antimicrobial potential of metabolites extracted from bacterial symbionts associated with marine...
TRANSCRIPT

ORIGINAL ARTICLE
Antimicrobial potential of metabolites extracted frombacterial symbionts associated with marine sponges incoastal area of Gulf of Mannar Biosphere, IndiaS. Skariyachan, A. G. Rao, M.R. Patil, B. Saikia, V. Bharadwaj KN and J. Rao GS
Department of Biotechnology, R & D Centre, Dayananda Sagar College of Engineering, Bangalore, Karnataka, India
Significance and Impact of the Study: This is the first study demonstrating antimicrobial potential offlurophoric and chromophoric metabolites extracted from bacterial biosymbionts associated with mar-ine sponges. Our study has significant scope as Indian coastal area especially harbours vast varieties ofsponges with novel secondary metabolites-producing organisms. The natural metabolites extracted fromsponge-derived bacteria pave novel therapeutic remedy against various pathogens when most of themare emerged as extreme drug resistant superbugs.
Keywords
chromophoric, endosymbiotic bacteria,
fluorophoric, Gulf of Mannar, marine
sponges, secondary metabolites, therapeutic
potential.
Correspondence
Sinosh Skariyachan, Department of Biotech-
nology, R & D Centre, Dayananda Sagar
College of Engineering, Bangalore 560 078
Karnataka, India.
E-mail: [email protected]
2013/1529: received 30 July 2013, revised 14
October 2013 and accepted 15 October 2013
doi:10.1111/lam.12178
Abstract
Marine coastal areas of India have vast diversity of sponges which harbours
many endosymbiotic bacteria which are the source of many potential
antimicrobial metabolites. This study focuses the screening and
characterization of drug-producing bacteria symbiotically which are associated
with marine sponges collected from Gulf of Mannar, South Coast India. Six
different sponges were collected and they were identified on the basis of their
morphology. The drug-producing isolates were screened by agar overlay
method towards various clinical strains. The secondary metabolites were
characterized and were found to be quinones, alkaloids, flavanoids and flavonyl
glycosides. The metabolites showed significant inhibitory properties against
clinical strains that were further identified as chromophoric and fluorophoric
in nature. Ethyl acetate extracts of chromophore and floureophore substances
showed significant inhibitory properties against Methicillin resistant
Staphylococcus aureus (MRSA) and Salmonella typhi respectively. 16S rRNA
gene sequencing of theses isolates revealed that chomophore-producing strain
were closely related to Pseudomonas spp. RHLB12, isolated from Callyspongia
spp. and floureophore-producing bacteria was related to Bacillus licheniformis
T6-1 which was isolated from Haliclona spp. Hence, our study demonstrated
that antimicrobial metabolites extracted from symbiotic bacteria associated
with marine sponges have high therapeutic potential against many bacterial
pathogens including multidrug-resistant strains.
Introduction
Marine sponges are one of the richest sources of biologi-
cally active secondary metabolites with vast chemical
diversity. Various metabolites obtained from such sponges
have yielded pharmaceutical products such as novel anti-
inflammatory agents, anticancer agents and antibiotics
(Boopathy and Kathiresan 2010). The marine coastal
areas in India have vast diversity of marine sponges in
which Gulf of Mannar region is a bed for valuable
sponges. The Gulf of Mannar is located in the state of
Tamil Nadu on the South-eastern tip of India. Around
3600 species of marine fauna and flora have been identi-
fied in the Gulf of Mannar area consisting of 108 species
of sponges (Mol et al. 2010). Until now, more than 5000
different compounds have been isolated and identified
Letters in Applied Microbiology 58, 231--241 © 2013 The Society for Applied Microbiology 231
Letters in Applied Microbiology ISSN 0266-8254

from sponges with nearly 800 of them exhibiting antimi-
crobial properties. These compounds belonged to differ-
ent class of compounds such as terpenoids, alkaloids,
macrolides, polyethers, nucleoside derivatives and pep-
tides (Ravichandran et al. 2011; Dhinakaran et al. 2012).
Marine sponges harbour micro-organisms on their sur-
faces, in their canal systems and intercellular spaces, and
these contribute up to 40% of the total cellular content of
a sponge. Most of these micro-organisms are endosymbi-
otic bacteria and are probably major source of novel
secondary metabolites and other valuable compounds
(Petchi et al. 2013). These symbiotic bacteria may play
roles in digestion, waste removal and help in the nutri-
tional process, either by intracellular digestion or by
translocation of metabolites including photosynthesis,
nitrification and nitrogen fixation. They also stabilize the
sponge skeleton and participate in the host’s chemical
defence. This has greatly improved our knowledge of
sponge–microbe interactions. Symbiotic bacteria associ-
ated with sponges produce potential antimicrobial
compounds and bioactive compounds. Many bioactive
compounds extracted from Dendrilla nigra (Ivanova et al.
1999), Axinella donnani and Clathria gorgonoides
(Aishwarya et al. 2013) have significant antimicrobial
activities against various Gram-positive pathogenic bacte-
ria. Similarly, dichloromethane-methanol (1 : 1) extract
of the sponge Phycopsis sp. collected from Tuticorin coast
of India exhibited antibacterial activity (Venkateswarlu
and Biabani 1995). Many bacterial metabolites have been
isolated from the marine sponges, Cryptophycin I and
Chondramide isolated from Dysidea spp. and Jaspis spp.
respectively (Sabdono and Radjasa 2008). In addition to
therapeutic applications of bioactive compounds from
marine sponges against various bacteria, there are many
reports which reveal the antitumor drugs such as Kendar-
imide A, isolated from sponge Haliclona sp. has been
shown to reverse multidrug resistance in tumour cells
(Roser et al. 2005).
Recent studies revealed that endosymbionts associated
with sponge Amphimedon ochracea produced cytotoxic
compounds against HepG2 (hepatocellular carcinoma),
HCT (colon carcinoma) and MCF-7 (breast carcinoma)
cancer cell lines (Aboul-Ela et al. 2012), Proteobacteria
and novel Pseudomonas sp. associated with the sponge
Suberites domuncula produced cytotoxic compounds
against HeLa and PC12 cells (Thakur et al. 2005), Bacillus
megaterium and Pseudoalteromonas aurantia isolated from
Eupexaura curvata, Pseudoalteromonas piscicida isolated
from Hymeniacidon perleve and Psedoalteromonas rubra
isolated from sea water produce bioactive compounds
with wide range of antimicrobial spectrum (Zheng et al.
2005a,b). Similarly, Bacillus atrophaeus was isolated from
the marine sponge Dysidea avara in the South China Sea,
which can produce Bacillamide C and a new compound
of Neobacillamide A (Liu et al. 2012).
The use of molecular approaches for describing the
microbial diversity has greatly enhanced the knowledge of
these natural microbial communities associated with mar-
ine sponges. Cloning and sequencing of 16S rRNA genes
gives data which can be used to describe complete micro-
bial community, composition and phylogenetic relatives,
and they can indicate possible nutritional requirements
and physiological riches of many endosymbiotic microbes
(Webster et al. 2001) present in various sponges. This
study mainly focuses on the screening and characteriza-
tion of drug-producing bacteria that are symbiotically
associated with marine sponges found in South Coast of
India, Gulf of Mannar. The identification and character-
ization of secondary metabolites or similar bioactive com-
pounds isolated from symbiotically associated bacteria is
also studied. Further, antimicrobial properties of metabo-
lites isolated from endosymbiotic bacteria from sponges
against various bacterial pathogens, especially multidrug-
resistant strains have been performed in this study.
