cyp12a1, a mitochondrial cytochrome p450 from the house fly
TRANSCRIPT

CYP12A1, a Mitochondrial Cytochrome P450from the House Fly1
Victor M. Guzov,2 Gopalan C. Unnithan, Alexey A. Chernogolov,* and Rene Feyereisen3
Department of Entomology and Center for Toxicology, University of Arizona, Forbes 410, Tucson, Arizona 85721; and*Institute of Bioorganic Chemistry, Academy of Sciences of Belarus, 220141 Minsk, Belarus
Received May 22, 1998, and in revised form August 17, 1998
Eukaryotic P450 proteins are membrane proteinsfound predominantly in the endoplasmic reticulum. Invertebrates, several biosynthetic P450s are found in mi-tochondria as well. We cloned three putative insect mi-tochondrial P450s from larval house fly cDNA. TheseP450s are members of a new P450 family, CYP12. TheCYP12 proteins are most closely related to the mamma-lian mitochondrial P450 of the CYP11, CYP24, andCYP27 families. The most abundant cDNA, CYP12A1,was expressed in Escherichia coli and purified. NADPH-dependent reduction of CYP12A1 was rapid and efficientwith the bovine mitochondrial proteins adrenodoxin re-ductase and adrenodoxin as electron transfer partners.In contrast, house fly microsomal NADPH cytochromeP450 reductase reduced CYP12A1 only poorly. In a re-constituted system with the bovine mitochondrial elec-tron donors, CYP12A1 metabolized a variety of insec-ticides and other xenobiotics, but did not metabolizeecdysteroids, juvenoids, or fatty acids. Subcellular local-ization of CYP12A1 by immunogold histochemistry es-tablished the mitochondrial nature of this protein.CYP12A1 mRNA levels are constitutively higher in aninsecticide-resistant strain than in a susceptible strain,and this trait maps to chromosome II in the house fly,where the constitutive overexpression of the pesticide-metabolizing microsomal CYP6A1 also maps. Multiplemitochondrial P450s have evolved in insects and mayplay a role in the metabolism of xenobiotics in additionto their possibly ancestral functions in steroidogenesis.© 1998 Academic Press
Key Words: mitochondrial P450; heterologous expres-sion; immunogold histochemistry; pesticide metabolism.
The P450s4 constitute a large superfamily of hemeproteins sharing a common, ancient evolutionary ori-gin (1). Currently, more than 500 P450 sequences havebeen documented from eubacteria, archaebacteria,plants, fungi, and animals (2). These enzymes playcrucial roles in the biosynthetic and degradative path-ways for many endogenous compounds, such as steroidhormones, vitamins D3, bile acids, and eicosanoids (3).P450s have also received considerable attention be-cause of their role in xenobiotic metabolism, both bio-activation and detoxification (4).
The overwhelming majority of eukaryotic P450s arelocalized in the endoplasmic reticulum. Only five typesof P450 enzymes have been clearly identified in eu-karyotic mitochondria and cloned to date. P450scc(CYP11A) is the cholesterol side-chain cleavage en-zyme that initiates steroid hormone synthesis in ver-tebrates. The CYP11B enzymes are the 11b hydroxy-lase/18 hydroxylases found in adrenals and gonads.CYP24 is a vitamin D3 24-hydroxylase and CYP27A isa vitamin D3 25-hydroxylase and is also involved in thebiosynthesis of bile acids whereas CYP27B is the 25-hydroxyvitamin D3 1a-hydroxylase (4, 5). All of thesemitochondrial enzymes have known biosynthetic func-tions, and all have been documented solely in verte-brate species.
Whereas microsomal P450s rely on the flavoproteinNADPH:cytochrome P450 reductase as well as cyto-chrome b5 for their supply of reducing equivalents,mitochondrial P450s rely on two different proteins, a
1 The sequence data are deposited in GenBank under the Acces-sion Nos. U86618 (CYP12A1), U94698 (CYP12A2), and U94699(CYP12A3).
2 Present address: Monsanto Company, 700 Chesterfield ParkwayNorth, St. Louis, MO 63198.
3 To whom correspondence should be addressed at Department ofEntomology, University of Arizona, Forbes 410, PO Box 210036,Tucson, AZ 85721-0036. Fax: (520) 626-8058; E-mail: [email protected].
4 Abbreviations used: P450, cytochrome P450; RT, reverse tran-scription; DTT, dithiothreitol.
0003-9861/98 $25.00 231Copyright © 1998 by Academic PressAll rights of reproduction in any form reserved.
ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS
Vol. 359, No. 2, November 15, pp. 231–240, 1998Article No. BB980901

ferredoxin and an NADPH-dependent ferredoxin re-ductase (e.g., adrenodoxin and adrenodoxin reductasein adrenals). The electron transfer chain of mitochon-drial P450s is thus homologous to that found in somesoluble bacterial P450 systems (e.g., P450cam, puti-daredoxin, and putidaredoxin reductase) (4). The dif-ference in electron transfer chain for mitochondrialand microsomal P450, as well as the apparent special-ization of vertebrate mitochondrial P450s in biosyn-thetic functions, suggests a fundamental difference be-tween microsomal and mitochondrial P450s. Yet phy-logenetic trees (1, 2, 6) show mitochondrial P450s to bea branch monophyletic with microsomal P450s and notrelated to the bacterial P450s that utilize a homologouselectron transfer chain. Furthermore, there are reportsof the existence of mammalian mitochondrial P450sthat are biochemically and immunologically similar tomicrosomal P450 forms and that metabolize xenobiot-ics; these rely on mitochondrial electron donors but aretruncated forms of microsomal P450s transiting to theinner membrane of the mitochondria by a novel mech-anism (7, 8). The presence of P450s in insect mitochon-dria has been inferred from biochemical data on en-zyme activities related to the biosynthesis of ecdys-teroids, the insect molting hormones (9), and several“mitochondrial-like” P450 sequences from inverte-brates have been reported (10–12). Thus, more infor-mation is needed to understand the origin and evolu-tion of mitochondrial P450s in general and to provideunequivocal evidence on the subcellular localization ofa putative insect mitochondrial P450 in particular. Wefelt that the cloning and biochemical characterizationof a nonvertebrate mitochondrial P450 would be animportant step in this direction. We report here thecloning of three mitochondrial P450s from the house flyand show that one of them, CYP12A1, is capable ofmetabolizing xenobiotics and is overexpressed in aninsecticide-resistant strain.
