inhibition of mouse bladder tumor proliferation by murine interferon-γ and its synergism with...
TRANSCRIPT

[CANCER RESEARCH 44, 4377-4381, October 1984]
Inhibition of Mouse Bladder Tumor Proliferation by Murine lnterferon-7and Its Synergism with Interferon-/?1
Timothy L. Ratliff,2 Dov Kadmon, Amos Shapiro,3 Allan J. Jacobs, and W. D. W. Heston
Department of Surgery, Division of Urology, Washington University School of Medicine at The Jewish Hospital of St. Louis [T. L R., D. K,, A. S.]; Urologie OncologyResearch Laboratory, Department of Surgery, Division of Urology [W. D. W. H.], anöDepartment oÃObstetrics and Gynecology [A. J. J.], Washington University Schoolof Mediane, St. Louis, Missouri 63110
ABSTRACT
We studied the effect of interferon-7 (IFN-7) and mouse L-cellInterferon (IFN-0) on the proliferation of a mouse bladder tumor,MBT-2. A liquid culture clonogenic assay was used, and a linearrelationship was obtained between the number of cells platedand the number of colonies formed. When the cells were assayedin the presence of various doses of murine IFN-7 or IFN-0, colonyformation was inhibited in a dose-dependent manner. Partiallypurified IFN-7 was more effective than IFN-0 in inhibiting MBT-2colony formation in that IFN-/3 exhibited a 50% inhibition dose of
approximately 1000 units/ml, while the 50% inhibition dose forthe partially purified IFN-7 was approximately 70 units/ml. The50% inhibition dose for recombinant IFN-7 was 700 units/ml,
suggesting that multiple lymphokines were active in the partiallypurified preparation. Further studies with partially purified IFN-7
showed that the inhibitory effect was time dependent with themaximal effect observed after 48 hr of exposure in a 5-dayassay. Treatment of partially purified IFN-7 for 24 hr at pH 2.0
resulted in the abrogation of the antiproliferative effect. Studiesin which partially purified IFN-7 preparations were treated with amonoclonal antibody against IFN-7 also resulted in abrogation
of antiproliferative activity, confirming the nature of the antiproliferative agent to be IFN-7. Further studies showed that murinerecombinant IFN-7 also inhibited MBT-2 proliferation in a dose-dependent manner, confirming that IFN-7 alone mediates antiproliferative activity. Combinations of IFN-0 and recombinantIFN-7 acted synergistically in the inhibition of MBT-2 proliferation.
INTRODUCTION
IFN4 was first described in 1957 by Isaacs and Lindenmann(16). Since the initial discovery, 3 major IFN classes, IFN-«,IFN-ß,and IFN-7, have been identified based on antigenic andphysicochemical properties (25). These IFNs have been shownto exert multiple biological functions, including antiviral, antitu-
mor, and immunomodulatory activity, as well as direct inhibitionof cellular proliferation (6,11, 22, 24).
Although the antiproliferative effects of IFN-a and IFN-/3 have
been studied extensively, studies on the antiproliferative activityof IFN-7 are more limited. Both IFN-«and IFN-/3 have been
shown to inhibit cell proliferation in monolayer cultures as well
1This work was supported by USPHS Grant CA 28860 from the National Cancer
Institute through the National Bladder Cancer Project.2To whom requests for reprints should be addressed.3Present address: Department of Urotogy, Hadassah University Hospital, Jeru
salem, Israel.4The abbreviations used are: IFN, Interferon; PBS, phosphate-buffered saline
[0.1 M K2HPO<:15 M NaCI (pH 7.2)]; PHA, phytohemagglutinin; CHO, Chinesehamster ovary; IDM, 50% inhibition of cellular proliferation; MEM, minimal essentialmedium; VSV, vesicular stomatitis virus.
Received March 19,1984; accepted July 3,1984.
