effect of high salts concentrations on the growth of rhizobia and responses to added osmotica
TRANSCRIPT

Journal of Applied Microbiology 1999, 86, 889–898
Effect of high salts concentrations on the growth of rhizobiaand responses to added osmotica
H. Abdelmoumen 1, A. Filali-Maltouf 2, M. Neyra3, A. Belabed 4 andM. Missbah El Idrissi 1
1Laboratoire de Microbiologie Appliquee, Departement de Biologie, Faculte des Sciences, Universite Mohamed I,Oujda, 2Laboratoire de Microbiologie et de Biologie Moleculaire, Faculte des Sciences, Rabat, Morocco,3Laboratoire de Microbiologie, ORSTOM, Dakar, Senegal, and 4Laboratoire d’Ecophysiologie et de productionvegetales, Departement de Biologie, Faculte des Sciences, Oujda, Morocco
6925/98: received 14 October 1998, revised 6 January 1999 and accepted 11 January 1999
H. ABDELMOUMEN, A. FILALI-MALTOUF, M. NEYRA, A. BELABED AND M. MISSBAH EL IDRISSI.
1999. Twenty-eight reference strains, 79 rhizobia isolated from Trigonella foenum graecum(fenugreek), 26 strains isolated from root nodules of Ceratonia siliqua (carob tree), 30strains isolated from nodules of Adenocarpus decorticans and five isolated fromCytisus arboreus root nodules, were screened for their tolerance to increasedconcentrations of NaCl. Nine isolates of fenugreek were able to grow on medium containing10% NaCl and one strain still grew at 14%. The effect of a range of salts at 2·5 and5% (w/v) on the growth of rhizobia was assessed and it was shown that this effectdepended on the ion form and the strains used. In general, NaCl appeared to be a goodindicator of the tolerance of the strains to salts. The concentrations of the salts used werehigh and even at 5%, growth was not completely arrested in the less tolerant strains.Some substances, such as glutamate, proline, glycine betaine and CaCl2, were tested asosmotica. The effect of the two amino acids and betaine was confirmed with all thestrains used whereas the alleviating effect of CaCl2 was not observed in all strains.This salt had different effects on two isolates of fenugreek. These results revealed agreat diversity in salt tolerance, correlated with different responses to other stressconditions, which may be due to diversity in microbial ecology.
INTRODUCTION
Saline soils are common in regions of arid or semi-arid climatewhere transport of soluble salts to the ocean does not occurbecause of low rainfall (Hayward and Wadleigh 1949; Pillaiand Sen 1973; Webster and Wilson 1980). They are char-acterized by the presence of high levels of neutral salts in thesurface layers resulting from the capillary rise of water whenevaporation exceeds precipitation. In the flood plains ofrivers, low-lying lake margins and coastal plains, salinegroundwater within a few meters of the soil surface can be amajor contributory factor (Eaglesham and Ayanaba 1984).The predominant salts are usually sulphates and chlorides
Correspondence to: Dr M. Missbah El Idrissi, Laboratoire de MicrobiologieAppliquee, Departement de Biologie, Faculte des Sciences, Universite MohamedI, Oujda, Morocco (e-mail: [email protected]).
© 1999 The Society for Applied Microbiology
of sodium, calcium and sometimes magnesium, and smallquantities of carbonates and bicarbonates are often present.
Many legumes appear to offer potential for planting onsaline sites where secondary salinity is causing loss of agri-cultural land. In the developing countries, fast-growing leg-umes are required for both fodder and sustained fuelwoodproduction (Craig et al. 1991).
Leguminous plants growing in highly saline environmentsrequire both the free-living rhizobia and the host to be tol-erant to salt. Some workers (Bhardwaj 1975; Lauter et al.1981; Singleton et al. 1982; Kassem et al. 1985) have foundthat the upper limits for salinity tolerance of rhizobia appearto be higher than those of their host legumes.
Salt may affect symbiosis by its effects on the growth andsurvival of rhizobia in the soil, restrictions on root colon-ization, inhibition of processes of infection and nodule devel-

890 H. ABDELMOUMEN ET AL.
opment, or impairment of active nodule functioning. Theseeffects may be mediated through an effect of salt on the host,or through a specific effect on the micro-symbiont itself.
Rhizobia show marked variation in salt tolerance. A num-ber are inhibited by 100mmol l−1 salt (Singleton et al. 1982;Yelton et al. 1983; Zhang et al. 1991), but growth at saltconcentrations of more than 300mmol l−1 has been reportedfor strains of Sinorhizobium meliloti (Graham and Parker 1964;Sauvage et al. 1983), S. fredii (Yelton et al. 1983) and Rhi-zobium tropici (Graham 1992). Some alfalfa, Acacia, Prosopisand Leucaena strains will tolerate 500mmol l−1 NaCl (Sauv-age et al. 1983; Zhang et al. 1991).
The ability of organisms to adapt to fluctuations in theosmolarity of their surrounding is of fundamental importancefor their survival. In order to grow, cells must maintainpositive turgor (Booth et al. 1988). Sinorhizobium meliloti hasbeen extensively investigated, and the mechanisms governingturgor restoration of the bacterial cells growing under hyp-erosmotic conditions have been partially elucidated (Botsford1984; Le Rudulier and Bernard 1986; Smith et al. 1988; Bots-ford and Lewis 1990). Specific concomitant accumulation ofpotassium and glutamate ions was found to be the primaryresponse in S. meliloti (Botsford and Lewis 1990), and thesame mechanism has been found in most of the bacterialspecies investigated thus far (Booth et al. 1988).
The enhancement of growth of rhizobia from differentspecies, resulting from added glycine betaine and other beta-ines under saline conditions, has been reported (Sauvage et al.1983; Bernard et al. 1986). However, Botsford (1984) reportedthat glycine betaine did not ameliorate inhibition of thegrowth of S. meliloti in NaCl. In several rhizobial species,ectoine has also been reported to exhibit osmoprotectiveproperties without being accumulated (Talibart et al. 1994).Furthermore, the induction of salt tolerance by calcium hasbeen observed in R. leguminosarum bv. viciae (Chien et al.1991).
It should be noted that the principal sources of salt tol-erance reside in little-studied and under-collected organisms,poorly represented in most culture collections. This makes astrong case for continuing the collection of rhizobia andcharacterization of new isolates.
An investigation is reported here of the effect of highsodium chloride concentrations on the growth of some newlyisolated rhizobia, the effect of a range of salts on growth ofthese rhizobia, and the effect of externally added osmotica onsalt tolerance of rhizobia.
