monocrotaline pyrrol is cytotoxic and alters the patterns of gfap expression on astrocyte primary...
TRANSCRIPT

Toxicology in Vitro 22 (2008) 1191–1197
Contents lists available at ScienceDirect
Toxicology in Vitro
journal homepage: www.elsevier .com/locate / toxinvi t
Monocrotaline pyrrol is cytotoxic and alters the patterns of GFAP expressionon astrocyte primary cultures
R.A. Barreto a, C.S. Sousa a, V.D.A. Silva a, A.R. Silva a, E.S. Veloso b, S.D. Cunha c, M.F.D. Costa a,R.S. El-Bachá a, S.L. Costa a,*
a Laboratório de Neuroquímica e Biologia Celular, Instituto de Ciências da Saúde, Departamento de Biofunc�ão, Universidade Federal da Bahia – UFBA, Salvador, BA 40.110-100, Brazilb Laboratório de Pesquisa em Matéria Médica, Faculdade de Farmácia, Universidade Federal da Bahia – UFBA, Salvador, BA 40170-290, Brazilc Departamento de Química Orgânica, Instituto de Química, Universidade Federal da Bahia – UFBA, Salvador, BA 40.170-290, Brazil
a r t i c l e i n f o a b s t r a c t
Article history:Received 21 November 2007Accepted 23 March 2008Available online 7 April 2008
Keywords:DehydromonocrotalinePyrrolizidine alkaloidAstrocytesGFAP
0887-2333/$ - see front matter � 2008 Elsevier Ltd. Adoi:10.1016/j.tiv.2008.03.013
Abbreviations: CNS, central nervous system; CYP4dehydromonocrotaline; DMF, dimethylformamide; Gtein; IF, intermediate filaments; LDH, lactate dehydroMTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazlizidine alkaloids.
* Corresponding author. Tel.: +55 71 3283 8919; faE-mail address: [email protected] (S.L. Costa).
Dehydromonocrotaline (DHMC) is the main monocrotaline active cytochrome P450’s metabolite, and hasalready been assessed in the CNS of experimentally intoxicated rats. DHMC effects were here investigatedtoward rat astroglial primary cultures regarding cytotoxicity, morphological changes and regulation ofGFAP expression. Cells, grown in DMEM supplemented medium, were treated with 0.1–500 lM DHMC,during 24- and 72-h. According to MTT and LDH tests, DHMC was toxic to astrocytes after 24-h exposureat 1 lM, and induced membrane damages at 500 lM. Rosenfeld dying showed hypertrophic astrocytesafter 72-h exposure to 0.1–1 lM DHMC. GFAP immunocytochemistry and western immunoblot revealedan increase of GFAP labelling and expression, suggesting an astrogliotic reaction to low concentrations ofDHMC. At higher concentrations (10–500 lM), astrocytes shrank their bodies and retracted their pro-cesses, presenting a more polygonal phenotype and a weaker expression on GFAP labelling Nuclear chro-matin staining by Hoechst-33258 dye, revealed condensed and fragmented chromatin in an importantproportion (±30%) of the astrocytes exposed to 100–500 lM DHMC, suggesting signs of apoptosis. Ourresults confirm a cytotoxic and dose-dependant effect of DHMC on cultures of rat cortical astrocytes,leading to apoptotic figures. These effects might be related to the neurological damages and clinical signsobserved in animals intoxicated by Crotalaria.
� 2008 Elsevier Ltd. All rights reserved.
1. Introduction lar macromolecules such as DNA and proteins, forming adducts
Plants of the Crotalaria genus (Leguminosae) grow abundantlyin tropical and subtropical zones and are adopted in popular med-icine (Atal and Sawhney, 1973; Mattocks, 1986). As invasive plants,they are easily found in crops plantations and pastures (Cheecke,1988; Hoehne, 1939; Tokarnia et al., 2000), and may be acciden-tally ingested by humans and animals. These plants are rich inpyrrolizidine alkaloids (PAs), the main plant toxins to cause intox-ication in human and animals (Mattocks, 1986; Huxtable, 1990).However, it is well known that, to exert their toxic effect, PA’s needto be metabolised by the hepatic enzymatic system cytochromeP450 (CYP450), generating active metabolites as dehydroalkaloidsand pyrrols (Mattocks, 1986). These composites may alkylate cellu-
ll rights reserved.
50, cytochrome P450; DHMC,FAP, glial fibrillary acid pro-genase; MCT, monocrotaline;olium bromide; PAs, pyrro-
x: +55 71 3283 8884.
that can initiate an acute or chronic toxicity (Culvenor et al.,1962). Monocrotaline (MCT) is the major PA of Crotalaria retusaresponsible for harmful effects observed in different animal species(Cheecke, 1998). Despite this alkali is primarily considered hepato-toxic, a pneumotoxic, nefrotoxic, cardiotoxic, fetotoxic and clasto-genic effects were also related to its intoxication in animals(Mattocks, 1986; Thomas et al., 1996; Ribeiro et al., 1993; Cheecke,1998; Medeiros et al., 2000). In addition, species particularly sensi-ble to intoxication by MCT, such as horses, show a classic liverfibrosis and neurological symptoms (Kimberly Horse Disease)associated to a chronic exposure to the alkaloid (Rose et al.,1957; Nobre et al., 2004).
