gfap-gfp neural progenitors are antigenically homogeneous and anchored in their enclosed mosaic...
TRANSCRIPT

GFAP-GFP Neural Progenitors Are AntigenicallyHomogeneous and Anchored in TheirEnclosed Mosaic Niche
JEAN-CLAUDE PLATEL,1,2 VALERIE GORDON,1,2 TRISTAN HEINTZ,1,2 AND ANG�ELIQUE BORDEY1,2*1Department of Neurosurgery, Yale University, New Haven, Connecticut 06520 80822Department of Cellular and Molecular Physiology, Yale University, New Haven, Connecticut 06520 8082
KEY WORDSastrocytes; stem cells; neurogenesis; proliferation; migra-tion; transgenic; neuroblast; epidermal growth factor;Olig2; NG2; Mcm2
ABSTRACTStudy of the different stages of postnatal neurogenesisrelies on using antigenic markers and transgenic mice. Inparticular, neural stem cells that express GFAP are studiedusing mice expressing GFP under the human GFAP pro-moter (GFAP-GFP). However, it remains unclear whetherGFP and the commonly used progenitor markers label dif-ferent cell populations in the neurogenic subventricularzone (SVZ) and its rostral extension into the olfactory bulb(i.e. rostral migratory stream, RMS). Here, we found thatall GFP-fluorescent cells express GFAP, the radial gliamarker brain lipid-binding protein (BLBP), Lewis X (LeX),and the astrocytic marker GLAST. Faint GFP fluorescencecould be detected in a few cells expressing EGF receptors(EGFRs), Olig2, or S100, suggesting that GFAP-GFP cellsgenerate these diverse cell types. GFP-fluorescent cellswere slowly cycling, as shown by their long-term retentionof BrdU, and less than 10% expressed the proliferativemarkers Ki67 and Mcm2. The majority of EGFR-expressingcells and Olig2-expressing cells were cycling. NG2 andEGFR identified distinct progenitor populations while Olig2labeled a subset of EGFR-expressing cells. The entire neu-rogenic zone contained a mosaic of different cell types andwas ensheathed by processes of GFAP-expressing cells andNG2 cells. Finally, using time-lapse imaging in acute slices,we show that GFP-fluorescent cells are stationary withinthe SVZ. Our findings collectively highlight the cellularmosaic of the neurogenic niche, show that the slowly-cy-cling GFAP-expressing cells are stationary and generatedistinct intermediate progenitors. VVC 2008 Wiley-Liss, Inc.
INTRODUCTION
The past decade has seen an exponential increase inthe number of studies on postnatal neurogenesis in theforebrain, including the neurogenic subventricular zone(SVZ) along the lateral ventricle and the subgranularzone of the hippocampal dentate gyrus. The majority ofthese studies rely on the use of antigenic markers and/or transgenic mice to identify the cell populations at dif-ferent phases of cell development. However, it remainsunclear whether the commonly used markers and trans-genic fluorescent proteins label single or multiple cell
populations in the SVZ and its rostral extension into theolfactory bulb. Neural stem cells (also called neural pro-genitors) are thought to be a subpopulation of glialfibrillary acidic protein (GFAP)-expressing cells (alsocalled type B cells) that asymmetrically divide intotransit amplifying progenitors (type C cells), whichthemselves generate neuroblasts (Doetsch et al., 1999,2002; for review see Bordey, 2006; Chan et al., 2004;Lledo et al., 2006). Once born in the SVZ, neuroblastsmigrate along the RMS to reach the olfactory bulb (OB)where they differentiate into interneurons.
Immunostaining for the GFAP antigen is a commonmethod to examine whether GFAP-expressing cells co-express antigenic markers for different SVZ cell popula-tions and for proliferation. However, interpretation ofco-staining data with GFAP is difficult due to the fila-mentous appearance of the GFAP staining, which under-estimates the cell morphology. This is particularly truewhen co-staining for GFAP and the various proliferationmarkers (e.g. 5-bromo-20-deoxyuridine BrdU, minichro-mosome maintenance protein 2 Mcm2, and Ki67), whichresult in a nuclear stain. As an alternative to GFAP, weand others have used another astrocytic marker, gluta-mate-aspartate transporter (GLAST), which decoratesthe surface of GFAP-expressing cells (Bolteus and Bor-dey, 2004; Braun et al., 2003; Liu et al., 2006). Trans-genic GFAP-GFP mice have also been used to study neu-rogenesis (Liu et al., 2005, 2006). However, a thoroughexamination of the expression pattern of GFP, GFAP,GLAST, and other markers has not been performed. Theepidermal growth factor receptor (EGFR) is routinelyused to label transit amplifying progenitors (Doetschet al., 2002), while the microtubule-associated proteindoublecortin (DCX) exclusively labels neuroblasts in theSVZ and RMS in non-injured tissue (Gleeson et al.,1999; Nacher et al., 2001, Parent et al., 2006). Consider-
Additional Supporting Information may be found in the online version of thearticle.
Grant sponsor: National Institutes of Health; Grant numbers: NS048256,DC007681; Grant sponsor: Yale Brown-Coxe fellowship.
*Correspondence to: Ang�elique Bordey, Department of Neurosurgery, Yale Uni-versity School of Medicine, 333 Cedar Street, FMB 422, New Haven, CT 06520-8082, USA. E-mail: [email protected]
Received 13 March 2008; Accepted 12 June 2008
DOI 10.1002/glia.20735
Published online 25 July 2008 in Wiley InterScience (www.interscience.wiley.com).
GLIA 57:66–78 (2009)
VVC 2008 Wiley-Liss, Inc.

ing that the half-life of GFP is 1 day, it is possible thatGFP persists at low levels in the first progeny of GFAP-expressing progenitors (i.e. EGFR-expressing cells), butthis has not been examined. Both the Lewis X (LeX)antigen (also called CD15) and Mcm2 have been shownto label SVZ cells displaying stem cell characteristics(Capela and Temple, 2002; Imura et al., 2006; Maslovet al., 2004). It has also been shown that neuro-glial pro-teoglycan 2 (NG2)-expressing cells (also called oligoden-drocyte precursor cells) act as transit amplifying progen-itors and Olig2-expressing progenitors act as gliogenicprogenitors in neonatal mice (Aguirre et al., 2004; Mar-shall et al., 2005). Collectively, a thorough examinationof the cellular expression of these markers in GFAP-GFP mice remains to be done in both the SVZ and RMS.
Here, we examined the expression patterns of differ-ent markers, some of which are listed above, in GFAP-GFP mice. We confirmed that GFP-fluorescent cellsexpress GFAP. Nevertheless, faint GFP is visible insome EGFR- and Olig2-positive cells, suggesting thatthey are generated from GFAP-GFP cells. While GFP-proliferative cells are slowly cycling and 7% are activelycycling at any given time, 90% of EGFR-expressing cellsare proliferating. LeX, GLAST, and the radial gliamarker BLBP (Hartfuss et al., 2001) label GFP-fluores-cent cells and some EGFR-expressing cells. The SVZ is acellular mosaic containing populations of GFAP-, DCX-,S100-, NG2-, Olig2-, and EGFR-expressing cells. Thesedifferent populations, some of which display small over-lap (e.g. Olig2 and EGFR cells, or NG2 and S100), havedifferent proliferative potentials. GFAP-GFP mice thusrepresent a valuable tool to study GFAP-expressing pro-genitors, and in particular their behavior in acute slices.We provide additional evidence using time lapse imagingthat GFP-fluorescent cells are stationary, as expected fora vital population of self-renewing neural progenitors.
MATERIALS AND METHODSAnimals
Staining was performed in FVB/N (Charles River,USA) or GFAP-GFP mice (FVB/N-Tg(GFAPGFP)14Mes/J, donating investigator Dr. Albee Messing, JacksonLabs, USA) that were 23–60 days old. For Fig. 1, 100-lm-thick sections were prepared from paraformaldehydefixed brains from P25 transgenic mice expressing GFPunder the promoter of doublecortin (DCX, FVB/N back-ground, a kind gift from Dr. R. Miller, University of Chi-cago, originally Gensat, USA).
Immunohistochemistry and Proliferation
Mice were deeply anesthetized with pentobarbital (50mg/kg). The brain was then quickly removed and placedin 4% paraformaldehyde overnight at 4�C, then washed in1X PBS. The next day, 100-lm-thick slices were preparedusing a vibratome (Leica VTS 1000, Germany). Free-float-ing sections were blocked in Tris Buffered Saline (TBS)
containing 0.1% Triton X100 1 0.1% Tween-20 1 2%BSA, and incubated in the primary antibodies (Table 1)overnight at 4�C. After several washes in TBS containing0.1% Tween-20, slices were incubated with the appropri-ate secondary antibody (Alexa Fluor series at 1:1000,Invitrogen, USA, or Cyanine series at 1:500, JacksonLabs) for 1 hr at room temperature. For labeling slowlydividing cells, 50 lg/g of BrdU was intraperitoneallyinjected once a day for 5 days in P30 GFAP-GFP mice.Three weeks following the last BrdU injection, the brainwas fixed and 100-lm-thick sections prepared. Stainingfor BrdU and other markers as described above was pre-ceded by a treatment with 2N HCl for 45 min at roomtemperature. Because GFP fluorescence was quenched,we stained for GFP when co-staining for BrdU. Appropri-ate control staining such as the absence of the primaryantibodies was routinely performed (data not shown).
