novel endogenous peptide agonists of cannabinoid receptors
TRANSCRIPT

The FASEB Journal • Research Communication
Novel endogenous peptide agonists of cannabinoidreceptors
Ivone Gomes,* Julia S. Grushko,† Urszula Golebiewska,‡ Sascha Hoogendoorn,*Achla Gupta,* Andrea S. Heimann,§ Emer S. Ferro,� Suzanne Scarlata,‡
Lloyd D. Fricker,† and Lakshmi A. Devi*,1
*Department of Pharmacology and Systems Therapeutics, Mount Sinai School of Medicine, NewYork, New York, USA; †Department of Molecular Pharmacology, Albert Einstein College of Medicine,New York, New York, USA; ‡Department of Physiology and Biophysics, Stony Brook UniversityMedical Center, Stony Brook, New York, USA; §Proteimax Biotechnology Ltd., Cotia, Brazil; and�Department of Cell Biology and Development, Institute of Biomedical Sciences, University of SaoPaulo, Sao Paulo, Brazil
ABSTRACT Hemopressin (Hp), a 9-residue �-hemo-globin-derived peptide, was previously reported tofunction as a CB1 cannabinoid receptor antagonist (1).In this study, we report that mass spectrometry (MS)data from peptidomics analyses of mouse brain ex-tracts identified N-terminally extended forms of Hpcontaining either three (RVD-Hp�) or two (VD-Hp�)additional amino acids, as well as a �-hemoglobin-derived peptide with sequence similarity to that ofhemopressin (VD-Hp�). Characterization of the �-he-moglobin-derived peptides using binding and func-tional assays shows that in contrast to Hp, whichfunctions as a CB1 cannabinoid receptor antagonist,both RVD-Hp� and VD-Hp� function as agonists.Studies examining the increase in the phosphorylationof ERK1/2 levels or release of intracellular Ca2�
indicate that these peptides activate a signal transduc-tion pathway distinct from that activated by the endo-cannabinoid, 2-arachidonoylglycerol, or the classic CB1agonist, Hu-210. This finding suggests an additionalmode of regulation of endogenous cannabinoid recep-tor activity. Taken together, these results suggest thatthe CB1 receptor is involved in the integration ofsignals from both lipid- and peptide-derived signalingmolecules.—Gomes, I., Grushko, J. S., Golebiewska,U., Hoogendoorn, S., Gupta, A., Heimann, A. S., Ferro,E. S., Scarlata, S., Fricker, L. D., Devi, L. A. Novelendogenous peptide agonists of cannabinoid receptors.FASEB J. 23, 3020–3029 (2009). www.fasebj.org
Key Words: G-protein-coupled receptors � pain � analgesia� drug abuse
Cannabinoid receptors are members of the super-family of Gi/Go-coupled receptors. The major psycho-active component of Cannabis sativa, �9-tetrahydrocan-nabinol, binds to at least two types of cannabinoidreceptors, CB1 and CB2 (2). The CB1 receptor isexpressed primarily in the central nervous system (3),whereas the CB2 receptor is predominantly expressedin immune cells (4) and is also detectable in brainstem
neurons (5) and spinal cord (6). More recently, GPR55,which is highly expressed in large dorsal root ganglionneurons, has also been shown to function as a canna-binoid receptor (7). A number of studies have shownthat cannabinoid receptor activation leads to the inhi-bition of adenylyl cyclase activity, inhibition of calciumchannels and D-type potassium channels, increases inthe phosphorylation of mitogen-activated protein ki-nases, and activation of A-type and inwardly rectifyingpotassium channels (8–10). To date, two endogenousligands of cannabinoid receptors, anandamide and2-arachidonoylglycerol (2-AG), which are derived frommembrane lipids, have been well characterized (11–13). Several studies have proposed important roles forthe endocannabinoid system (consisting of the recep-tors and endogenous ligands) in many pathophysiolog-ical processes including Parkinson’s disease, Alzhei-mer’s disease, depression, inflammation, neuropathicpain, and obesity (11, 13–14).
The recent finding that hemopressin (Hp), a 9-resi-due peptide derived from the �1 chain of hemoglobin,functions as an antagonist of the CB1 receptor andexhibits antinociceptive activity (1) suggests the possi-bility that cannabinoid receptor activity could be mod-ulated by peptides derived from larger precursor pro-teins, which would significantly increase the complexityof the endocannabinoid system. Many neuropeptidesfunction in cell-cell communication, and most of theseneuropeptides are produced within the secretory path-way by selective proteases that cleave the peptide pre-cursors at specific, well-defined sites. After cleavage, thepeptides are stored within vesicles and secreted uponcell stimulation. In addition to these “classic” neu-ropeptides, a number of bioactive peptides have beenidentified that appear to be derived from cellular
1 Correspondence: Department of Pharmacology and Sys-tems Therapeutics, Mt. Sinai School of Medicine, 1 Gustave L.Levy Place, Box 1603, New York, NY 10029, USA. E-mail:[email protected]
doi: 10.1096/fj.09-132142
3020 0892-6638/09/0023-3020 © FASEB

proteins. However, these peptides do not appear to beproduced by selective cleavages at specific sites or to bestored within vesicles and released by stimulation. Hpseems to be an example of this “nonclassic” group ofneuropeptides. In addition to Hp, the � and � chains ofhemoglobin are precursors of peptides named he-morphins and neokyotorphin; these peptides inter-act with opiate receptors and angiotensin receptors(15–19). Interestingly, Hp functions as an antagonistof the CB1 receptor, is orally active, and causesantinociception (1).
Hp was previously identified in extracts of rat brainusing an affinity purification method involving bindingto endopeptidase 24.15. In the present study, we inves-tigated whether Hp or Hp-like peptides were majorpeptides in mouse brain extracts, without using anaffinity purification approach. Although the 9-residueHp was not found, two N-terminally extended forms ofHp were found, one of which was highly represented ina meta-analysis of peptidomics analyses. These longerforms of Hp were then characterized for activity towardCB receptors; interestingly, although the nine-residueHp is an antagonist of the CB1 receptor, the longerforms are agonists of this receptor. Furthermore, thesignaling pathways for the peptide-based agonists ap-pear to be distinct from the lipid-based and syntheticagonists of the CB1 receptor, implying that this recep-tor has a complex function of integration of a widevariety of signaling molecules.
MATERIALS AND METHODS
Materials
HEK-293 cells, Neuro 2A cells, and CHO cells were obtainedfrom American Type Culture Collection (Manassas, VA,USA). DMEM, F12 medium, penicillin-streptomycin, lipo-fectamine, and Fluo-4 NW calcium dye were obtained fromInvitrogen (Carlsbad, CA, USA). Chimeric G16/Gi3 constructwas a kind gift from Dr. R. Margolskee (Mount Sinai Schoolof Medicine, New York, NY, USA). Plasmid cDNAs for CB1cannabinoid and CB2 cannabinoid receptors and for GPR55were a kind gift from Dr. K. Mackie (Indiana University,Bloomington, IN, USA). Plasmid cDNAs for � and � opioidreceptors were a kind gift from Dr. Chris Evans (Universityof California, Los Angeles, CA, USA). [3H]CP55,940 andSR141716 were obtained from the National Institute on DrugAbuse Drug Supply Program (Bethesda, MD, USA). Antibod-ies to phosphorylated and total ERK1/2 were obtained fromCell Signaling Technology Inc. (Danvers, MA, USA). Thesecondary antibodies IRDye680-labeled anti-rabbit and IRDye800-labeled anti-mouse antibodies were from Li-COR (Lin-coln, NE, USA). The anti-rabbit antibody conjugated tohorseradish peroxidase (HRP) was from Amersham Bio-sciences Corp. (Piscataway, NJ, USA). RVD-Hp�, VD-Hp�,VD-Hp�, and plasmid cDNA for angiotensin 1 (AT1) recep-tors were from Proteimax (Sao Paulo, Brazil). All otherreagents were obtained from Sigma-Aldrich Corp. (St. Louis,MO, USA).
