irrigation of mediterranean crops with saline water: from physiology to management practices
TRANSCRIPT

www.elsevier.com/locate/agee
Agriculture, Ecosystems and Environment 106 (2005) 171–187
Irrigation of Mediterranean crops with saline water: from
physiology to management practices
N.V. Paranychianakisa, K.S. Chartzoulakisb,*
aNAGREF, Institute for Agricultural Research, 71307 Iraklio, Crete, GreecebNAGREF, Institute for Olives and Subtropical Plants, 73100 Chania, Crete, Greece
Abstract
Salinity is currently one of the most severe abiotic factors limiting agricultural production. The high rates of population
growth and global warming are expected to further exacerbate the threat of salinity, especially in areas with a semi-arid climate
as in the Mediterranean region. Salinity affects plant performance through the development of osmotic stress and disruption of
ion homeostasis, which in turn cause metabolic dysfunctions. Particular emphasis is given on the impacts of salinity on
photosynthesis because of its potential restrictions on plant growth and yield. The inhibition of photosynthesis under low to
moderate salinity stress appear to be mainly attributed to diffusional limitations (stomatal and mesophyll conductance), even for
salt-sensitive fruit trees such as citrus trees. In contrast, biochemical limitations to photosynthesis appear to occur only when
stress becomes heavy. A thorough understanding of the mechanisms conferring salt tolerance is therefore essential under the
expected climatic change, as it will enable the selection of salt-tolerant genotypes and the adoption of appropriate practices to
alleviate salinity impacts on agricultural production. In fruit trees, salt tolerance is mainly associated with their ability to restrict
salt accumulation in the leaves. Cell features of specific tissues, morphological factors and water-use efficiency regulate salt
accumulation in the shoot. Furthermore, most fruit trees display a rapid osmotic adjustment in response to salinity, which is
mainly attributed to the accumulation of inorganic ions and carbohydrates. Little information is available about the ability of
horticultural crops to detoxify reactive oxygen species and to synthesize compatible solutes and hence on the potential
contribution of these mechanism to induce salt tolerance in horticultural crops.
# 2004 Elsevier B.V. All rights reserved.
Keywords: Climate change; Horticultural crops; Mechanisms of salt tolerance; Photosynthesis; Water resources
1. Introduction
Water demand is increasing worldwide due to fast
population growth rates, improvement in living
* Corresponding author. Tel.: +30 821 97142;
fax: +30 821 93963.
E-mail address: [email protected] (K.S. Chartzoulakis).
0167-8809/$ – see front matter # 2004 Elsevier B.V. All rights reserved
doi:10.1016/j.agee.2004.10.006
standards, expansion of irrigation schemes and
global warming (IPCC, 1996; UN Population
Division, 1994). In regions affected by water
scarcity such as the Mediterranean basin, water
supplies are already degraded, or subjected to
degradation processes, which worsen the shortage
of water (Chartzoulakis et al., 2001; Attard et al.,
1996). Reduced water supplies induce restrictions on
.

N.V. Paranychianakis, K.S. Chartzoulakis / Agriculture, Ecosystems and Environment 106 (2005) 171–187172
water uses and allocation policies among different
user sectors. In such regions, the competition for
scarce water resources among users will inevitably
reduce the supplies of freshwater available for
crop irrigation. As a consequence, agriculture will
increasingly be forced to utilize marginal waters
such as brackish water or reclaimed effluent to meet
its increasing demands, which in turn increases the
risks of soil salinization and yield reduction.
Accumulation of salts in root zone affects plant
performance through the development of a water
deficit and the disruption of ion homeostasis (Zhu,
2001; Munns, 2002). These stresses change hormo-
nal status and impair basic metabolic processes
(Loreto et al., 2003; Munns, 2002) resulting in
inhibition of growth and reduction in yield (Maas,
1993; Prior et al., 1992; Paranychianakis et al.,
2004a). Depressed photosynthesis has been sug-
gested to be responsible for at least part of the growth
and yield reduction (Prior et al., 1992; Munns,
2002). Despite the vast number of studies dealing
with the impacts of salinity on photosynthesis of
horticultural crops, most of them fail to quantify the
nature of photosynthetic limitations. Stomatal
closure, arising from the osmotic component of
salinity, has been reported to be primarily respon-
sible for photosynthesis inhibition in some studies
(Paranychianakis et al., 2004b; Banuls and Primo-
Millo, 1995). Reductions in mesophyll conductance
due to salinity-induced anatomical changes in leaves
have also been suggested to contribute in photo-
synthesis inhibition in citrus (Citrus sp.) (Romero-
Aranda et al., 1998), grapevines (Vitis vinifera)
(Gibberd et al., 2003; Downton, 1977) and olive
trees (Olea europea) (Bongi and Loreto, 1989;
Loreto et al., 2003). Other studies have found strong
correlations between salt accumulation, in particular
Cl�, and photosynthesis reduction (Lloyd et al.,
1989; Walker et al., 1981; Chartzoulakis et al.,
2002), implying that non-stomatal factors dominate
in photosynthesis inhibition. In fact, such correla-
tions do not represent cause–effect relationships,
bringing into question the contribution of non-
stomatal limitations. Identifying the nature of non-
stomatal limitations of photosynthesis under stress
conditions is currently an active area of photosynth-
esis research (Medrano et al., 2002; Centritto et al.,
2003). Estimations based on the model of Farquhar
et al. (1980) suggest a reduction of Rubisco activity
even at moderate salinity levels (Loreto et al., 2003;
Rivelli et al., 2002), while in vitro assays show that
reductions in Rubisco activity and content occur
only under severe salt stress (Delfine et al., 1999).
However, Centritto et al. (2003) showed that
estimates of photosynthetic capacity based on A–
Ci curves without removing diffusional limitations
could lead to incorrect interpretations of the actual
limitations of photosynthesis.
In order to cope with salinity stress plants trigger
a variety of mechanisms, which differentiate sub-
stantially among plant species or genotypes. These
mechanisms operate in a coordinated manner both at
a cellular and a whole-plant level. In horticultural
crops, salt tolerance is associated with their ability to
restrict salt accumulation in leaves (Mullins et al.,
1996; White and Broadley, 2001). Damage in fruit
trees is closely associated with Cl� accumulation,
thus genotypes with enhanced ability to restrict Cl�
entry into shoots generally show a higher tolerance
(Antcliff et al., 1983; Banuls et al., 1997; Storey and
Walker, 1999). Salt accumulation in shoot depends
on cell features (Tester and Davenport, 2003),
morphological factors (Moya et al., 1999), tran-
spiration rate (Moya et al., 2003) and water-use
efficiency (Gibberd et al., 2003). The most horti-
cultural crops show a rapid osmotic adjustment in
response to salinity which is attributed to inorganic
ions and soluble carbohydrates (Walker et al., 1981;
Gucci et al., 1997; Lloyd et al., 1990), while a
limited number of studies deal with their ability
to synthesize compatible solutes (Lloyd et al.,
1990). Findings from annual crops show that
genotypes with effective antioxidant systems show
a superior performance in saline environments.
However, relatively little information is available
about the ability of horticultural crops to detoxify
reactive oxygen species. Arbona et al. (2003)
found that ‘Carrizo’ citrange, a salt-sensitive root-
stock, possesses an efficient defense system against
ROS generation. However, the change in salt
tolerance of certain genotypes in different areas
(Maas, 1993) and the inability of salt-tolerant cells to
generate tolerant plants (Tester and Davenport,
2003) show that our knowledge for the factors
induce salinity tolerance at a whole-plant level is
incomplete.
N.V. Paranychianakis, K.S. Chartzoulakis / Agriculture, Ecosystems and Environment 106 (2005) 171–187 173
2. Salinity and climate change in
Mediterranean region
Soil salinization is one of the most severe causes of
yield reduction in modern agriculture. On a worldwide
basis, salinity has already affected approximately
80 million ha of arable land (Ghassemi et al., 1995)
and still continues to increase (FAOSTAT, statistics
database, http://apps.fao.org/cgi-bin/nph-dp.pl). Other
estimates are considerably higher and indicate that up
to 50% of all irrigated land may be salt-affected
(Flowers, 1999). Irrigation with low-quality water
and/or improper management practices are the
principal causes of land salinization in the Mediterra-
nean.
2.1. Current situation
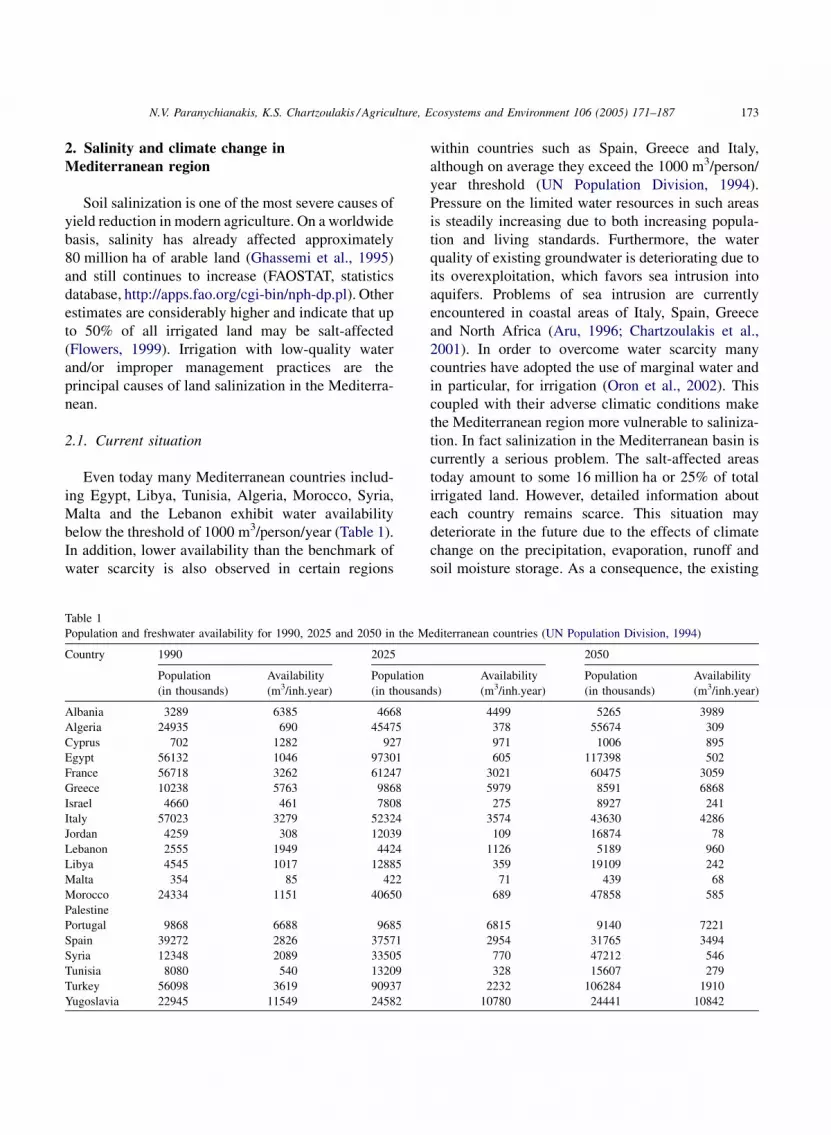
Even today many Mediterranean countries includ-
ing Egypt, Libya, Tunisia, Algeria, Morocco, Syria,
Malta and the Lebanon exhibit water availability
below the threshold of 1000 m3/person/year (Table 1).
In addition, lower availability than the benchmark of
water scarcity is also observed in certain regions
Table 1
Population and freshwater availability for 1990, 2025 and 2050 in the M
Country 1990 2025
Population
(in thousands)
Availability
(m3/inh.year)
Population
(in thousan
Albania 3289 6385 4668
Algeria 24935 690 45475
Cyprus 702 1282 927
Egypt 56132 1046 97301
France 56718 3262 61247
Greece 10238 5763 9868
Israel 4660 461 7808
Italy 57023 3279 52324
Jordan 4259 308 12039
Lebanon 2555 1949 4424
Libya 4545 1017 12885
Malta 354 85 422
Morocco 24334 1151 40650
Palestine
Portugal 9868 6688 9685
Spain 39272 2826 37571
Syria 12348 2089 33505
Tunisia 8080 540 13209
Turkey 56098 3619 90937
Yugoslavia 22945 11549 24582
within countries such as Spain, Greece and Italy,
although on average they exceed the 1000 m3/person/
year threshold (UN Population Division, 1994).
Pressure on the limited water resources in such areas
is steadily increasing due to both increasing popula-
tion and living standards. Furthermore, the water
quality of existing groundwater is deteriorating due to
its overexploitation, which favors sea intrusion into
aquifers. Problems of sea intrusion are currently
encountered in coastal areas of Italy, Spain, Greece
and North Africa (Aru, 1996; Chartzoulakis et al.,
2001). In order to overcome water scarcity many
countries have adopted the use of marginal water and
in particular, for irrigation (Oron et al., 2002). This
coupled with their adverse climatic conditions make
the Mediterranean region more vulnerable to saliniza-
tion. In fact salinization in the Mediterranean basin is
currently a serious problem. The salt-affected areas
today amount to some 16 million ha or 25% of total
irrigated land. However, detailed information about
each country remains scarce. This situation may
deteriorate in the future due to the effects of climate
change on the precipitation, evaporation, runoff and
soil moisture storage. As a consequence, the existing
editerranean countries (UN Population Division, 1994)
2050
ds)
Availability
(m3/inh.year)
Population
(in thousands)
Availability
(m3/inh.year)
4499 5265 3989
378 55674 309
971 1006 895
605 117398 502
3021 60475 3059
5979 8591 6868
275 8927 241
3574 43630 4286
109 16874 78
1126 5189 960
359 19109 242
71 439 68
689 47858 585
6815 9140 7221
2954 31765 3494
770 47212 546
328 15607 279
2232 106284 1910
10780 24441 10842