Results and discussion
Six different types of marine sponges were collected from
Olaikuda in Gulf of Mannar Bioshphere region, Ramesh-
warm, Tamil Nadu, India. Based on morphological fea-
tures of spicules and other specialized structures, the
collected marine sponges were identified as Cacaspongia
spp., Sigmadocia spp., Callyspongia spp., Dendrila spp.,
Gorgonid spp. and Haliclona spp. There are many reports
which reveal that these kinds of marine sponges are very
common in Indian coastal areas including Tuticorin coast
(Singla et al. 2013), Kavaratti Island, Lakshadweep archi-
pelago (Gopi et al. 2012) and Mandapam bay and Gulf of
Mannar (Velho-Pereira and Furtado 2012). Other reports
reveal that these marine sponges harbour various types of
symbiotically associated micro-organisms they are unique
sources of natural bioactive secondary metabolites with
pronounced pharmacological activities (Thomas et al.
2010; Hardoim et al. 2012; Velho-Pereira and Furtado
2012). For high microbial abundance (HMA) sponges,
around 38% of sponge wet weight is composed of bacte-
rial cells, and that such bacterial abundance surpasses that
of the seawater by 2–4 orders of magnitude (Taylor et al.
2007).
The bacteria which are symbiotically associated with the
collected sponge samples were isolated by Marine Zobell
agar. The cultural characteristics of the isolates were found
to be pale yellow coloured, medium sized, opaque and con-
vex colonies. The viable bacterial count in terms of colony
forming units (CFU g�1) ranged from 1�10 9 103 to
6�4 9 103, with an average of 3�85 9 103 CFU g�1. After
Letters in Applied Microbiology 58, 231--241 © 2013 The Society for Applied Microbiology232
Antimicrobial potential of metabolites extracted S. Skariyachan et al.

analysing the bacterial count from the sponge samples,
Sigmodocia spp. was found to have the highest CFU g�1
(Table 1).
The bacteria isolated from the sponges were character-
ized by standard microbiology techniques. Most of the
bacteria isolated from the sponges were found to be
Gram-positive bacilli and cocci. Few bacteria were found
to be Gram-negative rods (Fig. S1). Various studies on
antibiotic production in marine environment have
reported that around 36% of the isolates are Gram-nega-
tive rods (Beveridge 2001). To determine the characteris-
tics of the isolated bacteria, the sponge homogenate was
plated on differential and selective media. When plated
on MacConkey’s agar, the isolates from tissues were
found to be Gram-negative bacteria. MacConkey’s agar
indicates the presence of Gram-negative coliforms in the
sponges which are probably due to the presence of faecal
products from marine environments (Boopathy and
Kathiresan 2010). When the sponge homogenate was pla-
ted on Thiosulfate citrate bile salts sucrose (TCBS) agar,
it showed small, circular, golden yellow coloured, opaque
colonies indicating the presence of Vibrio spp. All sponges
showed the presence of Vibrio spp. Marine sediments and
sponges often harbours Vibrio spp. and optimum growth
parameter for Vibrio spp. is high salinity (West and
Colwell 1984). Studies have reported that Vibrio spp. are
predominant bacterial symbionts associated with marine
environment (Anand et al. 2006). When the sponge
homogenates were plated on Bromo thymol blue (BTB)
Lactose agar, the plates showed deep yellow coloured,
medium sized, circular colonies indicating the presence of
Staphylococcus spp. Such kinds of bacterial isolates were
observed from the tissue homogenates of Gorgonid spp.,
Haliclona spp., Cacaspongia spp., Callyspongia spp., Sigm-
odocia spp. and Dendrila spp. Similarly, circular, med-
ium-sized, good luxuriant, blue–green coloured colonies
were observed when the sponge homogenates were plated
on milk agar with cetrimide, which are the characteristic
features of Pseudomonas spp. Such types of colonies were
isolated from the marine sponge Callyspongia spp. Other
reports reveal that many species of Pseudomonas found as
biosymbiont with marine sponges and they are the poten-
tial sources of antimicrobial agents against drug resistant
pathogens (Marinho et al. 2009; Thomas et al. 2010).
Various types of symbiotically associated bacteria isolated
from the collected sponges are shown in Table 1. Maxi-
mum types of endosymbiotic bacteria were isolated from
the sponge Callyspongia spp. (Table 1).
The antimicrobial properties of marine isolates were
tested against MRSA, Proteus mirabilis, Klebsiella pneumo-
niae and Salmonella typhi by agar overlay method. A clear
zone of inhibition was observed on the soft agar mixed
with two test organisms (Fig. 1) indicating that marine
isolates have significant antimicrobial activities, which are
probably due to the production of bioactive secondary
metabolites. The drug-producing bacteria identified by
preliminary screening were further characterized by bio-
chemical tests. The microbiology characterizations of the
bacterial isolates from the marine sponges are shown in
Table 2. From our results, it is evident that the bioactive
metabolites produced by biosymbionts were extracellular
in nature. Most of the bacteria showed zones of inhibi-
tion against the two test organisms were isolated from the
sponges; Callyspongia spp. and Haliclona spp. Out of all,
the symbiotic isolates from Callyspongia spp. have signifi-
cant antimicrobial properties against MRSA. Similarly, the
bacteria isolated from Haliclona spp. showed significant
antimicrobial properties against clinical strain of Salm.
typhi. Studies have revealed that members of the Cally-
spongia spp. and Haliclona spp. are potential drug-bearers
and they have potential antimicrobial activities against
various pathogens (Ely et al. 2004; Aishwarya et al. 2013).
There are other reports where sponge-derived isolate of
Vibrio spp. have shown to be rich source of novel biologi-
cally active bacterial metabolites (Anand et al. 2006).
Similar studies demonstrated that marine derived Pseudo-
monas and Bacillus are sources of potential antimicrobial
substances (Anand et al. 2006; Devi et al. 2010).
The secondary metabolite-producing isolates were cul-
tured in Marine Zobel broth and minimum essential
media for extraction and purification of antimicrobial
compounds. The supernatant from appropriately incu-
bated culture was used for purification. The metabolite
from the supernatant was further purified by solvent
extraction method. The purified extract showed inhibitory
properties, zone of inhibition in the agar medium (well
Table 1 Total bacterial count (CFU g�1) and major types of symbiotic
bacteria isolated from selected marine sponges
Sponge
sample
Number of
bacteria
(CFU g�1) Major types of bacteria isolated
Dendrilla
spp.
5�7 9 103 Staphylococcus spp., Vibrio spp.
Cacaspongia
spp.
5 9 103 Staphylococcus spp., Bacillus spp.,
Vibrio spp.
Callyspongia
spp.
3�2 9 103 Staphylococcus spp., Pseudomonas spp.,
Klebsiella spp., Bacillus spp., Vibrio
spp.
Haliclona
spp.
9�1 9 103 Staphylococcus spp., Bacillus spp.,
Vibrio spp.
Sigmodocia
spp.
6�4 9 103 Staphylococcus spp., Bacillus spp.,
Vibrio spp.
Gorgonoid
spp.
1�8 9 103 Staphylococcus spp., Bacillus spp.,
Vibrio spp.
CFU, colony forming unit.