MATERIALS AND METHODS
Origin of the strains
Sixteen strains of rhizobia were obtained from the NifTalCenter Project (Hawaii, USA) and 12 were obtained from
© 1999 The Society for Applied Microbiology, Journal of Applied Microbiology 86, 889–898
USDA (USA) (Table 1). Eighty strains were isolated fromthe nodules of Trigonella foenum graecum (fenugreek) grownin different soils from the semi-arid to arid regions of north-east Morocco. Twenty-six strains of Rhizobium sp. fromCeratonia siliqua (carob tree), 30 strains newly isolated fromroot nodules of Adenocarpus decorticans and five strainsfrom Cytisus arboreus were also used.
All strains were maintained at 4 °C on Yeast Extract Man-nitol (YEM) agar slopes incorporating 3 g l−1 CaCO3 (Vincent1970).
Seeds sampling and germination
Seeds of Ceratonia siliqua and Trigonella foenum graecum wereobtained from pods sold (for medicinal purposes) in themarkets of Oujda. Seeds (and root nodules) of Adenocarpusdecorticans, used for effectiveness and infectivity trials, werecollected from fields in the mountains of Beni Znassen in thenorth-east of Oujda. Seeds (and nodules) of Cytisus arboreuswere collected from fields not far from the locality of Debdou(about 150 km to the south-east of Oujda), a semi-arid zonewith a mean rainfall of 350mm year−1 and mean minimumand maximum temperatures of 2 and 40 °C, respectively. Thesoil is alkaline with a pH of 8·32 0·1.
Before use, seeds of carob and A. decorticans were surface-sterilized by 5min exposure to 6% sodium hypochlorite and1 h exposure to concentrated H2SO4 for scarification, followedby thorough washing in sterile distilled water. To promotegermination, seeds were transferred to sterile water agar(0·6% w/v) plates and allowed to germinate at 26 °C in thedark. After germination, seedlings were aseptically trans-ferred as required.
Seeds of fenugreek were surface-sterilized by 5minexposure to 6% sodium hypochlorite and transferred to sterilewater agar plates. Seeds of C. arboreus were sterilized withsodium hypochlorite, steeped in a boiling bath for 30 s, thenplaced in ice overnight before transfer to water agar plates.
Tests of nodulation
All isolates were tested for nodulation of their original hosts.Seedlings were grown in plastic pots (diameter, 7 cm; height,14 cm) containing sterilized and nitrogen-free sand. Pots wereplaced in a growth chamber at 26 °C during the day and 20 °Cduring the night, illuminated for 14 h d−1, and watered witha dilution of Jensen nitrogen-free solution (Vincent 1970).Strains were grown on YEM broth and inoculated onto seed-lings. Nodulation was checked after 1–3 months.
GROWTH OF RHIZOBIA 891
Table 1 Effect of salt concentration (in g 100 mi−1) on the growth of rhizobia on YEMA medium—––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––
Salt concentration (%)—––––––––––––––––––––––––
Strains Hosts Origin Number 1 2 3 4 5—––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––Rhizobium etli USDA 9032 Phaseolus vulgaris USDA‡ 1 0† 0 0 0 0Sinorhizobium fredi USDA 205 Glycine max USDA 1 0 0 0 0 0Rhizobium galegae USDA 4128 Galegae orientalis USDA 1 0 0 0 0 0M. huakuii USDA 4779 Astragalus sinicus USDA 1 0 0 0 0 0Rhizobium leguminosarum bv phaseoli USDA 2671 Phaseolis vulgaris USDA 1 1 1 1 1 0Rhizobium leguminosarum bv trifoli USDA 2145 Trifolium pratens USDA 1 0 0 0 0 0Rhizobium leguminosarum bv viciae USDA 2370 Pisum sativum USDA 1 0 0 0 0 0Mesorhizobium loti USDA 3471 Lotus tenuis USDA 1 0 0 0 0 0S. meliloti 102F34 Melilotus sp. France§ 1 1 1 0 0 0S. meliloti USDA 1002 Melilotus sp. USDA 1 1 1 1 1 0Rhizobium tropici II. A USDA 9039 Phaseolus vulgaris USDA 1 0 0 0 0 0Rhizobium tropici II. B USDA 9030 Phaseolus vulgaris USDA 1 1 0 0 0 0
Rhizobium sp.Tal 1906 Glycine max NifTal 1 0 0 0 0 0
Center¶Tal 1884 Mimosa pudica Center 1 1 1 0 0 0Tal 169 Vigna unguiculata Center 1 0 0 0 0 0Tal 1826 Trifolium repens Center 1 0 0 0 0 0Tal 379 Glycine max Center 1 0 0 0 0 0Tal 634 Lens culinaris Center 1 0 0 0 0 0Tal 173 Canavalia gladiata Center 1 0 0 0 0 0Tal 833 Samanea saman Center 1 0 0 0 0 0Tal 183 Robinia pseudoacacia Center 1 0 0 0 0 0Tal 674 Sesbania rostrata Center 1 1 0 0 0 0Tal 480 Cicer arietinum Center 1 0 0 0 0 0Tal 1945 Sophora chrysophylla Center 1 0 0 0 0 0Tal 1770 Glyricidia sepium Center 1 0 0 0 0 0Tal 1983 Phaseolus vulgaris Center 1 0 0 0 0 0Tal 1950 Sophora chrysophylla Center 1 0 0 0 0 0Tal 1536 Albizia lebbek Center 1 0 0 0 0 0Rhizobium sp. Trigonella foenum graecum This study 79 79 79 42 43 18Rhizobium sp. Ceratonia siliqua This study 16 16 15 14 10 7Rhizobium sp. Adenocarpus decorticans This study 30 30 28 24 19 18Rhizobium sp. Cytisus arboreus This study 5 5 4 2 1 0—––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––
*Number of strains used.†Values denote number of strains giving positive growth.‡United States Department of Agriculture.§Faculte des Sciences, Nice, France.¶Niftal Project, Paia, Hawaii, USA.
© 1999 The Society for Applied Microbiology, Journal of Applied Microbiology 86, 889–898

892 H. ABDELMOUMEN ET AL.
Effect of NaCl on growth
The effect of salt on growth was assessed by two methods:(i) by first screening on TY agar (Beringer 1974) plates con-taining 1, 2, 3, 4 and 5% (w/v) NaCl. The plates wereinoculated with about 108 cells, and the growth was scoredafter 7 d at 28 °C; (ii) by determining the absorbance at 600 nmin MGS broth basal medium (Missbah et al. 1996) for selectedstrains Ad41, ChS10, Cyt8, S3G and S9D. The salt wasadded to the medium before autoclaving to give final con-centrations of 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 and 12% (w/v). Themedia were dispensed as 5ml aliquots into test tubes. Afterautoclaving, the tubes were inoculated with freshly preparedcultures of the appropriate strain in MGS to give an initialoptical density (O.D.) of 0·01–0·05.
