morphometric, elemental and biochemical characterisation of three haliclona species (demospongiae)...
TRANSCRIPT

Chemical Speciation and Bioavailability (2013), 25(1) 15
Doi: 10.3184/095422912X13496990950939
Morphometric, elemental and biochemical characterisation of three Haliclona species (Demospongiae) from the Gulf of Mannar, India
Srikanth Koigooraa, Ramjee Pallelaa,b, Chakra Reddy Nelakurtia, Madhavendra Sakunthala Sunkarac and Venkateswara Rao Janapalaa*
aToxicology Unit, Biology Division, Indian Institute of Chemical Technology, Hyderabad - 500607, IndiabDepartment of Chemistry and Institute of BioPhysico Sensor Technology, Pusan National University, Busan - 609735, South KoreacElectron Microscopy Division, Indian Institute of Chemical Technology, Hyderabad - 500607, India*E-mail: [email protected]
ABSTRACT
Marine sponges, in particular the Haliclona species (Demospongiae: Haplosclerida: Chalinidae) and/or their associated microorganisms, continue to yield an array of novel compounds. The aim of this work was the characterisation (morphometric, elemental and biochemical) of the three Haliclona species (Haliclona tenuiramosa, Haliclona cribricutis and Haliclona retiderma) from the Mandapam coast, Gulf of Mannar, India as an aid in the discrimination of the desired species for in vitro production of bioactive products and further ecobiotechnological applications. Diversity of the Haliclona species was analysed based on morphometric variables and elemental composition of the pinacoderm (outer epithelioid layer). Scanning electron microscope equipped with energy dispersive X-ray analysis did not reveal any change in elemental composition in spicules; however, the percentage of elemental composition in pinacoderm was significantly different among the species. The order and range of pinacoderm elemental composition were chlorine (36–51%), sodium (29–43%), silicon (0.72–2.2%), iron (0.46–0.73%). Aluminium (1.86–8.88%), calcium (0.92–16.19%) and sulfur (0.44–9.74%) varied greatly among the species. Total carbohydrate, protein and lipid contents also showed diversity among the species, i.e., carbohydrate content was high in H. tenuiramosa; proteins in H. cribricutis and lipids in H. retiderma, respectively. The order of ratio of spicule/protein content among the species were H. retiderma > H. cribricutis > H. tenuiramosa. The current approaches and results could direct an efficient application of discriminating sponge species in situ.
Keywords: marine sponges, Haliclona, morphometry, scanning electron microscope, spicules, biochemical composition, Gulf of Mannar
www.chemspecbio.co.uk
INTRODUCTION
Sponges (Porifera) are sessile creatures distributed in marine or freshwater from the deep sea to the coast and are dominant among marine species in number, especially of the deep sea habitat (Hooper and Van Soest, 2002; Yi et al., 2005). Sponges have a wide array of colours, textures, sizes, and shapes and, unlike all other marine invertebrates, they lack symmetry and have no true tissues or organs. These are composed of highly differentiated mesohyl containing spicules of different shapes and specialised cells of diverse functions (Jones, 1994). These sponges are divided into three classes based on the
type of spicules in their skeleton viz., bony (Calcarea), glass (Hexactenellida), and spongin (Demospongiae) (Diaz and Van Soest, 1994; Kerr and Lelly-Borges, 1994; Rodrigo et al., 1994; Ya-sheng, 1995; Hooper and Van Soest, 2002). The members of the class Demospongiae have a living tissue surrounding the pinacoderm, called the mesohyl, which contains a matrix composed of a striated protein spongin, an organic skeleton composed of collagen, and/or an inorganic skeleton composed of mineral spicules. This skeleton of gelatinous matrix, often embedded with foreign particles, causes identification problems in situ. Usually, the characteristic features of each species of demosponges are based on the climatic and environmental
CSB1100060.indd 15 2/6/2013 8:41:23 AM

16 Characterisation of three Haliclona species from the Gulf of Mannar
factors (Palumbi, 1984; Bell and Barnes, 2000). Moreover, the global distribution of these species has varied considerably depending on nutritional status of each species and the external environmental factors (Bell et al., 2002).
Marine sponges, especially the demosponges, have attracted significant attention from various scientific disciplines because of their possession of chemical molecules and proteins such as collagens (Venkateswarlu et al., 1994; Zhou and Molinski, 2003; Pallela et al., 2011a). As they produce various novel chemical molecules, sponges have been of great interest to chemists and have led to biotechnological applications and potential for drug discovery. Sponges can be considered as microbial fermenters that provide new avenues in marine microbiology and biotechnology, by harboring symbionts including bacteria, cyanobacteria, red algae, green algae, diatoms and dinoflagellates, fungi (Osinga et al., 1999; Hentschel et al., 2006; Wang, 2006). The association of symbionts mainly depends on the network of chambers and canals of sponges, where the canal formation and growth stage of the sponge larvae determines which symbiont to associate at specific regions of the sponge (Wilkinson, 1992; Osinga et al., 2001; Usher et al., 2005). Cell biologists have investigated sponges and their symbionts in order to understand their basic organisation on the cellular and skeletal level to procure biologically important molecules (Sipkema et al., 2005). However, knowledge of their morphometric, biochemical and ecological characteristics are so far limited (Rodrigo et al., 1994; Sipkema et al., 2003; Gunda and Janapala, 2009; Sipkema et al., 2009).