Letters in Applied Microbiology 58, 231--241 © 2013 The Society for Applied Microbiology 233
S. Skariyachan et al. Antimicrobial potential of metabolites extracted
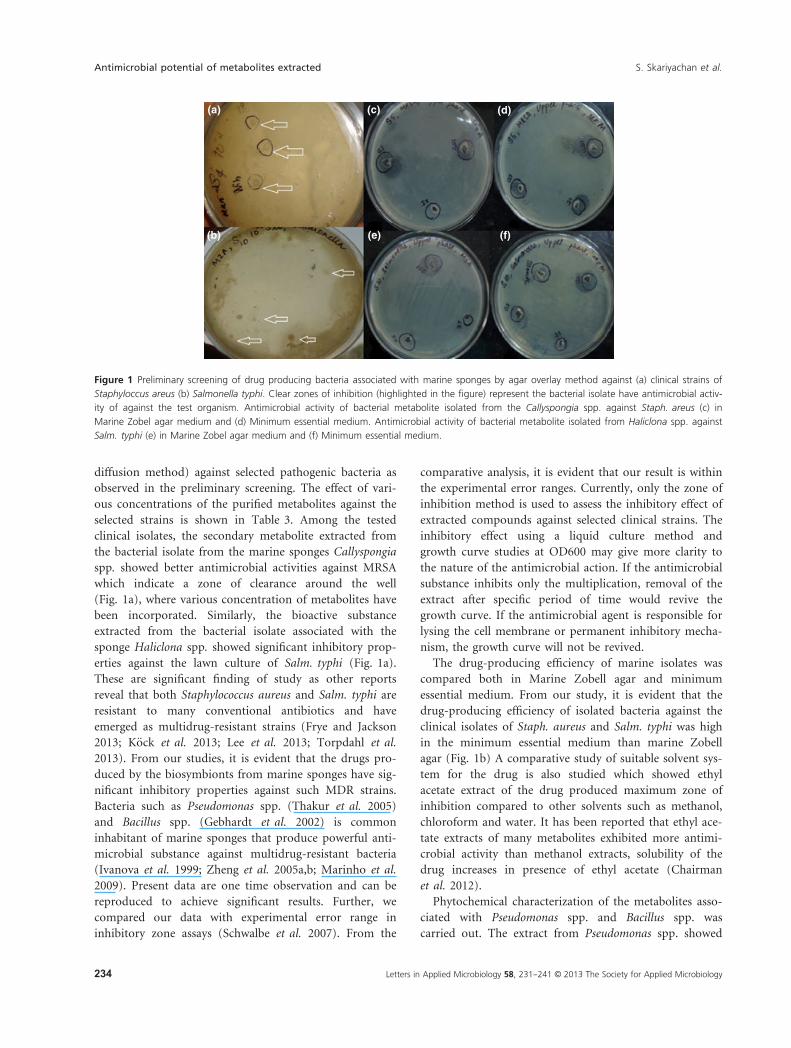
diffusion method) against selected pathogenic bacteria as
observed in the preliminary screening. The effect of vari-
ous concentrations of the purified metabolites against the
selected strains is shown in Table 3. Among the tested
clinical isolates, the secondary metabolite extracted from
the bacterial isolate from the marine sponges Callyspongia
spp. showed better antimicrobial activities against MRSA
which indicate a zone of clearance around the well
(Fig. 1a), where various concentration of metabolites have
been incorporated. Similarly, the bioactive substance
extracted from the bacterial isolate associated with the
sponge Haliclona spp. showed significant inhibitory prop-
erties against the lawn culture of Salm. typhi (Fig. 1a).
These are significant finding of study as other reports
reveal that both Staphylococcus aureus and Salm. typhi are
resistant to many conventional antibiotics and have
emerged as multidrug-resistant strains (Frye and Jackson
2013; K€ock et al. 2013; Lee et al. 2013; Torpdahl et al.
2013). From our studies, it is evident that the drugs pro-
duced by the biosymbionts from marine sponges have sig-
nificant inhibitory properties against such MDR strains.
Bacteria such as Pseudomonas spp. (Thakur et al. 2005)
and Bacillus spp. (Gebhardt et al. 2002) is common
inhabitant of marine sponges that produce powerful anti-
microbial substance against multidrug-resistant bacteria
(Ivanova et al. 1999; Zheng et al. 2005a,b; Marinho et al.
2009). Present data are one time observation and can be
reproduced to achieve significant results. Further, we
compared our data with experimental error range in
inhibitory zone assays (Schwalbe et al. 2007). From the
comparative analysis, it is evident that our result is within
the experimental error ranges. Currently, only the zone of
inhibition method is used to assess the inhibitory effect of
extracted compounds against selected clinical strains. The
inhibitory effect using a liquid culture method and
growth curve studies at OD600 may give more clarity to
the nature of the antimicrobial action. If the antimicrobial
substance inhibits only the multiplication, removal of the
extract after specific period of time would revive the
growth curve. If the antimicrobial agent is responsible for
lysing the cell membrane or permanent inhibitory mecha-
nism, the growth curve will not be revived.
The drug-producing efficiency of marine isolates was
compared both in Marine Zobell agar and minimum
essential medium. From our study, it is evident that the
drug-producing efficiency of isolated bacteria against the
clinical isolates of Staph. aureus and Salm. typhi was high
in the minimum essential medium than marine Zobell
agar (Fig. 1b) A comparative study of suitable solvent sys-
tem for the drug is also studied which showed ethyl
acetate extract of the drug produced maximum zone of
inhibition compared to other solvents such as methanol,
chloroform and water. It has been reported that ethyl ace-
tate extracts of many metabolites exhibited more antimi-
crobial activity than methanol extracts, solubility of the
drug increases in presence of ethyl acetate (Chairman
et al. 2012).
Phytochemical characterization of the metabolites asso-
ciated with Pseudomonas spp. and Bacillus spp. was
carried out. The extract from Pseudomonas spp. showed
(a) (c) (d)
(b) (f)(e)
Figure 1 Preliminary screening of drug producing bacteria associated with marine sponges by agar overlay method against (a) clinical strains of
Staphyloccus areus (b) Salmonella typhi. Clear zones of inhibition (highlighted in the figure) represent the bacterial isolate have antimicrobial activ-
ity of against the test organism. Antimicrobial activity of bacterial metabolite isolated from the Callyspongia spp. against Staph. areus (c) in
Marine Zobel agar medium and (d) Minimum essential medium. Antimicrobial activity of bacterial metabolite isolated from Haliclona spp. against
Salm. typhi (e) in Marine Zobel agar medium and (f) Minimum essential medium.
Letters in Applied Microbiology 58, 231--241 © 2013 The Society for Applied Microbiology234
Antimicrobial potential of metabolites extracted S. Skariyachan et al.
the presence of alkaloids, quinones, flavonyl glycosides
and flavanoids in marine Zobell broth. The extract from
Bacillus spp. showed the presence of carbohydrates,
proteins, lipids, alkaloids, quinones and flavonyl glyco-
sides. Bioactive compounds such as Micacocidin A, B, C
(antimycoplasmal) and C-I4, a cyclic dipeptide (chitinase
inhibitor) have been reported from marine Pseudomonas
spp. (Kobayashi et al. 1998). Many antibiotics including
cyclic peptides, cyclic lipopepides and novel thiopeptides
have been reported from marine Bacillus spp. (Nagai
et al. 2003).
