diversity of rhizobia nodulating wild shrubs of sicily and some neighbouring islands
TRANSCRIPT

Arch Microbiol (2008) 190:461–470
DOI 10.1007/s00203-008-0394-2ORIGINAL PAPER
Diversity of rhizobia nodulating wild shrubs of Sicily and some neighbouring islands
Massimiliano Cardinale · Angela Lanza · Maria Laura Bonnì · Salvatore Marsala · Anna Maria Puglia · Paola Quatrini
Received: 25 September 2007 / Revised: 19 May 2008 / Accepted: 26 May 2008 / Published online: 20 June 2008© Springer-Verlag 2008
Abstract Legume shrubs have great potential for rehabil-itation of semi-arid degraded soils in Mediterranean eco-systems as they establish mutualistic symbiosis with N-Wxing rhizobia. Eighty-eight symbiotic rhizobia were iso-lated from seven wild legume shrubs native of Sicily(Southern Italy) and grouped in operational taxonomicunits (OTU) by analysis of the ribosomal internal tran-scribed spacer (ITS) polymorphism. Partial sequencing of16S rRNA gene of representative isolates of each OTUrevealed that most Genisteae symbionts are related toBradyrhizobium canariense, B. japonicum and B. elkanii.Teline monspessulana was the only Genistea nodulated byMesorhizobium strains, and Anagyris foetida (Thermopsi-deae) was promiscuosly nodulated by Rhizobium, Meso-rhizobium, Agrobacterium and Bradyrhizobium strains.Analysis of the nodulation gene nodA assigned most Medi-terranean Genisteae bradyrhizobia to clade II but also toclades IV, I and III, which included, so far, sequences of(sub)tropical and Australian isolates. The high diversity andlow host speciWcity observed in most wild legumes isolatessuggest that preferential associations may establish in theWeld depending on diVerences in the beneWts conferred to
the host and on competition ability. Once identiWed, thesebeneWcial symbiosis can be exploited for rehabilitation ofarid, low productive and human-impacted soils of the Med-iterranean countries.
Keywords Rhizobium · Bradyrhizobium · 16SrDNA · Symbiotic genes · Mediterranean wild legumes · Genisteae · Thermopsideae · Soil rehabilitation · Nodule occupancy
Introduction
Global warming will lead the average temperature toincrease between 1.8 and 4°C until the 2099 (FourthAssessment Report of the Intergovernmental Panel onClimate Change-IPCC, Paris, February 2007, http://www.ipcc.ch//SPM2feb07.pdf). In the Mediterraneanregion the hot extremes are estimated to increase by 200–500% respect to the present in the coming decades(DiVenbaugh et al. 2007), strongly increasing the risk ofdesertiWcation.
Rehabilitation of semi-arid degraded soils in Mediterra-nean ecosystems with autochthonous shrub species is a rec-ommended practice (Requena et al. 2001). MostMediterranean wild species are mycorrhizal (Maremmaniet al. 2003), and legume shrubs have the additional beneWtto establish N-Wxing symbiosis with rhizobial soil bacteriathus determining a nitrogen input into the ecosystem (Cara-vaca et al. 2003).
The legume-rhizobia symbiosis shows, generally, a highdegree of host speciWcity based on the exchange of chemi-cal signals between the partners. ModiWed lipo-chitooligo-saccharides, known as Nod factors, act as speciWcmorphogenic signal molecules to induce root hair deforma-tion, division of root cortical cells and nodule formation.
Communicated by Ursula Priefer.
M. Cardinale · A. Lanza · M. L. Bonnì · S. Marsala · A. M. Puglia · P. Quatrini (&)Dipartimento di Biologia Cellulare e dello Sviluppo, University of Palermo, EdiWcio 16, Viale delle Scienze, 90128 Palermo, Italye-mail: [email protected]
Present Address:M. CardinaleInstitute for Environmental Biotechnology, Graz University of Technology, Petersgasse 12, 8010 Graz, Austria
123

462 Arch Microbiol (2008) 190:461–470
The lipo-chitooligosaccharide core is synthesized under thecontrol of the common nodABC genes with nodA encodingan acyltransferase responsible for the attachment of a fattyacid group to the oligosaccharide backbone. NodA varies inits speciWcity for diVerent fatty acid substrates, thus con-tributing to the host range speciWcity (Debellè et al. 2001).Other speciWc genes (nod, noe, nol) introduce other speciWcdecorations in the Nod factor (Moulin et al. 2004).
Inoculation with nitrogen-Wxing rhizobia and indigenousarbuscular mycorrhizal fungi enhances the establishment oflegume shrubs and increases soil fertility and quality ofdesertiWed ecosystems (Requena et al. 2001) and anthropo-genic soils (highly human-inXuenced soils as those derivedfrom “earth-moving” operations and those used as cover-ings of terminal phase municipal landWlls) (Quatrini et al.2003; Cardinale et al., in preparation). In semi-arid cli-mates, legume shrubs-rhizobia symbiosis should play thesame role as tree legumes-rhizobia in sub-tropical aridregions (Zahran 1999).
Despite the high potential for environmental application,little is known about rhizobia from wild woody legumesoriginating from the Mediterranean countries. Leguminousshrubs belonging to the genera Spartium, Cytisus, Genistaand Retama (Genisteae) were found nodulated in southernItaly and in western Spain by slow growing bradyrhizobia(Requena et al. 1997; Quatrini et al. 2002; Rodríguez-Eche-verría et al. 2003; Kalita et al. 2006). Symbionts fromperennial wild Genisteae and Loteae in Tunisia were,instead, Rhizobium and Sinorhizobium (Zakhia et al. 2004).Bradyrhizobium strains isolated in Europe from Genisteae(mainly Lupinus species), were recently found to form adistinct lineage, designated clade II, on nodulation genetrees (Stdpkowski et al. 2007). This clade is supposed tohave diversiWed in the Mediterranean region.
The aim of this work was to evaluate the diversity of rhi-zobia isolated from native Mediterranean wild legumeshrubs in Sicily and surrounding islands. We report of 88novel rhizobial isolates from Wve widespread and oneendemic species belonging to the Genisteae and one speciesbelonging to the Thermopsideae. The rhizobia were charac-terized by analysing the polymorphisms of the ribosomalinternal transcribed spacer (ITS) and by 16S gene sequenc-ing, as markers of the genomic background; nodA and otherhost speciWc nodulation genes were explored as determi-nants of host-range speciWcity.