as suspension cultures and cloned stem cell cultures (7, 14, 21,22). These IFNs are apparently not cytolytic, although some celldeath occurs when cultures are continuously exposed for morethan 48 hr (15). IFN-a and IFN-/3apparently inhibit DNA synthesisby shunting cells into the G0-Gi growth phase but do not signif
icantly alter RNA or protein synthesis (4, 19). Recent studiesusing electrophoretically pure (9) and recombinant IFN (8) preparations have shown that IFN-a and IFN-ßare the mediators of
the antiproliferative activity.Few studies have been performed assessing the effects of
IFN-7 on cellular proliferation. Ankel ef al. (1) showed that bothIFN-jS and IFN-7 inhibited the proliferation of mouse leukemiaL1210 cells. Moreover, IFN-7 was shown to inhibit the growth
of an L1210 subline that was resistant to the antiproliferativeeffects of IFN-0. The antiproliferative effect of IFN-7 was attributed to the presence of IFN-7 receptors on the IFN-/3-resistantL1210 cell line. Tyring ef al. (29) showed that IFN-7 was directlycytolytic to mouse leukemia cells, while IFN-0 inhibited prolifer
ation but was not cytolytic. Fleischmann (10) showed that apartially purified preparation of IFN-7 inhibited the proliferationof B-16 melanoma cells. Fleischmann suggested that IFN-7 wasa more potent inhibitor of cellular proliferation than was IFN-/3and further showed that IFN-7 and IFN-/3 inhibited proliferation
in a synergistic manner.Considerable variability in the susceptibility of tumor cells to
IFN-o and IFN-0 has been observed. These results range from
growth enhancement to substantial inhibition of cellular proliferation (30). In addition, differential inhibition of individual tumorsby IFN-a and IFN-/3 has been reported (3), suggesting that
individual tumors express varying susceptibilities to the antiproliferative effects of each class of IFN.
In this study, we evaluated the effects of IFN-0 and IFN-7 onthe proliferation of the MBT-2 mouse bladder tumor. Our resultssuggest that both IFN-/3 and IFN-7 inhibit MBT-2 proliferationwith partially purified IFN-7 being the more effective antiproliferative agent; however, studies with recombinant IFN-7 show thatthe concentration of recombinant IFN-7 required to produce 50%inhibition was equivalent to that of IFN-ß,suggesting that multiple
lymphokines were active in the partially purified preparation.Moreover, recombinant IFN-7 and IFN-0 in combination inhibitproliferation in a synergistic manner.
MATERIALS AND METHODS
Tumor. MBT-2 tumor cells were cultured in medium containing RPMI-
1640 solution supplemented with penicillin (100 units/ml), streptomycin(100 jig/ml), and 10% fetal calf serum (complete medium). Inoculation ofthe MBT-2 cell line s.c. produced tumors indistinguishable from MBT-2
tumors maintained by s.c. implanation. All tissue culture media andsupplements were purchased from K. C. Biologicals, Lenexa, KS.
OCTOBER 1984 4377
Research. on September 17, 2015. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

T. L Ratliff et al.
IFN Preparations. IFN-/3 was a gift from William E. Stewart II (Univer
sity of South Florida, School of Mediane, Tampa, FL), which wasprepared by stimulation of L929 cells with Newcastle disease virus andpurified to a specific activity of approximately 106 units/mg protein. IFN
produced by L-cells is a combination of both «and fi, with the ratio
produced being dependent on the method of induction and the purification schema, and is designated as IFN-/3.
Partially purified IFN-7 was prepared in our laboratory by stimulation
of either C3H/H or C57BL/6 splenocytes with PHA (5 ^g/ml; BurroughsWellcome). Supernatants were collected 24 hr later and concentrated oncontrol pore glass beads (Nuclear Electronics). Control pore glass beadconcentration effectively removed detectable PHA from the preparationand provided an IFN-7 preparation with a specific activity of approximately 10* units/mg protein. Mock IFN was prepared in an identical
manner except that splenocytes were cultured in the absence of PHA.Recombinant IFN-7 was kindly provided by Dr. Patrick M. Gray
(Genentech, Inc., South San Francisco, ÇA). Recombinant IFN-7 was
prepared from a mammalian cell expression system similar to the monkeyCOS ce!l/SV40 promoter system described for expression of humanIFN-7 (13) and murine IFN-7 (12). The expression plasmid was modifiedby insertion of gene-encoding dihydrofolate reducÃase, which allows
selection of transfected CHO cells in the presence of methotrexate (18,23). The recombinant murine IFN-7 used in these experiments consistedof serum-free supematants from CHO cells. Controls included serum-
free supernatants from nontransfected CHO cells. The antiviral activitywas monitored biweekly to monitor stability. The preparation was stablewhen stored at 4°.
Acid Treatment of IFN-7. An IFN-7 preparation was separated into 2aliquots containing sufficient quantities of IFN-7 for performance of the
clonogenic assay. One sample was dialyzed against glycine buffer (pH2.0) for 24 hr at 4°.The second sample was dialyzed against PBS under
identical conditions. Both samples were then dialyzed against RPMI-1640 and assayed for IFN activity. Acid treatment reduced IFN-7 levelsto <10 units/ml, while IFN-/3 titers were not affected significantly. ControlIFN-7 titers were not significantly affected by dialysis against PBS.
