monomeric aβ1–42 and rage: key players in neuronal differentiation
TRANSCRIPT

lable at ScienceDirect
Neurobiology of Aging 35 (2014) 1301e1308
Contents lists avai
Neurobiology of Aging
journal homepage: www.elsevier .com/locate/neuaging
Monomeric Ab1e42 and RAGE: key players in neuronaldifferentiation
Sabrina Piras a, Anna L. Furfaro a, Alessandra Piccini a, Mario Passalacqua a,Roberta Borghi b,c, Enrico Carminati b,c, Alessia Parodi d, Laura Colombo e,Mario Salmona e, Maria A. Pronzato a, Umberto M. Marinari a, Massimo Tabaton b,Mariapaola Nitti a,*aDepartment of Experimental Medicine, University of Genoa, Genoa, ItalybDepartment of Internal Medicine and Medical Specialities, University of Genoa, Genoa, Italyc IRCCS Azienda Ospedaliera Universitaria San MartinodIST, Istituto Nazionale Ricerca sul Cancro, Genoa, ItalydCenter of Excellence for Biomedical Research (CEBR), University of Genoa, Genoa, ItalyeDepartment of Molecular Biochemistry and Pharmacology, IRCCSdIstituto di Ricerche Farmacologiche “Mario Negri,” Milano, Italy
a r t i c l e i n f o
Article history:Received 11 October 2013Accepted 4 January 2014Available online 10 January 2014
Keywords:Retinoic acidAb1e42RAGEAMIGO-1Ab peptide molecular assembly
* Corresponding author at: Department of ExperimGenoa, 2, L.B. Alberti St, Genoa I-16132, Italy. Tel.:010 3538836.
E-mail address: [email protected] (M. Nitti
0197-4580/$ e see front matter � 2014 Elsevier Inc. Ahttp://dx.doi.org/10.1016/j.neurobiolaging.2014.01.002
a b s t r a c t
The aggregation of amyloid-b (Ab) peptides plays a crucial role in the onset and progression ofAlzheimer’s disease. Monomeric form of Ab, indeed, could exert a physiological role. Considering theanti-oligomerization property of all-trans retinoic acid (ATRA), the involvement of monomeric Ab1e42in ATRA-induced neuronal differentiation has been investigated. Four-day ATRA treatment increasesb-secretase 1 (BACE1) level, Ab1e42 production, and receptor for advanced glycation end-products(RAGE) expression. RAGE is a well-recognized receptor for Ab, and the block of both RAGE and Ab1e42with specific antibodies strongly impairs neurite formation in ATRA-treated cells. The involvement ofAb1e42 and RAGE in ATRA-induced morphologic changes has been confirmed treating undifferenti-ated cells with different molecular assemblies of peptide: 1 mM monomeric, but not oligomeric, Ab1e42 increases RAGE expression and favors neurite elongation. The block of RAGE completely preventsthis effect. Furthermore, our data underline the involvement of the RAGE-dependent adhesion mole-cule amphoterin-induced gene and open reading frame-1 as downstream effector of both ATRA andAb1e42. In conclusion, our findings identify a novel physiological role for monomeric Ab1e42 andRAGE in neuronal differentiation.
� 2014 Elsevier Inc. All rights reserved.
1. Introduction
Amyloid-b (Ab) peptides are generated by the endoproteolyticprocessing of transmembrane Ab precursor protein (APP)(Anderson et al., 1992; Haass et al., 1993) sequentially cleaved bythe b-secretase 1 (BACE1) (Vassar et al., 1999) and the g-secretase(Selkoe et al., 1996). Progressive cerebral accumulation of Ab is theprimary pathologic event of Alzheimer’s disease (AD) and, in theearly stages of AD, Ab increased accumulation impairs synapticplasticity and memory (Selkoe, 2001).
A series of evidence suggests that the composition andaggregation state of Ab may explain its impacts, showing thatneuronal toxicity is exerted by oligomeric and fibrillar form of Ab
ental Medicine, University ofþ39 010 3538831; fax: þ39
).
ll rights reserved.
(Haass and Selkoe, 2007; Kayed et al., 2003; Piccini et al., 2005;Selkoe, 2003). However, the monomeric soluble Ab, especiallyAb1e42, could have physiological effects because it has beendemonstrated to regulate synaptic plasticity and long-termpotentiation (Puzzo et al., 2011). Moreover, we have recentlyshown that 1 mM Ab1e42, in monomeric form, induces thetranscription of BACE1 promoting a positive feedback loop withno sign of cell damage (Piccini et al., 2012), further confirmingthat both BACE1 activation and Ab1e42 production can be part ofa signaling pathway involved in physiological functions. All-transretinoic acid (ATRA) has been shown to antagonize the oligo-merization of Ab in vitro, decreasing its cellular toxicity (Takasakiet al., 2011). Moreover, Ab deposition and neuronal loss in thebrain have been significantly decreased in AD transgenic micetreated with retinoic acid (Ding et al., 2008). However, its role inthe regulation of APP processing remains poorly understood.
The receptor for advanced glycation end-products (RAGE) is acell surface protein (Vlassara, 1992) and member of the

S. Piras et al. / Neurobiology of Aging 35 (2014) 1301e13081302
immunoglobulin superfamily (Neeper et al., 1992). It is a multi-ligand receptor, able to bind AGEs (Schmidt et al., 1996), ampho-terin (HMGB1) (Schmidt et al., 2000; Treutiger et al., 2003), S100Bprotein (Donato, 2007), and Ab (Yan et al., 1996). RAGE is upre-gulated in several pathologic conditions such as diabetes(Nogueira-Machado and Chaves, 2008; Vlassara, 2001), athero-sclerosis (Jandeleit-Dahm et al., 2008; Yamamoto et al., 2000), andinflammation (Ramasamyet al., 2008; Yan et al., 2008), and, in thecentral nervous system, it plays a crucial role in neuro-degeneration (Chen et al., 2007; Nitti et al., 2005; Vincent et al.,2007; Yan et al., 1996). However, it has been shown that RAGEhas also a role in the physiological regulation of cell homeostasis(Schmidt et al., 2007) and is involved in neurite outgrowth(Huttunen et al., 2000) and in neuronal differentiation (Huttunenet al., 2002). Recently, a new adhesion molecule involved in neu-rite elongation has been identified to be under RAGE control, theamphoterin-induced gene and open reading frame-1 (AMIGO-1)(Chen et al., 2006; Kajander et al., 2011; Kuja-Panula et al., 2003).