N.V. Paranychianakis, K.S. Chartzoulakis / Agriculture, Ecosystems and Environment 106 (2005) 171–187174
problems of water scarcity and quality will worsen and
countries in the southern and eastern Mediterranean
will be most affected.
2.2. Mediterranean and climate change
Various models have been developed to predict
climate change (IPCC, 1996). They predict an
increase in mean temperature, and changes in the
amounts and patterns of precipitation. However,
considerable uncertainty exists about the extent of
these changes.
Wingley (1992) predicted that a doubling of the
CO2 concentrations over the Mediterranean region
could cause warming of about 3.5 8C by the latter half
of the 21st century. Based on the runs of different
transient models, Rosenzweig and Tubiello (1997)
estimated that the temperature will rise by 1.4–2.6 8Cby 2020. It has also been predicted that by 2100
temperatures could have risen by up to 2.5–3 8C over
the Mediterranean sea, 3–4 8C over coastal areas and
4–4.5 8C over inland areas reaching its maximum
value, approximately 5.5 8C above Morocco (Cubasch
et al., 1996). With regards to precipitation, the
situation remains rather complicated. This is due to
the inability of global circulation models to predict
accurately regional rainfall (Palutikof and Wigley,
1996). However, there is a consensus that a decrease of
precipitation will occur in the part of Mediterranean
south of 40–458N and an increase in precipitation will
occur north of it (IPCC, 1996).
2.3. Effects of climatic change on water resources
availability and quality
Higher temperatures and population growth will
increase the demands for water in most Mediterranean
countries. Moreover, higher rates of evaporation
would cause rises in salt concentration in surface
water bodies, while rises in sea level would favor sea
intrusion into aquifers to coastal areas. It is estimated
that 1 m rise in the sea level will reduce water in the
main reservoir in Malta by 40% (Attard et al., 1996),
while in France, the salinity in the Vaccares is
expected to increase significantly (Corre, 1996).
Problems of sea intrusion would be further exacer-
bated in response to higher demand.
Under these conditions, freshwater resources
available for agriculture will decline quantitatively
and qualitatively. Water demands for irrigation are
projected to rise, bringing increased competition
between agriculture and other users. Therefore, the
use of lower-quality supplies will inevitably be
practiced for irrigation purposes in order to maintain
an economically viable agriculture. Many southern
and eastern countries of the Mediterranean (Algeria,
Cyprus, Morocco, Tunisia) have already experienced a
long drought. In these countries, the growth and yield
of crops were markedly reduced resulting in financial
consequences to their national economies.
3. Plant response to salinity
Salt accumulation in root zone causes the devel-
opment of osmotic stress and disrupts cell ion
homeostasis by inducing both the inhibition in uptake
of essential nutrients as K+, Ca2+ and NO3� and the
accumulation of toxic levels of Na+ and Cl�. In
addition, ROS can be generated (Zhu, 2001). These
stresses cause hormonal changes (Munns, 2002), alter
carbohydrate metabolism (Gao et al., 1998), reduce
the activity of certain enzymes (Munns, 1993) and
impair photosynthesis (Loreto et al., 2003). As a
consequence of these metabolic modifications and
dysfunctions, cell division and elongation decline or it
may be completely inhibited and cell death is
accelerated. At a whole-plant level, the impacts of
salinity are reflected through declines in growth,
reductions in yield and, in more acute cases, leaf
injuries are developed which can lead to the complete
defoliation of plants and their subsequent desiccation.
Munns (1993) suggested a two-phase model to
explain the response of plant growth to salinity. During
the first phase, growth reduction is ascribed to the
development of a water deficit, which prevails
immediately after the application of salinity treat-
ments. The second phase is due to accumulation of
salts in the shoot at toxic levels. This phase takes time
to develop depending on the intensity of stress and
plant tolerance to salinity. Under field conditions,
however, plant response to salinity may appreciably
deviate from this model. The gradual increase in soil
salinity in that case may result in a concurrent
occurrence of both osmotic and ionic effects of