The inaccessibility of the marine habitat for monitoring purposes causes problems for species identification and classification. Marine taxonomists have focused their attention on understanding the characteristic internal organisation of individual species. Species description and anatomical structure of sponges are needed to select targeted species to further study the culturing of sponge cells/fragments/whole sponges in situ as well as ex situ for the in vitro synthesis of novel molecules through approached biotechnology (Koopmans et al., 2009; de Caralt et al., 2010). Elemental composition in marine sponges
can also be used as biomarkers to assess pollution risks and ecosystem health in the ocean (Venkateswara Rao et al., 2006; Venkateswara Rao et al., 2009). In this paper, biochemical and morphometric variations of three Haliclona species (H. retiderma, H. tenuiramosa and H. cribricutis) of common habitat at Mandapam coast of Gulf of Mannar (GoM), were compared to assess the morphology of these species for our future collections and culturing their cells in vitro to produce secondary metabolites. Recently, similar ecotaxonomic analyses of three Petrosiidae species available in the same area of GoM have been reported (Rodrigo et al., 1994).
METHODOLOGY
Sampling
The test sponges were collected in the year 2009 during low tide times from the shallow sub-tidal coasts by snorkeling and skin-diving, at Mandapam coast (9° 10´ to 9° 50´ N; 78 10´ to 79° 07´ E) of Gulf of Mannar (GoM), India (Figure 1). Sponges were carefully collected by gently removing from the substratum to minimise the tissue damage, and were placed in plastic bags underwater, then transferred into large containers of aerated seawater for transporting to the laboratory. Before analysis, all the sponges were thoroughly cleaned by mechanical removal of foreign materials followed by repeated washing with artificial seawater (Millero, 1996).
The voucher specimens were submitted to National Institute of Oceanography (NIO), Goa for depository purpose, and were identified as H. retiderma (Dendy, 1922) (presently referred as Neopetrosia retiderma), H. tenuiramosa and H. cribricutis (Class: Demospongiae; Order: Haplosclerida and Family: Chalinidae) by Dr P. A. Thomas, Vizhinjam Research Centre of Central Marine Fisheries Research Institute (ICAR), Vizhinjam, Thiruvananthapuram, India (Thomas, 1998). Based on our earlier expeditions (2000 to 2009), it was noticed that these three types of Haliclona species exist abundantly at the Mandapam coast of GoM.
Figure 1 Location map of sponge sampling site (Mandapam region, Gulf of Mannar, Southeast Coast of India).
CSB1100060.indd 16 2/6/2013 8:41:24 AM

Srikanth Koigoora, Ramjee Pallela, Chakra Reddy Nelakurti, Madhavendra Sakunthala Sunkara and Venkateswara Rao Janapala 17
Morphological and architectural studies
Handmade tangential sections were prepared by using a razor blade to cut the sponge surface (pinacoderm). The pinacodermal sections of individual sponge species were observed with the help of video microscope (High scope - Compact Micro vision system: Model No. KH-2200 MD2) and their digital photographs were used to analyse the differences among the species.
Isolation and morphometry of spicules
Isolation of spicules was performed based on the modified method of Wang and Lavrov (2007). Specified pieces of individual species, consisting of two primary layers of cells (pinacoderm and choanoderm) and an inner cellular collagenous matrix (mesohyl) were used (a minimum of three replicates each) to digest with conc. HNO
3. Adequate amount
of sponge samples were allowed to stand for 2–3 hours until all the tissue pieces were dissolved and then heated gently over a Bunsen flame until the liquid was clear. These aliquots (n=5, each in 10 mL volume) of all the digested sponge samples were centrifuged at 5000g for 10 minutes and the precipitate was re-centrifuged after every wash with distilled water for three times. To clean up any additional organic matter sticking to the spicules, H
2O
2 was added and the last
cleaning was done with absolute ethanol and also to dilute the spicules.
The length and width of spicules (n=500) isolated from individual Haliclona species were calculated, by a compound microscope (POLYVAR, Reichert–Jung light microscope) attached to Ethovision–version 2.3 (Noldus Information Technology, The Netherlands) through a CCD camera (Sony CCD IRIS, Model No: SSC-M370CE). Various kinds of scleroid crystals were also observed for each individual species. The magnification was calibrated with the aid of ocular and stage micrometers (ERMA, Tokyo, Japan).
Analytical studies
Elemental analyses
The elemental composition of pinacoderm (Al, Ca, Cl, Fe, Na, S and Si) and spicules in different species of Haliclona (n=5) was analysed by HITACHI S-520 scanning electron microscope, equipped with energy dispersive X-ray analysis (EDXA). Samples were mounted on aluminium stubs with double adhesive tape, coated with gold in HITACHI HUS-SGB Vacuum and observed in Hitachi S-520 SEM. Then EDXA carried out with Oxford Link ISIS-300 detector calibrated with Cobalt standard at an acceleration voltage of 20 kV. Built-in standards were used for the effective quantification of each element available in spicules as well as the pinacoderm (Araújo et al., 2003; Pallela et al., 2011b).
Biochemical analyses
The sponge samples were processed individually (n=5) for estimating macromolecules such as carbohydrates,
proteins and lipid contents in individual species. The total carbohydrate content was assayed by the phenol–sulfuric acid method (Taylor, 1995) and the total protein content was estimated using folin–phenol reagent using bovine serum albumin as standard (Bradford, 1976). The total lipid content was measured by using 1:2 mixture of chloroform: methanol using the modified procedure of Brooks et al. (1998).
Statistical analyses
Data was expressed as mean (±SE) calculated for each variable of independent species (n=3 to 5). The suitability of the data for spicule analysis was evaluated to examine individual variables (morphological data) for the differences among test species by One-way analysis of variance (ANOVA). When populations were significantly different, multiple comparison post-hoc tests (Tukey’s HSD) were performed to see which populations differed from one another.
