differential glycosylation of polymeric and monomeric iga: a possible role in glomerular...
TRANSCRIPT

Differential Glycosylation of Polymeric and Monomeric IgA:A Possible Role in Glomerular Inflammation in IgANephropathy
Beatrijs D. Oortwijn,* Anja Roos,* Louise Royle,† Danielle J. van Gijlswijk-Janssen,*Maria C. Faber-Krol,* Jan-Willem Eijgenraam,* Raymond A. Dwek,† Mohamed R. Daha,*Pauline M. Rudd,† and Cees van Kooten**Department of Nephrology, Leiden University Medical Center, Leiden, The Netherlands; and †Glycobiology Institute,Department of Biochemistry, Oxford University, Oxford, United Kingdom
IgA nephropathy (IgAN) is characterized by mesangial deposition of polymeric IgA1 (pIgA1) and complement. Complementactivation via mannose-binding lectin and the lectin pathway is associated with disease progression. Furthermore, recentstudies have indicated a possible role for secretory IgA. IgAN is associated with abnormalities in circulating IgA, includingaberrant O-linked glycosylation. This study characterized and compared functional properties and N-linked glycosylation ofhighly purified monomeric IgA (mIgA) and pIgA from patients with IgAN and control subjects. Total serum IgA wasaffinity-purified from patients (n � 11) and control subjects (n � 11) followed by size separation. pIgA but not mIgA containedsecretory IgA, and its concentration was significantly higher in patients with IgAN than in control subjects. Both in patientswith IgAN and in control subjects, IgA binding to the GalNAc-specific lectin Helix Aspersa and to mannose-binding lectin wasmuch stronger for pIgA than for mIgA. Furthermore, binding of IgA to mesangial cells largely was restricted to polymeric IgA.Binding of pIgA to mesangial cells resulted in increased production of IL-8, predominantly with IgA from patients with IgAN.Quantitative analysis of N-linked glycosylation of IgA heavy chains showed significant differences in glycan compositionbetween mIgA and pIgA, including the presence of oligomannose exclusively on pIgA. In conclusion, binding and activationof mesangial cells, as well as lectin pathway activation, is a predominant characteristic of pIgA as opposed to mIgA.Furthermore, pIgA has different N-glycans, which may recruit lectins of the inflammatory pathway. These results underscorethe role of pIgA in glomerular inflammation in IgAN.
J Am Soc Nephrol 17: 3529–3539, 2006. doi: 10.1681/ASN.2006040388
P rimary IgA nephropathy (IgAN) is the most commonform of primary glomerulonephritis. The disease leadsto progressive renal failure in a substantial proportion
of patients. The hallmark of this disease is deposition of IgA inthe glomerular mesangium, together with markers of comple-ment activation (1,2). It generally is thought that this mesangialIgA mainly consists of IgA1 and is mostly polymeric (3). Thecomposition of polymeric forms of IgA (pIgA) in serum isdiverse and may include dimeric IgA, secretory IgA (SIgA),CD89 (Fc�RI)/IgA complexes, IgA immune complexes, andIgA–fibronectin complexes (4–6). Serum dimeric IgA consistsof two IgA molecules linked with J chain, whereas SIgA inaddition contains secretory component, derived from the mu-cosal epithelium.
Deposition of circulating IgA in the mesangium leads to renalinflammation, potentially involving direct interactions of IgA
with resident and infiltrating cells in the glomerulus, as well ascomplement activation. The inflammatory process results inrenal injury. Although the mechanism of IgA deposition in therenal mesangium of patients with IgAN has been a subject ofintensive research during the past decades, the pathogenesis ofIgAN is still incompletely characterized. A number of studiesprovided evidence for a mesangial IgA receptor, which is in-volved in mesangial cell activation by IgA in vitro (7–9).
Glomerular IgA deposition is associated with activation ofthe complement system (5), involving the alternative pathwayand the lectin pathway of complement (10). Recent studiesindicate deposition of mannose-binding lectin (MBL), a majorrecognition molecule of the lectin pathway of complement, in asubpopulation of patients in association with a more severerenal injury (10,11), whereas in vitro studies demonstrated bind-ing of MBL to polymeric serum IgA (12). Glomerular comple-ment activation can enhance renal injury via the proinflamma-tory effects of the complement activation cascade.
Studies in patients who had IgAN and received a renaltransplant showed recurrence of mesangial IgA deposition in ahigh number of cases (13). Conversely, the accidental transplan-tation of a kidney with mesangial IgA deposition into a recip-ient without IgAN resulted in spontaneous disappearance of
Received April 28, 2006. Accepted August 29, 2006.
Published online ahead of print. Publication date available at www.jasn.org.
B.D.O. and A.R. contributed equally to this work.
Address correspondence to: Dr. Cees van Kooten, LUMC, Department of Ne-phrology, C3-P, Albinusdreef 2, 2333 ZA Leiden, The Netherlands. Phone: �31-71-526-3964; Fax: �31-71-526-6868; E-mail: [email protected]
Copyright © 2006 by the American Society of Nephrology ISSN: 1046-6673/1712-3529

IgA deposits after transplantation (14). These studies stronglysuggest that IgAN is a systemic disease rather than a disease ofthe kidney.
On basis of these data, abnormalities in IgA are hypothesizedto be involved in the pathogenesis of IgAN. Therefore, circu-lating IgA from patients with IgAN has been studied exten-sively. Serum from patients with IgAN contains higher concen-trations of IgA (15,16). Recently, our group showed lowconcentrations of circulating SIgA in patients with IgAN andcontrol subjects, whereas patients with IgAN and a high serumconcentration of SIgA showed more hematuria (17). Further-more, SIgA accumulated in glomerular IgA deposits, suggest-ing a pathogenic role for SIgA in IgAN (17). Several studies alsofocused on IgA glycosylation, showing aberrant O-glycosyla-tion on circulating IgA from patients with IgAN, resulting inincreased Tn antigen (GalNAc�1-Ser/Thr) residues (18,19).This undergalactosylated IgA1 may lead to generation of cir-culating IgG–IgA1 complexes (20). O-linked glycans are presenton IgA1 but not IgA2, whereas IgA1 and IgA2 heavy chainsboth contain several N-glycosylation sites (21). The galactosy-lation of the N-glycans is not different between patients withIgAN and control subjects (22). However, the complete struc-ture of N-linked glycans on IgA has not been studied in IgAN.
Functional studies with purified IgA from patients withIgAN suggested an increased interaction of IgA with mesangialcells from patients with IgAN as compared with IgA fromhealthy individuals (23), although this still is controversial (24).Furthermore, after stimulation of mesangial cells with IgA frompatients with IgAN, the production of proinflammatory cyto-kines and chemokines was shown to be increased (25), possiblyinvolving (26,27) undergalactosylation of IgAN IgA (28).
The aim of our study was to characterize and compare themolecular composition and functional properties of monomericand polymeric serum IgA from patients with IgAN and controlsubjects. Therefore, we analyzed highly purified total serumIgA from patients and control subjects in a number of aspectsthat potentially are important in the pathogenesis of IgAN,including interaction with lectins and mesangial cells. The re-sults show clear functional differences between naturally oc-curring polymeric and monomeric serum IgA both for patientsand control subjects. The most obvious difference that wasnoted between IgA that was isolated from patients with IgANand from control subjects was an increased fraction of SIgA inpIgA from patients with IgAN. Furthermore, we demonstrate
that pIgA differs from monomeric IgA (mIgA) in its composi-tion of N-linked glycans.
Materials and MethodsParticipants
In this study, we obtained serum from 11 healthy volunteers and 11patients with primary IgAN (Table 1). All patients had biopsy-provenIgA nephropathy. None of these patients had clinical or laboratoryevidence of Henoch Schonlein purpura, systemic lupus erythematosus,or liver disease or received immunosuppressive therapy. A healthycontrol group was selected and matched for gender. The mean age ofthe control subjects was somewhat lower; however, we had no indica-tions that this affects the biochemical properties of IgA. Renal functionwas nonstable in five of the 11 patients, with serum creatinine rangingfrom 203 to 366 �mol/L. The study was approved by the ethicalcommittee of the Leiden University Medical Center. All individualsgave informed consent.