Staining was replicated at least in 4–5 slices from threedifferent mice. Z-stacks (ranging in thickness from 5 to20 lm) containing 5–10 Z-sections spaced by 1–2 lm wereacquired on a laser-scanning confocal microscope(Olympus FluoView 1000) with a 203 dry objective (N.A.0.75) or a 60x oil objective (N.A. 1.42). Each Z-stackobtained with the 203 objective contained the entiredorso-lateral SVZ. The number of proliferative cells (Ki67-negative and Mcm2-positive) was quantified in each Z-sec-tion. Images were analyzed using Imaris 4.0 (BitplaneAG, Switzerland) and reconstructed in ImageJ 1.39t (Free-ware, Wayne Rasband, NIH, USA) and Photoshop CS3
Fig. 1. Location of the SVZ and RMS. (A) Image of a sagittal sectionof a mouse brain from a postnatal day 25 DCX-GFP mouse. The imagewas reconstructed from 42 photographs taken with a 10x objective. Thewhite arrowhead points to the dorso-lateral SVZ. The arrow points tothe beginning of the RMS. (B) Images of coronal sections from a differ-ent brain at four different rostro-caudal levels shown by white lines onthe sagittal section in (A). Images were reconstructed from 1 to 4 photo-graphs of each coronal section. The lateral ventricle (LV), althoughsmall, is still visible in section number 3 from left. The white arrowspoint to the RMS or lateral ventricle. [Color figure can be viewed in theonline issue, which is available at www.interscience.wiley.com.]
67GFAP-GFP PROGENITORS STATIONED IN THEIR MOSAIC NICHE
GLIA

(Adobe, USA). To analyze the colocalization of the differentmarkers, the maximum intensity projection was firstobtained using Imaris to have a first overview of the colo-calized marker. Then, we consecutively observed each indi-vidual Z-section to confirm the co-localization.
Acute Slice Preparation and Time-LapseConfocal Imaging
Acute sagittal brain slices (300-lm-thick) containingthe SVZ and/or RMS from P23-29 GFAP-GFP mice wereprepared as we previously described (Bolteus and Bordey,2004; Bolteus et al., 2005) and in accordance with a proto-col approved by the Yale Animal Care and Use Commit-tee. Mice were deeply anesthetized and the brainsremoved. Slices were prepared with a Leica vibratomeand maintained in DMEM high glucose (Invitrogen, USA)at pH 7.4 when equilibrated with 95% O2/5% CO2 for1 hour before imaging. Slices were loaded 30 min in10 lM of SYTO59 (Invitrogen, USA,) at room temperatureand then washed for 10 min in DMEM. Slices were thenplaced in a flow-through chamber, held in position by anylon mesh glued to a U-shaped platinum wire and con-tinuously superfused with oxygenated DMEM. The solu-tion was stably heated to 37�C by using a series 20 plat-form and a SF-28 in-line heater (Warner Instruments,USA). Experiments were performed on an upright Olym-pus BX61WI microscope equipped with an Olympus Fluo-View 1000 confocal microscope and a water-immersionNomarski phase-contrast and fluorescence 60x objective(N.A. 0.9). Z-section images (20–30 sections spaced by2 lm) were acquired every 5–15 min with FluoView acqui-sition software and reconstructed with ImageJ off-line.
Data are expressed as mean 6 standard error. Statisti-cal analysis used a two-tailed t-test except where noted.
RESULTSThe Dorso-Lateral SVZ
The dorso-lateral SVZ could be mistaken for the RMSin sagittal sections because of the complex shape of thelateral ventricle, as illustrated in Fig. 1. In a sagittal
section from a DCX-GFP mouse (Fig. 1A), a stream ofGFP-fluorescent neuroblasts is visible under the corpuscallosum that was previously termed RMS by us andothers. However, using serial coronal sections (Fig. 1B)or whole mounts of the lateral ventricle (Supp. Info.Fig. 1, but see also Sawamoto et al., 2006), it becomesclear that this stream of cells corresponds to the dorso-lateral SVZ, which spreads medially (Fig. 1A, arrow-head). The chains of neuroblasts merge rostrally, result-ing in an enlarged RMS (Fig. 1A, arrow). Here, weexamined the antigenic properties, as well as the prolif-erative and migratory behavior of GFP-fluorescent cellsin the dorso-lateral SVZ (Bregma anteroposterior 0-1)and the RMS-OB from GFAP-GFP mice.
To define the limits of the dorso-lateral SVZ at the cel-lular level, we co-stained for DCX and nestin in GFAP-GFP mice (Fig. 2A,B). DCX immunostaining defines thecore of the SVZ while nestin staining marks the entireSVZ, including DCX-positive cells and GFP-fluorescentcells, as previously reported (Doetsch et al., 1997). SomeGFP-fluorescent cells are visible among the DCX core,where each individual GFP-fluorescent cell is known toensheath individual clusters (or chains) of neuroblasts(Doetsch et al., 1997). GFP-fluorescent cells are alsofound at the edge between the DCX-positive cells andthe striatum (arrows in Fig. 2B). These cells are alsonestin positive and are thus part of the SVZ.
GFP-Fluorescent Cells in the SVZ Comprise aSlowly Dividing Population of GFAP-Positive Cells
that Generate EGFR-Expressing Cells
To determine whether GFP fluorescence correspondsto GFAP expression in the postnatal SVZ, we performedimmunostaining for GFAP in coronal sections from post-natal day (P) 23 to P 60 GFAP-GFP mice (Fig. 3A–C).The filamentous nature of GFAP staining makes it diffi-cult to detect co-labeling and unambiguously ascertainwhether GFP and GFAP expression overlaps. Neverthe-less, we placed arrows to indicate the most obvious co-labeling between GFP and GFAP (Fig. 3C, whitearrows). We also noted that ependymal cells were not
TABLE 1. List of Antibodies with Their Concentrations
Antibody Species Concentration Source Catalog number
BLBP Rabbit 1/500 Chemicon AB9558BrdU-Alexa488 Mouse 1/200 Molecular probes A21303BrdU-Alexa647 Mouse 1/200 Molecular probes A21305DCX Goat 1/100 Santa Cruz SC 8066DCX Rabbit 1/100 Santa Cruz SC28939EGFR Sheep 1/100 Upstate antibodies 06-129GFAP-Cy3 Mouse IgG1 1/500 Sigma C9205GFAP Rabbit 1/1000 Dako Z0334Glast Guinea Pig 1/1000 Chemicon AB1782Ki67 Rabbit 1/200 Vector laboratories T0423Lex Mouse IgM 1/100 BD biosciences 55 9045Mcm2 Goat 1/500 Santa Cruz SC 9839Nestin Chicken 1/250 Novus NB 100-1604NG2 Rabbit 1/500 Chemicon AB5320Olig2 Rabbit 1/500 Santa Cruz SC 48817S100 Rabbit 1/200 Sigma S2644S100 Mouse 1/200 Chemicon MAB079-1
68 PLATEL ET AL.
GLIA
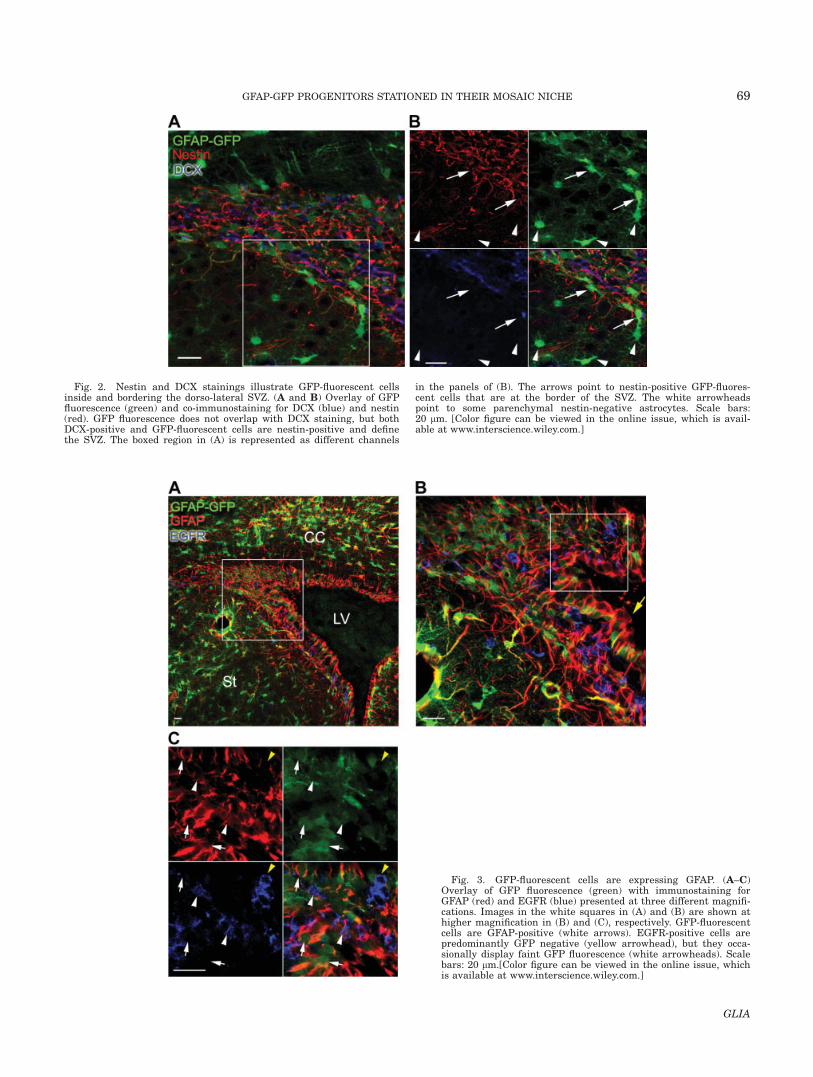
Fig. 2. Nestin and DCX stainings illustrate GFP-fluorescent cellsinside and bordering the dorso-lateral SVZ. (A and B) Overlay of GFPfluorescence (green) and co-immunostaining for DCX (blue) and nestin(red). GFP fluorescence does not overlap with DCX staining, but bothDCX-positive and GFP-fluorescent cells are nestin-positive and definethe SVZ. The boxed region in (A) is represented as different channels
in the panels of (B). The arrows point to nestin-positive GFP-fluores-cent cells that are at the border of the SVZ. The white arrowheadspoint to some parenchymal nestin-negative astrocytes. Scale bars:20 lm. [Color figure can be viewed in the online issue, which is avail-able at www.interscience.wiley.com.]