Glial cells and mainly astrocytes, the major cells of the glialfamily in the central nervous system (CNS), are essential for thenutritional and structural support to neurons in the CNS. Thesecells also participate in other functions such as immune responseand cerebral detoxification, displaying an active CYP450 system(Tardy, 1991; Letournel-Boulland et al., 1994; Montgomery,1994; Kandel, 2000). Thus, modifications of glial cells, and inparticular of those occurring on the astrocytic lineage, may consti-tute biological markers for many types of damages in the CNS. In a

1192 R.A. Barreto et al. / Toxicology in Vitro 22 (2008) 1191–1197
previous in vitro study over rat astrocyte primary cultures (Barretoet al., 2006), we observed that MCT induced changes on these cellsregarding their growth and morphology. These phenomena wereassociated with changes in the pattern of the glial fibrillary acidicprotein (GFAP), the main component of intermediate filaments(IF) in astrocytes, suggesting that these cells are involved in neuro-logical damages observed in an intoxicated animal.
It has been shown that, besides N-oxide derivatives, the mainhepatic CYP450 MCT metabolite is the dehydroalkaloid dehydro-monocrotaline (DHMC) (Mattocks, 1986). DHMC alkylates macro-molecules such as DNA and the cytoskeletal component actin,leading to formation of adducts which can initiate a chronic oracute intoxication (Huxtable, 1989; Hincks et al., 1991; Pereiraet al., 1998; Prakash et al., 1999). Moreover, MCT and DHMC werefound and quantified in the brain of experimentally intoxicatedrats, which demonstrates that both molecules are able to crossthe blood-brain barrier (Yan and Huxtable, 1995), or even thatDHMC might be generated after glial or neural CYP450 metabolismof MCT. In the present study, we investigated the cytotoxic effect ofDHMC, generated from MCT extracted from Crotalaria retusa, on ratastrocyte primary culture and its effect on the pattern and expres-sion levels of GFAP.
2. Material and methods
2.1. Astrocyte primary cultures
One-day-old postnatal Wistar rat pups used in this study wereobtained from the animal house facility of the Instituto de Ciênciasda Saúde, UFBA. Astrocyte cultures were prepared according toCookson and Pentreath (1994), as described (Silva et al., 2007).Briefly, cerebral hemispheres of newborn Wistar rat pups were iso-lated asseptically and meninges were removed. Neopallia were dis-sected out and then gently forced through a sterile 75 lm Nitexmesh. Cells were suspended in Dulbecco’s Modified Eagle’s Med-ium (DMEM, Cultilab, SP, Brazil), supplemented with 100 UI/mlpenicillin G, 100 lg/ml streptomycin, 2 mM L-glutamine, 0.011 g/lpyruvate, and 10% foetal calf serum (Cultilab, SP, Brazil), seededon polystyrene culture dishes (TPP, Switzerland), and incubatedin a humidified atmosphere with 5% CO2 at 37 �C. After 2 days, cul-tures were shaken for 5 min and the medium was changed to re-move microglial cells. As determined by immunocytochemistryfor the protein GFAP, astrocyte primary cultures presented about95% of GFAP-labelled astrocytes.
2.2. Alkaloids extraction
Monocrotaline was extracted and purified from an aqueous ex-tract from C. retusa seeds according to Culvenor and Smith (1957)method and previously described by Barreto et al. (2006), exceptthat hexane was used instead of petroleum ether and possible N-oxides forms were not converted into free bases. The attainmentof the DHMC was carried through the technique described by Mat-tocks et al. (1989). Briefly, 20 mg of MCT was weighed and dis-solved in 5 ml of chloroform. To this chloroform solution wasadded 5 ml of another chloroform solution contending 25 mg of3,4,5,6-tetrachloro-1,2-benzoquinone (o-chloranyl), the oxidantagent. Passed 2 min, this mixture was agitated vigorously per15 s with 1 ml of a cooled solution of 70% KOH and 2% borehidretoof sodium, being this last the reducing agent of the reaction. Theorganic phase was then transferred to a separation funnel, dehy-drated with sodium sulphate (Na2SO4) and adsorbed with coalafter activated. The chloroform phase was recrystallised in rotava-porizer, resulting in a practically pure dehidroalkaloid. The charac-terisation of the MCT was carried through nuclear magnetic
resonance of 13C and 1H, beyond spectre infrared ray. The DHMCwas confirmed by nuclear magnetic resonance of 1H.
2.3. Treatments
For treatments, DHMC was dissolved in dimethylformamide(DMF, Sigma, St. Louis, MO) forming 100 mM stock solution imme-diately before experiments. After 18 days in vitro, cells were trea-ted with DHMC at concentrations ranging from 0.1 to 500 lM for24-h and 72-h. The negative control group was treated with DMFdiluted in the culture medium at the higher equivalent volumeused in the treated group (0.1%), and showed no significant effecton analysed parameters compared to cells that did not receivethe diluent.
2.4. Cytotoxic effects
2.4.1. Induction of membrane damagesMembrane integrity was evaluated by measuring the lactate
dehydrogenase (LDH) activity in culture medium of control andtreated cells and evaluated as an index of cell damage. Cells weregrown in 40 mm in diameter plates (1.5 � 105 cells/plate) treatedwith 0.1–500 lM DHMC or with 0.1% DMF (control), for 24-h. Afterthat, the culture medium was removed and the LDH activity (UI/L)was measured according to manufacturer protocol (Doles, Goiás,Brazil). Three independent experiments were carried out for eachexperimental point.
2.4.2. Cell viabilityThe effect of DHMC on the metabolism of astrocyte primary cul-
tures was tested using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphe-nyltetrazolium bromide (MTT; Sigma, St. Louis, MO) test. Theexperiment was performed in 96 well plates (TPP Switzerland)(1 � 104 cells/well), and cells were incubated with 0.1–500 lMDHMC, or 0.1% DMF (control) for 24-h and 72-h. The cell viabilitywas quantified by the conversion of yellow MTT by mitochondrialdehydrogenases of living cells to purple MTT formazan (Hansenet al., 1989). Control and treated cells were incubated with MTTat a final concentration of 1 mg/ml for 2-h. Thereafter, cells werelysed with 20% (w/v) sodium dodecyl sulphate (SDS), 50% (v/v)dimethylformamide (DMF) (pH 4.7), and plates were kept over-night at 37 �C in order to dissolve formazan crystals. The opticaldensity of each sample was measured at 492 nm using a spectro-photometer (Thermo Plate-Reader). Three independent experi-ments were carried out with eight replicate wells for eachanalysis. Results from MTT test were expressed as percentages ofthe viability of the treated groups related to the control groups.
