occurrence, production, and applications of gellan: current state and perspectives
TRANSCRIPT

MINI-REVIEW
Occurrence, production, and applications of gellan: currentstate and perspectives
Arsenio M. Fialho & Leonilde M. Moreira &
Ana Teresa Granja & Alma O. Popescu &
Karen Hoffmann & Isabel Sá-Correia
Received: 2 March 2008 /Revised: 3 April 2008 /Accepted: 5 April 2008 /Published online: 28 May 2008# Springer-Verlag 2008
Abstract Bacterial exopolysaccharides (EPS) are productsof biotechnology that are of high interest due to theirrheological properties. This is the case of sphingans, agroup of structurally related EPS secreted by members ofthe genus Sphingomonas. Among these, gellan is amultifunctional gelling agent produced in high yields bythe non-pathogenic strain Sphingomonas elodea ATCC31461. In its native form, gellan is a linear anionic EPSbased on a tetrasaccharide repeat unit composed of twomolecules of D-glucose, one of L-rhamnose and one of D-glucuronic acid. The native gellan is partially esterified withacyl substituents (1 mol of glycerate and 0.5 mol of acetate)per repeat unit. Gellan has unique characteristics and hasmany applications, particularly in the food, pharmaceutical,and biomedical fields. This review summarizes currentknowledge on the structure and properties of gellan andprovides details about the biosynthesis of this exopolysac-charide. In addition, a highlight of the importance of gellanin industrial and medicinal applications is given.
Keywords Bacterial exopolysaccharides . Sphingans .
Gellan . Sphingomonas elodea
Sphingans: occurrence and structure
The ‘sphingans’ are structurally related exopolysaccharides(EPS) secreted by members of the genus Sphingomonas
(Pollock 1993). Strains of this genus are widely distributedin nature, having been isolated from many differentterrestrial and water habitats, as well as from plant rootsystems, clinical specimens, and other sources (White et al.1996; Reina et al. 1991). They have glycosphingolipids intheir outer membranes instead of lipopolysaccharide suchas in other Gram-negative bacteria (Kawasaki et al. 1994).Due to their biodegradative and biosynthetic capabilities,Sphingomonas have been utilized for a wide range ofbiotechnological applications, from bioremediation of en-vironmental contaminants (Coppotelli et al. 2008) toproduction of extracellular polymers such as sphingans(Pollock 1993). Members of the genus Sphingomonas andthe sphingans they produce include Sphingomonas elodea,ATCC 31461, which produces gellan (S-60); Sphingomo-nas sp. ATCC 31555, which produces welan (S-130);Sphingomonas sp. ATCC 31961, which produces rhamsan(S-194); Sphingomonas sp. ATCC 53159, which producesdiutan (S-657); and Sphingomonas sp. ATCC 31554,Sphingomonas sp. ATCC 31853, and Sphingomonas sp.ATCC 21423, which produce three different as yetunnamed polysaccharide, S-88, S-198, and S-7, respective-ly. The structures of all these sphingans polysaccharides areillustrated in Fig. 1. A phylogenetic analysis based on the16S ribosomal-RNA gene sequences of sphingan-producingstrains, as well as other Sphingomonas species and otherreference organisms, represented in Fig. 1a, indicates thatthe Sphingomonas strains that secrete sphingans clustertogether and are apart from the other representativemembers of the genus Sphingomonas.
Sphingans share the same linear tetrasaccharide back-bone structure (–X–glucose–glucuronic acid–glucose–X,where X is either L-rhamnose or L-mannose) to whichdistinct side groups are attached (Fig. 1b; Pollock 1993).The physical properties of sphingans vary considerably,
Appl Microbiol Biotechnol (2008) 79:889–900DOI 10.1007/s00253-008-1496-0
A. M. Fialho (*) : L. M. Moreira :A. T. Granja :A. O. Popescu :K. Hoffmann : I. Sá-CorreiaIBB—Institute for Biotechnology and Bioengineering,Centre for Biological and Chemical Engineering,Instituto Superior Técnico,Av. Rovisco Pais,1049-001 Lisbon, Portugale-mail: [email protected]
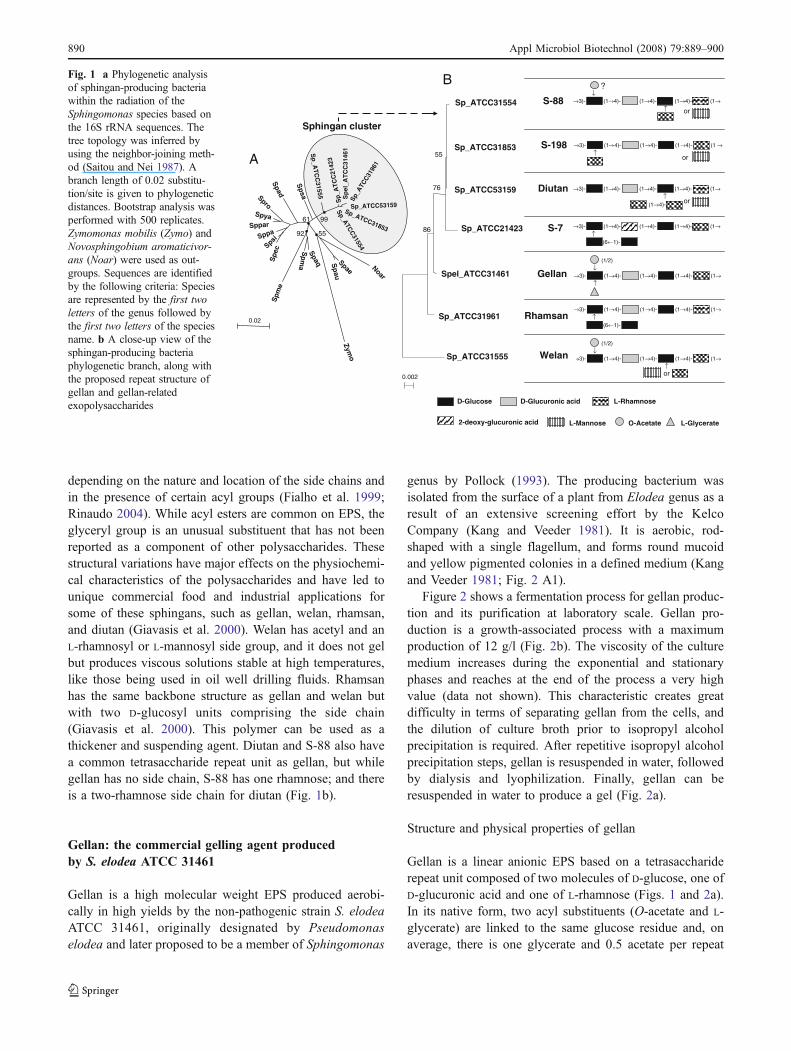
depending on the nature and location of the side chains andin the presence of certain acyl groups (Fialho et al. 1999;Rinaudo 2004). While acyl esters are common on EPS, theglyceryl group is an unusual substituent that has not beenreported as a component of other polysaccharides. Thesestructural variations have major effects on the physiochemi-cal characteristics of the polysaccharides and have led tounique commercial food and industrial applications forsome of these sphingans, such as gellan, welan, rhamsan,and diutan (Giavasis et al. 2000). Welan has acetyl and anL-rhamnosyl or L-mannosyl side group, and it does not gelbut produces viscous solutions stable at high temperatures,like those being used in oil well drilling fluids. Rhamsanhas the same backbone structure as gellan and welan butwith two D-glucosyl units comprising the side chain(Giavasis et al. 2000). This polymer can be used as athickener and suspending agent. Diutan and S-88 also havea common tetrasaccharide repeat unit as gellan, but whilegellan has no side chain, S-88 has one rhamnose; and thereis a two-rhamnose side chain for diutan (Fig. 1b).
Gellan: the commercial gelling agent producedby S. elodea ATCC 31461
Gellan is a high molecular weight EPS produced aerobi-cally in high yields by the non-pathogenic strain S. elodeaATCC 31461, originally designated by Pseudomonaselodea and later proposed to be a member of Sphingomonas
genus by Pollock (1993). The producing bacterium wasisolated from the surface of a plant from Elodea genus as aresult of an extensive screening effort by the KelcoCompany (Kang and Veeder 1981). It is aerobic, rod-shaped with a single flagellum, and forms round mucoidand yellow pigmented colonies in a defined medium (Kangand Veeder 1981; Fig. 2 A1).