The ethyl acetate extracts was spotted on the thin layer
chromatography (TLC) plates. On separation, a purple-
coloured compound was observed on the plate which
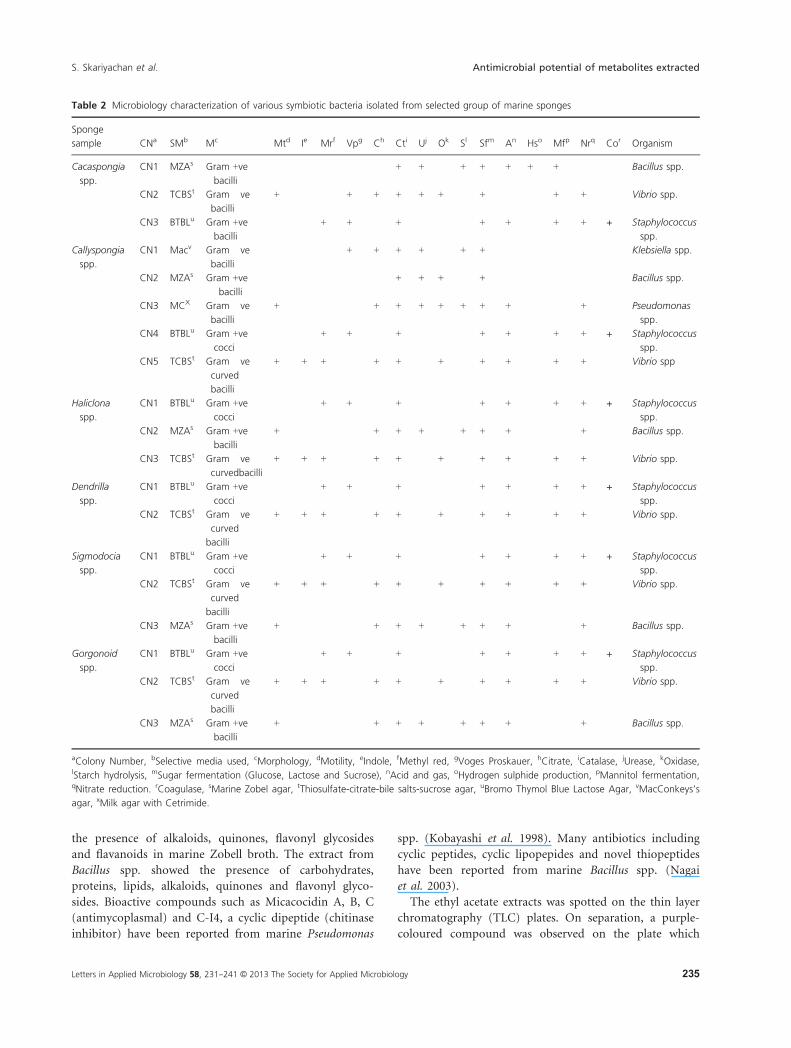
Table 2 Microbiology characterization of various symbiotic bacteria isolated from selected group of marine sponges
Sponge
sample CNa SMb Mc Mtd Ie Mrf Vpg Ch Cti Uj Ok Sl Sfm An Hso Mfp Nrq Cor Organism
Cacaspongia
spp.
CN1 MZAs Gram +ve
bacilli
� � � � � + + � + + + + + � � Bacillus spp.
CN2 TCBSt Gram �ve
bacilli
+ � � + + + + + � + � � + + � Vibrio spp.
CN3 BTBLu Gram +ve
bacilli
� � + + � + � � � + + � + + + Staphylococcus
spp.
Callyspongia
spp.
CN1 Macv Gram �ve
bacilli
� � � + + + + � + + � � � � � Klebsiella spp.
CN2 MZAs Gram +ve
bacilli
� � � � � + + + � + � � � � � Bacillus spp.
CN3 MCX Gram �ve
bacilli
+ � � � + + + + + + + � � + � Pseudomonas
spp.
CN4 BTBLu Gram +ve
cocci
� � + + � + � � � + + � + + + Staphylococcus
spp.
CN5 TCBSt Gram �ve
curved
bacilli
+ + + � + + � + � + + � + + � Vibrio spp
Haliclona
spp.
CN1 BTBLu Gram +ve
cocci
� � + + � + � � � + + � + + + Staphylococcus
spp.
CN2 MZAs Gram +ve
bacilli
+ � � � + + + � + + + � � + � Bacillus spp.
CN3 TCBSt Gram �ve
curvedbacilli
+ + + � + + � + � + + � + + � Vibrio spp.
Dendrilla
spp.
CN1 BTBLu Gram +ve
cocci
� � + + � + � � � + + � + + + Staphylococcus
spp.
CN2 TCBSt Gram �ve
curved
bacilli
+ + + � + + � + � + + � + + � Vibrio spp.
Sigmodocia
spp.
CN1 BTBLu Gram +ve
cocci
� � + + � + � � � + + � + + + Staphylococcus
spp.
CN2 TCBSt Gram �ve
curved
bacilli
+ + + � + + � + � + + � + + � Vibrio spp.
CN3 MZAs Gram +ve
bacilli
+ � � � + + + � + + + � � + � Bacillus spp.
Gorgonoid
spp.
CN1 BTBLu Gram +ve
cocci
� � + + � + � � � + + � + + + Staphylococcus
spp.
CN2 TCBSt Gram �ve
curved
bacilli
+ + + � + + � + � + + � + + � Vibrio spp.
CN3 MZAs Gram +ve
bacilli
+ � � � + + + � + + + � � + � Bacillus spp.
aColony Number, bSelective media used, cMorphology, dMotility, eIndole, fMethyl red, gVoges Proskauer, hCitrate, iCatalase, jUrease, kOxidase,lStarch hydrolysis, mSugar fermentation (Glucose, Lactose and Sucrose), nAcid and gas, oHydrogen sulphide production, pMannitol fermentation,qNitrate reduction. rCoagulase, sMarine Zobel agar, tThiosulfate-citrate-bile salts-sucrose agar, uBromo Thymol Blue Lactose Agar, vMacConkeys’s
agar, xMilk agar with Cetrimide.