RESULTS
Morphological and architectural studies
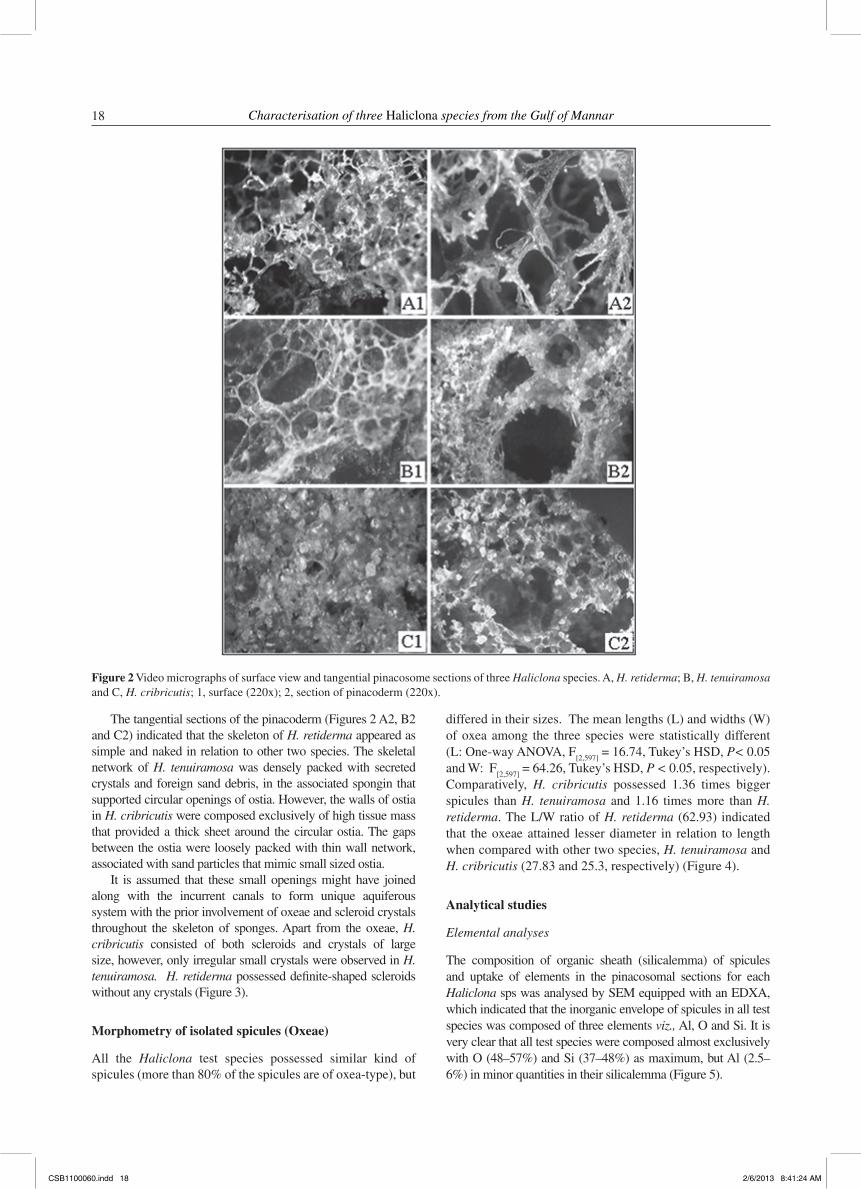
The three Haliclona species collected at Mandapam coast at same depth and environmental conditions. Much intraspecific differentiation was noticed from the gross morphological studies of the three test species. Specific arrangement of spicules combined with other tissue components offered a rigid and unique architecture to each of the individual species. Externally, the three Haliclona species appeared as arborescent and cushion shaped, more frequently visible with oscular mounds or chimneys to give a branching, tube-shaped to ramose morphotype. The outer ectosome of H. cribricutis, which functions as a defensive outer layer to impair settlement of organisms, was slightly elastic, appearing dark brownish black in colour; whereas, H. tenuiramosa showed a smooth surface, with a firm ectosomal layer of purple or violet. However, H. retiderma was fragile and brittle with oscula usually circular with the unispicular and isotropic surface. The video micrographs of the upper surface of individual test species indicated the differences in the architecture of ostia surrounded by varyingly thickened walls that were quite special for each species.
During comparative microscopic observations of the external surfaces of the sponges, very loose body texture was observed due to the peculiar arrangement of the spicules (by the impregnation of amorphous material leading to difference in the compartmental spaces) in H. retiderma followed by H. cribricutis and H. tenuiramosa. The external surface of all the Haliclona species were supported by the presence of different sizes of crystals. The surface of H. retiderma possessed small, bright and shiny crystals. Prominent crystals were observed to be embedded on the peripheries of ostia of H. cribricutis. There are significant interspecific changes in shape and form of ostia, which play a unique role in the makeup of aquiferous system among these test species (Figures 2 A1, B1 and C1).
CSB1100060.indd 17 2/6/2013 8:41:24 AM
18 Characterisation of three Haliclona species from the Gulf of Mannar
The tangential sections of the pinacoderm (Figures 2 A2, B2 and C2) indicated that the skeleton of H. retiderma appeared as simple and naked in relation to other two species. The skeletal network of H. tenuiramosa was densely packed with secreted crystals and foreign sand debris, in the associated spongin that supported circular openings of ostia. However, the walls of ostia in H. cribricutis were composed exclusively of high tissue mass that provided a thick sheet around the circular ostia. The gaps between the ostia were loosely packed with thin wall network, associated with sand particles that mimic small sized ostia.