Figure 2 shows a fermentation process for gellan produc-tion and its purification at laboratory scale. Gellan pro-duction is a growth-associated process with a maximumproduction of 12 g/l (Fig. 2b). The viscosity of the culturemedium increases during the exponential and stationaryphases and reaches at the end of the process a very highvalue (data not shown). This characteristic creates greatdifficulty in terms of separating gellan from the cells, andthe dilution of culture broth prior to isopropyl alcoholprecipitation is required. After repetitive isopropyl alcoholprecipitation steps, gellan is resuspended in water, followedby dialysis and lyophilization. Finally, gellan can beresuspended in water to produce a gel (Fig. 2a).
Structure and physical properties of gellan
Gellan is a linear anionic EPS based on a tetrasacchariderepeat unit composed of two molecules of D-glucose, one ofD-glucuronic acid and one of L-rhamnose (Figs. 1 and 2a).In its native form, two acyl substituents (O-acetate and L-glycerate) are linked to the same glucose residue and, onaverage, there is one glycerate and 0.5 acetate per repeat
0.02
Zymo
Noar
Spae
Spau
Spaq
Sp
ma
Spm
eS
pecSpsiSppa
Sppar
Spya
SproSpad
Spsa
Sp
_AT
CC
31555
Sp
_AT
CC
2142
3
Sp
el_A
TC
C31
461
Sp_A
TCC
3196
1
Sp_ATCC53159Sp_ATCC31853
Sp_ATC
C31554
Sp_ATCC31555
Sp_ATCC31961
Spel_ATCC31461
Sp_ATCC21423
Sp_ATCC53159
Sp_ATCC31853
Sp_ATCC31554
0.002
A
B
→ 3)- (1→ 4)- (1→ 4)- (1→ 4)- (1→
(1/2)↓
↑
→ 3)- (1→ 4)- (1→ 4)- (1→ 4)- (1→
(1/2)↓
↑ or
→ 3)- (1→ 4)- (1→ 4)- (1→ 4)- (1→ ↑
(6←1)-
→ 3)- (1 →4)- (1→ 4)- (1 →4)- (1 →↑
↑
or
?
→ 3)- (1 →4)- (1→ 4)- (1 →4)- (1→ ↑ or
↓
Gellan
S-198
Rhamsan
Welan
S-88
Diutan → 3)- (1→ 4)- (1→ 4)- (1→ 4)- (1→
or(1→ 4)-
S-7 → 3)- (1→ 4)- (1→ 4)- (1→ 4)- (1→ ↑
(6←1)-
99
55
61
92 86
76
55
Sphingan cluster
D-Glucose D-Glucuronic acid L-Rhamnose
2-deoxy-glucuronic acid L-Mannose O-Acetate L-Glycerate
Fig. 1 a Phylogenetic analysisof sphingan-producing bacteriawithin the radiation of theSphingomonas species based onthe 16S rRNA sequences. Thetree topology was inferred byusing the neighbor-joining meth-od (Saitou and Nei 1987). Abranch length of 0.02 substitu-tion/site is given to phylogeneticdistances. Bootstrap analysis wasperformed with 500 replicates.Zymomonas mobilis (Zymo) andNovosphingobium aromaticivor-ans (Noar) were used as out-groups. Sequences are identifiedby the following criteria: Speciesare represented by the first twoletters of the genus followed bythe first two letters of the speciesname. b A close-up view of thesphingan-producing bacteriaphylogenetic branch, along withthe proposed repeat structure ofgellan and gellan-relatedexopolysaccharides
890 Appl Microbiol Biotechnol (2008) 79:889–900
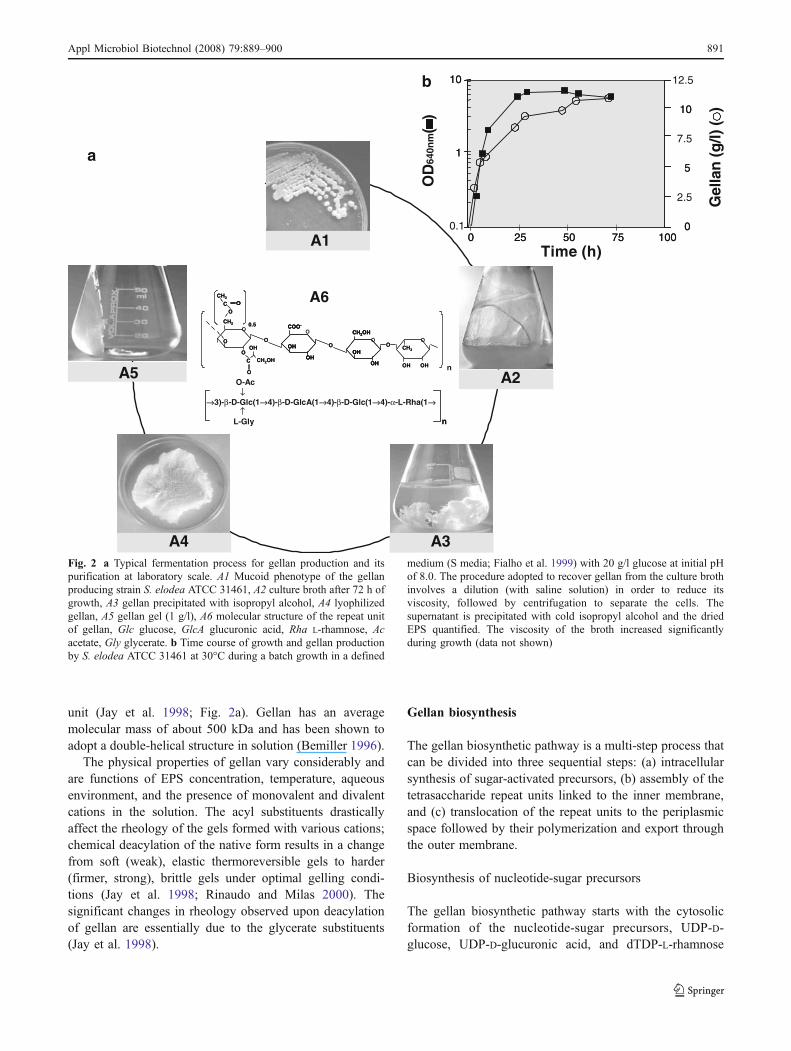
unit (Jay et al. 1998; Fig. 2a). Gellan has an averagemolecular mass of about 500 kDa and has been shown toadopt a double-helical structure in solution (Bemiller 1996).
The physical properties of gellan vary considerably andare functions of EPS concentration, temperature, aqueousenvironment, and the presence of monovalent and divalentcations in the solution. The acyl substituents drasticallyaffect the rheology of the gels formed with various cations;chemical deacylation of the native form results in a changefrom soft (weak), elastic thermoreversible gels to harder(firmer, strong), brittle gels under optimal gelling condi-tions (Jay et al. 1998; Rinaudo and Milas 2000). Thesignificant changes in rheology observed upon deacylationof gellan are essentially due to the glycerate substituents(Jay et al. 1998).
Gellan biosynthesis
The gellan biosynthetic pathway is a multi-step process thatcan be divided into three sequential steps: (a) intracellularsynthesis of sugar-activated precursors, (b) assembly of thetetrasaccharide repeat units linked to the inner membrane,and (c) translocation of the repeat units to the periplasmicspace followed by their polymerization and export throughthe outer membrane.