MATERIALS AND METHODS
Amplification of partial P450 cDNAs. Poly(A)1 RNA was ex-tracted from 4-day-old larvae of the diazinon-resistant “Rutgers”strain of the house fly, Musca domestica. About 1 mg of poly(A)1 RNAwas reverse transcribed using Superscript RNase H- Reverse Tran-scriptase (Life Technologies, Inc.) and an 18-mer oligo(dT) primer.The mRNA–cDNA duplex was used as template for PCR with Am-pliTaq polymerase (Perkin–Elmer) and a pair of degenerate oligonu-cleotide primers for two conserved regions of known mitochondrialP450s. The forward primer (for KETLRL(Y/H)P) was 59-AA(A/G)GA(A/G) ACI (C/T)TI (A/C)GI (C/T)T(C/G/T) (C/T)A(C/T) CC-39. Thereverse primer (for C(L/V)GRR(I/L)AE) was 59-TCI GCI A(A/G/T)IC(G/T)I C(G/T)(A/G/T) CCI A(A/C/G)(A/G) CA-39. After separation ofthe PCR products by electrophoresis on a 2% agarose gel, an area ofthe gel corresponding to DNA of the expected size (260–300 bp) andcontaining two visible bands was excised. The DNA was purifiedusing the Sephaglas BandPrep kit (Pharmacia Biotech Inc.), clonedinto pCR II (Invitrogen), and several clones were sequenced with theSequenase Version 2.0 DNA sequencing kit (U.S. Biochemical Corp.).
cDNA library construction and screening. Three RT-PCR cDNAclones encoding P450s (dTA2, dTa19, and dTa24) were used forscreening a house fly larval cDNA library (approximately 1 3 106
plaques) constructed with poly(A)1 RNA (6 mg) from 4-day old larvaeof the Rutgers strain using ZAP-cDNA Synthesis Kit (Stratagene).The 32P-labeled probes were generated using a Prime-It II kit (Strat-agene). Inserts were excised, recircularized into pBluescript plas-mids, and sequenced. Nucleotide sequences were analyzed with theBLAST program on the NCBI Blast Server (13).
Vector for heterologous expression of CYP12A1. The plasmid pC-Wori1 (kindly provided by Dr. F.W. Dahlquist, University of Oregon,Eugene, OR) was used for the expression of CYP12A1. This expres-sion plasmid contains an NdeI site adjacent to the ATG startingcodon. The 59-end of the CYP12A1 coding region was modified byPCR mutagenesis with Pfu polymerase (Stratagene) to introduce anNdeI site upstream of the ATG codon and to replace the second Ile(ATA) codon by Ala (GCT), a preferred second codon for expression ofthe lacZ gene (14). To facilitate purification of recombinant CYP12A1by immobilized metal affinity chromatography, the 39 end of theCYP12A1 cDNA was modified to introduce six His codons in front ofthe stop codon and a HindIII site downstream of the stop codon. Thesequence of the mutated cDNA was verified and cloned into NdeI/HindIII-digested pCWori1 vector to form a pC12His expression plas-mid which was used to transform the Escherichia coli strain BL21 forexpression experiments.
Expression in E. coli and purification of house fly CYP12A1. Anovernight culture of E. coli strain BL21 (pC12His) in Terrific Broth(15) modified with a mixture of trace elements (16) and containing200 mg ampicillin/ml was diluted 100-fold into the same medium.The flasks were shaken at 125 rpm and 30°C until the A600 of theculture was 1.0–1.5. Isopropyl-b-D-thiogalactopyranoside and d-ami-nolevulinic acid were then added to 0.1 mM and 0.5 mM, respec-tively, and the flasks were shaken at 125 rpm and 30°C for 16–20 h.The cells were pelleted, washed with 0.1 M Tris, pH 7.4, containing0.1 M NaCl, resuspended in 0.1 M Tris–HCl, pH 7.4, and storedfrozen at 280°C overnight. After thawing, phenylmethylsulfonylfluoride was added to a final concentration of 2 mM, and the cellswere sonicated on ice. The suspension was centrifuged at 100,000gfor 1 h, and the pellet was resuspended in 0.1 M Tris–HCl, pH 7.5,containing 20% glycerol to give a protein concentration of 3 mg/ml.Emulgen 911 (Kao Atlas, Japan) was then added to a final concen-tration of 1% (w/v), and the suspension was stirred for 1 h on ice. Thesuspension was centrifuged at 100,000g for 1 h, and the resultingsupernatant was loaded onto a His-bind (Novagen) column equili-brated with 20 mM Tris–HCl, pH 7.5, containing 5 mM imidazole,0.5 M NaCl, and 20% (v/v) glycerol (buffer A). The column waswashed with buffer A containing 0.1% (w/v) Emulgen 911, then with20 mM Tris–HCl, pH 7.5, containing 25 mM imidazole, 0.5 M NaCl,20% (v/v) glycerol, and 0.1% (w/v) Emulgen 911. The detergent wasreplaced by washing the column with 20 mM Tris–HCl, pH 7.5,containing 15 mM imidazole, 0.5 M NaCl, 20% glycerol, and 0.3%(w/v) sodium cholate. The protein was eluted with 20 mM Tris–HCl,pH 7.5, containing 200 mM imidazole, 0.5 M NaCl, 20% glycerol, and0.3% sodium cholate, and dialyzed against 0.1 M Tris–HCl, pH 7.5,containing 20% glycerol, 0.3% sodium cholate, 0.5 mM EDTA, and 1mM DTT, and stored frozen at 280°C.
Production and purification of antibodies. Antibodies againstCYP12A1 were raised in male white rabbits as described earlier (17)using antigen fixed in 10% polyacrylamide gel after SDS–PAGE (18),and the IgG fraction was isolated from the antiserum (19). Immuno-affinity purification of antibodies was performed using CYP12A1immobilized on CNBr-activated Sepharose (Pharmacia).
Electron microscope immunocytochemistry. Salivary glands fromlate 3rd instar larvae were fixed for 2 h in 0.2% glutaraldehyde:4%paraformaldehyde in 0.05 M sodium cacodylate buffer, pH 7.2, andembedded in LR White (20). Ultrathin sections on formvar-coated
232 GUZOV ET AL.

nickel grids were immunostained using affinity-purified primaryantibody (anti-CYP12A1) and a secondary antibody (goat anti-rabbitIgG)-coated 10-nm colloidal gold (AuroProbe EM GAR G10, Amer-sham). Grids with sections were treated according to the manufac-turer’s recommendations and then stained with 2% aqueous uranylacetate for 5 min. Sections were examined in a Philips CM 12electron microscope.