Cultures were grown at 28 °C in a rotary shaker at70 rev min−1 and the final O.D. was recorded after 6 d at600 nm.
Effect of a range of salts on the growth
Six different salts were used to determine their specific ioneffects on the growth of strains Ad41, ChS10, Cyt8, S3Gand S9D isolated, respectively, from Adenocarpus decorticans,Ceratonia siliqua, Cytisus arboreus, and the two last fromfenugreek root nodules. The salts were Na2SO4, NaCl,MgSO4.7H2O, MgCl2.6H2O, K2SO4 and KCl. The salts wereadded to the medium before autoclaving to give the con-centrations 0, 2·5 and 5% (w/v). After autoclaving, the mediadispensed as 5ml aliquots into test tubes were inoculated andthe final O.D. assessed as described for NaCl test.
Effect of externally added osmotica
Glutamate, proline, glycine betaine and CaCl2 were addedas filter-sterilized solutions to MGS medium (with 0·5 g l−1
NH4Cl2 instead of glutamate) containing 3 or 6% (w/v) NaClto give a final concentration of 0 or 10mmol l−1 glutamate.
Aliquots (50 ml) were dispensed into 250ml flasks andinoculated with freshly prepared cultures of rhizobia. Theywere incubated in a shaker (70 rev min−1) at 28 °C. The O.D.was assessed at 600 nm.
Salt (NaCl) was added to the medium before autoclavingto give concentrations of 3% for strains Cyt8, RhCS10 andS9D, or 6% for the salt-tolerant strains Ad41 and S3G.
RESULTS
Effect of NaCl on growth of strains
Strains used as standards showed a great variation in theirresponses to salt. Hence, strains USDA 4779, USDA 2370,USDA 9032, USDA 4128, USDA 2145, Tal 1906, Tal 169,
© 1999 The Society for Applied Microbiology, Journal of Applied Microbiology 86, 889–898
Tal 1826, Tal 173, Tal 833, Tal 183, Tal 480, Tal 1945, Tal1950 and Tal 1536 did not grow at salt concentration of 1%,whereas strains such as USDA 2671 and USDA 1002 grew at3% NaCl. Among all the standards used, only strain 2671still grew at 4% NaCl, but it was inhibited by 5% (Table 1).
Strains isolated from root nodules of Leguminosae fromnorth-east Morocco were tolerant to high salt concentrations,with 18 strains (out of 30) from Adenocarpus decorticans andseven strains (out of 26) isolated from Ceratonia siliqua tol-erant to 5% NaCl.
Some strains grew in NaCl concentrations up to 10%;among the strains isolated from nodules of Trigonella foenumgraecum, nine were able to grow at 10%, four grew at 12%and one grew at 14% (w/v) (Table 2).
Some strains, S8A, AdA1 and AD29, were inhibited by 2%(w/v) NaCl.
Some of the strains tolerant to 5% NaCl and to other stressconditions, such as pH, temperature and some heavy metals,were selected to test their response to much media with amuch higher NaCl concentration (Table 2).
The behaviour of the strains in salt-stressed medium wasalso assessed in MGS broth. It is interesting to note thatthere was a large decrease in cell number, expressed as O.D.,in salt concentrations, whereas growth on solid medium wasstill significant (Fig. 1).
Table 2 Tolerance of strains isolated from Trigonella foenumgraecum root nodules to NaCl concentrations up to 5% (w/v)—–––––––––––––––––––––––––––––––––––––––––––––––––––––
NaCl concentrations in g 100 ml−1
—––––––––––––––––––––––––––––––––––––––––Strains 5 6 7 8 9 10 12 14—–––––––––––––––––––––––––––––––––––––––––––––––––––––S3G ¦ ¦ ¦ ¦ ¦ ¦ ¦ −S9A ¦ ¦ − − − − − −S9C ¦ ¦ ¦ ¦ ¦ ¦ − −S10F ¦ ¦ ¦ ¦ ¦ ¦ ¦ ¦S10H ¦ ¦ ¦ ¦ ¦ ¦ − −S10J ¦ ¦ ¦ ¦ ¦ ¦ ¦ −S12D ¦ ¦ ¦ ¦ ¦ ¦ − −S17A ¦ ¦ − − − − − −S2TR ¦ ¦ ¦ ¦ ¦ ¦ − −S7TR ¦ ¦ ¦ ¦ ¦ ¦ − −S10TR ¦ ¦ ¦ ¦ ¦ ¦ − −S13TR ¦ ¦ ¦ ¦ ¦ ¦ − −S14TR ¦ ¦ ¦ ¦ ¦ ¦ ¦ −S22TR ¦ ¦ ¦ ¦ ¦ ¦ − −S37TR ¦ ¦ ¦ ¦ ¦ ¦ − −S39TR ¦ ¦ ¦ ¦ ¦ ¦ − −—–––––––––––––––––––––––––––––––––––––––––––––––––––––
¦, Colonies observed on YEM solidified medium.−, No growth observed.

GROWTH OF RHIZOBIA 893
Cyt 8
S3G
Salt concentration (g 100 ml–1 )
RhC S10 Inh
ibit
ion
of
gro
wth
(%
)
100
0
1%
S9D
Ad 41
80
60
40
20
2%4%
6%8%
10%
Fig. 1 Effect of salt concentration on the growth of rhizobia
Effect of a range of salts on rhizobia
Strains Ad41, RhCS10, Cyt8, S3G and S9D were used forthe remainder of the study. The three first represented,respectively, Adenocarpus decorticans, Ceratonia siliqua andCytisus arboreus isolates, whereas the last two were isolatesfrom Trigonella; S3G tolerates 12% NaCl whereas S9D issensitive to salt concentrations up to 4% (w/v).