Cell culture and transfection
HEK-293 cells were grown in DMEM containing 10% FBS and1% penicillin-streptomycin. Neuro 2A cells were grown in
DMEM and F12 medium (50:50) containing 10% FBS and 1%penicillin-streptomycin. CHO cells were grown in F12 me-dium containing 10% FBS and 1% penicillin-streptomycin.For receptor internalization studies CHO cells were trans-fected with myc-tagged CB1 cannabinoid receptors usingLipofectamine as per the manufacturer’s protocol (Invitro-gen). For experiments measuring increases in intracellularCa2� levels, HEK-293 cells were cotransfected with a chimericG16/Gi3 and either CB1 cannabinoid, CB2 cannabinoid, �2Aadrenergic, �2 adrenergic, � opioid, or � opioid receptors. Inthe case of AT1 receptors that signal through G�q, cells werenot cotransfected with the chimeric G16/Gi3.
Mass spectrometric identification of Hp-based peptides
Our composite database consists of MS data from peptidom-ics experiments (both published and unpublished) from�100 HPLC-MS runs using extracts from various regions ofmouse brain (20–31). Brain tissues were processed as de-scribed previously. Briefly, brain areas were pooled fromgroups of animals and frozen. For peptide extraction, thetissue pool was sonicated in ice-cold H2O and incubated in a70°C water bath for 20 min. The homogenate was then cooledon ice and acidified with 0.1 M HCl to a final concentrationof 10 mM HCl. The homogenates were kept on ice for 15 minand centrifuged at 15,000 g for 30 min at 4°C. The pH of thesupernatant was adjusted to 9.5 by addition of 0.2 M phos-phate buffer. The samples were then labeled with differentialisotopic trimethylammonium butyrate (TMAB) labels andanalyzed by liquid chromatography (LC)-tandem mass spec-trometry (MS/MS), as described previously (20–31).
Ligand binding and signaling studies
Membranes were prepared from the cerebellum and striatumof C57BL/6 mice brains as described previously (32). Forligand binding, membranes from mice cerebellum (50 �g)were incubated for 2 h at 30°C with 0.5 nM [3H]CP55,940 inthe absence or presence of increasing concentrations (0–1�M) of SR141716, RVD-Hp�, VD-Hp�, or VD-Hp� as de-scribed previously (32) in a buffer containing 50 mM Tris-Cl(pH 7.4), 5 mg/ml BSA (fatty acid free), 1 mM EDTA, 3 mMMgCl2, and protease inhibitor cocktail. At the end of theincubation period, membranes were filtered using GF/Bfilters (presoaked for 2 h at room temperature with 0.1%polyethyleneimine containing 0.2% fatty acid free BSA) andwashed 3 times with ice-cold 50 mM Tris-Cl (pH 7.4). Forguanosine 5�-O-(3-thio)triphosphate (GTPS) binding, mem-branes from striatum or cerebellum (10 �g) were incubatedwith the indicated doses of Hu-210, SR141716, RVD-Hp�, orVD-Hp� as described previously (32).
G�i16-facilitated Ca2� release
HEK-293 cells coexpressing a chimeric G16/Gi3 with eitherCB1 cannabinoid, CB2 cannabinoid, �2A adrenergic, �2 ad-renergic, � opioid, or � opioid receptors or HEK-293 cellsexpressing AT1 receptors were plated into poly-l-lysine-coated 96-well clear-bottom plates (40,000 cells/well). On thenext day, the growth medium was removed, and cells werewashed twice in HBSS buffer containing 20 mM HEPES. Cellswere incubated with Fluo-4 NW calcium dye (3 �M in 100 �l)for 1 h at 37°C. The different compounds (1 �M) were addedto the wells by the robotic arm of the FLEX Station, andintracellular Ca2� levels were measured for 300 s at excitation494 nm and emission 516 nm. In experiments examiningwhether cannabinoid receptor-mediated increases in intracel-lular Ca2� levels were blocked by the antagonist, cells were
3021ENDOGENOUS PEPTIDE AGONISTS TO CANNABINOID RECEPTORS

pretreated with SR141716 (10 �M) 30 min before addition ofthe different ligands.
Phosphorylation of MAP kinase (ERK)
HEK-293 cells expressing myc-tagged CB1 receptors, HA-tagged CB2 receptors, or GPR55 (2105 cells/well) weretreated for 5 min with 1 �M Hu-210, RVD-Hp�, VD-Hp�, orVD-Hp� in the absence or presence of 10 �M SR141716, andlevels of phosphorylated ERK1/2 were determined as de-scribed previously (1, 32). For dose-response studies, HEK-293 cells expressing CB1 receptors were treated for 5 min with0–10 �M Hu-210, RVD-Hp�, or VD-Hp�. For time coursestudies, HEK-293 cells expressing CB1 receptors were treatedwith 1 �M Hu-210, RVD-Hp�, or VD-Hp� for 0–20 min. Toobserve the effect of pertussis toxin (PTX) on phosphorylatedERK1/2 levels, HEK-293 cells expressing CB1 receptors werepretreated for 2 h at 37°C with 15 ng/ml (PTX), followed bya 5-min treatment with 1 �M Hu-210, RVD-Hp�, or VD-Hp�.
Neurite outgrowth assays
Neuro 2A cells endogenously expressing CB1 receptors weretreated for 16 h in medium containing 0.1% FBS with 1 �MHu-210, RVD-Hp�, VD-Hp�, or VD-Hp� in the absence orpresence of 10 �M SR141716, and neurite length was deter-mined as described previously (33, 34).
Receptor internalization studies
CHO cells expressing myc-tagged CB1 receptors (1–2105
cells /well) were plated onto a poly-l-lysine-coated 24-wellplate. On the next day, the growth medium was removed, andcells were treated with 1 �M Hu-210, RVD-Hp�, VD-Hp�, orVD-Hp� in the absence or presence of 10 �M SR141716 inDMEM for 60 min at 37°C. The plate was then placed on ice,and wells were washed 3 times with 500 �l of ice-cold PBS andsubjected to ELISA using 1:1000 of anti-myc polyclonal anti-body and 1:1000 of HRP-conjugated anti-rabbit secondaryantibody as described previously (32).