Materials and methods
Isolation of rhizobia
Rhizobia were isolated from seven species of wild Med-iterranean legumes (Anagyris foetida, Calicotome infesta,
Calicotome spinosa, Cytisus aeolicus, Genista aspalathoides,Spartium junceum, Teline monspessulana), either directlyfrom nodules of Weld grown plants or indirectly, from thenodules of plants seeded on soils sampled in diVerent areasof Sicily and surrounding islands. For the indirect isolation,seeds of the seven legume species were collected from wildplants, scariWed with sulphuric acid, rinsed in sterile waterand seeded. Root nodules were dissected from roots, rinsedthoroughly in water and 0.001% (v/v) Tween 80, surface-sterilized by immersion in 5% (v/v) sodium hypochlorite(3% active hypochlorite) for 30 min, and rinsed in sterilewater. Nodules were crushed on sterile plates, and streakedonto yeast-extract mannitol agar (YMA) plates containing0.0025% (w/v) Congo red. Plates were incubated at 28°Cfor 5–15 days. Individual colonies were picked and re-streaked on fresh plates. All of the strains were maintainedin YM broth containing 20% glycerol at ¡80°C.
Plant reinoculation tests
All the isolates from C. aeolicus and S. junceum were re-inoculated on seeds of the same species and some selectedstrains were used for cross inoculation experiments on S.junceum, C. aeolicus and Lupinus luteus as described inQuatrini et al. (2002) and Rodríguez-Echeverría et al.(2003).
Nodule occupancy analysis
Three Bradyrhizobium strains SjCL30, Sj9 and SjCb5 werechosen to analyse competition for nodule occupancy on S.junceum. ScariWed seeds were seeded in plastic pots onsterilised sandy soil and inoculated with the three strains,separately and in mixture, in the presence and in theabsence of the AMF Glomus intraradices (PremiertechBiotechnologies, Canada; 50 spores plant¡1). Each treat-ment consisted of 40 plants. After 5 months growth in thegreenhouse, three plants for each treatment were analysedfor the symbionts colonisation and nodule occupancy. Nod-ule occupancy by the three strains was identiWed based ontheir distinct PCR-RFLP proWles of the 16S–23S internallytranscribed spacer (ITS) with the restriction enzyme TaqI(Invitrogen). Plant height and basal stem diameter weremeasured on the other plants after 9 months growth.
DNA extraction and ampliWcation
DNA was extracted from 3-ml YMA cultures either usingthe lysozime-proteinase K-SDS protocol partially modiWed(Laguerre et al. 1992) or by direct lyses according to theprotocol of Muresu et al. (2005). All primers used for DNAampliWcation are listed in Table 1. The 16S–23S ribosomalspacers (Internal Transcribed Spacers, ITS) were ampliWed
123

Arch Microbiol (2008) 190:461–470 463
by PCR using ITSF/ITSReub primer set (Cardinale et al.2004) in a 30-�l volume reaction containing 1 �l of chro-mosomal DNA, 0.3 �M of each primer, 0.2 mM of dNTPsand 1.5 units of recombinant Taq DNA Polymerase (Invit-rogen, Life Technologies). The PCR was carried out underthe following conditions: 94°C for 5 min; 30 cycles of1 min at 94°C, 1 min at 55°C and 2 min at 72°C; 72°C for7 min. PCR products were compared on ethidium bromide-stained 1.5% agarose gel in order to identify diVerent ITSproWles. Isolates with identical ITS proWles were consid-ered belonging to the same operational taxonomic unit(OTU). The 16S rDNA gene of at least one isolate repre-sentative of each OTU was ampliWed using the fD1/rD1primer set (Weisburg et al. 1991) with the protocoldescribed above.
Several primer sets were used to amplify the nodA genesusing the protocols described by the authors (Haukka et al.1998; Sy et al. 2001; Moulin et al. 2001). A new primer set(NodAf2brad/NodAr2brad) was designed for the ampliWca-tion of nodA genes from some Bradyrhizobium isolates andused in a touch down PCR protocol with the following pro-gramme: an initial denaturation step at 94°C for 5 min; 16cycles with 1 min at 94°C, 1 min at annealing temperaturesdecreasing from 60 to 45°C (with 1°C decremental stepfrom cycle 2 to 16), 1 min at 72°C; 19 cycles with 1 min at95°C; 1 min at 45°C and 1 min at 72°C; a Wnal 7 min exten-sion at 72°C.
The genes nodZ and noeI were ampliWed using theprimer sets described in Table 1 and the PCR protocoldescribed above for ITS ampliWcation with annealing tem-
peratures of 52°C. For nolL ampliWcation the nolL primerset was used in a touchdown-PCR procedure with the fol-lowing programme: an initial denaturation step at 94°C for5 min; 9 cycles with 1 min at 94°C, 1 min at annealing tem-peratures decreasing from 48 to 40°C (with 1°C decremen-tal step from cycle 2 to 9), 1 min at 72°C; 27 cycles with1 min at 95°C; 1 min at 40°C and 1 min at 72°C; a Wnal7 min extension at 72°C.
All PCR products were visualized on ethidium bromide-stained 1.5% agarose gel. Bands of expected size wereexcised and puriWed by using GFX™ PCR DNA and GelBand PuriWcation Kit (Amersham biosciences, EnglandUK). PCR products were commercially sequenced. Theelectropherograms were checked with the programs EDIT-VIEW (http://www.appliedbiosystems.com/support/software/)and FINCHTV (http://www.geospiza.com/finchtv/index.htm). Unambiguous sequences were used in BLASTnsearches (Altschul et al. 1997, http://www.ncbi.nlm.nih.gov/blast/).
The sequences produced in this work are available inGenbank/EMBL/DDBJ database under the accession num-bers AM748939 to AM748974 (16S rRNA genes),AM748902 to AM748924 (nodA genes), AM748825 toAM748828 (nolL genes).