Antibody Neutralization. The monoclonal antibody to IFN-7 was
kindly provided by Dr. Robert D. Schreiber (Scripps Clinic and ResearchFoundation, La Jolla, CA). One ng of this antibody will neutralize approximately 10 units of IFN-7. Neutralization was performed as describedpreviously (20). Briefly, anti-IFN-7 in 5-fold excess was mixed with IFN-
7 sample, incubated 1 hr at room temperature, and added to the assaysystem.
IFN Assay. The IFN assay was performed as described previously(20). Briefly, freshly trypsinized L929 cells (10* cells/100 nO were addedto each well of a 96-well microtiter plate and incubated for 2 days at 37°
in 5% CO2. After incubation, serial 1:5 dilutions of each IFN sample wereprepared in serum-free MEM, and 100 ^l of each were added to appropriate wells. Controls included a laboratory IFN-0 standard (standardizedagainst NIH Reference Standard G002-904-511 ), VSV, and L929.
The plates were incubated 24 hr at 37°,after which they were washed
with MEM, and 200 plaque-forming units of VSV in MEM containing
2.5% serum were added to appropriate wells. The plates were incubatedat 37°for approximately 48 hr, which provided sufficient time for VSV
controls to show 100% cytopathic effect. IFN titers are reported as thereciprocal of the dilution providing 50% protection. One NIH referenceunit is equivalent to approximately 1 unit in our assay system.
In Vitro Clonogenic Assay. Single cell suspensions of MBT-2 cultures
were prepared by mechanical disaggregation (gentle pipeting followedby serial passage through progressively smaller needles), and viabilitywas determined by trypan blue exclusion.
For the clonogenic assay, monodispersed viable cells (3 x 103 cells/
dish) were plated in 60-mm dishes (Falcon No. 3002) in RPMI-1640
supplemented with 10% calf serum in a volume of 5 ml/dish. No agar orfeeder layers were used. The dishes were then incubated for 5 days at37°,7% CO2 in air, and saturation humidity. At the end of the incubation
period, the medium was discarded, and the adherent cells were dried in
air and fixed in methanol for 5 min. Following staining in Harris' hema-
toxylin (Fisher Scientific Co., Pittsburgh, PA) for 3 min, the dishes werewashed in tap water for 1 min and then dried. Distinct colonies (>50cells) were counted at x20.
A linear relationship was obtained between the number of viable cellsplated and the number of colonies formed after 5 days in culture, whenthe number of cells plated was in the range of 102 to 3 x 103 cells/dish
(Chart 1). Cloning efficiency (number of colonies formed/number of cellsplated) was approximately 8 to 18%.
IFN preparations were added to the cultures at the time of plating andremained for the duration of the assay unless stated otherwise. Thepercentage of inhibition was calculated as follows.
% of inhibition 1 -a-o
x 100
where a is control colony number, and b is experimental colony number.Because of reagent limitations, IFN-7 neutralization studies were
performed in 96-well microtiter plates (Flow Laboratories) as describedbelow. Viable MBT-2 cells were plated at a concentration of 104 cells/
well. IFN preparations were added, and cultures were incubated for 4days at 37°in 7% CO2 and saturated humidity. IFN-7 preparations were
neutralized immediately prior to addition to the antiproliferative assay asdescribed above. Controls included addition of anti-IFN-7 antibody only,
CHO control medium, and a standard medium control. After incubation,the plates were rinsed twice with PBS and stained with 0.5% solution ofcrystal violetfixative solution containing 5% formalin, 50% ethanol, and0.15 M NaCI. The plates were rinsed extensively with tap water and airdried overnight. The stain was extracted with 200 ^ of 2-methoxy-
ethanol, and the absorbance was determined on a spectrophotometerat 570 nm. The anti-IFN-7 and CHO control medium had no effect onthe proliferation of MBT-2 cells. The percentage of inhibition was calcu
lated as above from absorbance values.
RESULTS
Effect of Partially Purified IFN-7 and IFN-0 on MBT-2 ColonyFormation. In these experiments, 3 x 103 MBT-2 cells were
plated and exposed for 5 days to IFN (62 to 1000 units/ml). Theresults of a representative experiment are shown in Chart 2A.Control cultures were grown in the presence of a mock IFN-7preparation which consistently yielded colony counts similar tothose obtained in untreated control cultures (see Chart 4). Theresults show that the ID50for partially purified IFN-7 was approximately 70 units/ml, while the IDso for IFN-/3 was approximately1000 units/ml. The inhibitory activity of the partially purified IFN-7 preparation approached 100% in multiple experiments (>10).