In this study, we show that retinoic acid upregulating BACE1increases Ab1e42 generation. Ab1e42, through the activation ofRAGE and the upregulation of AMIGO-1, maintains neuriteoutgrowth and elongation, a specific feature of cell differentiation.
2. Materials and methods
2.1. Cell culture, differentiation, and treatments
SH-SY5Y neuroblastoma cells were cultured in RPMI 1640medium (Euroclone, Milan, Italy) supplemented with 10% fetalbovine serum (Euroclone), 2 mM glutamine (Sigma-Aldrich,Milan, Italy), 1% penicillin/streptomycin (Sigma-Aldrich), and 1%amphotericin B (Sigma-Aldrich), subcultured every 5 days at 1:5,and maintained in 5% CO2 humid atmosphere. Cells were differ-entiated by growth in the same medium supplemented with 10mM ATRA (Sigma-Aldrich) for 4 and 6 days. Some samples wereincubated with anti-RAGE blocking antibody (10 mg/mL mouseanti-RAGE; Chemicon International, Temecula, CA, USA) andanti-Ab1e42 antibody (2 mg/mL mouse anti-Ab1e42; Immuno-Biological Laboratories, Japan).
2.2. Preparation of Ab1e42 monomeric and oligomeric assemblies
Monomeric and oligomeric assemblies of Ab1e42 were pre-pared as previously described (Manzoni et al., 2009; Piccini et al.,2012).
2.3. Treatment of SH-SY5Y with Ab peptides
Aliquots of lyophilized monomeric and oligomeric assembliesAb1e42 were kept at �80 �C and dissolved at the time of theexperiment, as described (Manzoni et al., 2009). Cells were treatedwith 1 mM monomeric Ab1e42 in serum-free medium for 1 hourand then harvested for RNA and protein analyses.
2.4. Detection of cell death by fluorescence-activated cell sorteranalysis
After treatment, cells were stained with Annexin V-fluoresceinisothiocyanate and propidium iodide according to the manufac-turer’s instructions (BioVision, Mountain View, CA, USA). Stainedsamples were analyzed using FACSCanto II flow cytometer (BectonDickinson Italia [BD], Milan, Italy) equipped by FacsDiva software(BD). At least 10,000 events were analyzed. Each experiment wasperformed 3 times.
2.5. RNA extraction and semiquantitative real-time polymerasechain reaction
Total RNA was extracted using Trizol reagent (Life Technolo-gies, Carlsbad, CA, USA) according to the manufacturer’s in-structions and was then reverse transcribed into complementaryDNA (cDNA) by random hexamer primers and SuperScriptTM IIReverse Transcriptase (Life Technologies). Amplification of cDNAby polymerase chain reaction was performed using polymerasechain reaction (PCR) Master Mix 2� (Fermentas-Dasit, Milan,Italy) and specific primers for human AMIGO-1. Glyceraldehyde3-phosphate dehydrogenase (GAPDH) expression was used as thehousekeeping gene. Primer sequences used (Tib Mol Biol, Genoa,Italy) were AMIGO-1, forward 50-CCA AGT AAT GAA CGG GTG CT-30; reverse 50-GCT GGA AGG CTT CTC TAC AC-30 (315 bp) andGAPDH, forward 50-GTCTTCACCACCATGGAGAA-30; reverse 50-ATCCACAGTCTTCTGGGTGG-30 (266 bp). PCR products wereseparated by electrophoresis on 2% agarose gel prestained withethidium bromide, visualized under ultraviolet light, and quan-tified by densitometric analysis using a specific software (GelDoc;Bio-Rad, Milan, Italy).
2.6. Quantitative real-time PCR analysis
Primers and probes for BACE1 were designed with the aid ofPrimer Express software (Applied Biosystems). To avoid amplifica-tion of contaminating genomic DNA, primers and probes werelocated on adjacent exons.
Set primers and probe for BACE1 were forward 50-TGG AGG GCTTCT ACG TTG TCT T-30, reverse 50-GCT GCC GTC CTG AAC TCA TC-30,and probe 6FAM-50-TTG GCT TTG CTG TCA GCG CTT GC-30 TAMRA.
Set primers and probe for RAGE full length were forward 50-TGTGTG GCC ACC CAT TCC A-30, reverse 50-TCC CAC AGA GCC TGC AGTT-30, and probe 6FAM-50-AGG AAA GCC GTG CTG c TCA GCA TCAGCA TC-30 TAMRA.
Primers and probes for b-actin were obtained from a pre-developed assay on demand (Applied Biosystems). Five microlitersof the resulting cDNA dilution was used for quantitative PCRamplification performed, in duplicate, on the Prism 7900HT In-strument (Applied Biosystems) using the fluorescent TaqManmethod. The BACE1 messenger RNA (mRNA) quantities werenormalized to the control gene and were expressed in relation to acalibrator sample.
The levels of transcripts BACE1 and b-actin in each sample weredetermined using the standard curve. The standard curve was ob-tained with serial dilutions (106 to 10 molecules) of the calibrator,control plasmids containing cloned sequences of ABL gene (Ipsogen,Marseille, France).
2.7. Subcellular fractioning
Cytosolic and membrane fractions were obtained as previ-ously described (Nitti et al., 2010). Briefly, cells were lysed in 10mM N-2-hydroxyethyl piperazine-N-N-2 ethane sulfonic acid(Sigma-Aldrich) buffer pH 7.5, containing 0.25M sucrose, 5 mMethylenedinitrilo-tetraacetic acid (Sigma-Aldrich), and 10 mMmercaptoethanol (Sigma-Aldrich), supplemented with proteaseinhibitors (Roche Diagnostics). Samples were sonicated 4 � 1000,kept for 20 �C in ice, and centrifuged at 100,000� g for 30 mi-nutes. Cytosolic fraction (supernatant) was collected, and thepellet (membrane fraction) was dissolved in the same buffersupplemented with 0.3% Triton X-100 and centrifuged at100,000� g for 30 minutes. The supernatant was collected, andthe protein content was measured using Lowry assay.
