melatonin protects the cytochrome p450 system through a novel antioxidant mechanism
TRANSCRIPT

Ma
MMa
Ub
a
ARRAA
KACFMRR
1
wpnrcea[ne
mS1f
0d
Chemico-Biological Interactions 185 (2010) 208–214
Contents lists available at ScienceDirect
Chemico-Biological Interactions
journa l homepage: www.e lsev ier .com/ locate /chembio int
elatonin protects the cytochrome P450 system throughnovel antioxidant mechanism
aría Eugenia Leteliera,∗, José Jara-Sandovala, Alfredo Molina-Berríosa,ario Faúndezb, Paula Aracena-Parksa, Felipe Aguileraa
Laboratory of Pharmacology and Toxicology, Department of Pharmacological and Toxicological Chemistry, School of Chemical and Pharmaceutical Sciences,niversidad de Chile, Santiago 8380492, ChileLaboratory of Pharmacology and Toxicology, Department of Pharmacy, School of Chemistry, Pontificia Universidad Católica de Chile, Santiago 7820436, Chile
r t i c l e i n f o
rticle history:eceived 20 November 2009eceived in revised form 2 March 2010ccepted 9 March 2010vailable online 17 March 2010
eywords:ntioxidantytochrome P450e3+/ascorbate
a b s t r a c t
Melatonin, an endogenous hormone, is used as an antioxidant drug in doses quite higher than the endoge-nous circulating levels of this hormone. Hepatic endoplasmic reticulum contains the cytochrome P450(CYP450) system, which catalyzes one biotransformation pathway of melatonin; this organelle is alsoone of the main sources of reactive oxygen species in cells. Therefore, we proposed that the antioxidantactivity of this hormone may have a biological relevance in the organelle where it is biotransformed. Toevaluate this postulate, we used Fe3+/ascorbate, an oxygen free radical generating system that leads tolipid peroxidation, loss of protein-thiol content, and activation of UDP-glucuronyltransferase in rat livermicrosomes. We found that mM concentrations of melatonin prevented all these oxidative phenom-ena. We also found that Fe3+/ascorbate leads to structural alterations in the CYP450 monooxygenase,
elatoninat liver microsomeseactive oxygen species
the enzyme that binds the substrate in the CYP450 system catalytic cycle, probably through direct oxida-tion of the protein, and also inhibited p-nitroanisole O-demethylation, a reaction catalyzed by the CYP450system. Notably, melatonin prevented both phenomena at �M concentrations. We provide evidence sug-gesting that melatonin may be oxidized by oxygen free radicals. Thus, we postulate that melatonin maybe acting as an oxygen free radical scavenger, and Fe3+/ascorbate-modified melatonin would be directlyprotecting the CYP450 system through an additional specific mechanism. Pharmacological relevance of
ssed
this phenomenon is discu. Introduction
Melatonin (N-acetyl-5-methoxytryptamine) is a hormonehich synthesis occurs in the suprachiasmatic nucleus of theineal gland and is controlled by the photoperiod [1,2]. Endoge-ous melatonin peaks at sub-nM levels in a cyclic manner and itselease is mainly implicated in the regulation of the sleep/wakeycle and hormonal regulation of sexual development [3]. Sev-ral preclinical and clinical studies suggest that melatonin may be
suitable antioxidant drug for the treatment of several diseases4], particularly in sleep-wake disorders [5,6], depression [7] andeurodegenerative disorders [8,9]. Noteworthy, when melatonin isxogenously administered, circulating levels reach orders of mag-
∗ Corresponding author at: Laboratory of Pharmacology and Toxicology, Depart-ent of Pharmacological and Toxicological Chemistry, Chemical and Pharmaceutical
ciences School, Universidad de Chile, Sergio Livingstone Pohlhammer (ex-Olivos)007, Independencia, Santiago 8380492, Chile. Tel.: +56 2 9782885;ax: +56 2 9782996.
E-mail address: [email protected] (M.E. Letelier).
009-2797/$ – see front matter © 2010 Elsevier Ireland Ltd. All rights reserved.oi:10.1016/j.cbi.2010.03.020
.© 2010 Elsevier Ireland Ltd. All rights reserved.
nitude different from those reached through endogenous release[10] and may elicit additional functions on systems other than theCNS [11].
Antioxidant function of melatonin has been associated to itscapacity to scavenge reactive oxygen/nitrogen species (ROS/RNS)[12]. In fact, several studies show that melatonin is more effectivein protecting against oxidative damage than other antioxidants,including vitamin E, glutathione and mannitol [13–16]. On theother hand, circulating concentrations of melatonin following oraladministration are capable to increase both mRNA levels and theactivities of antioxidant enzymes [17].