Letters in Applied Microbiology 58, 231--241 © 2013 The Society for Applied Microbiology 235
S. Skariyachan et al. Antimicrobial potential of metabolites extracted
indicates the presence of a chromophore and the remain-
ing compounds of interest were colourless. The colourless
fractions are further visualized under UV lamp which
indicate that the extract also contains few fluorophoric
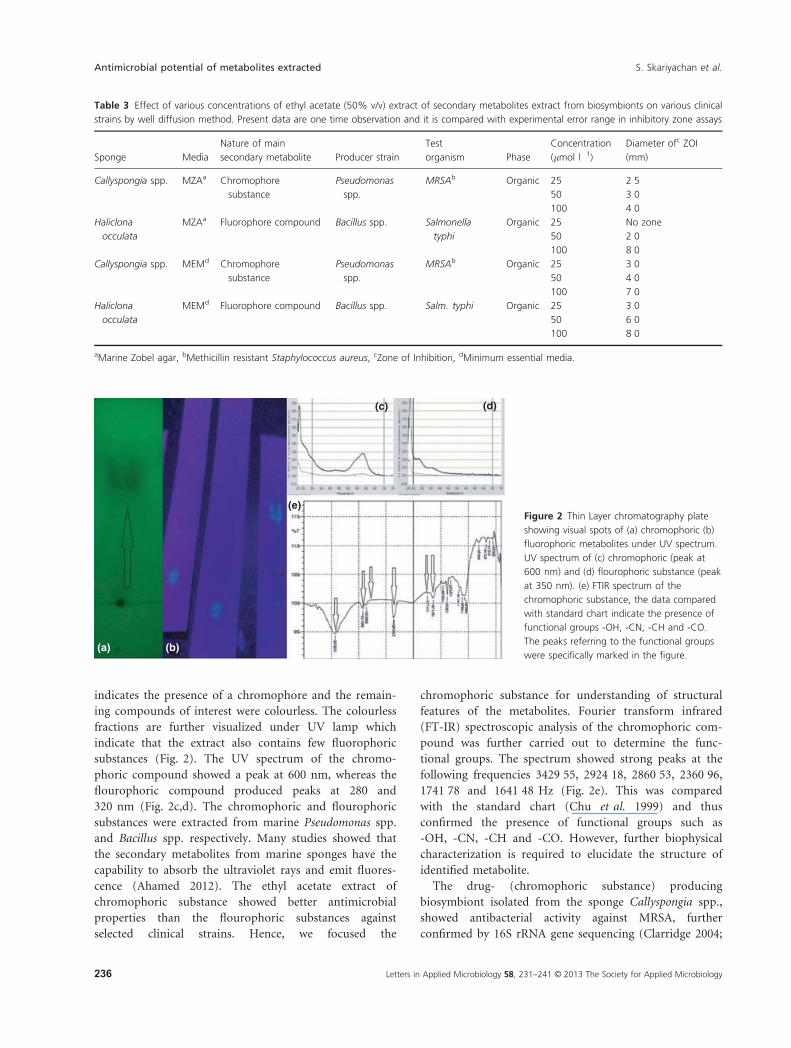
substances (Fig. 2). The UV spectrum of the chromo-
phoric compound showed a peak at 600 nm, whereas the
flourophoric compound produced peaks at 280 and
320 nm (Fig. 2c,d). The chromophoric and flourophoric
substances were extracted from marine Pseudomonas spp.
and Bacillus spp. respectively. Many studies showed that
the secondary metabolites from marine sponges have the
capability to absorb the ultraviolet rays and emit fluores-
cence (Ahamed 2012). The ethyl acetate extract of
chromophoric substance showed better antimicrobial
properties than the flourophoric substances against
selected clinical strains. Hence, we focused the
chromophoric substance for understanding of structural
features of the metabolites. Fourier transform infrared
(FT-IR) spectroscopic analysis of the chromophoric com-
pound was further carried out to determine the func-
tional groups. The spectrum showed strong peaks at the
following frequencies 3429�55, 2924�18, 2860�53, 2360�96,1741�78 and 1641�48 Hz (Fig. 2e). This was compared
with the standard chart (Chu et al. 1999) and thus
confirmed the presence of functional groups such as
-OH, -CN, -CH and -CO. However, further biophysical
characterization is required to elucidate the structure of
identified metabolite.
The drug- (chromophoric substance) producing
biosymbiont isolated from the sponge Callyspongia spp.,
showed antibacterial activity against MRSA, further
confirmed by 16S rRNA gene sequencing (Clarridge 2004;
Table 3 Effect of various concentrations of ethyl acetate (50% v/v) extract of secondary metabolites extract from biosymbionts on various clinical
strains by well diffusion method. Present data are one time observation and it is compared with experimental error range in inhibitory zone assays
Sponge Media
Nature of main
secondary metabolite Producer strain
Test
organism Phase
Concentration
(lmol l�1)
Diameter ofc ZOI
(mm)
Callyspongia spp. MZAa Chromophore
substance
Pseudomonas
spp.
MRSAb Organic 25 2�550 3�0100 4�0
Haliclona
occulata
MZAa Fluorophore compound Bacillus spp. Salmonella
typhi
Organic 25 No zone
50 2�0100 8�0
Callyspongia spp. MEMd Chromophore
substance
Pseudomonas
spp.
MRSAb Organic 25 3�050 4�0100 7�0
Haliclona
occulata
MEMd Fluorophore compound Bacillus spp. Salm. typhi Organic 25 3�050 6�0100 8�0
aMarine Zobel agar, bMethicillin resistant Staphylococcus aureus, cZone of Inhibition, dMinimum essential media.
(a) (b)
(c) (d)
(e)Figure 2 Thin Layer chromatography plate
showing visual spots of (a) chromophoric (b)
fluorophoric metabolites under UV spectrum.
UV spectrum of (c) chromophoric (peak at
600 nm) and (d) flourophoric substance (peak
at 350 nm). (e) FTIR spectrum of the
chromophoric substance, the data compared
with standard chart indicate the presence of
functional groups -OH, -CN, -CH and -CO.
The peaks referring to the functional groups
were specifically marked in the figure.
Letters in Applied Microbiology 58, 231--241 © 2013 The Society for Applied Microbiology236
Antimicrobial potential of metabolites extracted S. Skariyachan et al.

Janda and Abbott 2007). The sequence obtained was
compared with known sequences in the GenBank data-
base, identified by BLAST search; indicate closest phyloge-
netic relationship with 99% sequence identity (Hentschel
et al. 2001) towards Pseudomonas spp. RHLB12. Similarly,
the drug- (flourophoric substance) producing biosymbi-
ont isolated from the sponge Haliclona spp., showed anti-
bacterial activity against clinical strain of Salm. typhi, was
also characterized by 16S rRNA gene sequencing. From
the BLAST result we noticed that the sequence showed 96%
sequence identity with Bacillus licheniformis T6-1. The
16S rRNA sequences of Pseudomonas spp. RHLB12 were
deposited to GenBank database and can be accessed by
the accession number KF225558.1.
Materials and methods
Description of the sampling spot
The area under study was a beach by name Olaikuda in
Gulf of Mannar, Marine National Park, Rameshwaram,
Tamil Nadu, India. Gulf of Mannar is a Bio-sphere
Reserve which is developed for preserving aquatic life. It
comprises 21 islands running almost parallel to the coast-
line between latitude 8°47′ N and 9°15′N and longitude
78°12′E and 79°14′E with three distinct marine ecosys-
tems namely corals, sea grass and mangroves.
Collection, transportation and storage of sponge samples
The sponge samples were collected from bottom set crab
nets, for the harvest of edible crabs at a depth of 7 feet.
The net were deployed in the sea at 4 PM and was
retrieved at 6 AM the following morning on January 12,
2013. The sponge samples were carefully removed from
the wires of the crab net and washed with sea water to
remove the sand and adhered debris. The samples were
kept in different zip lock bags in an ice box. The samples
were transported to the laboratory with seawater under
required aseptic precautions (Dhinakaran et al. 2012).
The samples were aseptically stored at �4°C for 12 h. All
the samples were processed on the very next day and the
duration of this study was 6 months.
Isolation of symbiotic bacteria associated with marine
sponges
The sponges were washed thoroughly with sterile sea water
and homogenized in sponge dissociation medium (Anand
et al. 2006). The homogenate was serially diluted and pla-
ted on marine Zobell agar 2216 (Anand et al. 2006) by
pour plate technique (Geldreich et al. 1972). All the plates
were incubated at 37°C for 24 h. Subsequently, the
symbiotic bacteria that are associated the sponges were
selectively isolated using TCBS agar, Milk agar with cetri-
mide, BTB lactose agar and MacConkey’s agar (Hi-media,
Mumbai, India). The total number of endosymbiotic bac-
teria (CFU g�1) associated with each sponge sample was
determined by digital colony counter (Labtronics, India).