Fig. 3. GFP-fluorescent cells are expressing GFAP. (A–C)Overlay of GFP fluorescence (green) with immunostaining forGFAP (red) and EGFR (blue) presented at three different magnifi-cations. Images in the white squares in (A) and (B) are shown athigher magnification in (B) and (C), respectively. GFP-fluorescentcells are GFAP-positive (white arrows). EGFR-positive cells arepredominantly GFP negative (yellow arrowhead), but they occa-sionally display faint GFP fluorescence (white arrowheads). Scalebars: 20 lm.[Color figure can be viewed in the online issue, whichis available at www.interscience.wiley.com.]
69GFAP-GFP PROGENITORS STATIONED IN THEIR MOSAIC NICHE
GLIA
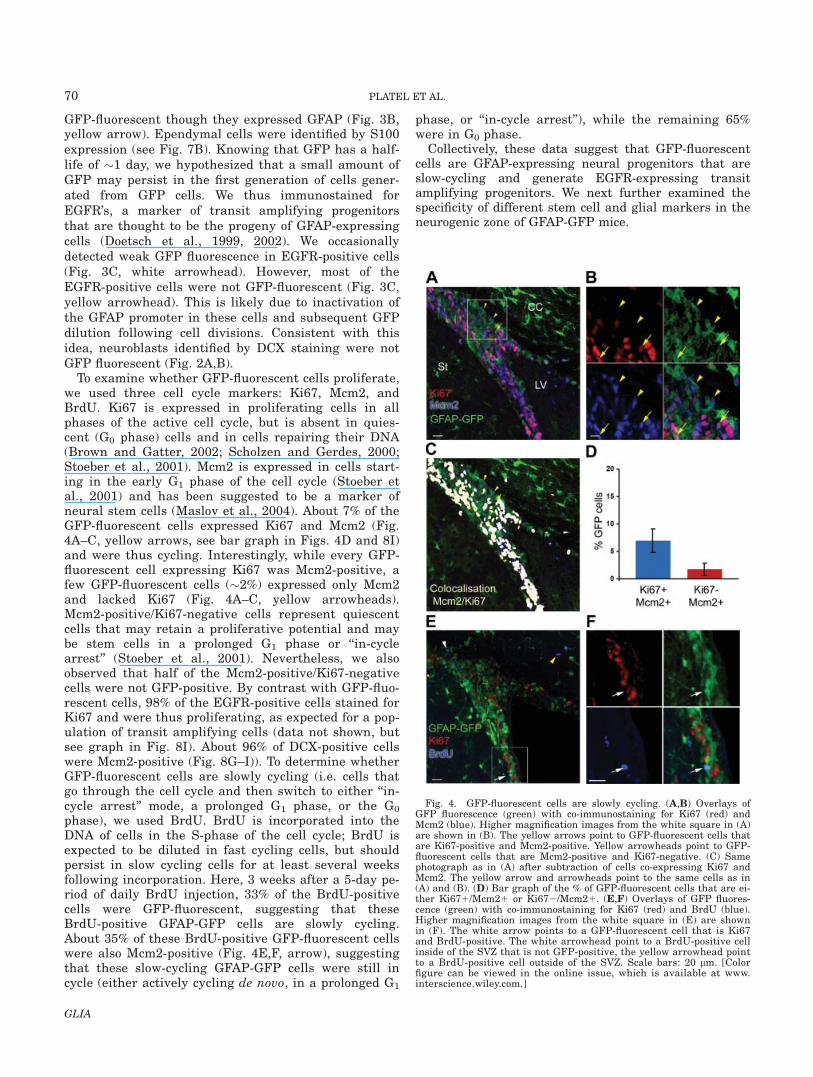
GFP-fluorescent though they expressed GFAP (Fig. 3B,yellow arrow). Ependymal cells were identified by S100expression (see Fig. 7B). Knowing that GFP has a half-life of �1 day, we hypothesized that a small amount ofGFP may persist in the first generation of cells gener-ated from GFP cells. We thus immunostained forEGFR’s, a marker of transit amplifying progenitorsthat are thought to be the progeny of GFAP-expressingcells (Doetsch et al., 1999, 2002). We occasionallydetected weak GFP fluorescence in EGFR-positive cells(Fig. 3C, white arrowhead). However, most of theEGFR-positive cells were not GFP-fluorescent (Fig. 3C,yellow arrowhead). This is likely due to inactivation ofthe GFAP promoter in these cells and subsequent GFPdilution following cell divisions. Consistent with thisidea, neuroblasts identified by DCX staining were notGFP fluorescent (Fig. 2A,B).
To examine whether GFP-fluorescent cells proliferate,we used three cell cycle markers: Ki67, Mcm2, andBrdU. Ki67 is expressed in proliferating cells in allphases of the active cell cycle, but is absent in quies-cent (G0 phase) cells and in cells repairing their DNA(Brown and Gatter, 2002; Scholzen and Gerdes, 2000;Stoeber et al., 2001). Mcm2 is expressed in cells start-ing in the early G1 phase of the cell cycle (Stoeber etal., 2001) and has been suggested to be a marker ofneural stem cells (Maslov et al., 2004). About 7% of theGFP-fluorescent cells expressed Ki67 and Mcm2 (Fig.4A–C, yellow arrows, see bar graph in Figs. 4D and 8I)and were thus cycling. Interestingly, while every GFP-fluorescent cell expressing Ki67 was Mcm2-positive, afew GFP-fluorescent cells (�2%) expressed only Mcm2and lacked Ki67 (Fig. 4A–C, yellow arrowheads).Mcm2-positive/Ki67-negative cells represent quiescentcells that may retain a proliferative potential and maybe stem cells in a prolonged G1 phase or ‘‘in-cyclearrest’’ (Stoeber et al., 2001). Nevertheless, we alsoobserved that half of the Mcm2-positive/Ki67-negativecells were not GFP-positive. By contrast with GFP-fluo-rescent cells, 98% of the EGFR-positive cells stained forKi67 and were thus proliferating, as expected for a pop-ulation of transit amplifying cells (data not shown, butsee graph in Fig. 8I). About 96% of DCX-positive cellswere Mcm2-positive (Fig. 8G–I)). To determine whetherGFP-fluorescent cells are slowly cycling (i.e. cells thatgo through the cell cycle and then switch to either ‘‘in-cycle arrest’’ mode, a prolonged G1 phase, or the G0
phase), we used BrdU. BrdU is incorporated into theDNA of cells in the S-phase of the cell cycle; BrdU isexpected to be diluted in fast cycling cells, but shouldpersist in slow cycling cells for at least several weeksfollowing incorporation. Here, 3 weeks after a 5-day pe-riod of daily BrdU injection, 33% of the BrdU-positivecells were GFP-fluorescent, suggesting that theseBrdU-positive GFAP-GFP cells are slowly cycling.About 35% of these BrdU-positive GFP-fluorescent cellswere also Mcm2-positive (Fig. 4E,F, arrow), suggestingthat these slow-cycling GFAP-GFP cells were still incycle (either actively cycling de novo, in a prolonged G1
phase, or ‘‘in-cycle arrest’’), while the remaining 65%were in G0 phase.
Collectively, these data suggest that GFP-fluorescentcells are GFAP-expressing neural progenitors that areslow-cycling and generate EGFR-expressing transitamplifying progenitors. We next further examined thespecificity of different stem cell and glial markers in theneurogenic zone of GFAP-GFP mice.
Fig. 4. GFP-fluorescent cells are slowly cycling. (A,B) Overlays ofGFP fluorescence (green) with co-immunostaining for Ki67 (red) andMcm2 (blue). Higher magnification images from the white square in (A)are shown in (B). The yellow arrows point to GFP-fluorescent cells thatare Ki67-positive and Mcm2-positive. Yellow arrowheads point to GFP-fluorescent cells that are Mcm2-positive and Ki67-negative. (C) Samephotograph as in (A) after subtraction of cells co-expressing Ki67 andMcm2. The yellow arrow and arrowheads point to the same cells as in(A) and (B). (D) Bar graph of the % of GFP-fluorescent cells that are ei-ther Ki671/Mcm21 or Ki672/Mcm21. (E,F) Overlays of GFP fluores-cence (green) with co-immunostaining for Ki67 (red) and BrdU (blue).Higher magnification images from the white square in (E) are shownin (F). The white arrow points to a GFP-fluorescent cell that is Ki67and BrdU-positive. The white arrowhead point to a BrdU-positive cellinside of the SVZ that is not GFP-positive, the yellow arrowhead pointto a BrdU-positive cell outside of the SVZ. Scale bars: 20 lm. [Colorfigure can be viewed in the online issue, which is available at www.interscience.wiley.com.]