Orally administered melatonin is biotransformed by liverenzymes, such as the cytochrome P450 (CYP450) system, mainlylocated in the endoplasmic reticulum [18,19], and sulfotransferase[11,20], located in the cytosol. In addition to its contributionto melatonin catabolism, the CYP450 system is also one of the
main sources of reactive oxygen species generation in the cell[21–23]. Oxidative conditions may alter the biological functionof biomolecules occurring in the endoplasmic reticulum, such asphospholipids and enzymatic systems involved in the biotrans-formation of drugs [24–29]. Thus, we proposed that the hepatic
gical
ec
tgFiclUF
sbodsAlirt
c(id
2
2
n(gmnApw
2
teat1batA
2
[bonaHts
M.E. Letelier et al. / Chemico-Biolo
ndoplasmic reticulum is a sub-cellular organelle where melatoninan be metabolized and also exerts its antioxidant properties.
To evaluate this postulate, we studied the ability of melatonino prevent the oxidative damage elicited by Fe3+/ascorbate, an oxy-en free radical generating system through Haber–Weiss and/orenton reactions, on biomolecules of rat liver microsomes (exper-mental preparation enriched in endoplasmic reticulum). At mMoncentrations, melatonin inhibited the oxidation of microsomalipids and protein-thiol groups, and the oxidative activation ofDP-glucuronyltransferase (UDPGT), all phenomena elicited bye3+/ascorbate.
Remarkably, �M concentrations of melatonin protected thepectral properties of the CYP450 monooxygenase, enzyme thatinds the substrate in the first and limiting step of the catalytic cyclef the CYP450 system. In addition, melatonin also protected the O-emethylation of p-nitroanisole, a reaction catalyzed by the CYP450ystem, from the oxidative damage promoted by Fe3+/ascorbate.pparently, melatonin can be modified by Fe3+/ascorbate, most
ikely through oxidation; we provide evidence that melatonin, ints Fe3+/ascorbate-modified form, may bind to a crucial histidineesidue in the CYP450 monooxygenase, protecting this enzyme andhe catalytic activity of the CYP450 system.
Differences found in the concentrations at which melatoninompletely protects from oxidation liver microsomal biomoleculesmM) and, specifically, the CYP450 system (�M) raise the possibil-ty that melatonin exerts its protective antioxidant activity throughifferent mechanisms.
. Materials and methods
.1. Chemicals
Bovine serum albumin (BSA), CHELEX-100, 5,5′-dithiobis-(2-itrobenzoic acid) (Ellman’s reagent, DTNB), 2,4-dithiothreitolDTT), Folin–Ciocalteu’s reagent, glucose-6-phosphate (G-6-P),lucose-6-phosphate dehydrogenase, GSH, l-cysteine, l-histidine,elatonin, NADP, sodium ascorbate, p-nitroanisole (PNA), p-
itrophenol (PNP), and UDPGA were obtained from Sigma–Aldrich.ll other chemicals used were of analytical grade. All solutions wererepared in 50 mM phosphate buffer at pH 7.4, previously treatedith CHELEX-100.
.2. Animals
Adult male Sprague–Dawley rats (200–250 g), maintained athe vivarium of the School of Chemical and Pharmaceutical Sci-nces (Universidad de Chile, Santiago, Chile) were used. Rats werellowed free access to pelleted food, maintained with controlledemperature (22 ◦C) and photoperiod (lights on from 07:00 to9:00 h). All procedures were performed using protocols approvedy the Institutional Ethical Committee of the School of Chemicalnd Pharmaceutical Sciences, Universidad de Chile, and accordingo the guidelines of the Guide for the Care and Use of Laboratorynimals (NRC, USA).
.3. Liver microsomal fraction
Microsomal fractions were prepared according to Letelier et al.27]. Rats were fasted for 15 h with water ad libitum, and sacrificedy decapitation. Livers were perfused in situ with four volumesf 25 ml 0.9% (w/v) NaCl, excised, and placed on ice. All homoge-
ization and fractionation procedures were performed at 4 ◦C andll centrifugations were performed using either a Suprafuge 22eraeus centrifuge or an XL-90 Beckmann ultracentrifuge. Liverissue (9–11 g wet weight), devoid of connective and vascular tis-ue, was homogenized with five volumes of 0.154 M KCl, with eight
Interactions 185 (2010) 208–214 209
strokes in a Dounce Wheaton B homogenizer. Homogenates werecentrifuged at 9000 × g for 15 min and sediments were discarded.Supernatants were then centrifuged at 105,000 × g for 60 min. Pel-lets (microsomes) were stored at −80 ◦C until use. Total microsomalprotein was determined according to Lowry et al. [30].