N.V. Paranychianakis, K.S. Chartzoulakis / Agriculture, Ecosystems and Environment 106 (2005) 171–187 175
salinity. Furthermore, the composition of irrigation
water and the salt tolerance of the crop may affect the
contribution of each component in growth reduction.
During irrigation with waters of a high ECw value but
with relatively low concentrations of Na+ and Cl�, the
osmotic component will prevail over the ionic. In such
cases, drought-tolerant species will display a better
performance. In contrast, when water with high
concentrations of Na+ and Cl� is used for irrigation
of salt-sensitive crops or genotypes, the ionic
component will dominate. Although field studies
did not enable us the possibility to distinguish between
osmotic and ionic effects of salinity on performance of
horticultural crops it is most probable that the osmotic
component prevails in such conditions. Declines in
growth rate of Soultanina grapevines, planted in large
pots and grown under field conditions, were correlated
with corresponding reductions in predawn leaf water
potential (Cpd) (Fig. 1) indicating that osmotic effect
of salinity was the main cause of growth reduction.
Exposure of citrus genotypes to salinity treatments of
different composition (40 mM Na+, Cl� or NaCl) and
Fig. 1. Growth rate (A) and predawn leaf water potential variation
(B) in vines irrigated with recycled water and freshwater during the
1998 growing season.
isotonic solution of nutrients showed that growth
reduction was mainly due to the osmotic effect. In
‘Macrophylla’ however ionic effects were also
detected (Ruiz et al., 1999).
Generally, yield is reduced by salinity to a lesser
extent than growth. Although some relationships have
been established to describe the response of yield to
salinity, based on single studies or the compilation of
data from many studies (Maas and Hoffman, 1977;
Maas, 1993; Shalhevet and Levy, 1990), yield losses
often dramatically deviates from those predicted by
models. Possible causes are differences in salt
tolerance among genotypes or rootstocks, cumulative
effects of salinity on plant performance over the years,
soil type, environmental conditions and applied
management practices.
4. Salt stress and photosynthesis
4.1. Photosynthesis and growth
Whether decreased photosynthesis is the cause of
growth reduction due to a lower availability of
assimilates to growing sinks remains a matter of
controversy (Munns, 2002). This may be due to: (a) an
inability of single leaf photosynthesis to reflect net
carbon gain at a whole-plant level, (b) a divergent
response to salinity among plant species or genotypes
and (c) differences in the length of exposure and the
intensity of salt stress.
In the sort-term hormonal signals arising from
abscisic acid, biosynthesis appear to dominate in
growth reduction over water deficit, ionic imbalances
or decreased production of assimilates and that
response appears to be uniform for both annual
species and horticultural crops. This is supported by
the greater sensitivity of growth reduction either to salt
or water stress than photosynthesis (Paranychianakis,
2001). In the long-term, however, reduced availability
of carbohydrates may contribute in growth reduction.
Except the reductions in photosynthesis rate, salinity-
induced leaf senescence and abscission and the
inability of plants to produce new leaves may result
in assimilates starvation. The growth of salt-stressed
citrus recovered after the application of Ca2+ a
response ascribed to ameliorative effects of Ca2+,
on leaf abscission and hence, on the maintenance of

N.V. Paranychianakis, K.S. Chartzoulakis / Agriculture, Ecosystems and Environment 106 (2005) 171–187176
greater photosynthetic leaf area (Romero-Aranda
et al., 1998).
Reductions in net assimilation rate per unit of leaf
area were strongly correlated with depressed growth
implying that reduced production of assimilates has a
significant role in growth inhibition under saline
conditions (Lovelock and Ball, 2002). In addition, the
lower starch content measured at the end of growing
season in the tissues of grapevines irrigated with saline
effluent (Table 2) further supports the hypothesis that
inadequate supplies of assimilates contribute to
growth reduction (Paranychianakis, 2001). Reduced
supplies of starch in grapevines are also reported by
Prior et al. (1992). This is especially crucial for
horticultural crops since assimilates availability in old
wood affects the following season growth and the
potential for yield. More concrete evidence for the
possible involvement of reduced assimilates on
growth inhibition of salt-treated plants is provided
by plants grown under elevated CO2 (Mavrogiano-
poulos et al., 1999; Maggio et al., 2002). The
stimulation of growth of salt-stressed plants grown
under a CO2 environment implies that salinity-
induced reduction of photosynthesis reduces growth
through its effects on assimilates production.
It is not only net gain of carbon, but also the
availability of assimilates to growing sinks that may
affect its potential utilization for growth. Assimilates
in salt-treated plants may be preferentially directed for
osmotic adjustment, biosynthesis of compatible
solutes, repair of the salinity-caused damage and
the maintenance of basic metabolic processes at the
expense of growth. Thus, despite the increases in leaf
carbohydrates which is a common response of salt-
stressed plants (Gao et al., 1998; Walker et al., 1981),
growth may eventually be limited due to the reduced
availability of assimilates to growing sinks. Altera-
tions in the activity of specific enzymes may reduce
Table 2
The effects of water quality on root, trunk and stems content of soluble
Organs Carbohydrates (% dw)
Recycled water Freshw
Roots (<2 mm) 2.13 1.86
Roots (>2 mm) 1.68 1.66
Trunk 2.24 2.26
Stems 2.67 b 2.99 a
Any two means not followed by the same letter are significantly differen
assimilate utilization by growing tips. Gao et al.
(1998) found reduced activity of the enzyme acid
invertase in salt-stressed tomato plants, which may
have inhibited growth through the limited sucrose
utilization. In addition, assimilates’ accumulation in
leaves of salt-stressed grapevines and olives (Walker
et al., 1981; Downton and Loveys, 1981; Tattini et al.,
1996) resulting either from impairments in carbohy-
drate metabolism or for osmotic adjustment may result
in feedback repression on the Calvin cycle causing a
further reduction in photosynthesis and hence in
growth.
4.2. Photosynthesis and salinity
Reduction in photosynthesis of horticultural crops
grown in saline environments can be attributed to
reductions in stomatal or mesophyll conductance and
biochemical limitations. The relative contribution of
these limitations remains obscure and often contrast-
ing. This may due to the technical constrains when
assessing biochemical limitations, to difficulties to
separate between the osmotic and ionic effects of
salinity or to differences among species or genotypes.
Thus, findings suggest either Na+ and/or Cl� toxicities
(Banuls and Primo-Millo, 1992; Fisarakis et al., 2001;
Banuls et al., 1997; Lloyd et al., 1990) or stomatal
closure (Paranychianakis et al., 2004b; Banuls and
Primo-Millo, 1995) as the main causes of photo-
synthesis reduction on horticultural crops.
4.2.1. Stomatal limitations
The strong correlations between leaf Cl� and/or
Na+ content and photosynthesis rate as well as the
maintenance of turgor in salt-stressed plants have led
in the conclusion that biochemical limitations
dominate in the reduction of photosynthesis in salt-
stressed fruit trees. However, these correlations do not
carbohydrates and starch (adapted from Paranychianakis, 2001)
Starch (% dw)
ater Recycled water Freshwater
9.93 b 11.43 a
8.16 b 9.98 a
6.50 b 6.82 a
5.41 b 5.93 a
t at P < 0.05 with Tukey’s significant difference.

N.V. Paranychianakis, K.S. Chartzoulakis / Agriculture, Ecosystems and Environment 106 (2005) 171–187 177
Fig. 2. Relationship between photosynthesis (Pn) and predawn leaf
water potential for grapevines irrigated with recycled water (open
circles) and freshwater (filled squares) (adapted from Paranychia-
nakis et al., 2004b).
Fig. 3. Relationship between photosynthesis and CO2 draw-down
from ambient (Ca) to the chloroplast (Cc) concentration (adapted
from Loreto et al., 2003). Different symbols represent different olive
cultivars.
represent cause–effect relationships. In addition, it is
now widely accepted that stomatal closure is not due
to turgor loss, but it is a highly regulated response to
salinity (Munns, 1993). Abscisic acid biosynthesis and
its transfer to shoots, and the accumulation of
carbohydrates, K+, Ca2+ and Cl� in guard cells are
involved in stomatal closure (Robinson et al., 1997;
Talbott and Zeiger, 1998). The same relationships
between Cpd and gas exchange found for salt-stressed
and non-stressed grapevines imply that decreased
photosynthesis is attributed to stomatal closure arising
from the osmotic component of salinity (Fig. 2)
(Paranychianakis et al., 2004b). A significant correla-
tion between stomatal conductance and leaf water
potential in salt-treated citrus was also reported by
Banuls and Primo-Millo (1995). Based on the lack of
significant effect of salinity on midday leaf water
potential (Cmd), Walker et al. (1997) attributed the
decline of photosynthesis rather to an ion imbalance
than to Cl� toxicity or to water deficit development.
However, Cmd is not as reliable a parameter as Cpd for
assessing plant water status and it is consistent with
the absence of any significant effect of salinity on Cmd
of grapevines despite the significant reduction on Cpd
(Paranychianakis et al., 2004b). The rapid recovery of
photosynthesis after the relief of salt stress despite leaf
salt content remaining unchanged or even slightly
increasing (Walker et al., 1981; Fisarakis et al., 2001;
Tattini et al., 1995) provides further evidence that
stomatal limitations dominate in photosynthesis
reduction. Likewise, leaf gas exchange of citrus was
not affected by foliar application of NaCl even until
leaf burns began to develop (Romero-Aranda and
Syvertsen, 1996).
4.2.2. Mesophyll limitations
Both water and salt stress may cause changes in
leaf anatomy, which in turn can reduce the diffusion of
CO2 to chloroplasts. Decreases in mesophyll con-
ductance of salt-stressed fruit trees resulting from
increases in leaf thickness, reductions in intercellular
air spaces and the lower volume/area ratio of cells
have been associated with decreased photosynthesis
(Bongi and Loreto, 1989; Downton, 1977; Romero-
Aranda et al., 1998). These anatomical changes appear
to be genotype-dependent (Romero-Aranda et al.,
1998; Loreto et al., 2003). A recent study on olive trees
revealed that the sensitivity of photosynthesis to salts
was higher for cultivars with inherently higher rates of
photosynthesis (Loreto et al., 2003). This effect cannot
be explained either by the observed reduction of
photochemical efficiency or by changes in Rubisco
activity of salt-stressed leaves. The strong relationship
between photosynthesis and mesophyll conductance
or high CO2 drawn-down (Fig. 3), found both in
cultivars with inherently low photosynthesis and in
salt-stressed plants of all cultivars, suggests that the
low CO2 chloroplast concentration is the limiting
factor of photosynthesis in olive trees. These findings