70 PLATEL ET AL.
GLIA

Both LeX and BLBP Are Expressed, ThoughNon-Exclusively, in GFP-Fluorescent Cells
The LeX antigen, which is the trisaccharide 3-fucosyl-N-acetyllactosamine (Gooi et al., 1981), also known asSSEA-1 (stage-specific embryonic antigen 1) or CD15(leucocyte cluster of differentiation 15), is expressed inSVZ cells displaying stem cell characteristics (Capelaand Temple, 2002; Imura et al., 2006). However, LeX isalso expressed by various cell types in the SVZ and inother locations (Bartsch and Mai, 1991; Capela andTemple, 2002). All GFP-fluorescent cells in the SVZwere LeX-positive (Supp. Info. Fig. 2A,B, yellow arrow-head). Similarly, mature astrocytes outside of the SVZstained strongly for LeX (Supp. Info. Fig. 2B, whitearrow). All of the EGFR-positive cells expressed LeX(Supp. Info. Fig. 2A,B, white arrowhead). The pattern ofLeX staining is widespread and punctate, and is there-fore not recommended for co-localization studies usingnuclear stains.
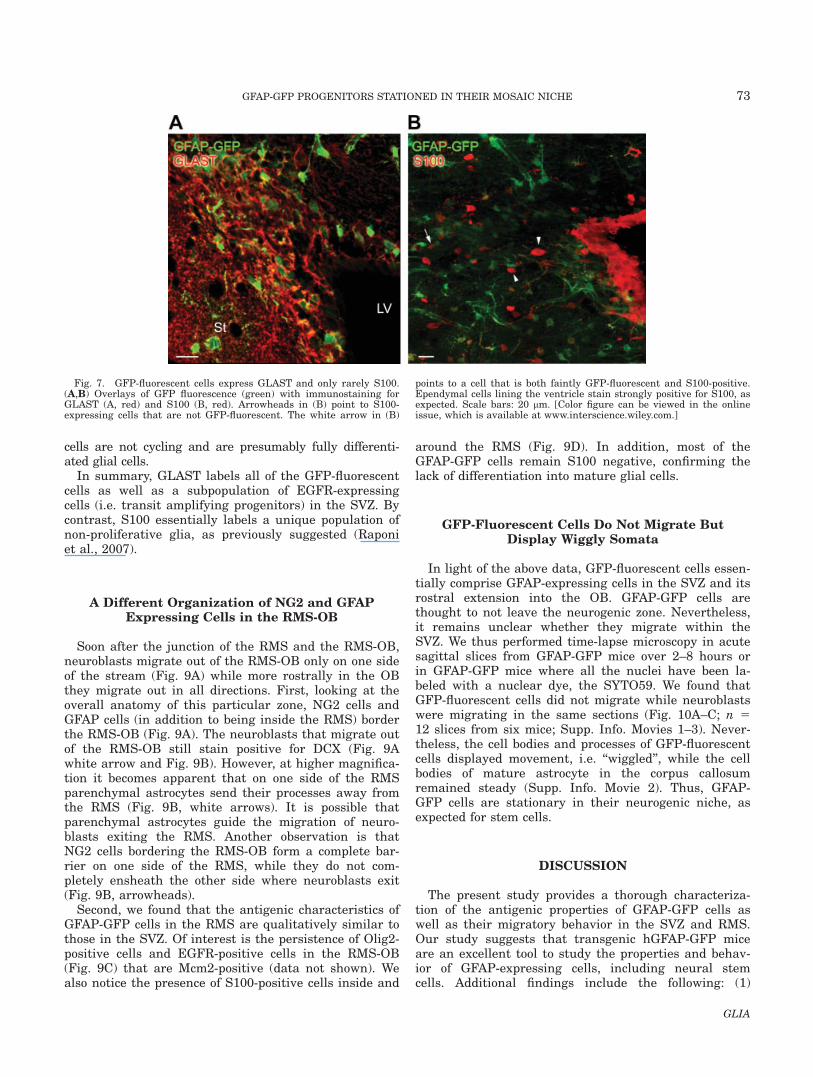
GFAP-expressing cells of the SVZ derive from radialglia and display many of their features (Bolteus et al.,2005; Liu et al., 2006). We thus immunostained forBLBP, a marker of radial glia (Hartfuss et al., 2001)shown to be in GFAP-expressing cells in the SVZ (Per-etto et al., 2005). All of the GFP-fluorescent cells includ-ing EGFR-positive cells were BLBP-positive (Fig. 5A,B,white arrow and white arrowhead, respectively). Sur-prisingly, a few BLBP-positive cells expressed neitherGFP nor EGFR (Fig. 5B, yellow arrowhead).
Collectively, these results show that BLBP and LeXare not exclusive markers of GFAP-expressing cells inthe SVZ, as they label both GFAP-GFP cells and transitamplifying progenitors. Nevertheless, BLBP may be ad-vantageous for labeling proliferative cells belonging tothese two populations due to its diffuse staining overthe cell body, permitting complete overlap with the nu-clear stains from proliferative markers.
NG2 Cells Do Not Express Either GFP or EGFR,While a Subset of Olig2 Cells Are Faintly
GFP-Fluorescent and Proliferating
NG2, a member of the chondroitin sulfate proteogly-can family, is a well-known marker of oligodendrocyteprecursors (Nishiyama et al., 1999). In addition, it hasbeen shown to label bipotential progenitors in the neo-natal SVZ that co-label with EGFR’s, but not with GFAP(Aguirre and Gallo, 2004; Aguirre et al., 2004). Here,most of the NG2 cells were found surrounding the SVZ,which was defined by nestin staining (Supp. Info. Fig.5A,B). NG2 cells at the border of the SVZ also sendprocesses inside this zone (Fig. 8C and arrowhead on8F). Nevertheless, a few NG2 cells with large arboriza-tion (2-4 cells per 20-lm-optical sections) were alsofound inside the SVZ (Fig. 6A,B). A few NG2 cells werefaintly GFP-fluorescent (Fig. 6B, white arrow) and didnot co-label with EGFR-positive cells (Supp. Info. Fig.3A). NG2 also stains pericytes that can be distinguished
from NG2 cells by their simple morphology and attach-ment to blood vessels (Supp. Info. Fig. 3A, white arrow).In this latter figure, EGFR-expressing cells are oftenfound in close proximity to blood vessels. NG2 cells didnot express DCX and were generally GLAST-negative(Supp. Info. Fig. 3B). But NG2 cells occasionally co-stained for S100 (Supp. Info. Fig. 3C). Finally, we foundthat processes of NG2 cells often surround cycling cellsbut only 18% of NG2 cells themselves stained positivefor Mcm2 (Fig. 8C,F,I).
The basic helix-loop-helix transcription factor Olig2 isexpressed in SVZ gliogenic progenitors (Marshall et al.,2005; Menn et al., 2006). Although Olig2-expressingcells can generate astrocytes in neonates (Marshall etal., 2005), they are not supposed to express GFAP, i.e.GFP in our transgenic mice. The majority of the Olig2-positive cells in the SVZ and surrounding tissue werenot GFP-fluorescent (Fig. 6C,D, arrowheads). However,a few Olig2-positive cells in the SVZ displayed faintGFP fluorescence (Fig. 6D, arrow), suggesting that theymay have been directly generated from GFAP-express-ing cells. We noticed that the GFP-fluorescent Olig2-pos-itive cells did not express EGFR while some Olig2-posi-tive cells with no GFP-fluorescence were EGFR-positive(Supp. Info. Fig. 4A,B), suggesting that they may consti-tute a population of transit amplifying progenitors.Finally, 57% of the Olig2-positive cells were Mcm2-posi-tive in the SVZ (Fig. 8B, arrowheads in E, and I), sug-gesting that they are actively dividing.