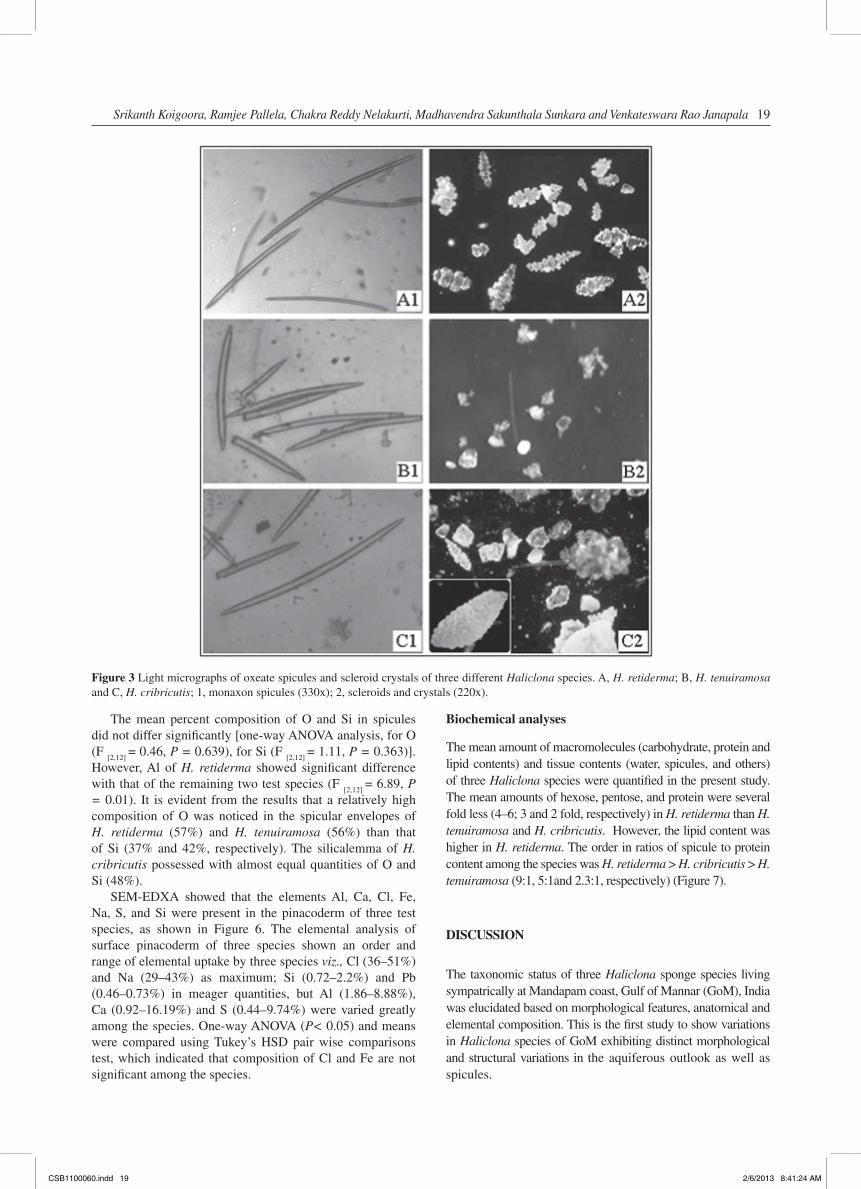
It is assumed that these small openings might have joined along with the incurrent canals to form unique aquiferous system with the prior involvement of oxeae and scleroid crystals throughout the skeleton of sponges. Apart from the oxeae, H. cribricutis consisted of both scleroids and crystals of large size, however, only irregular small crystals were observed in H. tenuiramosa. H. retiderma possessed definite-shaped scleroids without any crystals (Figure 3).
Morphometry of isolated spicules (Oxeae)
All the Haliclona test species possessed similar kind of spicules (more than 80% of the spicules are of oxea-type), but
differed in their sizes. The mean lengths (L) and widths (W) of oxea among the three species were statistically different (L: One-way ANOVA, F
[2,597] = 16.74, Tukey’s HSD, P< 0.05
and W: F[2,597]
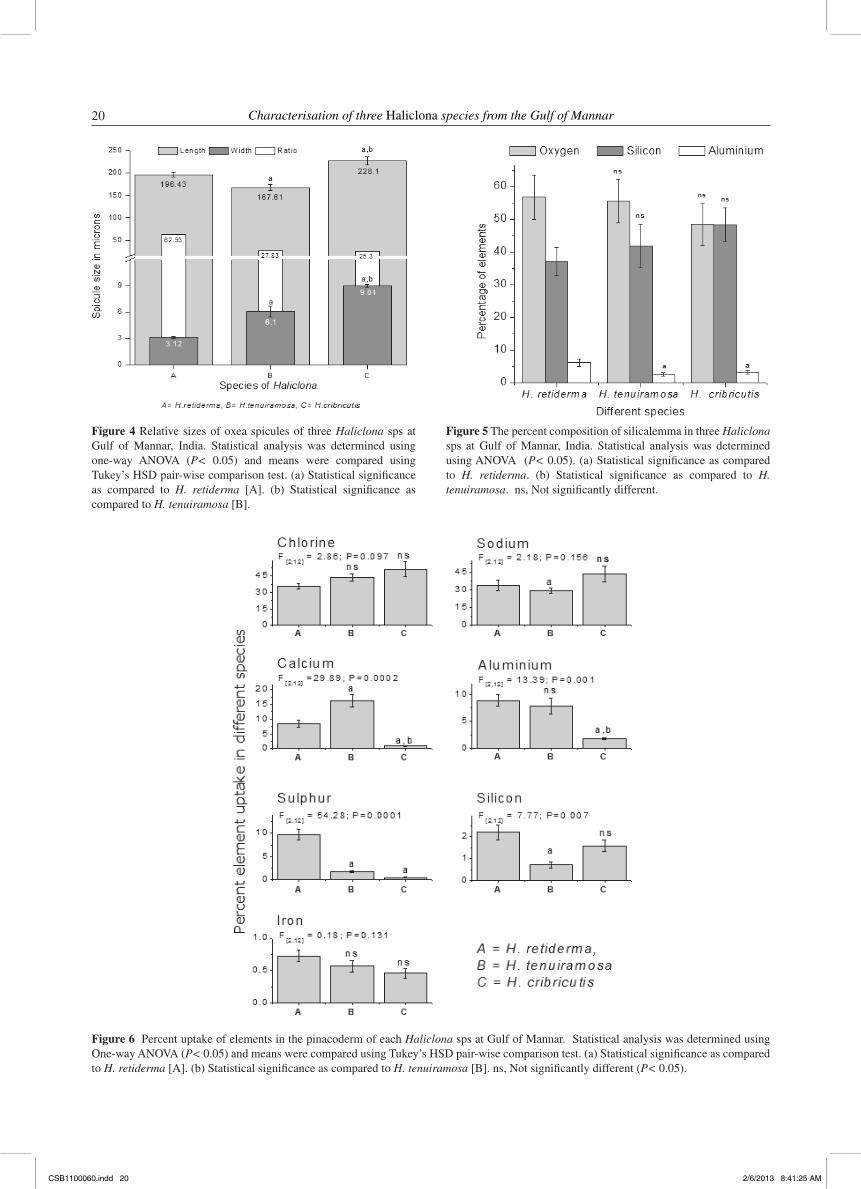
= 64.26, Tukey’s HSD, P < 0.05, respectively). Comparatively, H. cribricutis possessed 1.36 times bigger spicules than H. tenuiramosa and 1.16 times more than H. retiderma. The L/W ratio of H. retiderma (62.93) indicated that the oxeae attained lesser diameter in relation to length when compared with other two species, H. tenuiramosa and H. cribricutis (27.83 and 25.3, respectively) (Figure 4).
Analytical studies
Elemental analyses
The composition of organic sheath (silicalemma) of spicules and uptake of elements in the pinacosomal sections for each Haliclona sps was analysed by SEM equipped with an EDXA, which indicated that the inorganic envelope of spicules in all test species was composed of three elements viz., Al, O and Si. It is very clear that all test species were composed almost exclusively with O (48–57%) and Si (37–48%) as maximum, but Al (2.5–6%) in minor quantities in their silicalemma (Figure 5).
Figure 2 Video micrographs of surface view and tangential pinacosome sections of three Haliclona species. A, H. retiderma; B, H. tenuiramosa and C, H. cribricutis; 1, surface (220x); 2, section of pinacoderm (220x).
CSB1100060.indd 18 2/6/2013 8:41:24 AM
Srikanth Koigoora, Ramjee Pallela, Chakra Reddy Nelakurti, Madhavendra Sakunthala Sunkara and Venkateswara Rao Janapala 19
The mean percent composition of O and Si in spicules did not differ significantly [one-way ANOVA analysis, for O (F
[2,12] = 0.46, P = 0.639), for Si (F
[2,12] = 1.11, P = 0.363)].
However, Al of H. retiderma showed significant difference with that of the remaining two test species (F
[2,12] = 6.89, P
= 0.01). It is evident from the results that a relatively high composition of O was noticed in the spicular envelopes of H. retiderma (57%) and H. tenuiramosa (56%) than that of Si (37% and 42%, respectively). The silicalemma of H. cribricutis possessed with almost equal quantities of O and Si (48%).