2.5. Determination of morphological changes
2.5.1. Rosenfeld’s staining and immunocytochemistryMorphological changes were primarily assessed by analysing
the Rosenfeld’s staining. All control and treated cells seeded onpolystyrene culture dishes of 40 mm in diameter (1.5 � 105 cells/plate) were rinsed three times with PBS (without Ca2+ and Mg2+)and fixed for 10 min with methanol at –20 �C. Fixed cells werestained by the protocol established by Rosenfeld (Rosenfeld,1947). The Rosenfeld’s reagent (1 ml) was added and incubatedfor 20 min at room temperature. Thereafter, the plates were rinsedwith water, air-dried, analysed in an optic phase microscope (Ni-kon TS-100) and photographed using a digital camera (Nikon E-4300).
Morphological changes were also studied by the immunocyto-chemistry patterns for the specific cytoskeletal protein GFAP. Allcontrol and treated cells seeded on polystyrene culture plates of40 mm in diameter (1.5 � 105 cells/plate) were rinsed three times

Fig. 1. Cytotoxicity assay on astrocyte primary cultures treated with the equivalentvolume of the vehicle of dilution (DMF) or with the monocrotaline pyrrol DHMC. (A)Measure of LDH activity (mean ± SD) on the culture medium of cells after 24-hexposure to DMF (0.1 %) or 0.1–1 lM DHMC. (B) MTT test of cells after 24-h and 72-h exposure to LDH activity on the culture medium of cells exposed to DMF (0.1%) or0.1–1 lm DHMC; values for vehicle control samples were set at 100%; *P < 0.05%.
R.A. Barreto et al. / Toxicology in Vitro 22 (2008) 1191–1197 1193
with PBS and fixed with cold methanol at �20 �C for 10 min. Non-specific binding of antibody reagents was blocked by pre incubat-ing the plates with 3% bovine serum albumin (BSA) in PBS. Cellswere incubated with rabbit polyclonal anti-GFAP (1/500 in PBS,DAKO, Denmark) overnight and then with tetramethylrhodamineisothiocyanate conjugated goat anti-rabbit IgG antibody (1/250 inPBS, Sigma, St. Louis, MO) for 30 min at room temperature. Nuclearchromatin of fixed cells was stained with the fluorescent dye Hoe-chst 33258 (Sigma, St. Louis, MO), at a final concentration of 5 lg/ml in PBS, for 10 min at room temperature in a dark chamber.Thereafter, cells were analyzed using an epifluorescent microscope(Olympus BX70) and photographed. Always, 10 randomised repre-sentative fields were analysed.
2.5.2. Protein assay and western immunoblotGFAP expression was also investigated by western immunoblot.
Total protein content was determined in control or treated cellsseeded on polystyrene culture dishes of 40 mm in diameter(1.5 � 105 cells/plate). After 72-h exposure, cells were rinsed twicewith PBS, lysed and harvested in a 2% (w/v) SDS, 2 mM ethylene gly-col-bis [b-aminoethyl ether]-N,N,N0N0-tetraacetic acid (EGTA), 4 MUrea, 0.5% (v/v) Triton X-100, 62.5 mM Tris–HCl buffer (pH 6.8),supplemented with 1 ll/ml of a cocktail of proteases inhibitors (Sig-ma, St. Louis, MO). Protein content was determined by the methodof Lowry et al. (1951), using a protein assay reagent kit (Bio-Rad,Hercules, CA). For western immunoblot analysis, 10 lg proteinswere loaded onto a discontinuous 4% stacking and 10% runningSDS polyacrylamide gel (SDS–PAGE). Electrophoresis was per-formed at 200 V for 45 min. Proteins were then transferred to anitrocellulose membrane (Bio-Rad, Hercules, CA, USA), at 100 Vfor 1-h. Equal protein loading was confirmed by staining the mem-branes with Ponceau Red (Sigma). Thereafter, membranes wereblocked for 1-h at room temperature in 20 mM Tris-buffered saline(pH 7.5), containing 0.05% Tween 20 (TBS-T) and 5% powdered skimmilk. Subsequently, membranes were incubated with rabbit anti-GFAP (1:1,000, SIGNET, Dedham, MA) or for 1-h, diluted in TBS-Tcontaining 1% powdered skim milk. Conjugated alkaline phospha-tase goat anti-rabbit IgG or goat anti-mouse IgG (1:5000 in TBS-T,Bio-Rad, Hercules, CA) were used as secondary antibodies. Immu-noreactive bands were visualized using AP conjugated substrateKit (Bio-Rad, Hercules, CA) according to manufacturer’s instruc-tions. Quantification was obtained by scanning densitometry (Scan-Jet 4 C – HP) of three independent experiments, and analyzed withImageJ 1.33u (Wayne Rasband, National Institute of Helth, USA).Antibody specificity and linearity of the densitometric analysis sys-tem was assessed by serial dilutions of total protein from cells incontrol conditions within a range of 5–20 lg of protein per lane.
2.6. Statistical analysis
Results are expressed as mean ± standard deviation. One wayANOVA followed by the Student–Newman–Keuls test was usedto determine the statistical differences among groups differing inonly one parameter. Student’s t-test was used to compare twogroups and values of P < 0.05 were considered as significant.