Glial Markers GLAST and S100 Are NotExclusively Expressed in GFP-Fluorescent Cells
We and others have used GLAST as a marker ofGFAP-GFP cells in the SVZ (Bolteus and Bordey, 2004;Braun et al., 2003; Liu et al., 2006), but it remainsunclear whether GLAST labels only GFP-fluorescentcells. We confirmed that all of the GFP-fluorescent cellswere GLAST-positive (Fig. 7A). However, GLAST wasalso expressed in some of the EGFR-expressing cells(Supp. Info. Fig. 3D, arrowhead). Another commonlyused astrocytic marker is S100, which belongs to theS100 family of EF-band calcium binding proteins(Baudier et al., 1986). However, S100 is also found inNG2 cells in the parenchyma (Deloulme et al., 2004;Hachem et al., 2005), but was not found in the lateralSVZ (Raponi et al., 2007). A few cells expressing S100were found in the dorso-lateral portion of the SVZ in ev-ery section examined (Fig. 7B, see Supp. Info. Fig. 3C).Although the majority of the S100-positive-cells werenot GFP-positive (Fig. 7B, arrowhead), some cells werefaintly positive for both S100 and GFP (Fig. 7B, arrow).We also observed that some S100-positive cells borderingthe SVZ co-stained for NG2 (see Supp. Info. Fig. 3C).Finally, we found that 13% of S100-positive cells wereMcm2-positive. These cells also faintly expressed GFP(Fig. 8A,D, white arrowheads), suggesting that they arederived from GFAP-GFP cells. Thus, most S100-expressing
71GFAP-GFP PROGENITORS STATIONED IN THEIR MOSAIC NICHE
GLIA
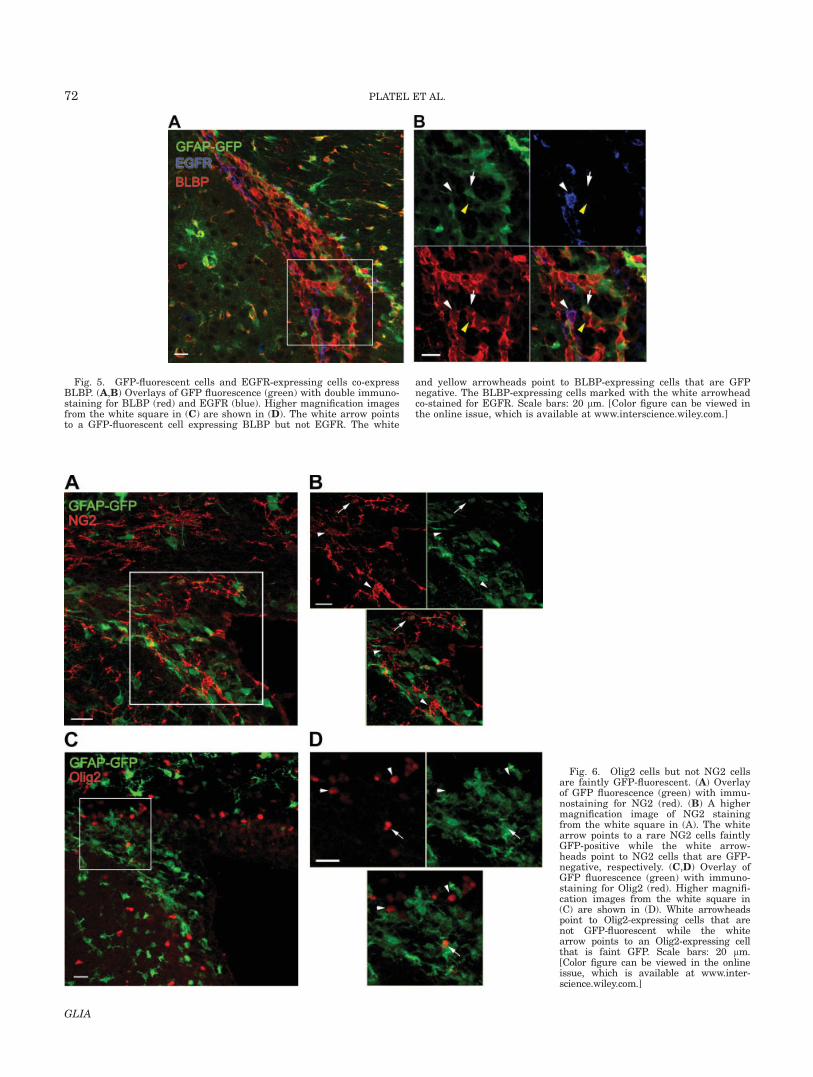
Fig. 6. Olig2 cells but not NG2 cellsare faintly GFP-fluorescent. (A) Overlayof GFP fluorescence (green) with immu-nostaining for NG2 (red). (B) A highermagnification image of NG2 stainingfrom the white square in (A). The whitearrow points to a rare NG2 cells faintlyGFP-positive while the white arrow-heads point to NG2 cells that are GFP-negative, respectively. (C,D) Overlay ofGFP fluorescence (green) with immuno-staining for Olig2 (red). Higher magnifi-cation images from the white square in(C) are shown in (D). White arrowheadspoint to Olig2-expressing cells that arenot GFP-fluorescent while the whitearrow points to an Olig2-expressing cellthat is faint GFP. Scale bars: 20 lm.[Color figure can be viewed in the onlineissue, which is available at www.inter-science.wiley.com.]
Fig. 5. GFP-fluorescent cells and EGFR-expressing cells co-expressBLBP. (A,B) Overlays of GFP fluorescence (green) with double immuno-staining for BLBP (red) and EGFR (blue). Higher magnification imagesfrom the white square in (C) are shown in (D). The white arrow pointsto a GFP-fluorescent cell expressing BLBP but not EGFR. The white
and yellow arrowheads point to BLBP-expressing cells that are GFPnegative. The BLBP-expressing cells marked with the white arrowheadco-stained for EGFR. Scale bars: 20 lm. [Color figure can be viewed inthe online issue, which is available at www.interscience.wiley.com.]
72 PLATEL ET AL.
GLIA
cells are not cycling and are presumably fully differenti-ated glial cells.
In summary, GLAST labels all of the GFP-fluorescentcells as well as a subpopulation of EGFR-expressingcells (i.e. transit amplifying progenitors) in the SVZ. Bycontrast, S100 essentially labels a unique population ofnon-proliferative glia, as previously suggested (Raponiet al., 2007).
A Different Organization of NG2 and GFAPExpressing Cells in the RMS-OB
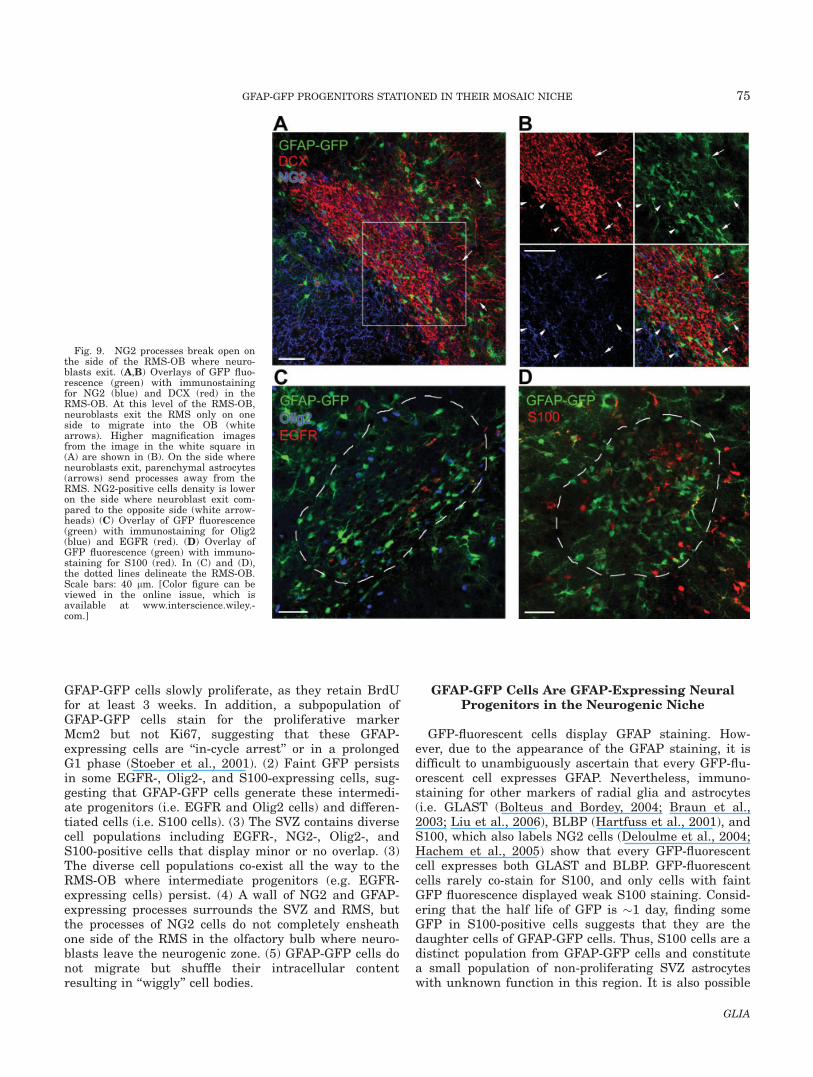
Soon after the junction of the RMS and the RMS-OB,neuroblasts migrate out of the RMS-OB only on one sideof the stream (Fig. 9A) while more rostrally in the OBthey migrate out in all directions. First, looking at theoverall anatomy of this particular zone, NG2 cells andGFAP cells (in addition to being inside the RMS) borderthe RMS-OB (Fig. 9A). The neuroblasts that migrate outof the RMS-OB still stain positive for DCX (Fig. 9Awhite arrow and Fig. 9B). However, at higher magnifica-tion it becomes apparent that on one side of the RMSparenchymal astrocytes send their processes away fromthe RMS (Fig. 9B, white arrows). It is possible thatparenchymal astrocytes guide the migration of neuro-blasts exiting the RMS. Another observation is thatNG2 cells bordering the RMS-OB form a complete bar-rier on one side of the RMS, while they do not com-pletely ensheath the other side where neuroblasts exit(Fig. 9B, arrowheads).
Second, we found that the antigenic characteristics ofGFAP-GFP cells in the RMS are qualitatively similar tothose in the SVZ. Of interest is the persistence of Olig2-positive cells and EGFR-positive cells in the RMS-OB(Fig. 9C) that are Mcm2-positive (data not shown). Wealso notice the presence of S100-positive cells inside and
around the RMS (Fig. 9D). In addition, most of theGFAP-GFP cells remain S100 negative, confirming thelack of differentiation into mature glial cells.