IgA PurificationIgA was precipitated from serum using (NH4)2SO4 at 50% saturation,
followed by affinity chromatography using HisA43 (mAb against hu-man IgA; provided by Dr. J. van den Born, Free University MedicalCenter, Amsterdam, The Netherlands) coupled to cyanogen bromide–activated Sepharose 4FF (Amersham, Roosendaal, The Netherlands). Acolumn of 15 ml was loaded with � globulin precipitate correspondingto 10 ml of serum, using 0.5� PBS as a running buffer, followed bywashing with 90 ml 0.5� PBS. Fractions of 3 ml were collected. Forremoval of nonspecifically bound proteins, the column was washedwith 70 ml of 1 M NaCl. Finally, bound IgA was eluted with 100 ml of0.1 M glycine/0.3 M NaCl (pH 2.8). Fractions were neutralized with 1M Tris (pH 8.0). Fractions that contained IgA, as assessed by ELISA(29), were pooled, dialyzed against PBS that contained 2 mM EDTA,and applied to a mixture of protein G/anti-human IgM (HB57)-BiogelA5 to remove residual contaminating IgG and IgM, followed by con-centration and size separation with a HiLoad 16/60 HR200 Superdexprep grade gel filtration column (120 ml; Amersham Pharmacia), run in50 mM NH4HCO3. Fractions were assessed for the presence of IgA andtotal protein. On basis of the protein profile, IgA-containing fractionswere pooled into pIgA (eluted at 44 to 50 ml) and mIgA (eluted at 50 to60 ml). These pools were analyzed for total IgA, IgA1, and IgA2 contentusing ELISA (29). The percentage of pIgA was quantified using calcu-lation of the area under the curve on the basis of the gel filtrationprofile.
MBL Binding ELISAMBL was purified from pooled plasma that was obtained from
healthy human donors, as described previously (12), resulting in a
Table 1. Clinical characteristics of the patients with IgAN and control subjectsa
Characteristic Patients with IgAN Control Subjects
n 11 11Male/female 8/3 9/2Mean age (yr) 48 33Serum creatinine (�mol/L; mean �range�) 190 (82 to 366) NDProteinuria (0 to 3�; median) 1� NDErythrocyturia (0 to 5�; median) 1� ND
aIgAN, IgA nephropathy; ND, not determined.
3530 Journal of the American Society of Nephrology J Am Soc Nephrol 17: 3529–3539, 2006

preparation of MBL in complex with its associated serine proteases(MASP). MBL binding was studied by ELISA, in which 5 �g/ml IgA orhuman serum albumin (HSA) as a control was coated, followed byblocking with PBS/BSA, incubation with MBL (2 �g/ml), and detectionof MBL binding as described (12). For inhibition experiments, MBL waspreincubated with MgEGTA (10 mM), d-mannose, or l-mannose (100mM; Sigma, St. Louis, MO).
Activation of C4 via the Lectin PathwayActivation of C4 by MBL-MASP complexes was measured as de-
scribed previously (12). In brief, incubation of MBL–MASP complexeson coated IgA was followed by incubation with purified C4 and detec-tion of C4 binding.
Helix Aspersa BindingIgA was assessed for binding to biotinylated Helix Aspersa (HAA;
Sigma) lectin, known to recognize terminal GalNAc. NUNC Maxisorpplates were coated with 5 �g/ml IgA or HSA as a control, in carbonatebuffer (pH 9.6), overnight at room temperature. After washing withPBS/Tween and blocking for 1 h with PBS/1% BSA, wells were incu-bated with 5 �g/ml biotinylated HAA in PBS/1% BSA/0.05% Tween.Binding of HAA was detected with horseradish peroxidase–conjugatedstreptavidin (Zymed, Invitrogen, Brech, The Netherlands). Enzymeactivity of horseradish peroxidase was developed using 2,2�-azino-di(3-thylbenstialozone) (ABTS; Sigma). The OD at 415 nm was measured.
SIgA ELISATo quantify SIgA levels in isolated IgA, we used a sandwich ELISA
specific for SIgA as described previously (17). Briefly, plates werecoated with a mAb against secretory component (NI194-4), followed byincubation with IgA and detection of IgA binding.
Glycosidase Treatment of IgADetection of undergalactosylated IgA with lectins could be hampered
by the presence of sialic acids. To get a clear and full picture of thegalactosylation, we treated IgA with neuraminidase and checked forbinding to HAA and MBL. IgA (5 �g/ml) and HSA were coated,followed by blocking and subsequent incubation with 100 mM sodiumacetate (pH 5.0), with or without 10 mU/ml neuraminidase from Ar-throbacter ureafaciens (Roche, Mannheim, Germany), for 3 h at 37°C.Subsequently, HAA and MBL binding were assessed as describedabove.
Cell CultureNormal human mesangial cells (Cambrex, Walkersville, MD) were
expanded according to the protocol provided by the manufacturer inmesangial cell basal medium with supplements (Cambrex). Experi-ments with normal human mesangial cells were performed in RPMIwith 10% FCS, 1% nonessential amino acids, 0.5% transferrin/insulin/selenium, 1% sodium pyruvate, and 1% l-glutamine (all purchasedfrom Life Technologies, Paisley, Scotland). AMC11, a spontaneouslygrowing mesangial cell line of adult human origin (provided by Prof.Holthofer, Helsinki, Finland), was cultured in DMEM with 10% FCS.Cells were harvested by trypsinization.
Flow CytometryCells were washed with FACS buffer (0.5� PBS that contained 1%
BSA/2.8% glucose/0.01% NaN3) and incubated with mIgA and pIgA.After incubation for 1 h at 4°C, cells were washed and incubated for 1 hat 4°C with monoclonal anti-IgA mAb 4E8 (IgG1) (29). IgA binding wasvisualized using PE-conjugated goat anti-mouse IgG1 (Southern Bio-
technology, Birmingham, AL), and fluorescence intensity was assessedby flow cytometry (FACSCalibur, Cell Quest Software; BD Biosciences,Alphen aan den Rijn, The Netherlands). Dead cells, identified by pro-pidium iodide uptake, were excluded from analysis.
Cytokine AnalysisProduction of IL-8 and monocyte chemoattractant protein-1 (MCP-1)
was measured in supernatants of cultured mesangial cells. Before stim-ulation, cells were transferred to 96-wells plates (Costar, Corning, NY)at a density of 15 � 103 cells per well and cultured overnight in culturemedium with 0.5% serum. Cells were cultured in the presence orabsence of mIgA and pIgA for 72 h, in concentrations as indicated. Theconcentration of IL-8 and MCP-1 in culture supernatants was measuredby ELISA as described previously (30,31).
N-Glycan AnalysisThe IgA heavy chains were isolated on SDS-PAGE under reducing
conditions and visualized by Coomassie staining. The N-glycans werereleased from these excised gel bands by PNGase F, labeled with thefluorophore 2-aminobenzamide, and analyzed by normal-phase HPLCwith exoglycosidase sequencing as described previously (32).
Identification of Gel Bands by Mass SpectrometryThe Coomassie-stained IgA heavy-chain bands from an SDS-PAGE
gel were excised and in-gel digested with trypsin (sequencing grade;Roche) as described previously (32).
Statistical AnalysesStatistical analysis was performed using the Mann-Whitney test and
the Wilcoxon signed rank test. The Spearman rank correlation coeffi-cient was used to analyze correlations. Differences in N-glycan com-position were evaluated using the t test. Differences were consideredstatistically significant at P � 0.05.
ResultspIgA Binds Better to Mesangial Cells and Induces MoreCytokine Production after Activation of Mesangial Cells
Human IgA was purified with an anti-IgA affinity column(Figure 1A). In the flow-through, no IgA was detectable,whereas IgA was eluted by acid elution. The fractions thatcontained IgA were pooled. Purified IgA was applied to a gelfiltration column (Figure 1B), and fractions that contained pIgAand mIgA were pooled as indicated.
To examine possible functional differences between mIgAand pIgA from patients with IgAN and control subjects, weinvestigated the binding of IgA to mesangial cells, as well as thecytokine response after stimulation. pIgA showed a 5.9-foldhigher binding to mesangial cells than mIgA (P � 0.0003), butthere was no difference between patients with IgAN and con-trol subjects (Figure 2A).