Biosynthesis of nucleotide-sugar precursors
The gellan biosynthetic pathway starts with the cytosolicformation of the nucleotide-sugar precursors, UDP-D-glucose, UDP-D-glucuronic acid, and dTDP-L-rhamnose
1
10
0 25 50 75 1000
5
10
0.1
1
10
0 25 50 75 1000
2.5
5
7.5
10
12.5
Time (h)
Gel
lan
(g
/l) (
) O
D64
0nm
( )
A6
A1
A2
A3A4
A5
O
O
CH2
C
O
OH
CH2OH
OC O
CH3
0.5
OOH
OH
COO-
OH
OH
CH2OH
O
OH OH
CH3O
O
O
O
CH2
C
O
OH
CH2OH
OC OO
CH3
0.5
OOH
OH
COO-
O
OH
OH
COO-
OH
OH
CH2OHO
OH
OH
CH2OH
OO
OH OH
CH3O
n
O-Ac ↓
→ 3)-β-D-Glc(1→ 4)-β-D-GlcA(1→ 4)-β-D-Glc(1→ 4)-α -L-Rha(1→ ↑
L-Gly nn
b
a
Fig. 2 a Typical fermentation process for gellan production and itspurification at laboratory scale. A1 Mucoid phenotype of the gellanproducing strain S. elodea ATCC 31461, A2 culture broth after 72 h ofgrowth, A3 gellan precipitated with isopropyl alcohol, A4 lyophilizedgellan, A5 gellan gel (1 g/l), A6 molecular structure of the repeat unitof gellan, Glc glucose, GlcA glucuronic acid, Rha L-rhamnose, Acacetate, Gly glycerate. b Time course of growth and gellan productionby S. elodea ATCC 31461 at 30°C during a batch growth in a defined
medium (S media; Fialho et al. 1999) with 20 g/l glucose at initial pHof 8.0. The procedure adopted to recover gellan from the culture brothinvolves a dilution (with saline solution) in order to reduce itsviscosity, followed by centrifugation to separate the cells. Thesupernatant is precipitated with cold isopropyl alcohol and the driedEPS quantified. The viscosity of the broth increased significantlyduring growth (data not shown)
Appl Microbiol Biotechnol (2008) 79:889–900 891

(Fig. 3). The enzymes required for the synthesis of thesesugar nucleotides are phosphoglucomutase (PgmG), UDP-glucose pyrophosphorylase (UgpG), UDP-glucose dehy-drogenase (UgdG), TDP-glucose pyrophosphorylase(RmlA), dTDP-D-glucose-4,6-dehydratase (RmlB), dTDP-6-deoxy-D-glucose-3,5-epimerase (RmlC), and dTDP-6-deoxy-L-mannose dehydrogenase (RmlD; Fig. 3; Sá-Correiaet al. 2002). Glucose-1-phosphate occupies a key positionfrom which two routes derive, one leading to UDP-D-glucose and UDP-D-glucuronic acid and the other leadingto dTDP-L-rhamnose (Fig. 3). The identification andbiochemical characterization of genes/enzymes involved inthe formation of the following gellan precursors werecarried out: the pgmG gene, encoding a phosphoglucomu-tase (EC 5.4.2.2), which catalyses the reversible conversionof glucose-6-phosphate into glucose-1-phosphate (Videiraet al. 2000); the ugpG gene, encoding a glucose-1-phosphate uridilyltransferase (or UDP-glucose pyrophos-phorylase; EC 2.7.7.9), which catalyses the reversibleconversion of glucose-1-phosphate and UTP into UDP-D-glucose and diphosphate (Marques et al. 2003); the ugdGgene, encoding a UDP-glucose dehydrogenase, whichconverts UDP-D-glucose into UDP-D-glucuronic acid(Granja et al. 2007); and rmlA, the first gene of the 4-generml cluster, which encodes a glucose-1-phosphate thymidi-lyltransferase (or TDP-glucose pyrophosphorylase; EC2.7.7.24) that converts glucose-1-phosphate and TTP intoTDP-D-glucose, necessary for the formation of dTDP-L-rhamnose (Silva et al. 2005; Fig. 3).
The genes pgmG, ugpG, and ugdG, required for theformation of the gellan precursors UDP-D-glucose and UDP-D-glucuronic acid, are not colocalized and are located outsidethe gellan biosynthetic gel clusters (Fig. 4a; Sá-Correia et al.2002). These gel clusters, described below, comprise genes
coding for enzymes involved in the synthesis of dTDP-L-rhamnose (RmlA, B, C, D), specific glycosyltransferases,and proteins required for gellan polymerization and export(Sá-Correia et al. 2002; Harding et al. 2004).
Assembly of the tetrasaccharide unit, polymerization,and export of the polymer
The synthesis of the sugar precursors is followed by theformation of the repeat unit by sequential transfer of thesugar donors to an activated lipid carrier by committedglycosyltransferases and by gellan polymerization andexport (Fig. 4). Most of these gellan-specific processes arecatalyzed by enzymes encoded by the gel clusters of genesthat contain at least 22 genes (Fig. 4a; Harding et al. 2004;Sá-Correia et al. 2002).
Gellan tetrasaccharide repeat units are assembled on alipid carrier comprising the C55-isoprenylphosphate carrier(Fig. 4b, step 2). The priming glycosyltransferase isencoded by the gene gelB, which is homologous to spsBfrom Sphingomonas sp. ATCC 31554 (Harding et al. 2004).SpsB was demonstrated to be a glucosyl-isoprenylphos-phate transferase that transfers glucose-1-phosphate fromUDP-glucose to the lipid carrier (Pollock et al. 1994;Yamazaki et al. 1996). The addition of the second sugar,glucuronic acid, from UDP-glucuronic acid into theglucosyl-α-pyrophosphorylpolyprenol intermediate is cata-lyzed by GelK (Videira et al. 2001). By transferring genesbetween related strains of Sphingomonas and by observingwhether sphingan synthesis was restored by the polysac-charide-negative mutants, Pollock et al. (1998) proposedthe gene spsL for encoding a glucosyl-(β1–4)-glucuronosyltransferase. Due to the strong homology between spsL andgelL genes, it is possible that the glycosyltransferase GelL
Glucose-1-P
PhosphoglucomutasepgmG, EC 5.4.2.2
UDP-D-Glucose
dTDP-D-GlucoseGlucose-6-P
UDP-D-Glucuronic acid
UDP-glucose pyrophosphorylaseugpG, EC 2.7.7.9
UDP-glucose dehydrogenase
ugdG, EC 1.1.1.22
dTDP-glucose pyrophosphorylasermlA; EC 2.7.7.24
dTDP-D-glucose-4,6-dehydratase
rmlB; EC 4.2.1.46
dTDP-6-deoxy-D-glucose-3,5-epimerase
rmlC; EC: 5.1.3.13
dTDP-6-deoxy-L-mannose dehydrogenase
rmlD; EC: 1.1.1.133
dTDP-L-Rhamnose
GlucoseFig. 3 Proposed pathwayleading to the nucleotide-sugarprecursors, UDP-D-glucose,UDP-D-glucuronic acid, anddTDP-L-rhamnose, involved ingellan biosynthesis with glu-cose as the substrate(Sá-Correia et al. 2002)
892 Appl Microbiol Biotechnol (2008) 79:889–900

catalyzes the addition of the third sugar (glucose) to therepeat unit. Finally, the enzyme that adds the last sugar tothe tetrasaccharide repeat unit, rhamnose, is putativelyencoded by the remaining gene with homology to glyco-syltransferases, gelQ, although no experimental evidence isavailable (Harding et al. 2004; Sá-Correia et al. 2002). Theassembly of the repeat unit will only be complete after theaddition of the substituent groups, acetate and glycerate, tothe first glucose residue. These reactions are catalyzed bytwo enzymes having acetyltransferase and glyceryltransfer-ase activities, respectively. Despite the mutations intro-duced into genes with unknown function from the gel
cluster I, no apparent negative effects on gellan productionor changes in its composition were detected (Harding et al.2004). These genes encoding for the repeat-unit-modifyingenzymes are probably elsewhere in the genome of S. elodeaATCC 31461.
The next step in polysaccharide biosynthesis is thepolymerization of the repeat units to form longer chains.The general Wzy-dependent pathway for polymerization ofpolysaccharides in bacteria proposes a common mechanismin which a transporter (termed Wzx) translocates the repeatunit from the cytoplasmic side to the periplasmic side of theinner membrane, and a polymerase (termed Wzy) joins
Cyto
pla
sm
Peri
pla
sm
IM
PgmG; UgpG; UgdG;
Rml A, B, C, D
UDP-Glucose UDP-Glucuronic acid dTDP-L-Rhamnose Lipid carrier Phosphate L-Glycerate O - Acetate
OM
BStep 1 Step 2 Step 3
Synthesis of nucleotide
sugar precursors
Assembly of
tetrasaccharide repeat-units Polymerization and export
GelBGelK
GelLGelQ
?