Correlation of the Time of Exposure to IFN-7 to Its InhibitoryEffect. In these experiments, MBT-2 cultures were treated with500 units of IFN-7 for 3, 6, 24, 48, or 120 hr. At the terminationof each exposure time, cultures were washed free of IFN-7
UX> IOOO 20OO 300ONUMBER VIABLE MBT-2 CELLS CULTURED
Chart 1. Linear relationshipbetween the number of MBT-2 cells plated and thenumber of colonies in a clonogenicassay. Points, mean; oars, S.E.
4378 CANCER RESEARCH VOL. 44
Research. on September 17, 2015. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from
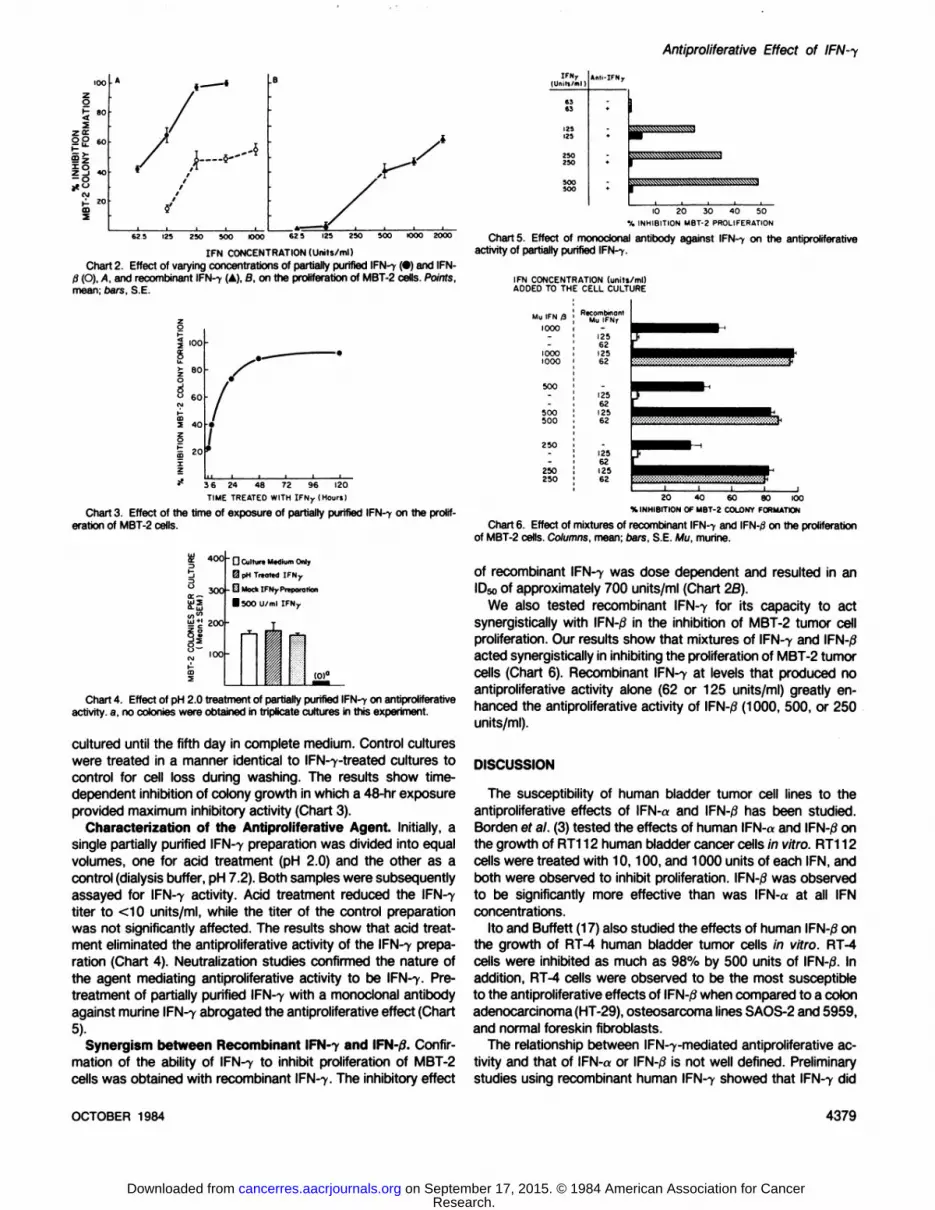
Antiproliferative Effect of
« 80
zu:2£ 60
55
IFNr(Unin/ml)
625 125 25O 5OO OOO 125 250 500 1000 2OOO
IFN CONCENTRATION(Units/ml)Chart 2. Effect of varying concentrations of partially purified IFN-i (•)and IFIM-
/3 (O), A, and recombinant IFN-r (A), B, on the proliferation of MBT-2 cells. Points,
mean; bars, S.E.