Intracellular Ca2� release in individual cells
Neuro 2A, HEK-293, or HEK-293 cells expressing HA-CB1were plated in Mattek chambers for 24 h to achieve 50–80%confluence. Before the experiment, cells were starved inserum-free medium from 6 h to overnight. Cells were incu-bated in HBSS medium with 1 �M Calcium Orange for 45min, washed 3 times in medium, and subsequently incubatedin medium without Calcium Orange for 30 min, followed by3 additional washes with HBSS medium. Prolonged washing isneeded to remove dye nonspecifically bound to the plasmamembrane. Cells were imaged on a Zeiss LSM510 laserscanning confocal microscope (Carl Zeiss, Thornwood, NY,USA). From one dish, several cells in one field of view wererecorded. Calcium Orange was excited with a 543-nm HeNelaser line, and the emission spectrum was recorded using along-pass 560 filter. Images were recorded before treatment(to measure Fmin). Cells were treated with 1 �M ligand; aftertreatment, a time course of fluorescence intensity was re-corded for 30 min. For Fmax estimation cells were treated with5 �g/ml calcimycin (A23187).
Free calcium concentration was calculated from the follow-ing equation:
�Ca2��free � Kd�F � Fmin
Fmax � F�
where Kd is the dissociation constant of the calcium indicator(185 nM for Calcium Orange), F is fluorescence intensity atgiven time point, Fmin is the fluorescence intensity in theabsence of stimulator, and Fmax is the intensity in the pres-ence of calcium ionophore. Average intensity of many pixelsinside a given cell was calculated using Zeiss software. Datapoints were averaged and se was calculated using Sigma Plot(Systat Software, Inc., San Jose, CA, USA).
Statistical analysis
The results are expressed as the mean se. Statisticalcomparisons were done using analysis of variance (ANOVA)followed by the Bonferroni multiple comparison test. P �0.05 indicated a significant difference. Statistical analyses ofdata were generated using GraphPad Prism 4.02 (GraphPadSoftware Inc., San Diego, CA, USA).
RESULTS
Identification of endogenous forms of Hp
We have previously shown that Hp, a nonapeptidederived from the � chain of hemoglobin, functions asan antagonist of CB1 cannabinoid receptors (1). Acentral question is whether Hp represents an endoge-nous peptide that is normally present in the brain. Toaddress this question, we searched for the Hp sequencein a database consisting of MS results from �100peptidomics analyses of mouse brain extracts. We didnot find Hp, but we did identify longer N-terminallyextended Hp peptides containing 11- and 12-aa resi-dues, VDPVNFKLLSH (VD-Hp�) and RVDPVNFKLLSH(RVD-Hp�), respectively, by MS/MS (Fig. 1). RVD-Hp�was detected in many of the brain regions examined,such as hypothalamus, nucleus accumbens, olfactorybulb, cerebellum, prefrontal cortex, and striatum, andVD-Hp� was detected in olfactory bulb, hypothalamus,and cerebellum (data not shown). Supplemental Table1 summarizes all of the neuropeptides, hemoglobin-derived peptides, and other representative peptidesthat were detected in at least 5 distinct samples.Whereas VD-Hp� was found in only 8 of the samples,RVD-Hp� was identified in 47 different samples, repre-senting one of the more frequently detected mousebrain peptides (Supplemental Table 1). In addition tothese �-hemoglobin-derived forms of Hp, we also founda number of other fragments of �-hemoglobin, al-though none were found as many times as RVD-Hp�(Supplemental Table 1). Two �-hemoglobin-derivedpeptides were also detected in our analysis; one corre-sponds to the previously identified bioactive peptidenamed VV-hemorphin, and the other has substantialsequence homology to Hp (Supplemental Table 1).Based on the nomenclature system for hemorphins andHp, we have named this �-hemoglobin-derived Hp-likepeptide VD-hemopressin� (VD-Hp�).
Agonistic activity of RVD-Hp�, VD-Hp�, and VD-Hp�at CB1 cannabinoid receptors
Because it was previously shown that Hp functions as anantagonist/inverse agonist of CB1 cannabinoid recep-
3022 Vol. 23 September 2009 GOMES ET AL.The FASEB Journal � www.fasebj.org

tors, the effect of RVD-Hp�, VD-Hp�, and VD-Hp� onsignaling by cannabinoid receptors was evaluated byexamining their effect on receptor-mediated increasesin intracellular Ca2� levels in heterologous cells ex-pressing a variety of family A G-protein-coupled recep-tors (GPCRs) along with a promiscuous G protein (Fig. 2).Treatment with RVD-Hp� and VD-Hp� alone causedan increase in intracellular Ca2� levels in cells express-ing CB1 cannabinoid receptors and to a lesser extent incells expressing CB2 cannabinoid receptors, whereasVD-Hp� was equally effective at CB1 and CB2 cannabi-noid receptors (Fig. 2). None of the three peptidesinduced increases in intracellular Ca2� levels in cellsexpressing � and � opioid, �2A and �2 adrenergic, orAT1 receptors (Fig. 2). Moreover, RVD-Hp� , VD-Hp�,and VD-Hp�-mediated increases in intracellular Ca2�
levels in CB1-expressing cells could be blocked bypretreating cells with the cannabinoid antagonistSR141716 (VD-Hp�-mediated increases in intracellularCa2� levels were only partially blocked by SR141716)(Fig. 3). We also examined the ability of extended Hppeptides to bind to cannabinoid receptors in cerebellarmembranes. We found that RVD-Hp� can displace thebinding of the cannabinoid ligand, [3H]CP55,940, withnanomolar affinity although to a lesser extent than the
classic CB1 antagonist, SR141716 (Supplemental Fig.1). Similar results were obtained with VD-Hp� andVD-Hp� (not shown). Taken together, these resultssuggest that, in contrast to Hp that functions as anantagonist (1), RVD-Hp� and VD-Hp� function asagonists of cannabinoid receptors.
Previous studies have shown that treatment withagonists of CB1 receptors can induce neurite outgrowthin cells endogenously expressing these receptors (33,34). We used this functional assay to measure theagonistic activity of RVD-Hp�, VD-Hp�, and VD-Hp�.As expected, these peptides increased the number ofcells with neurites in a manner similar to that of thecannabinoid agonist, Hu-210 (Fig. 4 and SupplementalFig. 2). We found that the RVD-Hp�- and VD-Hp�-mediated increases in neurite outgrowth could beblocked by pretreatment with the antagonist, SR141716(Fig. 4). Activation of CB1 receptors has also beenshown to lead to rapid receptor endocytosis (35, 36).We examined the ability of RVD-Hp�, VD-Hp�, andVD-Hp� to induce receptor endocytosis in CHO cellsexpressing myc-tagged CB1 receptors. The classic agonist,Hu-210, as well as RVD-Hp�, VD-Hp�, and VD-Hp�induce CB1 receptor internalization; however, only theRVD-Hp�- and VD-Hp�-induced CB1 receptor internal-
Figure 1. Mass spectrometric identification ofendogenous RVD-Hp�. Peptides were extractedfrom microwave-irradiated mouse brain, labeledwith TMAB isotopic labels, purified by microfil-tration using a 10-kDa exclusion unit, and ana-lyzed by LC/MS. In this example, the tandemmass spectrum is shown for the quadruplycharged ion with an m/z of 420.5 and a monoiso-topic mass of 1423.79 Da (after subtraction of themass of the hydrogenated form of the TMABisotopic tag). Inset: an expanded y axis for theindicated m/z range. The observed ions representb-series, y-series, and internal ions formed second-ary to the cleavage of the D-P bond. Fragmenta-tion ions usually lose the trimethylamine moiety.However, the doubly charged b3 ion shows asubstantial peak for the fragment that retainedthe trimethylamine; this is indicated as b3
2� � 59.Conversely, the parent ion that lost the trimeth-ylamine moiety is indicated as MH4� � 59. amu,atomic mass units.