Phylogenetic analysis
A 16S rDNA phylogenetic tree was constructed by using16S rDNA partial sequences from at least one representa-tive isolate for each OTU and sequences of rhizobial reference
Table 1 Primers for DNA ampliWcation and respective target genes used in this work
Primer Sequence 5�–3� Target gene Target organism(s) Reference
ITSF GTCGTAACAAGGTAGCCGTA 16S rRNA Most Eubacteria Cardinale et al. (2004)
ITSReub GCCAAGGCATCCACC 23S rRNA
fD1 AGAGTTTGATCCTGGCTCAG 16S rRNA Most Eubacteria Weisburg et al. (1991)
rD1 AAGGAGGTGATCCAGCC
NodA1 TGCRGTGGAARNTRNNCTGGGAAA nodA Rhizobium-Sinorhizobium Haukka et al. (1998)
NodA2 GGNCCGTCRTCRAAWGTCARGTA
nodA1f TGCRGTGGAARNTRBVYTGGG nodA Most rhizobia Sy et al. (2001)
nodAb1r GGNCCGTCRTCRAASGTCARGTA
Moul-nodAFor TCACARCTCKGGCCCGTTCCG nodA Most rhizobia and Burkholderia Moulin et al. (2001)
Moul-nodARev TGGGCSGGNGCNAGRCCBGA
NodAf2brad CRGTGGAGSSTDYGYTGGGA nodA Bradyrhizobium This work
NodAr2brad TCAHARCTCHGGHCCRTTCC
nodZfor YGGYGAYTGYCTSTGGTC nodZ Most rhizobia This work
nodZrev CGCTNGTBGGVGGAAAVC
noeIfor TGAYCCCGTSATYCTGG noeI Rhizobium, Brabyrhizobium-Sinorhizobium
This work
noeIrev CRTCACCYTCCACTSCG
nolLfor ATGGCRATAAGYGGATA nolL Rhizobium, Mesorhizobium-Bradyrhizobium
This work
nolLrev RCCCARMRARCATCAGAA
123

464 Arch Microbiol (2008) 190:461–470
strains. A 615-nucleotides multialignment was obtainedusing CLUSTALX (Thompson et al. 1994; http://bips.u-strasbg.fr/fr/Documentation/ClustalX/). The PHYLIP pack-age (Felsenstein, 2004; http://evolution.genetics.washing-ton.edu/phylip.html) was used to infer all phylogenetictrees. A distance matrix was calculated with DNADISTsoftware and the Neighbour-Joining phylogenetic trees(Saitou and Nei 1987) were constructed with NEIGHBORsoftware. Maximum likelihood trees were constructed withDNAML software (Felsenstein and Churchill 1996). AllnodA sequences obtained from at least one representativeisolate for each OTU were aligned with both referencestrain sequences and all best aligned sequences retrieved byBLAST alignment. A 291-nucleotides multialignment wasused for inferring the trees as described above for the 16SrDNA trees. All trees were statistically tested by perform-ing 1,000 bootstrap resamplings of the data and visualizedwith the TREEVIEW program (Page 1996; http://taxonomy.zoology.gla.ac.uk/rod/treeview.html).
Results
Isolation and identiWcation of rhizobia from Mediterranean wild legumes
We analysed Mediterranean legume shrubs in diVerentareas of Sicily and some of the surrounding islands (theAeolians Archipelagos, and Pantelleria Island) (Table 2).All investigated plants are native woody shrubs that belongto the Papilionoideae in the tribe Genisteae except Anagyrisfoetida, belonging to the Thermopsideae. C. aeolycus is anendemic species of the Aeolian Archipelagos (Conte et al.1998) seriously threatened and among the priority speciesfollowing the E.U. Directive 92/43. EVective nodulation inthe Weld was observed in all species including Anagyrisfoetida, Cytisus aeolicus and Genista aspalatoides forwhich it is reported here for the Wrst time. By using directisolation and trap host cultures we obtained 88 novel rhizo-bial isolates.
The new isolates from Anagyris foetida, Cytisus aeoli-cus, Calicotome infesta, Calicotome spinosa, Genistaaspalathoides, Spartium junceum and Teline monspessul-ana were grouped by PCR-ITS in 21 diVerent operationaltaxonomic units (OTU) (Table 2, letters A–U). Some OTUswere found in common among the diVerent plant speciessuggesting a high degree of promiscuity. 16S rRNA gene ofone or more representatives for each OTU, was sequencedtogether with representative strains of a collection previ-ously isolated from Spartium junceum (Table 2, Quatriniet al. 2002).
BLASTn search results are shown in Table 2. Twenty-Wve partial sequences of the representative strains for each
OTU were chosen to construct a phylogenetic tree, includ-ing also reference sequences (Fig. 1). The 615 nucleotidesincluded positions 121–729 of the 16S gene of B. japoni-cum USDA 110 (complete genome accession number:BA000040, region:1582226…1529715).
One group of isolates from the Genisteae C. aeolycus, G.aspalathoides, C. infesta, C. spinosa and S. junceum, isgrouped with B. canariense and B. japonicum strains in anot well resolved branch. Other isolates form an indepen-dent cluster, closer to B. elkanii, that is well supported byboth neighbour joining and likelihood bootstrap values(group B, Quatrini et al. 2002). Most isolates from A. foet-ida and T. monspessulana cluster with Mesorhizobium spe-cies and some of them form a clearly separate group withinthe M. ciceri lineage. One isolate from A. foetida and onefrom C. spinosa cluster with the fast growing Rhizobiumleguminosarum.
Phylogenetic analysis of nodulation genes
We obtained a nodA PCR product of the expected size fromall Bradyrhizobium strains analysed with at least one cou-ple of primers described in Table 1. No ampliWcation prod-uct was obtained from the Rhizobium isolates of A. foetidaand C. spinosa.
Our isolates were grouped in six clusters (Fig. 2) withthe Wrst four including Bradyrhizobium strains and twoother clusters formed by Mesorhizobium strains and includ-ing A. foetida and T. monspessulana isolates.
In the genus Bradyrhizobium the gene nodA is monophy-letic, despite the high sequence diversity (Moulin et al.2004), and forms seven clades (I-VII). The largest Brady-rhizobium cluster formed by our Genisteae isolates corre-sponds to clade II that comprises strains typical forbradyrhizobia from genistoid plants (Genisteae and Loteae)(Stdpkowski et al. 2003, 2005, 2007). PCR ampliWcationrevealed the presence of nodZ in representative strains ofthis Bradyrhizobium lineage while nolL was found only insome of them (Fig. 2). Nucleotide sequences of nolLshowed 98–73% identity with isolates from Lupinus angus-tifolius and Ornithopus compressus, respectively (Stdpkow-sky et al. 2005). None of the clade II isolates could beampliWed in the noeI gene (Fig. 2).