GelSGelG GelC GelE
GelD
Gellan
gelQ
gelI
gelK
gelL
gelJ
gelF
gelD
gelC
gelE
gelM
gelN
atrD
atrB
gelB
rmlA
rmlC
rmlB
rmlD
Orf
302
pgm
G
ugpG
ugdG
gel cluster_region I A
Polymerization/ Export
Glycosyltransferase ABC - transporterUnknown function dTDP – L-rhamnose biosynthesis
Phosphoglucomutase UDP-Glucose pyrophosphorylase UDP-Glucose dehydrogenase
gelR
gelS
gelG
Lyase
gel cluster
region IV
Translocase
Gellan
region III
gelA
Two component regulatory protein
Fig. 4 a Organization of the gellan biosynthetic gel clusters (regions Iand IV) and gellan region III from S. elodea ATCC 31461 (Sá-Correiaet al. 2002; Harding et al. 2004). Arrows represent ORFs, and genedesignations are indicated above the arrows. Known or putative genefunctions are indicated. b A model for biosynthesis and assembly ofgellan. Starting at the left side, activated sugar nucleotides aresequentially added to a lipid carrier to form repeat units of thepolysaccharide. The undecaprenyl pyrophosphate (und-PP)-linkedtetrasaccharide repeat unit is assembled at the interface between the
cytoplasm and the inner membrane by sequential activity of GelB,GelK, GelL, and GelQ glycosyltransferases. These newly synthesizedund-PP-linked repeat units are then translocated across the membranein a process requiring GelS. These repeat units provide the substratefor GelG polymerization and also require GelC/GelE proteins,possibly by forming oligomers that connect polymerization to exportand regulate polysaccharide chain length. Gellan chains are thenexported by GelD, which may act as a channel. IM Inner membrane,OM outer membrane
Appl Microbiol Biotechnol (2008) 79:889–900 893

together the repeat units by successive transfer of the nascentpolymer from its lipid carrier (which is then recycled) to thereducing end of the new lipid-linked repeat unit (Whitfield2006). The determination of length distribution of thesynthesized polysaccharide chains is controlled by a familyof proteins termed polysaccharide co-polymerases (PCPs).PCPs have, in addition to the membrane/periplasmicdomain, a cytoplasmic domain of protein tyrosine kinases,and the prototype of this family is Wzc from Escherichiacoli (Whitfield 2006). The export of the polysaccharide ismediated by integral outer membrane proteins from theouter membrane auxiliary (OMA) family, which form aprotein channel through which the polysaccharide chainsaccess the cell surface (Collins and Derrick 2007; Whitfield2006). The genes involved in the process of gellanpolymerization and export to the cell surface have beenidentified (Harding et al. 2004; Moreira et al. 2004; Sá-Correia et al. 2002). The genes encoding the gellanbiosynthetic protein homologues to Wzx and Wzy arelocated in the gel cluster region IV and are named gelS andgelG, respectively (Fig. 4a: Harding et al. 2004). Yamazakiet al. (1996) reported that inactivation of spsS or spsG geneexpression blocked sphingan S-88 synthesis, indicating thatthese genes are essential for polysaccharide production.Due to the homologies to SpsS and SpsG, we propose GelSas the translocase of the repeat units and GelG as thepolymerase (Fig. 4b, step 3).
As referred above, the extent of polysaccharide chainlength is dictated by the PCP enzymes, which interact bothwith the polymerase enzyme and the secretion machinery,as was shown recently by determination of 3D structure ofsome PCP members (Collins et al. 2007; Tocilj et al. 2008;Whitfield 2006). PCP enzymes involved in capsular or EPSsystems in Gram-negative bacteria are a single peptide withthe N-terminal being probably involved in chain lengthregulation and with a tyrosine kinase activity domain in theC-terminal region (Whitfield 2006). The role of tyrosinephosphorylation remains unknown, but the lack of phos-phorylation in proteins such as Wzc from E. coli or ExoPfrom Sinorhizobium meliloti results in drastic reduction ofhigh molecular weight polysaccharide biosynthesis(Niemeyer and Becker 2001; Wugeditsch et al. 2001).The PCP enzyme in S. elodea is encoded by the genes gelCand gelE. Contrastingly to Gram-negative bacteria, the pairGelC/GelE exhibit a genetic organization similar to the onedescribed for Gram-positive bacteria, being composed oftwo independent polypeptides (Fig. 4b, step 3). GelC showsthe typical PCP N- and C-terminal transmembrane helicesseparated by a segment with a predicted coiled-coil regionlocated in the periplasm (Moreira et al. 2004). The deletionof gelC or gelE genes from S. elodea resulted in a non-mucoid phenotype and the total absence of gellan from theculture supernatant.
The lack of gellan biosynthesis in the absence of GelEalso suggests an important role for this protein, probably byregulating GelC activity. In fact, GelE seems to have ATP-binding activity, but the introduction of several pointmutations in the Walker A or Walker B ATP-binding motifsdid not prevent wild-type levels of gellan biosynthesis.Despite the several attempts, it was not possible to assess atyrosine autokinase activity to GelE (Moreira et al. 2004).Nevertheless, the tyrosine residue at position 198 of GelEappears to be essential for the synthesis of high molecularweight gellan since in its absence, both the amount andviscosity of the EPS produced were much reduced whencompared to the wild-type polymer level production andrheological properties. Whether the tyrosine residue atposition 198 is important for the structure/stability of GelEin vivo or is important for phosphorylation remains to beclarified. A computational analysis of GelE structurereveals a potential amphiphatic helix at the C-terminalregion, which may be involved in the association of GelE tothe plasma membrane and therefore interacting with GelC.
The next step, the export process of gellan chains, ismediated by the OMA protein homologue encoded by thegene gelD. The deletion of gelD from S. elodea preventsgellan biosynthesis, consistent with the crucial role of thisprotein in EPS export (Fig. 4b, step 3). Overall, the proteinsintervenient in the polymerization and secretion of gellanshare sequence and possibly structural similarities to othercharacterized proteins involved in capsular and exopoly-saccharide biosynthesis, namely from E. coli, but the exactmechanisms of action of the Gel proteins still has to bedetermined.
Despite the characterization of many of the gel geneproducts, there are still some genes with an unprovedfunction. Harding et al. (2004) were unable to mutate gelF,concluding that either this mutation was lethal or themutation exert a polarity effect on downstream genes sincegelF is the first gene in what appears to be an operon of sixgenes (Fig. 4a). A mutation in gelI gene affected theamount of gellan produced but not its composition(Harding et al. 2004). Sequence analysis of protein GelIshowed N-terminal lipoprotein attachment site implicating apossible localization at the cell outer membrane. Further-more, GelI sequence shows weak homology to peptidyl-prolyl cis–trans isomerases, which are enzymes involved infolding of proteins located at the periplasm or outermembrane. These predicted characteristics and the pheno-type of decreased gellan production suggests a possibleinvolvement in processing proteins, with a role in gellansecretion. Similarly, a deletion of gelM-gelN resulted inlower amount of gellan and lower broth viscosity, suggest-ing that these proteins might affect the length or attachmentof the polysaccharide (Harding et al. 2004). GelM andGelN are homologous to EpsH and EpsI involved in
894 Appl Microbiol Biotechnol (2008) 79:889–900

methanolan biosynthesis by Methylobacillus sp. strain 12S,which were recently proposed to form a protein exportsorting system, analogous to the LPXTG/sortase system ofGram-positive bacteria (Haft et al. 2006). It is then possiblethat GelM/GelN are involved in protein export, namelyproteins targeted to the periplasm or outer membrane andwith a possible role in gellan secretion/attachment. Thehomologue of GelJ in Sphingomonas sp. ATCC 31554 isessential for sphingan biosynthesis, but its function isunknown (Yamazaki et al. 1996). Similar searches indicatelow identity with AAA ATPases, a family of proteins thatperform chaperone-like functions that assist in the assem-bly, operation, or disassembly of protein complexes. AtrDand AtrB showed homology to the ABC-transporter andmembrane fusion protein, respectively, of a type I secretionsystem. Mutations in each of these genes did not preventsphingan S-88 biosynthesis, but the role, if any, inpolysaccharide production, remains unknown (Yamazakiet al. 1996).