3. Results
3.1. Cell viability
Firstly, the effect of DHMC on the cellular viability was deter-mined by the measure of LDH activity in the culture medium aftershort time exposure (24-h). It revealed no effects on membraneintegrity of glial cells exposed to 0.1–100 lM DHMC (Fig. 1A). How-ever, when cells were exposed to 500 lM DHMC, the levels of LDH
activity in the culture medium significantly increased (P < 0.05).The exposure of the cells during 24-h to the dilution vehicle DMFin equivalent volume to the adopted concentrations, did not showany significant alteration in the analyzed parameters when com-pared with cultures that were not exposed to this solvent.
Moreover, DHMC altered the mitochondrial function of astro-cytes, as revealed by MTT test. No significant changes could be ob-served after 24-h treatment of the cells with 0.1 lM DHMC whencompared to the control (0.1% DMF) (Fig. 1B). But exposure to 1–500 lM DHMC caused a significant (P < 0.05) and dose-dependantreduction of cell viability (Fig. 1B). However, after 72-h experi-ment, a significant (P < 0.05) reduction of cell viability was evi-denced only at the higher concentrations of DHMC (100–500 lM)tested.
3.2. Cell morphology
The morphology and GFAP immunostaining pattern of cellstreated with the vehicle DMF (0.1%) or with DHMC (0.1–500 lM)after 72-h were investigated and are shown in Figs. 2 and 3.
Rosenfeld staining of the cells showed that the majority ofastrocytes in control conditions (0.1% DMF) present a flat/polygo-nal phenotype, with some cells presenting cellular processes(Fig. 2). However, after 72-h exposure, morphological changeswere evidenced in astrocytes exposed to DHMC, depending onthe concentration adopted. Astrocytes exposed to 0.1–1 lM DHMCshowed an enlarged cell body and extending processes (Fig. 2B–C).On the other hand, cells treated with higher concentrations, espe-

Fig. 2. Photomicrograph of astrocyte primary cultures in control conditions (0.1 % DMF) or after 72-h exposure to 0.1–500 lm DHMC (B–F), and staining by Rosenfeld’sstaining. Objective 20 � 0.70, Scale bars = 100 lm.
1194 R.A. Barreto et al. / Toxicology in Vitro 22 (2008) 1191–1197
cially at 100–500 lM DHMC, altered the characteristic astrocyticmorphology, shrinking their cell body and retracting their extend-ing processes (Fig. 2D–F).
Astrocyte cultures immunolabelled for GFAP in control condi-tions (0.1% DMF) present a monolayer of large flat cells with astar-like shape (Fig. 3A) and a GFAP labeling distributed in cellbody and its processes. Astrocytes exposed to low concentrationsof DHMC (0.1–1 lM) presented an enhanced GFAP labelling andan increased GFAP expression, with an enlargement of cell bodyand a strong tendency of forming enlarged processes (Fig. 3B–C).Some polygonal cells with enlargement of cell body were also evi-dent in cultures treated with the intermediary concentration of10 lM DHMC (Fig. 3D). However, with the increase in DHMC con-centration (10–500 lM), GFAP labelling revealed astrocytes with amore polygonal phenotype. Changes in the intracellular localisa-tion of GFAP labelling were also observed, occurring mainly inthe perinuclear compartment. A general weaker immunolabellingand even the apparition of intracellular labelled aggregates werealso common founds on treatment with higher concentrations ofDHMC (Fig. 3D–F).
3.3. GFAP steady state level
GFAP expression was also examined by western immunoblot-ting, 72-h post-treatment with DHMC (Fig. 4). GFAP is showed as
an immunoreactive band of 49 kDa. Astrocytes treated with 0.1–1 lM DHMC over expressed GFAP, compared to control. In themeantime, a strong decrease in GFAP expression level was ob-served on astrocytes after exposure to 10, 100 or 500 lM DHMC.
3.4. Hoechst staining
Nuclear chromatin staining by Hoechst-33258 dye, revealedcondensed and fragmented chromatin in an important proportion(±30%) of astrocytes exposed to 10–500 lM DHMC, which mightbe an indicative of apoptosis (Fig. 5).