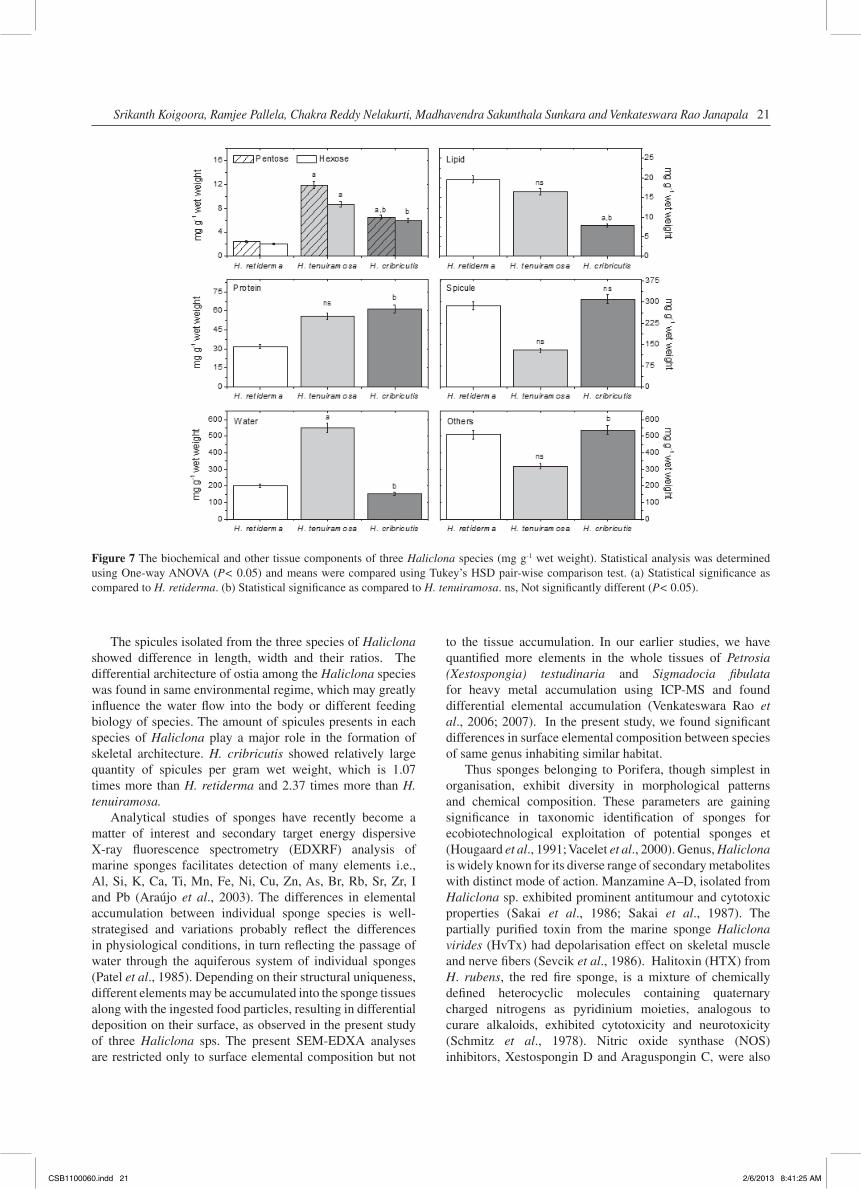
SEM-EDXA showed that the elements Al, Ca, Cl, Fe, Na, S, and Si were present in the pinacoderm of three test species, as shown in Figure 6. The elemental analysis of surface pinacoderm of three species shown an order and range of elemental uptake by three species viz., Cl (36–51%) and Na (29–43%) as maximum; Si (0.72–2.2%) and Pb (0.46–0.73%) in meager quantities, but Al (1.86–8.88%), Ca (0.92–16.19%) and S (0.44–9.74%) were varied greatly among the species. One-way ANOVA (P< 0.05) and means were compared using Tukey’s HSD pair wise comparisons test, which indicated that composition of Cl and Fe are not significant among the species.
Biochemical analyses
The mean amount of macromolecules (carbohydrate, protein and lipid contents) and tissue contents (water, spicules, and others) of three Haliclona species were quantified in the present study. The mean amounts of hexose, pentose, and protein were several fold less (4–6; 3 and 2 fold, respectively) in H. retiderma than H. tenuiramosa and H. cribricutis. However, the lipid content was higher in H. retiderma. The order in ratios of spicule to protein content among the species was H. retiderma > H. cribricutis > H. tenuiramosa (9:1, 5:1and 2.3:1, respectively) (Figure 7).
DISCUSSION
The taxonomic status of three Haliclona sponge species living sympatrically at Mandapam coast, Gulf of Mannar (GoM), India was elucidated based on morphological features, anatomical and elemental composition. This is the first study to show variations in Haliclona species of GoM exhibiting distinct morphological and structural variations in the aquiferous outlook as well as spicules.
Figure 3 Light micrographs of oxeate spicules and scleroid crystals of three different Haliclona species. A, H. retiderma; B, H. tenuiramosa and C, H. cribricutis; 1, monaxon spicules (330x); 2, scleroids and crystals (220x).
CSB1100060.indd 19 2/6/2013 8:41:24 AM
20 Characterisation of three Haliclona species from the Gulf of Mannar
Figure 4 Relative sizes of oxea spicules of three Haliclona sps at Gulf of Mannar, India. Statistical analysis was determined using one-way ANOVA (P< 0.05) and means were compared using Tukey’s HSD pair-wise comparison test. (a) Statistical significance as compared to H. retiderma [A]. (b) Statistical significance as compared to H. tenuiramosa [B].
Figure 5 The percent composition of silicalemma in three Haliclona sps at Gulf of Mannar, India. Statistical analysis was determined using ANOVA (P< 0.05). (a) Statistical significance as compared to H. retiderma. (b) Statistical significance as compared to H. tenuiramosa. ns, Not significantly different.