Figure 2. Longer Hp peptides are able to in-crease CB receptor-mediated intracellular Ca2�
levels. HEK-293 cells (40,000 cells/well) coex-pressing chimeric G16/Gi3 and individual recep-tors (except in the case of AT1 receptors) weretreated with 1 �M concentrations of receptoragonists, RVD-Hp�, VD-Hp�, or VD-Hp�, andintracellular Ca2� levels were determined. RFU,relative fluorescence units.
3023ENDOGENOUS PEPTIDE AGONISTS TO CANNABINOID RECEPTORS

ization can be blocked by the cannabinoid receptorantagonist, SR141716 (Fig. 5). Taken together, theseresults indicate that RVD-Hp� and VD-Hp� behave asselective agonists of CB1 cannabinoid receptors.
The ability of RVD-Hp�, VD-Hp�, and VD-Hp� tomediate phosphorylation of ERK1/2 in heterologouscells expressing CB1 receptors was examined. We foundthat all three peptides enhance phospho-ERK (pERK)levels in cells expressing CB1 receptors; the enhance-ment of pERK levels by RVD-Hp� and VD-Hp� but not byVD-Hp� is significantly blocked by SR141716 (Fig. 6A andSupplemental Fig. 3). Next, we focused our studies onRVD-Hp� and VD-Hp� because they exhibited CB1receptor selectivity. To explore this selectivity further,the ability of these peptides to transduce signals via CB2or GPR55 receptors was examined. Treatment of cellsexpressing either CB2 or GPR55 did not lead to sub-stantial increases in pERK levels by RVD-Hp� (increases
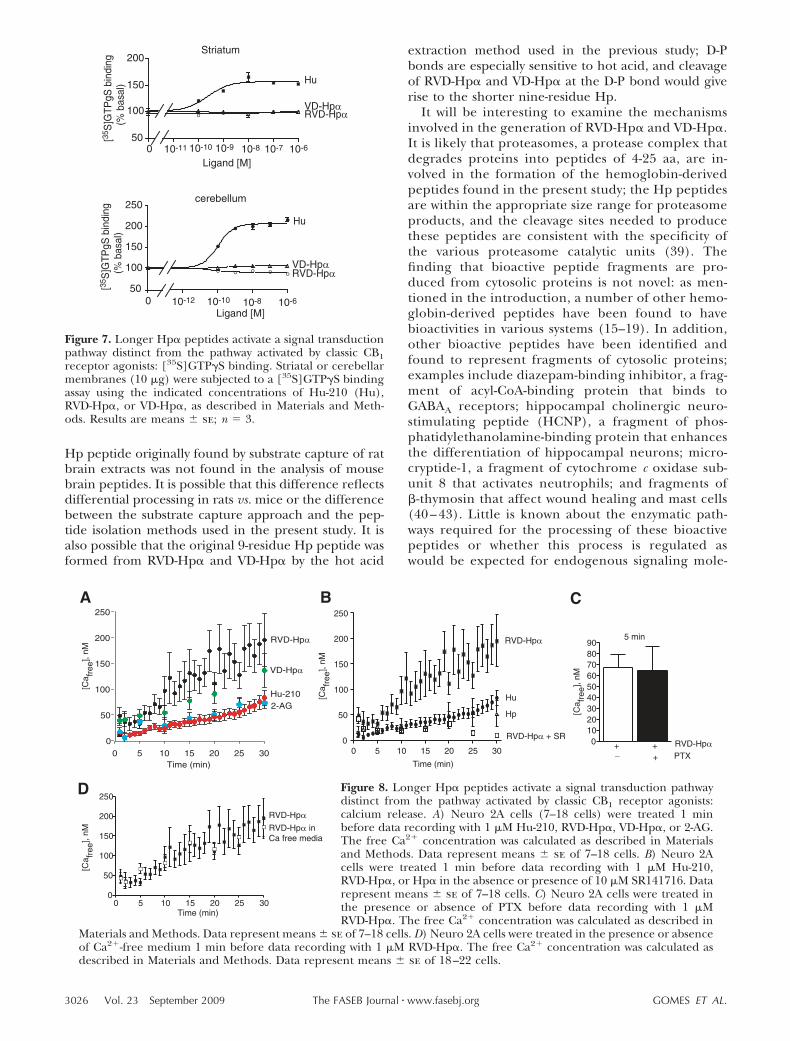
were found with VD-Hp� in cells expressing CB2 recep-tors, although they were not statistically significant)(Fig. 6A). Next, we focused on RVD-Hp� and com-pared the time course of ERK phosphorylation by thepeptide ligand with that of the classic CB1 ligand,Hu-210 (Fig. 6B). Both the level and rate of increase inpERK were lower in the case of RVD-Hp� comparedwith that of Hu-210 (Fig. 6B). The increase in pERKwith Hu-210 at early time points (3 and 5 min) is greatlyreduced by PTX, whereas the increase in pERK due toRVD-Hp� at 5 min is decreased by only 30% (Fig. 6B).This result, together with the different time course,suggests that stimulation of CB1 receptors by the pep-tide ligand leads to activation of a signaling pathwaydistinct from the classic G-protein-mediated pathway.The differences in signaling by the peptide ligands andHu-210 were further explored using a G-protein activa-tion assay. In both cerebellar and striatal membranes,although Hu-210 treatment caused a robust increase in[35S]GTPS binding, treatment with the peptide li-gands did not (Fig. 7). Taken together, these resultssuggest that the peptide ligands stimulate CB1 recep-tors via a mechanism distinct from Hu-210.
To explore the peptide agonist-mediated CB1 recep-tor signaling, we examined the dynamics of Ca2�
release in the absence of promiscuous G protein. InNeuro 2A cells that endogenously express CB1 recep-tors as well as in HEK-293 cells expressing recombinantreceptors, treatment with the peptide ligands leads to asustained increase in Ca2� release that is faster andmore robust than treatment with either the endocan-nabinoid 2-AG, or the classic agonist Hu-210 (Fig. 8A).Under the same conditions, activation of the endoge-nous adenosine receptors in Neuro 2A cells with 1 �MATP leads to a rapid (within seconds) and transientincrease in Ca2� release (data not shown). Further-more, the RVD-Hp�-mediated Ca2� release is blockedby SR141716 (Fig. 8B). In addition, we found that PTX
Figure 3. Longer Hp peptide-mediated calcium release isblocked by the CB1 receptor antagonist. HEK-293 cells(40,000 cells/well) coexpressing chimeric G16/Gi3 and CB1receptors were treated with 1 �M HU-210 (Hu), RVD-Hp�,VD-Hp�, or VD-Hp� in the absence or presence of 10 �MSR141716 (SR), and intracellular Ca2� levels were deter-mined. Results are means se; n � 3. RFU, relative fluores-cence units.