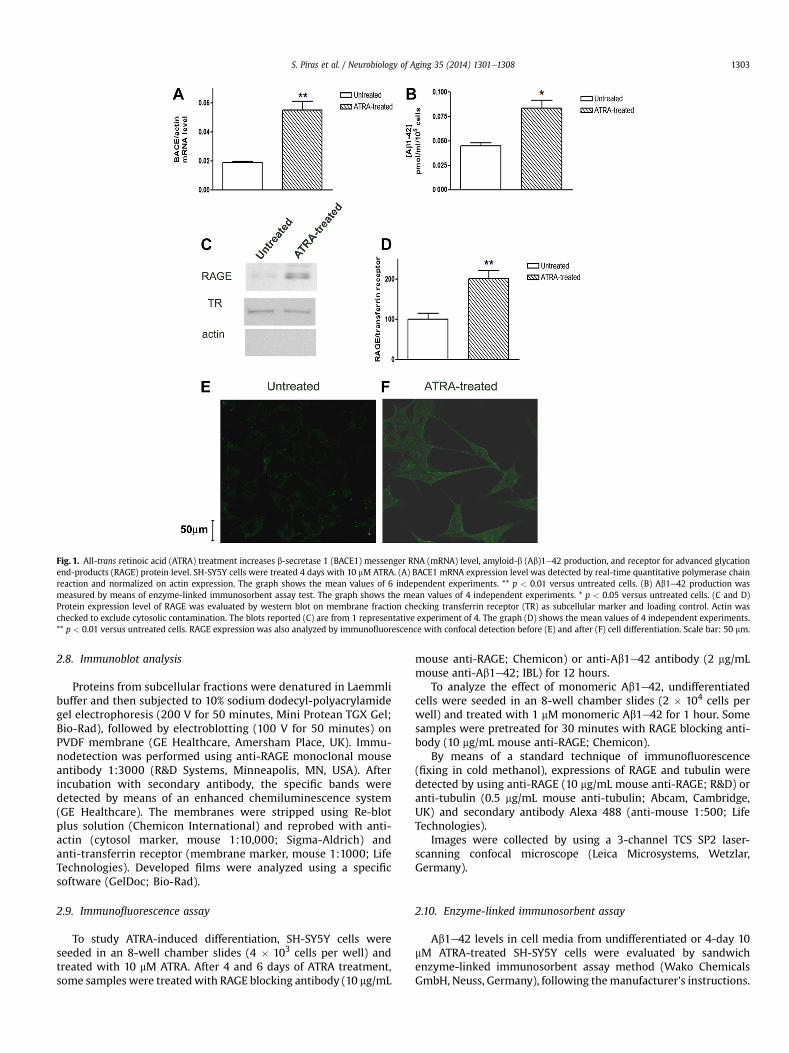
Fig. 1. All-trans retinoic acid (ATRA) treatment increases b-secretase 1 (BACE1) messenger RNA (mRNA) level, amyloid-b (Ab)1e42 production, and receptor for advanced glycationend-products (RAGE) protein level. SH-SY5Y cells were treated 4 days with 10 mM ATRA. (A) BACE1 mRNA expression level was detected by real-time quantitative polymerase chainreaction and normalized on actin expression. The graph shows the mean values of 6 independent experiments. ** p < 0.01 versus untreated cells. (B) Ab1e42 production wasmeasured by means of enzyme-linked immunosorbent assay test. The graph shows the mean values of 4 independent experiments. * p < 0.05 versus untreated cells. (C and D)Protein expression level of RAGE was evaluated by western blot on membrane fraction checking transferrin receptor (TR) as subcellular marker and loading control. Actin waschecked to exclude cytosolic contamination. The blots reported (C) are from 1 representative experiment of 4. The graph (D) shows the mean values of 4 independent experiments.** p < 0.01 versus untreated cells. RAGE expression was also analyzed by immunofluorescence with confocal detection before (E) and after (F) cell differentiation. Scale bar: 50 mm.
S. Piras et al. / Neurobiology of Aging 35 (2014) 1301e1308 1303
2.8. Immunoblot analysis
Proteins from subcellular fractions were denatured in Laemmlibuffer and then subjected to 10% sodium dodecyl-polyacrylamidegel electrophoresis (200 V for 50 minutes, Mini Protean TGX Gel;Bio-Rad), followed by electroblotting (100 V for 50 minutes) onPVDF membrane (GE Healthcare, Amersham Place, UK). Immu-nodetection was performed using anti-RAGE monoclonal mouseantibody 1:3000 (R&D Systems, Minneapolis, MN, USA). Afterincubation with secondary antibody, the specific bands weredetected by means of an enhanced chemiluminescence system(GE Healthcare). The membranes were stripped using Re-blotplus solution (Chemicon International) and reprobed with anti-actin (cytosol marker, mouse 1:10,000; Sigma-Aldrich) andanti-transferrin receptor (membrane marker, mouse 1:1000; LifeTechnologies). Developed films were analyzed using a specificsoftware (GelDoc; Bio-Rad).
2.9. Immunofluorescence assay
To study ATRA-induced differentiation, SH-SY5Y cells wereseeded in an 8-well chamber slides (4 � 103 cells per well) andtreated with 10 mM ATRA. After 4 and 6 days of ATRA treatment,some samples were treatedwith RAGE blocking antibody (10 mg/mL
mouse anti-RAGE; Chemicon) or anti-Ab1e42 antibody (2 mg/mLmouse anti-Ab1e42; IBL) for 12 hours.
To analyze the effect of monomeric Ab1e42, undifferentiatedcells were seeded in an 8-well chamber slides (2 � 104 cells perwell) and treated with 1 mM monomeric Ab1e42 for 1 hour. Somesamples were pretreated for 30 minutes with RAGE blocking anti-body (10 mg/mL mouse anti-RAGE; Chemicon).
By means of a standard technique of immunofluorescence(fixing in cold methanol), expressions of RAGE and tubulin weredetected by using anti-RAGE (10 mg/mL mouse anti-RAGE; R&D) oranti-tubulin (0.5 mg/mL mouse anti-tubulin; Abcam, Cambridge,UK) and secondary antibody Alexa 488 (anti-mouse 1:500; LifeTechnologies).
Images were collected by using a 3-channel TCS SP2 laser-scanning confocal microscope (Leica Microsystems, Wetzlar,Germany).
2.10. Enzyme-linked immunosorbent assay
Ab1e42 levels in cell media from undifferentiated or 4-day 10mM ATRA-treated SH-SY5Y cells were evaluated by sandwichenzyme-linked immunosorbent assay method (Wako ChemicalsGmbH, Neuss, Germany), following themanufacturer’s instructions.