Figure 6 Percent uptake of elements in the pinacoderm of each Haliclona sps at Gulf of Mannar. Statistical analysis was determined using One-way ANOVA (P< 0.05) and means were compared using Tukey’s HSD pair-wise comparison test. (a) Statistical significance as compared to H. retiderma [A]. (b) Statistical significance as compared to H. tenuiramosa [B]. ns, Not significantly different (P< 0.05).
CSB1100060.indd 20 2/6/2013 8:41:25 AM
Srikanth Koigoora, Ramjee Pallela, Chakra Reddy Nelakurti, Madhavendra Sakunthala Sunkara and Venkateswara Rao Janapala 21
The spicules isolated from the three species of Haliclona showed difference in length, width and their ratios. The differential architecture of ostia among the Haliclona species was found in same environmental regime, which may greatly influence the water flow into the body or different feeding biology of species. The amount of spicules presents in each species of Haliclona play a major role in the formation of skeletal architecture. H. cribricutis showed relatively large quantity of spicules per gram wet weight, which is 1.07 times more than H. retiderma and 2.37 times more than H. tenuiramosa.
Analytical studies of sponges have recently become a matter of interest and secondary target energy dispersive X-ray fluorescence spectrometry (EDXRF) analysis of marine sponges facilitates detection of many elements i.e., Al, Si, K, Ca, Ti, Mn, Fe, Ni, Cu, Zn, As, Br, Rb, Sr, Zr, I and Pb (Araújo et al., 2003). The differences in elemental accumulation between individual sponge species is well-strategised and variations probably reflect the differences in physiological conditions, in turn reflecting the passage of water through the aquiferous system of individual sponges (Patel et al., 1985). Depending on their structural uniqueness, different elements may be accumulated into the sponge tissues along with the ingested food particles, resulting in differential deposition on their surface, as observed in the present study of three Haliclona sps. The present SEM-EDXA analyses are restricted only to surface elemental composition but not
to the tissue accumulation. In our earlier studies, we have quantified more elements in the whole tissues of Petrosia (Xestospongia) testudinaria and Sigmadocia fibulata for heavy metal accumulation using ICP-MS and found differential elemental accumulation (Venkateswara Rao et al., 2006; 2007). In the present study, we found significant differences in surface elemental composition between species of same genus inhabiting similar habitat.