Enzymatic activity assays. The CYP12A1 activity was assayedwith at least two preparations of the purified enzyme in 50 mM Hepes,pH 7.4, containing 15 mM MgCl2, 0.1 mM EDTA (assay buffer), at 30°Cin a final volume of 0.1–1 ml. The reaction mixture contained substrate,NADPH-regenerating system (200 mM NADPH, 2 mM glucose 6-phos-phate, 3 units/ml glucose-6-phosphate dehydrogenase), CYP12A1, ad-renodoxin, and adrenodoxin reductase (0.2–0.5 mM, 1–5 mM, and 0.2–1mM, respectively). Prior to assay, the proteins were preincubated for 15min on ice in the presence of 0.3 mg/ml L-a-dilauroyl-sn-glycero-3-phosphocholine and 0.025% (w/v) Emulgen 911 in assay buffer (enzymemixture). One vol of enzyme mixture was then incubated for 2 min at30°C with 8 vol of assay buffer, and the reaction was started by theaddition of 1 vol of NADPH-regenerating system.
Aldrin, heptachlor, diazinon, lauric acid, and cholesterol (100 mM)metabolism were analyzed as described (21–24). N-Demethylation ofamitraz (1 mM nominal) was assayed by formaldehyde productionusing the Nash reagent (25). Products of progesterone and testoster-one (200 mM) were identified by HPLC-MS (Analytical Core, South-west Environmental Health Sciences Center) and quantitated byTLC (26). 7-Alkoxycoumarins were obtained commercially or synthe-sized by the Synthetic Core, Southwest Environmental Health Sci-ences Center. Dealkylation to 7-hydroxycoumarin formed was quan-titated as described (27). Hydroprene and methoprene metabolismwas analyzed by GC-FID.
For metabolism of ecdysteroids the reaction mixture contained100–200 mM substrate in a final volume 0.5 ml. The reaction prod-ucts were analyzed by reversed-phase HPLC.
Expression of CYP12A1 in various strains of the house fly. mRNAwas isolated from adult abdomens as described (28). A 0.25-kb-longriboprobe was generated by subcloning a ClaI/XhoI-fragment (encodingthe N-terminus of the protein) of the CYP12A1 cDNA into pBluescriptII vector followed by XhoI digestion and 32P-labeling using MAXIscriptin vitro transcription kit (Ambion) with T3 RNA Polymerase. Ribonu-clease protection assay was performed using HybSpeed RPA kit (Am-bion) with 2 mg mRNA per reaction. Data were quantitated using aPhosphorImager (Molecular Dynamics) and expressed as relativemRNA level normalized to that of the Rutgers strain.
Other procedures. Recombinant bovine adrenodoxin and adreno-doxin reductase were purified from corresponding E. coli expressionstrains as described before (29, 30). Concentrations of adrenodoxinand adrenodoxin reductase were determined from absorbance spec-tra (31, 32). Enzymatic reduction of CYP12A1 was measured bydifference spectrophotometry at 4°C. The assay buffer (see above)was bubbled with helium. Proteins were mixed with buffer in thepresence of an oxygen-scavenging system (0.14 mM glucose oxidase,3000 U catalase/ml, and 100 mM glucose). The sample cuvette wasbubbled with CO. The reaction was started by addition of NADPH(200 mM) and NADPH-regenerating system to each cuvette. Differ-ence spectra from 390 to 500 nm were then recorded (33).
RESULTS
Cloning of House Fly Mitochondrial P450 cDNAs
Conserved sequences around the putative ferredox-in- (34) and heme-binding sites of known mitochondrialP450s were chosen to design degenerate oligonucleo-tide primers for RT-PCR from larval mRNA of thehouse fly, M. domestica. Cloning of the PCR-products
and sequencing of several individual clones revealedthe presence of three different P450 fragments show-ing significant sequence identity to known mitochon-drial P450s. These were used to screen a house flylarval cDNA library yielding two full-length cDNAsand a partial cDNA. The deduced amino acid sequencesof these cDNAs (Fig. 1) were highly similar (75–79%identity to each other) but had only ,22% identity toCYP6A1 and to other insect P450s then available inGenBank. These sequences have been assigned as thefirst members of new P450 family named CYP12, andthe GenBank Accession Nos. are CYP12A1–U86618;CYP12A2–U94698, and CYP12A3–U94699. Sequencecomparison with known microsomal and mitochondrialP450s demonstrated that CYP12 proteins cluster in aphylogenetic tree with known mitochondrial P450sfrom vertebrate species and with three putative mito-chondrial P450 sequences from invertebrates, CYP10,CYP12B1, and an unnamed P450 sequence from Cae-norhabditis elegans (Fig. 2). The plethora of P450 se-quences available made it necessary, for clarity, tochoose a subset for inclusion in the analysis. However,the topology (Fig. 2) of the cladogram demonstrating amonophyletic origin of mitochondrial P450s was ob-tained by both Neighbor Joining and UPGMA methodsand was not affected by the inclusion of additionalparalogous sequences. The CYP12A1 cDNA was themost abundant in our house fly larval cDNA libraryand was chosen for heterologous expression in E. coliand functional characterization.
Expression in E. coli and Purification of CYP12A1
A plasmid for expression of CYP12A1 in E. coli wasconstructed. Even in the presence of the heme precur-sor d-aminolevulinic acid in the growth medium, theexpression level was moderate (60–80 nmol P450/Lculture) making purification of P450 by conventionalmethods difficult. Therefore, the CYP12A1 cDNA wasmutagenized by the addition of six His (CAT) codons atthe 39-end of the coding sequence. The His-tagged pro-tein (expression level 60–100 nmol/L culture) was pu-rified using immobilized metal affinity chromatogra-phy to near electrophoretic homogeneity (data notshown) with a specific P450 content of about 5 nmol/mgprotein, indicating that a significant part of the proteinwas isolated as apoprotein.