Another gene putatively involved in the regulation ofgellan biosynthesis was identified in another region of theS. elodea genome and was named gel region III (Fig. 4a;Harding et al. 2004). This gene encodes protein GelA,which is homologous to both sensor kinase and responseregulatory proteins, and may encode a positive regulator forgellan biosynthesis. Mutants that map in gel region IIIseemed to be non-mucoid on solid medium and producing alow amount of polysaccharide in liquid medium (Harding etal. 2004).
Comparison of sphingan gene clusters and encoded proteins
The complete sequence of three clusters of genes involvedin sphingan synthesis is presently known. These are thesphingan S-88 sps cluster from Sphingomonas sp. ATCC31554, the gellan gel cluster from S. elodea ATCC 31461,and the diutan dps cluster from Sphingomonas sp. ATCC53159. These clusters share a common core of genes thatincludes all the genes between spsQ/gelQ/dpsQ and rmlD(Fig. 5) and encode products involved in the synthesis ofnucleotide-sugar precursors for L-rhamnose, the glycosyl-transferases required for the four-sugar repeat unit back-bone, and gene products involved in polymerization andexport of the polysaccharide. The genes encoding theenzymes involved in the synthesis of the nucleotide-sugarprecursors for UDP-glucose, UDP-glucuronic acid, andGDP-mannose are elsewhere in the genome of the threestrains.
As referred above, diutan, gellan, and sphingan S-88have the same four-sugar backbone but different sidechains, with one rhamnose for sphingan S-88, tworhamnoses for diutan, and no side chain for gellan. Thisdifference is evident in the amount of glycosyltransferasesrequired for the assembly of the repeat unit. Therefore,gellan needs GelB, GelK, GelL, and GelQ; sphingan S-88needs SpsB, SpsK, SpsL, SpsQ, and a rhamnosyltransferasepossibly encoded by urf31; diutan needs DpsB, DpsK,DpsL, DpsQ, urf31, and another unknown glycosyltransfer-ase (Fig. 5; Coleman et al. 2008). Another difference
Gellan (gel)clusters
S-88 (sps) cluster
Diutan (dps)cluster
gelQ
gelI
gelK
gelL
gelJ
gelF
gelD
gelC
gelE
gelM
gelN
atrD
atrB
gelB
rmlA
rmlC
rmlB
rmlD
urf3
4
spsG
spsS
spsR
spsQ
spsI
spsK
spsL
spsJ
spsF
spsD
spsC
spsE
spsM
spsN
atrD
atrB
spsB
rmlA
rmlC
rmlB
rmlD
urf3
1ur
f34
6884 87
7658
4364
53
8578
8884
89 8671
8986 86 85
Sp_31961
Sp_53159
Sp_31853
Sp_31554
Sp_21423
Spel_31461
Sp_31555
0.002
SphingansRegion IV Region I
dpsG
dpsS
dpsR
dpsQ
dpsI
dpsK
dpsL
dpsJ
dpsF
dpsD
dpsC
dpsE
dpsM
dpsN atrD
atrB
dpsB
rmlA
rmlC
rmlB
rmlD
urf3
1.4
urf3
4
gelR
gelS
gelG
urf3
1
8893 8663
5353
19
7989
7887 88 85
90 8994 97 96 98
8874 76
88
4458 52
dTDP–L-rhamnose biosynthesis Glycosyltransferase Polymerization/ Export ABC -transporter
Unknown functionLyase
Translocase
Protein identity (%)
Protein identity (%)
Fig. 5 Comparative analysis of the exopolysaccharide biosynthesisgene clusters from S. elodea ATCC 31461 (gellan gel cluster; Hardinget al. 2004), Sphingomonas sp. ATCC 31554 (S-88 sps cluster;Yamazaki et al. 1996), and from Sphingomonas sp. ATCC 53159
(diutan dps cluster; Coleman et al. 2008). Known or putative genefunctions are indicated. The percentages show amino acid sequenceidentity between the homologous genes
Appl Microbiol Biotechnol (2008) 79:889–900 895

between the three gene clusters is the location of theputative wzx homologue (spsS, gelS, and dpsS), the putativepolysaccharide polymerase wzy (spsG, gelG, and dpsG),and the putative lyase (spsR, gelR, and dpsR). In bothsphingan S-88 and diutan gene clusters, the genes spsG/spsS/spsR and dpsG/dpsS/dpsR are located in the vicinity ofspsQ/dpsQ, respectively, although with a different organi-zation and orientation. Contrastingly, the gelS/gelG/gelRare located in another region of the genome of S. elodeaATCC 31461 but in the same orientation as in the S-88 spsgene cluster (Fig. 5; Harding et al. 2004).
The comparison of the amino acid sequence of thepredicted proteins from the sps gene cluster to the gel anddps clusters reveals a closer proximity between the sps andthe dps clusters (Fig. 5). This may probably reflect the factthat sphingan S-88 and diutan share more common struc-tural features than gellan and also the fact that Sphigomonassp. ATCC 31554 is more closely related to S. elodea ATCC53159 at the phylogenetic level (Figs. 1 and 5).
Environmental and nutritional factors affectingthe synthesis of gellan
As for many other EPS (Welman and Maddox 2003;Vaningelgem et al. 2004), it is possible to manipulate gellanproduction yield and its chemical composition, structure, andmolecular mass by modifying environmental and nutritionalfactors such as temperature (Martins and Sá-Correia 1994),level of dissolved oxygen (Banik and Santhiagu 2006), andgrowth medium composition, in particular the carbon (Fialhoet al. 1999; Wang et al. 2006; Banik et al. 2007) and nitrogen(West and Strohfus 1998) sources.
Temperature is one of the most important parametersinfluencing the production of gellan. Gellan productionshows a maximal yield at 20–25°C, which is not coincidentwith the optimal range for growth and for maximal activityof biosynthetic enzymes (30–35°C; Martins and Sá-Correia1994). It was observed by Martins and Sá-Correia (1994)that the gellans from the low temperature range (20–25°C)exhibited higher molecular mass compared with those fromthe high temperature range (30–35°C). This is in agreementwith the proposed mechanism in which slow-growing cellsexhibit much slower cell wall synthesis and thus have adecreased need for isoprenoid phosphate. As a conse-quence, more isoprenoid phosphate could be available forEPS synthesis (Sutherland 2001).
Cheese whey and molasses contain high sugar concen-trations and have been successfully used as raw materialsfor low-cost fermentative production of gellan (Fialho et al.1999; Banik et al. 2007). The results obtained areconsidered of industrial value because the poluent powerof these industrial wastes is greatly reduced. Cheese whey
is the by-product of cheese manufacture and contains mostof the lactose of milk. Comparison of gellan biosynthesisby S. elodea ATCC 31461 in a defined medium containingglucose or lactose (5–30 g/l) as the carbon source and indiluted cheese whey indicates that the alteration of thegrowth medium markedly affects the EPS yield, acylsubstitution level, polymer rheological properties, andsusceptibility to enzymatic degradation (Fialho et al.1999). Cheese whey and glucose-derived gellan had similaracyl contents but differ in their acetate and glycerate levels(Fialho et al. 1999).
Various organic (urea, peptone, soybean) and inorganic(NH4Cl, NH4NO3, NaNO3) nitrogen sources support gellanproduction by S. elodea ATCC 31461 (West and Strouhfus1998). The results revealed that the nature and theconcentration of the nitrogen source markedly affect thegellan yield. It is recognized that the use of organiccomplex sources led to high gellan production in compari-son to ammonium sulphate (West and Strouhfus 1998).
A response surface experimental design was used todetermine the influence of various nutrient supplements ongellan production using molasses. Among the 20 variablestested, statistical analysis revealed that molasses, tryptone,casaminoacid, disodium hydrogen orthophosphate, andmanganese chloride showed significant effect on gellanproduction (Banik et al. 2007).
The role of fermentor hydrodynamics on gellan fermen-tation kinetics and the rheological properties of the EPSsynthesized are also studied. Banik and Santhiagu (2006)reported that aeration exerts a critical influence on theaverage molecular mass of the gellan produced.