** 36 24 48 72 96 120
TIME TREATED WITH IFNy (HoursI
Chart 3. Effect of the time of exposure of partially purified IFN-f on the proliferation of MBT-2 cells.
£400L*»uj
5<n"jy*'
zooZo0^«
'»Ot^-<DS^
G CultureMédiumOnly0
pH Treated IFNy
- O MachIFNj-Preparation•
500 U/mlIFNr_,E|i(0)«H
Chart 4. Effect of pH 2.0 treatment of partially purified IFN--»-on antiproliferative
activity, a, no colonies were obtained in triplicate cultures in this experiment.
cultured until the fifth day in complete medium. Control cultureswere treated in a manner identical to IFN-7-treated cultures tocontrol for cell loss during washing. The results show time-dependent inhibition of colony growth in which a 48-hr exposure
provided maximum inhibitory activity (Chart 3).Characterization of the Antiproliferative Agent. Initially, a
single partially purified IFN-7 preparation was divided into equal
volumes, one for acid treatment (pH 2.0) and the other as acontrol (dialysis buffer, pH 7.2). Both samples were subsequentlyassayed for IFN-7 activity. Acid treatment reduced the IFN-7
titer to <10 units/ml, while the titer of the control preparationwas not significantly affected. The results show that acid treatment eliminated the antiproliferative activity of the IFN-7 prepa
ration (Chart 4). Neutralization studies confirmed the nature ofthe agent mediating antiproliferative activity to be IFN-7. Pre-treatment of partially purified IFN-7 with a monoclonal antibodyagainst murine IFN-7 abrogated the antiproliferative effect (Chart
5).Synergism between Recombinant IFN-7 and IFN-/1 Confir
mation of the ability of IFN-7 to inhibit proliferation of MBT-2cells was obtained with recombinant IFN-7. The inhibitory effect
125125
»nli-IFN,.
10 20 30 40 50% INHIBITION MBT-2 PROLIFERATION
Chart 5, Effect of monoclonal antibody against IFN-7 on the antiproliferativeactivity of partially purified IFN-f.
IFN CONCENTRATION (units/ml)ADDED TO THE CELL CULTURE
Mu IFN /31000
10001000
500500
250250
I20 40 60 80 100
% INHIBITION OF MBT-2 COLONY FORMATION
Chart 6. Effect of mixtures of recombinant IFN-> and IFN-0 on the proliferationof MBT-2 cells. Columns, mean; oars, S.E. Mu, murine.
of recombinant IFN-7 was dose dependent and resulted in an
IDsoof approximately 700 units/ml (Chart 2B).We also tested recombinant IFN-7 for its capacity to act
synergistically with IFN-|8 in the inhibition of MBT-2 tumor cellproliferation. Our results show that mixtures of IFN-7 and IFN-0acted synergistically in inhibiting the proliferation of MBT-2 tumorcells (Chart 6). Recombinant IFN-7 at levels that produced no
antiproliferative activity alone (62 or 125 units/ml) greatly enhanced the antiproliferative activity of IFN-/3 (1000, 500, or 250
units/ml).
DISCUSSION
The susceptibility of human bladder tumor cell lines to theantiproliferative effects of IFN-a and IFN-0 has been studied.Borden ef al. (3) tested the effects of human IFN-a and IFN-0 on
the growth of RT112 human bladder cancer cells in vitro. RT112cells were treated with 10,100, and 1000 units of each IFN, andboth were observed to inhibit proliferation. IFN-0 was observedto be significantly more effective than was IFN-a at all IFN
concentrations.Ito and Buffett (17) also studied the effects of human IFN-0 on
the growth of RT-4 human bladder tumor cells in vitro. RT-4cells were inhibited as much as 98% by 500 units of IFN-/3. Inaddition, RT-4 cells were observed to be the most susceptibleto the antiproliferative effects of IFN-0 when compared to a colonadenocarcinoma(HT-29), osteosarcoma lines SAOS-2 and 5959,
and normal foreskin fibroblasts.The relationship between IFN-7-mediated antiproliferative ac
tivity and that of IFN-a or IFN-/3 is not well defined. Preliminarystudies using recombinant human IFN-7 showed that IFN-7 did
OCTOBER 1984 4379
Research. on September 17, 2015. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