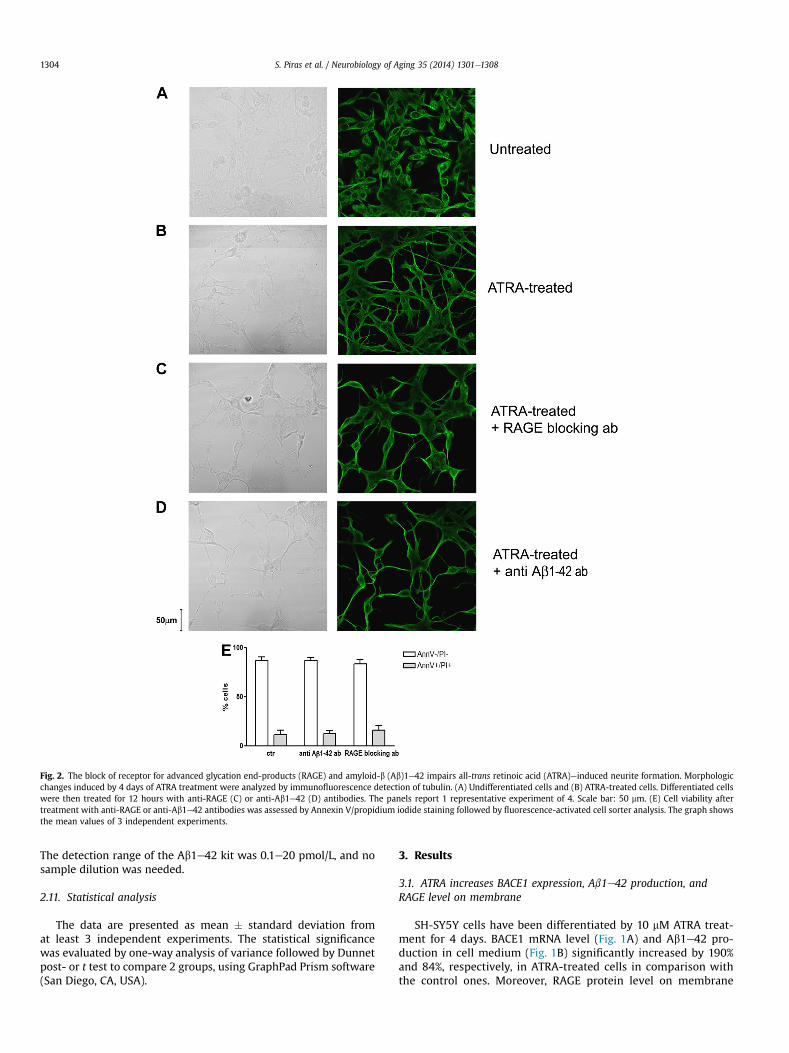
Fig. 2. The block of receptor for advanced glycation end-products (RAGE) and amyloid-b (Ab)1e42 impairs all-trans retinoic acid (ATRA)einduced neurite formation. Morphologicchanges induced by 4 days of ATRA treatment were analyzed by immunofluorescence detection of tubulin. (A) Undifferentiated cells and (B) ATRA-treated cells. Differentiated cellswere then treated for 12 hours with anti-RAGE (C) or anti-Ab1e42 (D) antibodies. The panels report 1 representative experiment of 4. Scale bar: 50 mm. (E) Cell viability aftertreatment with anti-RAGE or anti-Ab1e42 antibodies was assessed by Annexin V/propidium iodide staining followed by fluorescence-activated cell sorter analysis. The graph showsthe mean values of 3 independent experiments.
S. Piras et al. / Neurobiology of Aging 35 (2014) 1301e13081304
The detection range of the Ab1e42 kit was 0.1e20 pmol/L, and nosample dilution was needed.
2.11. Statistical analysis
The data are presented as mean � standard deviation fromat least 3 independent experiments. The statistical significancewas evaluated by one-way analysis of variance followed by Dunnetpost- or t test to compare 2 groups, using GraphPad Prism software(San Diego, CA, USA).
3. Results
3.1. ATRA increases BACE1 expression, Ab1e42 production, andRAGE level on membrane
SH-SY5Y cells have been differentiated by 10 mM ATRA treat-ment for 4 days. BACE1 mRNA level (Fig. 1A) and Ab1e42 pro-duction in cell medium (Fig. 1B) significantly increased by 190%and 84%, respectively, in ATRA-treated cells in comparison withthe control ones. Moreover, RAGE protein level on membrane
S. Piras et al. / Neurobiology of Aging 35 (2014) 1301e1308 1305
subcellular fractions increased by 80% in differentiated cells incomparison with undifferentiated ones (Fig. 1C and D). Theenhanced level of RAGE on the membrane has been confirmed bythe immunofluorescence analysis of RAGE in untreated (Fig. 1E) orATRA-treated (Fig. 1F) cells. RAGE appeared to be mainly locatedon cell membranes in differentiated cells, whereas undifferenti-ated cells showed only a slight cytosolic staining.
3.2. The block of RAGE or Ab1e42 impairs neurite elongation inATRA-treated cells
Four-day ATRA treatment induced SH-SY5Y differentiationwith evident neurite outgrowth and elongation compared withuntreated cells, as shown by immunofluorescence staining fortubulin (Fig. 2A and B). Moreover, 12-hour treatment both withanti-RAGE and anti-Ab1e42 antibodies reduced neurite branch-ing and thickness (Fig. 2C and D). No changes of cell viability havebeen observed in cells treated with both antibodies in compari-son with control cells (Fig. 3E). All the previously mentionedresults have been confirmed with 6 days of ATRA treatment (datanot shown).