GFP-Fluorescent Cells Do Not Migrate ButDisplay Wiggly Somata
In light of the above data, GFP-fluorescent cells essen-tially comprise GFAP-expressing cells in the SVZ and itsrostral extension into the OB. GFAP-GFP cells arethought to not leave the neurogenic zone. Nevertheless,it remains unclear whether they migrate within theSVZ. We thus performed time-lapse microscopy in acutesagittal slices from GFAP-GFP mice over 2–8 hours orin GFAP-GFP mice where all the nuclei have been la-beled with a nuclear dye, the SYTO59. We found thatGFP-fluorescent cells did not migrate while neuroblastswere migrating in the same sections (Fig. 10A–C; n 512 slices from six mice; Supp. Info. Movies 1–3). Never-theless, the cell bodies and processes of GFP-fluorescentcells displayed movement, i.e. ‘‘wiggled’’, while the cellbodies of mature astrocyte in the corpus callosumremained steady (Supp. Info. Movie 2). Thus, GFAP-GFP cells are stationary in their neurogenic niche, asexpected for stem cells.
DISCUSSION
The present study provides a thorough characteriza-tion of the antigenic properties of GFAP-GFP cells aswell as their migratory behavior in the SVZ and RMS.Our study suggests that transgenic hGFAP-GFP miceare an excellent tool to study the properties and behav-ior of GFAP-expressing cells, including neural stemcells. Additional findings include the following: (1)
Fig. 7. GFP-fluorescent cells express GLAST and only rarely S100.(A,B) Overlays of GFP fluorescence (green) with immunostaining forGLAST (A, red) and S100 (B, red). Arrowheads in (B) point to S100-expressing cells that are not GFP-fluorescent. The white arrow in (B)
points to a cell that is both faintly GFP-fluorescent and S100-positive.Ependymal cells lining the ventricle stain strongly positive for S100, asexpected. Scale bars: 20 lm. [Color figure can be viewed in the onlineissue, which is available at www.interscience.wiley.com.]
73GFAP-GFP PROGENITORS STATIONED IN THEIR MOSAIC NICHE
GLIA
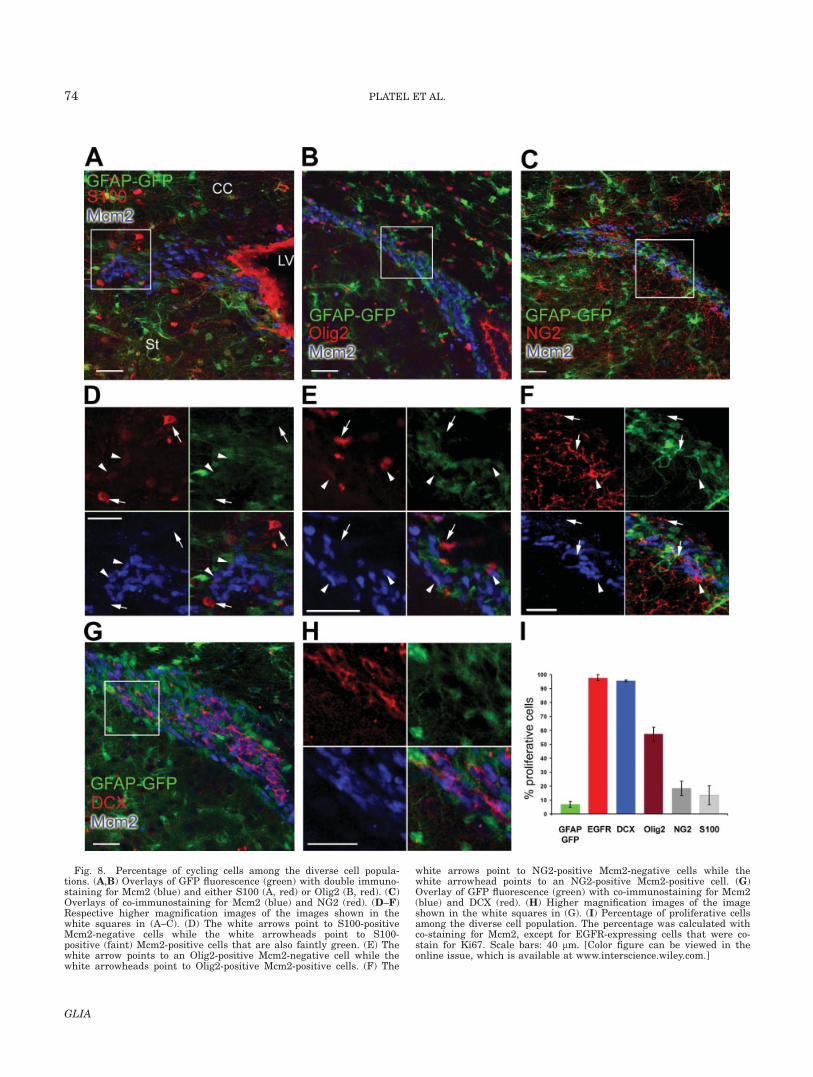
Fig. 8. Percentage of cycling cells among the diverse cell popula-tions. (A,B) Overlays of GFP fluorescence (green) with double immuno-staining for Mcm2 (blue) and either S100 (A, red) or Olig2 (B, red). (C)Overlays of co-immunostaining for Mcm2 (blue) and NG2 (red). (D–F)Respective higher magnification images of the images shown in thewhite squares in (A–C). (D) The white arrows point to S100-positiveMcm2-negative cells while the white arrowheads point to S100-positive (faint) Mcm2-positive cells that are also faintly green. (E) Thewhite arrow points to an Olig2-positive Mcm2-negative cell while thewhite arrowheads point to Olig2-positive Mcm2-positive cells. (F) The
white arrows point to NG2-positive Mcm2-negative cells while thewhite arrowhead points to an NG2-positive Mcm2-positive cell. (G)Overlay of GFP fluorescence (green) with co-immunostaining for Mcm2(blue) and DCX (red). (H) Higher magnification images of the imageshown in the white squares in (G). (I) Percentage of proliferative cellsamong the diverse cell population. The percentage was calculated withco-staining for Mcm2, except for EGFR-expressing cells that were co-stain for Ki67. Scale bars: 40 lm. [Color figure can be viewed in theonline issue, which is available at www.interscience.wiley.com.]
74 PLATEL ET AL.
GLIA
GFAP-GFP cells slowly proliferate, as they retain BrdUfor at least 3 weeks. In addition, a subpopulation ofGFAP-GFP cells stain for the proliferative markerMcm2 but not Ki67, suggesting that these GFAP-expressing cells are ‘‘in-cycle arrest’’ or in a prolongedG1 phase (Stoeber et al., 2001). (2) Faint GFP persistsin some EGFR-, Olig2-, and S100-expressing cells, sug-gesting that GFAP-GFP cells generate these intermedi-ate progenitors (i.e. EGFR and Olig2 cells) and differen-tiated cells (i.e. S100 cells). (3) The SVZ contains diversecell populations including EGFR-, NG2-, Olig2-, andS100-positive cells that display minor or no overlap. (3)The diverse cell populations co-exist all the way to theRMS-OB where intermediate progenitors (e.g. EGFR-expressing cells) persist. (4) A wall of NG2 and GFAP-expressing processes surrounds the SVZ and RMS, butthe processes of NG2 cells do not completely ensheathone side of the RMS in the olfactory bulb where neuro-blasts leave the neurogenic zone. (5) GFAP-GFP cells donot migrate but shuffle their intracellular contentresulting in ‘‘wiggly’’ cell bodies.
GFAP-GFP Cells Are GFAP-Expressing NeuralProgenitors in the Neurogenic Niche
GFP-fluorescent cells display GFAP staining. How-ever, due to the appearance of the GFAP staining, it isdifficult to unambiguously ascertain that every GFP-flu-orescent cell expresses GFAP. Nevertheless, immuno-staining for other markers of radial glia and astrocytes(i.e. GLAST (Bolteus and Bordey, 2004; Braun et al.,2003; Liu et al., 2006), BLBP (Hartfuss et al., 2001), andS100, which also labels NG2 cells (Deloulme et al., 2004;Hachem et al., 2005) show that every GFP-fluorescentcell expresses both GLAST and BLBP. GFP-fluorescentcells rarely co-stain for S100, and only cells with faintGFP fluorescence displayed weak S100 staining. Consid-ering that the half life of GFP is �1 day, finding someGFP in S100-positive cells suggests that they are thedaughter cells of GFAP-GFP cells. Thus, S100 cells are adistinct population from GFAP-GFP cells and constitutea small population of non-proliferating SVZ astrocyteswith unknown function in this region. It is also possible
Fig. 9. NG2 processes break open onthe side of the RMS-OB where neuro-blasts exit. (A,B) Overlays of GFP fluo-rescence (green) with immunostainingfor NG2 (blue) and DCX (red) in theRMS-OB. At this level of the RMS-OB,neuroblasts exit the RMS only on oneside to migrate into the OB (whitearrows). Higher magnification imagesfrom the image in the white square in(A) are shown in (B). On the side whereneuroblasts exit, parenchymal astrocytes(arrows) send processes away from theRMS. NG2-positive cells density is loweron the side where neuroblast exit com-pared to the opposite side (white arrow-heads) (C) Overlay of GFP fluorescence(green) with immunostaining for Olig2(blue) and EGFR (red). (D) Overlay ofGFP fluorescence (green) with immuno-staining for S100 (red). In (C) and (D),the dotted lines delineate the RMS-OB.Scale bars: 40 lm. [Color figure can beviewed in the online issue, which isavailable at www.interscience.wiley.-com.]