T. L. Ratliff et al.
not inhibit mitogen-induced proliferation of peripheral blood lymphocytes, although both IFN-«and IFN-0 were inhibitory (5).Ankel ef al. (1) compared the antiproliferative activities of IFN-,8and IFN-7 using L1210 tumor cells. Two variants were used,
one which was sensitive, L1210S, to the antiproliferative effectsof IFN-/3 and the other, L1210R, which was resistant to theantiproliferative effects of IFN-0. Both IFN-/3 and IFN-7 inhibited
L1210S proliferation in vitro, and the inhibitory activity for bothwas approximately 50% at 1000 units/ml. In addition, IFN-7 wasobserved to inhibit the proliferation of the IFN-0-resistant variant,
L1210R. These investigators attributed the differential susceptibility of L1210R to the presence or absence of cell surfacereceptors. Zhang ef al. (31) compared both human IFN-7 andmurine IFN-7 preparations with IFN-«of the respective species
for their relative antiproliferative activities against several celllines. The number of antiviral units required to produce an IDsowas lower for IFN-7 than either IFN-«or IFN-/3 for all cell linestested. Blalock ef al. (2) obtained similar results with human IFN-«,IFN-0, and IFN-7 against WISH and HEp-2 cells. IFN-7 was20 to 100 times more potent per antiviral unit than either IFN-«or IFN-0.
Our results show that both IFN-7 and IFN-,8 inhibit MBT-2stem cell proliferation. The inhibitory activity of both IFN-7 andIFN-fJ was dose dependent, and IFN-7-mediated inhibition required a minimum of 48-hr exposure for maximal expression.Partially purified IFN-7 appeared to mediate more antiproliferativeactivity based on antiviral units than did IFN-0. The IDso forpartially purified IFN-7 was approximately 70 units/ml, while theIDso for IFN-/3 was 1000 units/ml. Interestingly, the ID50 forrecombinant IFN-7 was 700 units/ml, which was 10-fold greater
than that of the partially purified preparation. This suggests thatthe partially purified preparation contains other lymphokines thatpotentiate the effects of IFN-7, although alone, they do not
appear to inhibit proliferation (Chart 5, antibody neutralizationstudy). In this regard, a recent report by Stone-Wolff ef al. (26)showed that the antiproliferative effects of human IFN-7 were
potentiated by lymphotoxin. Further studies will be required todetermine whether a similar relationship is present in our system.
Fleischmann and coworkers (10, 11) showed that partiallypurified murine IFN-7 potentiated the antitumor effects of IFN-/3both in vivo and in vitro. In the in vivo studies, IFN-7 (25 units/day), which alone had no effect on the growth of P388 lympho-cytic leukemia tumors, significantly enhanced the growth-inhibitory capacity of IFN-/3 (11). Similar synergism with partially purified IFN-7 was reported for in vitro antiproliferative assays (10).De Clercq ef al. (6) reported similar in vivo observations withmixtures of IFN-7 and IFN-,3against L1210 leukemia cells. These
investigators, however, suggested that the synergism occurredonly at suboptimal concentrations of IFN-0. Our results in an invitro cell proliferation assay using recombinant IFN-7 confirm thesynergistic effects of mixtures of partially purified murine IFN-7and IFN-0. The addition of recombinant IFN-7 in noninhibitoryconcentrations potentiates the antiproliferative activity of IFN-ß.
Unlike the in vivo data reported by De Clercq ef al. (6), synergismwas observed at both optimal and suboptimal concentrations ofIFN-/3.
Taken together, these results suggest that therapy with IFNseither alone or in combination may be an effective means oftreating bladder tumors; however, the uniformity of the susceptibility of bladder tumor cells to combinations of IFNs needs tobe established.
Previous studies have shown that various cell lines havedifferent sensitivities to the antiproliferative effects of IFN-o, IFN-jS,and IFN-7 as well as subtypes of IFN-«(3, 8, 28). Tsuruo ef
al. (28) examined the susceptibility of several mouse cell lines tothe antiproliferative activity of IFN-0. IFN concentrations ranging
from 3.3 to 10,000 units/ml were tested in an assay in whichcells were exposed to IFN-0 for 3 days and further cultured in
fresh medium for 2 days. The results are reported as the concentration of IFN-/3 required to inhibit cell growth by 50%. The
number of units required for 50% inhibition ranged from 10.5units for B16 melanoma to 10,000 units for a colon carcinoma,designated colon 26, while other cell lines were not sensitive tothe antiproliferative effects of IFN-,8.
