbertonGreenstone Belt, South Africa, Special Papers-Geological Society of America, 1999,pp. 1–36.
[54] K.L. French, C. Hallmann, J.M. Hope, P.L. Schoon, J.A. Zumberge, Y. Hoshino,C.A. Peters, S.C. George, G.D. Love, J.J. Brocks, R. Buick, R.E. Summons,Reappraisal of hydrocarbon biomarkers in Archean rocks, Proc. Natl. Acad. Sci.Unit. States Am. 112 (2015) 5915–5920.
[55] J.J. Brocks, G.D. Love, C.E. Snape, G.A. Logan, R.E. Summons, R. Buick, Release ofbound aromatic hydrocarbons from late Archean and Mesoproterozoic kerogensvia hydropyrolysis, Geochem. Cosmochim. Acta 67 (2003) 1521–1530.
[56] C.P. Marshall, G.D. Love, C.E. Snape, A.C. Hill, A.C. Allwood, M.R. Walter,M.J. Van Kranendonk, S.A. Bowden, S.P. Sylva, R.E. Summons, Structural char-acterization of kerogen in 3.4 Ga archaean cherts from the Pilbara Craton, westernAustralia, Precambrian Res. 155 (2007) 1–23.
[57] J.-P. Duda, V. Thiel, T. Bauersachs, H. Mißbach, M. Reinhardt, N. Schäfer,M.J. Van Kranendonk, J. Reitner, Ideas and perspectives: hydrothermally drivenredistribution and sequestration of early Archaean biomass–the “hydrothermal
pump hypothesis”, Biogeosciences 15 (2018) 1535–1548.[58] S. Derenne, F. Robert, A. Skrzypczak-Bonduelle, D. Gourier, L. Binet, J.-
N. Rouzaud, Molecular evidence for life in the 3.5 billion year old Warrawoonachert, Earth Planet. Sci. Lett. 272 (2008) 476–480.
[59] J. Lindsay, M. Brasier, N. McLoughlin, O. Green, M. Fogel, A. Steele, S. Mertzman,The problem of deep carbon—an Archean paradox, Precambrian Res. 143 (2005)1–22.
[60] F. Delarue, S. Derenne, K. Sugitani, F. Baudin, F. Westall, B. Kremer, R. Tartèse,A. Gonzalez, F. Robert, What is the meaning of Hydrogen-to-Carbon ratio de-termined in Archean organic matter? Org. Geochem. 122 (2018) 140–146.
[61] E. Muller, C. Thomazo, E.E. Stüeken, C. Hallmann, A. Leider, C. Chaduteau,R. Buick, F. Baton, P. Philippot, M. Ader, Bias in carbon concentration and δ13Cmeasurements of organic matter due to cleaning treatments with organic solvents,Chem. Geol. 493 (2018) 405–412.
[62] D.R. Lowe, Stromatolites 3,400-myr old from the archean of western Australia,Nature 284 (1980) 441–443.
[63] M. Walter, R. Buick, J. Dunlop, Stromatolites 3,400–3,500 Myr old from the northPole area, western Australia, Nature 284 (1980) 443–445.
[64] N. Noffke, Microbially Induced Sedimentary Structures, Encyclopedia ofAstrobiology, Springer, 2015, pp. 1565–1569.
[65] M. Homann, P. Sansjofre, M. Van Zuilen, C. Heubeck, J. Gong, B. Killingsworth,I.S. Foster, A. Airo, M.J. Van Kranendonk, M. Ader, S.V. Lalonde, Microbial lifeand biogeochemical cycling on land 3,220 million years ago, Nat. Geosci. 11(2018) 665.
[66] D.R. Lowe, Abiological origin of described stromatolites older than 3.2 Ga,Geology 22 (1994) 387–390.
[67] J.P. Grotzinger, D.H. Rothman, An abiotic model for stromatolite morphogenesis,Nature 383 (1996) 423.
[68] M.J. Van Kranendonk, G.E. Webb, B.S. Kamber, Geological and trace elementevidence for a marine sedimentary environment of deposition and biogenicity of3.45 Ga stromatolitic carbonates in the Pilbara Craton, and support for a reducingArchaean ocean, Geobiology 1 (2003) 91–108.