A second large nodA cluster is closely related to strainWSM1735, isolated from Rynchosia minima (Phaseoleae)in Australia and included in the nodA clade IV (Stdpkow-sky et al. 2005, 2007). Within this cluster, isolates from C.spinosa form a well supported group. Two other isolatesare included in clades I and III, with strains isolated also inAustralia (Stdpkowsky et al. 2005, 2007). We could notdetect the genes responsible for fucosylation and fucosemodiWcations (nodZ, nolL, noeI) in CaVcas27 and this sup-ports the aYliation of this strain to clade I, as all clade I
123

Arch Microbiol (2008) 190:461–470 465
Table 2 Rhizobia isolated from seven species of wild Mediterranean legumes and their phylogenetic aYliation
Isolatesa Host speciesb
Geographical originc
OTUd Aligned nte
Most closely related sequencef, accession number (% of identity)
Source
SjCL 1, 2, 3, 4, 5, 6, 7, 8, 9, 11, 12, 13, 14, 15, 15bis, 16, 17, 18, 19, 20, 21, 22, 23, 24, 24b, 26, 27, 30
SJ CL A 891 Bradyrhizobium japonicum strain BQ3, DQ133343 (99%)
This work
TmCAD 3, 4, 5, 7, 9 TM Pantelleria, TP B 919 Mesorhizobium amorphae strain ICMP 11721, AY491077 (100%)
This work
TmCAD 6, 8, 14 TM Pantelleria, TP C 864 Mesorhizobium sp. 98_RREM2003, AY238500 (100%)
This work
TmGib 4, 5, 6 TM Pantelleria, TP D 918 Mesorhizobium ciceri strain C-2/2, AY206686 (100%)
This work
AfPAR 4 AF PA E 866 Agrobacterium tumefaciens strain 204, EF217305 (100%)
This work
GasA GA Pantelleria, TP F 819 Bradyrhizobium japonicum isolate HA1, AF530468 (99%)
This work
AfLs 1, 2, 4 AF CL G 820 Rhizobium sp. H-4, AF279889 (99%) This work
AfLs 3 AF CL H 855 Mesorhizobium sp. CCNWGX022, DQ319189 (100%)
This work
AfPa 1 AF PA H 720 Mesorhizobium sp. STM 398, AY500256 (99%)
This work
CiE 2 CI CT I 868 Bradyrhizobium sp. MSDJ 5725, AF363148 (99%)
This work
GaPa 1 GA PA I This work
Af3B 1 AF PA J 631 Rhizobium sp. H-4, AF279889 (99%) This work
Af3B 2, 3, 4, 6, 7, 8, 9, 10, 11, 13, 14, 17, 19
AF PA K 831 Rhizobium sp. H-4, AF279889 (99%) This work
Cs2B 2 CS PA K This work
Af3B 5 AF PA L 760 Rhizobium sp. H-4, AF279889 (99%) This work
Af3B 15, 18 AF PA M This work
Cs2B 5 CS PA M 837 R. leguminosarum strain ICMP 14642, AY491062 (99%)
This work
Af3B 20 AF PA N 765 Rhizobium sp. H-4, AF279889 (99%) This work
Cs1B 1 CS PA O 438 Bradyrhizobium sp. Spain-6, AF461196 (97%)
This work
CaStro 2.2, 2.3, 5.1, 5.2, 5.3 CA Stromboli, ME O 816 Bradyrhizobium canariense, AY577427 (100%)
This work
CaVcas 4.1 CA Vulcano, ME O 499 Bacterium Ellin5232, AY234583 (100%) This work
Cs1B 2, 4, 5 CS PA P 863 Bradyrhizobium elkanii strain CCBAU 61184, EF061103 (99%)
This work
Cs1B 3, Cs2B 1 CS PA Q 708 Bradyrhizobium elkanii strain CCBAU 61184, EF061103 (99%)
This work
Af1B 1, 2 AF PA Q 814 Bradyrhizobium elkanii strain CCBAU 61184, EF061103 (99%)
This work
Cs2B 3, 4 CS PA R 863 Bradyrhizobium elkanii strain CCBAU 61184, EF061103 (99%)
This work
Af 3.6 AF PA S 798 Mesorhizobium sp. CCNWGX022, DQ319189 (100%)
This work
Af1B 3 AF PA T 829 Rhizobium leguminosarum strain PB161, EF525218 (99%)
This work
CaVcas 27, CaVcas 99 CA Vulcano, ME U 949 Bradyrhizobium japonicum strain JZ2, DQ133342 (100%)
This work
SjGb 2 SJ PA 2 879 Bacterium Ellin5232, AY234583 (100%) Quatrini et al. (2002)
123

466 Arch Microbiol (2008) 190:461–470
strains analysed so far lack nodZ (Stdpkowsy et al. 2007);moreover this strain was unable to form nodules on S. jun-ceum and L. luteus (Table 3).
Coherently with their phylogenetic aYliation most Mes-orhizobium isolates from Anagyris foetida formed a wellsupported group related to Mesorhizobium sp. N33 nodAsequence, isolated from Oxytropis arctobia that also nodu-lates Astragalus alpinus and Onobrychis viciifolia (Poinsotet al. 2001). All the nodA sequences of Teline monspessul-ana isolates clustered together, and were related to Meso-rhizobium loti.