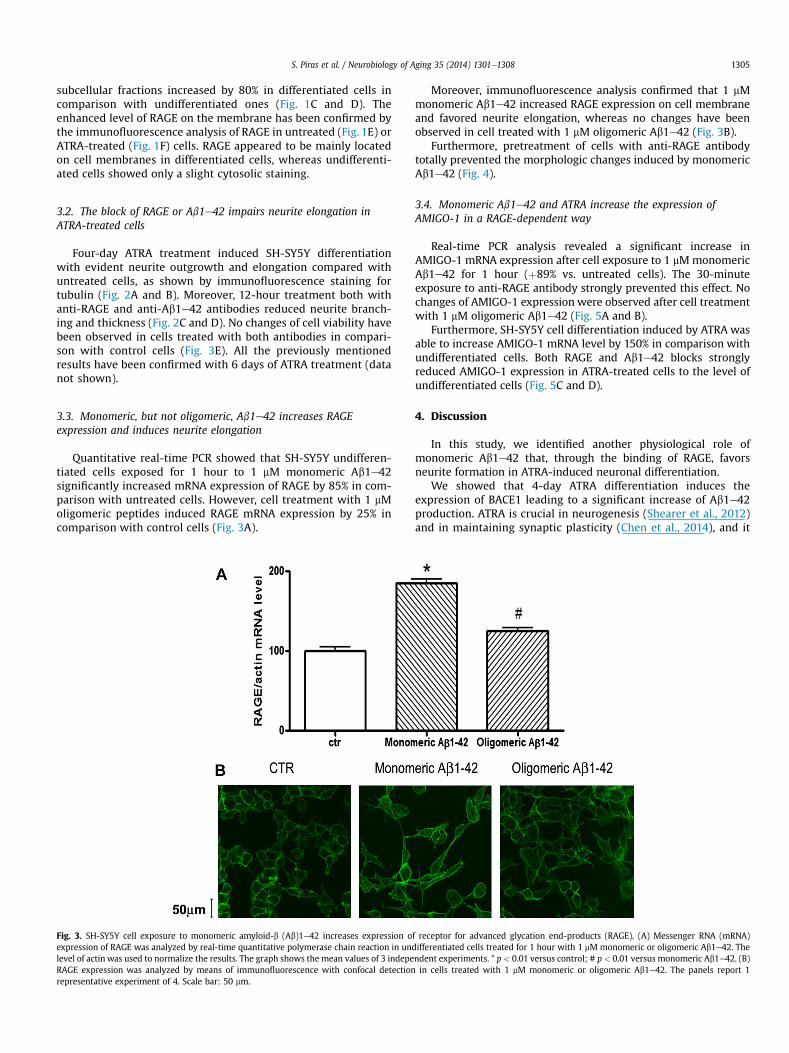
3.3. Monomeric, but not oligomeric, Ab1e42 increases RAGEexpression and induces neurite elongation
Quantitative real-time PCR showed that SH-SY5Y undifferen-tiated cells exposed for 1 hour to 1 mM monomeric Ab1e42significantly increased mRNA expression of RAGE by 85% in com-parison with untreated cells. However, cell treatment with 1 mMoligomeric peptides induced RAGE mRNA expression by 25% incomparison with control cells (Fig. 3A).
Fig. 3. SH-SY5Y cell exposure to monomeric amyloid-b (Ab)1e42 increases expression oexpression of RAGE was analyzed by real-time quantitative polymerase chain reaction in unlevel of actin was used to normalize the results. The graph shows the mean values of 3 indepeRAGE expression was analyzed by means of immunofluorescence with confocal detectionrepresentative experiment of 4. Scale bar: 50 mm.
Moreover, immunofluorescence analysis confirmed that 1 mMmonomeric Ab1e42 increased RAGE expression on cell membraneand favored neurite elongation, whereas no changes have beenobserved in cell treated with 1 mM oligomeric Ab1e42 (Fig. 3B).
Furthermore, pretreatment of cells with anti-RAGE antibodytotally prevented the morphologic changes induced by monomericAb1e42 (Fig. 4).
3.4. Monomeric Ab1e42 and ATRA increase the expression ofAMIGO-1 in a RAGE-dependent way
Real-time PCR analysis revealed a significant increase inAMIGO-1 mRNA expression after cell exposure to 1 mMmonomericAb1e42 for 1 hour (þ89% vs. untreated cells). The 30-minuteexposure to anti-RAGE antibody strongly prevented this effect. Nochanges of AMIGO-1 expression were observed after cell treatmentwith 1 mM oligomeric Ab1e42 (Fig. 5A and B).
Furthermore, SH-SY5Y cell differentiation induced by ATRA wasable to increase AMIGO-1 mRNA level by 150% in comparison withundifferentiated cells. Both RAGE and Ab1e42 blocks stronglyreduced AMIGO-1 expression in ATRA-treated cells to the level ofundifferentiated cells (Fig. 5C and D).
4. Discussion
In this study, we identified another physiological role ofmonomeric Ab1e42 that, through the binding of RAGE, favorsneurite formation in ATRA-induced neuronal differentiation.
We showed that 4-day ATRA differentiation induces theexpression of BACE1 leading to a significant increase of Ab1e42production. ATRA is crucial in neurogenesis (Shearer et al., 2012)and in maintaining synaptic plasticity (Chen et al., 2014), and it
f receptor for advanced glycation end-products (RAGE). (A) Messenger RNA (mRNA)differentiated cells treated for 1 hour with 1 mM monomeric or oligomeric Ab1e42. Thendent experiments. * p < 0.01 versus control; # p < 0.01 versus monomeric Ab1e42. (B)in cells treated with 1 mM monomeric or oligomeric Ab1e42. The panels report 1

Fig. 4. Monomeric amyloid-b (Ab)1e42 induces receptor for advanced glycation end-products (RAGE)edependent neurite elongation. RAGE expression was analyzed by immu-nofluorescence with confocal detection after treatment with 1 mMmonomeric Ab1e42 for 1 hour. Control samples were pretreated with RAGE blocking antibody for 30 minutes. Thepanels show 1 representative experiment of 4. Scale bar: 50 mm.
S. Piras et al. / Neurobiology of Aging 35 (2014) 1301e13081306
has been initially believed to stimulate the nonamyloidogenicprocessing of APP (Fahrenholz and Postina, 2006; Holback et al.,2008). However, in agreement with other studies (Koryakinaet al., 2009), we demonstrated a more complex effect of reti-noic acid on APP processing, confirming the upregulation ofBACE1 in differentiated cells, already demonstrated by Satoh andKuroda (2000) on NT2 cells. Moreover, we also show, for thefirst time, the increased production of Ab1e42 in retinoicacidetreated cells. Furthermore, it has been recently shown thatvitamin A and retinoic acid not only destabilize preformed Abfibrils (Ono et al., 2004) but also hamper the oligomerization andfibrillation of Ab, at least in vitro (Takasaki et al., 2011). Main-taining Ab in monomeric form is crucial to avoid Ab toxicity(Piccini et al., 2005; Selkoe, 2003). Recently, we showed that
Fig. 5. Amphoterin-induced gene and open reading frame-1 (AMIGO-1) is upregulated byadvanced glycation end-products (RAGE)edependent way. AMIGO-1 expression was analyelectrophoresis, and the relative intensities of the bands were normalized to glyceraldehydewith 1 mM monomeric or oligomeric Ab1e42 for 1 hour. Some samples were pretreated (30independent experiments. ** p < 0.01 versus control. x p < 0.01 versus monomeric Ab1e42. Texposed to 10 mM ATRA for 4 days and then treated for 12 hours with anti-RAGE or anti-Ab1e* p < 0.05 versus undifferentiated cells; # p < 0.05 versus ATRA-treated cells. The bands (D
monomeric Ab1e42 is able to upregulate BACE1 leading to apositive feedback loop that maintains Ab production without anysign of cell toxicity (Piccini et al., 2012), confirming that not onlyAb production but also BACE1 upregulation is not merely toxicand together could play a role in cell physiology.