[69] A.C. Allwood, B.S. Kamber, M.R. Walter, I.W. Burch, I. Kanik, Trace elementsrecord depositional history of an Early Archean stromatolitic carbonate platform,Chem. Geol. 270 (2010) 148–163.
[70] A.C. Allwood, M.R. Walter, B.S. Kamber, C.P. Marshall, I.W. Burch, Stromatolitereef from the early archaean era of Australia, Nature 441 (2006) 714.
[71] A.C. Allwood, J.P. Grotzinger, A.H. Knoll, I.W. Burch, M.S. Anderson,M.L. Coleman, I. Kanik, Controls on development and diversity of Early Archeanstromatolites, Proc. Natl. Acad. Sci. Unit. States Am. 106 (2009) 9548–9555.
[72] T.R. Bontognali, A.L. Sessions, A.C. Allwood, W.W. Fischer, J.P. Grotzinger,R.E. Summons, J.M. Eiler, Sulfur isotopes of organic matter preserved in 3.45-billion-year-old stromatolites reveal microbial metabolism, Proc. Natl. Acad. Sci.Unit. States Am. 109 (2012) 15146–15151.
[73] J.-P. Duda, M.J. Van Kranendonk, V. Thiel, D. Ionescu, H. Strauss, N. Schäfer,J. Reitner, A rare glimpse of Paleoarchean life: geobiology of an exceptionallypreserved microbial mat facies from the 3.4 Ga Strelley Pool Formation, WesternAustralia, PLoS One 11 (2016) e0147629.
[74] D.T. Flannery, A.C. Allwood, R.E. Summons, K.H. Williford, W. Abbey, E.D. Matys,N. Ferralis, Spatially-resolved isotopic study of carbon trapped in ∼3.43 GaStrelley Pool Formation stromatolites, Geochem. Cosmochim. Acta 223 (2018)21–35.
[75] N. Morag, K.H. Williford, K. Kitajima, P. Philippot, M.J. Van Kranendonk, K. Lepot,C. Thomazo, J.W. Valley, Microstructure-specific carbon isotopic signatures oforganic matter from ∼3.5 Ga cherts of the Pilbara Craton support a biologicorigin, Precambrian Res. 275 (2016) 429–449.
[76] S. Awramik, J. Schopf, M. Walter, Filamentous fossil bacteria from the archean ofwestern Australia, Precambrian Res. 20 (1983) 357–374.
[77] M.M. Walsh, D.R. Lowe, Filamentous microfossils from the 3,500-Myr-oldOnverwacht Group, barberton mountain land, South Africa, Nature 314 (1985)530–532.
[78] J.W. Schopf, B.M. Packer, Early archean (3.3-billion to 3.5-billion-year-old) mi-crofossils from warrawoona Group, Australia, Science 237 (1987) 70–73.
[79] R. Buick, Microfossil recognition in Archean rocks: an appraisal of spheroids andfilaments from a 3500 my old chert-barite unit at North Pole, Western Australia,Palaios (1990) 441–459.
[80] J.W. Schopf, Microfossils of the Early Archean Apex chert: new evidence of theantiquity of life, Science 260 (1993) [640]–[646].
[81] J.W. Schopf, A.B. Kudryavtsev, D.G. Agresti, T.J. Wdowiak, A.D. Czaja,Laser–Raman imagery of Earth's earliest fossils, Nature 416 (2002) 73–76.
[82] J.D. Pasteris, B. Wopenka, Necessary, but not sufficient: Raman identification ofdisordered carbon as a signature of ancient life, Astrobiology 3 (2003) 727–738.
[83] B. Wopenka, J.D. Pasteris, Structural characterization of kerogens to granulite-facies graphite; applicability of Raman microprobe spectroscopy, Am. Mineral. 78(1993) 533–557.
[84] O. Beyssac, B. Goffé, C. Chopin, J. Rouzaud, Raman spectra of carbonaceousmaterial in metasediments: a new geothermometer, J. Metamorph. Geol. 20(2002) 859–871.
[85] O. Beyssac, M. Lazzeri, Application of Raman spectroscopy to the study of gra-phitic carbons in the Earth Sciences. Applications of Raman spectroscopy to earthsciences and cultural heritage, EMU Notes in Mineralogy 12 (2012) 415–454.