EVectiveness and nodule occupancy
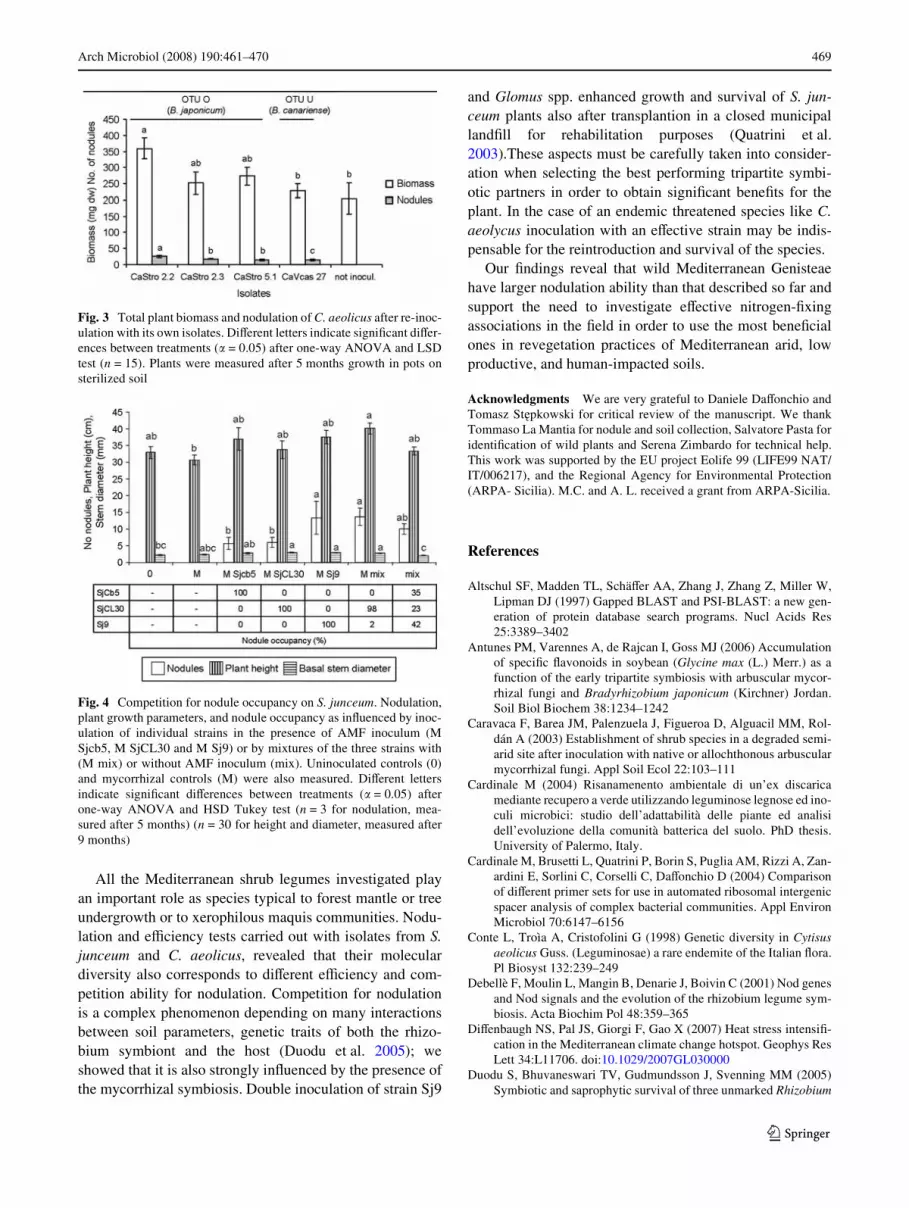
The isolates showed also diVerent infectivity and eVective-ness abilities. All the isolates from C. aeolicus, for exam-ple, could form nodules on their host but only strainCaStro2.2 was able to increase signiWcantly plant biomasscompared to the uninoculated controls (Fig. 3).
When three diVerent isolates from S. junceum were rein-oculated on the host species alone, or in mixture, in thepresence of the AMF G. intraradices they showed diVerenteYciency and also diVerent competition ability for noduleoccupancy (Fig. 4). The AMF was inoculated to simulatenatural soil conditions. Strain Sj9 was the most infectivewhen inoculated alone in the presence of the AMF (MSj9,13,3 nodules plant¡1) and also the most competitive fornodule occupancy when the three bacteria were inoculatedwithout the AMF (42.9% nodule occupancy), but SjCL30showed the highest competition for nodule occupancy
(99%) when co-inoculated in the presence of the AMF(Mmix). Both strains increased signiWcantly plant stemdiameter with the AMF compared to strain SjCb5(MSjCb5) and controls (M: only mycorrhizal inoculum;mix: only bacterial inoculum 0: not inoculated) (Fig. 4).
It is well known that the tripartite symbiosis enhancesnodulation and nodule activity and that symbiotic pathwaysactivated by both the bacterial and the fungal symbiontsshare common steps; however we found no reports aboutthe inXuence of AMF on strain competitivity. Flavonoidsrather than phosphorus appear to play a role as early signalsfor both rhizobia and AMF, and recent reports suggest thatspeciWc Xavonoids are produced and released into the rhi-zosphere as a function of the colonization process (Antuneset al. 2006). DiVerent rhizobial strains can respond diVer-ently to Xavonoids accumulation as inXuenced by the pres-ence of AMF; however further experiments are needed toconWrm this hypothesis.
Discussion
Most of the rhizobia isolated from Sicilian native andendemic shrub legumes are slow-growing strains, pheno-typically resembling Bradyrhizobium spp. All Genisteaeresulted nodulated by a highly diverse group of isolatesclose to B. japonicum and B. canariense with the onlyexception of Teline monspessulana nodulated by Mesorhiz-obium strains. As Teline is part of the Genisteae (Pardoet al. 2004), it might be assumed a geographic inXuence on
Table 2 continued
a The isolates underlined were sequenced for 16S rRNA geneb Two letter codes stands for the following plant hosts: AF Anagyris foetida; CA Cytisus aeolicus; CI Calicotome infesta; CS Calicotome spinosa;GA Genista aspalathoides; SJ Spartium junceum; TM Teline monspessulanac Two letter codes stand for the following cities in Sicily, Italy: CL Caltanissetta; CT Catania; ME Messina; PA Palermo; TP Trapani. Vulcano andStromboli belong to the Aeolian Archipelagos. Pantelleria is an island in the Strait of Sicilyd Strains were assigned to diVerent OTUs (indicated by letters) basing on polymorphisms in the ITS-PCR proWles. Previously isolated rhizobiawere grouped according to their 16S-ITS PCR-RFLP proWles (indicated by numbers, Quatrini et al. 2002)e Number of 16S rRNA gene nucleotides used in the alignement with sequences in the databasesf Highest sequence match with the closest 16S rRNA gene sequences of bacterial isolates available in public databases
Isolatesa Host speciesb
Geographical originc
OTUd Aligned nte
Most closely related sequencef, accession number (% of identity)
Source
Sj 1, 1EM, 4EM, 4, 8, 33a, 35b, 39b
SJ CT 3 676 Bradyrhizobium japonicum strain HMJ-03, DQ517957 (100%)
Quatrini et al. (2002)
SjCb 5, 7 SJ ME 3 784 Bradyrhizobium japonicum strain HMJ-03, DQ517957 (100%)
Quatrini et al. (2002)
Sj 26a, 26b, 27b, 27d SJ ME 6 532 Bradyrhizobium sp. Ai1a-2, AF514703 (99%) Quatrini et al. (2002)
Sj 14c, 15c SJ ME 7 689 Bradyrhizobium sp. lebi-3, AY490122 (99%) Quatrini et al. (2002)
Sj 5, 26e, 27a, 39a, 39d SJ ME 8 889 Bradyrhizobium sp. Ai1a-2, AF514703 (99%) Quatrini et al. (2002)
Sj 9 SJ PA 9 1,439 Bradyrhizobium sp. Cf1, DQ354622 (99%) Quatrini et al. (2002)
SjSa 1 SJ PA 11 551 Bradyrhizobiaceae bacterium NR111, DQ520809 (99%)
Quatrini et al. (2002)
123

Arch Microbiol (2008) 190:461–470 467
this diVerent symbiotic behaviour due the fact that all theisolates analysed were collected from Pantelleria island;isolates collected from other sites should be tested to con-Wrm this hypothesis.