Preliminary screening of drug-producing bacteria
Preliminary screenings of drug-producing bacterial iso-
lates were carried out by agar overlay method (Anand
et al. 2006). The test strains were gently overlaid using
3�5 mol l�1 soft agar over the marine isolates. The clinical
isolate of MRSA, Pr. mirabilis, Kl. pneumoniae and
Salm. typhi were used as the test organisms. The test
organisms were collected from Sagar Hospitals, Bangalore,
India. The soft agar was prepared by inoculating 1 mL
fresh cultures of test strain in 100 mol l�1 of soft agar
and mixed thoroughly. For marine strains, 1�5% NaCl
was added to the soft agar. The overlaid plates were incu-
bated at 37°C for 48 h. The zones of inhibition produced
by the marine isolates against the test organisms were
thereafter interpreted.
Secondary screening of drug-producing bacteria
Marine Zobell broth 2216 (Anand et al. 2006) and minimal
essential media (30 g l�1 sucrose, vitamin tablet and sea
water) were used for the extraction of drugs from marine
isolates. Ten ml culture of marine isolate was inoculated to
100 ml of marine Zobell broth and minimal essential media
and kept in a shaker incubator (Eppendorf) at 35°C for
7 days. The culture was centrifuged at 10 000 g for 20 min
and the supernatant was separated and stored at �4°C. Thesupernatant was further screened for antimicrobial activity
against the test organisms by well diffusion method at con-
centration ranges of 25–100 lmol l�1. The plates were
incubated at 37°C for 48 h and screened for antimicrobial
activities, presence of zone of inhibition.
Phytochemical analysis of secondary metabolites
produced by marine isolates
The supernatant from the marine isolates which showed
significant antimicrobial activities against the test organ-
ism were selected and transferred to separating funnels.
An equal mixture of supernatant and ethyl acetate, suit-
able solvent was poured to a separating funnel and the
separation process was carried out at room temperature
(Gokulkrishnan et al. 2011). The organic phase, contains
the antimicrobial metabolite, was collected and stored at
�4°C for further studies. The analysis for major classes of
antimicrobial substance present in the collected phase was
Letters in Applied Microbiology 58, 231--241 © 2013 The Society for Applied Microbiology 237
S. Skariyachan et al. Antimicrobial potential of metabolites extracted

carried out by standard phytochemical analysis. Meyer’s
and Wagner’s test (Miller et al. 2010) were used for the
detection of alkaloids; Molisch’s test (Usman et al. 2009)
was used for the detection of carbohydrate; Libermann
Burchard test (Aryantha et al. 2002) was used for the
detection of steroids; sulphuric acid test (Firdouse and
Alam 2011) was used for the detection of quinines; ferric
chloride test (Firdouse and Alam 2011) was used for the
detection of phenols; biuret, ninhydrin and Bradford
assays (Singh et al. 2012) were used for the detection of
proteins. Similarly, standard phytochemical screening was
performed for the detection of flavonyl glycosides, lipids
and flavanoids (Singh et al. 2012).
Purification and characterization of bioactive metabolites
Purification of the secondary metabolites, probable drugs,
was carried out by TLC. The organic extract was spotted
on silica gel plates (Merck Millipore, Mumbai, India)
using 30 : 70 ethyl acetate-hexane as solvent system. A
one-dimensional ascending technique was used to sepa-
rate the components present in the extract (Chairman
et al. 2012). The plates visualized under UV radiation by
short, middle and long wavelengths of 254 nm, 302 nm
and 600 nm respectively. The separated compounds were
eluted and subjected to Ultra Violet-Visible (UV-Vis)
spectral analysis using methanol as standard. A FT-IR
spectroscopic analysis was carried out to determine the
functional groups of bioactive compounds.
Antimicrobial testing of the identified compounds
An antimicrobial assay was further performed with the
identified compound to confirm the inhibitory properties.
Fractions of the identified compounds were diluted with
sterile distilled water. The antimicrobial solutions at vari-
ous concentrations were subjected to selected clinical iso-
lates using agar well diffusion method. All the plates were
incubated at 37°C for 48 h and the plates were screened
for zone of inhibition.
Microbial characterization of drug-producing isolates
The potent isolates selected from the preliminary screen-
ing were characterized by standard microbiology methods.
The morphological (Boobathy et al. 2009), physiological
and biochemical characteristics of each isolate were also
studied based on Bergey’s manual of systematic bacteriol-
ogy (Krieg et al. 2010). The biochemical tests performed
for the identification of the isolates include indole pro-
duction, methyl red, vogues prauskauer and citrate utili-
zation (Krieg, 2010), triple sugar iron test (Sulkin and
Willett 1940), starch hydrolysis (Colonna et al. 1992),
nitrate reduction (Moreno-Vivi�an et al. 1999), urease test
(MacFaddin 1980), catalase test (Miller et al. 2007),
oxidase test (Gordon and McLeod 1928) and hydrogen
sulphide production (Singh et al. 2012) tests.
Molecular characterization of drug-producing bacteria
The marine strain that produce antimicrobial secondary
metabolites was further characterized by 16S rRNA gene
sequencing (BioAxis DNA Research Centre Private Limited,
Hyderabad, India). The 16S rRNA gene from the selected
isolate was amplified using universal primers 16F (5′-AGA-GTTTGATCCTGGCTCAG-3′) and 16R (5′-AGA-GTTTGATCCTGGCTCAG-3′). A PCR was performed with
25 ml volume using 10 ng of genomic DNA, 19 reaction
buffer (0�01 mol l�1 Tris-HCl, pH 8�8 at 25°C, 0�0015 mol
l�1 MgCl2, 0�05 mol l�1 KCl and 0�1% Triton X-100), 0.
0�0004 mol l�1 (each) deoxynucleoside triphosphates and
0�5 U of DNA polymerase (New England Labs, Hitchin,
UK). The PCR was performed in an automated PCR system
9700 thermal cycler (Applied Biosystems, Foster City, CA,
USA) under the following conditions. The amplification
conditions were as follows 94°C for 1 min (denaturation),
55°C for 1 min (annealing), 72°C for 90 min (elongation)
and 72°C for 10 min final elongation. Expected PCR prod-
uct of around 1�5 kb was checked by electrophoresis of
5 ml of the PCR product on 1% agarose gel in 19 TBE buf-
fer and stained with 0�005 mol l�1 ethidium bromide. The
PCR product was precipitated by PEG-NaCl (20% PEG in
2�5 mol l�1 NaCl) precipitation at 37°C for 30 min. One
microlitre of purified PCR product was sequenced by auto-
mated sequence analyser (Applied Biosystems-3500, Capil-
lary sequencer). Further analysis of sequences was
performed by similarly searching tool, NCBI BLAST server
(http://www.ncbi.nlm.nih.gov/BLAST).
Acknowledgements
The authors acknowledge The Principal Chief Conserva-
tor of Forest, Govt of Tamil Nadu, India for grant per-
mission (C.No.WL5/11399/2013) for collecting the marine
sponges from Gulf of Mannar Biosphere, Ramesharam.
The authors are very much grateful to Dr. V. Deepak
Samuel, Programme Specialist, Energy and Environment
Unit, United Nations Development Programme, UNDP-
GEF, Gulf of Mannar Biosphere Reserve Trust, for his
immense support towards the collection and identifica-
tion of marine sponges.
Conflict of Interest
The authors are disclosing that there are no potential
sources of conflict.