Engineering sphingan production
Metabolic engineering represents one promising strategy toproduce modified gellans and/or to increase its production.Because gellan production is a growth-associated processwith a relatively low sugar conversion (40–50%), efforts toincrease its synthesis are likely to require strategies toenhance substrate conversion (Vartak et al. 1995). Thisstrategy was illustrated by Vartak et al (1995) in agenetically engineered S. elodea ATCC 31461 mutant inwhich the zwf gene encoding glucose-6-P-dehydrogenasewas interrupted, envisaging diverting carbon flow towardgellan synthesis. However, the use of this recombinantstrain did not significantly enhance the synthesis of gellan(Vartak et al. 1995). It has also been observed that theoptimal fermentation conditions for producing gellan arefavored by nutrient imbalance of C/N ratio. However, thesesame growth conditions favor the synthesis of considerableamounts of poly-β-hydroxybutyric acid (PHB), an intracel-lular energy storage product and a gellan-competing
896 Appl Microbiol Biotechnol (2008) 79:889–900

product. By random mutagenesis, the synthesis of PHB wasblocked, although no positive effect on gellan synthesis wasobserved (Baird and Cleary 1994).
As described above, significant progresses have beenmade in the genetic characterization of gellan biosynthesis.This allows the opportunity to evaluate the potential ofgenetic engineering for the controlled modification ofgellan production yield and the chemical composition andproperties of the biopolymers produced by recombinantbacteria. A few strategies have been applied to the earlysteps in gellan biosynthesis, i.e., precursor synthesis,aiming to rerouting the carbon flux. The individual over-expression of the pgmG gene (phosphoglucomutase) andthe ugpG gene (UDP-glucose pyrophosphorylase) result inan increase of enzyme-specific activities, which in turnincrease UDP-glucose synthesis in S. elodea ATCC 31461.However, the increased enzyme-specific activities did notsignificantly enhance the synthesis of gellan (Sá-Correia etal. 2002). Interestingly, in another experiment, the simulta-neous overexpression in S. elodea ATCC 31461 of pgmGand ugdG was reported to lead to a 20% increase in gellanyield and EPS viscosity (Sá-Correia et al. 2002). Similarresults was reported for the genetic manipulation of sugarprecursors of the sphingan S-7 synthesized by Sphingomo-nas sp. ATCC 21423 (Thorne et al. 2000).
While precursor synthesis has been proposed as potentialcontrolling points to enhance sphingan production yield,other metabolic engineering attempts were pursued. Thorneet al. (2000) reported that the simultaneous overexpressionof biosynthetic genes from the S-7 cluster, which code forspecific glycosyltransferases, assembly of the lipid-linkedcarbohydrate repeat unit, and secretion of the polymer,caused a 20% increase in the sphingan S-7 yield fromglucose and a larger increase in culture viscosity (Thorne etal. 2000). Recently, Coleman et al (2008) observed that arecombinant Sphingomonas ATCC 53159, harboring aplasmid containing 20 of the 24 biosynthetic genes fromthe diutan cluster, led to a significant increase in EPSviscosity yet only a small increase in diutan synthesis.Although these results are promising, the success of geneticengineering approaches still requires a more detailedunderstanding of sphingans biosynthesis, particularly abetter knowledge of the poorly understood steps and ofthe regulation and bottlenecks of sphingan pathways.
Gellan: a multifunctional gelling agent
Gellan has been approved in the US and in the EU for foodand medical usage as a gelling, stabilizing, and suspendingagent (Giavasis et al 2000). Compared with other poly-saccharides, gellan has many advantages such as anexcellent thermal and acid stability, adjustable gel elasticity
and rigidity, high transparency, and good flavor release.Gellan is commercially available in three forms: no, low,and high acyl content with the respective denominations ofGelrite®, Kelcogel® F, and Kelcogel® LT100. Gelrite® isused as a substitute of agar for the culture of thermophilicbacterial species and in plant tissue culture media (Lin andCasida 1984). Gelrite also has potential environmentalapplications such as in the biodegradation of gasoline(Moslemy et al. 2004) and for the transportation of gel-encapsulated bacteria for bioaugmentation of contaminatedaquifers (Moslemy et al. 2003). Kelcogels are food-gradegellans used as gelling agents in foods (it has E numberE418) and personal care applications (lotions, creams, andtoothpastes), being mainly used as a stabilizer andsuspending agent in a wide variety of applications in thefood industry (Sutherland 2001). Food products thattypically incorporate gellan include dessert gels, icingsand glazes, sauces, puddings, and microwavable foods.
Gellan applications in the biomedical field include itsuse as a pharmaceutical excipient for nasal, ocular, gastric,and colonic drug delivery applications (Sultana et al. 2006;Agnihotri et al. 2006; Jansson et al. 2005). Hydrocolloidbeads based on gellan have been shown to be useful forslow drug release (Gal and Nussinovitch 2007). A gellanfilm has been implanted for insulin delivery in diabetic rats,showing that gellan might be an ideal candidate in thedevelopment of protein delivery systems (Li et al. 2001).Gellan was also proved to be a suitable material for theconstruction of 3D scaffolds for tissue engineering(Ciardelli et al. 2005; Bertoni et al. 2006; Smith et al.2007). Ciardelli et al. (2005) show that gellan is a suitablematerial for the production of poly-(ɛ-caprolactone)-basedblends for tissue engineering scaffolds. To use a modifiedprotein for tissue engineering application, transglutaminasecrosslinked with gelatine has been incorporated in a gellanmatrix to enhance its stability in aqueous media (Bertoni etal. 2006). The first report of gellan alone being used in theimmobilization and culture of rat bone marrow cells wasevaluated by Smith et al. (2007), having shown that usingthe proper media, gellan has potential application as a 3Dculture matrix. This study shows that the gellan 3Dstructure provides a good environment and enhancescellular viability.
Each year, many gellan patents covering a wide range ofapplications are filed or published. A worldwide patentsearch was performed (http://ep.espacenet.com; March,2008) based on the keyword gellan. The search resultedin a list of approximately 890 issued patents and publishedapplications. A title keyword (gellan) search was performedto filter the results obtained to nearly 120 references. Eachof these references was examined, categorized into appli-cation fields, and some of them cited in Table 1. Asmentioned in Table 1, most of the gellan patents cover its
Appl Microbiol Biotechnol (2008) 79:889–900 897
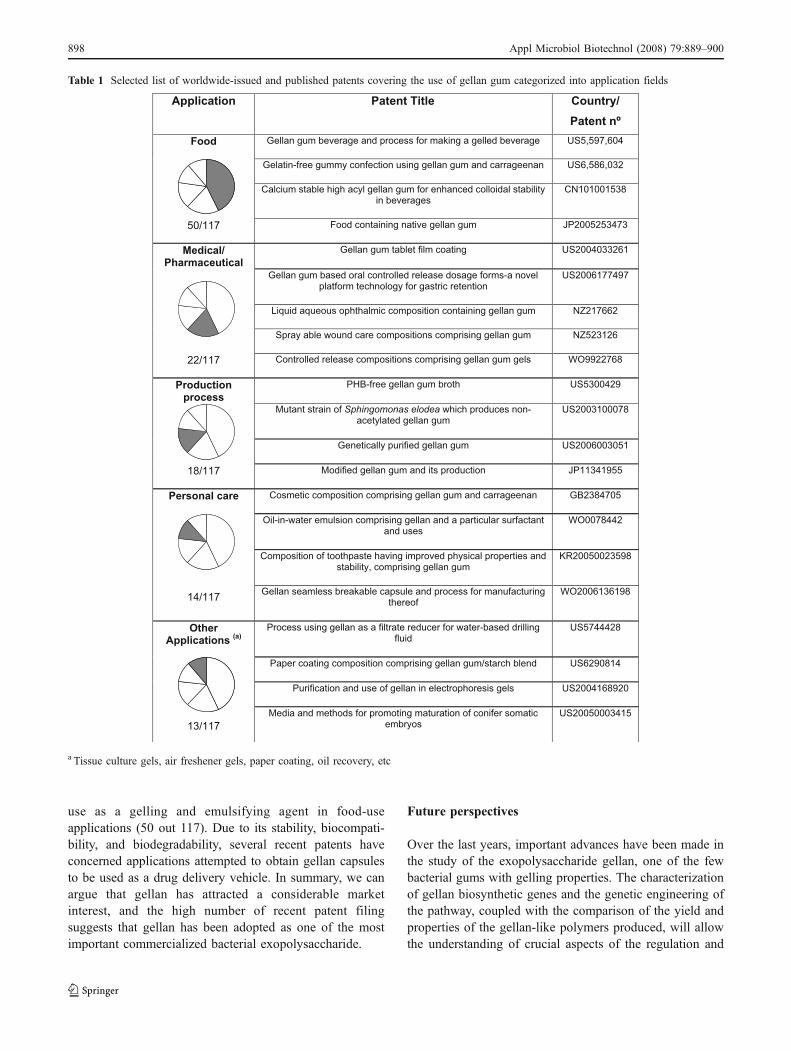
use as a gelling and emulsifying agent in food-useapplications (50 out 117). Due to its stability, biocompati-bility, and biodegradability, several recent patents haveconcerned applications attempted to obtain gellan capsulesto be used as a drug delivery vehicle. In summary, we canargue that gellan has attracted a considerable marketinterest, and the high number of recent patent filingsuggests that gellan has been adopted as one of the mostimportant commercialized bacterial exopolysaccharide.