N.V. Paranychianakis, K.S. Chartzoulakis / Agriculture, Ecosystems and Environment 106 (2005) 171–187178
indicate that salinity impacts on photosynthesis can be
reversed if the conductance to CO2 diffusion is
restored. This observation is of great importance with
respect to climate change. In fact, these results predict
a better performance for salt-stressed plants in a CO2-
rich world.
4.2.3. Biochemical limitations
Photosynthesis versus Ci curves have been widely
used to separate the biochemical from stomatal
limitations of photosynthesis. These curves often
show a decrease in apparent carboxylation efficiency
and hence on Rubisco activity even at moderate
salinity levels (Rivelli et al., 2002; Loreto et al., 2003).
However, such effects may due to technical constraints
as was recently shown in leaves of salt-treated olive
trees (Centritto et al., 2003). In that study the
preconditioning of salt-treated leaves to very low
CO2 to force the opening of stomata removed
limitations that implied biochemical impairment of
photosynthesis. These findings are in agreement with
those of in vitro assays which indicate that Rubisco
content and activity remain unchanged at moderate
salinity levels (Walker et al., 1981; Delfine et al.,
1999). Likewise, Medrano et al. (2002) investigating
the potential contribution of biochemical limitations
to photosynthesis reduction in water-stressed plants
found a similar relationship to that of Rubisco activity
assessed in vitro and gs when Pn–Ci curves were
converted to Pn–Cc and the apparent carboxylation
efficiency was recalculated. In addition, Rubisco is
maintained in excess in plant leaves since it may serve
for N storage. Thus, slight reductions in Rubisco
content may not limit photosynthetic capacity. Quick
et al. (1991) found that photosynthesis was reduced
only 6% when Rubisco was decreased by 60% in
tobacco plants. The lower contents of Rubisco were
compensated for by an increase in its activation
(60–100%), increases in its substrates, and a decrease
of its product.
4.3. Effects of salinity on photosynthesis under
climate change
The concentration of CO2 in the atmosphere is
increasing and is expected to double by the end of the
century. Plants grown under elevated CO2 environ-
ments show higher photosynthesis, reduced stomatal
conductance and improved water-use efficiency
(Drake et al., 1997). Thus, the performance of
horticultural crops grown under saline conditions
may be improved. Little information is available for
the performance of salt-stressed plants under condi-
tions of elevated CO2. Mavrogianopoulos et al. (1999)
reported that atmospheric CO2 concentrations of 800
and 1200 ppm stimulated photosynthesis in melons by
75 and 120%, respectively, in a range of salinity levels.
In another study, enhanced rates of photosynthesis in
response to a doubling of the atmospheric CO2
concentration were observed when plants exposed to
salinity levels of 25% seawater (Ball et al., 1997).
However, further increases in salinity level were not
resulted in differences in photosynthesis between
CO2-enriched and non-enriched plants implying that
biochemical limitations may prevail at higher salinity
levels. The increased rates of photosynthesis that are
observed under increased concentrations of CO2 may
be responsible for the better performance of salt-
treated tomato plants grown at 900 ppm compared to
those at 400 ppm. Plants grown at elevated CO2
exhibited a 60% greater threshold value for salinity
tolerance (Maggio et al., 2002). However, all the
above studies concentrate on the effects of increased
CO2 concentrations on plants grown in saline
environments without taking into consideration the
concurrent changes in temperature and the ozone
effects that may appreciably change their perfor-
mance. Thus, more information is needed to model the
performance of salinity-suffering crops with respect to
occurring climatic changes.
5. Mechanisms of salt tolerance
To cope with salinity plants trigger divergent
mechanisms that operate at a cellular and a
whole-plant level allowing their adaptation and
survival in saline environments. Differences in the
mechanisms plants posses determine their perfor-
mance under saline conditions. This review mainly
focuses on the processes regulating tolerance at a
whole-plant level such as salt uptake and transport to
the shoot, but some information is also given for
mechanisms operating at a cellular level such as
osmoregulation, compartmentation of salts and ROS
scavenging.

N.V. Paranychianakis, K.S. Chartzoulakis / Agriculture, Ecosystems and Environment 106 (2005) 171–187 179
5.1. Salt uptake and transport
In fruit species salinity tolerance has been
associated with their ability to restrict salts accumula-
tion in the shoot and particularly Cl�. Thus, leaf
Cl� content has been widely used as a criterion for
rating genotypes according to their ability to tolerate
salinity (Antcliff et al., 1983; Storey and Walker,
1999). The extent of salt accumulation in the shoot is
determined by the processes regulating net uptake
rate, loading to and reabsorption from the xylem,
and preferential distribution in particular organs or
tissues.
5.1.1. Salt net uptake
Net accumulation of salts into root is the result of
the balance between influx and efflux. These processes
are mainly regulated by ion channels and transporters
since the symplastic pathway appears to dominate for
both Na+ and Cl� entry (Tester and Davenport, 2003).
K+ channels and other non-selective cation channels
are considered responsible for Na+ uptake, while its
efflux is mediated by Na+/H+ antiporters (Blumwald et
al., 2000). In terms of Cl�, members of the ClC family,
various non-selective anion channels and Cl�/nH+
symporters appear to regulate Cl� accumulation into
root cells (Tyerman and Skerrett, 1999; White and
Broadley, 2001). In fruit trees such carriers have not
been identified yet, however, it can be inferred that
differences among genotypes or rootstocks in the
uptake and accumulation of salts (Chartzoulakis et al.,
2002; Moya et al., 2003; Romero-Aranda et al., 1998;
Walker et al., 1997) probably reflect differences in the
expression, the abundance or the properties of these
carriers. In addition, the rate of salt uptake by fruit
trees is dependent on their concentration in soil
solution (Moya et al., 2003; Storey, 1995) and
morphological factors such as the size of root system
and root to shoot ratio (Moya et al., 1999).
5.1.2. Root to shoot transport
Another control of salt accumulation in the shoot
can occur by minimizing salts’ efflux to the xylem or
by maximizing their reabsorption from the xylem at
the root or the stem. This means that parenchyma cells
have completely different properties from root cortical
cells since they need to maximize influx and minimize
efflux (Tester and Davenport, 2003). The mechanisms,
which regulate the removal of salts from xylem sap are
not fully understood.
Differences in the xylem Na+ and Cl� concentra-
tion assessed in citrus rootstocks (Zekri and Parsons,
1990; Walker et al., 1993b) may imply differences in
the rates of salt loading to xylem or in the mechanisms
they posses to reabsorb them back to xylem. The
increased accumulation of salts in leaves of Etrog
citron rootstock with increasing transpiration in
contrast to Rangpur lime rootstock in which salt
accumulation was not affected by transpiration rate
(Storey, 1995) may also imply differences in these
mechanisms among these rootstocks. The results of
this study also reveals the existence of a feedback
regulation of Cl� transport to shoot for Rangpur lime
rootstock which is probably modulated by leaf Cl�
content. Similarly, Elgazzar et al. (1965) found that
the shoot of Trifoliate orange was more effective to
restrict the transport of 22Na+ to leaves compared to
the Rough lemon. Furthermore, the ability of citrus to
reabsorb Na+ is highly depended on salt concentration
of the xylem sap (Elgazzar et al., 1965).
Preferential accumulation of both Na+ and Cl� in
the root has been often associated with lower salt
accumulation in the shoot. However, recent studies
question the contribution of this mechanism to induce
salt tolerance in the long-term or at high salinity
levels. Irrigation of grapevines with saline effluent did
not result in differences in Na+ accumulation in roots
(Fig. 4) at the end of the season compared to
freshwater-irrigated vines, while only slight differ-
ences were assessed in the case of Cl�. Significant
differences among citrus rootstocks to sequester salts
in roots were found only when irrigated with 30 and
60 mM NaCl, but these differences were eliminated
when salinity treatment increased to 90 mM NaCl
(Garcia-Sanchez et al., 2002). Similar results were
reported for olive trees by Chartzoulakis et al. (2002).
These studies indicate that sequestration of salts in
root can prevent salinity stress only at low salinity
levels or in the short-term.
Apart from differences in cell properties of
different tissues to control salt uptake and transloca-
tion in the shoot, morphological factors may also exert
an important role. Moya et al. (1999) found that
manipulation of the root to shoot ratio by applying root
pruning and defoliation affected Cl� accumulation in
the shoot. Increases in the root to shoot ratio and