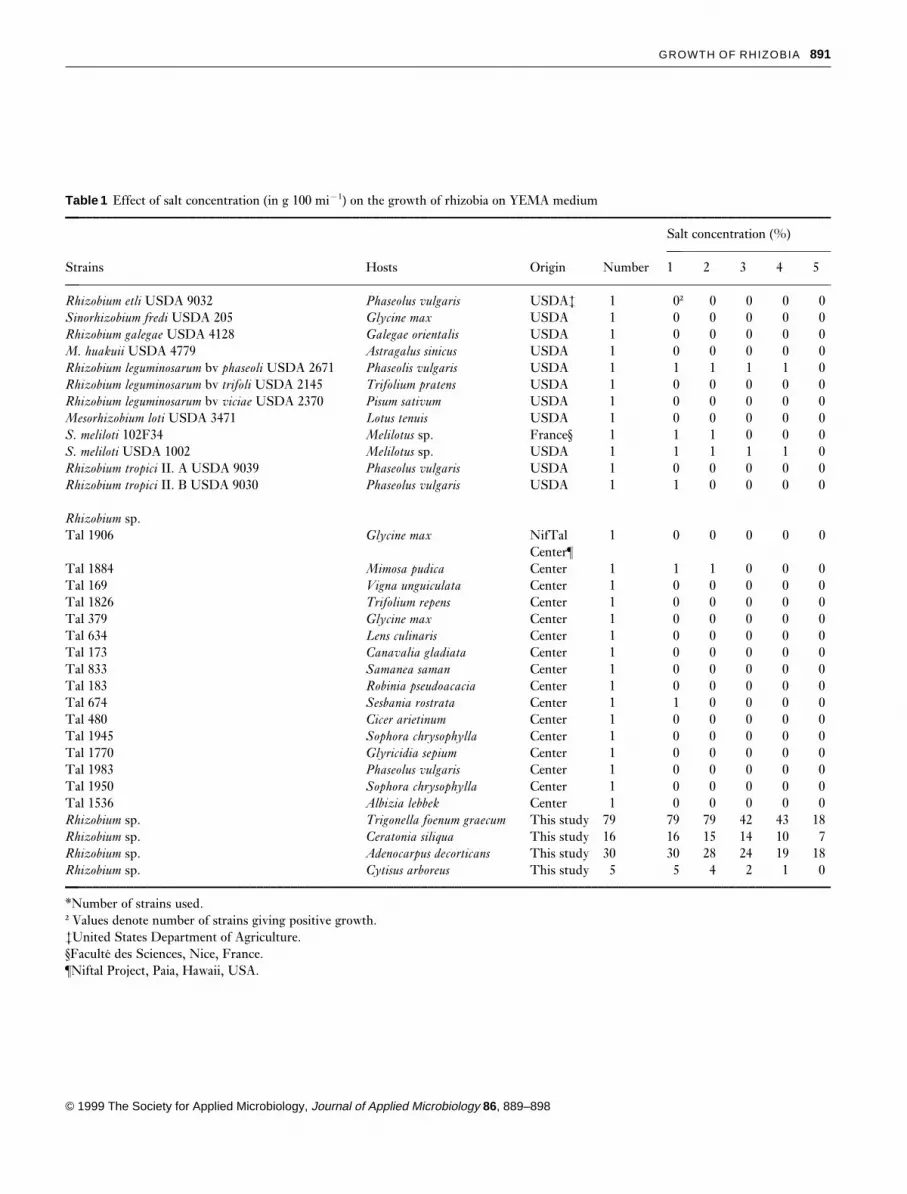
The effect of the different salts was much more inhibitoryat 5% than at 2·5% for all the strains (Fig. 2).
Ad41 was not inhibited by KCl at all, and the sulphateform of magnesium was the most inhibitory among the allsalts tested. It is notable that for this strain, the sulphateforms were much more inhibitory than the chloride forms.RhCS10 was strongly inhibited by 5% of both forms ofsodium; K2SO4 and KCl had the same effect. In a previouspaper, it was noted that on solid medium, this strain wastolerant to 4% NaCl but was very sensitive to KCl. StrainCyt8 was strongly inhibited by all the salts except MgSO4.Strain S3G was only slightly inhibited, if at all, by saltswhereas strain S9D was inhibited by 5% of most salts exceptmagnesium.
The behaviour of all five strains in Na2SO4-supplementedmedium was the same as in NaCl.
Response of the strains to added osmotica
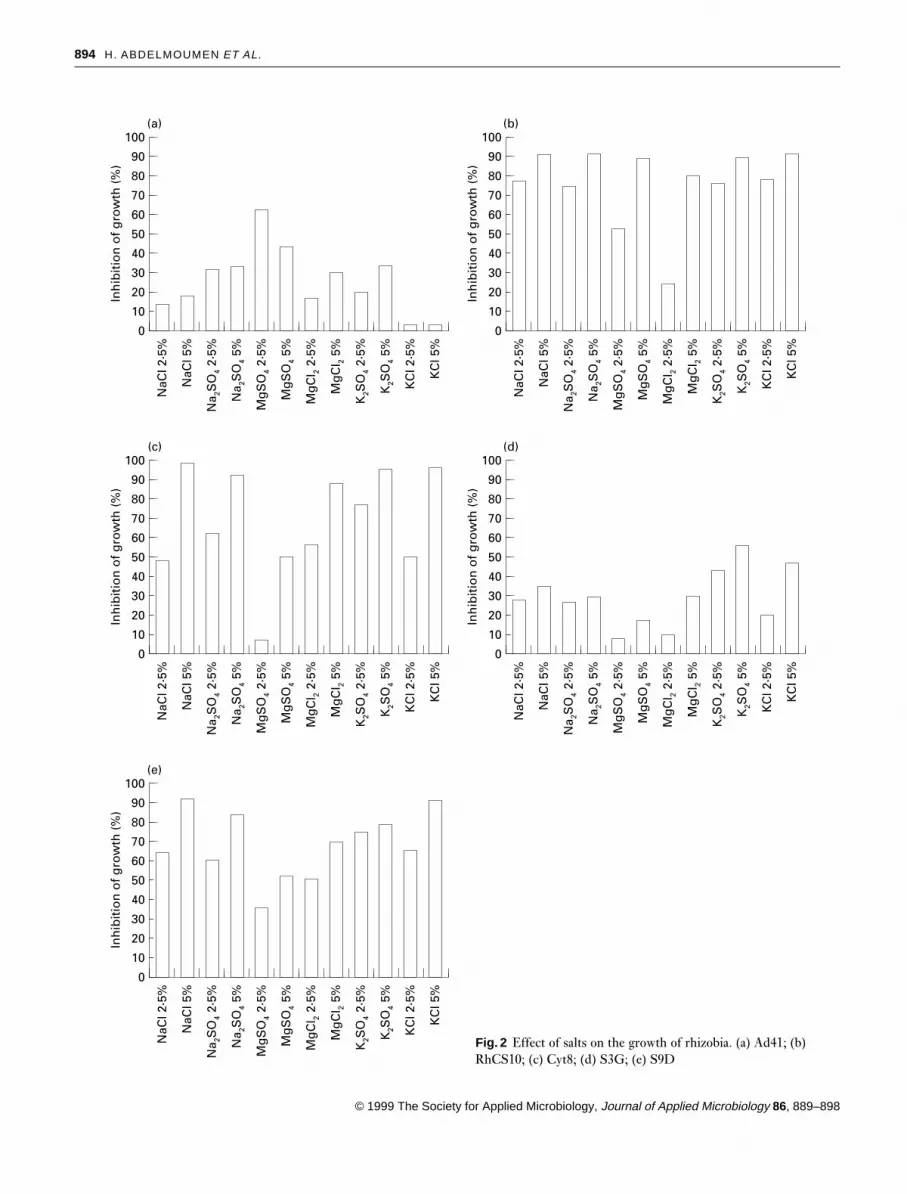
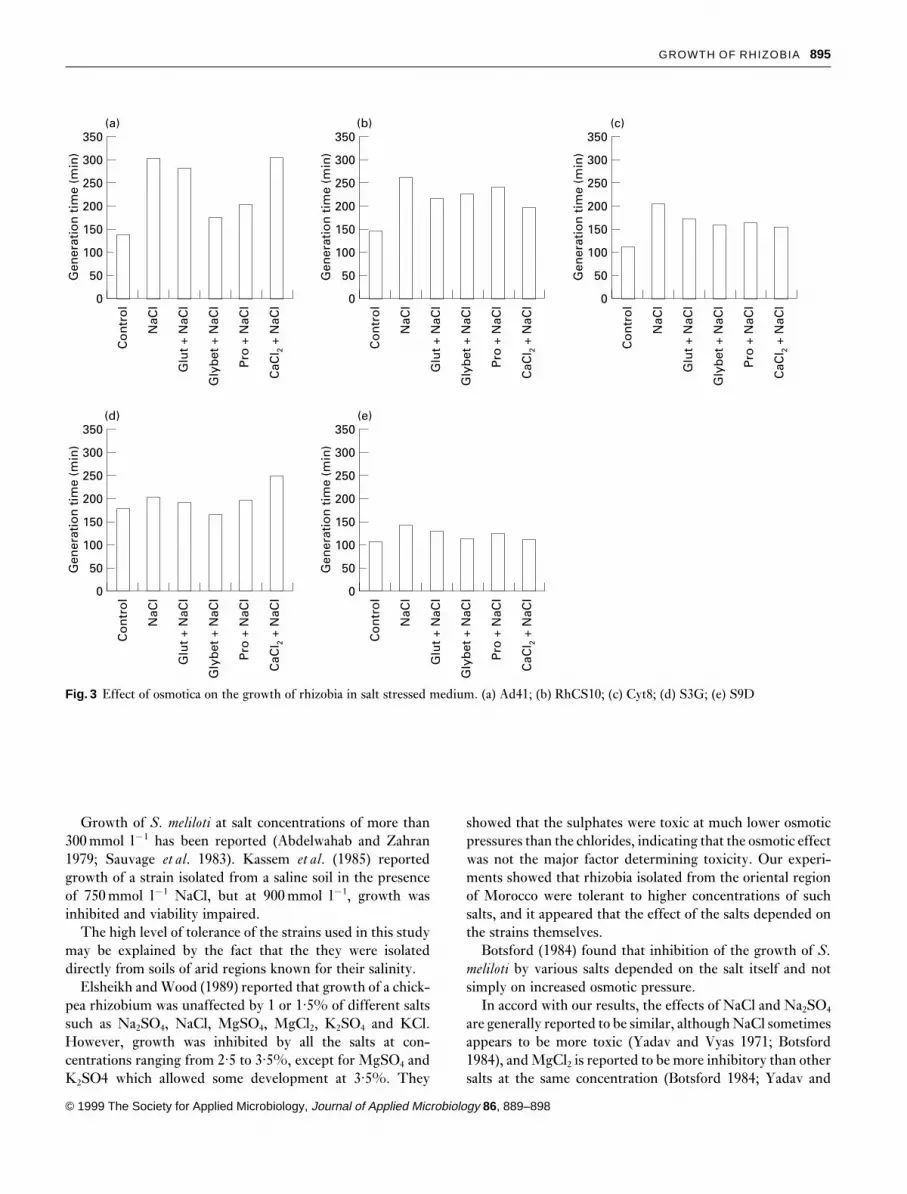
Effect of glutamate. Except for S9D, growth of the strainswas greatly enhanced in the presence of glutamate in MGSmedium without any salt. Glutamate improved the growthof all the strains in NaCl-containing media, but its effect wasmuch more remarkable in the case of strain S9D whichoutgrew the control (in MGS medium in which glutamatewas replaced by NH4Cl as nitrogen source).
© 1999 The Society for Applied Microbiology, Journal of Applied Microbiology 86, 889–898
For strains Cyt8, RhCS10 and S3G, the final biomass after3 d of incubation in salt- and glutamate-containing mediumwas higher than that of the control. This wass probably dueto the fact that glutamate was used by the cells as a nitrogenand/or carbon source in conditions where those sourcesbecame less available in the control medium (Fig. 3).