[86] F. Delarue, J.-N. Rouzaud, S. Derenne, M. Bourbin, F. Westall, B. Kremer,K. Sugitani, D. Deldicque, F. Robert, The Raman-derived carbonization continuum:a tool to select the best preserved molecular structures in Archean kerogens,Astrobiology 16 (2016) 407–417.
[87] N. Ferralis, E.D. Matys, A.H. Knoll, C. Hallmann, R.E. Summons, Rapid, direct and
J. Alleon and R.E. Summons Free Radical Biology and Medicine xxx (xxxx) xxx–xxx
8
non-destructive assessment of fossil organic matter via microRaman spectroscopy,Carbon 108 (2016) 440–449.
[88] S. Bernard, B. Horsfield, H.-M. Schulz, A. Schreiber, R. Wirth, T.T.A. Vu,F. Perssen, S. Könitzer, H. Volk, N. Sherwood, Multi-scale detection of organic andinorganic signatures provides insights into gas shale properties and evolution,Chem. Erde 70 (2010) 119–133.
[89] S. Bernard, B. Horsfield, H.-M. Schulz, R. Wirth, A. Schreiber, N. Sherwood,Geochemical evolution of organic-rich shales with increasing maturity: a STXMand TEM study of the Posidonia Shale (Lower Toarcian, northern Germany), Mar.Petrol. Geol. 31 (2012) 70–89.
[90] M.D. Brasier, O.R. Green, A.P. Jephcoat, A.K. Kleppe, M.J. Van Kranendonk,J.F. Lindsay, A. Steele, N.V. Grassineau, Questioning the evidence for Earth'soldest fossils, Nature 416 (2002) 76–81.
[91] M.D. Brasier, O.R. Green, J.F. Lindsay, N. McLoughlin, A. Steele, C. Stoakes,Critical testing of Earth's oldest putative fossil assemblage from the ∼3.5 Ga Apexchert, Chinaman Creek, Western Australia, Precambrian Res. 140 (2005) 55–102.
[92] J. Garcia-Ruiz, S. Hyde, A. Carnerup, A. Christy, M. Van Kranendonk, N. Welham,Self-assembled silica-carbonate structures and detection of ancient microfossils,Science 302 (2003) 1194–1197.
[93] D.L. Pinti, R. Mineau, V. Clement, Hydrothermal alteration and microfossil arte-facts of the 3,465-million-year-old Apex chert, Nat. Geosci. 2 (2009) 640.
[94] C.P. Marshall, J.R. Emry, A.O. Marshall, Haematite pseudomicrofossils present inthe 3.5-billion-year-old Apex chert, Nat. Geosci. 4 (2011) 240.
[95] M.D. Brasier, J. Antcliffe, M. Saunders, D. Wacey, Changing the picture of Earth'searliest fossils (3.5–1.9 Ga) with new approaches and new discoveries, Proc. Natl.Acad. Sci. Unit. States Am. 112 (2015) 4859–4864.
[96] D. Wacey, M. Saunders, C. Kong, A. Brasier, M. Brasier, 3.46 Ga Apex chert ‘mi-crofossils’ reinterpreted as mineral artefacts produced during phyllosilicate ex-foliation, Gondwana Res. 36 (2016) 296–313.
[97] J.W. Schopf, A.B. Kudryavtsev, Biogenicity of Earth's earliest fossils: a resolution ofthe controversy, Gondwana Res. 22 (2012) 761–771.
[98] J.W. Schopf, K. Kitajima, M.J. Spicuzza, A.B. Kudryavtsev, J.W. Valley, SIMSanalyses of the oldest known assemblage of microfossils document their taxon-correlated carbon isotope compositions, Proc. Natl. Acad. Sci. Unit. States Am. 115(2018) 53–58.
[99] M.-C. Sforna, M. Van Zuilen, P. Philippot, Structural characterization by Ramanhyperspectral mapping of organic carbon in the 3.46 billion-year-old Apex chert,Western Australia, Geochem. Cosmochim. Acta 124 (2014) 18–33.
[100] D. Wacey, M.R. Kilburn, M. Saunders, J. Cliff, M.D. Brasier, Microfossils of sul-phur-metabolizing cells in 3.4-billion-year-old rocks of Western Australia, Nat.Geosci. 4 (2011) 698–702.