Figure 4. Longer Hp� peptides induce neurite outgrowth inNeuro 2A cells. Neuro2A cells were treated for 16 h with 1 �MHu-210 (Hu), RVD-Hp�, or VD-Hp� in the absence orpresence of 10 �M SR141716 (SR), and the percentage ofcells extending neurites was determined. Results are means se; n � 3. **P � 0.01, ***P � 0.001; 1-way ANOVA.
Figure 5. Longer Hp peptides induce CB1 receptor internal-ization. CHO cells expressing myc-tagged CB1 receptors (1–2105 cells/well) were treated with 1 �M Hu-210 (Hu),RVD-Hp�, VD-Hp�, or VD-Hp� in the absence or presence of10 �M SR141716 (SR) and subjected to ELISA using 1:1000 ofanti-myc polyclonal antibody and 1:1000 of HRP-conjugatedanti-rabbit secondary antibody, as described in Materials andMethods. Results are means se of 3 experiments intriplicate. *P � 0.05, **P � 0.01, ***P � 0.001; 1-way ANOVA.
3024 Vol. 23 September 2009 GOMES ET AL.The FASEB Journal � www.fasebj.org

pretreatment does not block RVD-Hp�-mediated Ca2�
release at early time points (Fig. 8C), although partialblockade is observed at later time points (SupplementalFig. 4A). This finding suggests that the CB1 receptor-mediated Ca2� release is not via the G�i-mediatedpathway. Similar results were obtained when HEK cellstransfected with CB1 receptors were examined; RVD-Hp� caused a more robust increase in free intracellularCa2� than Hu-210 (Supplemental Fig. 4B). The pep-tide-mediated Ca2� release in the Neuro 2A cells is seenin the absence of extracellular calcium (Fig. 8D),indicating that the Ca2� release is from intracellularstores. These results are exciting and indicate that thepeptide agonists activate a signal transduction pathwaythat results in a sustained increase in Ca2� release,which is distinct from that seen with the endocanna-biniod 2-AG. Furthermore, the peptide-mediated Ca2�
signaling is distinct from that of the classic G�q-coupledreceptors that tends to be rapid and transient.
DISCUSSION
In this study, we show that N-terminally extended formsof Hp function as agonists of CB1 cannabinoid recep-
tors. Whereas previous studies have reported that CB1receptors can couple to different G proteins (37) andthat the same GPCR can activate distinct signaling onbeing activated by different ligands (38), the presentstudy, for the first time, demonstrates that two distinct“endogenous” ligands that differ in their chemicalnature (lipid vs. peptide) activate the same receptor toinitiate distinct signaling pathways.
The extended Hp peptides represent fairly abundantpeptides, based on the numbers of times each of thesepeptides were found in LC-MS analysis. Our compositedatabase of MS data represents �100 LC-mass spec-trometry runs from different samples of extracts ofvarious mouse brain regions; this database contains�1000 peptides that have been identified at least onceby MS/MS sequencing and that arise from eithersecretory pathway “neuropeptide” precursors or cytoso-lic proteins, such as the hemoglobin-derived Hps. Themost frequently found peptides were derived fromprecursors that are known to be expressed at relativelyhigh levels in mouse brain (Supplemental Table 1).RVD-Hp� was the most frequently detected �-hemoglo-bin-derived peptide, being found as many times assome of the more abundant neuropeptides. The 9-aa
Figure 6. Longer Hp� peptides mediate ERK phosphorylation.A) HEK-293 cells expressing myc-tagged CB1 receptors, HA-tagged CB2 receptors, or GPR55 were treated with 1 �M Hu-210(Hu), RVD-Hp�, or VD-Hp� in the absence or presence of 10�M SR141716 (SR), and levels of pERK1/2 were determined.Results are means se; n � 3. *P � 0.01, **P � 0.001; 1-wayANOVA. B) HEK-293 cells expressing CB1 receptors weretreated for 0–20 min with 1 �M Hu-210 or RVD-Hp� in theabsence or presence of PTX, and levels of phosphorylatedERK1/2 were determined as described in Materials and Meth-ods. Results are means se of 3 experiments in triplicate. **P �0.01; ***P � 0.001. Representative blots from 3 independentexperiments are shown.
3025ENDOGENOUS PEPTIDE AGONISTS TO CANNABINOID RECEPTORS
Hp peptide originally found by substrate capture of ratbrain extracts was not found in the analysis of mousebrain peptides. It is possible that this difference reflectsdifferential processing in rats vs. mice or the differencebetween the substrate capture approach and the pep-tide isolation methods used in the present study. It isalso possible that the original 9-residue Hp peptide wasformed from RVD-Hp� and VD-Hp� by the hot acid
extraction method used in the previous study; D-Pbonds are especially sensitive to hot acid, and cleavageof RVD-Hp� and VD-Hp� at the D-P bond would giverise to the shorter nine-residue Hp.
It will be interesting to examine the mechanismsinvolved in the generation of RVD-Hp� and VD-Hp�.It is likely that proteasomes, a protease complex thatdegrades proteins into peptides of 4-25 aa, are in-volved in the formation of the hemoglobin-derivedpeptides found in the present study; the Hp peptidesare within the appropriate size range for proteasomeproducts, and the cleavage sites needed to producethese peptides are consistent with the specificity ofthe various proteasome catalytic units (39). Thefinding that bioactive peptide fragments are pro-duced from cytosolic proteins is not novel: as men-tioned in the introduction, a number of other hemo-globin-derived peptides have been found to havebioactivities in various systems (15–19). In addition,other bioactive peptides have been identified andfound to represent fragments of cytosolic proteins;examples include diazepam-binding inhibitor, a frag-ment of acyl-CoA-binding protein that binds toGABAA receptors; hippocampal cholinergic neuro-stimulating peptide (HCNP), a fragment of phos-phatidylethanolamine-binding protein that enhancesthe differentiation of hippocampal neurons; micro-cryptide-1, a fragment of cytochrome c oxidase sub-unit 8 that activates neutrophils; and fragments of�-thymosin that affect wound healing and mast cells(40 – 43). Little is known about the enzymatic path-ways required for the processing of these bioactivepeptides or whether this process is regulated aswould be expected for endogenous signaling mole-
Figure 7. Longer Hp� peptides activate a signal transductionpathway distinct from the pathway activated by classic CB1receptor agonists: [35S]GTPS binding. Striatal or cerebellarmembranes (10 �g) were subjected to a [35S]GTPS bindingassay using the indicated concentrations of Hu-210 (Hu),RVD-Hp�, or VD-Hp�, as described in Materials and Meth-ods. Results are means se; n � 3.
Figure 8. Longer Hp� peptides activate a signal transduction pathwaydistinct from the pathway activated by classic CB1 receptor agonists:calcium release. A) Neuro 2A cells (7–18 cells) were treated 1 minbefore data recording with 1 �M Hu-210, RVD-Hp�, VD-Hp�, or 2-AG.The free Ca2� concentration was calculated as described in Materialsand Methods. Data represent means se of 7–18 cells. B) Neuro 2Acells were treated 1 min before data recording with 1 �M Hu-210,RVD-Hp�, or Hp� in the absence or presence of 10 �M SR141716. Datarepresent means se of 7–18 cells. C) Neuro 2A cells were treated inthe presence or absence of PTX before data recording with 1 �MRVD-Hp�. The free Ca2� concentration was calculated as described in
Materials and Methods. Data represent means se of 7–18 cells. D) Neuro 2A cells were treated in the presence or absenceof Ca2�-free medium 1 min before data recording with 1 �M RVD-Hp�. The free Ca2� concentration was calculated asdescribed in Materials and Methods. Data represent means se of 18–22 cells.