4. Discussion and conclusions
Among the major cell types of the CNS, astrocytes play an impor-tant role in brain homeostasis, detoxification, neuroprotection andimmune response against infections, trauma or toxicants (Tardy,1991; Letournel-Boulland et al., 1994; Montgomery, 1994; Coyleand Schwarcz, 2000; Kandel, 2000). Their activation, know as astro-cyte reactivity or astrogliosis, may interfere with neuronal cellhealth and integrity (Cookson et al., 1994; Coyle and Schwarcz,2000; Moises et al., 2002). Pyrrolizidine alkaloids such as MCT areone of the most potent vegetal toxins to human and animals (Roseet al., 1957; Gardiner et al., 1965; Gibbons et al., 1953; Curran et al.,1996; Hooper and Scanlan, 1977; Alfonso et al., 1993; Nobre et al.,
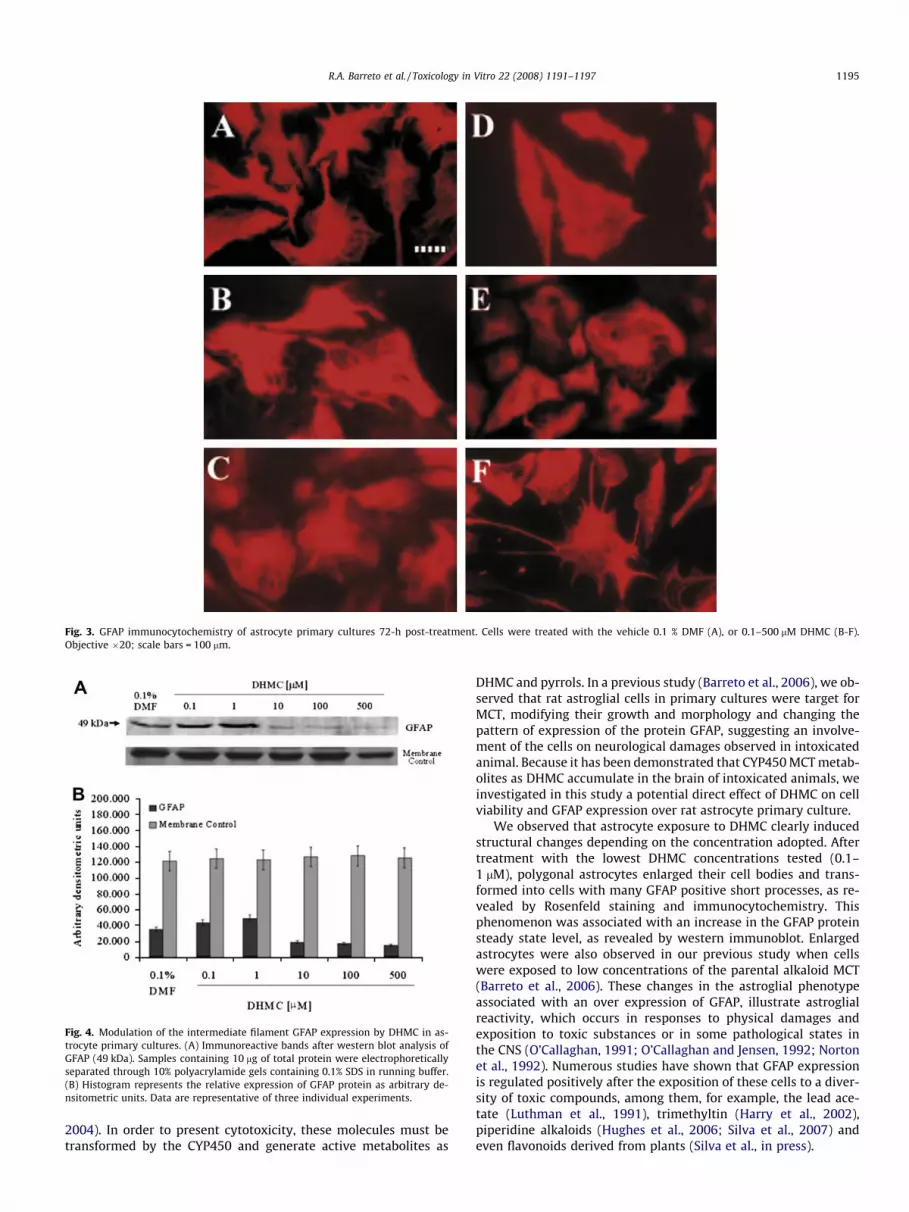
Fig. 3. GFAP immunocytochemistry of astrocyte primary cultures 72-h post-treatment. Cells were treated with the vehicle 0.1 % DMF (A), or 0.1–500 lM DHMC (B-F).Objective �20; scale bars = 100 lm.
Fig. 4. Modulation of the intermediate filament GFAP expression by DHMC in as-trocyte primary cultures. (A) Immunoreactive bands after western blot analysis ofGFAP (49 kDa). Samples containing 10 lg of total protein were electrophoreticallyseparated through 10% polyacrylamide gels containing 0.1% SDS in running buffer.(B) Histogram represents the relative expression of GFAP protein as arbitrary de-nsitometric units. Data are representative of three individual experiments.
R.A. Barreto et al. / Toxicology in Vitro 22 (2008) 1191–1197 1195
2004). In order to present cytotoxicity, these molecules must betransformed by the CYP450 and generate active metabolites as
DHMC and pyrrols. In a previous study (Barreto et al., 2006), we ob-served that rat astroglial cells in primary cultures were target forMCT, modifying their growth and morphology and changing thepattern of expression of the protein GFAP, suggesting an involve-ment of the cells on neurological damages observed in intoxicatedanimal. Because it has been demonstrated that CYP450 MCT metab-olites as DHMC accumulate in the brain of intoxicated animals, weinvestigated in this study a potential direct effect of DHMC on cellviability and GFAP expression over rat astrocyte primary culture.
We observed that astrocyte exposure to DHMC clearly inducedstructural changes depending on the concentration adopted. Aftertreatment with the lowest DHMC concentrations tested (0.1–1 lM), polygonal astrocytes enlarged their cell bodies and trans-formed into cells with many GFAP positive short processes, as re-vealed by Rosenfeld staining and immunocytochemistry. Thisphenomenon was associated with an increase in the GFAP proteinsteady state level, as revealed by western immunoblot. Enlargedastrocytes were also observed in our previous study when cellswere exposed to low concentrations of the parental alkaloid MCT(Barreto et al., 2006). These changes in the astroglial phenotypeassociated with an over expression of GFAP, illustrate astroglialreactivity, which occurs in responses to physical damages andexposition to toxic substances or in some pathological states inthe CNS (O’Callaghan, 1991; O’Callaghan and Jensen, 1992; Nortonet al., 1992). Numerous studies have shown that GFAP expressionis regulated positively after the exposition of these cells to a diver-sity of toxic compounds, among them, for example, the lead ace-tate (Luthman et al., 1991), trimethyltin (Harry et al., 2002),piperidine alkaloids (Hughes et al., 2006; Silva et al., 2007) andeven flavonoids derived from plants (Silva et al., in press).

