immunopathogenesis of psoriasis
TRANSCRIPT

Immunopathogenesis of psoriasis
Robert Sabat1, Sandra Philipp1, Conny Hoflich2, Stefanie Kreutzer1, Elizabeth Wallace1,
Khusru Asadullah3, Hans-Dieter Volk2, Wolfram Sterry4 and Kerstin Wolk1
1Interdisciplinary Group of Molecular Immunopathology, Dermatology ⁄ Medical Immunology, University Hospital Charite, Berlin, Germany;2Institute of Medical Immunology, University Hospital Charite, Berlin, Germany;3CRBA Dermatology, Schering AG, Berlin, Germany;4Department of Dermatology and Allergy, University Hospital Charite, Berlin, Germany
Correspondence: Dr Robert Sabat, Interdisciplinary Group of Molecular Immunopathology, Dermatology ⁄ Medical Immunology, Campus Charite
Mitte, University Hospital Charite, Chariteplatz 1, Berlin D-10117, Germany, Tel.: +49 30 450 518 009, Fax: +49 30 450 518 964,
e-mail: [email protected]
Accepted for publication 11 July 2007
Abstract: Psoriasis is a chronic skin disease that affects about
1.5% of the Caucasian population and is characterized by typical
macroscopic and microscopic skin alterations. Psoriatic lesions are
sharply demarcated, red and slightly raised lesions with silver-
whitish scales. The microscopic alterations of psoriatic plaques
include an infiltration of immune cells in the dermis and
epidermis, a dilatation and an increase in the number of blood
vessels in the upper dermis, and a massively thickened epidermis
with atypical keratinocyte differentiation. It is considered a fact
that the immune system plays an important role in the
pathogenesis of psoriasis. Since the early 1990s, it has been
assumed that T1 cells play the dominant role in the initiation and
maintenance of psoriasis. However, the profound success of anti-
tumor necrosis factor-a therapy, when compared with T-cell
depletion therapies, should provoke us to critically re-evaluate the
current hypothesis for psoriasis pathogenesis. Recently made
discoveries regarding other T-cell populations such as Th17 and
regulatory T cells, dendritic cells, macrophages, the keratinocyte
signal transduction and novel cytokines including interleukin
(IL)-22, IL-23 and IL-20, let us postulate that the pathogenesis of
psoriasis consists of distinct subsequent stages, in each of them
different cell types playing a dominant role. Our model helps to
explain the varied effectiveness of the currently tested immune
modulating therapies and may enable the prediction of the success
of future therapies.
Key words: autoimmunity – cytokine – inflammation
Please cite this paper as: Immunopathogenesis of psoriasis. Experimental Dermatology 2007; 16: 779–798.
Introduction
Psoriasis vulgaris (a.k.a. psoriasis in the rest of this review)
is a common, chronic, relapsing skin disease. It is charac-
terized by macroscopic (clinical) and corresponding micro-
scopic (histological) skin alterations and leads to
considerable impairment of the quality of life of the
affected patients. Moreover, special forms of psoriasis (e.g.
arthropathic form) can be accompanied by severe extra-
cutaneous changes. In addition to the disease frequency
and patients’ disability, researchers have concentrated on
psoriasis because of the postulated pathogenesis commonal-
ity of this disorder with other chronic immune-mediated
inflammatory diseases. In fact, similar reactions are hypoth-
esized to contribute to the initiation and maintenance of
diseases such as rheumatoid arthritis and Crohn’s disease,
and it is hoped that an improved understanding of the
pathogenesis of psoriasis will aid in the understanding of
other chronic inflammatory diseases and lead to novel
treatment options.
Psoriasis is a worldwide occurring disease. In the Cauca-
sian population, the prevalence is about 1.5% (1,2). It
means that in Europe alone, more than 7 million people
are afflicted with this disease. In other ethnic groups such
as the Japanese, the prevalence of psoriasis is much lower
(3,4). Epidemiological studies revealed that a distinct group
of disorders is quite frequently associated with psoriasis,
e.g. rheumatoid arthritis, colitis, diabetes, metabolic syn-
drome and hypertension. In contrast, atopic dermatitis and
allergies are less frequently seen to be associated with psori-
asis compared with normal rates of occurrence (5). Inter-
estingly, in contrast to atopic dermatitis, the reported
incidence of psoriasis has not increased in the last 20–
30 years. Men and women are affected by psoriasis at the
same rate. The first manifestation of the disorder usually
occurs around the age of 20 or between 50 and 60. How-
ever, it must be emphasized that psoriasis can manifest
itself at any age. According to Henseler and Christophers,
psoriasis can be differentiated into two subgroups: type I,
which begins before age 40, and type II, which begins after
DOI:10.1111/j.1600-0625.2007.00629.x
www.blackwellpublishing.com/EXDReview Article
ª 2007 The Authors
Journal compilation ª 2007 Blackwell Munksgaard, Experimental Dermatology, 16, 779–798 779
age 40 (6). Type I psoriasis, which accounts for approxi-
mately 75% of all psoriasis patients, is associated with a
more severe course of disease, limited success of treatment,
increased prevalence of certain human leucocyte antigen
(HLA)-types and stronger hereditary ties. Although the
inheritance pattern is currently still unclear, genetic dispo-
sition appears to play an important role in the susceptibil-
ity to develop psoriasis. This view is based on three
observations. First, the likelihood of developing psoriasis is
raised when first-grade relatives suffer from the disease.
The risk is about 20% if one parent has psoriasis, and is
about 75% if both parents are affected. If one monozygotic
twin suffers from psoriasis, the probability is more than
55% that the other will be affected too (4,7). Second, psori-
asis is associated with certain HLA-types (HLA-Cw6, HLA-
B13, HLA-B17, HLA-Bw57 and HLA-DR4). People with
HLA-Cw6, for example, have a 10-fold higher risk of dis-
ease (4,5). Thirdly, several psoriasis susceptibility loci have
been described. The PSORS1 in the major histocompatibil-
ity complex (MHC) region on chromosome 6 (6p21)
appears to be associated with most cases of psoriasis. Inter-
estingly, some genes from this region are associated also
with other immune diseases (rheumatoid arthritis, colitis
and diabetes) (3,8).
An infection with b-haemolytic streptococci often pre-
cedes the first manifestation of psoriasis (9). The course of
the disease is then chronic whereby the disease length,
intensity and recurrence are very different between patients
and sometimes even in the same patient. Consecutive new
exacerbations can be triggered by mechanical irritation (so-
called Koebner reaction; new lesions emerge at locations
that are mechanically irritated), medications (e.g. b-recep-
tor blockers, lithium, chloroquine, non-steroidal anti-
inflammatory agents, tetracyclines and interferons) and
infections (e.g. viral or bacterial infections) (10–12). Inter-
estingly, a positive Koebner reaction predicts subsequent
disease activity (10).
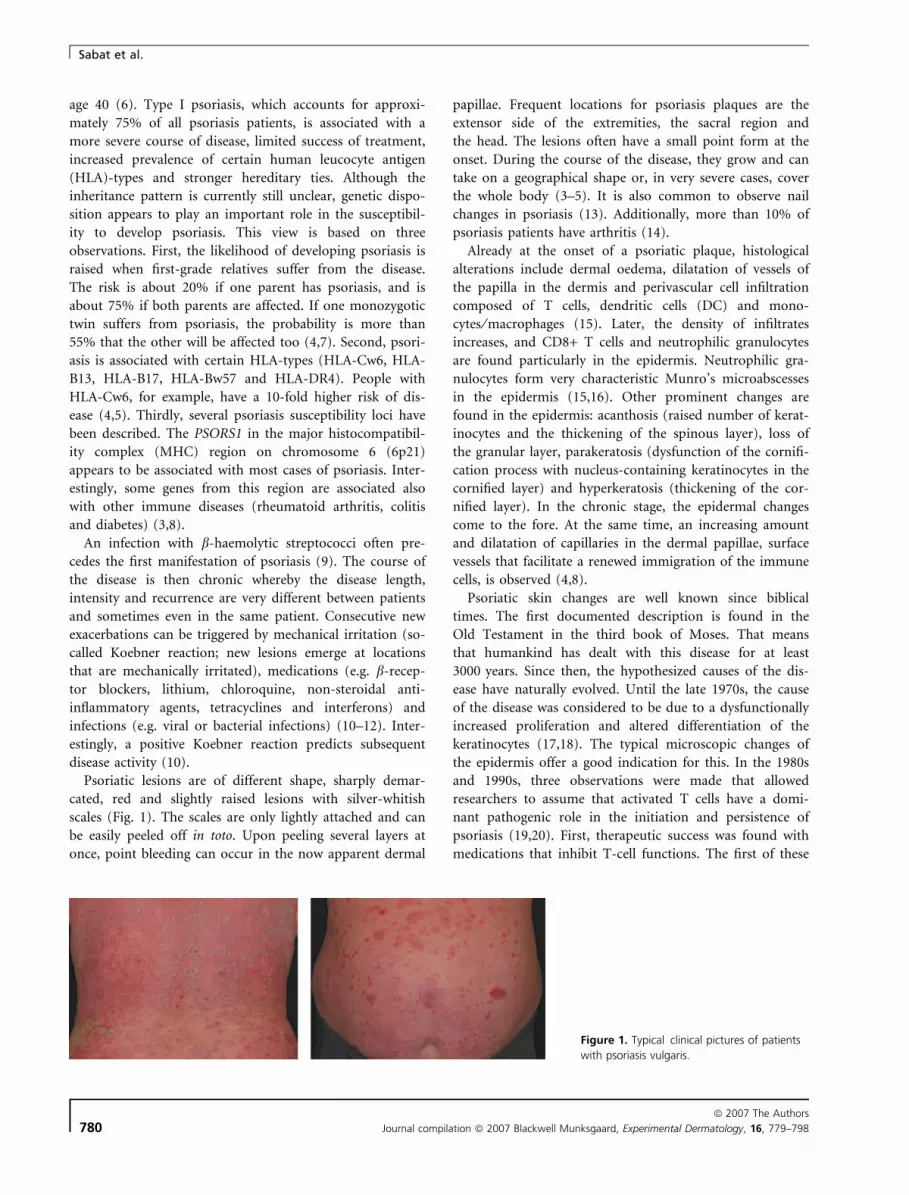
Psoriatic lesions are of different shape, sharply demar-
cated, red and slightly raised lesions with silver-whitish
scales (Fig. 1). The scales are only lightly attached and can
be easily peeled off in toto. Upon peeling several layers at
once, point bleeding can occur in the now apparent dermal
papillae. Frequent locations for psoriasis plaques are the
extensor side of the extremities, the sacral region and
the head. The lesions often have a small point form at the
onset. During the course of the disease, they grow and can
take on a geographical shape or, in very severe cases, cover
the whole body (3–5). It is also common to observe nail
changes in psoriasis (13). Additionally, more than 10% of
psoriasis patients have arthritis (14).
Already at the onset of a psoriatic plaque, histological
alterations include dermal oedema, dilatation of vessels of
the papilla in the dermis and perivascular cell infiltration
composed of T cells, dendritic cells (DC) and mono-
cytes ⁄ macrophages (15). Later, the density of infiltrates
increases, and CD8+ T cells and neutrophilic granulocytes
are found particularly in the epidermis. Neutrophilic gra-
nulocytes form very characteristic Munro’s microabscesses
in the epidermis (15,16). Other prominent changes are
found in the epidermis: acanthosis (raised number of kerat-
inocytes and the thickening of the spinous layer), loss of
the granular layer, parakeratosis (dysfunction of the cornifi-
cation process with nucleus-containing keratinocytes in the
cornified layer) and hyperkeratosis (thickening of the cor-
nified layer). In the chronic stage, the epidermal changes
come to the fore. At the same time, an increasing amount
and dilatation of capillaries in the dermal papillae, surface
vessels that facilitate a renewed immigration of the immune
cells, is observed (4,8).
Psoriatic skin changes are well known since biblical
times. The first documented description is found in the
Old Testament in the third book of Moses. That means
that humankind has dealt with this disease for at least
3000 years. Since then, the hypothesized causes of the dis-
ease have naturally evolved. Until the late 1970s, the cause
of the disease was considered to be due to a dysfunctionally
increased proliferation and altered differentiation of the
keratinocytes (17,18). The typical microscopic changes of
the epidermis offer a good indication for this. In the 1980s
and 1990s, three observations were made that allowed
researchers to assume that activated T cells have a domi-
nant pathogenic role in the initiation and persistence of
psoriasis (19,20). First, therapeutic success was found with
medications that inhibit T-cell functions. The first of these
Figure 1. Typical clinical pictures of patients
with psoriasis vulgaris.
Sabat et al.
ª 2007 The Authors
780 Journal compilation ª 2007 Blackwell Munksgaard, Experimental Dermatology, 16, 779–798

pharmaceutical products was cyclosporin A, a substance
that diminishes T-cell proliferation and cytokine produc-
tion (21). An improvement of psoriasis was also observed
after treatment with other T-cell modulating drugs: anti-
CD4 antibodies (22,23) and a fusion protein composed of
interleukin (IL)-2 and fragments of diphtheria toxin (24).
Second, psoriatic lesions healed when patients received a
haematopoietic stem cell transplant, necessary because of
an unrelated disease, from a non-psoriatic donor. On the
other hand, if an otherwise healthy patient receives bone
marrow from a donor with psoriasis, the patient also fre-
quently develops psoriasis (25,26). Thirdly, in a severe
combined immunodeficiency mouse that received a trans-
plant of uninvolved skin taken from a patient with psoria-
sis, psoriasis-like alterations of the transplanted skin could
be observed if autologous blood immune cells were acti-
vated in vitro and injected into this skin (27–29). Interest-
ingly, when skin grafts from healthy individuals were
transplanted and autologous blood immune cells were
injected, no conversion to psoriasis-like plaques was found
(29).
Many new insights have been gained over the last few
years that, in our opinion, should change the view of the
pathogenesis of psoriasis as a T1-mediated skin disease.
These new results include: (i) the observations of a pro-
found success of anti-tumor necrosis factor (TNF)-a ther-
apy in psoriasis patients, (ii) the knowledge regarding the
role of several types of immune cells such as DCs, Th17
cells, cd T cells, natural killer (NK) T cells and regulatory
T cells, (iii) the knowledge about the effects of signal trans-
duction activation in keratinocytes (e.g. STAT3) and (iv)
the findings regarding new mediators such as IL-22, IL-23
and IL-20. All of these new insights prompted us to pro-
pose a novel model of psoriasis pathogenesis. Our model is
based on numerous research results, although at some
points the facts are tied together by conjecture. These con-
jectures are clear identified as such in the text. According
to our model, the onset of the disease is similar to an
immune reaction, which is composed of three phases: a
sensitization phase, a silent phase and an effector phase
(Fig. 2). During the sensitization phase, DCs process and
present antigens and subsequently induce the development
of skin infiltrating effector ⁄ memory Th17 and T1 cells. It is
important to note that the sensitization phase is not
accompanied by any skin alterations. The sensitization
phase is then followed by a silent phase of variable length.
Each cycle of the reoccurring effector phase can be differ-
entiated into three subsequent stages: (i) skin infiltration of
immune cells, (ii) immune cell activation in the skin and
(iii) keratinocyte response. The ‘keratinocyte response’
leads to an overshot reaction of these tissue cells that is
reminiscent of an overshot regeneration process (like
wound-healing). After a successful treatment, the effector
phase transposes into the silent phase (Fig. 1). After a
while, a renewed effector phase follows. In our model, dif-
ferent cell types contribute significantly to the onset of pso-
riasis at different time points. This should be taken into
consideration when developing new therapies. Currently,
most patients are treated during the ‘keratinocyte response’
stage of our model, because this is the only stage of psoria-
sis pathogenesis in which skin alterations are visible. We
hypothesize that in this stage T cells, macrophages and
DCs are initially mainly responsible for the alteration of
keratinocytes. However, over time, macrophages, DCs and
later tissue cells also play a dominant role. In the final
stages of the ‘keratinocyte response’, T cells take on only a
minor role.
In subsequent sections of this review we describe in
detail every stage of our model.
Sensitization phase – antigen processingand presentation
The initial step of every specific immune reaction to an
antigen (Ag) is performed by professional antigen present-
ing cells (APCs) such as DCs or macrophages. At the onset
of psoriasis, the initial step may comprise the recognition
and uptake of (exogenous) Ag by tissue-guarding, imma-
ture DCs, the migration of these cells to the T-cell areas of
secondary lymphatic organs, the processing of the Ag and
the presentation of selected Ag fragments (peptides) on the
DCs’ cell surface MHC class II (MHC II) molecules to
T cells. In this process, the composition of the antigen-
processing proteases in DCs as well as the DCs’ MHC type
determines the nature of the presented peptides and
thereby, in the end, the specificity of the effector and mem-
ory T cells that is generated in the sensitization phase.
After the Ag uptake, DCs undergo a maturation process
if they encounter inflammatory conditions (because of
cytokines like TNF-a or IL-1b) or microbial products that
stimulate DCs via Toll-like receptors (30,31). They upregu-
late the functional CCR7 chemokine receptor on their sur-
face and migrate into the T-cell area of the regional lymph
nodes via afferent lymphatic vessels following a gradient ofFigure 2. Schematic diagram of the three phases of our model of
psoriasis pathogenesis.
Immunopathogenesis of psoriasis
ª 2007 The Authors
Journal compilation ª 2007 Blackwell Munksgaard, Experimental Dermatology, 16, 779–798 781