Human tumor cell lines also have been shown to vary in theirresponse to IFN. Von Hoff ef al. (30) tested the inhibitory effectsof human IFN-/3 on 62 tumors with a variety of origins in a softagar clonogenic assay. Cells were exposed to IFN-j3 (1000 units/
ml), which was included in the soft agar medium containing thecells. IFN-0 enhanced the growth of 3 tumors by 39, 82, and
141% of controls. Others also have shown enhanced tumorgrowth in the presence of IFN-0 as well as IFN-«(27). The overall
response reported by Von Hoff ef al. (30) varied considerablywith inhibition ranging from 0 to 90% in a series of 21 tumorsfor which dose-response data were obtained. Of these, 10 of 21were not inhibited by IFN-0, while 4 expressed 2 to 10% inhibi
tion, one was inhibited approximately 20%, 3 approximately40%, one approximately 60%, and 2 approximately 80 to 90%.
In conclusion, our results show that partially purified preparations of IFN-7 are more effective than IFN-,8 in inhibiting theproliferation of MBT-2 cells. Antibody neutralization studies demonstrated that such inhibition required IFN-7. A minimum expo
sure time of 48 hr was required for maximal inhibitory activity.Recombinant IFN-7 also was inhibitory, demonstrating that no
other factors are required for the antiproliferative effect. Furthermore, recombinant IFN-7 potentiated the antiproliferative effectof IFN-,8.
REFERENCES
1. Ankel, H., Krishnamurti,C., Besancan,F., Stefanos, S., and Falcoff, E. Mousefibroblast (type I) and immune (type II) interférons:pronounced differences inaffinity for gangliosides and in antiviral and antigrowth effects on mouseleukemiaL1210R cells. Proc. Nati. Acad. Sci. USA, 77: 2528-2532,1980.
2. Blalock, J. E., Georgiades,J. A., Langford, M. P., and Johnson, H. M. Purifiedhuman immune interferon has more potent anticellularactivity than fibroblastsor leukocyte interferon. Cell. Immunol.,49: 390-394,1980.
3. Borden, E. C., Mögen,T. F., and Voelkel, J. G. Comparativeantiproliferativeactivity in vitro of natural interferons-a and -0 for diploid and transformedhumancells. Cancer Res., 42: 4948-4953,1982.
4. Brouty-Boye, D., Macieira-Coelho,A., Fiszman, M., and Gresser, I. Effect ofinterferon on macromolecularsynthesis in L1210 cells in vitro. Int. J. Cancer12: 250-258,1973.
5. Czamiecki, C. W., and Fennie, C. W. Potentiatkjn of cell growth inhibitoryeffects with combinations of human recombinant alpha, beta, and gammainterferons (abstract). Proceedingsof the Third Annual InternationalCongressfor Interferon Research,p. 39. New York: Mary Ann LJebert,Inc., 1982.
6. De Clercq, E., Zhang, Z-X., and Huygen, K. Synergismin the antitumor effectsof type I and type II interferon in mice inoculated with leukemia L1210 cells.Cancer Lett., i 5: 223-228, 1982.
7. Epstein, L. B.. and Marcus, S. G. Review of experience with interferon anddrug sensitivity testing of ovarian carcinoma in semisolidagar culture. CancerChemother. Pharmacol.,6: 273-277, 1981.
8. Evinger, M., Maeda, S., and Pestka, S. Recombinanthuman leukocyte interferon produced in bacteria has antiproliferative activity. J. Bioi. Chem 256:2113-2114,1981.
9. Evinger, M., Rubinstein, M., and Pestka, S. Antiproliferative and antiviralactivitiesof human leukocyte interferons. Arch. Biochem. Biophys.,210: 319-329,1981.
4380 CANCER RESEARCH VOL. 44
Research. on September 17, 2015. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

10. Fleisctimann, W. R. Potentiation of direct anticellular activity of mouse interferons: mutual synergism and interferon concentration dependence. Cancer Res.,42:869-875,1982.
11. Fleischmann, W. R., Kleyn, K. M., and Baron, S. Potentiation of antitumoreffect of virus-induced interferon by mouse immune interferon preparations. J.Nati. Cancer Inst., 65: 963-966,1980.
12. Gray, P. W., and Goeddel, D. V. Cloning and expression of murine immuneinterferon cDNA. Proc. Nati. Acad. Sci. USA. 80: 5842-5846,1983.
13. Gray, P. W., Leung, D. W., Pennica, D., Yelverton, E., NajarÃan,R., Semonsen,C. C., Derynck, R., Sherwood, P. J., Wallace, D. M., Berger, S. L., Levinson,A. D., and Goeddel, D. V. Expression of human immune ¡nterferoncDNA in E.coli and monkey cells. Nature (Lond.), 295: 503-508,1982.