Many hypothesis have been made about potential Ab receptors,including NMDA receptor (Bi et al., 2002), AMPA receptor (Zhaoet al., 2010), low-density lipoprotein receptorerelated protein(Bu et al., 2006), and alpha7 nicotinic acetylcholine receptor(Wang et al., 2000), and a prominent role has been suggested forRAGE (Chen et al., 2007; Giri et al., 2002; Yan et al., 1996, 2009).We show that RAGE expression is increased in ATRA-treated cellsand is crucial for cell differentiation: indeed, the exposure ofdifferentiated cells to RAGE blocking antibody induces the same
all-trans retinoic acid (ATRA) and monomeric amyloid-b (Ab)1e42 in an receptor forzed by real-time polymerase chain reaction (PCR). PCR products were separated by3-phosphate dehydrogenase (GAPDH) expression. (A and B) SH-SY5Y cells were treatedminutes) with RAGE blocking antibody. The bar graph (A) shows the mean values of 3he bands (B) are representative of 3 independent experiments. (C and D) SH-SY5Y were42 antibodies. The bar graph (C) shows the mean values of 3 independent experiments.) are representative of 3 independent experiments.

S. Piras et al. / Neurobiology of Aging 35 (2014) 1301e1308 1307
morphologic changes observed in differentiated cells exposed toanti-Ab antibody, impairing neurite formation. The involvement ofRAGE-mediated signaling in cell differentiation has already beenshown in other articles, totally in agreement with our data(Huttunen et al., 2002; Wang et al., 2008).
To verify the role of Ab in cell differentiation, we exposed un-differentiated cells to Ab1e42. There is high biological variability ofAb effects among different batches of commercially available pep-tides, leading to heterogeneous and discordant results about thephysiological effect of Ab peptide that, in these preparations, canreach different molecular assemblies. To avoid this step, we used apreparation of Ab1e42 containing mostly monomeric species(Manzoni et al., 2009) and maintained low concentration (1 mM)and short timeframe (1 hour), to minimize the generation of toxicoligomeric forms (Piccini et al., 2012).
Our results show that monomeric Ab1e42 is able to induceRAGE mRNA level significantly more than oligomeric Ab1e42 inundifferentiated cells. Moreover, monomeric and not oligomericAb1e42 increases RAGE protein expression on cell membrane andinduces neurite elongation, further confirming that Ab molecularassembly is crucial in determining the effects of peptides.
In addition, we show that neurite elongation induced bymonomeric Ab1e42 is exerted in RAGE-dependent way: indeed,cell pretreatment with anti-RAGE antibody completely abolishedmorphologic changes induced by Ab1e42.
RAGE involvement in neurite elongation has already beendemonstrated (Kim et al., 2012; Wang et al., 2008), but, for the firsttime, we point out the role of monomeric Ab1e42 as a putativeligand for RAGE and the involvement of both molecules in ATRA-induced differentiation.
To identify a possible molecular target of Ab-RAGE differenti-ating signal, we investigated on AMIGO, a family of transmembraneproteins that, under RAGE control, trigger the elongation andfasciculation of neurites (Chen et al., 2006, 2012; Kajander et al.,2011).
We demonstrated that AMIGO-1 expression is upregulated byATRA treatment and the block of both Ab1e42 and RAGE reducesAMIGO-1 expression to the level of undifferentiated cells. More-over, we showed thatmonomeric but not oligomeric Ab1e42 is ableto increase AMIGO-1 mRNA level in a RAGE-dependent waybecause the block of RAGE totally prevents the effect of monomericAb1e42.
5. Conclusions
In this study, we show that neuronal differentiation induced byATRA involves the generation of Ab1e42 and the activation ofRAGE-mediated signaling, leading to neurite elongation throughthe upregulation of AMIGO-1 adhesion molecule. We proved thatthe aggregation state is crucial in determining the effect of Ab1e42showing a prodifferentiating role of monomeric but not oligomericAb1e42. Providing evidence of a novel physiological role formonomeric Ab1e42, we point out that strategies of AD therapyshould be focused not only in reduction of Ab amount but also ininhibition of Ab aggregation.
Disclosure statement
Authors have no conflicts of interest to declare.
Acknowledgements
The study is supported by grants from MIUR-PRIN 20125S38FA(Dr Mariapaola Nitti), Fondazione CARIGE (Professor Pronzato),Genoa University, and Banca Intesasanpaolo (Dr Mario Salmona).
The kind gift of FMOC amino acids by Flamma Spa (Bergamo, Italy)is gratefully acknowledged.
References
Anderson, J.P., Chen, Y., Kim, K.S., Robakis, N.K., 1992. An alternative secretasecleavage produces soluble Alzheimer amyloid precursor protein containing apotentially amyloidogenic sequence. J. Neurochem. 59, 2328e2331.
Bi, X., Gall, C.M., Zhou, J., Lynch, G., 2002. Uptake and pathogenic effects of amyloidbeta peptide 1-42 are enhanced by integrin antagonists and blocked by NMDAreceptor antagonists. Neuroscience 112, 827e840.
Bu, G., Cam, J., Zerbinatti, C., 2006. LRP in amyloid-beta production and metabolism.Ann. N. Y. Acad. Sci. 1086, 35e53.
Chen, L., Lau, A.G., Sarti, F., 2014. Synaptic retinoic acid signaling and homeostaticsynaptic plasticity. Neuropharmacology 78, 3e12.
Chen, X., Walker, D.G., Schmidt, A.M., Arancio, O., Lue, L.F., Yan, S.D., 2007. RAGE: apotential target for Abeta-mediated cellular perturbation in Alzheimer’s dis-ease. Curr. Mol. Med. 7, 735e742.
Chen, Y., Aulia, S., Li, L., Tang, B.L., 2006. AMIGO and friends: an emerging family ofbrain-enriched, neuronal growth modulating, type I transmembrane proteinswith leucine-rich repeats (LRR) and cell adhesion molecule motifs. Brain Res.Rev. 51, 265e274.
Chen, Y., Hor, H.H., Tang, B.L., 2012. AMIGO is expressed in multiple brain cell typesand may regulate dendritic growth and neuronal survival. J. Cell Physiol. 227,2217e2229.
Ding, Y., Qiao, A., Wang, Z., Goodwin, J.S., Lee, E.S., Block, M.L., Allsbrook, M.,McDonald, M.P., Fan, G.H., 2008. Retinoic acid attenuates beta-amyloid depo-sition and rescues memory deficits in an Alzheimer’s disease transgenic mousemodel. J. Neurosci. 28, 11622e11634.