75GFAP-GFP PROGENITORS STATIONED IN THEIR MOSAIC NICHE
GLIA
that other SVZ cells do not express GFAP but belong tothe astrocyte family (see discussion by (Kimelberg,2004)). In particular, some SVZ cells express the radialglial cell marker BLBP but not GFP (see Fig. 5). Itremains to be determined whether these cells belong tothe same family of astrocytes. The schematic diagram inFig. 11A summarizes our findings.
GFP-fluorescent cells generate transit amplifying pro-genitors that are a minority of the cells in the SVZ andare thought to express EGFRs (Doetsch et al., 1997,2002). Consistent with GFAP-GFP cells being neuralstem cells, a minority of EGFR-expressing cells dis-played weak GFP fluorescence and expressed GLASTand BLBP. While the GLAST staining can be difficult tointerpret because it marks the cell surface, BLBP stain-ing is unambiguous and stains the entire cell. About 7%of GFAP-GFP cells were actively cycling, or proliferative,at the time of brain fixation as shown with Ki67 stain-ing. We also found that GFAP-GFP cells were slowly cy-cling as expected for neural progenitors. Interestingly,we found that some GFP-fluorescent cells express Mcm2but not Ki67. This again is in agreement with GFAP-GFP cells being neural stem cells because Mcm2-posi-tive/Ki67-negative cells have been suggested to be eitherin ‘‘cycle arrest’’ or in a prolonged G1 phase (Stoeberet al., 2001). It remains to be determined whether thesecells behave as neural stem cells in the SVZ andwhether this staining pattern could be used as a markerof GFP-fluorescent, quiescent stem cells. Similarly, it
remains unclear whether the non-cycling GFP-fluores-cent cells possess the ability to re-enter the cell cycleand behave as neural stem cells.
A Mosaic Niche with Diverse Cell Populations
Using markers of bipotential and glial progenitors(NG2, (Aguirre and Gallo, 2004; Aguirre et al., 2004))and Olig2 (Marshall et al., 2005; Menn et al., 2006),respectively), we found that GFP-fluorescent cells didnot overlap with NG2. A minority of Olig2 cells dis-played faint GFP fluorescence, suggesting that theseOlig2 cells were direct daughter cells of GFAP-GFP cells,as previously suggested (Menn et al., 2006). This findingindicates that a lineage restricted to glial progenitorsmay exist from GFAP-GFP cells that bypasses EGFR-expressing cells. However, some cells expressed bothEGFR and Olig2 but not GFP, suggesting that someEGFR-expressing transit amplifying progenitors, maygenerate Olig2-expressing cells (glia-restricted progeni-tors) in addition to DCX-expressing neuroblasts. Lineagestudies would be required to definitively addresswhether two distinct lineages exist from GFAP-GFPcells.
The majority of EGFR-expressing cells (�98%) andOlig-2 expressing cells (57%) were cycling, whileonly 19% of NG2 cells both inside and surrounding theSVZ were cycling. Normalized to the number of Mcm2-
Fig. 10. GFP-fluorescent cells are anchored in the SVZ, but wiggletheir somata and processes. (A) Photographs of GFP-fluorescent cells inthe RMS (prior to the entry into the OB). (B) Higher magnification pho-tographs of GFP-fluorescent cells (top panels) shown in (A) and overlayof GFP fluorescence onto DIC image (bottom panels) taken from a time-lapse imaging movie at the different time-points indicated on theimages. Cells were imaged in an acute sagittal slice from a P29 GFAP-
GFP mouse. Images were acquired every 5 min for 1.5 hr. (C) Photo-graphs of GFP-fluorescent cells in the RMS overlaid with SYTO59nucleic acid dye staining. The slice was recorded for 1 hour. The arrow-head indicate a cell (probably a neuroblast) moving next to a stationaryGFP-positive astrocyte. Scale bars: (A) 30 lm, (B) 10 lm, (C) 20 lm.[Color figure can be viewed in the online issue, which is available atwww.interscience.wiley.com.]
76 PLATEL ET AL.
GLIA
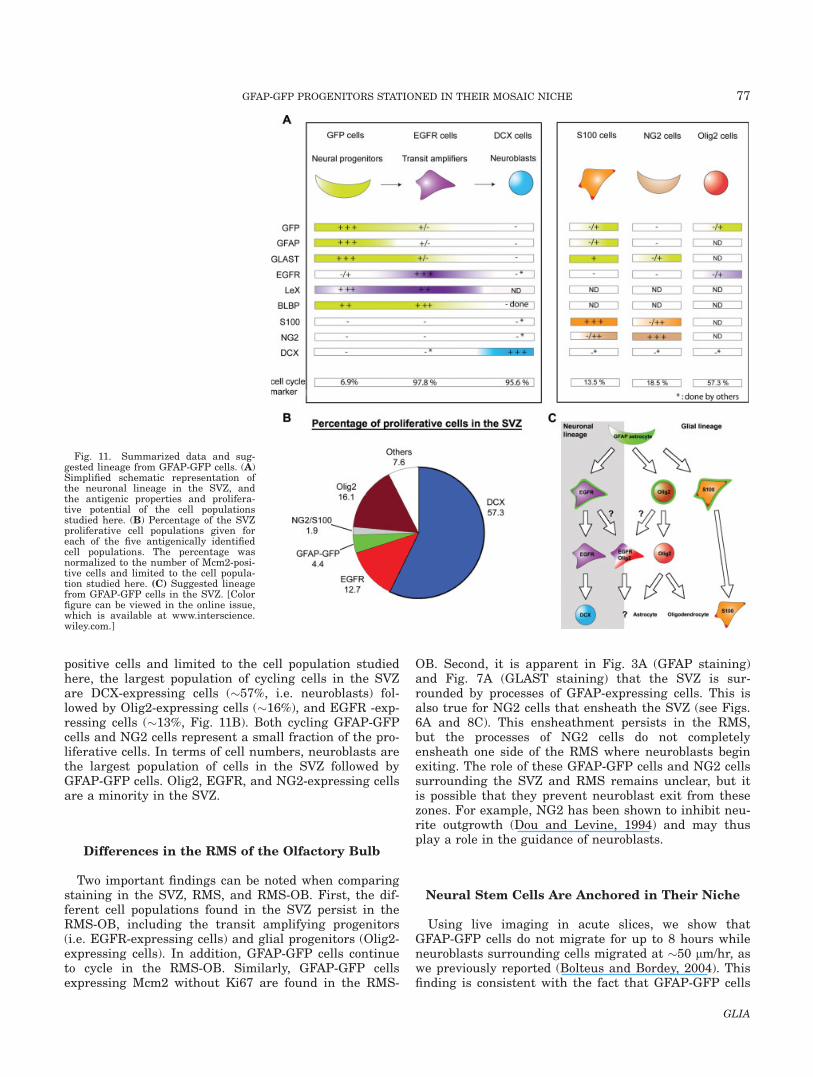
positive cells and limited to the cell population studiedhere, the largest population of cycling cells in the SVZare DCX-expressing cells (�57%, i.e. neuroblasts) fol-lowed by Olig2-expressing cells (�16%), and EGFR -exp-ressing cells (�13%, Fig. 11B). Both cycling GFAP-GFPcells and NG2 cells represent a small fraction of the pro-liferative cells. In terms of cell numbers, neuroblasts arethe largest population of cells in the SVZ followed byGFAP-GFP cells. Olig2, EGFR, and NG2-expressing cellsare a minority in the SVZ.
Differences in the RMS of the Olfactory Bulb
Two important findings can be noted when comparingstaining in the SVZ, RMS, and RMS-OB. First, the dif-ferent cell populations found in the SVZ persist in theRMS-OB, including the transit amplifying progenitors(i.e. EGFR-expressing cells) and glial progenitors (Olig2-expressing cells). In addition, GFAP-GFP cells continueto cycle in the RMS-OB. Similarly, GFAP-GFP cellsexpressing Mcm2 without Ki67 are found in the RMS-
OB. Second, it is apparent in Fig. 3A (GFAP staining)and Fig. 7A (GLAST staining) that the SVZ is sur-rounded by processes of GFAP-expressing cells. This isalso true for NG2 cells that ensheath the SVZ (see Figs.6A and 8C). This ensheathment persists in the RMS,but the processes of NG2 cells do not completelyensheath one side of the RMS where neuroblasts beginexiting. The role of these GFAP-GFP cells and NG2 cellssurrounding the SVZ and RMS remains unclear, but itis possible that they prevent neuroblast exit from thesezones. For example, NG2 has been shown to inhibit neu-rite outgrowth (Dou and Levine, 1994) and may thusplay a role in the guidance of neuroblasts.
Neural Stem Cells Are Anchored in Their Niche
Using live imaging in acute slices, we show thatGFAP-GFP cells do not migrate for up to 8 hours whileneuroblasts surrounding cells migrated at �50 lm/hr, aswe previously reported (Bolteus and Bordey, 2004). Thisfinding is consistent with the fact that GFAP-GFP cells
Fig. 11. Summarized data and sug-gested lineage from GFAP-GFP cells. (A)Simplified schematic representation ofthe neuronal lineage in the SVZ, andthe antigenic properties and prolifera-tive potential of the cell populationsstudied here. (B) Percentage of the SVZproliferative cell populations given foreach of the five antigenically identifiedcell populations. The percentage wasnormalized to the number of Mcm2-posi-tive cells and limited to the cell popula-tion studied here. (C) Suggested lineagefrom GFAP-GFP cells in the SVZ. [Colorfigure can be viewed in the online issue,which is available at www.interscience.wiley.com.]