N.V. Paranychianakis, K.S. Chartzoulakis / Agriculture, Ecosystems and Environment 106 (2005) 171–187180
Fig. 4. Na+ distribution in the different organs of grapevines irrigated with recycled (open bars, EC: 1.9 dS/m, Na+: 264 mg/l, Cl�: 436 mg/l) vs.
freshwater (solid bars, EC: 0.6 dS/m, Na+: 72 mg/l, Cl�: 118 mg/l) (adapted from Paranychianakis, 2001). Bars without letters imply non-
significant differences.
Fig. 5. Distribution of Cl� in leaves of different age according to its
availability in irrigation water (adapted from Paranychianakis,
2001). Bars without letters imply non-significant differences.
reductions in leaf biomass favored Cl� accumulation
in the shoot.
5.1.3. Distribution of salts within shoot
Preferential allocation of both Na+ and Cl� in old
leaves is crucial for salinity tolerance in glycophytes.
This may due to the rapid growth rates of young leaves
and the low transpiration rate. It is also possible that
salts are preferentially removed from sap moving to
actively growing organs (Tester and Davenport, 2003).
Marschner (1995) reported that 22Na+ which moved
out from source leaf did not reach into growing regions
of the roots or the shoot. It should be stressed,
however, that genotype and the intensity or the
duration of stress could substantially alter the pattern
of salts distribution within the shoot. Vines grafted on
41B rootstock showed a preferential accumulation of
Na+ to old leaves independent of salt availability,
while vines grafted on 1103P and 110R did not show
any differentiation in the allocation of Na+ with leaf
age (Fig. 4) (Paranychianakis, unpublished data). In
the same study Cl� distribution with leaf age did not
change in leaves of vines irrigated with saline effluent
in contrast to those irrigated with freshwater (Fig. 5)
implying that there is a threshold content for leaves to
sequester Cl�. After this threshold concentration is
exceeded Cl� is translocated uniformly in the younger
leaves. Moreover, salt distribution may change within
a given leaf. Preferential accumulation of Na+ to leaf
epidermical cells has been observed (Karley et al.,
2000). Such an allocation pattern for Na+ is of
paramount importance for maintaining photosynthetic
efficiency of mesophyll cells.

N.V. Paranychianakis, K.S. Chartzoulakis / Agriculture, Ecosystems and Environment 106 (2005) 171–187 181
5.2. Intercellular compartmentation
There is of substantial number of studies in the
literature showing that neither Na+ nor Cl� contents in
leaves is related to salt tolerance. This lack of
correlation probably results from differential ability of
various plant species or genotypes to distribute salts
within cell organelles. Sodium compartmentation into
vacuole appears to constitute the most effective
mechanism of plant cells to handle efficiently high
concentrations of salts and to prevent their toxic
effects on cytoplasm. Na+ compartmentation is
regulated by Na+/H+ antiporters (Hasegawa et al.,
2000). The overexpression of genes encoding Na+/H+
antiporters in different plant species induced the
tolerance of plants to salinity. Zhang and Blumwald
(2001) reported that tomato plants (Lycopersicum
solanum) overexpressing an Arabidopsis vacuolar
Na+/H+ antiporter were able to grow and produce
fruits even at concentrations of 200 mM NaCl. Leaf
Na+ and Cl� contents reached values of 20-fold to
those of wild-type plants.
5.3. Osmotic adjustment
Accumulation of solutes is a universal response of
stressed plants grown. Enhanced levels of osmolytes
in certain taxonomic groups such as xerophytes and
halophytes, known for their outstanding ability to
withstand adverse environmental conditions, imply
their crucial role in plant adaptation and survival in
water-deficient environments. Accumulation of
solutes results in osmotic adjustment favoring water
absorption and retention, which may maintain plant
growth and photosynthesis. Most horticultural crops
display a rapid osmotic adjustment in response to
salinity, which is ascribed mainly to ions and/or
carbohydrates accumulation (Downton and Loveys,
1981; Lloyd et al., 1990; Banuls and Primo-Millo,
1992; Gucci et al., 1997). Relative little information
is available for the ability of these species to
accumulate compatible solutes. Compatible solutes
apart from their contribution in osmotic adjustment
may have a protective role in protein structure and
photosynthesis. They probably act as osmoprotec-
tants and ROS scavengers. Engineering plant species
with genes inducing the biosynthesis of compatible
solutes such as D-ononitol or sorbitol has been
associated with higher rates of photosynthesis
(Sheveleva et al., 1997; Gao et al., 2001). Citrus
accumulate mainly proline and proline betaine and it
is dependent on genotype (Lloyd et al., 1990; Banuls
and Primo-Millo, 1992; Walker et al., 1993a, 1993b),
however, the contribution of these solutes in
conferring salt tolerance and photosynthesis of
citrus remains questionable (Lloyd et al., 1989).
Apparently, more research should be conducted
on this field to elucidate the role of compatible
solutes in conferring salt tolerance in horticultural
crops.
5.4. Reactive oxygen species scavenging
The generation of ROS in salt-stressed plants is
mainly induced from pathways alternative to
photosynthesis and photorespiration, from photo-
synthetic apparatus and from mitochondrial respira-
tion and may result in peroxidation of membrane
lipids, oxidation of proteins and disruption of PSII
(Mittler, 2002; Zhu, 2001; Nishiyama et al., 2001).
Thus, plants with more effective antioxidant systems
will display a superior performance in saline
environments. Little information is available about
the extent that ROS may contribute to salinity-
induced damage in horticultural crops. Differences
in the salt tolerance between two mulberry geno-
types (Morrus alba L.) were associated with
differences in the activity of enzymes involved in
ROS detoxification (Subhakar et al., 2001). ‘Carrizo’
citrange, sensitive to salinity rootstock, responded to
salt-induced oxidative stress by increasing enzy-
matic and non-enzymatic antioxidant defense pro-
portionally to the intensity of stress resulting in low
levels of malondialdehyde content (Arbona et al.,
2003).
6. Management practices
In order to mitigate the impacts of the use of low-
quality waters on the productivity of agricultural
crops, intense management practices should be
adopted. These practices can be separated into three
distinct categories: (a) irrigation management strate-
gies, (b) plant cultural practices and (c) the selection of
salt-tolerant genotypes.

N.V. Paranychianakis, K.S. Chartzoulakis / Agriculture, Ecosystems and Environment 106 (2005) 171–187182
6.1. Irrigation practices
Irrigation management practices aim for the
efficient use of saline water. This can be achieved
by maintaining salt accumulation in the root zone at
levels lower than the threshold values. Above these
values, a reduction of yield is observed. Such practices
are: (a) proper irrigation scheduling, (b) efficient
leaching of salts, (c) selection of irrigation method and
(d) establishing artificial drainage.
6.1.1. Irrigation scheduling
The term ‘irrigation scheduling’ includes both the
estimation of the irrigation requirements of the given
crop and the application of the appropriate irrigation
intervals. The establishment of an appropriate irrigation
schedule under saline water irrigation is much more
complicated than when freshwater is applied, due to
limited or completely absent information on the water-
use of salt-stressed plants. Critical questions that arise
are if the salinity changes the consumptive use of
irrigated plants and what are the leaching requirements
(LR) that should be included in the crop water
demands for waters of variable quality. Successful
irrigation with low-quality water requires relationships
that relate yield to water consumption. To establish such
relationships, two different approaches can be used.
Field experimentation, which is an expensive and time-
consuming procedure, or the use of mathematical
models to describe the soil-water–crop system.
Plant growth response under saline irrigation is a
function of the salts’ concentration in soil solution, in
particular Na+ and Cl�, and the matrix potential of the
soil. Thus, maintaining adequate soil-water avail-
ability is essential to restrict the damage of salt
accumulation. This can be achieved by increasing
irrigation frequency. After irrigation, soil moisture
content is high and the salt concentration or osmotic
pressure of the soil solution approaches their minimal
values.
6.1.2. Irrigation method
Irrigation method applied for saline irrigation may
have a great influence on salt accumulation and
distribution in the soil profile and hence on crop
production. Sprinkler irrigation with saline water may
cause injury if applied to plants with high rates of
foliar salt absorption, and the injury risk is greater if
irrigation is practiced during the daytime when the
evaporation rate is high. Trickle or drip irrigation is
recommended as it keeps the soil moisture continu-
ously high at the root zone, maintaining a low salt
concentration level. Common problems associated
with drip irrigation are the need to remove the
accumulating salts from the wetting front and the
avoidance of drippers clogging. The use of subsurface
drip irrigation (SDI) appears to be an ideal method for
irrigation with saline water. Irrigation of pears (Prunus
sp.) with saline water (ECw = 4.4 dS/m) through SDI
increased yield compared to surface drip irrigation
(Oron et al., 2002). In addition, the depth that emitters
are located appears to be a critical parameter since it
affects salt distribution in the root zone and therefore
the intensity of stress.
6.1.3. Leaching requirements
The amount of water (in terms of a fraction of the
applied water) that must be applied in excess to the
crop in order to control salts is referred to as ‘leaching
requirements’ (LR) and can be calculated, for drip
irrigation, from the following formula (Ayers and
Westcot, 1985):
LR ¼ ECw
5ECe � ECw(1)
where ECw is the electrical conductivity (dS/m) of the
irrigation water and ECe the electrical conductivity
(dS/m) of the saturation extract. ECe is the average soil
salinity tolerated by the crop. Depending on the crop
and the salinity of the water and soil, a 15–20%
leaching fraction is commonly recommended.
6.1.4. Establishing artificial drainage
When saline water is used for irrigation, existing
drainage problems greatly complicate water manage-
ment for salinity control. Temporary or permanent
high water tables (1.5 m or less) make the control of
salts difficult since their leaching is ineffective. A
more effective way for controlling the salinity
problems associated with a high water table is to
establish artificial drainage.
6.2. Cultural practices
Cultural practices may dramatically improve the
performance of crops grown in saline environments. In