Supernatants of the cells that were stimulated with IgA for72 h were tested for production of the chemokines IL-8 andMCP-1. The IL-8 production was significantly higher after stim-ulation of mesangial cells with pIgA than after stimulation withmIgA (P � 0.010), whereas IL-8 production was undetectable incultures without IgA (Figure 2B). Furthermore, IL-8 productiontended to be higher upon stimulation with pIgA from patientswith IgAN compared with that from control subjects (P �
0.077). A significant correlation was observed between binding
J Am Soc Nephrol 17: 3529–3539, 2006 Differential Glycosylation of Polymeric and Monomeric IgA 3531
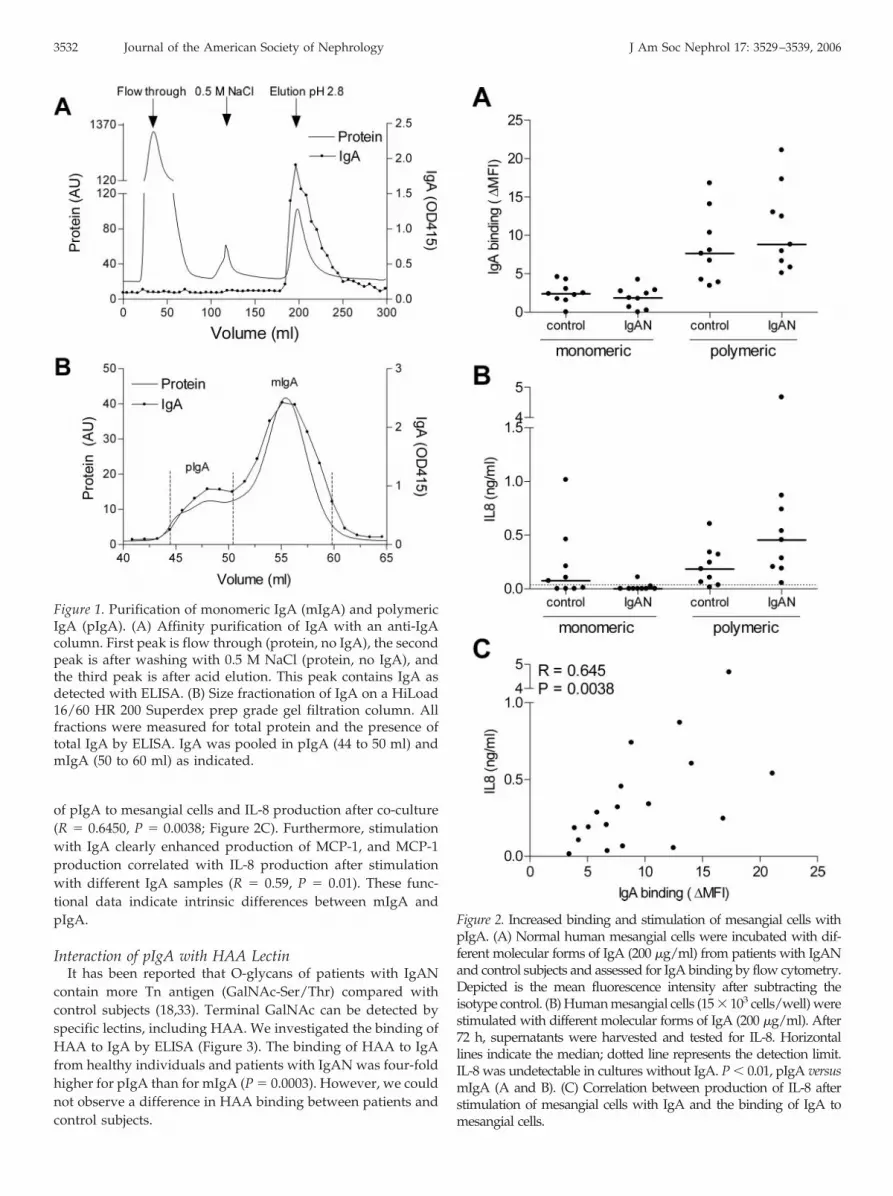
of pIgA to mesangial cells and IL-8 production after co-culture(R � 0.6450, P � 0.0038; Figure 2C). Furthermore, stimulationwith IgA clearly enhanced production of MCP-1, and MCP-1production correlated with IL-8 production after stimulationwith different IgA samples (R � 0.59, P � 0.01). These func-tional data indicate intrinsic differences between mIgA andpIgA.
Interaction of pIgA with HAA LectinIt has been reported that O-glycans of patients with IgAN
contain more Tn antigen (GalNAc-Ser/Thr) compared withcontrol subjects (18,33). Terminal GalNAc can be detected byspecific lectins, including HAA. We investigated the binding ofHAA to IgA by ELISA (Figure 3). The binding of HAA to IgAfrom healthy individuals and patients with IgAN was four-foldhigher for pIgA than for mIgA (P � 0.0003). However, we couldnot observe a difference in HAA binding between patients andcontrol subjects.
Figure 2. Increased binding and stimulation of mesangial cells withpIgA. (A) Normal human mesangial cells were incubated with dif-ferent molecular forms of IgA (200 �g/ml) from patients with IgANand control subjects and assessed for IgA binding by flow cytometry.Depicted is the mean fluorescence intensity after subtracting theisotype control. (B) Human mesangial cells (15 � 103 cells/well) werestimulated with different molecular forms of IgA (200 �g/ml). After72 h, supernatants were harvested and tested for IL-8. Horizontallines indicate the median; dotted line represents the detection limit.IL-8 was undetectable in cultures without IgA. P � 0.01, pIgA versusmIgA (A and B). (C) Correlation between production of IL-8 afterstimulation of mesangial cells with IgA and the binding of IgA tomesangial cells.
Figure 1. Purification of monomeric IgA (mIgA) and polymericIgA (pIgA). (A) Affinity purification of IgA with an anti-IgAcolumn. First peak is flow through (protein, no IgA), the secondpeak is after washing with 0.5 M NaCl (protein, no IgA), andthe third peak is after acid elution. This peak contains IgA asdetected with ELISA. (B) Size fractionation of IgA on a HiLoad16/60 HR 200 Superdex prep grade gel filtration column. Allfractions were measured for total protein and the presence oftotal IgA by ELISA. IgA was pooled in pIgA (44 to 50 ml) andmIgA (50 to 60 ml) as indicated.
3532 Journal of the American Society of Nephrology J Am Soc Nephrol 17: 3529–3539, 2006
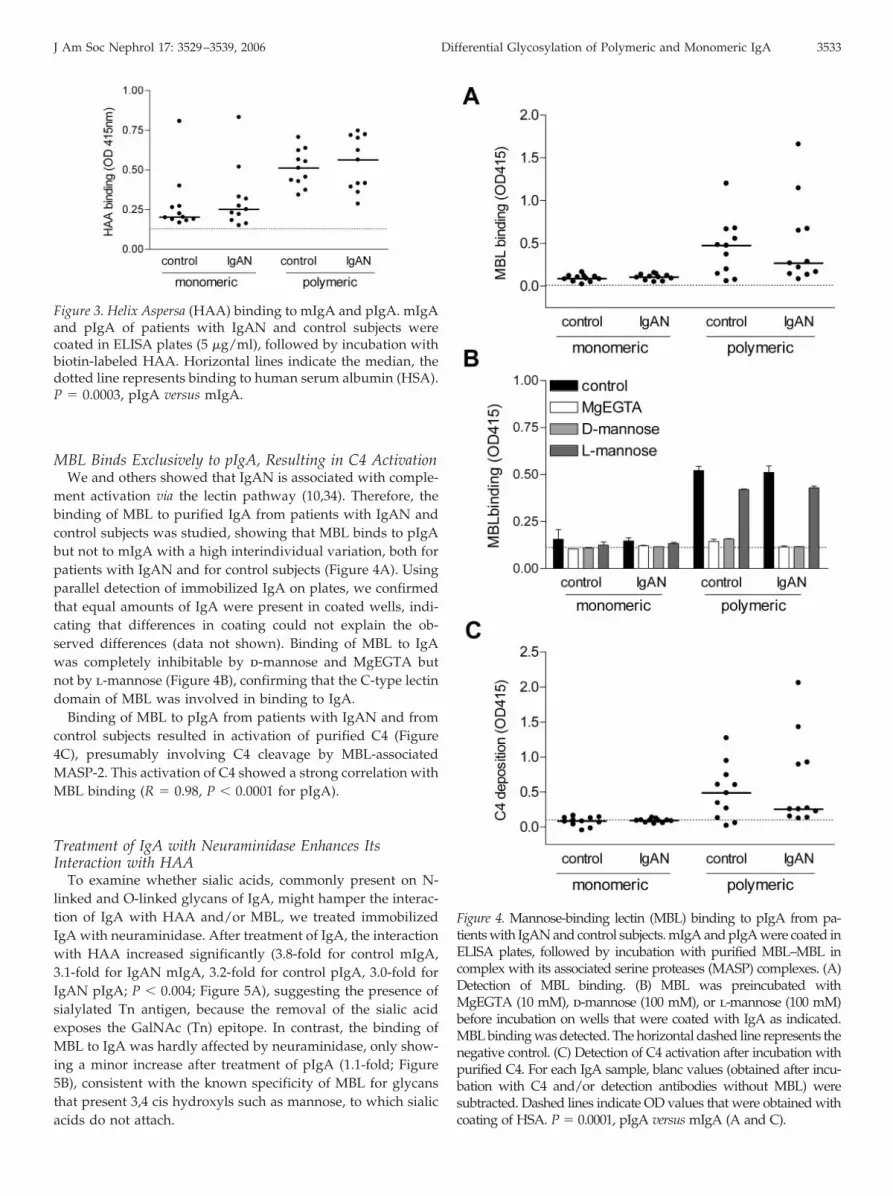
MBL Binds Exclusively to pIgA, Resulting in C4 ActivationWe and others showed that IgAN is associated with comple-
ment activation via the lectin pathway (10,34). Therefore, thebinding of MBL to purified IgA from patients with IgAN andcontrol subjects was studied, showing that MBL binds to pIgAbut not to mIgA with a high interindividual variation, both forpatients with IgAN and for control subjects (Figure 4A). Usingparallel detection of immobilized IgA on plates, we confirmedthat equal amounts of IgA were present in coated wells, indi-cating that differences in coating could not explain the ob-served differences (data not shown). Binding of MBL to IgAwas completely inhibitable by d-mannose and MgEGTA butnot by l-mannose (Figure 4B), confirming that the C-type lectindomain of MBL was involved in binding to IgA.