Effect of glycine betaine. Glycine betaine stimulated thegrowth of strains Cyt8, RhCS10 and S3G without any addedNaCl, but did not stimulate growth of Ad41 or S9D. Theosmoprotective effect of this substance was observed in theall five strains, as its presence in the media containing NaClprevented the inhibitory effect of the salt (Fig. 3).
Cyt8 showed a higher final biomass (in 3 d) when cultivatedin MGS salt-stressed medium containing glycine betaine thanin the control medium.
Effect of proline. Except for strain S9D, growth of all strainswas improved in the presence of proline in media withoutany added salt. The action of this amino acid was similar tothat of glycine betaine, i.e. the addition of proline enhancedthe growth of all the strains in the presence of NaCl. Thefinal biomass of strains Cyt8, RhCS10 and S3G was higherthan the control (Fig. 3).
Effect of CaCl2. Calcium slightly stimulated growth of strainsS3G and Ad41 but inhibited growth of strain S9D. Thephenomenon of external Ca2¦ alleviating salt stress wasclearly observed in strain S9D; calcium chloride increased itsgrowth in the presence of 3% NaCl.
Calcium slightly enhanced growth of strains RhCS10,Ad41 and Cyt8 in the presence of sodium (Fig. 3), but anopposite effect was obtained with strain S3G, although thefinal O.D. reached that of the control.
DISCUSSION
Rhizobia isolated from soils of the north-east region of Mor-occo differ in their response to salt stress. The level of tol-erance of these strains was generally very high in comparisonwith the reference strains used in this study and to the levelsreported in the literature.
In general, the effect of NaCl seems to be a good indicatorof the response of rhizobia to the different salts.
It has been reported that tolerance of different rhizobia toNaCl varies from 0·1 to 0·75mol l−1 (Tu 1981; Le Rudulierand Bouillard 1983; Kassem et al. 1985; Bernard et al. 1986).Graham and Parker (1964) showed that fast-growing, acid-producing rhizobia were generally more salt tolerant than theslow-growing, alkali-producing strains.