N.V. Paranychianakis, K.S. Chartzoulakis / Agriculture, Ecosystems and Environment 106 (2005) 171–187 183
this section, emphasis is given to practices such as the
application of water or soil amendments, improved
fertilization schedule and other miscellaneous prac-
tices.
6.2.1. Water or soil amendments
Soil permeability problems can be prevented or
corrected by using soil or water amendments.
Improved soil permeability can be achieved if either
the sodium in the irrigation water is lowered or the
calcium and magnesium concentration will increase.
However, at present there are no economically viable
processes for removing salts from irrigation water.
Chemicals can be added to the soil or irrigation water
to increase calcium concentration and to improve the
sodium/calcium ratio. Gypsum, sulfur or sulfuric acid
are the most commonly used soil amendments, while
gypsum, sulfuric acid and sulfur dioxide are used as
water amendments. Rates of gypsum application to
soil commonly range from 2 to 20 ton/ha, but amounts
as high as 40 ton/ha can be used in areas with
extremely high sodium content. Mulching may also be
beneficial on saline soils since it reduces evaporation
from the soil surface and/or encourages downward
flux of soil water.
6.2.2. Improved fertilization schedule
Inhibition of the uptake of essential nutrients by
salinity may result in severe reductions in yield,
depending on plant species. Supplemental fertiliza-
tion, particularly of K+, Ca2+, NO3� and in some cases
micronutrients, lead to a recovery of physiological
parameters and stimulates growth (Cramer and
Nowak, 1992; Marschner, 1995; Zhu, 2001). In
addition, application of K+ and Ca2+ may also
improve plant performance by reducing the uptake
of salts (Romero-Aranda et al., 1998).
Table 3
Potential use of such resources for citrus, grapevines and olive trees irrig
Water classification TDS (ppm) EC (dS/m
Freshwater <500 <0.6
Slightly brackish 500–1000 0.6–1.5
Brackish 1000–2000 1.5–3.0
Moderately saline 2000–5000 3.0–8.0
Saline 5000–10000 8.0–15
Highly saline 10000–35000 15.0–45
6.2.3. Miscellaneous practices
The introduction of arbuscular mycorrhizae has
been found to improve the performance of plants
grown in saline environments (Ruiz-Lozano et al.,
1996). The beneficial effects of arbuscular mycor-
rhizae are associated with improved nutrition and
better water absorption (Ruiz-Lozano and Azcon,
1995). The better performance of salt-treated Lactuca
sativa plants inoculated with arbuscular mycorrhizae
was associated with increased photosynthesis and
water-use efficiency (Ruiz-Lozano et al., 1996). Other
techniques, like the foliar application of polyamines or
glycine betaine, appear also to provide promising
results for their commercial application in the future to
improve the performance of salt-stressed plants.
6.3. Salt tolerance of different plant species and
genotypes
The selection of plant-tolerant plant species or
genotypes is a common practice to reduced losses of
yield under saline conditions. Threshold values of
salinity tolerance for citrus, grapevines and olive trees
are given in Table 3. Declines in water quality below
the threshold values reported in Table 3 do not
preclude their potential use for irrigation of the
considered crops, however the adoption of both
intense management practices and the use of salt-
tolerant genotypes is recommended to maintain crop
productivity in acceptable levels and to ensure land
sustainability. It should be stressed, however, that the
threshold values reported in Table 3 is indicative since
they may considerably vary among different cultivars
or rootstocks (Antcliff et al., 1983; Storey and Walker,
1999; Chartzoulakis et al., 2002). A classification of
various genotypes of citrus, grapevines and olives is
shown in Table 4 based on published studies.
ation
) Crop Threshold EC (dS/m)
Citrus 1.1–1.4
Grapevines 1.4–3.0
Olives 1.8–2.5
.0
.0

N.V. Paranychianakis, K.S. Chartzoulakis / Agriculture, Ecosystems and Environment 106 (2005) 171–187184
Table 4
Salt tolerance rating of various cultivars and rootstocks of citrus, grapevines and olives
Tolerant genotypes
Citrus Sunki mandarin, Cleopatra mandarin, Rangpur lime
Grapevines Ramsey, Dogridge, French Colombard
Olives Picual, Canivano, Jabaluna, Nevadillo, Ardequina, Frantoio, Kalamata, Lianolia Kerkiras, Megaritiki,
Korthreiki
Moderately tolerant genotypes
Citrus Sampson tangelo, Rough lemon, Sour orange, Ponkan mandarin
Grapevines 110R, 1103P, 140R, Chenin Blanc, Grenache, Soultanina
Olives Chorruo, Changlot Real, Verdial de velez, Gordal Sevillana, Oblonga, Blanqueta, Alameno, Manzanillo, Redondil,
Hojiblanca, Canivano Negro, Zorzariega, Picudo, Coratina, Maraiolo, Maurino, Koroneiki, Mastoidis, Amphisis,
Valanolia, Adramitini
Sensitive genotypes
Citrus Troyer citrange, Trifoliate orange, Rusk citrange, Sweet orange
Grapevines 41B, SO4, Muscat of Alexandria, Barbera, Ribier
Olives Pajarero, Chetoui, Calego, Cobancosa, Meski, Leccino, Throumbolia, Chondrolia Chalkidikis, Agouromanaki
Salt tolerance in each category is comparative for the given crop and does not imply similar salt tolerance among the different crops.
Furthermore, in woody plants salt tolerance of a
given genotype can display significant variations from
one area to another. Such variations have been
reported for citrus (Maas, 1993) and grapevines
(Downton, 1977; Antcliff et al., 1983; Arbabzadeh and
Dutt, 1987). Causes responsible for this variation are
differences in environmental factors (soil fertility, soil
physical conditions and climatic factors) which are
met from one region to another and plant genetic
diversity.
7. Conclusions and future research needs
Population growth and global warming will
substantially impact the availability and quality of
existing freshwater supplies. As a consequence, the
risk of land salinization will further threaten agri-
cultural production, particularly in areas with a semi-
arid or arid climate. However, more detailed studies
are needed to quantify the temporal and spatial effects
of climate change on water resources. Such informa-
tion is of paramount importance to adopt appropriate
management practices to minimize the salinization of
agricultural land and the impacts of salinity on crops’
productivity.
Decreased photosynthesis may represent a serious
constrain for current’s season growth and yield. In
addition, the lower amounts of assimilates in
permanent organs of perennial plants may be
responsible for the progressive decline in their
performance and their reduced fertility. Diffusional
limitations appear to dominate in photosynthesis
reduction at low to moderate salinity levels even for
plant species sensitive to salinity. This implies that the
performance of salt-stressed plants will be improved
from the expected increase in CO2 concentration.
However, results of plants growing under elevated
CO2 environments are limited and often confusing.
The situation becomes even more complex if we take
into account the concurrent increase in temperature
and the reductions in the availability of water
resources and nutrients.
Since global warming is expected to increase the
salt-affected land, the need for a thorough under-
standing of the mechanisms determining salt tolerance
in plants becomes more crucial in order to maintain
agricultural production within economically viable
levels. Despite the exciting progress, which has been
performed the last decade in terms of the identification
of genes inducing salinity tolerance, signaling and
biochemical adjustments as well as the mechanisms
operating at a whole-plant level, our knowledge is
incomplete. This is confirmed by the differential
response to salinity of a given genotype in different
areas and by the inability of salt-tolerant cells to
generate tolerant plants. Therefore, a better under-
standing of the interactions among genetic traits,
climatic conditions and management practices is
required.