Binding of MBL to pIgA from patients with IgAN and fromcontrol subjects resulted in activation of purified C4 (Figure4C), presumably involving C4 cleavage by MBL-associatedMASP-2. This activation of C4 showed a strong correlation withMBL binding (R � 0.98, P � 0.0001 for pIgA).
Treatment of IgA with Neuraminidase Enhances ItsInteraction with HAA
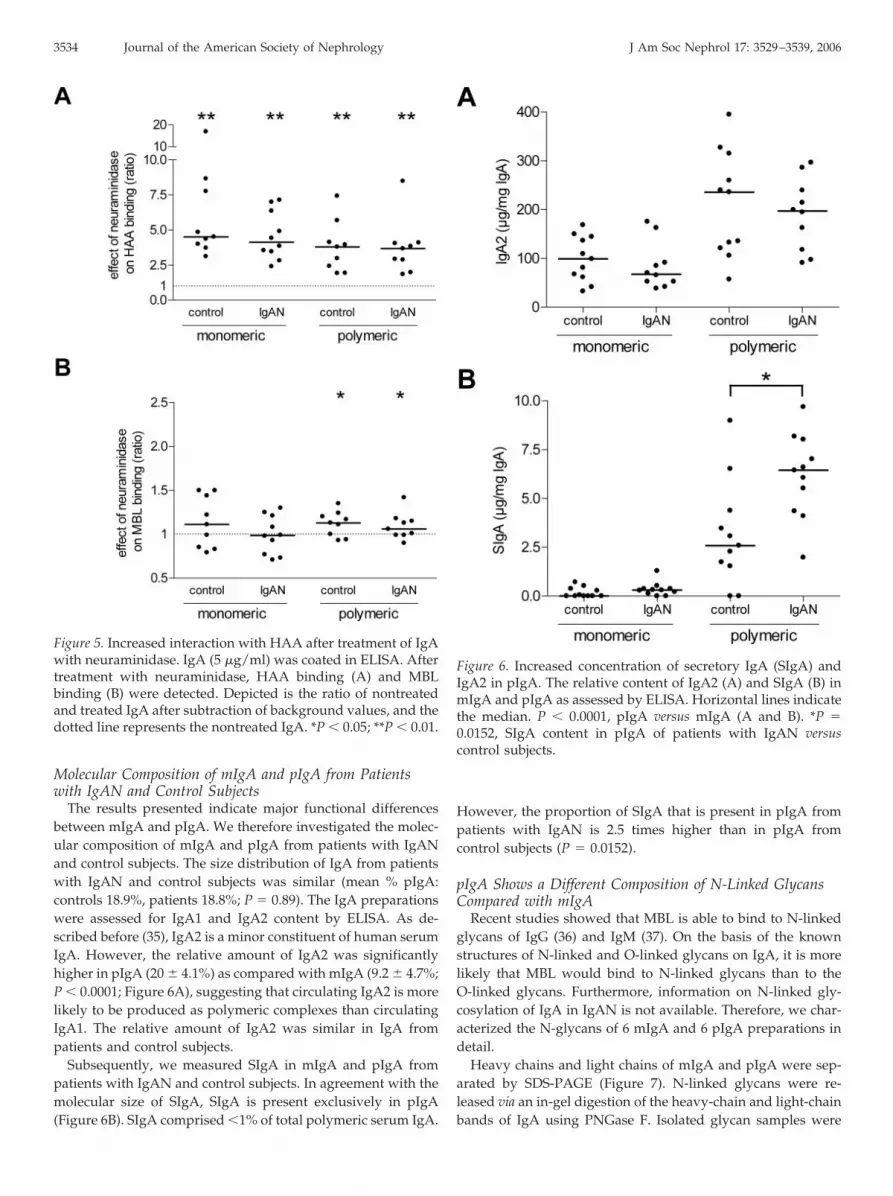
To examine whether sialic acids, commonly present on N-linked and O-linked glycans of IgA, might hamper the interac-tion of IgA with HAA and/or MBL, we treated immobilizedIgA with neuraminidase. After treatment of IgA, the interactionwith HAA increased significantly (3.8-fold for control mIgA,3.1-fold for IgAN mIgA, 3.2-fold for control pIgA, 3.0-fold forIgAN pIgA; P � 0.004; Figure 5A), suggesting the presence ofsialylated Tn antigen, because the removal of the sialic acidexposes the GalNAc (Tn) epitope. In contrast, the binding ofMBL to IgA was hardly affected by neuraminidase, only show-ing a minor increase after treatment of pIgA (1.1-fold; Figure5B), consistent with the known specificity of MBL for glycansthat present 3,4 cis hydroxyls such as mannose, to which sialicacids do not attach.
Figure 4. Mannose-binding lectin (MBL) binding to pIgA from pa-tients with IgAN and control subjects. mIgA and pIgA were coated inELISA plates, followed by incubation with purified MBL–MBL incomplex with its associated serine proteases (MASP) complexes. (A)Detection of MBL binding. (B) MBL was preincubated withMgEGTA (10 mM), d-mannose (100 mM), or l-mannose (100 mM)before incubation on wells that were coated with IgA as indicated.MBL binding was detected. The horizontal dashed line represents thenegative control. (C) Detection of C4 activation after incubation withpurified C4. For each IgA sample, blanc values (obtained after incu-bation with C4 and/or detection antibodies without MBL) weresubtracted. Dashed lines indicate OD values that were obtained withcoating of HSA. P � 0.0001, pIgA versus mIgA (A and C).
Figure 3. Helix Aspersa (HAA) binding to mIgA and pIgA. mIgAand pIgA of patients with IgAN and control subjects werecoated in ELISA plates (5 �g/ml), followed by incubation withbiotin-labeled HAA. Horizontal lines indicate the median, thedotted line represents binding to human serum albumin (HSA).P � 0.0003, pIgA versus mIgA.
J Am Soc Nephrol 17: 3529–3539, 2006 Differential Glycosylation of Polymeric and Monomeric IgA 3533
Molecular Composition of mIgA and pIgA from Patientswith IgAN and Control Subjects
The results presented indicate major functional differencesbetween mIgA and pIgA. We therefore investigated the molec-ular composition of mIgA and pIgA from patients with IgANand control subjects. The size distribution of IgA from patientswith IgAN and control subjects was similar (mean % pIgA:controls 18.9%, patients 18.8%; P � 0.89). The IgA preparationswere assessed for IgA1 and IgA2 content by ELISA. As de-scribed before (35), IgA2 is a minor constituent of human serumIgA. However, the relative amount of IgA2 was significantlyhigher in pIgA (20 � 4.1%) as compared with mIgA (9.2 � 4.7%;P � 0.0001; Figure 6A), suggesting that circulating IgA2 is morelikely to be produced as polymeric complexes than circulatingIgA1. The relative amount of IgA2 was similar in IgA frompatients and control subjects.
Subsequently, we measured SIgA in mIgA and pIgA frompatients with IgAN and control subjects. In agreement with themolecular size of SIgA, SIgA is present exclusively in pIgA(Figure 6B). SIgA comprised �1% of total polymeric serum IgA.
However, the proportion of SIgA that is present in pIgA frompatients with IgAN is 2.5 times higher than in pIgA fromcontrol subjects (P � 0.0152).
pIgA Shows a Different Composition of N-Linked GlycansCompared with mIgA
Recent studies showed that MBL is able to bind to N-linkedglycans of IgG (36) and IgM (37). On the basis of the knownstructures of N-linked and O-linked glycans on IgA, it is morelikely that MBL would bind to N-linked glycans than to theO-linked glycans. Furthermore, information on N-linked gly-cosylation of IgA in IgAN is not available. Therefore, we char-acterized the N-glycans of 6 mIgA and 6 pIgA preparations indetail.
Heavy chains and light chains of mIgA and pIgA were sep-arated by SDS-PAGE (Figure 7). N-linked glycans were re-leased via an in-gel digestion of the heavy-chain and light-chainbands of IgA using PNGase F. Isolated glycan samples were
Figure 5. Increased interaction with HAA after treatment of IgAwith neuraminidase. IgA (5 �g/ml) was coated in ELISA. Aftertreatment with neuraminidase, HAA binding (A) and MBLbinding (B) were detected. Depicted is the ratio of nontreatedand treated IgA after subtraction of background values, and thedotted line represents the nontreated IgA. *P � 0.05; **P � 0.01.