894 H. ABDELMOUMEN ET AL.
100
0
Inh
ibit
ion
of
gro
wth
(%
)
50
NaC
l 2·5
%90
80
70
60
40
30
20
10
(a)
NaC
l 5%
Na
2S
O4 2
·5%
Na
2S
O4 5
%
Mg
SO
4 2
·5%
Mg
SO
4 5
%
Mg
Cl 2
2·5
%
Mg
Cl 2
5%
K2S
O4 2
·5%
K2S
O4 5
%
KC
l 2·5
%
KC
l 5%
100
0
Inh
ibit
ion
of
gro
wth
(%
)
50
NaC
l 2·5
%
90
80
70
60
40
30
20
10
(b)
NaC
l 5%
Na
2S
O4 2
·5%
Na
2S
O4 5
%
Mg
SO
4 2
·5%
Mg
SO
4 5
%
Mg
Cl 2
2·5
%
Mg
Cl 2
5%
K2S
O4 2
·5%
K2S
O4 5
%
KC
l 2·5
%
KC
l 5%
100
0
Inh
ibit
ion
of
gro
wth
(%
)
50
NaC
l 2·5
%
90
80
70
60
40
30
20
10
(c)
NaC
l 5%
Na
2S
O4 2
·5%
Na
2S
O4 5
%
Mg
SO
4 2
·5%
Mg
SO
4 5
%
Mg
Cl 2
2·5
%
Mg
Cl 2
5%
K2S
O4 2
·5%
K2S
O4 5
%
KC
l 2·5
%
KC
l 5%
100
0
Inh
ibit
ion
of
gro
wth
(%
)
50
NaC
l 2·5
%
90
80
70
60
40
30
20
10
(d)
NaC
l 5%
Na
2S
O4 2
·5%
Na
2S
O4 5
%
Mg
SO
4 2
·5%
Mg
SO
4 5
%
Mg
Cl 2
2·5
%
Mg
Cl 2
5%
K2S
O4 2
·5%
K2S
O4 5
%
KC
l 2·5
%
KC
l 5%
100
0
Inh
ibit
ion
of
gro
wth
(%
)
50
NaC
l 2·5
%
90
80
70
60
40
30
20
10
(e)
NaC
l 5%
Na
2S
O4 2
·5%
Na
2S
O4 5
%
Mg
SO
4 2
·5%
Mg
SO
4 5
%
Mg
Cl 2
2·5
%
Mg
Cl 2
5%
K2S
O4 2
·5%
K2S
O4 5
%
KC
l 2·5
%
KC
l 5%
Fig. 2 Effect of salts on the growth of rhizobia. (a) Ad41; (b)RhCS10; (c) Cyt8; (d) S3G; (e) S9D
© 1999 The Society for Applied Microbiology, Journal of Applied Microbiology 86, 889–898
GROWTH OF RHIZOBIA 895
350
0
Gen
era
tio
n t
ime (
min
)
200
Co
ntr
ol
300
250
150
100
50
(a)
NaC
l
Glu
t +
NaC
l
Gly
bet
+ N
aC
l
Pro
+ N
aC
l
CaC
l 2 +
NaC
l
350
0
Gen
era
tio
n t
ime (
min
)
200
Co
ntr
ol
300
250
150
100
50
(b)
NaC
l
Glu
t +
NaC
l
Gly
bet
+ N
aC
l
Pro
+ N
aC
l
CaC
l 2 +
NaC
l
350
0
Gen
era
tio
n t
ime (
min
)
200
Co
ntr
ol
300
250
150
100
50
(c)
NaC
l
Glu
t +
NaC
l
Gly
bet
+ N
aC
l
Pro
+ N
aC
l
CaC
l 2 +
NaC
l
350
0
Gen
era
tio
n t
ime (
min
)
200
Co
ntr
ol
300
250
150
100
50
(d)
NaC
l
Glu
t +
NaC
l
Gly
bet
+ N
aC
l
Pro
+ N
aC
l
CaC
l 2 +
NaC
l
350
0
Gen
era
tio
n t
ime (
min
)
200
Co
ntr
ol
300
250
150
100
50
(e)N
aC
l
Glu
t +
NaC
l
Gly
bet
+ N
aC
l
Pro
+ N
aC
l
CaC
l 2 +
NaC
l
Fig. 3 Effect of osmotica on the growth of rhizobia in salt stressed medium. (a) Ad41; (b) RhCS10; (c) Cyt8; (d) S3G; (e) S9D
Growth of S. meliloti at salt concentrations of more than300mmol l−1 has been reported (Abdelwahab and Zahran1979; Sauvage et al. 1983). Kassem et al. (1985) reportedgrowth of a strain isolated from a saline soil in the presenceof 750mmol l−1 NaCl, but at 900mmol l−1, growth wasinhibited and viability impaired.
The high level of tolerance of the strains used in this studymay be explained by the fact that the they were isolateddirectly from soils of arid regions known for their salinity.
Elsheikh and Wood (1989) reported that growth of a chick-pea rhizobium was unaffected by 1 or 1·5% of different saltssuch as Na2SO4, NaCl, MgSO4, MgCl2, K2SO4 and KCl.However, growth was inhibited by all the salts at con-centrations ranging from 2·5 to 3·5%, except for MgSO4 andK2SO4 which allowed some development at 3·5%. They
© 1999 The Society for Applied Microbiology, Journal of Applied Microbiology 86, 889–898
showed that the sulphates were toxic at much lower osmoticpressures than the chlorides, indicating that the osmotic effectwas not the major factor determining toxicity. Our experi-ments showed that rhizobia isolated from the oriental regionof Morocco were tolerant to higher concentrations of suchsalts, and it appeared that the effect of the salts depended onthe strains themselves.
Botsford (1984) found that inhibition of the growth of S.meliloti by various salts depended on the salt itself and notsimply on increased osmotic pressure.
In accord with our results, the effects of NaCl and Na2SO4
are generally reported to be similar, although NaCl sometimesappears to be more toxic (Yadav and Vyas 1971; Botsford1984), and MgCl2 is reported to be more inhibitory than othersalts at the same concentration (Botsford 1984; Yadav and

896 H. ABDELMOUMEN ET AL.
Vyas 1971; Elsheikh and Wood 1989). However, Amara andMiller (1986) reported magnesium ions to be the least inhibi-tory to growth.