[101] D. Wacey, N. McLoughlin, M.J. Whitehouse, M.R. Kilburn, Two coexisting sulfurmetabolisms in a ca. 3400 Ma sandstone, Geology 38 (2010) 1115–1118.
[102] K. Sugitani, K. Lepot, T. Nagaoka, K. Mimura, M. Van Kranendonk, D.Z. Oehler,M.R. Walter, Biogenicity of morphologically diverse carbonaceous microstructuresfrom the ca. 3400 Ma Strelley Pool Formation, in the Pilbara Craton, westernAustralia, Astrobiology 10 (2010) 899–920.
[103] K. Sugitani, K. Mimura, T. Nagaoka, K. Lepot, M. Takeuchi, Microfossil assemblagefrom the 3400 Ma Strelley Pool Formation in the Pilbara Craton, westernAustralia: results form a new locality, Precambrian Res. 226 (2013) 59–74.
[104] K. Sugitani, K. Mimura, M. Takeuchi, K. Lepot, S. Ito, E. Javaux, Early evolution oflarge micro‐organisms with cytological complexity revealed by microanalyses of3.4 Ga organic-walled microfossils, Geobiology 13 (2015) 507–521.
[105] K. Sugitani, T. Kohama, K. Mimura, M. Takeuchi, R. Senda, H. Morimoto,Speciation of paleoarchean life demonstrated by analysis of the morphologicalvariation of lenticular microfossils from the Pilbara Craton, Australia,Astrobiology 18 (2018) 1057–1070.
[106] K. Lepot, K.H. Williford, T. Ushikubo, K. Sugitani, K. Mimura, M.J. Spicuzza,J.W. Valley, Texture-specific isotopic compositions in 3.4 Gyr old organic mattersupport selective preservation in cell-like structures, Geochem. Cosmochim. Acta112 (2013) 66–86.
[107] D.Z. Oehler, M.M. Walsh, K. Sugitani, M.-C. Liu, C.H. House, Large and robustlenticular microorganisms on the young Earth, Precambrian Res. 296 (2017)112–119.
[108] J. Alleon, S. Bernard, C. Le Guillou, O. Beyssac, K. Sugitani, F. Robert, Chemicalnature of the 3.4 Ga Strelley Pool microfossils, Geochemical Perspectives Letters 7(2018) 37–42.
[109] D. Wacey, N. Noffke, M. Saunders, P. Guagliardo, D.M. Pyle, Volcanogenic pseudo-fossils from the ∼3.48 Ga Dresser Formation, Pilbara, western Australia,Astrobiology 18 (2018) 539–555.
[110] S. Bernard, B. Horsfield, Thermal maturation of gas shale systems, Annu. Rev.Earth Planet Sci. 42 (2014) 635–651.
[111] D.E. Briggs, R.E. Summons, Ancient biomolecules: their origins, fossilization, androle in revealing the history of life, Bioessays 36 (2014) 482–490.
[112] J. Alleon, S. Bernard, C. Le Guillou, J. Marin-Carbonne, S. Pont, O. Beyssac,K.D. McKeegan, F. Robert, Molecular preservation of 1.88 Ga Gunflint organicmicrofossils as a function of temperature and mineralogy, Nat. Commun. 7 (2016)11977.
[113] J. Alleon, S. Bernard, C. Le Guillou, D. Daval, F. Skouri-Panet, S. Pont, L. Delbes,F. Robert, Early entombment within silica minimizes the molecular degradation ofmicroorganisms during advanced diagenesis, Chem. Geol. 437 (2016) 98–108.
[114] M. Igisu, T. Yokoyama, Y. Ueno, S. Nakashima, M. Shimojima, H. Ohta,S. Maruyama, Changes of aliphatic C–H bonds in cyanobacteria during experi-mental thermal maturation in the presence or absence of silica as evaluated byFTIR microspectroscopy, Geobiology 16 (2018) 412–428.
[115] W. Altermann, J. Kazmierczak, Archean microfossils: a reappraisal of early life on
Earth, Res. Microbiol. 154 (2003) 611–617.[116] K. Sugitani, K. Grey, T. Nagaoka, K. Mimura, M.R. Walter, Taxonomy and bio-
genicity of archaean spheroidal microfossils (ca. 3.0 Ga) from the mount gold-sworthy–mount grant area in the northeastern Pilbara Craton, western Australia,Precambrian Res. 173 (2009) 50–59.