Fig. 5. Determination of apoptois induced by DHMC in astrocyte primary cultures. Hoechst-33258 staining of nuclear chromatin was performed 72-h post-treatment incontrol condition (0.1% DMF) (A), and in cultures of cells treated with 10 lM (B), 100 lM (C) or 500 lM DHMC (D), showing nuclear condensation and fragmentation (arrows).Objective 20 � 0.70; scale bars = 10 lm.
1196 R.A. Barreto et al. / Toxicology in Vitro 22 (2008) 1191–1197
While an increase in GFAP production may be a sign of astrogl-iosis, reactive injury, and even neurodegeneration (Tardy, 1991;Coyle and Schwarcz, 2000; Costa et al., 2002), a decrease in its lev-els may signify abnormal synaptogenesis and neurotransmission(O’Callaghan, 1991; Rajkowska et al., 2002). Astrocytes reacted tothe highest dose of DHMC (10–500 lM) with a lost of typical astro-cyte star-like phenotype and retraction of the cell body, associatedwith changes on GFAP pattern of immunostaining or reduction ofits expression. GFAP is the major IF of the cytoskeletal of differen-tiated astrocytes (Gomes et al., 1999), and its assembly as polymershas an important role in determining the cellular shape. Consider-ing that PA’s metabolites are able to alkilate macromolecules likethe cytoskeletal protein actin (Wilson et al., 1998), we suggest thatDHMC may be interacting with GFAP, changing its assembly andstability. Down-regulation on GFAP expression in astrocyte pri-mary cultures exposed to the parental alkaloid MCT was also ob-served in our previous study (Barreto et al., 2006). Generation ofDHMC by astrocytes through CYP450 might be a pivotal eventfor GFAP disruption induction.
Associated with these data, the analysis of the morphologic as-pects of the cell nuclei with the intercalating Hoechst-33258,showed a condensation and fragmentation of the chromatin inastrocytes treated with the highest DHMC concentrations adopted,and mainly at 500 lM DHMC. It has been clearly demonstrated indifferent cell cultures, such as hepatic, pulmonary and vascular celllines (Hincks et al., 1991; Kim et al., 1995; Pereira et al., 1998; Kos-ogof et al., 2001; Thomas et al., 1996; Wilson et al., 1998), that MCT(and others PAs) metabolites can also alkilate macromolecules ofDNA (Wang et al., 2005; Xia et al., 2006) and induce genotoxicity.Therefore, changes in astrocytes DNA integrity observed afterexposure to DHMC, might be a result of DNA alkilation or even asign that cells entered in apoptosis. To clarify this, complementarystudies about the potential of DHMC on inducing genotoxicity and/or apoptosis might be performed.
The cleavage of tetrazoline ring in MTT involves the mitochon-drial succinate dehydrogenase and depends on the activity of therespiratory chain and the redox state of the mitochondria. Shorttime exposure (24-h) to 1–500 lM DHMC, as shown by MTT test,caused cytotoxicity on astrocytes. In in vitro studies, the measure
of the LDH activity in the culture medium is another useful param-eter to investigate cell toxicity. Although DHMC was toxic to glialcells at concentrations greater than 1 lM, as evidenced by theMTT assay, only at 500 lM and after 24-h it induced a significantincrease in LDH activity on the culture medium. These results,associated with chromatin condensation and fragmentation as wellas disruption of GFAP, observed after 10–500 lM DHMC exposure,indicates that, besides necrosis, the MCT metabolite induces loss ofDNA and cytoskeletal stability in astrocytes. This then leads to achange in cellular metabolism and viability, resulting in a pro-grammed cell death as apoptosis. Concerning the fundamentalroles of astrocytes on the CNS function and homeostasis, the im-pact of DHMC on these cells may have an impact on neurons sta-bility and may be related to damages observed in intoxicatedanimals.
References
Alfonso, H.A., Sanchez, L.M., Figueredo, M.A., Gómez, B.C., 1993. Intoxication dueCrotalaria retusa and C. spectabillis in chickens and geese. Veterinary and HumanToxicology 32, 539.
Atal, C.K., Sawhney, R.S., 1973. The pyrrolizidine alkaloids from Indian Crotalarias.Indian Journal of Pharmacology 35, 1–12.
Barreto, R.A., HugheS, J.B., Souza, C.S., Silva, V.D.A., Silva, A.R., Velozo, E.S., Batatinha,M.J.M., Costa, M.F.D., El-Bachá, R.S., Costa, S.L., 2006. The pyrrolizidine alkaloidmonocrotaline, extracted from Crotalaria retusa, interfere on cellular growth,alters GFAP expression and induces morphological changes on astrocyteprimary cultures. Brazilian Journal of Animal Health and Production 7 (2),112–127.
Cheecke, P.R., 1988. Toxicity and metabolism of pyrrolizidine alkaloids. Journal ofAnimal Science 66, 2343–2350.
Cookson, M.R., McClean, R., Williams, S.P., Davenport-Jones, J., Egan, C., O’Hare, S.,1994. Use of astrocytes for in vitro neurotoxicity testing. Toxicology in Vitro 8(4), 817–819.
Cookson, M.R., Pentreath, V.W., 1994. Alterations in the glial fibrillary acidic proteincontent of primary astrocyte cultures for evaluation of glial cell toxicity.Toxicology In vitro 8 (3), 351–359.
Costa, S.L., Planchenault, T., Charrière-Bertrand, C., Mouchel, Y., Fages, C., Juliano, S.,2002. Astroglial permissivity for neurotic outgrowth in neuron-astrocytecocultures depends on regulation of laminin bioavailability. Glia 37, 105–113.
Coyle, J.T., Schwarcz, R., 2000. Mind glue: implications of glial cell biology forpsychiatry. Archives of General Psychiatry 57 (1), 90–93.
Culvenor, C.C.J., Dann, A.T., Dick, A.T., 1962. Alkylation as the mechanism by whichthe hepatotoxic pyrrolizidine alkaloids act on cell nuclei. Nature 195 (4841),570–573.