Glutamate enhanced the growth of all the strains in thepresence of NaCl, probably because of its accumulation (con-comitant with potassium accumulation) in the cytoplasmwhich plays a key role in turgor restoration of bacterial cellsgrowing under hyperosmotic conditions (Hua et al. 1982; Yapand Lim 1983; Yelton et al. 1983; Yang and Li 1989; Talibartet al. 1994).
Effectively, in different bacterial species, the intracellularconcentration of this amino acid was found to be proportionalto the external medium osmolarity. It is considerably aug-mented in Gram-negative bacteria as a consequence of hyper-osmotic shock (Booth 1993; Csonka 1989; Talibart et al.1994). Moreover, potassium glutamate has been consideredto be a signal for the activation of several systems involved inthe osmotic response of enteric bacteria (Ramirez et al. 1989).
However, some authors found no genuine effect of glu-tamate with NaCl. Elsheikh and Wood (1989) reported noimprovement in the growth of chickpea rhizobia with theaddition of glutamate to 340mmol l−1 NaCl. A similar resultwas reported for S. meliloti (Botsford 1984). Furthermore,Gonzalez-Gonzalez et al. (1990) demonstrated little changein the specific activity of the enzymes involved in glutamatesynthesis with 400mmol l−1 NaCl in the medium.
Nevertheless, the effect of glycine betaine does not seemto be general and depends on the species of rhizobia used; LeRudulier and Bernard (1986) found that the addition of gly-cine betaine significantly reduced the doubling time of strainsof S. meliloti following exposure to NaCl, whereas no ben-eficial effect, or only a slight improvement in growth, wasfound for Bradyrhizobium strains (Bernard et al. 1986;Talibart et al. 1994).
On exposure to NaCl, salt-tolerant rhizobia increase theirrate of uptake of glycine betaine; they are able to synthesizethis substance more rapidly from choline but show decreasedcatabolism of glycine betaine (Bernard et al. 1986; Le Rud-ulier and Bernard 1986; Smith et al. 1988).
Le Rudulier and Bouillard (1983) reported that Gram-negative bacteria are entirely dependent on exogenous prolinefor its accumulation in high concentrations during osmoticstress, an event which will relieve the increase in ionicstrength (Smith 1985; Booth et al. 1988). However, Measures(1975) found that osmotic stress resulted in large increasesin the intracellular levels of proline in a large variety ofbacteria.
In rhizobia isolated from Acacia root nodules (Craig et al.1991), proline was either not detected or was detected innegligible amounts, whereas the addition of proline did notinfluence stationary phase cell density of Rh. leguminosarumbv. viciae C1204b in the presence of NaCl (Chien et al.1991).
© 1999 The Society for Applied Microbiology, Journal of Applied Microbiology 86, 889–898
In bacteria, it is likely that compatible solutes exert theireffects at two levels. First, it has been well established thatthe solutes can interact with proteins in the presence of highsalt concentrations and stabilize their conformation, leadingto restoration of activity (Pollard and Wyn-Jones 1979; Arak-awa and Timoshev 1983). The second effect of compatiblesolutes is that their accumulation by the cell leads directly tothe release of potassium ions and a lowering of the cytoplasmicionic strength (Booth et al. 1988). Thus, there is a synergismbetween the direct effect of betaine or proline on proteinstructure and the indirect effect of lowering ionic strength.
Calcium was reported to have a beneficial action on rhizobiaunder stress conditions. Reeve et al. (1993) and Howiesonet al. (1992) reported that calcium ameliorated the growth ofS. meliloti in acid pH media. Chien et al. (1991) showed thatCaCl2 can induce NaCl tolerance in the free-living micro-symbiont Rh. leguminosarum bv. viciae C1204b and suggestedthat Ca2¦-induced alleviation of NaCl toxicity was due to aNa¦/Ca2¦ interaction rather than to an osmoprotectiveeffect of this divalent cation. Our results show that the effectof calcium chloride depends on the nature of the strain.
In conclusion, it appears that the effect of salts on thegrowth of rhizobia depends on the strains themselves, as theresponses of the different strains used were disparate, evenwithin isolates from the same host legume collected from thesame soil in the same region.
The high levels of saline stress to which strains were sub-mitted, and to which they showed a great resistance, suggestthat various ways may be used in concert by cells to withstandthese conditions.
REFERENCES
Abdelwahab, A.M. and Zahran, H.H. (1979) Salt tolerance of Rhi-zobium species in broth cultures. Zeitschrift fur Allgemande Mikro-biologie 19, 681–685.
Amara, D.S. and Miller, R.H. (1986) Effect of moisture and salt onselected Rhizobium phaseoli strains. Mircen Journal 2, 373–382.
Arakawa, T. and Timoshev, S.N. (1983) Preferential interactions ofproteins with solvent components in aqueous amino acids solu-tions. Archives of Biochemistry and Biophysics 224, 169–177.
Beringer, J.E. (1974) Factor transfer in Rhizobium leguminosarum.Journal of General Microbiology 84, 188–198.
Bernard, T., Pocard, J., Perroud, B. and Le Rudulier, D. (1986)Variation in the response of salt stressed Rhizobium strains tobetaines. Archives of Microbiology 143, 359–364.
Bhardwaj, K.K. (1975) Survival and symbiotic characteristics ofRhizobium in saline-alkali soils. Plant and Soil 43, 377–385.
Booth, I.R. (1993) Regulation of gene expression during osmo-regulation: the role of potassium glutamate as a secondary signalof osmotic stress. In Alkali Cation Transport Systems in Prokaryotesed. Baker, E.P. pp. 309–331. Boca Raton: CRC Press.
Booth, I.R., Cairney, J., Sutherland, L. and Higgins, C.F. (1988)

GROWTH OF RHIZOBIA 897
Enteric bacteria and osmotic stress: an integrated homeostaticsystem. Journal of Applied Bacteriology Symposium (Suppl.) 35S–49S.
Botsford, J.L. (1984) Osmoregulation in Rhizobium meliloti:inhibition of growth by salts. Archives of Microbiology 137, 124–127.
Botsford, J.L. and Lewis, T.A. (1990) Osmoregulation in Rhizobiummeliloti: production of glutamic acid in response to osmotic stress.Applied and Environmental Microbiology 56, 488–494.
Chien, C.T., Shetty, K., Mortimer, M. and Orser, C.S. (1991)Calcium induced salt tolerance in Rhizobium leguminosarum bvviciae strain C1204b. FEMS Microbiology Letters 83, 219–224.
Craig, G.F., Atkins, C.A. and Bell, D.T. (1991) Effect of the salinityon growth of four strains of Rhizobium and their infectivity andeffectiveness on two species of Acacia. Plant and Soil 133, 253–262.
Csonka, L.N. (1989) Physiological and genetic responses of bacteriato osmotic stress. Microbiological Reviews 53, 121–147.