Thus sponges belonging to Porifera, though simplest in organisation, exhibit diversity in morphological patterns and chemical composition. These parameters are gaining significance in taxonomic identification of sponges for ecobiotechnological exploitation of potential sponges et (Hougaard et al., 1991; Vacelet et al., 2000). Genus, Haliclona is widely known for its diverse range of secondary metabolites with distinct mode of action. Manzamine A–D, isolated from Haliclona sp. exhibited prominent antitumour and cytotoxic properties (Sakai et al., 1986; Sakai et al., 1987). The partially purified toxin from the marine sponge Haliclona virides (HvTx) had depolarisation effect on skeletal muscle and nerve fibers (Sevcik et al., 1986). Halitoxin (HTX) from H. rubens, the red fire sponge, is a mixture of chemically defined heterocyclic molecules containing quaternary charged nitrogens as pyridinium moieties, analogous to curare alkaloids, exhibited cytotoxicity and neurotoxicity (Schmitz et al., 1978). Nitric oxide synthase (NOS) inhibitors, Xestospongin D and Araguspongin C, were also
Figure 7 The biochemical and other tissue components of three Haliclona species (mg g-1 wet weight). Statistical analysis was determined using One-way ANOVA (P< 0.05) and means were compared using Tukey’s HSD pair-wise comparison test. (a) Statistical significance as compared to H. retiderma. (b) Statistical significance as compared to H. tenuiramosa. ns, Not significantly different (P< 0.05).
CSB1100060.indd 21 2/6/2013 8:41:25 AM

22 Characterisation of three Haliclona species from the Gulf of Mannar
isolated and purified to 100% from sponge, Haliclona exigua, which is rarely available at GoM of India (Venkateswara Rao et al., 1998). A new cyclic metabolite, haliclamide, has been isolated from the Vanuatu marine sponge Haliclona sp. exhibited cytotoxicity against the NSCLC-N6 human bronchopulmonary non-small-cell-lung carcinoma cell lines (Randazzo et al., 2001).
As the demand for pharmacologically potent natural products is constantly increasing, various attempts for biotechnological production of sponge metabolites have been made. Sponges are known to possess strong regenerative capacities and small masses of sponge tissue are able to grow and regenerate into healthy sponges (Simpson, 1984). This potency has been used for cultivation of sponge tissue samples in both half-open systems and open sea aquaculture on a broad range of sponge species (Osinga et al., 1999). It is known that species belonging to the same genera may not produce same molecules and hence, the architecture of individual species may be an important aspect to pursue along with isolation of novel molecules. Hence, the species distribution patterns, morphological and anatomical variations within the species may influence the production of different kinds of therapeutic agents and as an alternative potential source of collagen (Gunda and Janapala, 2009; Luter and Duckworth, 2010; Pallela et al., 2011a). Our recent reports on H. tenuiramosa, Neopetrosia similis, Petrosia (Xestospongia) testudinaria and Petrosia nigricans also suggest the use of marine sponges as bioindicators and biomonitors for assessing pollution parameters and chemical speciation in any sustainable ecosystem (Rodrigo et al., 1994; Venkateswara Rao et al., 2009).
ACKNOWLEDGEMENTS
The authors are thankful to Department of Biotechnology (DBT), Government of India for financial assistance, and also to the Director, IICT for providing the facilities and his constant encouragement. Our special thanks to Dr P. A. Thomas from Vizhinjam Research Centre of Central Marine Fisheries Research Institute (CMFRI-ICAR), for the identification of sponges. The authors SK, RP and CRN thank Council for Scientific and Industrial Research (CSIR), for providing Senior Research Fellowship (CSIR-SRF). The authors SK and RP contributed equally to this paper.
REFERENCES
Araújo, M., Conceição, A., Barbosa, T., Lopes, M. and Humanes, M. (2003) X-Ray Spectrom., 32, 428-433.
Bell, J. and Barnes, D. (2000) J. Mar. Biol. Assoc. UK, 80, 707-718.Bell, J., Barnes, D. and Turner, J. (2002) Mar. Biol., 140, 75-81.Bradford, M. (1976) Anal. Biochem.,72, 248-254.Brooks, S. P. J., Ratnayake, W. M. N., Lampi, B. J. and Hollywood,
R. (1998) J. Agricult. Food Chem., 46, 4214-4217.
de Caralt, S., Sánchez-Fontenla, J., Uris, M. and Wijffels, R. (2010) Mar. Drugs, 8, 1731-1742.
Diaz, M. and Van Soest, R. (1994) The Plakinidae: a systematic review. In: Van Soest, R.W.M., Van Kempen, Th.M.G. and Braekman, J.-C. (eds), Sponges in time and space, pp. 93-110. Balkema Press, Rotterdam.
Gunda, V. and Janapala, V. (2009) Cell Tiss. Res., 337, 527-535.Hentschel, U., Usher, K. and Taylor, M. (2006) FEMS Microbiol.
Ecol., 55, 167-177.Hooper, J. and Van Soest, R. (2002) In: Hooper, J.N.A. and van Soest,
R.W.M. (eds), Systema Porifera: a guide to the classification of sponges, pp. 15-18. Kluwer Acadmeic/Plenum Publishers, New York.