R.A. Barreto et al. / Toxicology in Vitro 22 (2008) 1191–1197 1197
Culvenor, C.C.J., Smith, L.W., 1957. The alkaloids of Crotalaria retusa L.. AustralianJournal of Chemistry 10, 464–473.
Curran, J.M., Sutherland, R.J., Peet, R.L., 1996. A screening test for subclinical liverdisease in horses affected by pyrrolizidine alkaloid toxicosis. AustralianVeterinary Journal 74 (3), 236–240.
Gardiner, M.R., Royce, R., Bokor, A., 1965. Studies on Crotalaria crispata, a newlyrecognised cause of Kimberley horse disease. Journal of comparative pathologyand bacteriology 89, 43–53.
Gibbons, W.J., Durr, E.H., Cox, S.A., 1953. An outbreak of cirrosis of the liver inhorses. North American Veterinary 34, 556–558.
Gomes, F.C.A., Paulin, D., Moura Neto, V., 1999. Glial fibrillary acidic protein (GFAP):modulation by growth factors and its implication in astrocyte differentiation.Brazilian Journal of Medical and Biological Research 32, 619–631.
Hansen, M.B., Nielsen, S.E., Berg, K., 1989. Re-examination, and further developmentof a precise and rapid dye method for measuring cell growth/cell kill. Journal ofImmunological Methods 119, 203–210.
Harry, G.J., Tyler, K., Lefebvre d’Hellencourt, C., Tilson, H.A., Maier, W.E., 2002.Morphological alterations and elevations in tumor necrosis factor-a, interleukin(IL)-1a, and IL-6 in mixed glia cultures following exposure to trimethyltin:modulation by proinflammatory cytokine recombinant proteins andneutralizing antibodies. Toxic and Applied Pharmacology 180, 205–218.
Hincks, J.R., Kim, H.Y., Segall, H.J., Molyneux, R.J., Stermitz, F.R., Coulombe, R.J., 1991.DNA cross-linking in mammalian cells by pyrrolizidine alkaloids: structure–activity relationships. Toxicology Applied Pharmacology 111, 90–98.
Hoehne, F.C., 1939. Leguminosas papilionaceas: Crotalaria e Lupinus. In: Hoehne,F.C. (Ed.), Plantas e Substâncias Vegetais Tóxicas e Medicinais. siam, São Paulo,p. 137–444.
Hooper, P.T., Scanlan, W.A., 1977. Crotalaria retusa poisoning in pigs and poultry.Australian Veterinary Journal 53, 109–114.
Hughes, J.B., Souza, J.S., Barreto, R.A., Silva, A.R., Souza, C.S., Silva, V.D.A., Souza,B.M.P.S., Freitas, S.R.V.B., Costa, M.F.D., El-Bachá, R.S., Batatinha, M.J.M., Tardy,M., Costa, S.L., 2006. Cytotoxic effects of an extract containing alkaloidsobtained from Prosopis juliflora Sw. D.C. (Algaroba) pods on glial cells. BrazilianJournal of Veterinary Research and Animal Science 6, 31–41.
Huxtable, R.J., 1990. Activation and pulmonary toxicity of pyrrolizidine alkaloids.Pharmacology and Therapeutics 47, 371.
Huxtable, R.J., 1989. Human health implications of pyrrolizidine alkaloids and herbscontaining them. In: Cheecke, P.R. (Ed.), Toxicants of Plants Origin. Boca Raton,Florida.
Kandel, E.R., 2000. Nerve cells and behavior. In: Kandel, E.R., Schwartz, J.H., Jessell,T.M. (Eds.), Principles of neural science. fourth ed. New York.
Kim, H.Y., Stermitz, F.R., Coulombe Jr., R.A., 1995. Pyrrolizidine alkaloid-inducedDNA-protein cross-links. Carcinogenesis 16 (11), 2691–2697.
Kosogof, C., Tepea, J.J., Williams, R.M., 2001. DNA cross-linking by a photo triggeredpyrrolic progenitor developed from monocrotaline. Tetrahedron Letters 42,6641–6643.
Letournel-Boulland, M.L., Fages, C., Rolland, B., Tardy, M., 1994. Lipopolysaccharides(LPS) up-regulate the IL-1-mRNA and downregulate the glial fibrillary acidicprotein (GFAP) and glutamine synthetase (GS)-mRNAs in astroglial primaryculture. European Cytokine Network 5 (1), 51–56.
Lowry, O.H., Rosenbroug, N.J., Farr, A.L., Randall, R.J., 1951. Proteinmeasurement with the Folin phenol reagent. Journal of Biology Chemistry193, 265–275.
Luthman, J., Olson, L., Bjorklund, H., Henschen, A., Hoffer, B., Oskarsson, A., 1991.Combined lead acetate and disulfiram treatment-induced alterations of glialfibrillary acidic protein (GFA) immunoreactive astrocytes in brain smears.Toxicology 65 (3), 333–346.
Mattocks, A.R., 1986. Toxicology of pyrrolizidine alkaloids in animal. In: Chemistryand Toxicology of Pyrrolizidine Alkaloids. Academic Press, London, New York,pp. 191–219.
Mattocks, A.R., Jukes, R., Brown, J., 1989. Simple procedures for preparing putativetoxic metabolites of pyrrolizidine alkaloids. Toxicon 27 (5), 561–567.
Medeiros, R.M.T., Górniak, S.L., Guerra, J.L., 2000. Fetotoxicity and reproductiveeffects of monocrotaline in pregnant rats. Journal of Ethnopharmacology 69,181–188.