Subcellular Localization
Analysis of CYP12A1 expression at various develop-mental stages by Western blotting with anti-CYP12A1antibodies revealed that the highest level of CYP12A1immunoreactive proteins are seen during the late 3rdlarval instar with high expression levels in the Mal-pighian tubules and salivary glands. Ribonuclease pro-
233INSECT MITOCHONDRIAL P450

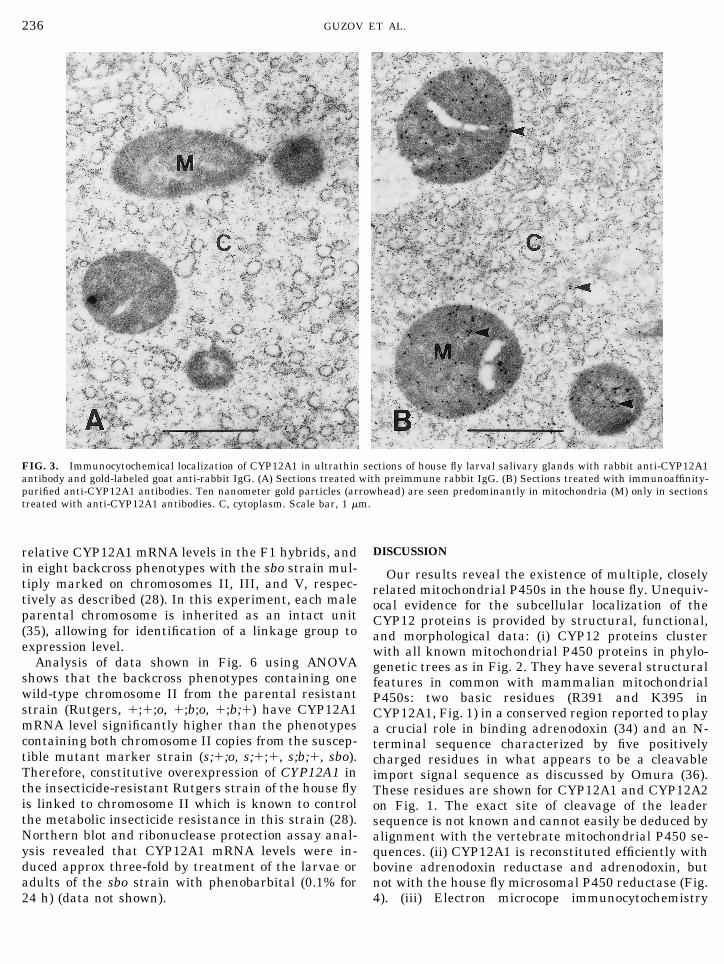
tection assays confirmed that CYP12A1 is expressed inthe fat body and in the gut of 4-day-old larvae (V. M.Guzov and R. Feyereisen, unpublished observations).Salivary glands of late 3rd instar larvae were used forimmunochemical localization of CYP12A1 because thistissue is easier to prepare for this technique. On sec-tions treated with anti-CYP12A1, gold particles werelocalized primarily in mitochondria (Fig. 3). The lowdensity of gold particles in the cytoplasm may repre-sent nascent CYP12A1 polypeptide synthesized in the
endoplasmic reticulum and not yet transferred to mi-tochondria, or alternatively, cross-reaction with otherP450s present in the endoplasmic reticulum. No goldparticles were observed in control sections treated withpre-immune IgG (Fig. 3).
Enzymatic Reduction of CYP12A1
Mammalian mitochondrial P450s depend on two re-dox partners, an FAD-containing ferredoxin reductase
FIG. 1. Alignment of the deduced amino acid sequences of CYP12A1, 2, and 3. The 1 represent positive charges in the putativemitochondrial targeting sequences of CYP12A1 and CYP12A2, the arrows represent the position of the oligonucleotides used for PCR cloning,and the conserved, underlined basic residues are orthologous to those of the putative adrenodoxin binding site of mammalian mitochondrialP450 proteins (34).
234 GUZOV ET AL.

and an iron-sulfur ferredoxin to transfer electrons fromNADPH to P450. Insect homologs of these mitochon-drial proteins are not yet available. Therefore, we usedthe mammalian mitochondrial electron transfer part-ners (recombinant bovine adrenodoxin reductase andadrenodoxin) to reconstitute a functional electron-transfer chain with house fly CYP12A1.
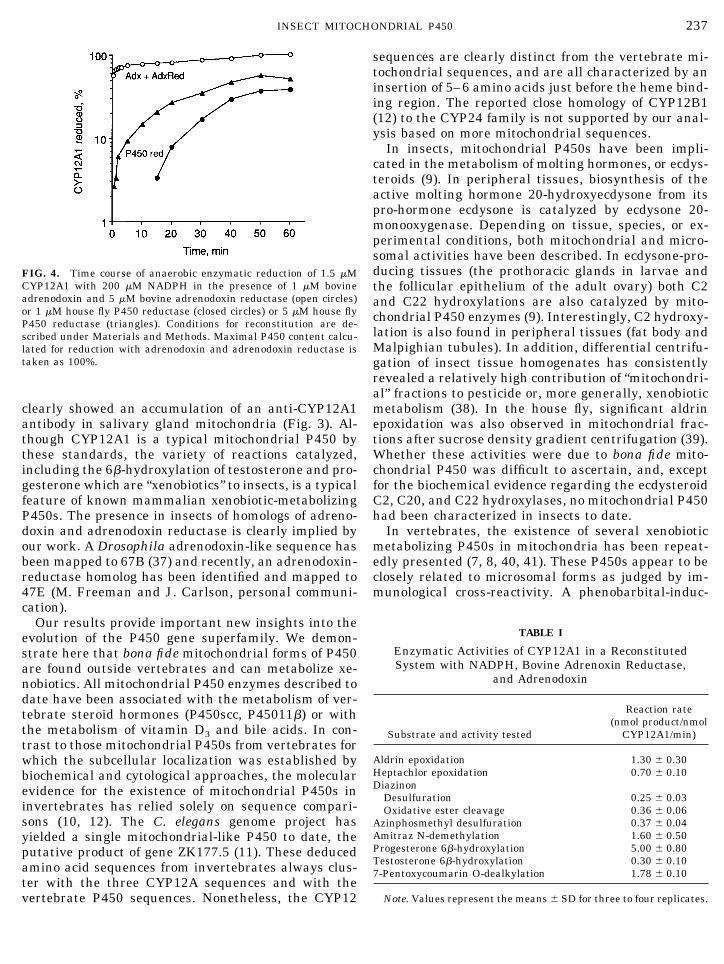
Enzymatic reduction of recombinant CYP12A1 un-der anaerobic conditions was monitored by formationof the carbonyl–ferrous complex with a peak at 450 nmin CO-difference spectra. Two types of NADPH-depen-dent electron partners were used to reduce CYP12A1:(1) recombinant bovine mitochondrial adrenodoxin re-ductase and adrenodoxin and (2) recombinant house flymicrosomal P450 reductase. Spectra were recorded at4°C because rates of reduction were too fast to allowaccurate readings at room temperature. Figure 4shows that over 70% of CYP12A1 was reduced withbovine mitochondrial electron partners within 2 min,while reduction with the microsomal electron partner,P450 reductase, occurred very slowly, even though thisis a protein from the same species (house fly). Thus,house fly mitochondial P450 CYP12A1 can be effi-
ciently reduced by NADPH in the presence of bovinemitochondrial electron transfer partners. This recon-stituted system was therefore used to study the sub-strate specificity of CYP12A1.