In some recent studies Genistoid plants (Papilionoideae:Genisteae and Loteae) were found nodulated by four Brady-rhizobium lineages (Jarabo-Lorenzo et al. 2003; Vinuesaand Silva 2004; Vinuesa et al. 2005; Kalita et al. 2006); twoof these lineages were proposed as a novel species B. canar-iense and its sister species B. japonicum biovar genistearum(Vinuesa et al. 2005). One group of Genisteae isolates wasintermingled with these novel proposed species in the 16Sphylogenetic tree although the diVerent clades are not welldelineated. Interestingly, a second group of isolates clusterson a separate branch closer to B. elkanii suggesting it maybelong to a new species or biovar. To our knowledge,this is the Wrst report about isolation of strains belonging toB. elkanii in Europe. Further investigations, based on the
phylogeny of other houskeeping genes (i.e. recA, glnII,atpD) are needed to conWrm these results.
Anagyris foetida, the only Thermopsidea analysed, waspromiscuously nodulated by Rhizobium, Mesorhizobium,Agrobacterium and Bradyrhizobium strains. Promiscuoussymbioses are considered widespread and more ancestralthan restricted host ranges that represent specialization andare limited to speciWc niches (Perret et al. 2000). The highdegree of symbiotic promiscuity of A. foetida does not cor-respond to enhanced plant beneWts as we detected low or nobeneWts following inoculation with a mixture of strainsherein decribed (Cardinale 2004).
According to nodA phylogeny most of our Genisteae iso-lates belong to clade II supporting the common evolution-ary history of rhizobia from Genisteae legumes and thehypothesis that this clade diversiWed in the Mediterraneanregion (Stdpkowski et al. 2007). The group is also charac-terized by the presence of nodZ and the absence of noeI
Fig. 1 Neighbor joining tree showing phylogenetic relation-ships between 615 bp-16S se-quences of rhizobial isolates and type strains. Numbers on nodes are, respectively, Neighbor Join-ing and Maximum Likelihood bootstrap support values (%) of 1,000 re-samplings. OTU opera-tional taxonomic unit
123

468 Arch Microbiol (2008) 190:461–470
involved in methylation of fucose, which has never beendetected in clade II (Stdpkowski et al. 2007). The fact thatnolL, involved in acetylation of Nod factor fucose residues,was present in only some of our Genisteae bradyrhizobiabelonging to nodA clade II and absent in some other leadsto the hypothesis that fucose acetylation is not necessary forNod factor recognition by Genisteae or that its sequence ismore divergent than those of other nodulation genes.
A consistent group of Sicilian isolates from Genisteaehowever, clusters also with clade IV and two strains withclades I and III. Most of clade IV and III strains belong tothe branch closer to B. elkanii, in the 16S tree. Clades I, IIIand IV are considered of tropical and Australian origin(Stdpkowski et al. 2005, 2007) and their presence in Sicilycould be explained by the introduction of Australian nativeAcacia spp. into the Mediterranean region. This hypothesisseems to be corroborated, at least for clade I, by the highsequence similarity (99%) of strain CaVcas27 16S genewith that of rhizobia recently isolated from the root nodulesof A. mollissima and A. melanoxylon plants in Sicily (Qua-trini and Cardinale, unpublished results). However this isthe Wrst study of a large collection of Genisteae strains iso-lated from a broad range of wild shrubs in the Mediterra-nean basin—the Genisteae center of diversity. Theabundance of European strains in clade IV suggests torevise the hypothesis of the Australian origin of this clade,that was originally based on phylogenetic studies of Lupi-nus isolates and a few European Genisteae symbionts.
Fig. 2 Maximum likelihood tree showing the phylogenetic relationships between 293 bp-nodA sequences of rhizobial iso-lates and type strains. Numbers on nodes are, respectively, Neighbor Joining and Maximum Likelihood bootstrap support values (%) of 1,000 re-sam-plings. PCR products from nodZ, nolL and noeI (+) were obtained in this work and from Stdpkowski et al. 2003a, Moulin et al. 2004b, Stepkowsky et al. 2005c. (¡) indicates lack of ampliWcation. OTU operational taxonomic unit
Table 3 Cross inoculation tests of some C. aeolycus and S. junceumisolates
Bacterial isolate
nodA clade
Nod+ on
S. junceum C. aeolicus L. luteus
Sj15c II + + +
SjGb2 II + + +
CaStro 2.2 II + + +
SjCb5 II + + ¡CaVcas 27 I ¡ + ¡
123
Arch Microbiol (2008) 190:461–470 469
All the Mediterranean shrub legumes investigated playan important role as species typical to forest mantle or treeundergrowth or to xerophilous maquis communities. Nodu-lation and eYciency tests carried out with isolates from S.junceum and C. aeolicus, revealed that their moleculardiversity also corresponds to diVerent eYciency and com-petition ability for nodulation. Competition for nodulationis a complex phenomenon depending on many interactionsbetween soil parameters, genetic traits of both the rhizo-bium symbiont and the host (Duodu et al. 2005); weshowed that it is also strongly inXuenced by the presence ofthe mycorrhizal symbiosis. Double inoculation of strain Sj9
and Glomus spp. enhanced growth and survival of S. jun-ceum plants also after transplantion in a closed municipallandWll for rehabilitation purposes (Quatrini et al.2003).These aspects must be carefully taken into consider-ation when selecting the best performing tripartite symbi-otic partners in order to obtain signiWcant beneWts for theplant. In the case of an endemic threatened species like C.aeolycus inoculation with an eVective strain may be indis-pensable for the reintroduction and survival of the species.