3026 Vol. 23 September 2009 GOMES ET AL.The FASEB Journal � www.fasebj.org

cules. Preliminary data from our laboratory suggest apossible role of global ischemia in the up-regulationof VD-Hp� and RVD-Hp� (data not shown). A recentstudy reported an increase in a large number ofhemoglobin-derived peptides that was induced by a3- or 10-min delay between death and microwaveirradiation (44) and a previous article from ourlaboratory listed VD-Hp� as one of the protein-derived peptides that is much more abundant inbrains that were not subjected to microwave irradia-tion as opposed to those that were irradiated (20).Because a delay in processing of brain tissue afterdecapitation is a well-accepted model of global ische-mia (45), this result provides further support to theidea that ischemia may regulate the breakdown ofhemoglobin into hemoglobin-derived peptides suchas VD-Hp� and RVD-Hp�.
Additional areas for further studies are to identifycell types that produce the hemoglobin and also toidentify the pathway by which intracellular peptidesand proteins are secreted. Although hemoglobin istraditionally viewed as being specific to erythrocytes,other cell types produce � and/or � hemoglobin.Both � and � hemoglobin mRNA and protein werefound in human endometrium, with all epithelialcells and most stromal cells being positive for immu-noreactive protein (46). A microarray screen de-tected � and � hemoglobin mRNA in mouse lens(47). Within the lung, alveolar epithelial type II cellsand Clara cells were found to express � and �hemoglobin mRNA and protein (48). Hemoglobinexpression was found to decrease when alveolarepithelial type II cells differentiated into type I cells,illustrating specificity of expression in type II but notin type I alveolar epithelial cells (49). �minor hemo-globin expression was induced in macrophages whenthe cells were stimulated to produce nitric oxide(50). Expression of both chains of hemoglobin wasfound in the mesangial region of rat glomeruli inkidney and in mesangial cells in vitro; this renalhemoglobin expression was increased with hypoxia(51). Furthermore, there has recently been a reportof � and � hemoglobin in neurons and glia (52).Thus, it is possible that the hemoglobin-derivedpeptides detected in our peptidomics analysis ofmouse brain extracts arise from neurons or glia andnot from erythrocytes present in the brain tissue.Because hemoglobin lacks a signal peptide, any celltype producing this protein will express it in thecytosol or related compartments and not in theclassic secretory pathway. For the hemoglobin-de-rived peptides to interact with receptors on distantcells, the peptides need to be secreted. The secretionof cytosolic proteins has been well documented;examples include the various interleukins as well asthymosin, HCNP and its precursor protein, andintracellular enzymes such as endopeptidases 24.15and 24.16 (53–56). Despite the large number ofcytosolic proteins/peptides known to be secreted,the precise mechanisms remain elusive.
Taken together, the data in this study are consis-tent with the notion that RVD-Hp� and VD-Hp�represent endogenous neuromodulatory peptideswith CB1 receptor activity. The amino acid sequenceof RVD-Hp� is highly conserved from reptiles tomammals, and only one conservative difference isfound among mammals (RVDPVNFKLLSH vs. RVDPVNFKFLSH); both of these peptides function simi-larly in their binding to CB1 receptors (data notshown). In contrast to the previously identified Hppeptide, which is an antagonist (or inverse agonist)at the CB1 cannabinoid receptors, the peptides iden-tified in the present study that contain 2 or 3additional N-terminal amino acids are agonists. An-other example of bioactive peptides being eitheragonists or antagonists of the same receptor, depend-ing on peptide length, is that of �-endorphin (57).Another finding from these studies is that longerHp-derived peptides activate signaling pathways dis-tinct from the classic CB1 receptor agonist, Hu-210,in that 1) the peptides do not induce increases in[35S]GTPS binding, whereas Hu-210 does; 2) thelevel and rate of increase in phosphorylation ofERK1/2 by these peptides are different from those ofHu-210; 3) the levels of intracellular Ca2� release bythese peptides are much more robust than thoseinduced by Hu-210; and 4) the effects of the peptideson signaling are partially blocked by PTX in contrastto Hu-210, which is completely blocked. The Ca2�
release generated by longer Hp-derived peptidesinvolves a longer time scale (30 min) than generallyseen after activation of G�q receptors (seconds). Theobservations that peptide-mediated Ca2� release isblocked by the CB1 antagonist, is partially blocked byPTX, and involves release of Ca2� from intracellularstores suggest that the longer Hp peptides activate aG� pathway. Because the classic CB1 agonist, Hu-210,and the endogenous agonist, 2-AG, only cause a smallincrease in Ca2� release, the robust and sustainedlevel observed with longer Hp peptides suggests anunexpected lack of receptor down-regulation that iscurrently under investigation. The finding that RVD-Hp� and VD-Hp� possess unique agonistic activity atCB1 receptors provides additional tools to under-stand how the endocannabinoid system is modulatedas well as novel candidates to be developed as thera-peutic agents in the treatment of pathological con-ditions involving cannabinoid receptors.
This work was supported by NIH grants (DA019521 toL.A.D., GM05313 to S.S., GM071558 to L.A.D. and S.S., andDA04494 and DK51271 to L.D.F.), by a Zwanenberg Founda-tion Fellowship (to S.H.), by the Fundacao de Amparo aPesquisa do Estado de Sao Paulo (Grants 04/04933-2 to E.S.F.and 04/14258-0 to A.S.H.), and by Conselho Nacional deDesenvolvimento Científico e Tecnologico (E.S.F.). Massspectrometry was performed through Rede de Proteoma doEstado de Sao Paulo in the Laboratorio Nacional de LuzSincrotron, Campinas, Sao Paulo, Brazil. We thank Dr. K.Mackie (Indiana University, Bloomington, IN, USA) for theplasmid cDNAs for CB1 cannabinoid and CB2 cannabinoidreceptors and for GPR55. We also thank Emeline Maillet and
3027ENDOGENOUS PEPTIDE AGONISTS TO CANNABINOID RECEPTORS

Robert Margolskee for help with analysis using the FLEXStation.