CCL21 and CCL19 (ligands for CCR7) (32–34) thereby car-
rying the previously uptaken Ag with them and drastically
reducing their capacity to further absorb Ags (35,36).
Moreover, several changes occur regarding the MHC II
pathway. In immature DCs, most MHC II resides intracel-
lularly. Upon DC maturation, a redistribution of MHC II
occurs in the cells leading to high expression on the cell
surface. Some studies provided evidence that the intracellu-
lar accumulation of MHC II in immature DCs is caused by
the inefficient formation of mature complexes of MHC II
and antigenic peptides that are caused by a minimal cellu-
lar lysosomal activity. In fact, it has been shown that in
immature DCs, the (at least partially intact) Ag as well as
the MHC II, the latter being associated with the chaperonic
invariant chain fragment p10, resided within the cell in
specialized endosomal ⁄ lysosomal compartments, co-local-
ized with HLA-DM, inactive hydrolytic proteases, and pro-
tease inhibitors such as cystatin C (37–41). The invariant
chain is known to stabilize the conformation of freshly syn-
thesized MHC II, to protect it from premature peptide
loading by blocking its binding groove with its class-II-
associated invariant-chain peptide (CLIP) portion, and to
retain the MHC II in the endosomal compartments with
the help of two sorting signals in its cytoplasmic portion
(42). Increased protease activity upon DC maturation may
be achieved by the decreased production of protease inhibi-
tors and the activation of the vacuolar ATPase (38,40), and
leads to splitting of both the invariant chain and the Ag.
The splitting of the invariant chain occurs in a defined
sequence of steps (43). The limiting step is the last cleavage
whereby the endosomal retention signal sequence is split-
off from the p10 fragment to leave the MHC II peptide-
binding, groove-blocking CLIP fragment. In DCs, this last
cleavage is thought to be carried out by the cysteine endo-
peptidase cathepsin S (44,45). Like the processing of the
invariant chain, the degradation of the uptaken Ag upon
DC maturation is a process in which numerous proteases,
including cathepsins and legumain, play a role (42). The
end result is approximately 12–25 amino acid long pep-
tides. These peptides are exchanged for CLIP, provided a
certain affinity for the MHC II concerned exists. This most
probably takes place with the help of the non-classical
MHC II molecule HLA-DM (43). The assembled peptide-
loaded MHC II can then be transported to the surface of
the DC. The additional mechanism for the intracellular
accumulation of MHC II in immature DCs may be the
increased turnover of cell surface MHC II peptide com-
plexes because of their increased endocytosis (46). In fact,
the half-life of the MHC II at the cell surface is about 10 h
in immature DCs, and increases to more than 100 days
after DC maturation (47).
Interestingly, the synthesis of further MHC a and b chains,
after a transient upregulation, is inhibited in mature DCs
(47,48). This has been shown to be caused by the reduced
expression of the transcriptional regulator CIITA that is
caused by the epigenetic modulation of the promoters of the
encoding gene (49). Like the downregulated Ag uptake, the
reduced MHC II synthesis may also prevent the presentation
of newly encountered Ags. This prevention, in turn, in con-
nection with the low turnover of cell surface MHC II, may
cause DCs to retain a long-lasting, selective memory of the
special, previously uptaken Ag as proven by its capacity to
stimulate T cells even several days later (47,48). MHC II pep-
tide complexes on the DC surface can be recognized by
CD4+ T cells via the T cell receptor (TCR) given the fact that
this TCR is specific for the MHC–peptide complex. The
increased cell surface expression of peptide-loaded MHC II is
also accompanied in DCs by strengthened expression of
adhesion and co-stimulatory molecules (see below).
Although the presentation of peptides derived from Ags
from the extracellular environment typically occurs via the
MHC II pathway as described above, DCs are endowed
with the capacity to cross-present those Ags on MHC I
molecules leading to the additional, initial activation of
CD8+ T cells (50).
At this point, there remains at least one question that is
crucial for the understanding of the pathogenesis of psoria-
sis, namely which Ag is responsible for initial T-cell prim-
ing in the majority of patients.
The answer should be revealed when the reactivity of T
cells in psoriatic lesions can be identified. To this purpose,
the Fry group isolated T cells from psoriatic lesions of
acute guttate and chronic plaque psoriasis and established
T-cell lines from these cells. They could show that these
T-cell lines were reactive against isolates from streptococci
(51,52). A streptococcal origin of the primary Ag(s) would
be in line with the fact that first manifestation and relapses
and aggravations of psoriasis are often linked to infections
with streptococci, especially b-haemolytic Streptococcus
pyogenes (9,11). At this point, it should be emphasized that
neither the portion of psoriasis patients with skin infil-
trated T cells against streptococcal Ags nor the proportion
of such T cells within the whole T-cell infiltrate is currently
known. In addition to streptococcal Ags, other Ags, even a
few endogenous Ags, were postulated to trigger psoriasis.
Interestingly, it has been shown in experimental models
that DCs can break tolerance to endogenous antigens via
bystander activation under certain circumstances. In this
case, it would make no sense to look for exogenous ‘cross-
reactive’ antigens as a trigger of psoriasis.
Sensitization phase – generation ofeffector and memory T cells
The conversion from naıve to effector and memory T
cells that occurs in secondary lymphatic organs under the
Sabat et al.
ª 2007 The Authors
782 Journal compilation ª 2007 Blackwell Munksgaard, Experimental Dermatology, 16, 779–798

guidance of DCs is necessary for T cells to obtain their
functions and their ability to immigrate into tissues. The
naıve T cells permanently circulate between blood and
secondary lymphatic organs, such as lymph nodes and
tonsils. In T-cell areas of secondary lymphatic organs, they
congregate with mature DCs. A naıve CD4+ T cell whose
TCR has a high enough affinity for the respective MHC
II–peptide complex, sticks to the respective DC and forms
a so-called immunological synapsis. In order to become
activated, the T cell now needs three signals: the first sig-
nal has already been delivered by interaction between
TCR and MHC II–peptide complex. The second signal is
given by the so called co-stimulatory molecules. The third
signal for T-cell activation is delivered by soluble media-
tors (Fig. 3).
The most important co-stimulatory molecules expressed
by DCs are those of the B7 family: CD86 (B7-2), CD80
(B7-1), B7h, PD-L1 and PD-L2 as well as CD40 (53,54).
CD86 and CD80 interact with the T-cell molecules CD28
and CD152 (cytotoxic T-lymphocyte-associated protein 4,
CTLA-4). CD28 shows a large constitutive expression in T
cells, particularly in naıve T cells. Engagement of CD28
reduces the required number of triggered TCRs for T-cell
activation and allows activation of T cells by low affinity
ligands (55). Additionally, it is important for stable IL-2
production and IL-2Ra (CD25) expression, and it prevents
the induction of unresponsiveness and apoptosis of T cells
after TCR stimulation (55,56). Hence, CD86 functions as a
major co-stimulatory molecule in DCs and is critical for
full activation, particularly of naıve T cells. CD152 does
not appear to be expressed on resting T cells, but is trans-
ported from intracellular clathrin vesicles to the cell surface
following TCR stimulation (57). Engagement of CD152
inhibits TCR- and CD28-mediated signal transduction,
increases the threshold for activation of these cells and
represses the cell cycle (58). Therefore, it counteracts acute
T-cell responses but is also essential for the formation of
memory T cells (otherwise all cells would differentiate into
effector cells with short life times). The counterpart for
B7h (DC) on T cells is ICOS, for PD-L1 and PD-L2 on T
cells is PD-1, and for CD40 on T cells is CD154 (CD40
ligand). The immunological synapsis is stabilized by adhe-
sion molecules expressed by DCs and T cells (59). Among
the most important are (i) the interactions between inter-
cellular adhesion molecule 1 (ICAM-1; CD54) (DC) and
lymphocyte function-associated antigen-1 (LFA-1;
CD11a ⁄ CD18; aL:b2) (T cells) and (ii) the interactions
between lymphocyte function-associated antigen 3 (LFA-3;
CD58) (DC) and CD2 (T cells).
The third signal mentioned above essentially influences
the mode of action of the effector and memory T cells,
which evolve from these activated T cells (Fig. 3). Accord-
ing to what we actually know, naıve CD4+ T cells can be
polarized into four different directions: Th1, Th2, Th17
and regulatory T cells. If IL-12 (p35 ⁄ p40) is present dur-
ing activation of naıve T cells, they are polarized into Th1
cells through activation of the transcription factors STAT4
and T-bet. The generation of Th1 cells is supported by
gamma interferon (IFN-c), which increases the expression
of the specific receptor chain for IL-12 (IL-12Rb2) (60).
During repeated activation, Th1 cells primarily produce
IFN-c, IL-22, IL-26, GM-CSF and TNF-b. If IL-4 is pres-
ent, naıve T cells are polarized into Th2 cells through
activation of the transcription factors STAT6 and GATA-3
(60). These T cells secret IL-4, IL-5 and IL-13 during
repeated activation. The activation of naıve T cells with
mature DCs in the presence of IL-6 and transforming
growth factor (TGF)-b upregulated the receptor for IL-23
(p19 ⁄ p40) and together with this cytokine induced Th17
cell development (61,62). Th17 cells primarily produce IL-
6, IL-17 and IL-22. Th17 development is independent of
STAT1, STAT4 and STAT6 signalling. The Th1-cell induc-
ing cytokines IL-12 and IFN-c actively suppress the devel-
opment of Th2 and Th17 cells. Conversely, the Th2-cell
inducing cytokine IL-4 inhibits the development of Th1
and Th17 cells (61,62). Regulatory T cells are thought to
APC Th1
Th2
Th17
Treg
NaïveCD4+T-cell
CD54
CD86
CD40 CD154
CD28
CD4
CD3
CD2
TCR
LFA-3
LFA-1T-bet
IFN-γ, TNF-β, IL-22
IFN-γ, IL-12
IL-4, IL-5, IL-13IL-4
IL-6, IL-17, IL-22
IL-6, TGF-β
IL-10, TGF-β
IL-23
IL-10, TGF-β
GATA-3
RORγt
Foxp3
MHC II
Figure 3. The activation of naıve CD4+ T
cells by antigen-presenting cells (APCs) leads
to the development of different CD4+ T-cell
lineages with distinct properties. The first and
second signal delivered by the APC to the T
cell is responsible for the generation of
effector ⁄ memory T cells. The T-cell lineage is
dependent on the cytokine milieu present
during the T-cell activation.
Immunopathogenesis of psoriasis
ª 2007 The Authors
Journal compilation ª 2007 Blackwell Munksgaard, Experimental Dermatology, 16, 779–798 783

be responsible for limiting T-cell immune responses,
partly by production of immunosuppressive cytokines,
such as IL-10 and TGF-b. However, there is still a rather
broad discussion on the generation and maintenance of
regulatory T cells (60).
Preliminary results describe distinct patterns of cell-sur-
face molecule expression allowing phenotypical distinction
between Th1, Th2, Th17 and regulatory T cells. CCR5 and
CXCR3 expression are thought to characterize Th1-cells
whereas CCR3 and CCR4 expression are assigned to Th2-
cells (63–65). However, Th17 T cells may, also be CCR4+.
Regulatory T cells seem to be contained in the bright posi-
tive CD25 fraction of CD4+ T cells (66).
Independent of its polarization into a Th1, Th2, or Th17
cell, a successfully activated naıve T cell develops into
either an effector T cell (TE), an effector memory T cell
(TEM) or a central memory T cell (TCM) (67). Effector T
cells immediately home into inflamed tissue, display their
effector functions and die. TEM cells recirculate between
blood and peripheral tissues and rapidly produce effector
cytokines upon restimulation, but they have a poor prolif-
erative capacity. In contrast, TCM cells mainly recirculate
between blood and lymph nodes and have limited effector
functions, but proliferate and become effectors following
secondary stimulation. For peripheral blood CD4+ and
CD8+ T cells, the expression of the lymph node homing
receptors CCR7 and CD62 L in combination with CD45RA
expression may be able to characterize these subsets pheno-
typically: in CD4+ T cells CD45RA+ ⁄ CCR7+ ⁄ CD62
L+ cells represent the naıve compartment,
CD45RA) ⁄ CCR7) ⁄ CD62 L cells represent the TEM com-
partment, and CD45RA) ⁄ CCR7+ ⁄ CD62 L+ cells represent
the TCM compartment. Among CD8+ T cells, effector T
cells seem to add to the CD45RA+ ⁄ CCR7) ⁄ CD62 L) frac-
tion (68,69).
A subset of successfully activated T cells expresses the
so-called cutaneous lymphocyte-associated antigen (CLA).
CLA, a defined carbohydrate epitope (so-called sialyl-
Lewis(x)), guides leucocytes to inflamed skin. With help
from fucosyltransferase VII, CLA can be placed on top of
P-selectin glycoprotein ligand-1 (PSGL-1) (70). PSGL-1 is
a cell surface molecule expressed constitutively on all
peripheral T cells (70). Mice transgenically deficient in
fucosyltransferase VII show a reduced percentage of skin
homing T cells and other leucocytes (71). Very recent
results show that CD43, a sialomucin also constitutively
expressed on T cells, can also be crowned with the CLA
epitope (72). Interestingly, CLA-modified PSGL-1 binds
both E- and P-selectin, whereas CLA modified CD43 only
binds E-selectin. However, the mechanisms by which DCs
induce the expression of CLA on a defined subset of acti-
vated T cells are still widely unknown. DCs may induce the
expression of tissue-specific homing receptors on T cells
congruent to the organ from which the DC comes. An
exception may be made for tonsillar DCs that present
streptococcal Ags corresponding to the localization of
streptococcal infection in the upper respiratory tract and
the tonsils. These DCs, however, seem to generate some
skin-homing T cells as, like in the skin-draining lymph
nodes, 5–10% of tonsillar T cells express CLA. Most inter-
estingly, TCR spectra-types and sequencing of TCR rear-
rangements provided evidence of shared T-cell clonality of
tonsillar and skin lesional CLA+ T cells, but not CLA-
tonsillar and peripheral T cells in psoriatic patients (73). In
contrast to the generation of skin-homing T cells in tonsils,
the generation of such cells could occur directly in skin-
draining lymph nodes because of a cutaneous presence of
streptococcal Ags that may reach the skin to some extent
during streptococcal infection (74).
Effector phase – skin infiltration of T andother immune cells
If psoriatic plaques develop, the first microscopically visi-
ble events are the superficial perivascular infiltration of
lymphocytes and monocytic cells and the dilation of the
blood vessels in the dermal papillae (15). The passage of
leucocytes from the blood vessels into tissue occurs in five
steps (Fig. 4). In these processes, endothelial cells play a
decisive role.
In the first step, leucocytes roll along the blood vessel
wall. Rolling reduces the flow velocity of the leucocytes and
is mediated by the interaction between P- and E-selectin
expressed by endothelial cells and selectin ligands expressed
CCR4
E-Selectin
CLA
CCL17
LFA-1
VLA-4
CD54 CD106
Fibronectin
CXCL9 CXCR3
Rolling Triggering Adhesion Diapedesis Migration
Figure 4. Five steps of skin infiltration of T
cells. Only the adhesion molecules and
chemokines, which are most important for
the immigration of T cells in the skin, are
presented.
Sabat et al.
ª 2007 The Authors
784 Journal compilation ª 2007 Blackwell Munksgaard, Experimental Dermatology, 16, 779–798