Substrate Specificity of CYP12A1
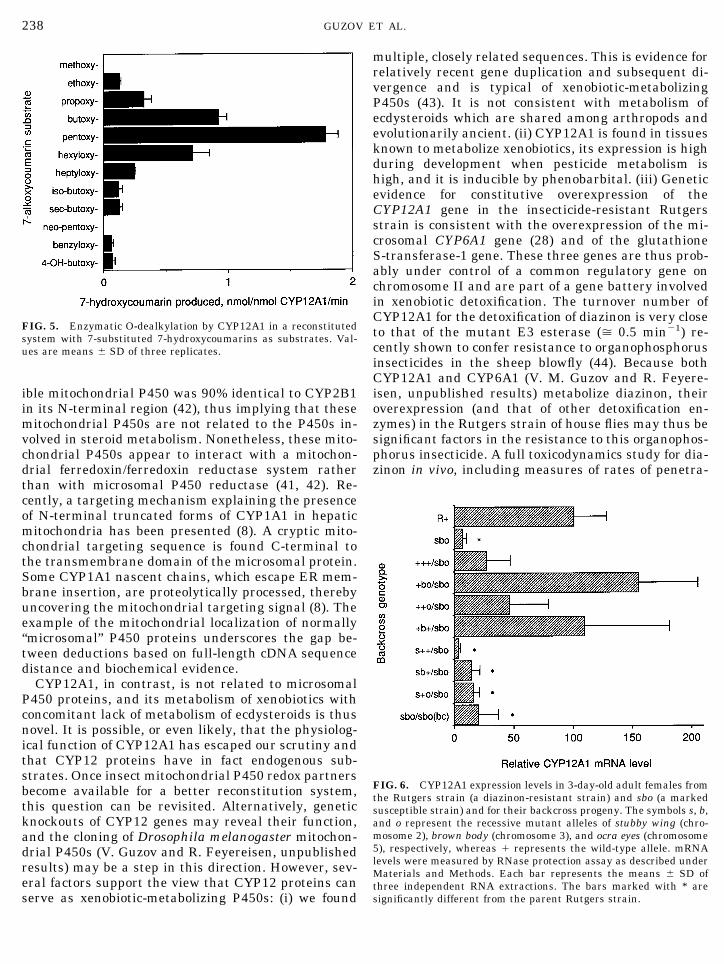
Table I summarizes the nature and extent ofCYP12A1-dependent metabolism in our reconstitutedsystem. CYP12A1 metabolizes a number of insecticidesand several common P450 substrates which are allxenobiotics for insects. It catalyzes different types ofchemical reactions such as hydroxylation, epoxidation,N-demethylation, O-dealkylation, desulfuration, andoxidative ester cleavage. HPLC-MS showed that 6bhydroxylation was the major pathway of testosteroneand progesterone metabolism. Hydroxylation at 16band at an unknown position were minor pathways oftestosterone metabolism, whereas 16a, 11a, and 17awere minor sites of progesterone hydroxylation.
The O-dealkylation of 7-substituted 7-hydroxycou-marins was studied in greater detail (Fig. 5). The high-est activity was observed with 7-pentoxycoumarin withlower turnover numbers for smaller and larger sub-stituents. Both geometry and hydrophobicity of thesubstituent was important for enzymatic activity, be-cause radical branching or incorporation of a hydroxylgroup drastically reduced O-dealkylation activity. In-terestingly, the alkoxyresorufin analogs of the couma-rins tested above, including pentoxyresorufin, were notmetabolized significantly under our conditions. Also,little or no (, 0.1 nmol/nmol CYP12A1/min) metabo-lism of the pesticides DDT, cypermethrin, or propoxur,or of aminopyrine and benzphetamine was observed.
Several potential endogenous substrates were as-sayed. These included cholesterol, lauric acid, ecdys-teroids (ecdysone, 20-hydroxyecdysone, 2-deoxyecdys-one, 2b,3b,14a-trihydroxy-5b-cholest-7-en-6-one, and2b,3b-dihydroxy-5b-cholest-7-en-6-one), and the ana-logs of juvenile hormone, methoprene, and hydroprene.No activity was detected with any of these compounds.In addition, whereas steroids such as progesterone andpesticides such as aldrin gave type I spectral interac-tions with CYP12A1, no type I spectrum was detect-able with cholesterol or ecdysteroids (data not shown).
Overexpression of CYP12A1 in an Insecticide-Resistant Strain
Ribonuclease protection assay of mRNA extractedfrom adult flies of the insecticide-resistant Rutgers andsusceptible sbo strains revealed that CYP12A1 is con-stitutively overexpressed in the Rutgers strain by ap-prox five-fold (Fig. 6). In order to investigate the ge-netic control of this elevated expression, we assayed
FIG. 2. Phylogenetic tree of representative P450s. The three housefly CYP12 sequences were aligned with the sequences of P450 pro-teins representing vertebrate mitochondrial P450s (rat CYP11A andB, human CYP24 and CYP27A and B), the sequences of the putativemitochondrial CYP10 from the pond snail, Lymnaea stagnalis,CYP12B1 from Drosophila acanthoptera, and the putative product ofthe ZK177.5 gene from C. elegans as well as with microsomal P450sequences (house fly CYP6A1, human CYP3A4, tobacco hornwormCYP4M2, mouse CYP1A1 and rat CYP2B1), and the bacterialP450cam (CYP101). The Clustal alignment was used to generate atree by the Neighbour-Joining method.
235INSECT MITOCHONDRIAL P450
relative CYP12A1 mRNA levels in the F1 hybrids, andin eight backcross phenotypes with the sbo strain mul-tiply marked on chromosomes II, III, and V, respec-tively as described (28). In this experiment, each maleparental chromosome is inherited as an intact unit(35), allowing for identification of a linkage group toexpression level.
Analysis of data shown in Fig. 6 using ANOVAshows that the backcross phenotypes containing onewild-type chromosome II from the parental resistantstrain (Rutgers, 1;1;o, 1;b;o, 1;b;1) have CYP12A1mRNA level significantly higher than the phenotypescontaining both chromosome II copies from the suscep-tible mutant marker strain (s;1;o, s;1;1, s;b;1, sbo).Therefore, constitutive overexpression of CYP12A1 inthe insecticide-resistant Rutgers strain of the house flyis linked to chromosome II which is known to controlthe metabolic insecticide resistance in this strain (28).Northern blot and ribonuclease protection assay anal-ysis revealed that CYP12A1 mRNA levels were in-duced approx three-fold by treatment of the larvae oradults of the sbo strain with phenobarbital (0.1% for24 h) (data not shown).