REFERENCES
1. Heimann, A. S., Gomes, I., Dale, C. S., Pagano, R. L., Gupta, A.,de Souza, L. L., Luchessi, A. D., Castro, L. M., Giorgi, R., Rioli,V., Ferro, E. S., and Devi, L. A. (2007) Hemopressin in aninverse agonist of CB1 cannabinoid receptors. Proc. Natl. Acad.Sci. U. S. A. 104, 20588–20593
2. Begg, M., Pacher, P., Batkai, S., Osei-Hyiaman, D., Offertaler, L.,Mo, F. M., Liu, J., and Kunos, G. (2005) Evidence for novelcannabinoid receptors. Pharmacol. Ther. 106, 133–145
3. Matsuda, L. A., Lolait, S. J., Brownstein, M. J., Young, A. C., andBonner, T. I. (1990) Structure of a cannabinoid receptor andfunctional expression of the cloned cDNA. Nature 346, 561–564
4. Munro, S., Thomas, K. L., and Abu-Shaar, M. (1993) Molecularcharacterization of a peripheral receptor for cannabinoids.Nature 365, 61–65
5. Van Sickle, M. D., Duncan, M., Kingsley, P. J., Mouihate, A.,Urbani, P., Mackie, K., Stella, N., Makriyannis, A., Piomelli, D.,Davison, J. S., Marnett, L. J., Di Marzo, V., Pittman, Q. J., Patel,K. D., and Sharkey, K. A. (2005) Identification and functionalcharacterization of brainstem cannabinoid CB2 receptors. Sci-ence 310, 329–332
6. Zhang, J., Hoffert, C., Vu, H. K., Groblewski, T., Ahmad, S., andO’Donnell, D. (2003) Induction of CB2 receptor expression inthe rat spinal cord of neuropathic but not inflammatory chronicpain models. Eur. J. Neurosci. 17, 2750–2754
7. Lauckner, J. E., Jensen, J. B., Chen, H. Y., Lu, H. C., Hille, B.,and Mackie, K. (2008) GPR55 is a cannabinoid receptor thatincreases intracellular calcium and inhibits M current. Proc.Natl. Acad. Sci. U. S. A. 105, 2699–2704
8. Pertwee, R. G. (1997) Pharmacology of cannabinoid CB1 andCB2 receptors. Pharmacol. Ther. 74, 129–180
9. Pertwee, R. G., and Ross, R. A. (2002) Cannabinoid receptorsand their ligands. Prostaglandins Leukot. Essent. Fatty Acids 66,101–121
10. Howlet, A. C., and Mukhopadhyay, S. (2000) Cellular signaltransduction by anandamide and 2-arachidonoylglycerol. Chem.Phys. Lipids 108, 53–70
11. Di Marzo, V., and Petrosino, S. (2007) Endocannabinoids andthe regulation of their levels in health and disease. Curr. Opin.Lipidol. 18, 129–140
12. Battista, N., Fezza, F., Finazzi-Agro, A., and Maccarrone, M.(2006) The endocannabinoid system in neurodegeneration.Ital. J. Biochem. 55, 283–289
13. Boyd, S. T. (2006) The endocannabinoid system. Pharmacother-apy 26, 218S–221S
14. Iversen, L., and Chapman, V. (2002) Cannabinoids: a realprospect for pain relief. Curr. Opin. Pharmacol. 2, 50–55
15. Brantl, V., Gramsch, C., Lottspeich, F., Mertz, R., Jaeger, K. H.,Herz, A. (1986) Novel opioid peptides derived from hemoglo-bin: hemorphins. Eur. J. Pharmacol. 125, 309–310
16. Liebmann, C., Schrader, U., and Brantl, V. (1997) Opioidreceptor affinities of the blood-derived tetrapeptides hemor-phin and cytochrophin. Eur. J. Pharmacol. 166, 523–526
17. Moeller, I., Lew, R. A., Mendelsohn, F. A., Smith, A. I., Brennan,M. E., Tetaz, T. J., and Chai, S. Y. (1997) The globin fragmentLVV-hemorphin-7 is an endogenous ligand for the AT4 recep-tor in the brain. J. Neurochem. 68, 2530–2537
18. Lee, J., Mustafa, T., McDowall, S. G., Mendelsohn, F. A.,Brennan, M., Lew, R. A., Albiston, A. L., and Chai, S. Y. (2003)Structure-activity study of LVV-hemorphin-7: angiotensin AT4receptor ligand and inhibitor of insulin-regulated aminopepti-dase. J. Pharmacol. Exp. Ther. 305, 205–211
19. Fukui, K., Shiomi, H., Takagi, H., Hayashi, K., Kiso, Y., andKitagawa, K. (1983) Isolation from bovine brain of a novelanalgesic peptapeptide, neo-kyotorphin, containing the Tyr-Arg(kyotorphin) unit. Neuropharmacology 22, 191–196
20. Che, F. Y., Lim, J., Pan, H., Biswas, R., and Fricker, L. D. (2005)Quantitative neuropeptidomics of microwave-irradiated mousebrain and pituitary. Mol. Cell. Proteomics 4, 1391–1405
21. Che, F. Y., Zhang, X., Berezniuk, I., Callaway, M., Lim, J., andFricker, L. D. (2007) Optimization of neuropeptide extractionfrom the mouse hypothalamus. J. Proteome Res. 6, 4667–4676
22. Fricker, L. D. (2007) Neuropeptidomics to study peptide pro-cessing in animal models of obesity. Endocrinology 148, 4185–4190
23. Pan, H., Che, F. Y., Peng, B., Steiner, D. F., Pintar, J. E., andFricker, L. D. (2006) The role of prohormone convertase-2 inhypothalamic neuropeptide processing: a quantitative neu-ropeptidomic study. J. Neurochem. 98, 1763–1777
24. Decaillot, F. M., Che, F. Y., Fricker, L. D., and Devi, L. A. (2006)Peptidomics of Cpefat/fat mouse hypothalamus and striatum:effect of chronic morphine administration. J. Mol. Neurosci. 28,277–284
25. Che, F. Y., Vathy, I., and Fricker, L. D. (2006) Quantitativepeptidomics in mice: effect of cocaine treatment. J. Mol. Neuro-sci. 28, 265–275
26. Lim, J., Berezniuk, I., Che, F. Y., Parikh, R., Biswas, R., Pan, H.,and Fricker, L. D. (2006) Altered neuropeptide processing inprefrontal cortex of Cpefat/fat mice: implications for neuropep-tide discovery. J. Neurochem. 96, 1169–1181
27. Fricker, L. D., Lim, J., Pan, H., and Che, F. Y. (2006) Peptido-mics: identification and quantitation of endogenous peptides inneuroendocrine tissues. Mass Spectrom. Rev. 25, 327–344
28. Pan, H., Nanno, D., Che, F. Y., Zhu, X., Salton, S. R., Steiner,D. F., Fricker, L. D., and Devi, L. A. (2005) Neuropeptideprocessing profile in mice lacking prohormone convertase-1.Biochemistry 44, 4939–4948
29. Che, F. Y., Biswas, R., and Fricker, L. D. (2005) Relativequantitation of peptides in wild-type and Cpefat/fat mouse pitu-itary using stable isotopic tags and mass spectrometry. J. MassSpectrom. 40, 227–237
30. Che, F. Y., and Fricker, L. D. (2005) Quantitative peptidomics ofmouse pituitary: comparison of different stable isotopic tags. J.Mass Spectrom. 40, 238–249
31. Che, F. Y., Yuan, Q., Kalinina, E., and Fricker, L. D. (2005)Peptidomics of Cpefat/fat mouse hypothalamus: effects of fooddeprivation and exercise on peptide levels. J. Biol. Chem. 280,4451–4461