by leucocytes (PSGL-1, CLA-modified PSGL-1 and CLA-
modified CD43). In healthy controls, endothelial cells of
cutaneous postcapillary venules express low amounts of P-
selectin molecules (75). In the uninvolved skin of patients
with psoriasis, endothelial cells express both P- and E-selec-
tin, and its expression is much more enhanced in psoriatic
skin (75). One of the actions of TNF-a is the early release
of so-called Weibel-Palade bodies containing P-selectin.
Additionally, TNF-a induces the synthesis and surface
expression of E-selectin molecules.
Very important for the tissue homing is the production
of chemokines, which exist not only as soluble mediators
but also bound to glycosaminoglycans on endothelial cells
at the site of inflammation (76,77). In the second step of
skin infiltration, the immune cells rolling along the blood
vessel wall recognize chemokines presented by endothelial
cells and are activated. It was postulated that the chemoki-
ne CCL27 (ligand for CCR10) is highly relevant for migra-
tion of T cells into psoriatic skin (78) although other
chemokines seem to play a predominant role (76,77,79–
81). Reiss et al. show that the inhibition of CCL27 has no
effect on lymphocyte recruitment into skin in the psoriatic
mouse model, but only the additional blockage of the
interaction between CCL17 and CCR4 abrogates skin
recruitment in this model (80). Moreover, most skin-infil-
trating lymphocytes in allergic delayed-type hypersensitivity
express CCR4, but only about 10% express CCR10 (77). In
psoriasis lesions, the majority of CD4+ T cells and about
half of the CD8+ T cells expressed CCR4 and CCR6 (81).
Interestingly, CCL17, the ligand for CCR4, was expressed
by TNF-a or IFN-c-stimulated dermal endothelial cells but
not by activated keratinocytes (81,82). CXCR3 is expressed
by about one-third of CD4+ and CD8+ T cells infiltrating
psoriatic skin lesions. CXCR3 may be particularly impor-
tant for the migration of CD8+ T cells into the epidermis,
as its ligands CXCL9 and CXCL10 are expressed by TNF-aor IFN-c-stimulated keratinocytes (82).
The next step of skin infiltration is characterized by the
formation of tight adhesions between endothelial cells and
immune cells (Fig. 4). This is achieved by integrins
expressed on immune cells and their ligands expressed on
endothelial cells. Chemokines induce the integrin-depen-
dent adhesion of immune cells to endothelial cells and
cause the rapid arrest of the former under physiological
flow (76). CD11a ⁄ CD18 (LFA-1) seems to be the most
important integrin for skin homing. It binds to CD54 and
CD102 (ICAM-2) expressed by endothelial cells. Another
integrin system, consisting of very late antigen (VLA)-4
(a4:b1) expressed by T cells and vascular cell adhesion mol-
ecule 1 (CD106) expressed by endothelial cells, does seem
to be less important in this process. In fact, the vast major-
ity of intra-epidermal T cells in psoriatic skin lesions
expressed CD11a ⁄ CD18, whereas only 58% of CD4+ and
85% of CD8+ T cells expressed VLA-4 (81). The endothe-
lium-presented chemokines triggered instantaneous exten-
sion of bent CD11a ⁄ CD18, which led to an increased
affinity of CD11a ⁄ CD18 – CD54 binding (83). Although
VLA-4 affinity is not altered upon chemokine signalling,
subsequent VLA-4 clustering at the leucocyte–substrate
contact zone results in enhanced leucocyte avidity to
CD106 (84). Resting endothelial cells only express CD102.
TNF-a and IFN-c, but not IL-6 or GM-CSF, induced the
expression of CD54 on endothelial cells (85,86).
The passage of immune cells through the endothelial
wall is called diapedesis and is probably performed by
pores formed between endothelial cells (Fig. 4). It seems
that this step is also dependent on integrins and, within
the integrin system, more on CD11a ⁄ CD18 than on VLA-4
(87). In fact, the intensity of endothelial cell expression of
CD54, the CD11a ⁄ CD18 ligand, correlated with the degree
of dermal inflammation (88). A small amount of peripheral
T cells also migrates into non-inflamed skin (see TEM cells).
This is made possible by constitutive expression of P-selec-
tin and CD102 on resting dermal endothelial cells. Interest-
ingly, Lowes et al. showed that in non-lesional psoriatic
skin, the number of T cells is higher than in skin from
healthy participants (89). In psoriatic skin lesions, CD8+ T
cells primarily home into the epidermis, while CD4+ cells
are mainly present in the dermis (90,91). The reasons for
these different anatomical homing patterns may be caused
by the varied expression of chemokine receptors (such as
CXCR3, see above) and integrins (such as CD103) on
CD4+ and CD8+ T cells. In fact, 80% of epidermal CD8+
T cells expressed the integrin CD103 (aE:b7). In contrast,
only 5% of the dermal T cells and less than 1% of periph-
eral blood lymphocytes derived from psoriatic patients or
healthy participants were CD103-positive (92,93). CD103
binds epidermal E-cadherin (94). Interestingly, in vitro cul-
ture experiments showed that CD103 was preferentially
expressed on CD8+ T cells after stimulation with anti-CD3
monoclonal antibodies. Co-culture with TGF-b and IL-4
upregulated the CD103 expression on T cells, whereas
IL-12 downregulated it (92,93).
Other immune cells such as NK cells, monocytes ⁄ macro-
phages, DCs and neutrophilic granulocytes also use a
mechanism similar to the one described above for T cells
that migrate into the skin. However, they are often guided
to the skin by other chemotactic factors and can also use
other adhesion molecules. In fact, skin-derived NK cells
have been shown to express high levels of the chemokine
receptors CXCR3 and CCR5, and intermediate amounts of
CXCR1, CCR6 and CCR8. In line with that, prompt migra-
tion of these cells could be induced in vitro with CXCL10
and CCL5, and to a lower extent with CCL20 and CCL4
(95). The complement component C5a is apparently the
most important of these chemotactic factors that capture
Immunopathogenesis of psoriasis
ª 2007 The Authors
Journal compilation ª 2007 Blackwell Munksgaard, Experimental Dermatology, 16, 779–798 785

the myeloid DCs in psoriatic lesions (96). Langerhans cells
immigrate into psoriatic lesions most likely because of
CCL20 (ligand for CCR6). Keratinocytes produce CCL20
after stimulation and CCL20 is upregulated in lesional pso-
riatic skin (97). For homing of neutrophilic granulocytes,
interaction between the integrin MAC-1 (CD11b ⁄ CD18;
aM:b2), expressed by neutrophils, and CD90, expressed by
endothelial cells, does play an important role (98). In the
end, neutrophils migrate into the stratum corneum and
form microabscesses characteristic for psoriatic skin lesions.
CXCL8 and complement factors (C5a) may also play an
important role in this immigration.
Mostly as a consequence of the infiltration, significantly
more immune cells are found in psoriatic lesions than in
healthy skin. Interestingly, the numbers of T cells, macro-
phages and DCs are similar (89,99).
Not surprisingly, CD4+ and CD8+ T cells of psoriatic
skin lesions have activation and memory phenotypes
(90,91,100). T cells from the lesional epidermis expressed
HLA-DR (86%), CD69 (59%), CD25 (55%) (all activation
markers), and were CD45RA negative (91%) (memory phe-
notype). T cells from lesional dermis showed similar char-
acteristics except for CD69 (17%) (101). CD69 is a surface
molecule that is expressed by T cells for only a short time
after activation. Regarding cytokine phenotypes, Schlaak
et al. already described in 1994 that T-cell clones generated
from lymphocytes taken from psoriatic skin partially exhib-
ited a T1 cytokine secretion profile (102). The authors
demonstrated that such clones produced large amounts of
IL-2 and IFN-c but only a little or no IL-4, IL-10 and
TNF-a (102). A later study demonstrated that about 40%
of skin CD4+ and CD8+ T cells were able to produce IFN-
c, IL-2 and TNF-a after ex vivo stimulation (103). Recently,
our group generated quantitative data on mRNA expres-
sion of 20 cytokines demonstrating that of all T-cell cyto-
kines investigated, IL-22 and IL-17 had the highest
expression in lesional psoriatic skin (104,105) (R. Sabat,
unpublished data). The expression of IL-22 was approxi-
mately 10 times higher than that of IFN-c (105). Interest-
ingly, we also found IL-22 in the blood of patients with
psoriasis (in contrast to IFN-c), and the level of it in the
blood correlated with the severity of disease (106). IL-22 is
a member of the IL-10 IFN family (107). It can generally
be produced by activated T and NK cells, but not by other
immune cells or tissue cells (105,108). The main source of
IL-22 in psoriatic lesions should be the effector ⁄ memory
Th17 and Th1 cells (108–110). As soon as the late 1990s, it
was determined that the majority of the CD4+ and CD8+
T-cell clones derived from lesional psoriatic skin expressed
IL-17 mRNA, suggesting that skin-infiltrating T cells pro-
duce this cytokine (111). In fact, Chan et al. very recently
demonstrated elevated IL-17 expression in psoriatic skin
lesions (112). Our measurements showed that this elevated
expression was in the same magnitude as IL-22 (104,105)
(R. Sabat, unpublished data). All these data suggest that
the majority of CD4+ T cells in lesional psoriatic skin may
be IL-22 and IL-17-producing T cells. Interestingly, Cargill
et al. very recently found that single-nucleotide polymor-
phisms in the IL-23 receptor gene are associated with pso-
riasis, supporting the role of Th17 T cells in psoriasis
pathogenesis (113).
Since more than 15 years, it has been known that the
number of macrophages is increased in psoriatic skin
lesions (114–116). We recently found that the numbers of
macrophages (CD68+) and T cells in diseased skin are very
similar (R. Sabat, unpublished data). The number of DCs
in psoriatic skin lesions is apparently only about half the
amount of T cells (117) (R. Sabat, unpublished data). In
normal skin, two populations of DCs in particular can be
found; namely the Langerhans cells that are present in the
epidermis and the dermal DCs that are located in the der-
mis (118). Psoriatic lesional skin contains unaltered num-
bers of Langerhans cells (119) and at least two other DC
populations. In contrast to normal skin, there are also
inflammatory dendritic epidermal cells (CD11c+ ⁄ CD1a+ ⁄Lin) ⁄ CD123)) and small amounts of plasmacytoid DCs
(CD11c) ⁄ CD1a) ⁄ Lin) ⁄ CD123+) (117,119,120). It should
also be mentioned that a high number of CD11c+ APCs
can be found in diseased skin. This population consists of
DCs (89) and probably mainly macrophages (121).
In psoriatic lesions, the amount of NK T cells is also
increased in comparison with normal skin (122). NK T
cells are a separate line of T cells, which are known for (i)
their expression of NK-cell receptors, (ii) their extremely
restricted TCR repertoire (mostly Va24 and Vb11) and
(iii) their activation by glycolipids presented via CD1d
(123). NK T cells can be CD4+ or CD4- ⁄ CD8. Approxi-
mately 5% of the cells infiltrated in the psoriatic lesions
are NK cells (95).
Effector phase – immune cell activation inthe skin
The immune cells may be activated after migration into the
skin. In the dermis and epidermis, different APC popula-
tions such as macrophages and various types of DCs can
stimulate T cells, and vice versa. T cells that have migrated
into the epidermis may additionally be activated by kerati-
nocytes. T cells may proliferate as a consequence of activa-
tion in the skin (124,125). Psoriatic lesions even
demonstrate some characteristics of lymph nodes. The acti-
vation of T cells in the skin can be negatively influenced by
resistant intraepithelial T cells and regulatory T cells.
Already in 1994, Nestle et al. demonstrated that DCs
from psoriatic plaques were much more effective stimula-
tors of spontaneous T-cell proliferation than blood-derived
Sabat et al.
ª 2007 The Authors
786 Journal compilation ª 2007 Blackwell Munksgaard, Experimental Dermatology, 16, 779–798