Donato, R., 2007. RAGE: a single receptor for several ligands and different cellularresponses: the case of certain S100 proteins. Curr. Mol. Med. 7, 711e724.
Fahrenholz, F., Postina, R., 2006. Alpha-secretase activationdan approach toAlzheimer’s disease therapy. Neurodegener. Dis. 3, 255e261.
Giri, R., Selvaraj, S., Miller, C.A., Hofman, F., Yan, S.D., Stern, D., Zlokovic, B.V.,Kalra, V.K., 2002. Effect of endothelial cell polarity on beta-amyloid-inducedmigration of monocytes across normal and AD endothelium. Am. J. Physiol.Cell Physiol. 283, C895eC904.
Haass, C., Hung, A.Y., Schlossmacher, M.G., Oltersdorf, T., Teplow, D.B., Selkoe, D.J.,1993. Normal cellular processing of the beta-amyloid precursor protein resultsin the secretion of the amyloid beta peptide and related molecules. Ann. N. Y.Acad. Sci. 695, 109e116.
Haass, C., Selkoe, D.J., 2007. Soluble protein oligomers in neurodegeneration: les-sons from the Alzheimer’s amyloid beta-peptide. Nat. Rev. Mol. Cell Biol. 8,101e112.
Holback, S., Adlerz, L., Gatsinzi, T., Jacobsen, K.T., Iverfeldt, K., 2008. PI3-K- and PKC-dependent up-regulation of APP processing enzymes by retinoic acid. Biochem.Biophys. Res. Commun. 365, 298e303.
Huttunen, H.J., Kuja-Panula, J., Sorci, G., Agneletti, A.L., Donato, R., Rauvala, H., 2000.Coregulation of neurite outgrowth and cell survival by amphoterin and S100proteins through receptor for advanced glycation end products (RAGE) activa-tion. J. Biol. Chem. 275, 40096e40105.
Huttunen, H.J., Kuja-Panula, J., Rauvala, H., 2002. Receptor for advanced glycationend products (RAGE) signaling induces CREB-dependent chromograninexpression during neuronal differentiation. J. Biol. Chem. 277, 38635e38646.
Jandeleit-Dahm, K., Watson, A., Soro-Paavonen, A., 2008. The AGE/RAGE axis indiabetes-accelerated atherosclerosis. Clin. Exp. Pharmacol. Physiol. 35, 329e334.
Kajander, T., Kuja-Panula, J., Rauvala, H., Goldman, A., 2011. Crystal structure androle of glycans and dimerization in folding of neuronal leucine-rich repeatprotein AMIGO-1. J. Mol. Biol. 413, 1001e1015.
Kayed, R., Head, E., Thompson, J.L., McIntire, T.M., Milton, S.C., Cotman, C.W.,Glabe, C.G., 2003. Common structure of soluble amyloid oligomers impliescommon mechanism of pathogenesis. Science 300, 486e489.
Kim, J., Wan, C.K., O’Carroll, S.J., Shaikh, S.B., Nicholson, L.F., 2012. The role of re-ceptor for advanced glycation end products (RAGE) in neuronal differentiation.J. Neurosci. Res. 90, 1136e1147.
Koryakina, A., Aeberhard, J., Kiefer, S., Hamburger, M., Kuenzi, P., 2009. Regulation ofsecretases by all-trans-retinoic acid. FEBS J. 276, 2645e2655.
Kuja-Panula, J., Kiiltomaki, M., Yamashiro, T., Rouhiainen, A., Rauvala, H., 2003.AMIGO, a transmembrane protein implicated in axon tract development, de-fines a novel protein family with leucine-rich repeats. J. Cell Biol. 160, 963e973.
Manzoni, C., Colombo, L., Messa, M., Cagnotto, A., Cantù, L., Del Favero, E.,Salmona, M., 2009. Overcoming synthetic Abeta peptide aging: a new approachto an age-old problem. Amyloid 16, 71e80.
Neeper, M., Schmidt, A.M., Brett, J., Yan, S.D., Wang, F., Pan, Y.C., Elliston, K., Stern, D.,Shaw, A., 1992. Cloning and expression of a cell surface receptor for advancedglycosylation end products of proteins. J. Biol. Chem. 267, 14998e15004.
Nitti, M., d’Abramo, C., Traverso, N., Verzola, D., Garibotto, G., Poggi, A., Odetti, P.,Cottalasso, D., Marinari, U.M., Pronzato, M.A., Domenicotti, C., 2005. Central roleof PKCdelta in glycoxidation-dependent apoptosis of human neurons. FreeRadic. Biol. Med. 38, 846e856.
Nitti, M., Furfaro, A.L., Cevasco, C., Traverso, N., Marinari, U.M., Pronzato, M.A.,Domenicotti, C., 2010. PKC delta and NADPH oxidase in retinoic acid-inducedneuroblastoma cell differentiation. Cell Signal. 22, 828e835.

S. Piras et al. / Neurobiology of Aging 35 (2014) 1301e13081308
Nogueira-Machado, J.A., Chaves, M.M., 2008. From hyperglycemia to AGE-RAGEinteraction on the cell surface: a dangerous metabolic route for diabetic pa-tients. Expert Opin. Ther. Targets 12, 871e882.
Ono, K., Yoshiike, Y., Takashima, A., Hasegawa, K., Naiki, H., Yamada, M., 2004.Vitamin A exhibits potent antiamyloidogenic and fibril-destabilizing effectsin vitro. Exp. Neurol. 189, 380e392.
Piccini, A., Russo, C., Gliozzi, A., Relini, A., Vitali, A., Borghi, R., Giliberto, L.,Armirotti, A., D’Arrigo, C., Bachi, A., Cattaneo, A., Canale, C., Torrassa, S.,Saido, T.C., Markesbery, W., Gambetti, P., Tabaton, M., 2005. Beta-amyloid isdifferent in normal aging and in Alzheimer disease. J. Biol. Chem. 280,34186e34192.
Piccini, A., Borghi, R., Guglielmotto, M., Tamagno, E., Cirmena, G., Garuti, A.,Pollero, V., Cammarata, S., Fornaro, M., Messa, M., Colombo, L., Salmona, M.,Perry, G., Tabaton, M., 2012. Beta-amyloid 1-42 induces physiological tran-scriptional regulation of BACE1. J. Neurochem. 122, 1023e1031.
Puzzo, D., Privitera, L., Fa’, M., Staniszewski, A., Hashimoto, G., Aziz, F., Sakurai, M.,Ribe, E.M., Troy, C.M., Mercken, M., Jung, S.S., Palmeri, A., Arancio, O., 2011.Endogenous amyloid-beta is necessary for hippocampal synaptic plasticity andmemory. Ann. Neurol. 69, 819e830.