Future perspectives
Over the last years, important advances have been made inthe study of the exopolysaccharide gellan, one of the fewbacterial gums with gelling properties. The characterizationof gellan biosynthetic genes and the genetic engineering ofthe pathway, coupled with the comparison of the yield andproperties of the gellan-like polymers produced, will allowthe understanding of crucial aspects of the regulation and
Table 1 Selected list of worldwide-issued and published patents covering the use of gellan gum categorized into application fields
Application Country/
Patent nº
Food Gellan gum beverage and process for making a gelled beverage US5,597,604
Gelatin-free gummy confection using gellan gum and carrageenan US6,586,032
Calcium stable high acyl gellan gum for enhanced colloidal stability in beverages
CN101001538
50/117 Food containing native gellan gum JP2005253473
Medical/ Pharmaceutical
Gellan gum tablet film coating US2004033261
Gellan gum based oral controlled release dosage forms-a novel platform technology for gastric retention
US2006177497
Liquid aqueous ophthalmic composition containing gellan gum NZ217662
Spray able wound care compositions comprising gellan gum NZ523126
22/117 Controlled release compositions comprising gellan gum gels WO9922768
Production process
PHB-free gellan gum broth US5300429
Mutant strain of Sphingomonas elodea which produces non-acetylated gellan gum
US2003100078
Genetically purified gellan gum US2006003051
18/117 Modified gellan gum and its production JP11341955
Personal care Cosmetic composition comprising gellan gum and carrageenan GB2384705
Oil-in-water emulsion comprising gellan and a particular surfactant and uses
WO0078442
Composition of toothpaste having improved physical properties and stability, comprising gellan gum
KR20050023598
14/117 Gellan seamless breakable capsule and process for manufacturing
thereof WO2006136198
Other Applications
(a)
Process using gellan as a filtrate reducer for water-based drilling fluid
US5744428
Paper coating composition comprising gellan gum/starch blend US6290814
Purification and use of gellan in electrophoresis gels US2004168920
13/117
Media and methods for promoting maturation of conifer somatic embryos
US20050003415
Patent Title
a Tissue culture gels, air freshener gels, paper coating, oil recovery, etc
898 Appl Microbiol Biotechnol (2008) 79:889–900

bottlenecks of the gellan pathway. This will provideguidance to the engineering of bacterial EPS opening thedoor to their tailoring. Specifically, in the area of healthfood research, it would be of interest to create gellan-relatedelastic polysaccharide that produces gels that melt at 30–35°C (mouth temperature). In the field of medicinalapplications, new and promising gellan applications arebeginning to emerge. For example, gellan was recently provedto be a suitable material for the construction of 3D scaffoldsfor tissue engineering (Smith et al. 2007). Preliminary resultshave also indicated the possibility of using gellan as a drugdeliver vehicle and a stabilizer for bioactive anticancerpeptides (Fialho AM, unpublished results).
Acknowledgments Part of the work in the author’s laboratory wassupported by FEDER, the POCTI Programme, and Fundação para aCiência e a Tecnologia (FCT), Portugal (grants POCTI/BIO/58401/2004, PTDC/BIA-MIC/65210-2006, PTDC/AGR-AAM/66977/2006and PTDC/QUI/67925/2006) and PhD fellowships to A. O. Popescuand K. Hoffmann.
References
Agnihotri SA, Jawalkar SS, Aminabhavi TM (2006) Controlledrelease of cephalexin through gellan gum beads: effect offormulation parameters on entrapment efficiency, size, and drugrelease. Eur J Pharm Biopharm 63:249–261
Baird JK, Cleary JM (1994) PHB-Free gellan gum broth US Patent5300429
Banik RM, Santhiagu A (2006) Improvement in production andquality of gellan gum by Sphingomonas paucimobilis under highdissolved oxygen tension levels. Biotechnol Lett 28:1347–1350
Banik RM, Santhiagu A, Upadhyay SN (2007) Optimization ofnutrients for gellan gum production by Sphingomonas paucimo-bilis ATCC-31461 in molasses based medium using responsesurface methodology. Bioresour Technol 98:792–797
Bemiller JN (1996) Structure–property relationships of water-solublepolysaccharides. J Appl Glycosci 43:377–384
Bertoni F, Barbani N, Giusti P, Ciardelli G (2006) Transglutaminasereactivity with gelatine: perspective applications in tissueengineering. Biotechnol Lett 28:697–702
Ciardelli G, Chiono V, Vozzi G, Pracella M, Ahluwalia A, Barbani N,Cristallini C, Giusti P (2005) Blends of poly-(epsilon-caprolactone)and polysaccharides in tissue engineering applications. Biomacro-molecules 6:1961–1976
Coleman RJ, Patel YN, Harding NE (2008) Identification andorganization of genes for diutan polysaccharide synthesis fromSphingomonas sp. ATCC 53159. J Ind Microbiol Biotechnol35:263–274
Collins RF, Derrick JP (2007) Wza: a new structural paradigm forouter membrane secretory proteins? Trends Microbiol 15:96–100
Collins RF, Beis K, Dong C, Botting CH, McDonnell C, Ford RC,Clarke BR, Whitfield C, Naismith JH (2007) The 3D structure ofa periplasm-spanning platform required for assembly of group 1capsular polysaccharides in Escherichia coli. Proc Natl Acad SciU S A 104:2390–2395
Coppotelli BM, Ibarrolaza A, Del Panno MT, Morelli IS (2008)Effects of the inoculant strain Sphingomonas paucimobilis20006FA on soil bacterial community and biodegradation inphenanthrene-contaminated soil. Microb Ecol 55:173–183
Fialho AM, Martins LO, Donval ML, Leitão JH, Ridout MJ, Jay AJ,Morris VJ, Sá-Correia I (1999) Structures and properties ofgellan polymers produced by Sphingomonas paucimobilis ATCC31461 from lactose compared with those produced from glucoseand from cheese whey. Appl Environ Microbiol 65:2485–2491
Gal A, Nussinovitch A (2007) Hydrocolloid carriers with filler inclusionfor diltiazem hydrochloride release. J Pharm Sci 96:168–178
Giavasis I, Harvey LM, McNeil B (2000) Gellan gum. Crit RevBiotechnol 20:177–211
Granja AT, Popescu A, Marques AR, Sá-Correia I, Fialho AM (2007)Biochemical characterization and phylogenetic analysis of UDP-glucose dehydrogenase from the gellan gum producer Sphingo-monas elodea ATCC 31461. Appl Microbiol Biotechnol76:1319–1327
Haft DH, Paulsen IT, Ward N, Selengut JD (2006) Exopolysaccharide-associated protein sorting in environmental organisms: the PEP-CTERM/EpsH system. Application of a novel phylogeneticprofiling heuristic. BMC Biology 4:29
Harding NE, Patel YN, Coleman RJ (2004) Organization of genesrequired for gellan polysaccharide biosynthesis in Sphingomonaselodea ATCC 31461. J Ind Microbiol Biotechnol 31:70–82
Jansson B, Hägerström H, Fransén N, Edsman K, Björk E (2005) Theinfluence of gellan gum on the transfer of fluorescein dextranacross rat nasal epithelium in vivo. Eur J Pharm Biopharm59:557–564