DCs or DCs from skin of healthy individuals. Antibody-
blocking studies demonstrated involvement of HLA-DR,
CD28 ligands and LFA-1 in this stimulation (126). New
studies hinted at an important role of plasmacytoid DCs
during the development of psoriasis. In a xenograft model
of human psoriasis, blocking IFN-a signalling or inhibiting
the ability of plasmacytoid DCs to produce IFN-a pre-
vented the T-cell proliferation and development of psoriasis
(127). Interestingly, Eriksen et al. very recently showed that
psoriatic T cells have increased and prolonged responses to
IFN-a with respect to the level of STAT activation, when
compared with infiltrating T cells from the skin of non-
psoriatic donors. This increased IFN-a-induced signalling
led to an enhanced binding of STAT4 to the IFN-c pro-
moter and an enhanced IFN-c production, as well as to
inhibition of T-cell growth (128).
An important question for the understanding of the pso-
riasis pathogenesis is how monocytes ⁄ macrophages and
DCs activate the immigrated T cells in the skin. We assume
that cytokines (in particular IL-23 and IL-6) produced by
these APCs play a dominate role. In fact, myeloid DCs
(CD83+) and in particular monocytes ⁄ macrophages
(CD68+) separated from psoriatic skin lesions expressed IL-
23, and IL-23 (a Th17-cell-inducing cytokine) expression
was increased in psoriasis. In contrast, IL-12 (a Th1-cell-
inducing cytokine) expression was not upregulated (121).
Another question is whether a certain antigen is required
for the activation of T cells in the skin, and if so which one.
Some studies propose a mechanism for the reactivation
of psoriasis-relevant T cells that were primarily directed
against streptococcal components. Although some spread-
ing of streptococcal components into the skin has been
reported (74), these Ags do not seem to persist in psoriatic
lesions and do not seem to play a role in the activation of
the immigrated T cells. This is also supported by the fact
that antibiotic treatment or tonsillectomy seldom improves
the course of the psoriasis (129,130). An attractive basis for
the resolution of this discrepancy may be the concept of
cross-reactivity of the T cells, primarily directed against
streptococcal Ags, with auto-Ags. This may be possible in
principle as a certain number of potentially autoreactive T
cells seem to persist in the organism despite the negative
T-cell selection process in the thymus. The model of cross-
reactivity requires that one or several microbial structures
are shared by structures of the human tissue which is, in
the case of psoriasis, the skin [molecular mimicry (131)].
Indeed, initial database searches identified high structural
similarities between streptococcal M proteins, which are
major streptococcal virulence factors, and type I keratins
(132). Both types of proteins share the a-helical coiled-
structure formed by hepta-peptide repeat patterns. More
recently, several studies demonstrated responses of periph-
eral blood T cells from psoriatic patients but not healthy
participants to several synthetic peptides corresponding to
shared sequence motifs present in M proteins and type I
keratins. One example is sequences comprising the ALE-
EAN motif common in both streptococcal M6 and K17
(133,134). Among them, Gudmundsdottir et al., using the
146-K17 peptide, demonstrated that out of 17 patients and
17 control persons tested, 13 patients responded to this
peptide compared with four controls, and the responses
were significantly stronger in the patient group (133).
Although all type I keratins share significant sequence
similarity, K17 seem to be a preferred candidate for the
psoriasis relevant auto-Ag. The auto-Ag is expected to be
present in skin (and may be in the synovial compart-
ments), but not in uninvolved tissues. Indeed, K17 has
been shown to be present in skin, but not in buccal
mucosa. Under normal conditions, however, cutaneous
K17 expression is missing except for the hear follicles, nail
beds, sweat glands and sebaceous glands. Its expression is
induced in suprabasal keratinocytes in psoriatic lesions
(135,136). In vitro, K17 expression could be induced in the
keratinocytes HaCat cell line by IFN-c, and to a lesser
extent, by TGF-a, whereas other pro-inflammatory cyto-
kines such as IL-1, IL-6, IL-8 and IL-18 had no inducing
effect (137,138). Regarding the expressional regulation of
K17, one would argue that the K17 expression is not the
cause but rather a secondary effect induced by activated T
cells. However, there may be one argument supporting a
primary involvement of K17: psoriasis often starts with the
affection of the scalp, where K17 is physiologically present.
In the epidermis, migrated T cells can also be activated
by keratinocytes. The keratinocytes have been shown to
express MHC class II molecules and CD54 after exposure
to IFN-c (139). Such activated keratinocytes were able to
induce T-cell proliferation by bacterial-derived super-Ags
(such as staphylococcal enterotoxin A and B), which could
be significantly and partially inhibited by mAbs against
LFA-1 and by mAb against MHC class II, respectively, but
not by mAbs against the CD28 ligands (140). Interestingly,
these IFN-c-treated keratinocytes are not able to support
T-cell proliferation to alloantigens (140). This can be taken
to mean that such cells impair antigen presentation via
MHC II. However, results from Griffiths et al. showed that
the presence of intra-epidermal lymphocytes was not corre-
lated with keratinocyte HLA-DR expression (88). Addition-
ally, the restricted clonality of the CLA+ T cells that are
present within the lesional skin in psoriatic patients (73),
as well as the disease’s familiarity coupled to certain pre-
vailing MHC types argues against the involvement of
super-Ags and for the involvement of classical antigenic
proteins of the streptococci. It should be noted, however,
that bacterial super-Ags possibly contribute to skin inflam-
mation through direct activation of keratinocytes, probably
by binding to MHC II (141).
Immunopathogenesis of psoriasis
ª 2007 The Authors
Journal compilation ª 2007 Blackwell Munksgaard, Experimental Dermatology, 16, 779–798 787

Keratinocytes may also play a role in the activation of
NK T cells through CD1d. CD1d is expressed by keratino-
cytes in normal skin at a relatively low-level and confined
to upper-level keratinocytes immediately beneath the lipid-
rich cornified layer. In psoriatic plaques, there is an over-
expression of CD1d. CD1d could also be rapidly induced
on keratinocytes in normal skin by physical trauma that
disrupted barrier function. Keratinocytes also displayed
enhanced CD1d following exposure to IFN-c in vitro. Com-
bining CD1d+ keratinocytes with human NK T-cell clones
resulted in the clustering of NK-T cells, and, while no sig-
nificant proliferation ensued, NK T cells became activated
to produce large amounts of IFN-c (142).
The activation of T cells in the skin should be inhibited
by two subpopulations of T cells. In normal mouse skin,
and also in low amounts in human skin, intraepithelial T
cells can be found. The biology of these cells is poorly
understood. They express the cd TCR and often show less
TCR variability, in comparison with T cells that express the
ab TCR. This distinction is especially evident in mice and
several other mammals as well as in humans albeit to a les-
ser degree. Recent studies in laboratory animals have indi-
cated the capacity of cd T cells to play major roles in the
maintenance of the epidermal barrier, the regulation of
cutaneous inflammation, and the protection against cutane-
ous neoplasms (143). Interestingly, the intraepithelial T
cells are able to inhibit ab T cells and prevent dermatitis in
the mouse model (144). In his presentation at the 4th
International Congress ‘Psoriasis from Gene to Clinic’ in
London in 2005, Hayday hypothesized that following a
chronic activation of cd T cells in psoriasis, the cells
become ‘exhausted’ which then leads to a lack of inhibition
of ab T cells in the skin. The second subpopulation of T
cells that can inhibit Th17 and Th1 cells is the regulatory T
cells. New data suggests that these cells are dysfunctional in
patients with psoriasis. In fact, regulatory CD4+ T cells
from peripheral blood from patients with psoriasis have
been shown to be deficient in their suppressor activity
(145). In this study, regulatory T cells were also isolated
from the site of inflammation, the psoriatic plaques, and
were analysed. At calculated ratios of regulatory T cells to
effector T cells found to be present in the skin, the psoriat-
ic regulatory T-cell population demonstrated decreased
suppression of effector T cells.
The skin-infiltrating monocytes ⁄ macrophages and DCs
are apparently activated by IFN-c produced by T, NK T
and NK cells, and possibly by heat shock protein produced
by keratinocytes. These cells then may begin to produce
TNF-a, IL-6, IL-18, IL-19, IL-20 and IL-23. Interestingly,
the biological activity of IL-1 is also not enhanced in psori-
atic lesions. Dermal macrophages in the papillary dermis
could be the main source of TNF-a in psoriatic skin. After
staining sections of a psoriatic lesion, Nickoloff et al. found
also TNF-a concentrated in keratinocytes and in intra-epi-
dermal Langerhans cells, although it was not found after
staining of endothelial cells, mast cells or dermal DCs
(146). In a recent study, Lowes et al. showed TNF-a stain-
ing in CD11+ APCs (89). As already noted above, this cell
population probably comprises DCs and, in particular,
macrophages. It should also be mentioned that some other
manuscripts describe the keratinocytes as a mention-worthy
cellular source of TNF-a in psoriatic lesions (75,147).
Effector phase – keratinocyte response
Keratinocytes can be activated during the initiation of pso-
riatic lesions mainly by mediators produced by T1 cells
(IFN-c and IL-22). However, we postulated that over time
the mediators of Th17 cells (IL-6, IL-17 and IL-22), fol-
lowed by those of macrophages and DCs (TNF-a, IL-6, IL-
18, IL-19 and IL-20) (Wolk et al., unpublished data) as
well as lastly mediators produced by keratinocytes such as
TGF-a, nerve growth factor (NGF), IL-19, IL-20, and by
stromal cells in the dermis such as keratinocyte growth fac-
tor (KGF), insulin-like growth factor 1, and fibroblast
growth factor 10, become increasingly important (Fig. 5).
Activation of keratinocytes leads to (i) increased prolifera-
tion of these cells and (ii) alteration of their maturation.
Moreover, activated keratinocytes produce numerous varied
mediators that can cause further immigration of immune
cells, activate stromal cells in the dermis, and induce angio-
genesis.
The keratinocyte proliferation in psoriatic lesions is
raised almost 50-fold. Until today, it has not been possible
to identify the mediator(s), which is clearly responsible for
this massive increase. Immune cells could partly be respon-
sible for the proliferation. Hancock et al. could show that
activated and non-activated T cells release factors that
could increase keratinocyte proliferation (148). The same
group also found that suppressive molecules were produced
preferentially by monocyte cultures. Bata-Csorgo et al.
demonstrated that CD4+ T cells, cloned from lesional pso-
riatic skin and stimulated by immobilized anti-CD3 plus
fibronectin, promoted psoriatic uninvolved keratinocyte
proliferation via soluble factors (149). The search for the
T-cell mediator that causes this development has been dis-
appointing to date. Of the T-cell-produced mediators that
have been investigated, mediators such as IFN-c and TGF-bappear to inhibit the proliferation of keratinocytes and oth-
ers appear to have no effect on proliferation or they
increased proliferation of keratinocytes at only high con-
centrations (IL-6, IL-8) (148,150). As has been documented
in many studies, the strong inhibitory impact of IFN-c on
keratinocyte proliferation suggests that this cytokine does
not play a role in any of the marked changes in the kerati-
nocytes in psoriasis. At this point, it should be interjected
Sabat et al.
ª 2007 The Authors
788 Journal compilation ª 2007 Blackwell Munksgaard, Experimental Dermatology, 16, 779–798

that a discrepancy between the presence of IFN-c in psori-
atic lesions and the increased proliferation of keratinocytes
does not necessarily exist. For this discrepancy, there may
be two explanations. First, the expression of IFN-c in psori-
atic lesions is only slightly increased in contrast to other
cytokines (104,105). Second, the psoriatic keratinocytes
show abnormal signalling in the IFN-c pathway. Support-
ing the second explanation, Jackson et al. demonstrated
that psoriatic keratinocytes showed a reduced induction of
IRF-1 and STAT1a activation after stimulation with IFN-c,
compared with normal keratinocytes (151). Nevertheless,
IFN-c can play a role in psoriasis, particularly in the early
stages of the effector phase: (i) by increasing the immigra-
tion of immune cells into the skin (by induction of numer-
ous chemokines in keratinocytes such as CCL2, CXCL2,
CXCL9, CXCL10 and CXCL11), (ii) by activating mono-
cytes, macrophages, DCs and endothelial cells, and (iii)
by upregulating MHC I, MHC II, CD1d and CD54 in
keratinocytes.
Coming back to the increased proliferation of keratino-
cytes, it was interesting that products from neutrophilic
granulocytes were identified that could be responsible for
the increased proliferation of keratinocytes. Human leuco-
cyte elastase induced proliferation in murine keratinocytes
in concentrations that can be found on the skin surface of
psoriatic lesions (152). Moreover, daily topical application
of human leucocyte elastase on hairless mice induced a
concentration-dependent epidermal hyperproliferation. His-
tologic analysis revealed marked vasodilation but, interest-
ingly, no inflammatory infiltrates (152). However, anti-
CXCL8 therapy in psoriatic patients significantly reduced
the amount of neutrophilic granulocytes in the diseased
skin, but had no impact on the macroscopic skin alteration
(153). This contradicts an important role of neutrophilic
granulocytes in the induction of the keratinocyte response.
The impressive therapeutic effects of the neutralization
of TNF-a in psoriasis let us assume that this cytokine
strongly, although indirectly, increases the proliferation of
keratinocytes. As mentioned previously, macrophages in
the papillary dermis could be the main source of TNF-a in
the psoriatic skin (146). If this is the case, macrophages
appear to be the cell type mainly responsible for the psori-
atic skin alterations. Interestingly, two independent studies
very recently demonstrated in two different murine models
that activated macrophages are essential for chronic psoria-
sis-like skin inflammation (154,155). TGF-a could play a
role as an intermediate in the TNF-a-elicited increase of
keratinocyte proliferation. Keratinocytes themselves pro-
duce TGF-a and this mediator even induces its own gene
expression (156). TNF-a has been shown to induce TGF-ain keratinocytes (157) and TGF-a messenger RNA and pro-
tein were much more abundant in lesional psoriatic epider-
mis than in normal-appearing skin of psoriatic patients or
in normal epidermis (158,159). TGF-a is known to bind to
the epidermal growth factor receptor and to stimulate
proliferation of keratinocytes and accelerates epidermal
regeneration (160). In contrast to TGF-a, mRNA levels of
TGF-b1, which inhibits epithelial cell growth (161), are not
significantly different in normal, uninvolved, and lesional
psoriatic epidermis (158). Apparently, during the course of
a psoriatic lesion, TGF-a and other factors, which are
themselves produced by keratinocytes, play a role in kerati-
nocyte proliferation.
NGF may be one such mediator. NGF is able to increase
the proliferation of keratinocytes through high-affinity
NGF receptors (trk) (162,163). Even if the expression of
this receptor appeared to be decreased by keratinocytes of
non-lesional and lesional skin in patients with psoriasis
(164), it is still possible that NGF contributes to the
increased proliferation of these cells. Two facts speak in
favour of this: (i) NGF is expressed at high levels by kerati-
nocytes in lesional and non-lesional skin (165), and (ii)
high-affinity nerve growth factor receptor blockers improve
psoriasis-like skin alterations in the severe combined
immunodeficient mouse–human skin model (166). At this
point, it must be emphasized that the effects of neuropep-
tides such as NGF are not limited to their impact on the
proliferation of keratinocytes in psoriasis, but can also
influence the angiogenesis, T-cell activation and prolifera-
tion of cutaneous nerve cells.
Healthy skin
DC
T-cell
T-cell
Macrophage
Fibroblast
Endothelial cell
AP1AP2
VEGFKGF
TNF-α
TNF-α
TGF-α
IL-19
IL-6
IL-17
IL-22 IL-22
IL-8
IL-23
IL-20
Psoriatic skin
Neutrophil
Figure 5. Various cell populations and their mediators are responsible
for the ‘keratinocyte response’ stage of our model of psoriasis
pathogenesis. Only the cell types and mediators, which are most
important for the induction of hyper-proliferation and altered
maturation of keratinocytes, are presented.
Immunopathogenesis of psoriasis
ª 2007 The Authors
Journal compilation ª 2007 Blackwell Munksgaard, Experimental Dermatology, 16, 779–798 789