Ramasamy, R., Yan, S.F., Herold, K., Clynes, R., Schmidt, A.M., 2008. Receptor foradvanced glycation end products: fundamental roles in the inflammatoryresponse: winding the way to the pathogenesis of endothelial dysfunction andatherosclerosis. Ann. N. Y Acad. Sci. 1126, 7e13.
Satoh, J., Kuroda, Y., 2000. Amyloid precursor protein beta-secretase (BACE) mRNAexpression in human neural cell lines following induction of neuronal differ-entiation and exposure to cytokines and growth factors. Neuropathology 20,289e296.
Schmidt, A., Kuhla, B., Bigl, K., Munch, G., Arendt, T., 2007. Cell cycle related signalingin Neuro2a cells proceeds via the receptor for advanced glycation end products.J. Neural Transm. 114, 1413e1424.
Schmidt, A.M., Hori, O., Cao, R., Yan, S.D., Brett, J., Wautier, J.L., Ogawa, S.,Kuwabara, K., Matsumoto, M., Stern, D., 1996. RAGE: a novel cellular receptor foradvanced glycation end products. Diabetes 45 (suppl 3), S77eS80.
Schmidt, A.M., Yan, S.D., Yan, S.F., Stern, D.M., 2000. The biology of the receptor foradvanced glycation end products and its ligands. Biochim. Biophys. Acta 1498,99e111.
Selkoe, D.J., Yamazaki, T., Citron, M., Podlisny, M.B., Koo, E.H., Teplow, D.B., Haass, C.,1996. The role of APP processing and trafficking pathways in the formation ofamyloid beta-protein. Ann. N. Y. Acad. Sci. 777, 57e64.
Selkoe, D.J., 2001. Alzheimer’s disease results from the cerebral accumulation andcytotoxicity of amyloid beta-protein. J. Alzheimers Dis. 3, 75e80.
Selkoe, D.J., 2003. Folding proteins in fatal ways. Nature 426, 900e904.Shearer, K.D., Stoney, P.N., Morgan, P.J., McCaffery, P.J., 2012. A vitamin for the brain.
Trends Neurosci. 35, 733e741.Takasaki, J., Ono, K., Yoshiike, Y., Hirohata, M., Ikeda, T., Morinaga, A., Takashima, A.,
Yamada, M., 2011. Vitamin A has anti-oligomerization effects on amyloid-betain vitro. J. Alzheimers Dis. 27, 271e280.
Treutiger, C.J., Mullins, G.E., Johansson, A.S., Rouhiainen, A., Rauvala, H.M.,Erlandsson-Harris, H., Andersson, U., Yang, H., Tracey, K.J., Andersson, J.,Palmblad, J.E., 2003. High mobility group 1 B-box mediates activation of humanendothelium. J. Intern. Med. 254, 375e385.
Vassar, R., Bennett, B.D., Babu-Khan, S., Kahn, S., Mendiaz, E.A., Denis, P.,Teplow, D.B., Ross, S., Amarante, P., Loeloff, R., Luo, Y., Fisher, S., Fuller, J.,Edenson, S., Lile, J., Jarosinski, M.A., Biere, A.L., Curran, E., Burgess, T., Louis, J.C.,Collins, F., Treanor, J., Rogers, G., Citron, M., 1999. Beta-secretase cleavage ofAlzheimer’s amyloid precursor protein by the transmembrane aspartic proteaseBACE. Science 286, 735e741.
Vincent, A.M., Perrone, L., Sullivan, K.A., Backus, C., Sastry, A.M., Lastoskie, C.,Feldman, E.L., 2007. Receptor for advanced glycation end products activationinjures primary sensory neurons via oxidative stress. Endocrinology 148,548e558.
Vlassara, H., 1992. Receptor-mediated interactions of advanced glycosylation endproducts with cellular components within diabetic tissues. Diabetes 41 (suppl 2),52e56.
Vlassara, H., 2001. The AGE-receptor in the pathogenesis of diabetic complications.Diabetes Metab. Res. Rev. 17, 436e443.
Wang, H.Y., Lee, D.H., D’Andrea, M.R., Peterson, P.A., Shank, R.P., Reitz, A.B., 2000.Beta-amyloid(1-42) binds to alpha7 nicotinic acetylcholine receptor with highaffinity. Implications for Alzheimer’s disease pathology. J. Biol. Chem. 275,5626e5632.
Wang, L., Li, S., Jungalwala, F.B., 2008. Receptor for advanced glycation end products(RAGE) mediates neuronal differentiation and neurite outgrowth. J. Neurosci.Res. 86, 1254e1266.
Yamamoto, Y., Yamagishi, S., Yonekura, H., Doi, T., Tsuji, H., Kato, I., Takasawa, S.,Okamoto, H., Abedin, J., Tanaka, N., Sakurai, S., Migita, H., Unoki, H., Wang, H.,Zenda, T., Wu, P.S., Segawa, Y., Higashide, T., Kawasaki, K., Yamamoto, H., 2000.Roles of the AGE-RAGE system in vascular injury in diabetes. Ann. N. Y. Acad. Sci.902, 163e170; discussion 170e172.
Yan, S.D., Chen, X., Fu, J., Chen, M., Zhu, H., Roher, A., Slattery, T., Zhao, L.,Nagashima, M., Morser, J., Migheli, A., Nawroth, P., Stern, D., Schmidt, A.M., 1996.RAGE and amyloid-beta peptide neurotoxicity in Alzheimer’s disease. Nature382, 685e691.
Yan, S.D., Bierhaus, A., Nawroth, P.P., Stern, D.M., 2009. RAGE and Alzheimer’s dis-ease: a progression factor for amyloid-beta-induced cellular perturbation?J. Alzheimers Dis. 16, 833e843.
Yan, S.F., Ramasamy, R., Schmidt, A.M., 2008. Mechanisms of disease: advancedglycation end-products and their receptor in inflammation and diabetes com-plications. Nat. Clin. Pract. Endocrinol. Metab. 4, 285e293.
Zhao, W.Q., Santini, F., Breese, R., Ross, D., Zhang, X.D., Stone, D.J., Ferrer, M.,Townsend, M., Wolfe, A.L., Seager, M.A., Kinney, G.G., Shughrue, P.J., Ray, W.J.,2010. Inhibition of calcineurin-mediated endocytosis and alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors prevents am-yloid beta oligomer-induced synaptic disruption. J. Biol. Chem. 285,7619e7632.