[117] E.J. Javaux, C.P. Marshall, A. Bekker, Organic-walled microfossils in 3.2-billion-year-old shallow-marine siliciclastic deposits, Nature 463 (2010) 934.
[118] F. Delarue, F. Robert, K. Sugitani, R. Tartèse, R. Duhamel, S. Derenne, Nitrogenisotope signatures of microfossils suggest aerobic metabolism 3.0 Gyr ago,Geochemical Perspectives Letters 7 (2018) 32–36.
[119] C. House, D. Oehler, K. Sugitani, K. Mimura, Carbon isotopic analyses of ca. 3.0 Gamicrostructures imply planktonic autotrophs inhabited Earth's early oceans,Geology 41 (2013) 651–654.
[120] K. Lepot, K. Benzerara, G.E. Brown Jr., P. Philippot, Microbially influenced for-mation of 2,724-million-year-old stromatolites, Nat. Geosci. 1 (2008) 118.
[121] K. Lepot, K. Benzerara, N. Rividi, M. Cotte, G.E. Brown Jr., P. Philippot, Organicmatter heterogeneities in 2.72 Ga stromatolites: alteration versus preservation bysulfur incorporation, Geochem. Cosmochim. Acta 73 (2009) 6579–6599.
[122] K. Lepot, K.H. Williford, P. Philippot, C. Thomazo, T. Ushikubo, K. Kitajima,S. Mostefaoui, J.W. Valley, Extreme 13C-depletions and organic sulfur contentargue for S-fueled anaerobic methane oxidation in 2.72 Ga old stromatolites,Geochem. Cosmochim. Acta 244 (2018) 522–547.
[123] D.T. Flannery, R.E. Summons, M.R. Walter, Archean Lakes as Analogues forHabitable Martian Paleoenvironments. From Habitability to Life on Mars, Elsevier,2018.
[124] J. Hayes, Geochemical evidence bearing on the origin of aerobiosis, a speculativehypothesis, The Earth's Earliest Biosphere: its Origin and Evolution, 1983.
[125] J. Hayes, Global methanotrophy at the Archean-Proterozoic transition, Early lifeon Earth (1994) 220–236.
[126] D.T. Flannery, A.C. Allwood, M.J. Van Kranendonk, Lacustrine facies dependenceof highly 13C-depleted organic matter during the global age of methanotrophy,Precambrian Res. 285 (2016) 216–241.
[127] S.P. Slotznick, W.W. Fischer, Examining archean methanotrophy, Earth Planet.Sci. Lett. 441 (2016) 52–59.
[128] J. Marin‐Carbonne, L. Remusat, M. Sforna, C. Thomazo, P. Cartigny, P. Philippot,Sulfur isotope's signal of nanopyrites enclosed in 2.7 Ga stromatolitic organic re-mains reveal microbial sulfate reduction, Geobiology 16 (2018) 121–138.
[129] D.A. Fike, C.L. Gammon, W. Ziebis, V.J. Orphan, Micron-scale mapping of sulfurcycling across the oxycline of a cyanobacterial mat: a paired nanoSIMS and CARD-FISH approach, ISME J. 2 (2008) 749.
[130] D.A. Fike, N. Finke, J. Zha, G. Blake, T.M. Hoehler, V.J. Orphan, The effect ofsulfate concentration on (sub) millimeter-scale sulfide δ34S in hypersaline cya-nobacterial mats over the diurnal cycle, Geochem. Cosmochim. Acta 73 (2009)6187–6204.
[131] J. Krissansen-Totton, R. Buick, D.C. Catling, A statistical analysis of the carbonisotope record from the Archean to Phanerozoic and implications for the rise ofoxygen, Am. J. Sci. 315 (2015) 275–316.
[132] J.R. Havig, T.L. Hamilton, A. Bachan, L.R. Kump, Sulfur and carbon isotopicevidence for metabolic pathway evolution and a four-stepped Earth system pro-gression across the Archean and Paleoproterozoic, Earth Sci. Rev. 174 (2017)1–21.
[133] T.W. Lyons, C.T. Reinhard, G.D. Love, S. Xiao, Geobiology of the Proterozoic Eon,Fundamentals of Geobiology, 2012, pp. 371–402.