Also, the major mediators that lead to altered maturation
of keratinocytes in psoriasis have not yet been clearly iden-
tified. In normal skin, the maturation of keratinocytes from
the basal layer to the cornified layer takes approximately
28 days. The last step of this maturation process is the ter-
minal differentiation. This is a particular apoptotic process
that results in the formation of the mechanically resistant
cornified layer (167). The manifold morphological changes
that take place during the transition from granular to cor-
nified cells include the synthesis of numerous proteins that
are encoded by the epidermal differentiation complex on
chromosome 1 (1q21), the aggregation of the keratin inter-
mediate filament network into macrofibrils, the synthesis of
extracellular lipids, and the dissolution of the nucleus and
other organelles (168). In psoriatic lesions, the maturation
is shortened to 5 days. This shortened maturation is associ-
ated with massively disrupted terminal differentiation of
keratinocytes and is mainly reflected by parakeratosis. Pso-
riatic lesions are also characterized by the absence of a
granular layer. Some markers, specific for granular layer in
normal skin such as involucrin and transglutaminase are
expressed in the spinous layer, while other granular mark-
ers such as filaggrin, are either absent or found in the para-
keratotic scales (169). Additionally, the expression of
proteins such as corneodesmosin, that is present in normal
skin in the cornified layer, is significantly increased and
also observed in the spinous layer in psoriatic lesions
(170). Furthermore, there is apparently a downregulation
of keratin K1 and K10, which are typically necessary for
terminal differentiation. Moreover, the production of extra-
cellular lipids is reduced and finally the retention of par-
tially differentiated keratinocytes causes typical psoriatic
scales.
We assume that IL-20 and IL-22 are the key mediators
that alter the terminal differentiation of psoriatic keratino-
cytes. As stated above, the main source of IL-22 in psoriatic
lesions should be the effector ⁄ memory Th17 and Th1 cells
(108–110). IL-22 affects tissue cells and not immune cells
(105,108). Keratinocytes are one of the most important
targets of these cytokines (105). IL-22 operates through a
receptor complex, which is composed of IL-22R1 and IL-
10R2 (171). IL-20 can affect keratinocytes via two different
receptor complexes (IL-20R1 ⁄ IL-20R2 and IL-22R1 ⁄ IL-
20R2). This sharing of identical receptors chains may not
be associated with binding competition or mutual limita-
tion of biological effects of the different mediators (172).
IL-22 regulates three functions of keratinocytes: (i) produc-
tion of antimicrobial proteins, (ii) differentiation and (iii)
migration (105,106,173).
In 2004, we could show that IL-22 strongly increases the
expression of the antimicrobially acting b-defensin 2 and
b-defensin 3 (105). It was the first discovered effect of IL-
22 on keratinocytes. More recently, we were able to show
that IL-22 also induces the expression of S100A7, S100A8
and S100A9 (106). The modes of action of b-defensins and
S100A proteins are different; S100A proteins act through
zinc sequestration, whereas b-defensins destabilize the
microbial membrane (174). This use of different mecha-
nisms suggests that IL-22 kills microbial pathogens very
efficiently and is therefore a very potent player in the anti-
microbial defense of the epidermis. Despite the troubled
integrity of the epidermis, cutaneous infections are not
common in psoriasis. Interestingly, the genes encoding
S100A7, S100A8 and S100A9 are located inside the epider-
mal differentiation complex.
More importantly for the psoriasis pathogenesis, IL-22
regulates the terminal differentiation of keratinocytes. In
fact, our data showed that IL-22 reduced the expression
of profilaggrin (FLG), K1, K10, calmodulin-like 5
(CALML5), keratinocyte differentiation-associated protein
(KDAP) and kallikrein 7 (KLK7) (106). Profilaggrin
(FLG) is known to be processed during terminal differen-
tiation by several proteases providing the N-terminal pep-
tide and several copies of filaggrin. The N-terminal
peptide translocates into the nucleus where it contributes
to the nuclear dissolution (175). Filaggrin binds to the
cytoplasmic keratin intermediate filament network and
aggregates it into macrofibrils (176). The keratins con-
tained in the macrofibrils are K1 and K10. Calmodulin-
like 5 is a calcium-binding protein that interacts with
transglutaminase 3, a key enzyme in the terminal differen-
tiation of keratinocytes (177). Transglutaminases are
known to crosslink many proteins in the cornified layer
(apart from keratins and filaggrin, these include loricrin
and involucrin), and contribute to the formation of the
cornified cell envelope (178,179). A serine protease
involved in the physiological detachment of corneocytes
from the stratum corneum is KLK7. It cleaves two adhe-
sive proteins from the extracellular part of corneodesmo-
somes, namely corneodesmosin and DSC1 (180). The
degradation of these proteins at the epidermal surface is
necessary for the physiological desquamation.
The last group of IL-22 sensitive genes is comprised of
genes that encode proteins responsible for cellular migra-
tion. IL-22 enhanced the expression of MMP1 and MMP3,
and reduced the expression of DSC1 (106). Metalloprotein-
ases constitute a family of structurally related zinc-depen-
dent neutral endopeptidases, which degrade the
extracellular matrix (181). MMP1 and MMP3 are inducible
and secreted, and their expressions are upregulated in psori-
atic lesions. MMP1 cleaves fibrillar type I collagen and is
needed to initiate keratinocyte migration. MMP3, a protease
with a wide range of substrate specificities, degrades old
multicellular actin networks thereby playing a role not only
in wound contraction but also in immune cell infiltration
of the skin (182). DSC1 is an adhesive desmosomal protein.
Sabat et al.
ª 2007 The Authors
790 Journal compilation ª 2007 Blackwell Munksgaard, Experimental Dermatology, 16, 779–798

Interestingly, two groups very recently described that
repeated cutaneous IL-23 application induced epidermal
hyperplasia with parakeratosis in mice (112,183). Moreover,
Zheng et al. demonstrated that these alterations were par-
tially dependent on IL-22 (183).
We postulate that IL-20 (and possibly IL-19 as well) has
effects similar to those of IL-22 because it acts through a
similar receptor (see above). We have demonstrated that
IL-20 is expressed in vitro in particular not only by kerati-
nocytes stimulated with pro-inflammatory cytokines but
also by activated monocytes (104,108). This is in line with
in situ hybridization data from the Kragballe group that
found IL-20 particularly in the suprapapillary epidermis in
psoriatic plaques close to macrophages (184). IL-20 affects
keratinocytes among other cellular targets (104,108). Inter-
estingly, TNF-a increases the sensitivity of keratinocytes to
IL-20 (185). Single-nucleotide polymorphisms of the IL-20
gene are associated with psoriasis (186,187). Even if the
effects of IL-20 on keratinocytes have not yet been
described in detail, the observations that (i) IL-20 is
strongly expressed in psoriatic skin (104,184,188,189) and
(ii) overexpression of IL-20 in transgenic mice causes skin
thickening with aberrant differentiation, let us assume that
IL-20 has an important role in psoriasis (185,190). Interest-
ingly, the skin aberrations in IL-20 transgenic mice are
without immune cell infiltration. This lets us postulate that
IL-20 could be a distal mediator of psoriasis pathogenesis.
IL-20 and IL-22 activate STAT3 in keratinocytes
(104,106). Here it may be interesting to mention that mice
that transgenically express a constitutively active STAT3
variant in keratinocytes have been shown to develop psori-
asis-like skin alterations (191). Furthermore, a transgenic
mice strain engineered with a deleted STAT3 gene in kerat-
inocytes developed relatively normal skin and hair follicles,
although the hair cycle and wound healing were severely
compromised (191).
Other characteristics of psoriatic lesions are the dilation
and increased number of dermal blood vessels. During
dilation, complement products and TNF-a are allowed to
play an important role. Angiogenesis is probably dependent
on vascular endothelial growth factor (VEGF) and angio-
protein 2. VEGF is a selective endothelial cell mitogen that
also enhances microvascular permeability. TGF-a induced
VEGF mRNA expression in cultured epidermal keratino-
cytes. Moreover, the hyperplastic epidermis of psoriatic
skin expresses strikingly increased amounts of VEGF, and
two VEGF receptors, kdr and flt-1, are overexpressed by
papillary dermal microvascular endothelial cells (192). An-
giopoietin 1, angiopoietin 2 and their receptor (Tie2) are
also upregulated in involved psoriatic skin compared with
uninvolved psoriatic skin and healthy skin. Angiopoietin 1
is expressed by stromal cells in the highly vascularized pap-
illary dermis and angiopoietin 2 is expressed by endothelial
cells in the vicinity of the proliferating epidermis that
abundantly expressed VEGF. VEGF was shown to enhance
angiopoietin 2 and Tie2 expression in dermal microvascu-
lar endothelial cell cultures (193). Angiopoietin 1 induces
Tie2 signalling as a receptor activator and maintains blood
vessel formation, whereas angiopoietin 2 destabilizes vessels
by blocking Tie2 signalling as an antagonist of angiopoietin
1 and acts with VEGF to initiate angiogenesis. In addition
to keratinocytes and stromal cells of the dermis, macro-
phages could also play a certain role in angiogenesis. By
the release of proteases, growth factors (basic fibroblast
growth factor, GM-CSF, TGF-a, IGF-I, PDGF and VEGF),
and other cytokines, activated macrophages have the capa-
bility to influence each phase of the angiogenic process
(194). Apparently, the direct effect of T cells on the new
angiogenesis is minimal. Actually, all IFNs dose- and time-
dependently inhibited the proliferation of endothelial cells
in vitro (195).
Conclusions
In this review, we postulate that psoriasis is an immunolog-
ically induced, overshot, regeneration-like reaction of the
skin in which different cells play a dominant role at differ-
ent stages. According to this hypothesis, the pathogenesis
of psoriasis can be basically subdivided into three phases:
(i) the sensitization phase, (ii) the silent phase and (iii) the
effector phase (Fig. 2). In the first phase, specific effector
Th17 and Th1 cells evolve from naıve T cells under the
influence of DCs in secondary lymphatic organs such as
the lymph nodes or tonsils (Fig. 3). In this process, DCs
should play a dominant role because they do not only
determine the specificity and the homing receptors but also
the characteristic functional phenotype of effector T cells
that determines their future action. As long as the sensitiza-
tion phase is not associated with an infection, it is clinically
unnoticeable and not characterized by any skin alterations.
Afterwards, a silent phase of variable duration occurs. The
third phase begins with the skin infiltration of various
immune cells (monocytes ⁄ macrophages, various subpopu-
lations of DCs, various subpopulations of T cells and neu-
trophilic granulocytes) (Fig. 4). The infiltration is made
easier by the fact that endothelial cells of the dermal blood
vessels in uninvolved skin from psoriasis patients already
express P- and E-selectin. Immigration is apparently initi-
ated by banal trauma or by invasion of a few microorgan-
isms (without clinical manifestation of infection) that leads
to the activation of local tissue macrophages, DCs and mast
cells. Finally, through the products of these cells and com-
plement components, endothelial cells are activated, which
then actively support the infiltration of immune cells
into the skin. Later in this phase, the immigrated immune
cells activate each other so that the T cells stimulate
Immunopathogenesis of psoriasis
ª 2007 The Authors
Journal compilation ª 2007 Blackwell Munksgaard, Experimental Dermatology, 16, 779–798 791

monocytes ⁄ macrophages and DCs through IFN-c and these
cells then activate the T cells through IL-23 and IL-6. At the
same time, keratinocytes are activated whereby they actively
support the infiltration of further immune cells through the
production of chemokines. As this phase continues, the
biology of the keratinocytes changes resulting in a massively
increased proliferation of these cells and their altered termi-
nal differentiation. Apparently, these changes are not
induced by IFN-c but rather by other T-cell cytokines (e.g.
IL-22, IL-17) and by mediators of macrophages ⁄ DCs (e.g.
TNF-a, IL-6). Still later in this phase, the mediators of mac-
rophages ⁄ DCs as well as the keratinocyte autocrine media-
tors (e.g. IL-20, TGF-a) and stromal cell mediators of the
dermis gain dominance in the activation of keratinocytes
(Fig. 5). In conclusion, the reaction of the keratinocytes
mimics an overshot wound-healing process. This model of
psoriasis pathogenesis is summarized in Fig. 6. A successful
therapy transposes the effector phase into the silent phase.
After a while, a renewed effector phase follows (Fig. 1).
Without an adequate treatment existing lesions are likely to
persist permanently in most cases.
We assume that two conditions must be met in order to
bring about the onset of psoriasis: the presence of activated
immune cells with a specific function ⁄ phenotype in the
skin, and the hyper-reactive dysfunction of keratinocytes.
The presence of activated immune cells in the skin is pre-
sumed to be dependent on the following factors: (i) the
generation of specific T1 and Th17 cells with skin-homing
receptors, (ii) the increased readiness of the endothelial
cells of dermal blood vessels in uninvolved skin to support
the infiltration of immune cells in the skin, (iii) the defi-
ciency of mechanisms in involved skin to inhibit the activa-
tion of immune cells. The preferred generation of T1 and
Th17 cells is probably contingent upon the orientation of
the immune system. In fact, patients with psoriasis also
commonly suffer from other inflammatory diseases (e.g.
rheumatoid arthritis, colitis). The increased readiness of the
endothelial cells of uninvolved skin could be because of a
genetic defect of these cells and ⁄ or could be because of a
hyper-reactivity of the tissue-localized immune cells such as
macrophages, DCs and mast cells that create a permanent
subinflammation condition in the skin without adequate
cause. The lack of mechanisms to inhibit immune cells in
involved skin could either be due to a functional deficiency
of regulatory cells (such as regulatory T cells or intraepithe-
lial T cells), a hyper-reactivity of the immune cells (that
may be due to a reduced effectiveness of intracellular nega-
tive regulators of cellular activation), and ⁄ or to a hyper-
reactivity of the keratinocytes and other tissue cells. It is
most probable that more than one of these causes exists
simultaneously. Some of them have been experimentally
confirmed (such as the deficiency of regulatory cells and
the hyper-reactivity of DCs in psoriatic lesions). We are
convinced that a hyper-reactive dysfunction of keratino-
cytes also exists and is a prerequisite for the initiation and
maintenance of psoriasis. This opinion is based on the exis-
tence of macroscopic (clinical) as well as microscopic (his-
tological) differences between chronic psoriasis and skin
Figure 6. Summary of our model of psoriasis pathogenesis.
Sabat et al.
ª 2007 The Authors
792 Journal compilation ª 2007 Blackwell Munksgaard, Experimental Dermatology, 16, 779–798