Letters in Applied Microbiology 58, 231--241 © 2013 The Society for Applied Microbiology238
Antimicrobial potential of metabolites extracted S. Skariyachan et al.

References
Aboul-Ela, H.M., Shreadah, M.A., Abdel-Monem, N.M.,
Yakout, G.A. and van Soest, R.W.M. (2012) Isolation,
cytotoxic activity and phylogenetic analysis of Bacillus spp.
bacteria associated with the red sea sponge Amphimedon
ochracea. Adv Biosci Biotechnol 3, 815–823.
Ahamed, N. (2012) Isolation and identification of secondary
metabolites producing organisms from marine sponge.
Discovery 1, 14–17.
Aishwarya, M.S., Lipton, A.P. and Sarika, A.R. (2013)
Phylogenetic appraisal of the drug bearing marine sponge
Callyspongia subarmigera (Ridley, 1884) from South India.
Indian J Geo-Mar Sci 42, 139–145.
Anand, T.P., Bhat, A.W., Shouche, Y.S., Roy, U., Siddharth, J.
and Sarma, S.P. (2006) Antimicrobial activity of marine
bacteria associated with sponges from the waters off the
coast of South East India. Microbiol Res 161, 252–262.
Aryantha, N.P., Adinda, A. and Kusmaningat, S. (2002)
Occurrence of triterpenoids and polysaccharides on
Ganoderma tropicum with Ganoderma lucidum as
reference. Aust Mycol 20, 123–129.
Beveridge, T.J. (2001) Use of the Gram stain in microbiology.
Biotech Histochem 76, 111–118.
Boobathy, S., Soundarapandian, P., Subasri, V., Vembu, N.
and Gunasundari, V. (2009) Bioactivities of protein
isolated from marine sponge, Sigmadocia fibulatus. Curr
Res J Biol Sci 1, 160–162.
Boopathy, N.S. and Kathiresan, K. (2010) Anticancer drugs
from marine flora: an overview. J Oncol 2010, 214186.
Chairman, K., RanjitSingh, A.J.A. and Ramesh, M. (2012)
Screening twelve species of sponges for biomedical activity
in Gulf of Mannar Tuticorin Coast. Int J Mar Sci 2, 43–50.
Chu, P.M., Guenther, F.R., Rhoderick, G.C. and Lafferty, W.J.
(1999) The NIST quantitative infrared database. J Res Natl
Inst Stand Technol 104, 59.
Clarridge, J. (2004) Impact of 16S rRNA Gene sequence analysis
for identification of bacteria on clinical microbiology and
infectious diseases. Clin Microbiol Rev 17, 840–862.
Colonna, P., Leloup, V. and Bul�eon, A. (1992) Limiting factors
of starch hydrolysis. Eur J Clin Nutr 46, S17–S32.
Devi, P., Wahidullah, S., Rodrigues, C. and Souza, L.D. (2010)
The sponge-associated bacterium Bacillus licheniformis
SAB1: a source of antimicrobial compounds. Mar Drugs 8,
1203–1212.
Dhinakaran, D.I., Manohari, V., Atchya, B., Tamilselvi, K. and
Lipton, A.P. (2012) Antifungal and cytotoxic activities of
some marine sponges collected from the South East Coast
of India. J Appl Pharm Sci 2, 52–55.
Ely, R., Supriya, T. and Naik, C.G. (2004) Antimicrobial
activity of marine organisms collected off the coast of
South East India. J Exp Mar Biol Ecol 309, 121–127.
Firdouse, S. and Alam, P. (2011) Phytochemical investigation
of extract of Amorphophallus campanulatus tubers. Int J
Phytomed 3, 32–35.
Frye, J.G. and Jackson, C.R. (2013) Genetic mechanisms of
antimicrobial resistance identified in Salmonella enterica,
Escherichia coli, and Enteroccocus spp. isolated from U.S.
food animals. Front Microbiol 4, 135.
Gebhardt, K., Schimana, J., Muller, J., Fielder, H.P.,
Kallenborn, H.G., Holzenk€ampfer, M., Krastel, P., Zeeck,
A. et al. (2002) Screening for biologically active
metabolites with endosymbiotic bacilli isolated from
arthropods. FEMS Microbiol Lett 217, 199–205.
Geldreich, E.E., Nash, H.D., Reasoner, D.J. and Taylor, R.H.
(1972) The necessity of controlling bacterial populations in
potable waters; community water supply. J Am Water
Works Assoc 64, 596–602.
Gokulkrishnan, K., Kusuma, S. and Boopalan, K. (2011)
Antimicrobial activity of marine bacteria isolated from the
Mangalore coast, West Coast of India. Recent Res Sci
Technol 3, 15–17.
Gopi, M., Kumaran, S., Kumar, T.T., Deivasigamani, B.,
Alagappan, K. and Prasad, S.G. (2012) Antibacterial
potential of sponge endosymbiont marine Enterobacter
spp. at Kavaratti Island, Lakshadweep archipelago. Asian
Pac J Trop Med 5, 142–146.
Gordon, J. and McLeod, J.W. (1928) Practical application of
the direct oxidase reaction in bacteriology. J Pathol
Bacteriol 31, 185–190.
Hardoim, C.C.P., Esteves, A.I.S., Pires, F.R., Goncalves, J.M.S.,
Cox, C.J., Xavier, R.J. and Costa, R. (2012) Phylogenetically
and spatially close marine sponges harbour divergent
bacterial communities. PLoS ONE 7, e53029.
Hentschel, U., Schmid, M., Wagner, M., Fieseler, L., Gernert,
C. and Hacker, J. (2001) Isolation and phylogenetic
analysis of bacteria with antimicrobial activities from the
Mediterranean sponges Aplysina aerophoba and Aplysina
cavernicola. FEMS Microbiol Ecol 35, 305–312.
Ivanova, E.P., Vysotskii, M.V., Svetashev, V.I.,
Nedashkovskayal, O.I., Gorshkoval, N.M., Mikhailovl,
V.V., Yumoto, N., Shigeri, Y., et al. (1999)
Characterization of Bacillus strains of marine origin. Int
Microbiol 2, 267–271.
Janda, J.M. and Abbott, S.L. (2007) 16S rRNA gene sequencing
for bacterial identification in the diagnostic laboratory:
pluses, perils, and pitfalls. J Clin Microbiol 45, 2761–2764.
Kobayashi, S., Hodaka, S., Kawamura, Y., Ozaki, M. and
Hayase, Y. (1998) Micacocidin A, B and C, novel
antimycoplasma agents from Pseudomonas spp. J Antibiot
(Tokyo) 51, 323–332.
K€ock, R., Schaumburg, F., Mellmann, A., K€oksal, M., Jurke, A.,
Becker, K. and Friedrich, A.W. (2013) Livestock-associated
methicillin-resistant Staphylococcus aureus (MRSA) as
causes of human infection and colonization in Germany.
PLoS ONE 8, e55040.
Krieg, N.R., Ludwig, W., Whitman, W.B., Hedlund, B.P.,
Paster, B.J., Staley, J.T., Ward, N., Brown, D. et al. (2010)
Bergey’s Manual of Systematic Bacteriology. New York, NY:
Springer.
Letters in Applied Microbiology 58, 231--241 © 2013 The Society for Applied Microbiology 239
S. Skariyachan et al. Antimicrobial potential of metabolites extracted

Lee, Y.J., Kim, K.S., Kwon, Y.K. and Tak, R.B. (2003)
Biochemical characteristics and antimicrobials
susceptibility of Salmonella gallinarum isolated in Korea.