32. Gomes, I., Filipovska, J., and Devi, L. A. (2003) Opioid receptoroligomerization: detection and functional characterization ofinteracting receptors. Methods Mol. Med. 84, 157–183
33. Jordan, J. D., He, J. C., Eungdamrong, N. J., Gomes, I., Ali, W.,Nguyen, T., Bivona, T. G., Philips, M. R., Devi, L. A., andIyengar, R. (2005) Cannabinoid receptor-induced neurite out-growth is mediated by Rap1 activation through G�o/i-triggeredproteasomal degradation of Rap1GAPII. J. Biol. Chem. 280,11413–11421
34. He, J. C., Gomes, I., Nguyen, T., Jayaram, G., Ram, P. T., Devi,L. A., and Iyengar, R. (2005) The G�o/i-coupled cannabinoidreceptor-mediated neurite outgrowth involves Rap regulation ofSrc and Stat3. J. Biol. Chem. 280, 33426–33434
35. Hsieh, C., Brown, S., Derleth, C., and Mackie, K. J. (1999)Internalization and recycling of the CB1 cannabinoid receptor.J. Neurochem. 73, 493–501
36. Coutts, A. A., Anavi-Goffer, S., Ross, R. A., MacEwan, D. J.,Mackie, K., Pertwee, R. G., and Irving, A. J. (2001) Agonist-induced internalization and trafficking of cannabinoid CB1receptors in hippocampal neurons. J. Neurosci. 21, 2425–2433
37. Lauckner, J. E., Hille, B., and Mackie, K. (2005) The cannabi-noid agonist WIN55,2-12-2 increases intracellular calcium viaCB1 receptor coupling to Gq/11 G proteins. Proc. Natl. Acad. Sci.U. S. A. 102, 19144–19149
38. Drake, M. T., Violin, J. D., Whalen, E. J., Wisler, J. W., Shenoy,S. K., and Lefkowitz, R. J. (2008) �-Arrestin-biased agonism and�2-adrenergic receptor. J. Biol. Chem. 283, 5669–5676
39. Goldberg, A. L. (2003) Protein degradation and protectionagainst misfolded or damaged proteins. Nature 426, 895–899
40. Costa, E., and Guidotti, A. (1991) Diazepam binding inhibitor(DBI): a peptide with multiple biological actions. Life Sci. 49,325–344
41. Goldstein, A. L., Hannappel, E., and Kleinman, H. K. (2005)Thymosin �4: actin-sequestering protein moonlights to repairinjured tissues. Trends Mol. Med. 11, 421–429
42. Ojika, K., Mitake, S., Tohdoh, N., Appel, S. H., Otsuka, Y.,Katada, E., and Matsukawa, N. (2000) Hippocampal cholinergicneurostimulating peptides (HCNP). Prog. Neurobiol. 60, 37–83
3028 Vol. 23 September 2009 GOMES ET AL.The FASEB Journal � www.fasebj.org

43. Mukai, H., Hokari, Y., Seki, T., Takao, T., Kubota, M., Matsuo,Y., Tsukagoshi, H., Kato, M., Kimura, H., Shimonishi, Y., Kiso, Y.,Nishi, Y., Wakamatsu, K., and Munekata, E. (2008) Discovery ofmitocryptide-1, a neutrophil-activating cryptide from healthyporcine heart. J. Biol. Chem. 283, 30596–30605
44. Skold, K., Svensson, M., Norrman, M., Sjogren, B., Svennings-son, P., and Andren, P. E. (2007) The significance of biochem-ical and molecular sample integrity in brain proteomics andpeptidomics: stathmin 2-20 and peptides as sample qualityindicators. Proteomics 7, 4445–4456
45. Bazinet, R. P., Lee, H. J., Felder, C. C., Porter, A. C., Rapoport,S. I., and Rosenberger, T. A. (2005) Rapid high energy micro-wave fixation is required to determine the anandamide (N-arachidonoylethanolamine) concentration of rat brain. Neuro-chem. Res. 30, 597–601
46. Dassen, H., Kamps, R., Punyadeera, C., Dijcks, F., de Goeij, A.,Ederveen, A., Dunselman, G., and Groothuis, P. (2008) Haemo-globin expression in human endometrium. Hum. Reprod. 23,635–641
47. Wride, M. A., Mansergh, F. C., Adams, S., Everitt, R., Minnema,S. E., Rancourt, D. E., and Evans, M. J. (2003) Expressionprofiling and gene discovery in the mouse lens. Mol. Vis. 9,360–396
48. Newton, D. A., Rao, K. M., Dluhy, R. A., and Baatz, J. E. (2006)Hemoglobin is expressed by alveolar epithelial cells. J. Biol.Chem. 281, 5668–5676
49. Bhaskaran, M., Chen, H., Chen, Z., and Liu, L. (2005) Hemo-globin is expressed in alveolar epithelial type II cells. Biochem.Biophys. Res. Commun. 333, 1348–1352
50. Liu, L., Zeng, M., and Stamler, J. S. (1999) Hemoglobininduction in mouse macrophages. Proc. Natl. Acad. Sci. U. S. A.96, 6643–6647
51. Nishi, H., Inagi, R., Kato, H., Tanemoto, M., Kojima, I., Son, D.,Fujita, T., and Nangaku, M. (2008) Hemoglobin is expressed by
mesangial cells and reduces oxidant stress. J. Am. Soc. Nephrol.19, 1500–1508
52. Richter, F., Meurer, B. H., Zhu, C., Madvedeva, V. P,. andChesselet, M. F. (2009) Neurons express hemoglobin � and �chains in rat and human brains. [E-pub ahead of print] J. Comp.Neurol. doi:10.1002/cne.22062
53. Slobodyansky, E., Guidotti, A., Wambebe, C., Berkovich, A., andCosta, E. (1989) Isolation and characterization of a rat braintriakontatetraneuropeptide, a posttranslational product of diaz-epam binding inhibitor: specific action at the Ro 5-4864 recog-nition site. J. Neurochem. 53, 1276–1284
54. Goumon, Y., Angelone, T., Schoentgen, F., Chasserot-Golaz, S.,Almas, B., Fukami, M. M., Langley, K., Welters, I. D., Tota, B.,Aunis, D., and Metz-Boutigue, M. H. (2004) The hippocampalcholinergic neurostimulating peptide, the N-terminal fragmentof the secreted phosphatidylethanolamine-binding protein, pos-sesses a new biological activity on cardiac physiology. J. Biol.Chem. 279, 13054–13064
55. Badamchian, M., Damavandy, A. A., Damavandy, H., Wadhwa,S. D., Katz, B., and Goldstein, A. L. (2007) Identification andquantification of thymosin �4 in human saliva and tears. Ann.N. Y. Acad. Sci. 1112, 458–465
56. Ferro, E. S., Carreno, F. R., Goni, C., Garrido, P. A., Guimaraes,A. O., Castro, L. M., Oliveira, V., Araujo, M. C., Rioli, V., Gomes,M. D., Fontenele-Neto, J. D., and Hyslop, S. (2004) The intra-cellular distribution and secretion of endopeptidases 24.15 (EC3.4.24.15) and 24.16 (EC 3.4.24.16). Protein Pept. Lett. 11,415–421
57. Millington, W. R., and Smith, D. L. (1991) The posttranslationalprocessing of �-endorphin in human hypothalamus. J. Neuro-chem. 57, 775–781
Received for publication February 17, 2009.Accepted for publication March 26, 2009.
3029ENDOGENOUS PEPTIDE AGONISTS TO CANNABINOID RECEPTORS