manifestations of chronic graft-versus-host disease
(GVHD). In both diseases, the first phase (the sensitization
phase) and the beginning of third phase (the effector
phase) of our model seem to be similar (generation of spe-
cific effector T cells, immune cell skin infiltration, produc-
tion of IFN-c by T1 cells). However, the results of the
third phase are completely different. In GVHD, the skin
does not show hyper-proliferation of keratinocytes or
enhanced desquamation. This means that although the
beginning of the immunological reaction may be similar,
the keratinocytes react differently in these two diseases.
This may be contingent upon genetically determined reac-
tion patterns of keratinocytes that should be different in
both diseases. Moreover, some gene products from the pso-
riasis-susceptibility loci regulate the differentiation of kerat-
inocytes (such as the genes from epidermal differentiation
complex in PSORS4) (8).
Our model of psoriasis explains the differences between
the observed therapeutic successes of various immune
treatments. It should be taken into account in which phase
of the pathogenesis these patients find themselves when
they are being treated. According to our model, patients
who already have existing macroscopic skin alterations are
currently in the last stage of the effector phase: in the
‘keratinocyte response’ stage. Consistent with this, therapies
that influence the beginning of the immune reaction have a
weak-efficiency over a short period. These are applications
that have the goals of (i) preventing the generation of
effector cells (CTLA-4-Ig), or (ii) inducing the emergence
of oppositionally polarized effector T cells (IL-10, and IL-
4). Such therapies can be useful only after a long treatment,
if indeed the already existing ‘pathological’ effector T cells
are no longer present. According to our model, also thera-
pies that influence the first stage of the effector phase (skin
infiltration of T and other immune cells) should have a
weak efficiency over a short period (196). In fact, despite
numerous attempts, no psoriasis therapy with medications
that interact with chemokines has been successful (197).
We postulate that therapies that act on the ‘keratinocyte
response’ stage will have the greatest success already after a
short treatment. According to our model, at the beginning
of this stage the ‘keratinocyte response’ is induced by T
cells and macrophages ⁄ DCs. As it continues, the macro-
phages ⁄ DCs and tissue cells are the main driving force.
Consistent with this, inhibition of T-cell function or T-cell
depletion is only successful in a portion of the patients
(198). In contrast, anti-TNF-a therapy appears to cause an
improvement in the skin alterations of all patients (198).
The anti-TNF-a therapy is apparently so effective for two
reasons; it leads to partial depletion of macrophages ⁄ DCs
and it neutralizes TNF-a, a cytokine that strengthens many
inflammatory processes of this immunologically induced,
overshot, regeneration-like reaction.
In the coming years, research will tell us to what
extent our model corresponds to the real pathogenesis of
psoriasis.
Acknowledgements
We would like to thank Dr Wolf-Dietrich Docke for very
helpful discussions and Sascha Rutz for the assistance in
the figure production. We also thank the German Ministry
of Education and Research (Bundesministerium fur
Bildung and Forschung) for their generous support.
References
1 Ferrandiz C, Bordas X, Garcia-Patos V, Puig S, Pujol R, Smandia A.
Prevalence of psoriasis in Spain (epiderma project: phase I). J Eur
Acad Dermatol Venereol 2001: 15: 20–23.
2 Nevitt G J, Hutchinson P E. Psoriasis in the community: prevalence,
severity and patients’ beliefs and attitudes towards the disease. Br
J Dermatol 1996: 135: 533–537.
3 Langley R G, Krueger G G, Griffiths C E. Psoriasis: epidemiology,
clinical features, and quality of life. Ann Rheum Dis 2005: 64 (Sup-
pl. 2): ii18–23; discussion ii24–25.
4 Schon M P, Boehncke W H. Psoriasis. N Engl J Med 2005: 352:
1899–1912.
5 Christophers E. Psoriasis – epidemiology and clinical spectrum. Clin
Exp Dermatol 2001: 26: 314–320.
6 Henseler T, Christophers E. Psoriasis of early and late onset: char-
acterization of two types of psoriasis vulgaris. J Am Acad Dermatol
1985: 13: 450–456.
7 Watson W, Cann H M, Farber E M, Nall M L. The genetics of psori-
asis. Arch Dermatol 1972: 105: 197–207.
8 Bowcock A M, Krueger J G. Getting under the skin: the immuno-
genetics of psoriasis. Nat Rev Immunol 2005: 5: 699–711.
9 Telfer N R, Chalmers R J, Whale K, Colman G. The role of strepto-
coccal infection in the initiation of guttate psoriasis. Arch Dermatol
1992: 128: 39–42.
10 Eyre R W, Krueger G G. Response to injury of skin involved and
uninvolved with psoriasis, and its relation to disease activity: Koeb-
ner and ‘reverse’ Koebner reactions. Br J Dermatol 1982: 106:
153–159.
11 Gudjonsson J E, Thorarinsson A M, Sigurgeirsson B, Kristinsson K
G, Valdimarsson H. Streptococcal throat infections and exacerba-
tion of chronic plaque psoriasis: a prospective study. Br J Dermatol
2003: 149: 530–534.
12 Tsankov N, Angelova I, Kazandjieva J. Drug-induced psoriasis. Rec-
ognition and management. Am J Clin Dermatol 2000: 1: 159–165.
13 Grover C, Reddy B S, Uma Chaturvedi K. Diagnosis of nail psoria-
sis: importance of biopsy and histopathology. Br J Dermatol 2005:
153: 1153–1158.
14 Zachariae H. Prevalence of joint disease in patients with psoriasis:
implications for therapy. Am J Clin Dermatol 2003: 4: 441–447.
15 Ragaz A, Ackerman A B. Evolution, maturation and regression of
lesions of psoriasis. New observations and correlation of clinical
and histologic findings. Am J Dermatopathol 1979: 1: 199–214.
16 Bos J D, Hulsebosch H J, Krieg S R, Bakker P M, Cormane R H.
Immunocompetent cells in psoriasis. In situ immunophenotyping by
monoclonal antibodies. Arch Dermatol Res 1983: 275: 181–189.
17 Hodge L, Comaish J S. Psoriasis: current concepts in management.
Drugs 1977: 13: 288–296.
Immunopathogenesis of psoriasis
ª 2007 The Authors
Journal compilation ª 2007 Blackwell Munksgaard, Experimental Dermatology, 16, 779–798 793

18 Voorhees J J. Pathophysiology of psoriasis. Annu Rev Med 1977:
28: 467–473.
19 Barker J N. Psoriasis as a T cell-mediated autoimmune disease.
Hosp Med 1998: 59: 530–533.
20 Ozawa M, Aiba S. Immunopathogenesis of psoriasis. Curr Drug
Targets Inflamm Allergy 2004: 3: 137–144.
21 Mueller W, Herrmann B. Cyclosporin A for psoriasis. N Engl J Med
1979: 301: 555.
22 Nicolas J F, Chamchick N, Thivolet J, Wijdenes J, Morel P, Revillard
J P. CD4 antibody treatment of severe psoriasis. Lancet 1991: 338:
321.
23 Prinz J, Braun-Falco O, Meurer M et al. Chimaeric CD4 monoclonal
antibody in treatment of generalized pustular psoriasis. Lancet
1991: 338: 320–321.
24 Gottlieb S L, Gilleaudeau P, Johnson R et al. Response of psoriasis
to a lymphocyte-selective toxin (DAB389IL-2) suggests a primary
immune, but not keratinocyte, pathogenic basis. Nat Med 1995: 1:
442–447.
25 Eedy D J, Burrows D, Bridges J M, Jones F G. Clearance of severe
psoriasis after allogenic bone marrow transplantation. BMJ 1990:
300: 908.
26 Gardembas-Pain M, Ifrah N, Foussard C, Boasson M, Saint Andre J P,
Verret J L. Psoriasis after allogeneic bone marrow transplantation.
Arch Dermatol 1990: 126: 1523.
27 Boehncke W H, Dressel D, Zollner T M, Kaufmann R. Pulling the
trigger on psoriasis. Nature 1996: 379: 777.
28 Nickoloff B J, Kunkel S L, Burdick M, Strieter R M. Severe combined
immunodeficiency mouse and human psoriatic skin chimeras. Vali-
dation of a new animal model. Am J Pathol 1995: 146: 580–588.
29 Wrone-Smith T, Nickoloff B J. Dermal injection of immunocytes
induces psoriasis. J Clin Invest 1996: 98: 1878–1887.
30 De Smedt T, Pajak B, Muraille E et al. Regulation of dendritic cell
numbers and maturation by lipopolysaccharide in vivo. J Exp Med
1996: 184: 1413–1424.
31 MacPherson G G, Jenkins C D, Stein M J, Edwards C. Endotoxin-
mediated dendritic cell release from the intestine. Characterization
of released dendritic cells and TNF dependence. J Immunol 1995:
154: 1317–1322.
32 MartIn-Fontecha A, Sebastiani S, Hopken U E et al. Regulation of
dendritic cell migration to the draining lymph node: impact on T
lymphocyte traffic and priming. J Exp Med 2003: 198: 615–621.
33 Sozzani S. Dendritic cell trafficking: more than just chemokines.
Cytokine Growth Factor Rev 2005: 16: 581–592.
34 Yanagihara S, Komura E, Nagafune J, Watarai H, Yamaguchi Y.
EBI1 ⁄ CCR7 is a new member of dendritic cell chemokine receptor
that is upregulated upon maturation. J Immunol 1998: 161:
3096–3102.
35 Garrett W S, Chen L M, Kroschewski R et al. Developmental control
of endocytosis in dendritic cells by Cdc42. Cell 2000: 102: 325–334.
36 Sallusto F, Cella M, Danieli C, Lanzavecchia A. Dendritic cells use
macropinocytosis and the mannose receptor to concentrate macro-
molecules in the major histocompatibility complex class II compart-
ment: downregulation by cytokines and bacterial products. J Exp
Med 1995: 182: 389–400.
37 Pierre P, Denzin L K, Hammond C et al. HLA-DM is localized to
conventional and unconventional MHC class II-containing endocytic
compartments. Immunity 1996: 4: 229–239.
38 Pierre P, Mellman I. Developmental regulation of invariant chain
proteolysis controls MHC class II trafficking in mouse dendritic
cells. Cell 1998: 93: 1135–1145.
39 Pierre P, Turley S J, Gatti E et al. Developmental regulation of
MHC class II transport in mouse dendritic cells. Nature 1997: 388:
787–792.
40 Trombetta E S, Ebersold M, Garrett W, Pypaert M, Mellman I. Acti-
vation of lysosomal function during dendritic cell maturation. Sci-
ence 2003: 299: 1400–1403.
41 Turley S J, Inaba K, Garrett W S et al. Transport of peptide-MHC
class II complexes in developing dendritic cells. Science 2000: 288:
522–527.
42 Watts C, Matthews S P, Mazzeo D, Manoury B, Moss C X. Aspar-
aginyl endopeptidase: case history of a class II MHC compartment
protease. Immunol Rev 2005: 207: 218–228.
43 Watts C. The exogenous pathway for antigen presentation on
major histocompatibility complex class II and CD1 molecules. Nat
Immunol 2004: 5: 685–692.
44 Nakagawa T Y, Brissette W H, Lira P D et al. Impaired invariant
chain degradation and antigen presentation and diminished colla-
gen-induced arthritis in cathepsin S null mice. Immunity 1999: 10:
207–217.
45 Shi G P, Villadangos J A, Dranoff G et al. Cathepsin S required for
normal MHC class II peptide loading and germinal center develop-
ment. Immunity 1999: 10: 197–206.
46 Villadangos J A, Schnorrer P, Wilson N S. Control of MHC class II
antigen presentation in dendritic cells: a balance between creative
and destructive forces. Immunol Rev 2005: 207: 191–205.
47 Cella M, Engering A, Pinet V, Pieters J, Lanzavecchia A. Inflamma-
tory stimuli induce accumulation of MHC class II complexes on
dendritic cells. Nature 1997: 388: 782–787.
48 Pure E, Inaba K, Crowley M T et al. Antigen processing by epider-
mal Langerhans cells correlates with the level of biosynthesis of
major histocompatibility complex class II molecules and expression
of invariant chain. J Exp Med 1990: 172: 1459–1469.
49 Landmann S, Muhlethaler-Mottet A, Bernasconi L et al. Maturation
of dendritic cells is accompanied by rapid transcriptional silencing
of class II transactivator (CIITA) expression. J Exp Med 2001: 194:
379–391.
50 Cresswell P, Ackerman A L, Giodini A, Peaper D R, Wearsch P A.
Mechanisms of MHC class I-restricted antigen processing and
cross-presentation. Immunol Rev 2005: 207: 145–157.
51 Baker B S, Bokth S, Powles A et al. Group A streptococcal antigen-
specific T lymphocytes in guttate psoriatic lesions. Br J Dermatol
1993: 128: 493–499.
52 Brown D W, Baker B S, Ovigne J M, Hardman C, Powles A V, Fry
L. Skin CD4+ T cells produce interferon-gamma in vitro in response
to streptococcal antigens in chronic plaque psoriasis. J Invest Der-
matol 2000: 114: 576–580.
53 Greenwald R J, Freeman G J, Sharpe A H. The B7 family revisited.
Annu Rev Immunol 2005: 23: 515–548.
54 Wang S, Chen L. T lymphocyte co-signaling pathways of the
B7-CD28 family. Cell Mol Immunol 2004: 1: 37–42.
55 Bachmann M F, McKall-Faienza K, Schmits R et al. Distinct roles
for LFA-1 and CD28 during activation of naive T cells: adhesion
versus costimulation. Immunity 1997: 7: 549–557.
56 Albanesi C, Scarponi C, Cavani A, Federici M, Nasorri F, Girolomoni
G. Interleukin-17 is produced by both Th1 and Th2 lymphocytes,
and modulates interferon-gamma- and interleukin-4-induced
activation of human keratinocytes. J Invest Dermatol 2000: 115:
81–87.
57 Oki S, Kohsaka T, Azuma M. Augmentation of CTLA-4 expression
by wortmannin: involvement of lysosomal sorting properties of
CTLA-4. Int Immunol 1999: 11: 1563–1571.
58 Krummel M F, Allison J P. CD28 and CTLA-4 have opposing effects
on the response of T cells to stimulation. J Exp Med 1995: 182:
459–465.
59 Sims T N, Dustin M L. The immunological synapse: integrins take
the stage. Immunol Rev 2002: 186: 100–117.
Sabat et al.
ª 2007 The Authors
794 Journal compilation ª 2007 Blackwell Munksgaard, Experimental Dermatology, 16, 779–798