Jay AJ, Colquhoun IJ, Ridout MJ, Brownsey GJ, Morris VJ, FialhoAM, Leitão JH, Sá-Correia I (1998) Analysis of structure andfunction of gellans with different substitution patterns. CarbohydrPolym 35:179–188
Kang KS, Veeder GT (1981) Fermentation process for preparation ofpolysaccharide S-60. U. S. Patent: 4, 377, 636, 1981
Kawasaki SR, Moriguchi R, Sekiya K, Nakai T, Ono E, Kume K,Kawahara K (1994) The cell envelope structure of the lipopoly-saccharide-lacking gram-negative bacterium Sphingomonas pau-cimobilis. J Bacteriol 176:284–290
Li J, Kamath K, Dwivedi C (2001) Gellan film as an implant forinsulin delivery. J Biomater Appl 15:321–343
Lin CC, Casida LE (1984) GELRITE as a gelling agent in media forthe growth of thermophilic microorganisms. Appl EnvironMicrobiol 47:427–429
Marques AR, Ferreira PB, Sá-Correia I, Fialho AM (2003) Charac-terization of the ugpG gene encoding a UDP-glucose pyrophos-phorylase from the gellan gum producer Sphingomonaspaucimobilis ATCC 31461. Mol Genet Genomics 268:816–824
Martins LO, Sá-Correia I (1994) Temperature profiles of gellan gumsynthesis and activities of biosynthetic enzymes. Biotechnol ApplBiochem 20:385–395
Moreira LM, Hoffmann K, Albano H, Becker A, Niehaus K, Sá-Correia I (2004) The gellan gum biosynthetic genes gelC andgelE encode two separate polypeptides homologous to theactivator and the kinase domains of tyrosine autokinases. J MolMicrobiol Biotechnol 8:43–57
Moslemy P, Guiot SR, Neufeld RJ (2004) Activated sludge encapsu-lation in gellan gum microbeads for gasoline biodegradation.Bioprocess Biosyst Eng 26:197–204
Moslemy P, Neufeld RJ, Millette D, Guiot SR (2003) Transport ofgellan gum microbeads through sand: an experimental evaluationfor encapsulated cell bioaugmentation. J Environ Manage69:249–59
Niemeyer D, Becker A (2001) The molecular weight distribution ofsuccinoglycan produced by Sinorhizobium meliloti is influencedby specific tyrosine phosphorylation and ATPase activity of thecytoplasmic domain of the ExoP protein. J Bacteriol 183:5163–5170
Pollock TJ (1993) Gellan-related polysaccharides and the genusSphingomonas. J Gen Microbiol 139:1939–1945
Appl Microbiol Biotechnol (2008) 79:889–900 899

Pollock TJ, Thorne L, Yamazaki M, Mikolajczak MJ, Armentrout RW(1994) Mechanism of bacitracin resistance in gram-negativebacteria that synthesize exopolysaccharides. J Bacteriol176:6229–6237
Pollock TJ, vanWorkumWA, Thorne L,MikolajczakMJ, Yamazaki M,Kijne JW, Armentrout RW (1998) Assignment of biochemicalfunctions to glycosyl transferase genes which are essential forbiossynthesis of exopolysaccharides in Sphingomonas strain S88and Rhizobium leguminosarum. J Bacteriol 180:586–593
Reina J, Bassa A, Llompart I, Borrell D (1991) Infections withPseudomonas paucimobilis. Report of four cases and review. RevInfect Dis 13:1072–1076
Rinaudo M (2004) Role of substituents on the properties of somepolysaccharides. Biomacromolecules 5:1155–1165
Rinaudo M, Milas M (2000) Gellan gum, a bacterial gelling polymer.In: Doxastakis G, Kiosseoglou V (eds) Novel macromolecules infood systems. Elsevier, Amsterdam, pp 239–263
Sá-Correia I, Fialho AM, Videira P, Moreira LM, Marques AR,Albano H (2002) Gellan gum biosynthesis in Sphingomonaspaucimobilis ATCC 31461: genes, enzymes and exopolysacchar-ide production engineering. J Ind Microbiol Biotechnol 29:170–176
Saitou N, Nei M (1987) The neighbor-joining method: a new methodfor reconstructing phylogenetic trees. Mol Biol Evol 4:406–425
Silva E, Marques AR, Fialho AM, Granja AT, Sá-Correia I (2005)Proteins encoded by Sphingomonas elodea ATCC 31461 rmlAand ugpG genes, involved in gellan gum biosynthesis, exhibitboth dTDP- and UDP-glucose pyrophosphorylase activities. ApplEnviron Microbiol 71:4703–4712
Smith AM, Shelton RM, Perrie Y, Harris JJ (2007) An initialevaluation of gellan gum as a material for tissue engineeringapplications. J Biomater Appl 22:241–254
Sultana Y, Aqil M, Ali A (2006) Ion-activated. Gelrite-based in situophthalmic gels of pefloxacin mesylate: comparison withconventional eye drops. Drug Deliv 13:215–219
Sutherland IW (2001) Microbial polysaccharides from Gram-negativebacteria. Int Dairy J 11:663–674
Thorne L, Mikolajczak MJ, Armentrout RW, Pollock TJ (2000)Increasing the yield and viscosity if exoplysaccharides secretedby Sphingomonas by augmentation of chromosomal genes withmultiple copies of cloned biosynthetic genes. J Ind MicrobiolBiotechnol 25:49–57
Tocilj A, Munger C, Proteau A, Morona R, Purins L, Ajamian E,Wagner J, Papadopoulos M, Van Den Bosch L, Rubinstein JL,Féthière J, Matte A, Cygler M (2008) Bacterial polysaccharideco-polymerases share a common framework for control ofpolymer length. Nat Struct Mol Biol 15:130–138
Vaningelgem F, Zamfir M, Adriany T, De Vuyst L (2004) Fermen-tation conditions affecting the bacterial growth and exopolysac-charide production by Streptococcus thermophilus ST 111 inmilk-based medium. J Appl Microbiol 97:1257–1273
Vartak NB, Lin CC, Cleary JM, Fagan MJ, Saier Jr MH (1995)Glucose metabolism in Sphingomonas elodea: pathway engi-neering via construction of a glucose-6-phosphate dehydrogenaseinsertion mutant. Microbiology 141:2339–2350
Videira PA, Cortes LL, Fialho AM, Sá-Correia I (2000) Identificationof the pgmG gene, encoding a bifuncional protein withphosphoglucomutase and phosphomannomutase activities, inthe gellan gum-producing strain Sphingomonas paucimobilisATCC 31461. Appl Environ Microbiol 66:2252–2258
Videira P, Fialho AM, Geremia RA, Breton C, Sá-Correia I (2001)Biochemical characterization of the beta-1,4-glucuronosyltrans-ferase GelK in the gellan gum-producing strain Sphingomonaspaucimobilis A.T.C.C. 31461. Biochem J 358:457–464
Wang X, Xu P, Yuan Y, Liu C, Zhang D, Yang Z, Yang C, Ma C(2006) Modeling for gellan gum production by Sphingomonaspaucimobilis ATCC 31461 in a simplified medium. Appl EnvironMicrobiol 72:3367–3374
Welman AD, Maddox IS (2003) Exopolysaccharides from lactic acidbacteria: perspectives and challenges. Trends Biotechnol 21:269–274
West TP, Strohfus B (1998) Effect of complex nitrogen sources upongellan production by Sphingomonas paucimobilis ATCC 31461.Microbios 94:145–152
White DC, Sutton SD, Ringelberg DB (1996) The genus Sphingomo-nas: physiology and ecology. Curr Opin Biotechnol 7:301–306
Whitfield C (2006) Biosynthesis and assembly of capsular polysac-charides in Escherichia coli. Annu Rev Biochem 75:39–68
Wugeditsch T, Paiment A, Hocking J, Drummelsmith J, Forrester C,Whitfield C (2001) Phosphorylation of Wzc, a tyrosine autoki-nase, is essential for assembly of group 1 capsular polysacchar-ides in Escherichia coli. J Biol Chem 276:2361–2371
Yamazaki M, Thorne L, Mikolajczak M, Armentrout RW, Pollock TJ(1996) Linkage of genes essential for synthesis of a polysaccharidecapsule in Sphingomonas strain S88. J Bacteriol 178:2676–2687
900 Appl Microbiol Biotechnol (2008) 79:889–900