Our Wndings reveal that wild Mediterranean Genisteaehave larger nodulation ability than that described so far andsupport the need to investigate eVective nitrogen-Wxingassociations in the Weld in order to use the most beneWcialones in revegetation practices of Mediterranean arid, lowproductive, and human-impacted soils.
Acknowledgments We are very grateful to Daniele DaVonchio andTomasz Stdpkowski for critical review of the manuscript. We thankTommaso La Mantia for nodule and soil collection, Salvatore Pasta foridentiWcation of wild plants and Serena Zimbardo for technical help.This work was supported by the EU project Eolife 99 (LIFE99 NAT/IT/006217), and the Regional Agency for Environmental Protection(ARPA- Sicilia). M.C. and A. L. received a grant from ARPA-Sicilia.
References
Altschul SF, Madden TL, SchäVer AA, Zhang J, Zhang Z, Miller W,Lipman DJ (1997) Gapped BLAST and PSI-BLAST: a new gen-eration of protein database search programs. Nucl Acids Res25:3389–3402
Antunes PM, Varennes A, de Rajcan I, Goss MJ (2006) Accumulationof speciWc Xavonoids in soybean (Glycine max (L.) Merr.) as afunction of the early tripartite symbiosis with arbuscular mycor-rhizal fungi and Bradyrhizobium japonicum (Kirchner) Jordan.Soil Biol Biochem 38:1234–1242
Caravaca F, Barea JM, Palenzuela J, Figueroa D, Alguacil MM, Rol-dán A (2003) Establishment of shrub species in a degraded semi-arid site after inoculation with native or allochthonous arbuscularmycorrhizal fungi. Appl Soil Ecol 22:103–111
Cardinale M (2004) Risanamenento ambientale di un’ex discaricamediante recupero a verde utilizzando leguminose legnose ed ino-culi microbici: studio dell’adattabilità delle piante ed analisidell’evoluzione della comunità batterica del suolo. PhD thesis.University of Palermo, Italy.
Cardinale M, Brusetti L, Quatrini P, Borin S, Puglia AM, Rizzi A, Zan-ardini E, Sorlini C, Corselli C, DaVonchio D (2004) Comparisonof diVerent primer sets for use in automated ribosomal intergenicspacer analysis of complex bacterial communities. Appl EnvironMicrobiol 70:6147–6156
Conte L, Troìa A, Cristofolini G (1998) Genetic diversity in Cytisusaeolicus Guss. (Leguminosae) a rare endemite of the Italian Xora.Pl Biosyst 132:239–249
Debellè F, Moulin L, Mangin B, Denarie J, Boivin C (2001) Nod genesand Nod signals and the evolution of the rhizobium legume sym-biosis. Acta Biochim Pol 48:359–365
DiVenbaugh NS, Pal JS, Giorgi F, Gao X (2007) Heat stress intensiW-cation in the Mediterranean climate change hotspot. Geophys ResLett 34:L11706. doi:10.1029/2007GL030000
Duodu S, Bhuvaneswari TV, Gudmundsson J, Svenning MM (2005)Symbiotic and saprophytic survival of three unmarked Rhizobium
Fig. 3 Total plant biomass and nodulation of C. aeolicus after re-inoc-ulation with its own isolates. DiVerent letters indicate signiWcant diVer-ences between treatments (� = 0.05) after one-way ANOVA and LSDtest (n = 15). Plants were measured after 5 months growth in pots onsterilized soil
Fig. 4 Competition for nodule occupancy on S. junceum. Nodulation,plant growth parameters, and nodule occupancy as inXuenced by inoc-ulation of individual strains in the presence of AMF inoculum (MSjcb5, M SjCL30 and M Sj9) or by mixtures of the three strains with(M mix) or without AMF inoculum (mix). Uninoculated controls (0)and mycorrhizal controls (M) were also measured. DiVerent lettersindicate signiWcant diVerences between treatments (� = 0.05) afterone-way ANOVA and HSD Tukey test (n = 3 for nodulation, mea-sured after 5 months) (n = 30 for height and diameter, measured after9 months)
123

470 Arch Microbiol (2008) 190:461–470
leguminosarum biovar trifolii strains introduced into the Weld.Environ Microbiol 7:1049–1058
Felsenstein J (2004) PHYLIP (Phylogeny Inference Package) version3.6 (Distributed by the author). Department of Genome Sciences,University of Washington, Seattle
Felsenstein J, Churchill GA (1996) A hidden Markov model approachto variation among sites in rate of evolution. Mol Biol Evol13:93–104
Haukka K, Lindstrom K, Young JPW (1998) Three phylogeneticgroups of nodA and nifH genes in Sinorhizobium and Mesorhizo-bium isolates from leguminous trees growing in Africa and LatinAmerica. Appl Environ Microbiol 64:419–426
Kalita M, Stdpkowski T, Lotocka B, Malek W (2006) Phylogeny ofnodulation genes and symbiotic properties of Genista tinctoriabradyrhizobia. Arch Microbiol 186:87–97
Jarabo-Lorenzo A, Perez-Galdona R, Donate-Correa J, Rivas R, Ve-lazquez E, Hernandez M, Temprano F, Martinez-Molina E, Ruiz-Argueso T, Leon-Barrios M (2003) Genetic diversity of bradyrhi-zobial populations from diverse geographic origins that nodulateLupinus spp. and Ornithopus spp. System Appl Microbiol26:611–623
Laguerre G, Mazurier SI, Amarger N (1992) Plasmid proWles and restric-tion fragment length polymorphism of Rhizobium leguminosarumbv. viciae in Weld populations. FEMS Microbiol Ecol 101:17–26
Maremmani A, Bedini S, Matonevic I, Tomei PE, Giovannetti M(2003) Type of mycorrhizal associations in two coastal nature re-serves of the Mediterranean basin. Mycorrhiza 13:33–40