Figure 6. Increased concentration of secretory IgA (SIgA) andIgA2 in pIgA. The relative content of IgA2 (A) and SIgA (B) inmIgA and pIgA as assessed by ELISA. Horizontal lines indicatethe median. P � 0.0001, pIgA versus mIgA (A and B). *P �0.0152, SIgA content in pIgA of patients with IgAN versuscontrol subjects.
3534 Journal of the American Society of Nephrology J Am Soc Nephrol 17: 3529–3539, 2006
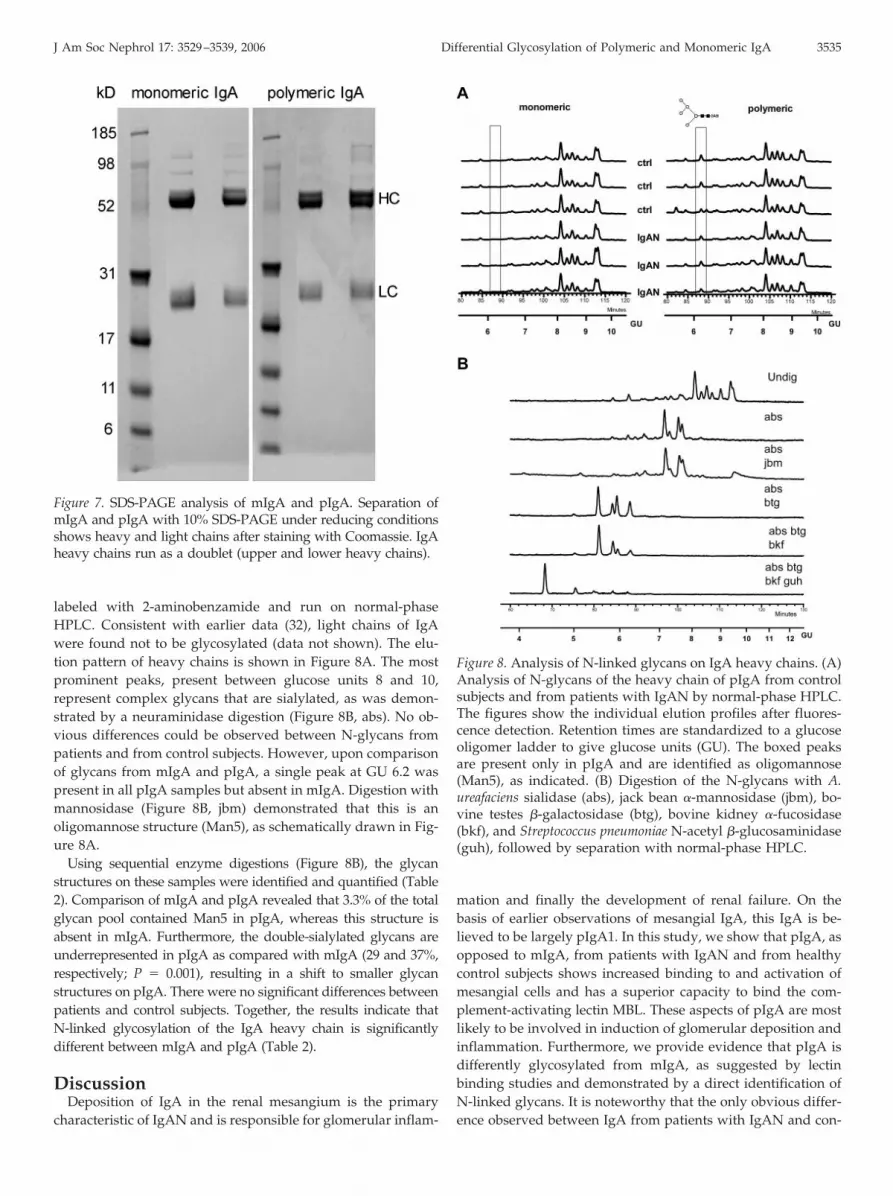
labeled with 2-aminobenzamide and run on normal-phaseHPLC. Consistent with earlier data (32), light chains of IgAwere found not to be glycosylated (data not shown). The elu-tion pattern of heavy chains is shown in Figure 8A. The mostprominent peaks, present between glucose units 8 and 10,represent complex glycans that are sialylated, as was demon-strated by a neuraminidase digestion (Figure 8B, abs). No ob-vious differences could be observed between N-glycans frompatients and from control subjects. However, upon comparisonof glycans from mIgA and pIgA, a single peak at GU 6.2 waspresent in all pIgA samples but absent in mIgA. Digestion withmannosidase (Figure 8B, jbm) demonstrated that this is anoligomannose structure (Man5), as schematically drawn in Fig-ure 8A.
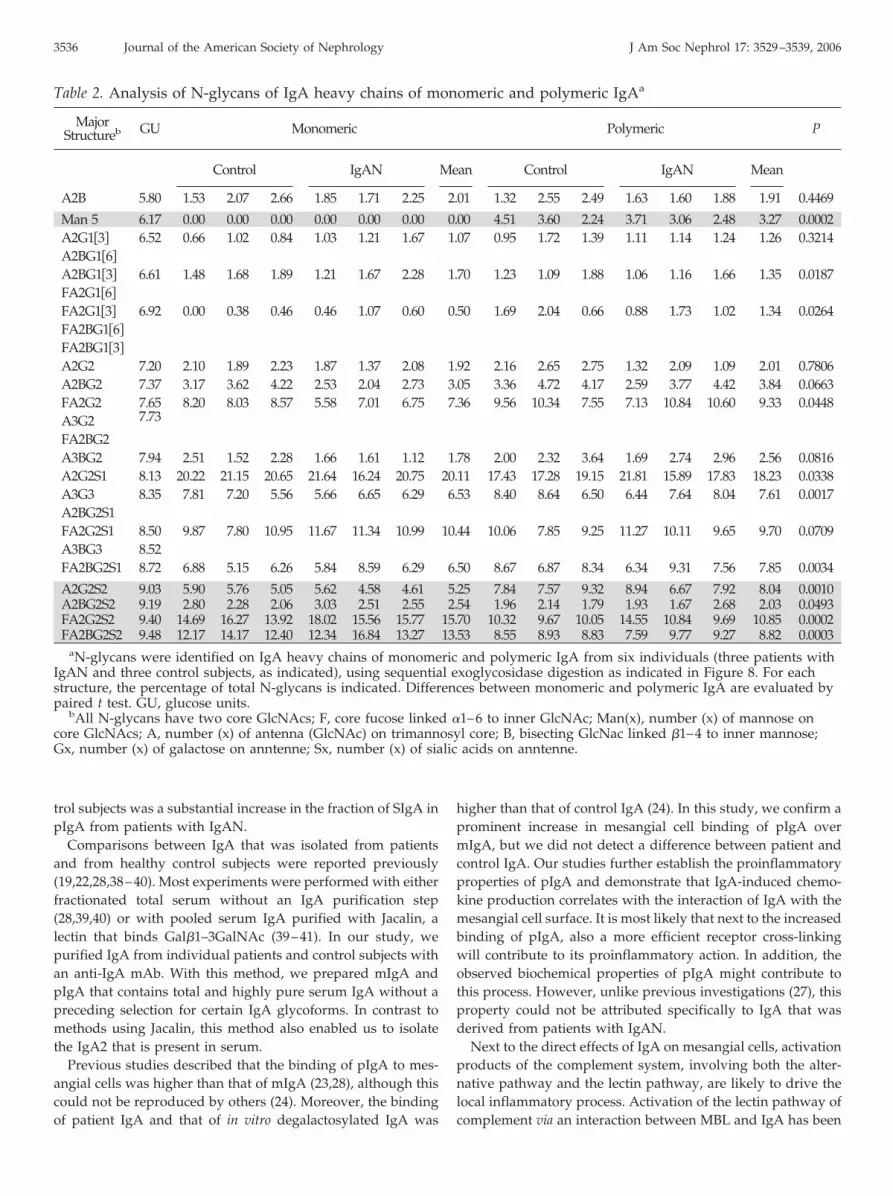
Using sequential enzyme digestions (Figure 8B), the glycanstructures on these samples were identified and quantified (Table2). Comparison of mIgA and pIgA revealed that 3.3% of the totalglycan pool contained Man5 in pIgA, whereas this structure isabsent in mIgA. Furthermore, the double-sialylated glycans areunderrepresented in pIgA as compared with mIgA (29 and 37%,respectively; P � 0.001), resulting in a shift to smaller glycanstructures on pIgA. There were no significant differences betweenpatients and control subjects. Together, the results indicate thatN-linked glycosylation of the IgA heavy chain is significantlydifferent between mIgA and pIgA (Table 2).