Hougaard, L., Christophersen, C., Nielsen, P., Klitgaard, A. and Tendal, O. (1991) Biochem. Systemat. Ecol., 19, 223-235.
Jones, W. (1994) In: van Soest, R.W.M., van Kempen, T.G. and Braekman J.C. (eds), Sponges in time and space, pp. 241-250. Balkema Press, Rotterdam.
Kerr, R. and Lelly-Borges, M. (1994) In: van Soest, R.W.M. van Kempen T.M.G. and Braekman J.C. (eds), Sponges in time and space, pp. 65-74. Balkema Press, Rotterdam.
Koopmans, M., Martens, D. and Wijffels, R. (2009) Mar. Drugs, 7, 787-802.
Luter, H. and Duckworth, A. (2010) Biochem. Systemat. Ecol., 38, 146-153.
Millero, F. (1996) Chemical oceanography, p. 469. CRC Press, Boca Raton.
Osinga, R., Armstrong, E., Grant Burgess, J., Hoffmann, F., Reitner, J. and Schumann-Kindel, G. (2001) Hydrobiologia, 461, 55-62.
Osinga, R., Tramper, J. and Wijffels, R. (1999) Mar. Biotechnol., 1, 509-532.
Pallela, R., Bojja, S. and Janapala, V. R. (2011a) Int. J. Biol. Macromol., 49, 85-92.
Pallela, R., Koigoora, S., Gunda, V. G., Sunkara, M. S. and Janapala, V. R. (2011b) Chem. Spec. Bioavailab., 23, 16-23.
Palumbi, S. (1984) Science, 225, 1478-1480.Patel, B., Balani, M. C. and Patel, S. (1985) Sci. Total Environ., 41,
143-152.Randazzo, A., Debitus, C. and Gomez-Paloma, L. (2001)
Tetrahedron, 57, 4443-4446.Rodrigo, A., Bergquist, P., Bergquist, P. and Reeves, R. (1994)
In: Van Soest, R. W.M., Van Kempen, T.M.G. and Braekman, J.C. (eds), Sponges in time and space: biology, chemistry, palaeontology. Proceedings of the 4th International Porifera Conference, pp. 47-54. Balkema Press, Rotterdam.
Sakai, R., Higa, T., Jefford, C. and Bernardinelli, G. (1986) J. Am. Chem. Soc., 108, 6404-6405.
Sakai, R., Kohmoto, S., Higa, T. and Jefford, C. (1987) Tetrahedron Lett., 28, 5493-5496.
Schmitz, F., Hollenbeak, K. and Campbell, D. (1978) J. Org. Chem.,43, 3916-3922.
Sevcik, C., Alvarez-Vasquez, F., Saavedra, J. and Cordovez, G. (1986) Toxicon, 24, 851-860.
Simpson, T.L. (1984) The cell biology of sponges, pp. 1-662. Springer-Verlag, New York.
Sipkema, D., Heilig, H., Akkermans, A., Osinga, R., Onji, J. and Wijffels, R. (2003) Mar. Biotechnol., 5, 443-449.
Sipkema, D., Holmes, B., Nichols, S. and Blanch, H. (2009)
CSB1100060.indd 22 2/6/2013 8:41:25 AM

Srikanth Koigoora, Ramjee Pallela, Chakra Reddy Nelakurti, Madhavendra Sakunthala Sunkara and Venkateswara Rao Janapala 23
Microbial Ecol., 58, 903-920.Sipkema, D., Osinga, R., Schatton, W., Mendola, D., Tramper, J. and
Wijffels, R. (2005) Biotechnol. Bioeng., 90, 201-222.Taylor, K. (1995) Appl. Biochem. Biotechnol. 53, 207-214.Thomas, P. (1998) Porifera, faunal diversity in India, pp. 28-36.
ENVIS Central Zoological Survey of India, Calcutta.Usher, K., Sutton, D., Toze, S., Kuo, J. and Fromont, J. (2005) Mar.
Freshwat. Res., 56, 125-132.Vacelet, J., Borchiellini, C., Perez, T., Bultel-Ponce, V., Brouard, J.
and Guyot, M. (2000) Zoosystema, 22, 313-326.Venkateswara Rao, J., Desaiah, D., Vig, P. and Venkateswarlu, Y.
(1998) Toxicology, 129, 103-112.Venkateswara Rao, J.V., Kavitha, P., Reddy, N. and Rao, T. (2006)
Chemosphere, 65, 634-638.Venkateswara Rao, J.V., Kavitha, P., Srikanth, K., Usman, P.K. and
Rao, T.G. (2007) Toxicol. Environ. Chem., 89, 487-498.
Venkateswara Rao, J., Srikanth, K., Pallela, R. and Gnaneshwar
Rao, T. (2009) Environ. Monit. Assess., 156, 451-459.
Venkateswarlu, Y., Reddy, M. V. R. and Rao, J. V. (1994) J. Nat.
Prod. 57, 1283-1285.
Wang, G. (2006) J. Industr. Microbiol. Biotechnol., 33, 545-551.
Wang, X. and Lavrov, D. (2007) Mol. Biol. Evolut., 24, 363-373.
Wilkinson, C. (1992) In: Reisser, W. (ed.), Algae and symbioses:
plants, animals, fungi, viruses, interactions explored, pp. 111-
115. Biopress Ltd, Bristol.
Ya-sheng, W. (1995) Acta Palaeontol. Sinica, 34, 381-392.
Yi, Q., Wei, Z., Hua, L., Xingju, Y. and Meifang, J. (2005) Chin. J.
Oceanol. Limnol., 23, 194-198.
Zhou, G. and Molinski, T. (2003) Mar. Drugs, 1, 46-53.
CSB1100060.indd 23 2/6/2013 8:41:25 AM