N.V. Paranychianakis, K.S. Chartzoulakis / Agriculture, Ecosystems and Environment 106 (2005) 171–187 185
References
Antcliff, A.J., Newman, H.P., Barret, H.C., 1983. Variation in
chloride accumulation in some American species of grapevine.
Vitis 22, 357–362.
Arbabzadeh, F., Dutt, G., 1987. Salt tolerance of grape rootstocks,
under greenhouse conditions. Am. J. Enol. Vitic. 38, 95–101.
Arbona, V., Flors, V., Jacas, J., Garcia-Agustin, P., Gomez-Gadenas,
A., 2003. Enzymatic and non-enzymatic antioxidant responses
of Carrizo citrange, a salt sensitive Citrus rootstock, to different
levels of salinity. Plant Cell Physiol. 44, 388–394.
Aru, A., 1996. The Rio Santa Lucia site: an integrated study of
desertification. In: Brandt, C.J., Thornes, J.B. (Eds.), Mediter-
ranean Desertification and Land Use. Wiley, Chichester, UK, pp.
189–206.
Attard, D.J., Axiak, V., Borg, S., Cachia, J., de Bono, G., Lanfranco,
E., Micallef, R.E., Mifsud, J., 1996. Implications of expected
climatic changes for Malta. In: Jeftic, L., Keckes, S., Pernetta, J.
(Eds.), Climate Change and the Mediterranean, vol. 2. Arnold,
UK, pp. 322–430.
Ayers, R.S., Westcot, D.W., 1985. Irrigation water quality. In:
Pettygrove, G.S., Asano, T. (Eds.), Irrigation with Reclaimed
Municipal Wastewater – A Guidance Manual. Lewis Publishers.
Ball, M.C., Cochrane, M.J., Rawson, H.M., 1997. Growth and water
use of the mangroves Rhizophora apiculata and R. stylosa in
response to salinity and humidity under ambient and elevated
concentrations of atmospheric CO2. Plant Cell Environ. 20,
1158–1166.
Banuls, J., Primo-Millo, E., 1992. Effects of chloride and sodium on
gas exchange parameters and water relations of Citrus plants.
Physiol. Plant. 86, 115–123.
Banuls, J., Primo-Millo, E., 1995. Effects of salinity on some Citrus
scion-rootstock combinations. Ann. Bot. 76, 97–102.
Banuls, J., Serna, M.D., Legaz, F., Talon, M., Primo-Millo, E., 1997.
Growth and gas exchange parameters of Citrus plants stressed
with different salts. J. Plant Physiol. 150, 194–199.
Blumwald, E., Aharon, G.S., Apse, M.P., 2000. Sodium transport in
plants. Biochim. Biophys. Acta 1465, 140–151.
Bongi, G., Loreto, F., 1989. Gas exchange properties of salt-stressed
olive (Olea europaea L.) leaves. Plant Physiol. 90, 1408–1416.
Centritto, M., Loreto, F., Chartzoulakis, K., 2003. The use of low
[CO2] to estimate diffusional and non-diffusional limitations of
photosynthetic capacity of salt-stressed olive saplings. Plant Cell
Environ. 26, 585–594.
Chartzoulakis, K., Loupassaki, M., Bertaki, M., Androulakis, I.,
2002. Effects of NaCl salinity on growth, ion content and CO2
assimilation rate of six olive cultivars. Sci. Hortic. 96, 235–247.
Chartzoulakis, K.S., Paranychianakis, N.V., Angelakis, A.N., 2001.
Water resources management in the island of Crete, Greece with
emphasis on the agricultural use. Water Policy 3, 193–205.
Corre, J.J., 1996. Implications des changements climatiques
etude de cas:Le Golfe du Lion (France). In: Jeftic, L., Milli-
man, J.D., Sestini, G. (Eds.), Climatic Change and the Medi-
terranean. Edward Arnold, London, pp. 328–427.
Cramer, G.R., Nowak, R.S., 1992. Supplemental manganese
improves the relative growth, net assimilation and photosyn-
thetic rates of salt-stressed barley. Physiol. Plant. 84, 600–605.
Cubasch, U., von Storch, H., Waszkewitz, J., Zorita, E., 1996.
Estimates of climate change in southern Europe derived
from dynamical climate model output. Clim. Res. 7, 129–
149.
Delfine, S., Alvino, A., Villani, M.C., Loreto, F., 1999. Restrictions
to CO2 conductance and photosynthesis in spinach leaves reco-
vering from salt stress. Plant Physiol. 119, 1101–1106.
Downton, W.J.S., 1977. Photosynthesis in salt stressed grapevines.
Aust. J. Plant Physiol. 4, 183–192.
Downton, W.J.S., Loveys, B.R., 1981. Abscisic acid content and
osmotic relations of salt-stressed grapevine leaves. Aust. J. Plant
Physiol. 8, 443–448.
Drake, A.M., Gonzalez-Meler, M., Long, S.P., 1997. More efficient
plants: a consequence of rising atmospheric CO2. Annu. Rev.
Plant Physiol. Mol. Biol. 48, 607–637.
Elgazzar, A., Wallace, A., Hemaidan, N., 1965. Sodium distribution
in rough lemon and trifoliate orange seedlings. Soil Sci. 99, 387–
391.
Farquhar, G.D., von Caemmerer, S., Berry, J.A., 1980. A biochem-
ical model of photosynthetic CO2 assimilation in leaves of C3
species. Planta 149, 78–90.
Fisarakis, I., Chartzoulakis, K., Stavrakas, D., 2001. Response of
Sultana vines (V. vinifera L.) on six rootstocks to NaCl
salinity exposure and recovery. Agric. Water Manage. 51, 13–
27.
Flowers, T.J., 1999. Salinisation and horticultural production. Sci.
Hortic. 78, 1–4.
Gao, Z., Sagi, M., Lips, S.H., 1998. Carbohydrate metabolism in
leaves and assimilate partitioning in fruits of tomato (Lycoper-
sicon esculentum L.) as affected by salinity. Plant Sci. 135, 149–
159.
Gao, M., Tao, R., Miura, K., Dandekar, A.M., Sugiura, A., 2001.
Transformation of Japanese persimmon (Diospyros kaki Thunb.)
with apple cDNA encoding NADP-dependent sorbitol-6-phos-
phate dehydrogenase. Plant Sci. 160, 837–845.
Garcia-Sanchez, F., Jifon, J.L., Carvajal, M., Syvertsen, J.P., 2002.
Gas exchange, chlorophyll, nutrient contents in relation to Na+
and Cl� accumulation in Sunburst mandarin grafted in different
rootstocks. Plant Sci. 162, 705–712.
Ghassemi, F., Jakeman, A.J., Nix, H.A., 1995. Salinization of Land
and Water Resources: Human Causes, Extent Management and
Case Studies. UNSW Press/CAB International, Sydney, Aus-
tralia/Wallingford, UK.
Gibberd, M.R., Walker, R.R., Condon, G.A., 2003. Whole-plant
transpiration efficiency of Sultana grapevine grown under saline
conditions is increased through the use of Cl-excluding root-
stock. Funct. Plant Biol. 30, 643–652.
Gucci, R., Lombardini, L., Tattini, M., 1997. Analysis of leaf water
relations in leaves of two olive (Olea europea) cultivars differing
in tolerance to salinity. Tree Physiol. 17, 13–21.
Hasegawa, P.M., Bressan, S.A., Zhu, J.K., Bohnert, H.J., 2000. Plant
cellular and molecular responses to high salinity. Annu. Rev.
Plant Physiol. Plant Mol. Biol. 51, 463–499.
IPCC, 1996. Houghton, J.T., Meira Filho, L.B., Callander, B.A.,
Harris, N., Kattenberg, A., Maskell, K. (Eds.), Climate Change
1995: The Science of Climate Change. Intergovernmental Panel
on Climate Change, Cambridge University Press, Cambridge.