77GFAP-GFP PROGENITORS STATIONED IN THEIR MOSAIC NICHE
GLIA

are neural stem cells that are derived from radial gliaand remain in their specific niche where they self-renew(Doetsch et al., 1999; Merkle et al., 2004, 2007). Never-theless, we found that GFAP-GFP cells display move-ment of their cell bodies that may represent shuffling oftheir DNA necessary for epigenetic regulation.
In conclusion, we show that GFP-fluorescent cells inthe SVZ and RMS express GFAP. Nevertheless, faintGFP is visible in some EGFR- and Olig2-positive cells,suggesting that they are generated from GFAP-GFPcells. The SVZ and the RMS are mosaics of different celltypes including GFAP-, DCX-, S100-, NG2-, Olig2-, andEGFR-expressing cells. These different populations,some of which display small overlap, have different pro-liferative potentials. This diversity adds to the alreadyknown diversity of regional stem cell fate (Merkle et al.,2007). Collectively, GFAP-GFP mice represent a valuabletool to study GFAP-expressing progenitors, and in par-ticular their behavior in acute slices.
ACKNOWLEDGMENTS
We thank Stephanie Young for thoroughly proof read-ing this article.
REFERENCES
Aguirre A, Gallo V. 2004. Postnatal neurogenesis and gliogenesis in theolfactory bulb from NG2-expressing progenitors of the subventricularzone. J Neurosci 24:10530–10541.
Aguirre AA, Chittajallu R, Belachew S, Gallo V. 2004. NG2-expressingcells in the subventricular zone are type C-like cells and contributeto interneuron generation in the postnatal hippocampus. J Cell Biol165:575–589.
Bartsch D, Mai JK. 1991. Distribution of the 3-fucosyl-N-acetyl-lactos-amine (FAL) epitope in the adult mouse brain. Cell Tissue Res263:353–366.
Baudier J, Glasser N, Gerard D. 1986. Ions binding to S100 proteins. I.Calcium- and zinc-binding properties of bovine brain S100alpha alpha, S100a (alpha beta), and S100b (beta beta) protein: Zn21
regulates Ca21 binding on S100b protein. J Biol Chem 261:8192–8203.
Bolteus AJ, Bordey A. 2004. GABA release and uptake regulate neuro-nal precursor migration in the postnatal subventricular zone. J Neu-rosci 24:7623–7631.
Bolteus AJ, Garganta C, Bordey A. 2005. Assays for measuring extrac-ellular GABA levels and cell migration rate in acute slices. Brain ResBrain Res Protoc 14:126–134.
Bordey A. 2006. Adult neurogenesis: Basic concepts of signaling. CellCycle 5:722–728.
Braun N, Sevigny J, Mishra SK, Robson SC, Barth SW, Gerstberger R,Hammer K, Zimmermann H. 2003. Expression of the ecto-ATPaseNTPDase2 in the germinal zones of the developing and adult ratbrain. Eur J Neurosci 17:1355–1364.
Brown DC, Gatter KC. 2002. Ki67 protein: The immaculate deception?Histopathology 40:2–11.
Capela A, Temple S. 2002. LeX/ssea-1 is expressed by adult mouse CNSstem cells, identifying them as nonependymal. Neuron 35:865–875.
Chan JA, Zhang H, Roberts PS, Jozwiak S, Wieslawa G, Lewin-KowalikJ, Kotulska K, Kwiatkowski DJ. 2004. Pathogenesis of tuberous scle-rosis subependymal giant cell astrocytomas: Biallelic inactivation ofTSC1 or TSC2 leads to mTOR activation. J Neuropathol Exp Neurol63:1236–1242.
Deloulme JC, Raponi E, Gentil BJ, Bertacchi N, Marks A, LabourdetteG, Baudier J. 2004. Nuclear expression of S100B in oligodendrocyteprogenitor cells correlates with differentiation toward the oligoden-
droglial lineage and modulates oligodendrocytes maturation. Mol CellNeurosci 27:453–465.
Doetsch F, Caille I, Lim DA, Garcia-Verdugo JM, Alvarez-Buylla A.1999. Subventricular zone astrocytes are neural stem cells in theadult mammalian brain. Cell 97:703–716.
Doetsch F, Garcia-Verdugo JM, Alvarez-Buylla A. 1997. Cellular compo-sition and three-dimensional organization of the subventricular ger-minal zone in the adult mammalian brain. J Neurosci 17:5046–5061.
Doetsch F, Petreanu L, Caille I, Garcia-Verdugo JM, Alvarez-Buylla A.2002. EGF converts transit-amplifying neurogenic precursors in theadult brain into multipotent stem cells. Neuron 36:1021–1034.
Dou CL, Levine JM. 1994. Inhibition of neurite growth by the NG2chondroitin sulfate proteoglycan. J Neurosci 14:7616–7628.
Gleeson JG, Lin PT, Flanagan LA, Walsh CA. 1999. Doublecortin is amicrotubule-associated protein and is expressed widely by migratingneurons. Neuron 23:257–271.
Gooi HC, Feizi T, Kapadia A, Knowles BB, Solter D, Evans MJ. 1981.Stage-specific embryonic antigen involves alpha 1 goes to 3 fucosy-lated type 2 blood group chains. Nature 292:156–158.
Hachem S, Aguirre A, Vives V, Marks A, Gallo V, Legraverend C. 2005.Spatial and temporal expression of S100B in cells of oligodendrocytelineage. Glia 51:81–97.
Hartfuss E, Galli R, Heins N, Gotz M. 2001. Characterization of CNSprecursor subtypes and radial glia. Dev Biol 229:15–30.
Imura T, Nakano I, Kornblum HI, Sofroniew MV. 2006. Phenotypic andfunctional heterogeneity of GFAP-expressing cells in vitro: Differen-tial expression of LeX/CD15 by GFAP-expressing multipotent neuralstem cells and non-neurogenic astrocytes. Glia 53:277–293.
Kimelberg HK. 2004. The problem of astrocyte identity. Neurochem Int45:191–202.
Liu X, Bolteus AJ, Balkin DM, Henschel O, Bordey A. 2006. GFAP-expressing cells in the postnatal subventricular zone display a uniqueglial phenotype intermediate between radial glia and astrocytes. Glia54:394–410.
Liu X, Wang Q, Haydar TF, Bordey A. 2005. Nonsynaptic GABA signal-ing in postnatal subventricular zone controls proliferation of GFAP-expressing progenitors. Nat Neurosci 8:1179–1187.
Lledo PM, Alonso M, Grubb MS. 2006. Adult neurogenesis and func-tional plasticity in neuronal circuits. Nat Rev Neurosci 7:179–193.
Marshall CA, Novitch BG, Goldman JE. 2005. Olig2 directs astrocyteand oligodendrocyte formation in postnatal subventricular zone cells.J Neurosci 25:7289–7298.
Maslov AY, Barone TA, Plunkett RJ, Pruitt SC. 2004. Neural stem celldetection, characterization, and age-related changes in the subven-tricular zone of mice. J Neurosci 24:1726–1733.
Menn B, Garcia-Verdugo JM, Yaschine C, Gonzalez-Perez O, RowitchD, Alvarez-Buylla A. 2006. Origin of oligodendrocytes in the subven-tricular zone of the adult brain. J Neurosci 26:7907–7918.
Merkle FT, Mirzadeh Z, Alvarez-Buylla A. 2007. Mosaic organization ofneural stem cells in the adult brain. Science 317:381–384.
Merkle FT, Tramontin AD, Garcia-Verdugo JM, Alvarez-Buylla A. 2004.Radial glia give rise to adult neural stem cells in the subventricularzone. Proc Natl Acad Sci USA 101:17528–17532.
Nacher J, Crespo C, McEwen BS. 2001. Doublecortin expression in theadult rat telencephalon. Eur J Neurosci 14:629–644.
Nishiyama A, Chang A, Trapp BD. 1999. NG21 glial cells: A novel glialcell population in the adult brain. J Neuropathol Exp Neurol58:1113–1124.
Parent JM, von dem BN, Lowenstein DH. 2006. Prolonged seizuresrecruit caudal subventricular zone glial progenitors into the injuredhippocampus. Hippocampus 16:321–328.
Peretto P, Giachino C, Aimar P, Fasolo A, Bonfanti L. 2005. Chain forma-tion and glial tube assembly in the shift from neonatal to adult subven-tricular zone of the rodent forebrain. J Comp Neurol 487:407–427.
Raponi E, Agenes F, Delphin C, Assard N, Baudier J, Legraverend C,Deloulme JC. 2007. S100B expression defines a state in which GFAP-expressing cells lose their neural stem cell potential and acquire amore mature developmental stage. Glia 55:165–177.
Sawamoto K, Wichterle H, Gonzalez-Perez O, Cholfin JA, Yamada M,Spassky N, Murcia NS, Garcia-Verdugo JM, Marin O, Rubenstein JL,Tessier-Lavigne M, Okano H, Alvarez-Buylla A. 2006. New neuronsfollow the flow of cerebrospinal fluid in the adult brain. Science311:629–632.
Scholzen T, Gerdes J. 2000. The Ki-67 protein: From the known andthe unknown. J Cell Physiol 182:311–322.
Stoeber K, Tlsty TD, Happerfield L, Thomas GA, Romanov S, BobrowL, Williams ED, Williams GH. 2001. DNA replication licensing andhuman cell proliferation. J Cell Sci 114:2027–2041.
78 PLATEL ET AL.
GLIA