DISCUSSION
Our results reveal the existence of multiple, closelyrelated mitochondrial P450s in the house fly. Unequiv-ocal evidence for the subcellular localization of theCYP12 proteins is provided by structural, functional,and morphological data: (i) CYP12 proteins clusterwith all known mitochondrial P450 proteins in phylo-genetic trees as in Fig. 2. They have several structuralfeatures in common with mammalian mitochondrialP450s: two basic residues (R391 and K395 inCYP12A1, Fig. 1) in a conserved region reported to playa crucial role in binding adrenodoxin (34) and an N-terminal sequence characterized by five positivelycharged residues in what appears to be a cleavableimport signal sequence as discussed by Omura (36).These residues are shown for CYP12A1 and CYP12A2on Fig. 1. The exact site of cleavage of the leadersequence is not known and cannot easily be deduced byalignment with the vertebrate mitochondrial P450 se-quences. (ii) CYP12A1 is reconstituted efficiently withbovine adrenodoxin reductase and adrenodoxin, butnot with the house fly microsomal P450 reductase (Fig.4). (iii) Electron microcope immunocytochemistry
FIG. 3. Immunocytochemical localization of CYP12A1 in ultrathin sections of house fly larval salivary glands with rabbit anti-CYP12A1antibody and gold-labeled goat anti-rabbit IgG. (A) Sections treated with preimmune rabbit IgG. (B) Sections treated with immunoaffinity-purified anti-CYP12A1 antibodies. Ten nanometer gold particles (arrowhead) are seen predominantly in mitochondria (M) only in sectionstreated with anti-CYP12A1 antibodies. C, cytoplasm. Scale bar, 1 mm.
236 GUZOV ET AL.
clearly showed an accumulation of an anti-CYP12A1antibody in salivary gland mitochondria (Fig. 3). Al-though CYP12A1 is a typical mitochondrial P450 bythese standards, the variety of reactions catalyzed,including the 6b-hydroxylation of testosterone and pro-gesterone which are “xenobiotics” to insects, is a typicalfeature of known mammalian xenobiotic-metabolizingP450s. The presence in insects of homologs of adreno-doxin and adrenodoxin reductase is clearly implied byour work. A Drosophila adrenodoxin-like sequence hasbeen mapped to 67B (37) and recently, an adrenodoxin-reductase homolog has been identified and mapped to47E (M. Freeman and J. Carlson, personal communi-cation).
Our results provide important new insights into theevolution of the P450 gene superfamily. We demon-strate here that bona fide mitochondrial forms of P450are found outside vertebrates and can metabolize xe-nobiotics. All mitochondrial P450 enzymes described todate have been associated with the metabolism of ver-tebrate steroid hormones (P450scc, P45011b) or withthe metabolism of vitamin D3 and bile acids. In con-trast to those mitochondrial P450s from vertebrates forwhich the subcellular localization was established bybiochemical and cytological approaches, the molecularevidence for the existence of mitochondrial P450s ininvertebrates has relied solely on sequence compari-sons (10, 12). The C. elegans genome project hasyielded a single mitochondrial-like P450 to date, theputative product of gene ZK177.5 (11). These deducedamino acid sequences from invertebrates always clus-ter with the three CYP12A sequences and with thevertebrate P450 sequences. Nonetheless, the CYP12
sequences are clearly distinct from the vertebrate mi-tochondrial sequences, and are all characterized by aninsertion of 5–6 amino acids just before the heme bind-ing region. The reported close homology of CYP12B1(12) to the CYP24 family is not supported by our anal-ysis based on more mitochondrial sequences.
In insects, mitochondrial P450s have been impli-cated in the metabolism of molting hormones, or ecdys-teroids (9). In peripheral tissues, biosynthesis of theactive molting hormone 20-hydroxyecdysone from itspro-hormone ecdysone is catalyzed by ecdysone 20-monooxygenase. Depending on tissue, species, or ex-perimental conditions, both mitochondrial and micro-somal activities have been described. In ecdysone-pro-ducing tissues (the prothoracic glands in larvae andthe follicular epithelium of the adult ovary) both C2and C22 hydroxylations are also catalyzed by mito-chondrial P450 enzymes (9). Interestingly, C2 hydroxy-lation is also found in peripheral tissues (fat body andMalpighian tubules). In addition, differential centrifu-gation of insect tissue homogenates has consistentlyrevealed a relatively high contribution of “mitochondri-al” fractions to pesticide or, more generally, xenobioticmetabolism (38). In the house fly, significant aldrinepoxidation was also observed in mitochondrial frac-tions after sucrose density gradient centrifugation (39).Whether these activities were due to bona fide mito-chondrial P450 was difficult to ascertain, and, exceptfor the biochemical evidence regarding the ecdysteroidC2, C20, and C22 hydroxylases, no mitochondrial P450had been characterized in insects to date.
In vertebrates, the existence of several xenobioticmetabolizing P450s in mitochondria has been repeat-edly presented (7, 8, 40, 41). These P450s appear to beclosely related to microsomal forms as judged by im-munological cross-reactivity. A phenobarbital-induc-
FIG. 4. Time course of anaerobic enzymatic reduction of 1.5 mMCYP12A1 with 200 mM NADPH in the presence of 1 mM bovineadrenodoxin and 5 mM bovine adrenodoxin reductase (open circles)or 1 mM house fly P450 reductase (closed circles) or 5 mM house flyP450 reductase (triangles). Conditions for reconstitution are de-scribed under Materials and Methods. Maximal P450 content calcu-lated for reduction with adrenodoxin and adrenodoxin reductase istaken as 100%.
TABLE I
Enzymatic Activities of CYP12A1 in a ReconstitutedSystem with NADPH, Bovine Adrenoxin Reductase,
and Adrenodoxin
Substrate and activity tested
Reaction rate(nmol product/nmol
CYP12A1/min)
Aldrin epoxidation 1.30 6 0.30Heptachlor epoxidation 0.70 6 0.10Diazinon
Desulfuration 0.25 6 0.03Oxidative ester cleavage 0.36 6 0.06
Azinphosmethyl desulfuration 0.37 6 0.04Amitraz N-demethylation 1.60 6 0.50Progesterone 6b-hydroxylation 5.00 6 0.80Testosterone 6b-hydroxylation 0.30 6 0.107-Pentoxycoumarin O-dealkylation 1.78 6 0.10
Note. Values represent the means 6 SD for three to four replicates.