14. Gresser, I., Brouty-Boye, D., Thomas, M. T., and Macieira-Coelho, A. Interferonand cell division. I. Inhibition of the multiplication of mouse leukemia L1210cells in vitro by interferon preparations. Proc. Nati. Acad. Sci. USA, 66:1052-
1058,1970.15. Gresser, I., and Tovey, M. G. Antitumor effects of interferon. Btochim. Biophys.
Acta, 516:231-247,1978.16. Isaacs, A., and LJndenmann, J. Virus interference: the ¡nterferon.Proc. R. Soc.
Lond. B Biol. Sci., 147: 258-267, 1957.17. Ito, M., and Buffett, R. F. Cytocidal effect of purified human fibroWast ¡nterferon
on tumor cells in vitro. J. Nati. Cancer Inst., 66: 819-825,1981.18. Lee, F., Mulligan, R., Berg, P., and Ringold, G. Glucocorticoids regulate
expression of dihydrofolate reducÃasecONA in mouse mammary tumour viruschimaeric plasmids. Nature (Lond.), 294: 228-232,1981.
19. Macieira-Coelho, A., Brouty-Boye, D., Thomas, M. T., and Gresser, I. Interferonand cell division. III. Effect of interferon on the division cycle of L1210 cells invitro. J. Cell. Biol., 48: 415-419, 1971.
20. McCool, R. E., Catalona, W. J., Langford, M. P., and Ratliff, T. L. Induction ofhuman gamma interferon by Protein A from Staphylococcus aureus. J. Interferon Res., 1: 473-481, 1981.
21. Panniers, L. R. V., and Clemens, M. J. Inhibition of cell division by interferon:
Antiproliferative Effect of IFN-y
changes in cell cycle characteristics and in morphology of Ehrlich ascites tumorcells in culture. J. Cell. Sci., 48: 259-279,1981.
22. Paucker, «.,Cantell, K., and Henle, W. Quantitative studies on viral interferencein suspended L cells. III. Effect of interfering viruses and interferon on thegrowth rate of cells. Virology, 17: 324-334,1962.
23. Simonsen, C. D., and Levinson, A. D. Isolation and expression of an alteredmouse dihydrofolate reducÃasecDNA. Proc. Nati. Acad. Sci. USA, 80: 2495-2499,1983.
24. Sonnenfeld, G., and Merigan, T. C. A regulatory role for interferon in immunity.Ann. N. Y. Acad. Sci., 77: 345-354,1979.
25. Stewart, W. E., II. Interferons: their purification and characterization. In: TheInterferon System. Berlin: Springer-Verlag, 1979.
26. Stone-Wolff, D. S., Yip, Y. K., Kelker, H. C., Le, J., Henrikson-Destefano, D.,Rubin, B. Y., Rinderknecht, E., Aggarwar, B. B., and Vilcek, J. Interrelationships of human interferon-gamma with lymphotoxin and monocyte cytotoxin.J. Exp. Med., 759: 828-843, 1984.
27. Taetle, R., Buick, R. N., and McCulloch, E. A. Effect of interferon on colonyformation in culture by blast cell progenitors in acute myeloblastic leukemia.Blood, 56: 549-552,1980.
28. Tsuruo, T., lida, H., Tsukagoski, S., Oku, T., and Kishlda, T. Different susceptibilities of cultured mouse cell lines to mouse interferon. Gann, 73: 42-47,1982.
29. Tyring, S., Kumpel, G. P., Fleischmann, W. R., and Baron, S. Direct cytolysisby partially purified preparations of immune ¡nterferon.Int. J. Cancer, 30: 59-64,1982.
30. Von Hoff, D. D., Gutterman, J., Portnoy, B., and Coltman, C. A., Jr. Activity ofhuman leukocyte interferon in a human tumor cloning system. Cancer Chem-other. Pharmacd., 8. 99-103,1982.
31. Zhang, Z-X., De Clercq, E., Heremans, H., Vertiaegen-Lewalle, M., and Con
tent, J. Antiviral and anticellular activities of human and murine type I and typeII interferons in human cells monosemic, disomic, and trisomie for chromosome21. Proc. Soc. Exp. Btol. Med., Õ70:103-111, 1982.
OCTOBER 1984 4381
Research. on September 17, 2015. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

1984;44:4377-4381. Cancer Res Timothy L. Ratliff, Dov Kadmon, Amos Shapiro, et al.
β and Its Synergism with Interferon-γInterferon-Inhibition of Mouse Bladder Tumor Proliferation by Murine
Updated version
http://cancerres.aacrjournals.org/content/44/10/4377
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on September 17, 2015. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from