DiscussionDeposition of IgA in the renal mesangium is the primary
characteristic of IgAN and is responsible for glomerular inflam-
mation and finally the development of renal failure. On thebasis of earlier observations of mesangial IgA, this IgA is be-lieved to be largely pIgA1. In this study, we show that pIgA, asopposed to mIgA, from patients with IgAN and from healthycontrol subjects shows increased binding to and activation ofmesangial cells and has a superior capacity to bind the com-plement-activating lectin MBL. These aspects of pIgA are mostlikely to be involved in induction of glomerular deposition andinflammation. Furthermore, we provide evidence that pIgA isdifferently glycosylated from mIgA, as suggested by lectinbinding studies and demonstrated by a direct identification ofN-linked glycans. It is noteworthy that the only obvious differ-ence observed between IgA from patients with IgAN and con-
Figure 7. SDS-PAGE analysis of mIgA and pIgA. Separation ofmIgA and pIgA with 10% SDS-PAGE under reducing conditionsshows heavy and light chains after staining with Coomassie. IgAheavy chains run as a doublet (upper and lower heavy chains).
Figure 8. Analysis of N-linked glycans on IgA heavy chains. (A)Analysis of N-glycans of the heavy chain of pIgA from controlsubjects and from patients with IgAN by normal-phase HPLC.The figures show the individual elution profiles after fluores-cence detection. Retention times are standardized to a glucoseoligomer ladder to give glucose units (GU). The boxed peaksare present only in pIgA and are identified as oligomannose(Man5), as indicated. (B) Digestion of the N-glycans with A.ureafaciens sialidase (abs), jack bean �-mannosidase (jbm), bo-vine testes �-galactosidase (btg), bovine kidney �-fucosidase(bkf), and Streptococcus pneumoniae N-acetyl �-glucosaminidase(guh), followed by separation with normal-phase HPLC.
J Am Soc Nephrol 17: 3529–3539, 2006 Differential Glycosylation of Polymeric and Monomeric IgA 3535
trol subjects was a substantial increase in the fraction of SIgA inpIgA from patients with IgAN.
Comparisons between IgA that was isolated from patientsand from healthy control subjects were reported previously(19,22,28,38–40). Most experiments were performed with eitherfractionated total serum without an IgA purification step(28,39,40) or with pooled serum IgA purified with Jacalin, alectin that binds Gal�1–3GalNAc (39–41). In our study, wepurified IgA from individual patients and control subjects withan anti-IgA mAb. With this method, we prepared mIgA andpIgA that contains total and highly pure serum IgA without apreceding selection for certain IgA glycoforms. In contrast tomethods using Jacalin, this method also enabled us to isolatethe IgA2 that is present in serum.
Previous studies described that the binding of pIgA to mes-angial cells was higher than that of mIgA (23,28), although thiscould not be reproduced by others (24). Moreover, the bindingof patient IgA and that of in vitro degalactosylated IgA was
higher than that of control IgA (24). In this study, we confirm aprominent increase in mesangial cell binding of pIgA overmIgA, but we did not detect a difference between patient andcontrol IgA. Our studies further establish the proinflammatoryproperties of pIgA and demonstrate that IgA-induced chemo-kine production correlates with the interaction of IgA with themesangial cell surface. It is most likely that next to the increasedbinding of pIgA, also a more efficient receptor cross-linkingwill contribute to its proinflammatory action. In addition, theobserved biochemical properties of pIgA might contribute tothis process. However, unlike previous investigations (27), thisproperty could not be attributed specifically to IgA that wasderived from patients with IgAN.
Next to the direct effects of IgA on mesangial cells, activationproducts of the complement system, involving both the alter-native pathway and the lectin pathway, are likely to drive thelocal inflammatory process. Activation of the lectin pathway ofcomplement via an interaction between MBL and IgA has been
Table 2. Analysis of N-glycans of IgA heavy chains of monomeric and polymeric IgAa
MajorStructureb GU Monomeric Polymeric P
Control IgAN Mean Control IgAN Mean
A2B 5.80 1.53 2.07 2.66 1.85 1.71 2.25 2.01 1.32 2.55 2.49 1.63 1.60 1.88 1.91 0.4469
Man 5 6.17 0.00 0.00 0.00 0.00 0.00 0.00 0.00 4.51 3.60 2.24 3.71 3.06 2.48 3.27 0.0002A2G1�3� 6.52 0.66 1.02 0.84 1.03 1.21 1.67 1.07 0.95 1.72 1.39 1.11 1.14 1.24 1.26 0.3214A2BG1�6�
A2BG1�3� 6.61 1.48 1.68 1.89 1.21 1.67 2.28 1.70 1.23 1.09 1.88 1.06 1.16 1.66 1.35 0.0187FA2G1�6�
FA2G1�3� 6.92 0.00 0.38 0.46 0.46 1.07 0.60 0.50 1.69 2.04 0.66 0.88 1.73 1.02 1.34 0.0264FA2BG1�6�
FA2BG1�3�
A2G2 7.20 2.10 1.89 2.23 1.87 1.37 2.08 1.92 2.16 2.65 2.75 1.32 2.09 1.09 2.01 0.7806A2BG2 7.37 3.17 3.62 4.22 2.53 2.04 2.73 3.05 3.36 4.72 4.17 2.59 3.77 4.42 3.84 0.0663FA2G2 7.65
7.738.20 8.03 8.57 5.58 7.01 6.75 7.36 9.56 10.34 7.55 7.13 10.84 10.60 9.33 0.0448
A3G2FA2BG2A3BG2 7.94 2.51 1.52 2.28 1.66 1.61 1.12 1.78 2.00 2.32 3.64 1.69 2.74 2.96 2.56 0.0816A2G2S1 8.13 20.22 21.15 20.65 21.64 16.24 20.75 20.11 17.43 17.28 19.15 21.81 15.89 17.83 18.23 0.0338A3G3 8.35 7.81 7.20 5.56 5.66 6.65 6.29 6.53 8.40 8.64 6.50 6.44 7.64 8.04 7.61 0.0017A2BG2S1FA2G2S1 8.50 9.87 7.80 10.95 11.67 11.34 10.99 10.44 10.06 7.85 9.25 11.27 10.11 9.65 9.70 0.0709A3BG3 8.52FA2BG2S1 8.72 6.88 5.15 6.26 5.84 8.59 6.29 6.50 8.67 6.87 8.34 6.34 9.31 7.56 7.85 0.0034
A2G2S2 9.03 5.90 5.76 5.05 5.62 4.58 4.61 5.25 7.84 7.57 9.32 8.94 6.67 7.92 8.04 0.0010A2BG2S2 9.19 2.80 2.28 2.06 3.03 2.51 2.55 2.54 1.96 2.14 1.79 1.93 1.67 2.68 2.03 0.0493FA2G2S2 9.40 14.69 16.27 13.92 18.02 15.56 15.77 15.70 10.32 9.67 10.05 14.55 10.84 9.69 10.85 0.0002FA2BG2S2 9.48 12.17 14.17 12.40 12.34 16.84 13.27 13.53 8.55 8.93 8.83 7.59 9.77 9.27 8.82 0.0003
aN-glycans were identified on IgA heavy chains of monomeric and polymeric IgA from six individuals (three patients withIgAN and three control subjects, as indicated), using sequential exoglycosidase digestion as indicated in Figure 8. For eachstructure, the percentage of total N-glycans is indicated. Differences between monomeric and polymeric IgA are evaluated bypaired t test. GU, glucose units.
bAll N-glycans have two core GlcNAcs; F, core fucose linked �1–6 to inner GlcNAc; Man(x), number (x) of mannose oncore GlcNAcs; A, number (x) of antenna (GlcNAc) on trimannosyl core; B, bisecting GlcNac linked �1–4 to inner mannose;Gx, number (x) of galactose on anntenne; Sx, number (x) of sialic acids on anntenne.
3536 Journal of the American Society of Nephrology J Am Soc Nephrol 17: 3529–3539, 2006

shown before (12), and in this study, we confirm and extendthese data by showing that binding of MBL is a commonfeature of pIgA but not mIgA that was isolated from differentdonors. Because ligand recognition by MBL requires multiplypresented carbohydrates, MBL binding could be favored by thestructure of pIgA. Binding of MBL leads to activation of C4presumably via activation of the C4-cleaving enzyme MASP-2.In a healthy situation, the binding of MBL to pIgA could beinvolved in host defense. However, in IgAN, lectin pathwayactivation via pIgA is unfavorable (10).