[134] J. Farquhar, H. Bao, M. Thiemens, Atmospheric influence of Earth's earliest sulfurcycle, Science 289 (2000) 756–758.
[135] A. Bekker, Great Oxygenation Event, Encyclopedia of astrobiology, 2014, pp. 1–9.[136] G. Izon, A.L. Zerkle, I. Zhelezinskaia, J. Farquhar, R.J. Newton, S.W. Poulton,
J.L. Eigenbrode, M.W. Claire, Multiple oscillations in Neoarchaean atmosphericchemistry, Earth Planet. Sci. Lett. 431 (2015) 264–273.
[137] G. Luo, S. Ono, N.J. Beukes, D.T. Wang, S. Xie, R.E. Summons, Rapid oxygenationof Earth's atmosphere 2.33 billion years ago, Science Advances 2 (2016)e1600134.
[138] A.L. Zerkle, S.W. Poulton, R.J. Newton, C. Mettam, M.W. Claire, A. Bekker,C.K. Junium, Onset of the aerobic nitrogen cycle during the great oxidation event,Nature 542 (2017) 465.
[139] G. Luo, C.K. Junium, G. Izon, S. Ono, N.J. Beukes, T.J. Algeo, Y. Cui, S. Xie,R.E. Summons, Nitrogen fixation sustained productivity in the wake of the pa-laeoproterozoic great oxygenation event, Nat. Commun. 9 (2018) 978.
[140] E. Stüeken, R. Buick, A. Schauer, Nitrogen isotope evidence for alkaline lakes onlate Archean continents, Earth Planet. Sci. Lett. 411 (2015) 1–10.
[141] W.W. Fischer, S. Schroeder, J.P. Lacassie, N.J. Beukes, T. Goldberg, H. Strauss,U.E. Horstmann, D.P. Schrag, A. Knoll, Isotopic constraints on the late archeancarbon cycle from the transvaal supergroup along the western margin of thekaapvaal Craton, South Africa, Precambrian Res. 169 (2009) 15–27.
[142] R. Buick, B. Rasmussen, B. Krapez, Archean oil: evidence for extensive hydro-carbon generation and migration 2.5-3.5 Ga, AAPG (Am. Assoc. Pet. Geol.) Bull.82 (1998) 50–69.
[143] B. Rasmussen, Evidence for pervasive petroleum generation and migration in 3.2and 2.63 Ga shales, Geology 33 (2005) 497–500.
[144] D.J. Des Marais, H. Strauss, R.E. Summons, J. Hayes, Carbon isotope evidence forthe stepwise oxidation of the Proterozoic environment, Nature 359 (1992) 605.
[145] D.J. Des Marais, Isotopic evolution of the biogeochemical carbon cycle during thePrecambrian, Rev. Mineral. Geochem. 43 (2001) 555–578.
[146] J.L. Eigenbrode, K.H. Freeman, R.E. Summons, Methylhopane biomarker hydro-carbons in Hamersley Province sediments provide evidence for Neoarcheanaerobiosis, Earth Planet. Sci. Lett. 273 (2008) 323–331.
J. Alleon and R.E. Summons Free Radical Biology and Medicine xxx (xxxx) xxx–xxx
9
[147] J. Li, S. Bernard, K. Benzerara, O. Beyssac, T. Allard, J. Cosmidis, J. Moussou,Impact of biomineralization on the preservation of microorganisms during fossi-lization: an experimental perspective, Earth Planet. Sci. Lett. 400 (2014) 113–122.
[148] A. Picard, A. Kappler, G. Schmid, L. Quaroni, M. Obst, Experimental diagenesis oforgano-mineral structures formed by microaerophilic Fe (II)-oxidizing bacteria,Nat. Commun. 6 (2015) 6277.
[149] A. Picard, M. Obst, G. Schmid, F. Zeitvogel, A. Kappler, Limited influence of Si onthe preservation of Fe mineral‐encrusted microbial cells during experimental di-agenesis, Geobiology 14 (2015) 276–292.
[150] J. Miot, S. Bernard, M. Bourreau, F. Guyot, A. Kish, Experimental maturation ofArchaea encrusted by Fe-phosphates, Sci. Rep. 7 (2017) 16984.
J. Alleon and R.E. Summons Free Radical Biology and Medicine xxx (xxxx) xxx–xxx
10