J Vet Sci 4, 161–166.
Liu, F., Sun, W., Su, F., Zhou, K. and Li, Z. (2012) Draft
genome sequence of the sponge-associated strain Bacillus
atrophaeus C89, a potential producer of marine drugs.
J Bacteriol 194, 4454.
MacFaddin, J.F. (1980) Biochemical Tests for Identification of
Medical Bacteria, pp. 173–183. Baltimore, MD: Williams &
Wilkins.
Marinho, P.R., Moreira, A.P., Pellegrino, F.L., Muricy, G.,
Bastos Mdo, C., Santos, K.R., Giambiagi-de Marval, M.
and Laport, M.S. (2009) Marine Pseudomonas putida: a
potential source of antimicrobial substances against
antibiotic-resistant bacteria. Mem Inst Oswaldo Cruz 104,
678–682.
Miller, J.M., Krisher, K., Holmes, H.T. (2007) General
principles of specimen collection and handling. In
Manual of Clinical Microbiology, 9th edn ed. Murray,
P.R., Baron, E.J., Jorgensen, J.H., Pfaller M.A., et al.,
p. 43. Washington, DC: American Society for
Microbiology.
Miller, J.H., Singh, A.J. and Northcote, P.T. (2010)
Microtubule-stabilizing drugs from marine sponges: focus
on peloruside A and zampanolide. Mar Drugs 8,
1059–1079.
Mol, V.P.L., Raveendran, T.V., Abhilash, K.R. and
Parameswaran, P.S. (2010) Inhibitory effect of Indian
sponge extracts on bacterial strains and larval settlement of
the barnacle, Balanus amphitrite. Int Biodeterior Biodegrad
64, 506–510.
Moreno-Vivi�an, C., Cabello, P., Mart�ınez-Luque, M., Blasco,
R. and Castillo, F. (1999) Prokaryotic nitrate reduction:
molecular properties and functional distinction
among bacterial nitrate reductases. J Bacteriol 181,
6573–6584.
Nagai, K., Kamigiri, K., Arao, N., Suzumura, K., Kawano, Y.,
Yamaoka, M., Zhang, H. and Watanabe, M. (2003) YM-
266183 and YM-266184, novel thiopeptide antibiotics
produced by Bacillus cereus isolated from marine sponge.
J Antibiot (Tokyo) 56, 123–128.
Petchi, R.R., Vijaya, C. and Parasuraman, S. (2013) Anti-
arthritic activity of ethanolic extract of Tridax procumbens
(Linn.) in Sprague Dawley rats. Pharmacognosy Res 5,
113–117.
Ravichandran, S., Wahidullah, S., D’Souza, L. and
Anbuchezhian, R.M. (2011) Antimicrobial activity of
marine sponge Clathria indica (Dendy, 1889). Bioorg Khim
37, 483–489.
Roser, D.J., Ashbolt, N., Ho, G., Mathew, K., Nair, J., Ryken-
Rapp, D. and Toze, S. (2005) Hydrogen sulphide
production tests and the detection of groundwater faecal
contamination by septic seepage. Water Sci Technol 51,
291–300.
Sabdono, A. and Radjasa, O.R. (2008) Microbial symbionts in
marine sponges: marine natural product factory. J Coast
Dev 11, 57–62.
Schwalbe, R., Steele-Moore, L. and Goodwin, A.C. (2007)
Antimicrobial Susceptibility Testing Protocols, pp. 91–109.
Boca Raton, FL: CRC Press.
Singh, D., Singh, P., Gupta, A., Solanki, S., Sharma, E. and
Nema, R. (2012) Qualitative estimation of the presence of
bioactive compound in Centella Asiatica: an important
medicinal plant. Int J Life Sci Med Res 2, 5–7.
Singla, N., Bansal, N., Gupta, V. and Chander, J. (2013)
Outbreak of Salmonella typhi enteric fever in sub-urban
area of North India: a public health perspective. Asian Pac
J Trop Med 6, 167–168.
Sulkin, S.E. and Willett, J.C. (1940) A triple sugar-ferrous
sulfate medium for use in identification of enteric
organisms. J Lab Clin Med 25, 649–653.
Taylor, M.W., Radax, R., Steger, D. and Wagner, M. (2007)
Sponge-associated microorganisms: evolution, ecology, and
biotechnological potential. Microbiol Mol Biol Rev 71, 295–
347.
Thakur, A.N., Thakur, N.L., Indap, M.M., Pandit, R.A., Datar,
V.V. and Muller, W.E.G. (2005) Antiangiogenic,
antimicrobial, and cytotoxic potential of sponge-associated
bacteria. Mar Biotechnol 7, 245–252.
Thomas, T.R., Kavlekar, D.P. and LokaBharathi, P.A. (2010)
Marine drugs from sponge-microbe association-a review.
Mar Drugs 8, 1417–1468.
Torpdahl, M., Lauderdale, T.L., Liang, S.Y., Li, I., Wei, S.H.
and Chiou, C.S. (2013) Human isolates of Salmonella
enterica serovar typhimurium from Taiwan displayed
significantly higher levels of antimicrobial resistance than
those from Denmark. Int J Food Microbiol 161,
69–75.
Usman, H., Abdulrahman, F. and Usman, A. (2009)
Qualitative phytochemical screening and in vitro
antimicrobial effects of methanol stem bark extract of
Ficus thonningii (Moraceae). Afr J Tradit Complement
Altern Med 6, 289–295.
Velho-Pereira, S. and Furtado, I. (2012) Antibacterial
activity of halophilic bacterial bionts from marine
invertebrates of Mandapam, India. Indian J Pharm Sci
74, 331–338.
Venkateswarlu, Y. and Biabani, M.A.F. (1995) Phycopsisenone
A new phenolic secondary metabolic from the sponge
Physopsis spp. J Nat Prod 58, 269–270.
Webster, N.S., Wilson, K.J., Blackall, L.L. and Hill, R.T. (2001)
Phylogenetic diversity of bacteria associated with the
marine sponge Rhopaloeides odorabil. Appl Environ
Microbiol 67, 434–444.
West, P.A. and Colwell, R.R. (1984) Identification and
Classification of Vibrionaceae – An Overview, pp. 285–363.
New York, NY: Wiley & Sons.
Zheng, L., Chen, H., Han, X., Lin, W. and Yan, X. (2005a)
Antimicrobial screening and active compound isolation
Letters in Applied Microbiology 58, 231--241 © 2013 The Society for Applied Microbiology240
Antimicrobial potential of metabolites extracted S. Skariyachan et al.

from marine bacterium NJ6-3-1 associated with the
sponge Hymeniacidon perleve. World J Microbiol Biotechnol
21, 201–206.
Zheng, L., Yan, X., Xu, J., Chen, H. and Lin, W. (2005b)
Hymeniacidon perleve, associated bioactive Pseudomonas
spp. NJ-6-3-1. Prikl Biokhim Mikrobiol 41, 35–39.
Supporting Information
Additional Supporting Information may be found in the
online version of this article:
Figure S1 Microscopic view of drug producing bacte-
rial isolates from marine sponges after Gram Staining.
Letters in Applied Microbiology 58, 231--241 © 2013 The Society for Applied Microbiology 241
S. Skariyachan et al. Antimicrobial potential of metabolites extracted