Eaglesham, A.R.J. and Ayanaba, A. (1984) Tropical stress ecologyof Rhizobia, root nodulation and legume fixation. In CurrentDevelopments in Biological Nitrogen Fixation ed. Subba Rao, N.S.1–35. London: E. Arnold.
Elsheikh, E.A.E. and Wood, M. (1989) Response of chickpea andsoybean rhizobia to salt: osmotic and specific ion effects of salts.Soil Biology and Biochemistry 21, 889–895.
Gonzalez-Gonzalez, R., Botsford, J.L. and Lewis, T. (1990) Osmo-regulation in Rhizobium meliloti: characterization of enzymes inglutamate synthesis. Canadian Journal of Microbiology 36, 469–474.
Graham, P.H. (1992) Stress tolerance in Rhizobium and Brady-rhizobium and nodulation under adverse soil conditions. CanadianJournal of Microbiology 38, 475–484.
Graham, P.H. and Parker, C.A. (1964) Diagnostic features in thecharacterization of the root nodule bacteria of legumes. Plant andSoil 20, 383–396.
Hayward, H.E. and Wadleigh, C.H. (1949) Plant growth on salineand alkali soils. Advances in Agronomy 1, 1–38.
Howieson, J.G., Robson, A.D. and Abbot, L.K. (1992) Calciummodifies pH effects on the growth of acid tolerant and acidsensitive Rhizobium meliloti. Australian Journal of Agriculture andResearch 43, 765–772.
Hua, S.T., Tsai, V.T., Lichens, G.H. and Noma, A.T. (1982)Accumulation of amino acids in Rhizobium sp. strain WR1001 inresponse to sodium chloride salinity. Applied and EnvironmentalMicrobiology 44, 135–140.
Kassem, M., Cappelano, A. and Gounot, A.M. (1985) Effet duchlorure de sodium sur la croissance in vitro, l’infectivite etl’efficience de Rhizobium meliloti. Mircen Journal 1, 63–75.
Lauter, D.J., Munns, D.N.. and Clarkin, K.L. (1981) Salt responseof chickpea as influenced by N supply. Agronomy Journal 73, 961–966.
Le Rudulier, D.A. and Bernard, T. (1986) Salt tolerance in Rhi-zobium: a possible role for betaines. FEMS Microbiology Review39, 67–72.
Le Rudulier, D.A. and Bouillard, L. (1983) Glycine betaine anosmotic effector in Klebsiella pneumoniae and other members of
© 1999 The Society for Applied Microbiology, Journal of Applied Microbiology 86, 889–898
Enterobacteriaceae. Applied and Environmental Microbiology 46,152–159.
Measures, J.C. (1975) Role of amino acids in osmoregulation of non-halophilic bacteria. Nature 257, 398–400.
Missbah El Idrissi, M., Auajjar, N., Belabed, A., Dessaux, Y. andFilali-Maltouf, A. (1996) Characterization of rhizobia isolatedfrom carob tree (Ceratonia siliqua). Journal of Applied Bacteriology80, 165–173.
Pillai, R.N. and Sen, A. (1973) Salt tolerance of Rhizobium fromDolichos lablab. Zentralblatt fur Bakteriologie 128, 538–542.
Pollard, A. and Wyn-jones, R.G. (1979) Enzyme activities in con-centrated solutions of glycine betaine and other solutes. Planta144, 291–298.
Ramirez, R.M., Prince, W.S., Bremer, E. and Villarejo, M. (1989)In vitro reconstitution of osmoregulated expression of ProU ofEscherichia coli. Proceedings of the National Academy of SciencesUSA 86, 1153–1157.
Reeve, W.R., Tiwari, R.P., Dilworth, M.J. and Gleen, A.R. (1993)Calcium affects the growth and survival of Rhizobium meliloti.Soil Biology and Biochemistry 25, 581–586.
Sauvage, D., Hamelin, J. and Larher, F. (1983) Glycine betaine andother structurally related compounds improve the salt toleranceof Rhizobium meliloti. Plant Science Letters 31, 291–302.
Singleton, P.W., Swaify, S.A. and Bohlool, B.B. (1982) Effect ofsalinity on Rhizobium growth and survival. Applied and Environ-mental Microbiology 44, 884–890.
Smith, L.T. (1985) Characterization of a glutamyl kinase from E.coli that confers proline over production and osmotic tolerance.Journal of Bacteriology 164, 1088–1093.
Smith, L.T., Pocard, J.A., Bernard, T. and Le Rudulier, D.A.(1988) Osmotic control of betaine biosynthesis and degradationin Rhizobium meliloti. Journal of Bacteriology 170, 3142–3149.
Sutherland, L., Cairney, J., Elmore, M.J., Booth, I.R. and Higgins,C.F. (1986) Osmotic regulation of transcription: induction of theproU betaine transport gene is determined by the accumulationof intracellular potassium. Journal of Bacteriology 168, 805–814.
Talibart, R., Jebbar, M., Gouesbet, G. et al. (1994) Osmoadaptationin rhizobia: ectoine-induced salt tolerance. Journal of Bacteriology176, 5210–5217.
Tu, J.C. (1981) Effect of salinity on Rhizobium root hair interaction,nodulation and growth of soybean. Canadian Journal of PlantScience 61, 231–239.
Vincent, J.M. (1970). A Manual for the Practical Study of RootNodule Bacteria. IBP Handbook no. 15. Oxford: Blackwell Scien-tific Publications.
Webster, C.C. and Wilson, P.N. (1980) Agriculture in the Tropics.London: Longman.
Yadav, N.K. and Vyas, S.R. (1971) Response of root nodule bacteriato saline, alkaline and acid conditions. Indian Journal of Agri-cultural Sciences 41, 875–881.
Yang, S.S. and Li, J.L. (1989) The construction of salt tolerant higheffective strains of soyabean rhizobia. Acta Microbiologica Sinica29, 107–112.
Yap, S.F. and Lim, S.T. (1983) Response of Rhizobium UMKL20to sodium chloride stress. Archives of Microbiology 135, 224–228.

898 H. ABDELMOUMEN ET AL.
Yelton, M.M., Yang, S.S., Edie, S.A. and Lim, S.T. (1983) Charac-terzation of an effective salt tolerant fast growing strain of Rhi-zobium japonicum. Journal of General Microbiology 129, 1537–1547.
© 1999 The Society for Applied Microbiology, Journal of Applied Microbiology 86, 889–898
Zhang, X., Harper, R., Karsisto, M. and Lindstrom, K. (1991)Diversity of Rhizobium bacteria isolated from the root nodules ofleguminous trees. International Journal of Systematic Bacteriology41, 104–113.