60 Weaver C T, Harrington L E, Mangan P R, Gavrieli M, Murphy K
M. Th17: an effector CD4 T cell lineage with regulatory T cell ties.
Immunity 2006: 24: 677–688.
61 Harrington L E, Hatton R D, Mangan P R et al. Interleukin 17-produc-
ing CD4+ effector T cells develop via a lineage distinct from the T
helper type 1 and 2 lineages. Nat Immunol 2005: 6: 1123–1132.
62 Park H, Li Z, Yang X O et al. A distinct lineage of CD4 T cells regu-
lates tissue inflammation by producing interleukin 17. Nat Immunol
2005: 6: 1133–1141.
63 Bonecchi R, Bianchi G, Bordignon P P et al. Differential expression
of chemokine receptors and chemotactic responsiveness of type 1
T helper cells (Th1s) and Th2s. J Exp Med 1998: 187: 129–134.
64 Loetscher P, Uguccioni M, Bordoli L et al. CCR5 is characteristic of
Th1 lymphocytes. Nature 1998: 391: 344–345.
65 Till S, Jopling L, Wachholz P et al. T cell phenotypes of the normal
nasal mucosa: induction of Th2 cytokines and CCR3 expression by
IL-4. J Immunol 2001: 166: 2303–2310.
66 Banham A H, Powrie F M, Suri-Payer E. FOXP3+ regulatory T cells:
current controversies and future perspectives. Eur J Immunol 2006:
36: 2832–2836.
67 Sallusto F, Geginat J, Lanzavecchia A. Central memory and effector
memory T cell subsets: function, generation and maintenance.
Annu Rev Immunol 2004: 22: 745–763.
68 Bingaman A W, Patke D S, Mane V R et al. Novel phenotypes and
migratory properties distinguish memory CD4 T cell subsets in lym-
phoid and lung tissue. Eur J Immunol 2005: 35: 3173–3186.
69 Schwendemann J, Choi C, Schirrmacher V, Beckhove P. Dynamic
differentiation of activated human peripheral blood CD8+ and
CD4+ effector memory T cells. J Immunol 2005: 175: 1433–1439.
70 Fuhlbrigge R C, Kieffer J D, Armerding D, Kupper T S. Cutaneous
lymphocyte antigen is a specialized form of PSGL-1 expressed on
skin-homing T cells. Nature 1997: 389: 978–981.
71 Weninger W, Ulfman L H, Cheng G et al. Specialized contributions
by alpha(1,3)-fucosyltransferase-IV and FucT-VII during leukocyte
rolling in dermal microvessels. Immunity 2000: 12: 665–676.
72 Fuhlbrigge R C, King S L, Sackstein R, Kupper T S. CD43 is a ligand
for E-selectin on CLA+ human T cells. Blood 2006: 107: 1421–1426.
73 Diluvio L, Vollmer S, Besgen P, Ellwart J W, Chimenti S, Prinz J C.
Identical TCR beta-chain rearrangements in streptococcal angina
and skin lesions of patients with psoriasis vulgaris. J Immunol
2006: 176: 7104–7111.
74 Weisenseel P, Prinz J C. Incidental detection of S. pyogenes-DNA
in psoriatic skin by PCR. Arch Dermatol Res 2005: 296: 573–576.
75 Terajima S, Higaki M, Igarashi Y, Nogita T, Kawashima M. An
important role of tumor necrosis factor-alpha in the induction of
adhesion molecules in psoriasis. Arch Dermatol Res 1998: 290:
246–252.
76 Campbell J J, Haraldsen G, Pan J et al. The chemokine receptor
CCR4 in vascular recognition by cutaneous but not intestinal mem-
ory T cells. Nature 1999: 400: 776–780.
77 Soler D, Humphreys T L, Spinola S M, Campbell J J. CCR4 versus
CCR10 in human cutaneous TH lymphocyte trafficking. Blood
2003: 101: 1677–1682.
78 Homey B, Alenius H, Muller A et al. CCL27-CCR10 interactions
regulate T cell-mediated skin inflammation. Nat Med 2002: 8:
157–165.
79 Baekkevold E S, Wurbel M A, Kivisakk P et al. A role for CCR4 in
development of mature circulating cutaneous T helper memory cell
populations. J Exp Med 2005: 201: 1045–1051.
80 Reiss Y, Proudfoot A E, Power C A, Campbell J J, Butcher E C. CC
chemokine receptor (CCR)4 and the CCR10 ligand cutaneous T
cell-attracting chemokine (CTACK) in lymphocyte trafficking to
inflamed skin. J Exp Med 2001: 194: 1541–1547.
81 Teraki Y, Miyake A, Takebayashi R, Shiohara T. Homing receptor
and chemokine receptor on intraepidermal T cells in psoriasis vul-
garis. Clin Exp Dermatol 2004: 29: 658–663.
82 Rottman J B, Smith T L, Ganley K G, Kikuchi T, Krueger J G. Poten-
tial role of the chemokine receptors CXCR3, CCR4, and the inte-
grin alphaEbeta7 in the pathogenesis of psoriasis vulgaris. Lab
Invest 2001: 81: 335–347.
83 Shamri R, Grabovsky V, Gauguet J M et al. Lymphocyte arrest
requires instantaneous induction of an extended LFA-1 conforma-
tion mediated by endothelium-bound chemokines. Nat Immunol
2005: 6: 497–506.
84 Grabovsky V, Feigelson S, Chen C et al. Subsecond induction of
alpha4 integrin clustering by immobilized chemokines stimulates
leukocyte tethering and rolling on endothelial vascular cell adhesion
molecule 1 under flow conditions. J Exp Med 2000: 192: 495–
506.
85 Detmar M, Tenorio S, Hettmannsperger U, Ruszczak Z, Orfanos C E.
Cytokine regulation of proliferation and ICAM-1 expression of
human dermal microvascular endothelial cells in vitro. J Invest Der-
matol 1992: 98: 147–153.
86 Ruzicka T, Sommerburg C, Braun-Falco O et al. Efficiency of acitre-
tin in combination with UV-B in the treatment of severe psoriasis.
Arch Dermatol 1990: 126: 482–486.
87 Ding Z, Xiong K, Issekutz T B. Chemokines stimulate human T lym-
phocyte transendothelial migration to utilize VLA-4 in addition to
LFA-1. J Leukoc Biol 2001: 69: 458–466.
88 Griffiths C E, Voorhees J J, Nickoloff B J. Characterization of inter-
cellular adhesion molecule-1 and HLA-DR expression in normal and
inflamed skin: modulation by recombinant gamma interferon and
tumor necrosis factor. J Am Acad Dermatol 1989: 20: 617–629.
89 Lowes M A, Chamian F, Abello M V et al. Increase in TNF-alpha
and inducible nitric oxide synthase-expressing dendritic cells in pso-
riasis and reduction with efalizumab (anti-CD11a). Proc Natl Acad
Sci U S A 2005: 102: 19057–19062.
90 Bos J D, Hagenaars C, Das P K, Krieg S R, Voorn W J, Kapsenberg
M L. Predominance of ‘memory’ T cells (CD4+, CDw29+) over
‘naive’ T cells (CD4+, CD45R+) in both normal and diseased
human skin. Arch Dermatol Res 1989: 281: 24–30.
91 Bovenschen H J, Seyger M M, Van de Kerkhof P C. Plaque psoriasis
vs. atopic dermatitis and lichen planus: a comparison for lesional
T-cell subsets, epidermal proliferation and differentiation. Br J Der-
matol 2005: 153: 72–78.
92 Pauls K, Schon M, Kubitza R C et al. Role of integrin alp-
haE(CD103)beta7 for tissue-specific epidermal localization of CD8+
T lymphocytes. J Invest Dermatol 2001: 117: 569–575.
93 Teraki Y, Shiohara T. Preferential expression of alphaEbeta7 inte-
grin (CD103) on CD8+ T cells in the psoriatic epidermis: regulation
by interleukins 4 and 12 and transforming growth factor-beta. Br J
Dermatol 2002: 147: 1118–1126.
94 Cepek K L, Shaw S K, Parker C M et al. Adhesion between epithe-
lial cells and T lymphocytes mediated by E-cadherin and the alpha
E beta 7 integrin. Nature 1994: 372: 190–193.
95 Ottaviani C, Nasorri F, Bedini C, de Pita O, Girolomoni G, Cavani
A. CD56brightCD16(-) NK cells accumulate in psoriatic skin in
response to CXCL10 and CCL5 and exacerbate skin inflammation.
Eur J Immunol 2006: 36: 118–128.
96 Mrowietz U, Koch W A, Zhu K et al. Psoriasis scales contain C5a
as the predominant chemotaxin for monocyte-derived dendritic
cells. Exp Dermatol 2001: 10: 238–245.
97 Dieu-Nosjean M C, Massacrier C, Homey B et al. Macrophage
inflammatory protein 3alpha is expressed at inflamed epithelial sur-
faces and is the most potent chemokine known in attracting lan-
gerhans cell precursors. J Exp Med 2000: 192: 705–718.
Immunopathogenesis of psoriasis
ª 2007 The Authors
Journal compilation ª 2007 Blackwell Munksgaard, Experimental Dermatology, 16, 779–798 795

98 Wetzel A, Wetzig T, Haustein U F et al. Increased neutrophil
adherence in psoriasis: role of the human endothelial cell receptor
Thy-1 (CD90). J Invest Dermatol 2006: 126: 441–452.
99 Bjerke J R, Krogh H K, Matre R. Characterization of mononuclear
cell infiltrates in psoriatic lesions. J Invest Dermatol 1978: 71:
340–343.
100 Servitje O, Bordas X, Seron D et al. Changes in T-cell phenotype
and adhesion molecules expression in psoriatic lesions after low-
dose cyclosporin therapy. J Cutan Pathol 1996: 23: 431–436.
101 Ferenczi K, Burack L, Pope M, Krueger J G, Austin L M. CD69,
HLA-DR and the IL-2R identify persistently activated T cells in psori-
asis vulgaris lesional skin: blood and skin comparisons by flow
cytometry. J Autoimmun 2000: 14: 63–78.
102 Schlaak J F, Buslau M, Jochum W et al. T cells involved in psoriasis
vulgaris belong to the Th1 subset. J Invest Dermatol 1994: 102:
145–149.
103 Austin L M, Ozawa M, Kikuchi T, Walters I B, Krueger J G. The
majority of epidermal T cells in psoriasis vulgaris lesions can pro-
duce type 1 cytokines, interferon-gamma, interleukin-2 and tumor
necrosis factor-alpha, defining TC1 (cytotoxic T lymphocyte) and
TH1 effector populations: a type 1 differentiation bias is also mea-
sured in circulating blood T cells in psoriatic patients. J Invest Der-
matol 1999: 113: 752–759.
104 Kunz S, Wolk K, Witte E et al. Interleukin (IL)-19, IL-20 and IL-24
are produced by and act on keratinocytes and are distinct from
classical ILs. Exp Dermatol 2006: 15: 991–1004.
105 Wolk K, Kunz S, Witte E, Friedrich M, Asadullah K, Sabat R. IL-22
increases the innate immunity of tissues. Immunity 2004: 21:
241–254.
106 Wolk K, Witte E, Wallace E et al. IL-22 regulates the expression of
genes responsible for antimicrobial defense, cellular differentiation
and mobility in keratinocytes: a potential role in psoriasis. Eur J
Immunol 2006: 36: 1309–1323.
107 Wolk K, Sabat R. Interleukin-22: a novel T- and NK-cell derived
cytokine that regulates the biology of tissue cells. Cytokine Growth
Factor Rev 2006: 17: 367–380.
108 Wolk K, Kunz S, Asadullah K, Sabat R. Cutting edge: immune cells
as sources and targets of the IL-10 family members? J Immunol
2002: 168: 5397–5402.
109 Chung Y, Yang X, Chang S H, Ma L, Tian Q, Dong C. Expression
and regulation of IL-22 in the IL-17-producing CD4+ T lympho-
cytes. Cell Res 2006: 16: 902–907.
110 Liang S C, Tan X Y, Luxenberg D P et al. Interleukin (IL)-22 and
IL-17 are coexpressed by Th17 cells and cooperatively enhance
expression of antimicrobial peptides. J Exp Med 2006: 203:
2271–2279.
111 Teunissen M B, Koomen C W, de Waal Malefyt R, Wierenga E A,
Bos J D. Interleukin-17 and interferon-gamma synergize in the
enhancement of proinflammatory cytokine production by human
keratinocytes. J Invest Dermatol 1998: 111: 645–649.
112 Chan J R, Blumenschein W, Murphy E et al. IL-23 stimulates epi-
dermal hyperplasia via TNF and IL-20R2-dependent mechanisms
with implications for psoriasis pathogenesis. J Exp Med 2006: 203:
2577–2587.
113 Cargill M, Schrodi S J, Chang M et al. A large-scale genetic associ-
ation study confirms IL12B and leads to the identification of IL23R
as psoriasis-risk genes. Am J Hum Genet 2007: 80: 273–290.
114 Cerio R, Griffiths C E, Cooper K D, Nickoloff B J, Headington J T.
Characterization of factor XIIIa positive dermal dendritic cells in
normal and inflamed skin. Br J Dermatol 1989: 121: 421–431.
115 Djemadji-Oudjiel N, Goerdt S, Kodelja V, Schmuth M, Orfanos C E.
Immunohistochemical identification of type II alternatively activated
dendritic macrophages (RM 3 ⁄ 1 + 3, MS-1+ ⁄ -, 25 F9-) in psoriatic
dermis. Arch Dermatol Res 1996: 288: 757–764.
116 van den Oord J J, de Wolf-Peeters C. Epithelium-lining macrophag-
es in psoriasis. Br J Dermatol 1994: 130: 589–594.
117 Guttman-Yassky E, Lowes M A, Fuentes-Duculan J et al. Major dif-
ferences in inflammatory dendritic cells and their products distin-
guish atopic dermatitis from psoriasis. J Allergy Clin Immunol
2007: 119: 1210–1217.
118 Angel C E, George E, Brooks A E, Ostrovsky L L, Brown T L, Dunbar P
R. Cutting edge: CD1a+ antigen-presenting cells in human dermis
respond rapidly to CCR7 ligands. J Immunol 2006: 176: 5730–5734.
119 Wollenberg A, Wagner M, Gunther S et al. Plasmacytoid dendritic
cells: a new cutaneous dendritic cell subset with distinct role in
inflammatory skin diseases. J Invest Dermatol 2002: 119: 1096–
1102.
120 Gilliet M, Conrad C, Geiges M et al. Psoriasis triggered by toll-like
receptor 7 agonist imiquimod in the presence of dermal plasmacy-
toid dendritic cell precursors. Arch Dermatol 2004: 140: 1490–
1495.
121 Lee E, Trepicchio W L, Oestreicher J L et al. Increased expression of
interleukin 23 p19 and p40 in lesional skin of patients with psoria-
sis vulgaris. J Exp Med 2004: 199: 125–130.
122 Cameron A L, Kirby B, Fei W, Griffiths C E. Natural killer and natu-
ral killer-T cells in psoriasis. Arch Dermatol Res 2002: 294: 363–
369.
123 Wilson S B, Delovitch T L. Janus-like role of regulatory iNKT cells in
autoimmune disease and tumour immunity. Nat Rev Immunol
2003: 3: 211–222.
124 Bata-Csorgo Z, Hammerberg C, Voorhees J J, Cooper K D. Flow
cytometric identification of proliferative subpopulations within
normal human epidermis and the localization of the primary
hyperproliferative population in psoriasis. J Exp Med 1993: 178:
1271–1281.
125 Boyman O, Hefti H P, Conrad C, Nickoloff B J, Suter M, Nestle F
O. Spontaneous development of psoriasis in a new animal model
shows an essential role for resident T cells and tumor necrosis fac-
tor-alpha. J Exp Med 2004: 199: 731–736.
126 Nestle F O, Turka L A, Nickoloff B J. Characterization of dermal
dendritic cells in psoriasis. Autostimulation of T lymphocytes and
induction of Th1 type cytokines. J Clin Invest 1994: 94: 202–209.
127 Nestle F O, Conrad C, Tun-Kyi A et al. Plasmacytoid predendritic
cells initiate psoriasis through interferon-alpha production. J Exp
Med 2005: 202: 135–143.
128 Eriksen K W, Lovato P, Skov L et al. Increased sensitivity to inter-
feron-alpha in psoriatic T cells. J Invest Dermatol 2005: 125: 936–
944.
129 Owen C M, Chalmers R J, O’Sullivan T, Griffiths C E. A systematic
review of antistreptococcal interventions for guttate and chronic
plaque psoriasis. Br J Dermatol 2001: 145: 886–890.
130 Wilson J K, Al-Suwaidan S N, Krowchuk D, Feldman S R. Treat-
ment of psoriasis in children: is there a role for antibiotic therapy
and tonsillectomy? Pediatr Dermatol 2003: 20: 11–15.
131 Prinz J C. Disease mimicry – a pathogenetic concept for T cell-
mediated autoimmune disorders triggered by molecular mimicry?
Autoimmun Rev 2004: 3: 10–15.
132 McFadden J, Valdimarsson H, Fry L. Cross-reactivity between strep-
tococcal M surface antigen and human skin. Br J Dermatol 1991:
125: 443–447.
133 Gudmundsdottir A S, Sigmundsdottir H, Sigurgeirsson B, Good M
F, Valdimarsson H, Jonsdottir I. Is an epitope on keratin 17 a major
target for autoreactive T lymphocytes in psoriasis? Clin Exp Immu-
nol 1999: 117: 580–586.
Sabat et al.
ª 2007 The Authors
796 Journal compilation ª 2007 Blackwell Munksgaard, Experimental Dermatology, 16, 779–798