237INSECT MITOCHONDRIAL P450
ible mitochondrial P450 was 90% identical to CYP2B1in its N-terminal region (42), thus implying that thesemitochondrial P450s are not related to the P450s in-volved in steroid metabolism. Nonetheless, these mito-chondrial P450s appear to interact with a mitochon-drial ferredoxin/ferredoxin reductase system ratherthan with microsomal P450 reductase (41, 42). Re-cently, a targeting mechanism explaining the presenceof N-terminal truncated forms of CYP1A1 in hepaticmitochondria has been presented (8). A cryptic mito-chondrial targeting sequence is found C-terminal tothe transmembrane domain of the microsomal protein.Some CYP1A1 nascent chains, which escape ER mem-brane insertion, are proteolytically processed, therebyuncovering the mitochondrial targeting signal (8). Theexample of the mitochondrial localization of normally“microsomal” P450 proteins underscores the gap be-tween deductions based on full-length cDNA sequencedistance and biochemical evidence.
CYP12A1, in contrast, is not related to microsomalP450 proteins, and its metabolism of xenobiotics withconcomitant lack of metabolism of ecdysteroids is thusnovel. It is possible, or even likely, that the physiolog-ical function of CYP12A1 has escaped our scrutiny andthat CYP12 proteins have in fact endogenous sub-strates. Once insect mitochondrial P450 redox partnersbecome available for a better reconstitution system,this question can be revisited. Alternatively, geneticknockouts of CYP12 genes may reveal their function,and the cloning of Drosophila melanogaster mitochon-drial P450s (V. Guzov and R. Feyereisen, unpublishedresults) may be a step in this direction. However, sev-eral factors support the view that CYP12 proteins canserve as xenobiotic-metabolizing P450s: (i) we found
multiple, closely related sequences. This is evidence forrelatively recent gene duplication and subsequent di-vergence and is typical of xenobiotic-metabolizingP450s (43). It is not consistent with metabolism ofecdysteroids which are shared among arthropods andevolutionarily ancient. (ii) CYP12A1 is found in tissuesknown to metabolize xenobiotics, its expression is highduring development when pesticide metabolism ishigh, and it is inducible by phenobarbital. (iii) Geneticevidence for constitutive overexpression of theCYP12A1 gene in the insecticide-resistant Rutgersstrain is consistent with the overexpression of the mi-crosomal CYP6A1 gene (28) and of the glutathioneS-transferase-1 gene. These three genes are thus prob-ably under control of a common regulatory gene onchromosome II and are part of a gene battery involvedin xenobiotic detoxification. The turnover number ofCYP12A1 for the detoxification of diazinon is very closeto that of the mutant E3 esterase (> 0.5 min21) re-cently shown to confer resistance to organophosphorusinsecticides in the sheep blowfly (44). Because bothCYP12A1 and CYP6A1 (V. M. Guzov and R. Feyere-isen, unpublished results) metabolize diazinon, theiroverexpression (and that of other detoxification en-zymes) in the Rutgers strain of house flies may thus besignificant factors in the resistance to this organophos-phorus insecticide. A full toxicodynamics study for dia-zinon in vivo, including measures of rates of penetra-
FIG. 5. Enzymatic O-dealkylation by CYP12A1 in a reconstitutedsystem with 7-substituted 7-hydroxycoumarins as substrates. Val-ues are means 6 SD of three replicates.
FIG. 6. CYP12A1 expression levels in 3-day-old adult females fromthe Rutgers strain (a diazinon-resistant strain) and sbo (a markedsusceptible strain) and for their backcross progeny. The symbols s, b,and o represent the recessive mutant alleles of stubby wing (chro-mosome 2), brown body (chromosome 3), and ocra eyes (chromosome5), respectively, whereas 1 represents the wild-type allele. mRNAlevels were measured by RNase protection assay as described underMaterials and Methods. Each bar represents the means 6 SD ofthree independent RNA extractions. The bars marked with * aresignificantly different from the parent Rutgers strain.
238 GUZOV ET AL.

tion and amounts of CYP12A1 per fly, would be neededto resolve this question. Nevertheless, this P450 geneis constitutively overexpressed in the resistant strain,and even if an endogenous function for CYP12A1 iseventually found, its activity towards xenobiotics mayhave a selective advantage. An artificial distinctionbetween “xenobiotic metabolism” and “physiologicalfunction” of P450 enzymes is no longer warranted, assuggested by Nebert (45). For example, the constitu-tively expressed microsomal CYP1B1 which is knownto metabolize diverse procarcinogens (46) has a (yetstill obscure) endogenous function revealed by map-ping mutations causing congenital glaucoma in hu-mans to the CYP1B1 gene (47). A similar “opportunis-tic” role for P450 in xenobiotic metabolism may beplayed by CYP12A1 and perhaps other mitochondrialP450s, even in vertebrates (48).
Further research will certainly uncover the mito-chondrial P450s involved in ecdysteroid biosynthesis inarthropods. In the pond snail, Lymnaea stagnalis, aP450 cDNA was obtained from the dorsal bodies, en-docrine glands that produce a gonadotropic hormone(10). Neither the product of these glands nor the func-tion of this P450 has been reported yet; its amino acidsequence places it along with CYP12 and the verte-brate mitochondrial P450s, so it may represent a mi-tochondrial P450 as well. The implication is that mito-chondrial P450s are involved in biosynthetic reactionsin both vertebrates and invertebrates and that thismay represent an ancestral feature of those P450s. Theselection pressure leading to the diversification of mi-tochondrial P450s and their involvment in xenobioticmetabolism will become clearer when more informa-tion regarding the phylogenetic distribution and func-tion of mitochondrial P450 becomes available.
ACKNOWLEDGMENTS
We thank Frederick W. Dahlquist (University of Oregon) for a giftof plasmid pCWori1, Michael R. Waterman (Vanderbilt University)for the adrenodoxin and adrenodoxin reductase E. coli expressionstrains, Martin Taylor for help with statistical analysis of the data,and Rosemarie Rosell for advice on immunogold staining. This workwas supported by National Institutes of Health Grants GM 39014 toR.F. and by NIEHS Center Grant ES 06694. It was presented in partat the International Symposium on P450 Biodiversity, Woods Hole,1995.
REFERENCES
1. Nelson, D., Koymans, L., Kamataki, T., Stegeman, J. J., Feyere-isen, R., Waxman, D., Waterman, M. R., Gotoh O., Coon, M. J.,Estabrook, R. W., Gunsalus, I. G., and Nebert, D. W. (1996)Pharmacokinetics 6, 1–42.