N.V. Paranychianakis, K.S. Chartzoulakis / Agriculture, Ecosystems and Environment 106 (2005) 171–187186
Karley, A.J., Leigh, R.A., Sanders, D., 2000. Differential ion
accumulation and ion fluxes in the mesophyll and epidermis
of barley. Plant Physiol. 122, 835–844.
Lloyd, J., Kriedemann, P.E., Aspinall, D., 1989. Comparative sen-
sitivity of Prior Lisbon lemon and Valencia orange trees to foliar
sodium and chloride concentrations. Plant Cell Environ. 12,
529–540.
Lloyd, J., Kriedemann, P.E., Aspinall, D., 1990. Contrasts between
Citrus species in response to salinisation: an analysis of photo-
synthesis and water relations for different rootstock scion com-
binations. Physiol. Plant. 78, 236–246.
Loreto, F., Centritto, M., Chartzoulakis, K., 2003. Photosynthetic
limitations in olive cultivars with different sensitivity to salt
stress. Plant Cell Environ. 26, 595–601.
Lovelock, C.E., Ball, M.C., 2002. Influence of salinity on photo-
synthesis of halophytes. In: Lauchli, A., Lutge, U. (Eds.), Sa-
linity: Environment–Plants–Molecules. Kluwer Academic
Publishers, pp. 315–340.
Maas, E.V., 1993. Salinity and citriculture. Tree Physiol. 12, 195–
216.
Maas, E.V., Hoffman, G.J., 1977. Crop salt tolerance: current
assessment. J. Irrig. Drain. Div. 103, 115–134.
Maggio, A., Dalton, F.N., Piccinni, G., 2002. The effects of elevated
carbon dioxide on static and dynamic indices for tomato salt
tolerance. Eur. J. Agron. 16, 197–206.
Marschner, H., 1995. Mineral Nutrition of Higher Plants, 2nd ed.
Academic Press, London.
Mavrogianopoulos, G.N., Spanakis, J., Tsikalas, P., 1999. Effect of
carbon dioxide enrichment and salinity on photosynthesis and
yield in melon. Sci. Hortic. 79, 51–63.
Medrano, H., Escalona, J.M., Bota, J., Gulias, J., Flexas, J., 2002.
Regulation of photosynthesis to progressive drought: stomatal
conductance as a reference parameter. Ann. Bot. 89, 895–
905.
Mittler, R., 2002. Oxidative stress, antioxidants and stress tolerance.
Trends Plant Sci. 7, 405–410.
Moya, J.L., Gomez-Cadenas, A., Primo-Millo, E., Talon, M., 2003.
Chloride absorption in salt-sensitive Carrizo citrange and salt-
tolerant Cleopatra mandarin citrus rootstocks is linked to water
use. J. Exp. Bot. 54, 825–833.
Moya, J.L., Primo-Millo, E., Talon, M., 1999. Morphological factors
determining salt tolerance in citrus seedlings: the shoot to root
ratio modulates passive root uptake of chloride ions and
their accumulation in leaves. Plant Cell Environ. 22, 1425–
1433.
Mullins, M.G., Bouquet, A., Williams, L.E., 1996. Biology of
Grapevine, 3rd ed. Press Syndicate of the University of Cam-
bridge, Cambridge, p. 239.
Munns, R., 1993. Physiological processes limiting plant growth in
saline soils: some dogmas and hypotheses. Plant Cell Environ.
16, 15–24.
Munns, R., 2002. Comparative physiology of salt and water stress.
Plant Cell Environ. 25, 239–250.
Nishiyama, Y., Yamamoto, H., Allakhverdiev, S.I., Inaba, M.,
Yokota, A., Murata, N., 2001. Oxidative stress inhibits the repair
of photodamage to the photosynthetic machinery. EMBO J. 20,
5587–5594.
Oron, G., DeMalach, Y., Gillerman, L., David, I., Lurie, S., 2002.
Effect of water salinity and irrigation technology on yield and
quality of pears. Biosyst. Eng. 81, 237–247.
Palutikof, J.P., Wigley, T.M.L., 1996. Developing climate
change scenarios for the Mediterranean region. In: Jeftic, L.,
Keckes, S., Pernetta, J.C. (Eds.), Climatic Change and
the Mediterranean, vol. 2. Edward Arnold, London, pp. 27–
55.
Paranychianakis, N.V., 2001. Influence of rootstock, irrigation level
and recycled water on the growth, nutrition and physiology of
Soultanina grapevines. Ph.D. Thesis. Agricultural University of
Athens, pp. 202.
Paranychianakis, N.V., Aggelides, S., Angelakis, A.N., 2004a.
Influence of rootstock, irrigation level and recycled water on
the growth and yield of Soultanina grapevines. Agric. Water
Manage. 69, 13–27.
Paranychianakis, N.V., Chartzoulakis, K.S., Angelakis, A.N., 2004b.
Influence of rootstock, irrigation level and recycled water on
water relations and gas exchange of Soultanina grapevines.
Environ. Exp. Bot. 52, 185–198.
Prior, L.D., Grieve, A.M., Slavish, P.G., Gullis, P.R., 1992. Sodium
chloride and soil texture interactions in irrigated field grown
Sultana grapevines. II. Plant mineral content, growth and phy-
siology. Aust. J. Agric. Res. 43, 1067–1084.
Quick, W.P., Schurr, U., Scheibe, R., Schultze, E.D., Rodermel,
S.R., Bogorad, L., Stitt, M., 1991. Decreased ribulose-1,5-bipho-
sphate carboxylase-oxygenase in transgenic tobacco trans-
formed with antisense rdcS: impacts on photosynthesis in
ambient growth conditions. Planta 183, 542–554.
Rivelli, A.R., Lovelli, S., Perniola, M., 2002. Effects of salinity
on gas exchange, water relations and growth of sunflower
(Helianthus annuus). Funct. Plant Biol. 29, 1405–1415.
Robinson, M.F., Viery, A.A., Sanders, D., Mansfiels, T.A., 1997.
How can stomata contribute to salt tolerance. Ann. Bot. 80, 387–
393.
Romero-Aranda, R., Moya, J.L., Tadeo, F.R., Legaz, F., Primo-
Millo, E., Talon, M., 1998. Physiological and anatomical dis-
turbances induced by chloride salts in sensitive and tolerant
citrus: beneficial and detrimental effects of cations. Plant Cell
Environ. 21, 1243–1253.
Romero-Aranda, R., Syvertsen, J.P., 1996. The influence of
foliar applied urea nitrogen and saline solutions on net gas
exchange of Citrus leaves. J. Am. Soc. Hortic. Sci. 121, 501–
506.
Rosenzweig, C., Tubiello, F.N., 1997. Impacts of global climate
change on Mediterranean agriculture: current methodologies
and future directions. An introductory essay. Mitigation Adapta-
tion Strategies for Global Change 1, 219–232.
Ruiz, D., Martinez, V., Cerda, A., 1999. Demarcating specific ion
(NaCl, Cl, Na) and osmotic effects in the response of two citrus
rootstocks to salinity. Sci. Hortic. 80, 213–224.
Ruiz-Lozano, J.M., Azcon, R., 1995. Hyphal contribution to water
uptake in mycorrhizal plants as affected by the fungal species
and water status. Physiol. Plant. 95, 472–478.
Ruiz-Lozano, J.M., Azcon, R., Gomez, M., 1996. Alleviation of salt
stress by arbuscular mycorrhizal Glomus species in Lactuca
sativa plants. Physiol. Plant. 98, 767–772.

N.V. Paranychianakis, K.S. Chartzoulakis / Agriculture, Ecosystems and Environment 106 (2005) 171–187 187
Shalhevet, J., Levy, Y., 1990. Citrus trees. In: Stewart, B.A., Nielsen,
D.R. (Eds.), Irrigation of Agricultural Crops. American Society
of Agronomy, Madison, WI, pp. 951–986.
Sheveleva, E., Chmara, W., Bohnert, H.J., Jensen, R.G., 1997.
Increased salt and drought tolerance by D-ononitol production
in transgenic Nicotiana tabacum L.. Plant Physiol. 115, 1211–
1219.
Storey, R., 1995. Salt tolerance, ion relations and the effect of root
medium on the response of citrus to salinity. Aust. J. Plant. Phys.
22, 101–114.
Storey, R., Walker, R.R., 1999. Citrus and salinity. Sci. Hortic. 78,
39–81.
Subhakar, C., Lakshmi, A., Giridarakumar, S., 2001. Changes in the
antioxidant efficacy in two high yielding genotypes of mulberry
(Morrus alba L.) under NaCl salinity. Plant Sci. 161, 613–619.
Talbott, L.D., Zeiger, E., 1998. The role of sucrose in guard cell
osmoregulation. J. Exp. Bot. 49, 329–337.
Tattini, M., Gucci, R., Coradeschi, M.A., Ponzio, C., Everard, J.D.,
1995. Growth, gas exchange and ion content in Olea europaea
plants during salinity stress and subsequent relief. Physiol. Plant.
95, 203–210.
Tattini, M., Gucci, R., Romani, A., Baldi, A., Everard, J.D., 1996.
Changes in non-structural carbohydrates in olive (Olea europea)
leaves during root zone salinity stress. Physiol. Plant. 98, 117–
124.
Tester, M., Davenport, P., 2003. Na+ tolerance and Na+ transport in
higher plants. Ann. Bot. 91, 503–527.
Tyerman, S.D., Skerrett, I.M., 1999. Root ion channels and salinity.
Sci. Hortic. 78, 175–235.
UN Population Division, 1994. World population prospects
(sustaining water: an update). The 1994 Revision. The UN,
New York.
Walker, R.P., Torokfalvy, E., Scott, N.S., Kriedemann, P.E., 1981.
An analysis of photosynthetic response to salt treatment in Vitis
vinifera. Aust. J. Plant Physiol. 8, 359–374.
Walker, R.R., Blackmore, D.H., Clingeleffer, P.R., Iakono, F., 1997.
Effect of salinity and Ramsey rootstock on ion concentrations
and carbon dioxide assimilation in leaves of drip-irrigated, field-
grown grapevines (Vitis vinifera L. cv. Sultana). Aust. J. Grape
Wine Res. 3, 66–74.
Walker, R.R., Blackmore, D.H., Sun, Q., 1993a. Carbon dioxide
assimilation and foliar ion concentrations in leaves of lemon
(Citrus limon L.) trees irrigated with NaCl or Na2SO4. Aust. J.
Plant Physiol. 20, 173–185.
Walker, R.R., Munns, R., Tonnet, M.L., 1993b. Xylem chloride and
sodium concentrations of salt treated citrus plants. In: Beilby,
M.J., Walker, N.A., Smith, J.R. (Eds.), Membrane Transport in
Plants and Fungi. University of Sydney, Sydney, pp. 490–
494.
White, P.J., Broadley, M.R., 2001. Chloride in soils and its uptake
and movement within the plant: a review. Ann. Bot. 88, 967–
988.
Wingley, T.M.L., 1992. Future climate of the Mediterranean Basin
with particular emphasis on changes in precipitation. In: Jeftic,
L., Milliman, J.D., Sestini, G. (Eds.), Climatic Change and the
Mediterranean. Arnold, London, pp. 15–44.
Zekri, M., Parsons, L.R., 1990. Response of split-root Sour orange
seedlings to NaCl and polyethylene glycol stresses. J. Exp. Bot.
41, 35–40.
Zhang, H.X., Blumwald, E., 2001. Transgenic salt tolerant tomato
plants accumulate salt in the foliage but not in fruits. Nat.
Biotechnol. 19, 765–768.
Zhu, J.K., 2001. Plant salt tolerance. Trends Plant Sci. 6,
66–71.