Moulin L, Munive A, Dreyfus B, Boivin-Masson C (2001) Nodulationof legumes by members of the �-subclass of Proteobacteria. Na-ture 411:948–950
Moulin L, Bena G, Boivin-Masson C, Stdpkowski T (2004) Phyloge-netic analyses of symbiotic nodulation genes support vertical andlateral gene co-transfer within the Bradyrhizobium genus. MolPhylogenet Evol 30:720–732
Muresu R, Sulas L, Polone E, Squartini A (2005) PCR primers basedon diVerent portions of insertion elements can assist genetic relat-edness studies, strain Wngerprinting and species identiWcation inrhizobia. FEMS Microbiol Ecol 54:445–453
Page RD (1996) TreeView: an application to display phylogenetictrees on personal computers. Comput Appl Biosci 12:357–358
Perret X, Staehelin C, Broughton WJ (2000) Molecular basis of sym-biotic promiscuity. Microbiol Mol Biol Rev 64:180–201
Pardo C, Cubas P, Tahiri H (2004) Molecular phylogeny and system-atics of Genista (Leguminosae) and related genera based onnucleotide sequences of nrDNA (ITS region) and cpDNA (trnL-trnF intergenic spacer). Plant System Evol 244:93–119
Poinsot V, Belanger E, Laberge S, Yang GP, Antoun H, Cloutier J,Treilhou M, Denarié J, Prome JC, Debellé F (2001) Unusual me-thyl-branched alpha, beta-unsaturated acyl chain substitutions inthe Nod Factors of an arctic rhizobium, Mesorhizobium sp. strainN33 (Oxytropis arctobia). J Bacteriol 183:3721–3728
Quatrini P, Scaglione G, Cardinale M, Caradonna F, Puglia AM (2002)Bradyrhizobium sp. nodulating the Mediterranean shrub SpanishBroom (Spartium junceum L.). J Appl Microbiol 92:13–21
Quatrini P, Scaglione G, Incannella G, Badalucco L, Puglia AM, LaMantia T (2003) Microbial inoculants on woody legumes to recov-er a municipal landWll site. Water Air Soil Pollut Focus 3:189–199
Requena N, Jimenez I, Toro M, Barea JM (1997) Interactions betweenplant-growth-promoting rhizobacteria (PGPR), arbuscularmycorrhizal fungi and Rhizobium spp. in the rhizosphere of An-thyllis cytisoides, a model legume for revegetation in mediterra-nean semi-arid ecosystems. New Phytol 136:667–677
Requena N, Perez-Solis E, Azcón-Aguilar C, JeVries P, Barea JM(2001) Management of indigenous plant-microbe symbioses aidsrestoration of desertiWed ecosystems. Appl Environ Microbiol67:495–498
Rodríguez-Echeverría S, Pérez-Fernández MA, Vlaar S, Finnan T(2003) Analysis of the legume-rhizobia symbiosis in shrubs fromcentral western Spain. J Appl Microbiol 95:1367–1374
Saitou N, Nei M (1987) The neighbor-joining method: a new methodfor reconstructing phylogenetic trees. Mol Biol Evol 4:406–425
Stdpkowski T, Swiderska A, Miedzinska K, Czaplinska M, SwiderskiM, Biesiadka J, Legocki AB (2003) Low sequence similarity andgene content of symbiotic clusters of Bradyrhizobium sp. WM9(Lupinus) indicate early divergence of “lupin” lineage in the ge-nus Bradyrhizobium. Antonie Van Leeuwenhoek 84:115–124
Stdpkowski T, Moulin L, Krzyzanska A, McInnes A, Law IJ, Howie-son J (2005) European origin of Bradyrhizobium populationsinfecting lupins and serradella in soils of Western Australia andSouth Africa. Appl Environ Microbiol 71:7041–7052
Stdpkowski T, Hughes CE, Law IJ, Markiewicz L, Gurda D, ChlebickaA, Moulin L (2007) DiversiWcation of lupine Bradyrhizobiumstrains: evidence from nodulation gene trees. Appl EnvironMicrobiol 73:3254–3264
Sy A, Giraud E, Jourand P, Garcia N, Willems A, de Lajudie P, Prin Y,Neyra M, Gillis M, Boivin-Masson C, Dreyfus B (2001) Methy-lotrophic Methylobacterium bacteria nodulate and Wx nitrogen insymbiosis with legumes. J Bacteriol 183:214–220
Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W: improv-ing the sensitivity of progressive multiple sequence alignmentthrough sequence weighting, position-speciWc gap penalties andweight matrix choice. Nucl Acids Res 22:4673–4680
Vinuesa P, Silva C (2004) Species delineation and biogeography ofsymbiotic bacteria associated with cultivated and wild legumes.In: Werner D (ed) Biological Resources and Migration. Springer,Berlin, pp 43–155
Vinuesa P, Leon-Barrios M, Silva C, Willems A, Jarabo-Lorenzo A,Perez-Galdona R, Werner D, Martinez-Romero E (2005) Brady-rhizobium canariense sp. nov., an acid-tolerant endosymbiontthat nodulates endemic genistoid legumes (Papilionoideae: Gen-isteae) from the Canary Islands, along with Bradyrhizobium ja-ponicum bv. genistearum, Bradyrhizobium genospecies alpha andBradyrhizobium genospecies beta. Int J Syst Evol Microbiol55:569–575
Weisburg W, Barns SM, Pelletier DA, Lane DJ (1991) 16S ribosomalDNA ampliWcation for phylogenetic study. J Bacteriol 173:697–703
Zahran H (1999) Rhizobium-legume symbiosis and nitrogen Wxationunder severe conditions and in an arid climate. Microbiol MolBiol Rev 63:968–989
Zakhia F, Jeder H, Domergue O, Willems A, Cleyet-Marel JC, GillisM, Dreyfus De, Lajudie P (2004) Characterisation of wild legumenodulating bacteria (LNB) in the infra-arid zone of Tunisia. Sys-tem Appl Microbiol 7:380–395
123