2. http://drnelson.utmem.edu/homepage.html3. Nebert, D. W. (1991) Mol. Endocrinol. 5, 1203–1214.4. Omura, T., Ishimura, Y., and Fuji-Kuriyama, Y. (Eds.) (1993)
Cytochrome P-450, Kodansha, Tokyo.
5. Shinki, T., Shimada, H., Wakino, S., Anasawa, H., Hayashi, M.,Saruta, T., DeLuca, H. F., and Suda, T. (1997) Proc. Natl. Acad.Sci. USA 94, 12920–12925.
6. Gotoh, O. (1993) in Cytochrome P-450 (Omura, T., Ishimura, Y.,and Fuji-Kuriyama, Y., Eds.), pp. 255–272, Kodansha, Tokyo.
7. Anandatheerthavarada, H. K., Addya, S., Dwivedi, R. S., Biswas,G., Mullick, J., and Avadhani, N. G. (1997) Arch. Biochem. Bio-phys. 339, 136–150.
8. Addya, S., Anandatheerthavarada, H. K., Biswas, G., Bhagwat,S. V., Mullick, J., and Avadhani, N. G. (1997) J. Cell. Biol. 139,589–599.
9. Feyereisen, R. (1999) Annu. Rev. Entomol. 44, 507–533.10. Teunissen, Y., Gerearts, W. P. M., van Herrehuizen, H., Planta,
R. J., and Joosse, J. (1992) J. Biochem. 112, 249–252.11. GenBank Accession No. U21321, gene ZK 177.5.12. Danielson, P. B., and Fogleman, J. C. (1997) Insect Biochem.
Mol. Biol. 27, 595–604.13. Gish, W., and States, D. J. (1993) Nat. Genet. 3, 266–272.14. Looman, A. C., Bodlaender, J., Comstock, L. J., Eaton, D.,
Thurani, R., de Boer, H. A., and Van Knippenberg, P. H. (1987)EMBO J. 6, 2489–2492.
15. Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989) MolecularCloning: A Laboratory Manual, 2nd ed., Cold Spring HarborLaboratory Press, Cold Spring Harbor, NY.
16. Bauer, S. D., and Shiloah, J. (1974) Biotechnol. Bioeng. 16,933–941.
17. Usanov S. A., Chernogolov, A. A., and Chashchin V. L. (1989)FEBS Lett. 255, 125–128.
18. Laemmli, U. K. (1970) Nature 227, 680–685.19. McKinney, M. M., and Parkinson, A. (1987) J. Immunol. Meth-
ods 96, 271–278.20. Newman, G. R. (1989) in Colloidal Gold: Principles, Methods,
and Applications (Hayat, M. A., Ed.), Vol. 2, pp. 47–73, AcademicPress, San Diego.
21. Andersen, J. F., Utermohlen, J. G., and Feyereisen, R. (1994)Biochemistry 33, 2171–2177.
22. Dunkov, B. C., Guzov, V. M., Mocelin, G., Shotkoski, F., Brun, A.,Amichot, M., ffrench-Constant, R. H., and Feyereisen, R. (1997)DNA Cell Biol. 11, 1345–1356.
23. Murataliev, M. B., and Feyereisen, R. (1996) Biochemistry 35,15029–15037.
24. Pikuleva, I. A., Mackman, R. L., Kagawa, N., Waterman M. R.,and Ortiz de Montellano, P. R.(1995) Arch. Biochem. Biophys.322, 189–197.
25. Werringloer, J. (1978) Methods Enzymol. 52C, 297–302.26. Waxman, D. J. (1991) Methods Enzymol. 206, 462–476.27. Matsubara, T., Otsubo, S., Yoshihara, E., and Touchi, A. (1983)
Jpn J. Pharmacol. 33, 41–56.28. Carino, F. A., Koener, J. F., Plapp, F. W., Jr., and Feyereisen, R.
(1994) Insect Biochem. Mol. Biol. 24, 411–418.29. Sagara, Y., Hara, T., Ariyasu, Y., Ando, F., Tokunaga, N., and
Horiuchi, T. (1992) FEBS Lett. 300, 208–212.30. Sagara, Y., Wada, A., Takata, Y., Waterman, M. R., Sekimizu,
K., and Horiuchi, T. (1993) Biol. Pharmacol. Bull. 16, 627–630.31. Huang, J. J., and Kimura, T. (1973) Biochemistry 12, 406 –
409.32. Chu, J.-W., and Kimura, T. (1973) J. Biol. Chem. 248, 2089–
2094.33. Omura, T., and Sato, R. (1964) J. Biol. Chem. 239, 2370–2378.34. Wada, A., and Waterman, M. R. (1992) J. Biol. Chem. 267,
22877–22882.
239INSECT MITOCHONDRIAL P450

35. Tsukamoto, M. (1964) Botyu-Kagaku 29, 51–59.36. Omura, T. (1993) in Cytochrome P450 (Schenkman, J. B., and
Greim, H., Eds.), pp. 61–69, Springer, Berlin.37. Pauli, D., and Tonka, C. H. (1987) J. Mol. Biol. 198, 235–240.38. Wilkinson, C. F., and Brattsten, L. B. (1972) Drug Metab. Rev. 1,
153–228.39. Feyereisen, R. (1983) Pest Biochem. Physiol. 19, 262–269.40. Niranjan, B. G., Rada, H., Shayiq, R. M., Jefcoate, C. R., and
Avadhani, N. G. (1988) J.Biol. Chem. 263, 575–580.41. Shayiq, R. M., and Avadhani, N. G. (1989) Biochemistry 28,
7546–7554.42. Shayiq, R. M., and Avadhani, N. G. (1990) Biochemistry 29,
866–873.
43. Gonzalez, F. J., and Nebert, D. W. (1990) Trends Genet. 6,182–186.
44. Newcomb, R. D., Campbell, P. M., Ollis, D. L., Cheah, E., Rus-sell, R. J., and Oakeshott, J. G.(1997) Proc. Natl. Acad. Sci. USA94, 7464–7468.
45. Nebert, D. W. (1991) Mol. Endocrinol. 5, 1203–1214.46. Shimada, T., Gillam, E. M., Sutter, T. R., Strickland, P. T.,
Guengerich, F. P., and Yamazaki, H. (1997) Drug Metab. Dispos.25, 617–622.
47. Stoilov, I., Akarsu, A. N., and Sarfarazi, M. (1997) Hum. Mol.Genet. 6, 641–647.
48. Lund, B. O., and Lund, J. (1995) J. Biol. Chem. 270, 20895–20897.
240 GUZOV ET AL.