Many studies on IgA from patients with IgAN focused onglycosylation. IgA is glycosylated extensively, via both N-link-ages (IgA1 and IgA2) and O-linkages (IgA1) (21). It was ob-served consistently that serum IgA from patients with IgANcontains smaller O-linked glycans, with less sialylation andgalactosylation, than IgA from healthy control subjects (18).Previous investigations suggested that this predominantly wasthe case for mIgA (40). Our experiments using HAA, a lectinthat is used commonly to detect terminal GalNAc on nongalac-tosylated O-linked glycans, suggested the presence of terminalGalNAc (Tn antigen) predominantly on pIgA, from both pa-tients and control subjects. This is in agreement with a previousstudy that showed reactivity of HAA with high molecularweight serum proteins (28) and with data that were providedby Leung et al. (40). Binding of HAA was strongly increased byneuraminidase treatment, suggesting a high frequency of non-galactosylated O-linked glycans on IgA that expose terminalGalNAc after enzymatic removal of sialic acid.
A detailed quantitative analysis of N-linked glycans on IgAheavy chains of mIgA and pIgA revealed several significantdifferences between these molecular forms of IgA. In this re-spect, pIgA consistently contained an oligomannose structurethat was undetectable on all mIgA preparations and showedsignificantly less glycans with two terminal sialic acid residues.Recent studies showed that MBL can bind to certain glycoformsof human IgM, involving GlcNAc-terminated glycans and oli-gomannose structures on the IgM heavy chains (37). Therefore,the presence of specific glycans on pIgA but not mIgA alsomight be involved in its recognition by MBL. At present, it isunknown whether a specific glycosylation pattern of the heavychain of pIgA is involved in the polymerization of IgA and/orwhether this merely is related to the conditions that are presentduring production of the different forms of IgA. Earlier studiesfrom our group indicated that polymeric serum IgA containsdimeric IgA linked with J chain, as well as complexes of mIgAlinked via other mechanisms (42). In the first case, polymeriza-tion takes place in the B cell, and the presence of oligomannose,which is a premature glycan structure, might suggest the en-doplasmic reticulum as a possible location for polymerization,which prevents further synthesis of the glycan structure bysteric hindrance. In the latter case, polymerization might takeplace outside the B cell.
Part of the observed differences in glycosylation betweenmIgA and pIgA also might be explained by the presence ofSIgA in the polymeric fraction of serum IgA, because the SIgAheavy-chain N-glycosylation is very different from that of mo-nomeric serum IgA (32). SIgA has approximately 8% oligom-
annose structures and 60% glycans with exposed GlcNAc with�15% for all glycans sialylated (32), compared with monomericserum IgA, in which most of the glycans are sialylated. How-ever, SIgA comprises only �1% of total polymeric serum IgA.
We observed that polymeric serum IgA contains more IgA2than mIgA. This could be because IgA2 polymerizes moreeasily than IgA1 or that IgA2-producing B cells preferentiallysecrete pIgA. Bone marrow–derived IgA as present in serumlargely is monomeric and of the IgA1 subclass, whereas muco-sal IgA largely is polymeric, containing J chain and secretorycomponent (43), and contains a substantial fraction of IgA2 (35).Therefore, an increased fraction of IgA2 and the presence ofSIgA in circulating pIgA may suggest its production by themucosal immune system. Quantitative measurement of thepresence of SIgA in polymeric serum IgA suggests that only aminor part of pIgA contains a secretory component. We hy-pothesize that this SIgA is derived from the mucosal surface.Circulating dimeric IgA without a secretory component couldbe produced partially in mucosal lymphoid tissue and directlytransported toward the circulation.
Although SIgA requires transepithelial transport for the at-tachment of a secretory component to dimeric IgA, the presenceof low concentrations of circulating SIgA has been describedbefore (17,44,45). Moreover, increased serum levels of SIgAhave been reported in various diseases (17,46–48). In contrastto previous studies, we now determined the SIgA concentrationin highly purified polymeric serum IgA. Our data demonstratea clear relative increase in SIgA in pIgA from patients withIgAN compared with control subjects. We recently reported apreferential interaction of SIgA with mesangial cells andshowed glomerular accumulation of SIgA in IgAN (17). There-fore, these results further support a role for SIgA in the patho-genesis of IgAN.
Taken together, the presented data suggest that a part ofcirculating pIgA has a mucosal origin. There is accumulatingevidence that the pathogenesis of IgAN is related to aberrantproduction of IgA. In this respect, in vivo studies indicated thatpatients with IgAN have a disturbed mucosal immune re-sponse, which was restricted to production of antibodies of theIgA1 subclass (49). Our observation that SIgA is increased inthe pIgA fraction of patients with IgAN further supports a rolefor abnormal mucosal immunity. Because IgAN is a slowlyprogressive disease, it is well conceivable that only a minorsubfraction of circulating IgA in patients with IgAN is abnor-mal and that this IgA gradually accumulates in the mesangialarea. We hypothesize that this abnormal IgA is derived at leastpartially from the mucosal immune system. Because our datastrongly indicate that large-sized IgA is especially able to in-teract with mesangial cells and to induce complement activa-tion, the gradual deposition of such proinflammatory IgA even-tually may lead to renal disease.
AcknowledgmentsThis work was financially supported by the Dutch Kidney Founda-
tion (C99.1822 to B.D.O.; PC95 to A.R.).We thank A. de Wilde (Leiden, The Netherlands) for technical assis-
tance, Dr. J. van der Born (Amsterdam, The Netherlands) for the supply
J Am Soc Nephrol 17: 3529–3539, 2006 Differential Glycosylation of Polymeric and Monomeric IgA 3537

of valuable reagents, and E. Shillington (Oxford, UK) for assistancewith glycan sequencing.
References1. Berger J, Hinglais N: Intercapillary deposits of IgA-IgG [in
French]. J Urol Nephrol (Paris) 74: 694–695, 19682. Donadio JV, Grande JP: IgA nephropathy. N Engl J Med
347: 738–748, 20023. Tomino Y, Sakai H, Miura M, Endoh M, Nomoto Y: De-
tection of polymeric IgA in glomeruli from patients withIgA nephropathy. Clin Exp Immunol 49: 419–425, 1982
4. Novak J, Julian BA, Tomana M, Mestecky J: Progress inmolecular and genetic studies of IgA nephropathy. J ClinImmunol 21: 310–327, 2001
5. Floege J, Feehally J: IgA nephropathy: Recent develop-ments. J Am Soc Nephrol 11: 2395–2403, 2000
6. van der Boog PJ, van Kooten C, de Fijter JW, Daha MR:Role of macromolecular IgA in IgA nephropathy. KidneyInt 67: 813–821, 2005
7. Moura IC, Centelles MN, Arcos-Fajardo M, Malheiros DM,Collawn JF, Cooper MD, Monteiro RC: Identification of thetransferrin receptor as a novel immunoglobulin (Ig)A1 re-ceptor and its enhanced expression on mesangial cells inIgA nephropathy. J Exp Med 194: 417–425, 2001
8. McDonald KJ, Cameron AJ, Allen JM, Jardine AG: Expres-sion of Fc alpha/mu receptor by human mesangial cells: Acandidate receptor for immune complex deposition in IgAnephropathy. Biochem Biophys Res Commun 290: 438–442,2002
9. van den Dobbelsteen ME, van der Woude FJ, SchroeijersWE, van den Wall Bake AW, Van Es LA, Daha MR: Bind-ing of dimeric and polymeric IgA to rat renal mesangialcells enhances the release of interleukin 6. Kidney Int 46:512–519, 1994
10. Roos A, Rastaldi MP, Calvaresi N, Oortwijn BD, Schlag-wein N, van Gijlswijk-Janssen DJ, Stahl GL, Matsushita M,Fujita T, van Kooten C, Daha MR: Glomerular activation ofthe lectin pathway of complement in IgA nephropathy isassociated with more severe renal disease. J Am Soc Nephrol17: 1724–1734, 2006