134 Johnston A, Gudjonsson J E, Sigmundsdottir H, Love T J, Valdi-
marsson H. Peripheral blood T cell responses to keratin peptides
that share sequences with streptococcal M proteins are largely
restricted to skin-homing CD8(+) T cells. Clin Exp Immunol 2004:
138: 83–93.
135 de Jong E M, van Vlijmen I M, van Erp P E, Ramaekers F C, Troya-
novski S M, van de Kerkhof P C. Keratin 17: a useful marker in
anti-psoriatic therapies. Arch Dermatol Res 1991: 283: 480–482.
136 Leigh I M, Navsaria H, Purkis P E, McKay I A, Bowden P E, Riddle P
N. Keratins (K16 and K17) as markers of keratinocyte hyperprolif-
eration in psoriasis in vivo and in vitro. Br J Dermatol 1995: 133:
501–511.
137 Bockelmann R, Horn T, Gollnick H, Bonnekoh B. Interferon-
gamma-dependent in vitro model for the putative keratin 17 auto-
immune loop in psoriasis: exploration of pharmaco- and gene-ther-
apeutic effects. Skin Pharmacol Physiol 2005: 18: 42–54.
138 Bonnekoh B, Huerkamp C, Wevers A et al. Up-regulation of kera-
tin 17 expression in human HaCaT keratinocytes by interferon-
gamma. J Invest Dermatol 1995: 104: 58–61.
139 Griffiths C E, Voorhees J J, Nickoloff B J. Gamma interferon
induces different keratinocyte cellular patterns of expression of
HLA-DR and DQ and intercellular adhesion molecule-I (ICAM-I)
antigens. Br J Dermatol 1989: 120: 1–8.
140 Nickoloff B J, Mitra R S, Green J et al. Accessory cell function of
keratinocytes for superantigens. Dependence on lymphocyte func-
tion-associated antigen-1 ⁄ intercellular adhesion molecule-1 interac-
tion. J Immunol 1993: 150: 2148–2159.
141 Travers J B, Hamid Q A, Norris D A et al. Epidermal HLA-DR and
the enhancement of cutaneous reactivity to superantigenic toxins
in psoriasis. J Clin Invest 1999: 104: 1181–1189.
142 Bonish B, Jullien D, Dutronc Y et al. Overexpression of CD1d by
keratinocytes in psoriasis and CD1d-dependent IFN-gamma pro-
duction by NK-T cells. J Immunol 2000: 165: 4076–4085.
143 Girardi M. Cutaneous biology of gammadelta T cells. Adv Dermatol
2004: 20: 203–215.
144 Girardi M, Lewis J, Glusac E et al. Resident skin-specific gam-
madelta T cells provide local, nonredundant regulation of cutane-
ous inflammation. J Exp Med 2002: 195: 855–867.
145 Sugiyama H, Gyulai R, Toichi E et al. Dysfunctional blood and tar-
get tissue CD4+CD25high regulatory T cells in psoriasis: mecha-
nism underlying unrestrained pathogenic effector T cell
proliferation. J Immunol 2005: 174: 164–173.
146 Nickoloff B J, Karabin G D, Barker J N et al. Cellular localization of
interleukin-8 and its inducer, tumor necrosis factor-alpha in psoria-
sis. Am J Pathol 1991: 138: 129–140.
147 Kristensen M, Chu C Q, Eedy D J, Feldmann M, Brennan F M,
Breathnach S M. Localization of tumour necrosis factor-alpha (TNF-
alpha) and its receptors in normal and psoriatic skin: epidermal
cells express the 55-kD but not the 75-kD TNF receptor. Clin Exp
Immunol 1993: 94: 354–362.
148 Hancock G E, Kaplan G, Cohn Z A. Keratinocyte growth regulation
by the products of immune cells. J Exp Med 1988: 168:
1395–1402.
149 Bata-Csorgo Z, Hammerberg C, Voorhees J J, Cooper K D. Kinetics
and regulation of human keratinocyte stem cell growth in short-
term primary ex vivo culture. Cooperative growth factors from
psoriatic lesional T lymphocytes stimulate proliferation among pso-
riatic uninvolved, but not normal, stem keratinocytes. J Clin Invest
1995: 95: 317–327.
150 Olaniran A K, Baker B S, Garioch J J, Powles A V, Fry L. A compari-
son of the stimulatory effects of cytokines on normal and psoriatic
keratinocytes in vitro. Arch Dermatol Res 1995: 287: 231–236.
151 Jackson M, Howie S E, Weller R, Sabin E, Hunter J A, McKenzie R
C. Psoriatic keratinocytes show reduced IRF-1 and STAT-1alpha
activation in response to gamma-IFN. FASEB J 1999: 13: 495–502.
152 Rogalski C, Meyer-Hoffert U, Proksch E, Wiedow O. Human leuko-
cyte elastase induces keratinocyte proliferation in vitro and in vivo.
J Invest Dermatol 2002: 118: 49–54.
153 Numerof R P, Asadullah K. Cytokine and anti-cytokine therapies
for psoriasis and atopic dermatitis. BioDrugs 2006: 20: 93–103.
154 Stratis A, Pasparakis M, Rupec R A et al. Pathogenic role for skin
macrophages in a mouse model of keratinocyte-induced psoriasis-
like skin inflammation. J Clin Invest 2006: 116: 2094–2104.
155 Wang H, Peters T, Kess D et al. Activated macrophages are essen-
tial in a murine model for T cell-mediated chronic psoriasiform skin
inflammation. J Clin Invest 2006: 116: 2105–2114.
156 Coffey R J Jr, Derynck R, Wilcox J N et al. Production and auto-
induction of transforming growth factor-alpha in human keratino-
cytes. Nature 1987: 328: 817–820.
157 Valyi-Nagy I, Jensen P J, Albelda S M, Rodeck U. Cytokine-induced
expression of transforming growth factor-alpha and the epidermal
growth factor receptor in neonatal skin explants. J Invest Dermatol
1992: 99: 350–356.
158 Elder J T, Fisher G J, Lindquist P B et al. Overexpression of trans-
forming growth factor alpha in psoriatic epidermis. Science 1989:
243: 811–814.
159 Higashiyama M, Matsumoto K, Hashimoto K, Yoshikawa K.
Increased production of transforming growth factor-alpha in psori-
atic epidermis. J Dermatol 1991: 18: 117–119.
160 Schultz G S, White M, Mitchell R et al. Epithelial wound healing
enhanced by transforming growth factor-alpha and vaccinia
growth factor. Science 1987: 235: 350–352.
161 Malkani A K, Baker B S, Garioch J J et al. Normal response to
tumor necrosis factor-alpha and transforming growth factor-beta
by keratinocytes in psoriasis. Exp Dermatol 1993: 2: 224–230.
162 Di Marco E, Mathor M, Bondanza S et al. Nerve growth factor
binds to normal human keratinocytes through high and low affin-
ity receptors and stimulates their growth by a novel autocrine loop.
J Biol Chem 1993: 268: 22838–22846.
163 Pincelli C, Sevignani C, Manfredini R et al. Expression and function
of nerve growth factor and nerve growth factor receptor on cul-
tured keratinocytes. J Invest Dermatol 1994: 103: 13–18.
164 Bull H A, Leslie T A, Chopra S, Dowd P M. Expression of nerve
growth factor receptors in cutaneous inflammation. Br J Dermatol
1998: 139: 776–783.
165 Fantini F, Magnoni C, Bracci-Laudiero L, Pincelli C T. Nerve growth
factor is increased in psoriatic skin. J Invest Dermatol 1995: 105:
854–855.
166 Raychaudhuri S P, Sanyal M, Weltman H, Kundu-Raychaudhuri S.
K252a, a high-affinity nerve growth factor receptor blocker,
improves psoriasis: an in vivo study using the severe combined
immunodeficient mouse-human skin model. J Invest Dermatol
2004: 122: 812–819.
167 Candi E, Schmidt R, Melino G. The cornified envelope: a model of
cell death in the skin. Nat Rev Mol Cell Biol 2005: 6: 328–340.
168 Fuchs E. Epidermal differentiation: the bare essentials. J Cell Biol
1990: 111: 2807–2814.
169 Bernard B A, Asselineau D, Schaffar-Deshayes L, Darmon M Y.
Abnormal sequence of expression of differentiation markers in
psoriatic epidermis: inversion of two steps in the differentiation
program? J Invest Dermatol 1988: 90: 801–805.
170 Allen M, Ishida-Yamamoto A, McGrath J et al. Corneodesmosin
expression in psoriasis vulgaris differs from normal skin and other
inflammatory skin disorders. Lab Invest 2001: 81: 969–976.
Immunopathogenesis of psoriasis
ª 2007 The Authors
Journal compilation ª 2007 Blackwell Munksgaard, Experimental Dermatology, 16, 779–798 797

171 Xie M H, Aggarwal S, Ho W H et al. Interleukin (IL)-22, a novel
human cytokine that signals through the interferon receptor-
related proteins CRF2-4 and IL-22R. J Biol Chem 2000: 275:
31335–31339.
172 Wolk K, Witte E, Reineke U et al. Is there an interaction between
interleukin-10 and interleukin-22? Genes Immun 2005: 6: 8–18.
173 Boniface K, Lecron J C, Bernard F X et al. Keratinocytes as targets
for interleukin-10-related cytokines: a putative role in the patho-
genesis of psoriasis. Eur Cytokine Netw 2005: 16: 309–319.
174 Harder J, Schroder J M. Psoriatic scales: a promising source for the
isolation of human skin-derived antimicrobial proteins. J Leukoc
Biol 2005: 77: 476–486.
175 Pearton D J, Dale B A, Presland R B. Functional analysis of the
profilaggrin N-terminal peptide: identification of domains that reg-
ulate nuclear and cytoplasmic distribution. J Invest Dermatol 2002:
119: 661–669.
176 Kuechle M K, Thulin C D, Presland R B, Dale B A. Profilaggrin
requires both linker and filaggrin peptide sequences to form gran-
ules: implications for profilaggrin processing in vivo. J Invest Der-
matol 1999: 112: 843–852.
177 Mehul B, Bernard D, Simonetti L, Bernard M A, Schmidt R. Identifi-
cation and cloning of a new calmodulin-like protein from human
epidermis. J Biol Chem 2000: 275: 12841–12847.
178 Fuchs E. Keratins and the skin. Annu Rev Cell Dev Biol 1995: 11:
123–153.
179 Fuchs E, Weber K. Intermediate filaments: structure, dynamics,
function and disease. Annu Rev Biochem 1994: 63: 345–382.
180 Caubet C, Jonca N, Brattsand M et al. Degradation of corneodes-
mosome proteins by two serine proteases of the kallikrein family,
SCTE ⁄ KLK5 ⁄ hK5 and SCCE ⁄ KLK7 ⁄ hK7. J Invest Dermatol 2004:
122: 1235–1244.
181 Nagase H, Woessner J F Jr. Matrix metalloproteinases. J Biol Chem
1999: 274: 21491–21494.
182 Bullard K M, Lund L, Mudgett J S et al. Impaired wound contraction
in stromelysin-1-deficient mice. Ann Surg 1999: 230: 260–265.
183 Zheng Y, Danilenko D M, Valdez P et al. Interleukin-22, a T(H)17
cytokine, mediates IL-23-induced dermal inflammation and acan-
thosis. Nature 2007: 445: 648–651.
184 Romer J, Hasselager E, Norby P L, Steiniche T, Thorn Clausen J,
Kragballe K. Epidermal overexpression of interleukin-19 and -20
mRNA in psoriatic skin disappears after short-term treatment with
cyclosporine a or calcipotriol. J Invest Dermatol 2003: 121:
1306–1311.
185 Blumberg H, Conklin D, Xu W F et al. Interleukin 20: discovery,
receptor identification and role in epidermal function. Cell 2001:
104: 9–19.
186 Koks S, Kingo K, Ratsep R, Karelson M, Silm H, Vasar E. Combined
haplotype analysis of the interleukin-19 and -20 genes: relationship
to plaque-type psoriasis. Genes Immun 2004: 5: 662–667.
187 Koks S, Kingo K, Vabrit K et al. Possible relations between the
polymorphisms of the cytokines IL-19, IL-20 and IL-24 and plaque-
type psoriasis. Genes Immun 2005: 6: 407–415.
188 Otkjaer K, Kragballe K, Funding A T et al. The dynamics of gene
expression of interleukin-19 and interleukin-20 and their receptors
in psoriasis. Br J Dermatol 2005: 153: 911–918.
189 Wei C C, Chen W Y, Wang Y C et al. Detection of IL-20 and its
receptors on psoriatic skin. Clin Immunol 2005: 117: 65–72.
190 Liu L, Ding C, Zeng W et al. Selective enhancement of multipoten-
tial hematopoietic progenitors in vitro and in vivo by IL-20. Blood
2003: 102: 3206–3209.
191 Sano S, Chan K S, Carbajal S et al. Stat3 links activated keratino-
cytes and immunocytes required for development of psoriasis in a
novel transgenic mouse model. Nat Med 2005: 11: 43–49.
192 Detmar M, Brown L F, Claffey K P et al. Overexpression of vascular
permeability factor ⁄ vascular endothelial growth factor and its
receptors in psoriasis. J Exp Med 1994: 180: 1141–1146.
193 Kuroda K, Sapadin A, Shoji T, Fleischmajer R, Lebwohl M. Altered
expression of angiopoietins and Tie2 endothelium receptor in pso-
riasis. J Invest Dermatol 2001: 116: 713–720.
194 Sunderkotter C, Steinbrink K, Goebeler M, Bhardwaj R, Sorg C.
Macrophages and angiogenesis. J Leukoc Biol 1994: 55: 410–
422.
195 Ruszczak Z, Detmar M, Imcke E, Orfanos C E. Effects of rIFN alpha,
beta and gamma on the morphology, proliferation and cell surface
antigen expression of human dermal microvascular endothelial cells
in vitro. J Invest Dermatol 1990: 95: 693–699.
196 Friedrich M, Bock D, Philipp S et al. Pan-selectin antagonism
improves psoriasis manifestation in mice and man. Arch Dermatol
Res 2006: 297: 345–351.
197 Zollner T M, Asadullah K, Schon M P. Targeting leukocyte traffick-
ing to inflamed skin: still an attractive therapeutic approach? Exp
Dermatol 2007: 16: 1–12.
198 Philipp S, Wolk K, Kreutzer S et al. The evaluation of psoriasis ther-
apy with biologics leads to a revision of the current view of the
pathogenesis of this disorder. Expert Opin Ther Targets 2006: 10:
817–831.
Sabat et al.
ª 2007 The Authors
798 Journal compilation ª 2007 Blackwell Munksgaard, Experimental Dermatology, 16, 779–798