Moises, H.W., Zoega, T., Gottesman II, 2002. The glial growth factors deficiency andsynaptic destabilization hypothesis of schizophrenia. BMC Psychiatry 2 (1), 8.
Montgomery, D.L., 1994. Astrocytes: form, function and roles in diseases. VeterinaryPathology 31, 145–167.
Nobre, V.M.T., Riet-Correa, F., Dantas, A.F.M., Barbosa Filho, J.M., Tabosa, I.M., 2004.Vasconcelos JS. Intoxicac�ão por Crotalaria retusa (Fabaceae) em equídeos nosemi-árido da Paraíba. Pesquisa Veterinária Brasileira 24 (3), 132–143.
Norton, W.T., Aquino, D.A., Hozumi, I., Chiu, F.C., Brosnan, C.F., 1992. Quantitativeaspects of reactive gliosis. A review. Neurochemical Research 17, 877–885.
O’Callaghan, J.P., 1991. Assessment of neurotoxicity: use of glial fibrillary acidicprotein as a biomarker. Biomedical and Environmental Sciences 4, 197–206.
O’Callaghan, J.P., Jensen, K.F., 1992. Enhanced expression of glial fibrillary acidicprotein and the cupric silver degeneration reaction can be used as sensitive andearly indicators of neurotoxicity. Neurotoxicology 13 (1), 113–122.
Pereira, T.N., Webb, R.I., Reilly, P.E.B., Seawright, A.A., Prakash, A.S., 1998.Dehydromonocrotaline generates sequence-selective N-7 guanine alkylationand heat and alkali stable multiple fragment DNA crosslinks. Nucleic AcidsResearch 26, 5441–5447.
Prakash, A.S., Pereira, T.N., Reilly, P.E., Seawright, A.A., 1999. Pyrrolizidine alkaloidsin human diet. Mutation Research 5, 53–67.
Rajkowska, G., Miguel-Hidalgo, J.J., Makkos, Z., Meltzer, H., Overholser, J.,Stockmeier, C., 2002. Layer-specific reductions in GFAP-reactive astrogliain the dorsolateral prefrontal cortex in schizophrenia. Schizophr Research 57(2–3), 127–138.
Ribeiro, L.R., Silva, A.R., Bautista, A.R., Costa, S.L., Sales, L.A., Rios, A.C., Salvadori,D.M.F., 1993. Clastogenic effect of extracts obtained from Crotalaria retusa L. andCrotalaria mucronata Desv. on mouse bone marrow cells. Mutation Research300, 253–258.
Rose, A.L., Gardner, C.A., Mc Connell, J.D., Bull, L.B., 1957. Field and experimentalinvestigations of ‘‘walk-about” disease of horses (Kimberly horse disease) inNorthern Australia. Crotalaria poisoning in horses. Australian Veterinary Journal33, 25–33.
Rosenfeld, G., 1947. Corante pancrômico para hematologia e citologia clínica: Novacombinac�ão dos componentes do May Grunwald e do Giemsa num só corantede emprego prático. Memória do Instituto Butantã 20, 329–335.
Silva, A.M.M., Silva, A.R., Pinheiro, A.M., Freitas, S.R.V.B., Silva, V.D.A., Souza, C.S.,Velozo, E.S., El-Bachá, R.S., Costa, M.F.D., Costa, S.L., 2007. Alkaloids fromProsopis juliflora leaves induce glial activation, cytotoxicity and stimulate noproduction. Toxicon 49 (5), 601–614.
Silva, A.R., Pinheiro, A.M., Souza, C.S., Freitas, S.R.V.B., Vasconcellos, V., Freire, S.M.,Velozo, E.S., Tardy, M., El-Bachá, R.S., Costa, M.F.D., Costa, S.L., in press. Theflavonoid rutin induces astrocyte and microglia activation and regulates TNF-alpha and NO release in primary glial cell cultures. Cell Biology and Toxicology.doi:10.1007/s10565-007-9017-y.
Tardy, M., 1991. Astrocyte et homeostasie. Médecine Science 8 (7), 799–804.Thomas, H.C., Lamé, M.W., Wilson, D.W., Segall, H.J., 1996. Cell cycle alterations
associated with covalent binding of monocrotaline pyrrole to pulmonary arteryendothelial cell DNA. Toxicology and Applied Pharmacology 141 (1), 319–329.
Tokarnia, C.H., Döbereiner, J., Peixoto, P.V., 2000. Plantas experimentalmente tóxicaspara bovinos, porém incerto se de interesse pecuário. In: Tokarnia, C.H.,Döbereiner, J., Peixoto, P.V. (Eds.), Plantas Tóxicas do Brasil, 2nd ed. Helianthus,Rio de Janeiro, pp. 240–242.
Wang, Y.P., Yan, J., Beger, R.D., Fu, P.P., Chou, M.W., 2005. Metabolic activation of thetumorigenic pyrrolizidine alkaloid, monocrotaline, leading to DNA adductformation in vivo. Cancer Letters 226, 27–35.
Wilson, D.W., Lamé, M.W., Dunston, S.K., Taylor, D.W., Segall, H.J., 1998.Monocrotaline pyrrole interacts with actin and increases thrombin-mediatedpermeability in pulmonary artery endothelial cells. Toxicology and AppliedPharmacology 152, 138–144.
Xia, Q., Chou, M.W., Edgar, J.A., Doergea, D.R., Fu, P.P., 2006. Formation of DHP-derived DNA adducts from metabolic activation of the prototype heliotridine-type pyrrolizidine alkaloid, lasiocarpine. Cancer Letters 231, 138–145.
Yan, C.C., Huxtable, R.J., 1995. The effect of the pyrrolizidine alkaloids,monocrotaline and tricodesmine, on tissue pyrrole binding and glutathionemetabolism in the rat. Toxicon 33, 627–634.