11. Endo M, Ohi H, Ohsawa I, Fujita T, Matsushita M, Fujita T:Glomerular deposition of mannose-binding lectin (MBL)indicates a novel mechanism of complement activation inIgA nephropathy. Nephrol Dial Transplant 13: 1984–1990,1998
12. Roos A, Bouwman LH, Gijlswijk-Janssen DJ, Faber-KrolMC, Stahl GL, Daha MR: Human IgA activates the com-plement system via the mannan-binding lectin pathway.J Immunol 167: 2861–2868, 2001
13. Berger J, Yaneva H, Nabarra B, Barbanel C: Recurrence ofmesangial deposition of IgA after renal transplantation.Kidney Int 7: 232–241, 1975
14. Sanfilippo F, Croker BP, Bollinger RR: Fate of four cadav-eric donor renal allografts with mesangial IgA deposits.Transplantation 33: 370–376, 1982
15. Schena FP, Pastore A, Ludovico N, Sinico RA, Benuzzi S,Montinaro V: Increased serum levels of IgA1-IgG immunecomplexes and anti-F(ab�)2 antibodies in patients with pri-mary IgA nephropathy. Clin Exp Immunol 77: 15–20, 1989
16. van der Boog PJ, van Kooten C, van Seggelen A, Mallat M,Klar-Mohamad N, de Fijter JW, Daha MR: An increased
polymeric IgA level is not a prognostic marker for progres-sive IgA nephropathy. Nephrol Dial Transplant 19: 2487–2493, 2004
17. Oortwijn BD, van der Boog PJ, Roos A, van der Geest RN,de Fijter JW, Daha MR, van Kooten C: A pathogenic rolefor secretory IgA in IgA nephropathy. Kidney Int 69: 1131–1138, 2006
18. Allen AC, Bailey EM, Barratt J, Buck KS, Feehally J: Anal-ysis of IgA1 O-glycans in IgA nephropathy by fluoro-phore-assisted carbohydrate electrophoresis. J Am SocNephrol 10: 1763–1771, 1999
19. Coppo R, Amore A: Aberrant glycosylation in IgA ne-phropathy (IgAN). Kidney Int 65: 1544–1547, 2004
20. Tomana M, Novak J, Julian BA, Matousovic K, Konecny K,Mestecky J: Circulating immune complexes in IgA ne-phropathy consist of IgA1 with galactose-deficient hingeregion and antiglycan antibodies. J Clin Invest 104: 73–81,1999
21. Mestecky J, Moro B, Underdown BJ: Mucosal immuno-globulins. In: Mucosal Immunology, 2nd ed., edited by OgraPL, Mestecky J, Lamm ME, Strober W, Bienenstock J,McGhee JR, San Diego, Academic Press, 1999, pp 133–152
22. Allen AC, Harper SJ, Feehally J: Galactosylation of N- andO-linked carbohydrate moieties of IgA1 and IgG in IgAnephropathy. Clin Exp Immunol 100: 470–474, 1995
23. Leung JC, Tang SC, Lam MF, Chan TM, Lai KN: Charge-dependent binding of polymeric IgA1 to human mesangialcells in IgA nephropathy. Kidney Int 59: 277–285, 2001
24. Wang Y, Zhao MH, Zhang YK, Li XM, Wang HY: Bindingcapacity and pathophysiological effects of IgA1 from pa-tients with IgA nephropathy on human glomerular mes-angial cells. Clin Exp Immunol 136: 168–175, 2004
25. Gomez-Guerrero C, Lopez-Armada MJ, Gonzalez E, EgidoJ: Soluble IgA and IgG aggregates are catabolized by cul-tured rat mesangial cells and induce production of TNF-alpha and IL-6, and proliferation. J Immunol 153: 5247–5255, 1994
26. Duque N, Gomez-Guerrero C, Egido J: Interaction of IgAwith Fc alpha receptors of human mesangial cells activatestranscription factor nuclear factor-kappa B and inducesexpression and synthesis of monocyte chemoattractantprotein-1, IL-8, and IFN-inducible protein 10. J Immunol159: 3474–3482, 1997
27. Leung JC, Tang SC, Chan LY, Tsang AW, Lan HY, Lai KN:Polymeric IgA increases the synthesis of macrophage mi-gration inhibitory factor by human mesangial cells in IgAnephropathy. Nephrol Dial Transplant 18: 36–45, 2003
28. Novak J, Vu HL, Novak L, Julian BA, Mestecky J, TomanaM: Interactions of human mesangial cells with IgA andIgA-containing immune complexes. Kidney Int 62: 465–475,2002
29. de Fijter JW, van den Wall Bake AW, Braam CA, Van EsLA, Daha MR: Immunoglobulin A subclass measurementin serum and saliva: Sensitivity of detection of dimericIgA2 in ELISA depends on the antibody used. J ImmunolMethods 187: 221–232, 1995
30. van Kooten C, Gerritsma JS, Paape ME, Van Es LA,Banchereau J, Daha MR: Possible role for CD40-CD40L inthe regulation of interstitial infiltration in the kidney. Kid-ney Int 51: 711–721, 1997
31. Taekema-Roelvink ME, Kooten C, Kooij SV, Heemskerk E,Daha MR: Proteinase 3 enhances endothelial monocyte
3538 Journal of the American Society of Nephrology J Am Soc Nephrol 17: 3529–3539, 2006

chemoattractant protein-1 production and induces in-creased adhesion of neutrophils to endothelial cells byupregulating intercellular cell adhesion molecule-1. J AmSoc Nephrol 12: 932–940, 2001
32. Royle L, Roos A, Harvey DJ, Wormald MR, Gijlswijk-Janssen D, Redwan E, Wilson IA, Daha MR, Dwek RA,Rudd PM: Secretory IgA N- and O-glycans provide a linkbetween the innate and adaptive immune systems. J BiolChem 278: 20140–20153, 2003
33. Odani H, Hiki Y, Takahashi M, Nishimoto A, Yasuda Y,Iwase H, Shinzato T, Maeda K: Direct evidence for de-creased sialylation and galactosylation of human serumIgA1 Fc O-glycosylated hinge peptides in IgA nephropathyby mass spectrometry. Biochem Biophys Res Commun 271:268–274, 2000
34. Matsuda M, Shikata K, Wada J, Sugimoto H, Shikata Y,Kawasaki T, Makino H: Deposition of mannan bindingprotein and mannan binding protein-mediated comple-ment activation in the glomeruli of patients with IgA ne-phropathy. Nephron 80: 408–413, 1998
35. Van Es LA, de Fijter JW, Daha MR: Pathogenesis of IgAnephropathy. Nephrology 3: 3–12, 1997
36. Malhotra R, Wormald MR, Rudd PM, Fischer PB, DwekRA, Sim RB: Glycosylation changes of IgG associated withrheumatoid arthritis can activate complement via the man-nose-binding protein. Nat Med 1: 237–243, 1995
37. Arnold JN, Wormald MR, Suter DM, Radcliffe CM, HarveyDJ, Dwek RA, Rudd PM, Sim RB: Human serum IgMglycosylation: Identification of glycoforms that can bind tomannan-binding lectin. J Biol Chem 280: 29080–29087, 2005
38. Julian BA, Novak J: IgA nephropathy: An update. CurrOpin Nephrol Hypertens 13: 171–179, 2004
39. Amore A, Cirina P, Conti G, Brusa P, Peruzzi L, Coppo R:Glycosylation of circulating IgA in patients with IgA ne-phropathy modulates proliferation and apoptosis of mes-angial cells. J Am Soc Nephrol 12: 1862–1871, 2001
40. Leung JC, Poon PY, Lai KN: Increased sialylation of poly-
meric immunoglobulin A1: Mechanism of selective glo-merular deposition in immunoglobulin A nephropathy?J Lab Clin Med 133: 152–160, 1999
41. Kabir S: Jacalin: A jackfruit (Artocarpus heterophyllus) seed-derived lectin of versatile applications in immunobiologi-cal research. J Immunol Methods 212: 193–211, 1998
42. van der Boog PJ, van Zandbergen G, de Fijter JW, Klar-Mohamad N, van Seggelen A, Brandtzaeg P, Daha MR,van Kooten C: Fc alpha RI/CD89 circulates in humanserum covalently linked to IgA in a polymeric state. J Im-munol 168: 1252–1258, 2002
43. Delacroix DL, Dive C, Rambaud JC, Vaerman JP: IgA sub-classes in various secretions and in serum. Immunology 47:383–385, 1982
44. Thompson RA, Asquith P, Cooke WT: Secretory IgA in theserum. Lancet 2: 517–519, 1969
45. Delacroix DL, Vaerman JP: A solid phase, direct competi-tion, radioimmunoassay for quantitation of secretory IgAin human serum. J Immunol Methods 40: 345–358, 1981
46. Mole CM, Renoult E, Bene MC, Seilles E, Kessler M, Re-villard JP, Faure GC: Serum secretory IgA and IgM andfree secretory component in IgA nephropathy. Nephron 71:75–78, 1995
47. Seilles E, Rossel M, Vuitton DA, Mercier M, Njoya O,Capron JP, Nalpas B, Gibey R, Revillard JP: Serum secre-tory IgA and secretory component in patients with non-cirrhotic alcoholic liver diseases. J Hepatol 22: 278–285, 1995
48. Rostoker G, Terzidis H, Petit-Phar M, Meillet D, Lang P,Dubert JM, Lagrue G, Weil B: Secretory IgA are elevated inboth saliva and serum of patients with various types ofprimary glomerulonephritis. Clin Exp Immunol 90: 305–311,1992
49. de Fijter JW, Eijgenraam JW, Braam CA, Holmgren J, DahaMR, Van Es LA, van den Wall Bake AW: Deficient IgA1immune response to nasal cholera toxin subunit B in pri-mary IgA nephropathy. Kidney Int 50: 952–961, 1996
J Am Soc Nephrol 17: 3529–3539, 2006 Differential Glycosylation of Polymeric and Monomeric IgA 3539