2.4. Microsomal lipid peroxidation
The extent of microsomal lipid peroxidation was estimated bydetermining TBARS, according to Letelier et al. [27]. Microsomes(1 mg protein/mL) were pre-incubated with increasing concentra-tions of melatonin (up to 1 mM) in 50 mM phosphate buffer, pH7.4, for 15 min at 25 ◦C. Then, mixtures were further incubated with600 �M FeCl3 and 1 mM sodium ascorbate for 20 min at 37 ◦C withconstant agitation. Blanks contained all the reagents but micro-somal protein. Proteins were precipitated by the addition of TCAto 6.7% (w/v) final concentration; samples were then centrifugedat 10,000 × g for 10 min in a Suprafuge 22 Heraeus centrifuge andsupernatants were collected. Thiobarbituric acid was added to thesupernatants to final 0.5% (w/v) and mixtures were incubatedfor 1 h at 50 ◦C under constant agitation. Following incubation,absorbance at 532 nm of samples was determined in a UV3 Uni-cam UV–VIS spectrophotometer, and TBARS were estimated usingthe ε532 = 156 mM−1 cm−1, and expressed as nmol TBARS/min/mgprotein.
2.5. Determination of microsomal thiol content
Thiol groups were titrated with DTNB as described by Letelieret al. [27]. For protection experiments, microsomes (1 mg pro-tein/mL) were pre-incubated with 0.1 or 1 mM melatonin in 50 mMphosphate buffer, pH 7.4 for 15 min at 25 ◦C. Then, mixtures werefurther incubated with 600 �M FeCl3 and 1 mM sodium ascorbate,for 30 min at 37 ◦C with constant agitation. Blanks contained allreagents but microsomal protein. For reversion experiments, mela-tonin (0.1 or 1 mM) was added following incubation of microsomeswith Fe3+ and ascorbate, under identical conditions. After incuba-tions, microsomal thiol content was titrated with 0.6 mM DTNBfor 1 h at 37 ◦C. Samples were then centrifuged at 10,000 × g for10 min in a Suprafuge 22 Heraeus centrifuge and supernatants werecollected. Absorbance of supernatants was determined at 412 nmin a UV3 Unicam UV–VIS spectrophotometer and thiol contentwas estimated on the basis of the equimolar apparition of 5-thio-2-nitrobenzoic acid (ε410 = 13,600 M−1 cm−1) and is presented asnmol thiols/min/mg protein.
2.6. Assay of UDPGT activity
This activity was determined using p-nitrophenol as substrate,according to the method described by Letelier et al. [27]. Micro-somes (2 mg protein/mL) were pre-incubated with 5 �M or 1 mMmelatonin for 15 min at 37 ◦C in 4 mM MgCl2 and 100 mM Tris–HClbuffer, pH 8.5. Then, mixtures were further incubated with 600 �MFeCl3, 1 mM sodium ascorbate for 30 min at 37 ◦C with constantagitation. Afterwards, UDPGT activity was assayed incubating thesesamples with 0.5 mM p-nitrophenol and 2 mM UDPGA. Blanks wereassayed in the absence of UDPGA. Reactions were stopped by theaddition of TCA to 5% (w/v) final concentration; samples werethen centrifuged at 10,000 × g for 10 min in a Suprafuge 22 Her-aeus centrifuge and supernatants were collected. NaOH was addedto the supernatants to final 0.5 M and absorbance at 410 nm was
measured in a UV3 Unicam UV–VIS spectrophotometer. Reactionrates were determined in conditions where product formationwas linearly dependent to time and protein concentration. Tocalculate UDPGT activity, a standard curve of p-nitrophenol wasgenerated.
2 gical Interactions 185 (2010) 208–214
2
Omnaw(ir5aU
2
uM1pw6BtbwHt0abogpc
2
6bf2c
2
dbuc
3
3l
pm1tb
Fig. 1. Microsomal lipoperoxidation promoted by Fe3+/ascorbate: effect of mela-tonin. Microsomes were pre-incubated for 15 min at 25 ◦C with increasing melatoninconcentrations (up to 1 mM) prior to incubation with the Fe3+/ascorbate for 20 min
◦
Previously, we have reported that rat liver UDP-glucuronyltransferase (UDPGT) activity is increased byFe3+/ascorbate through an oxidative mechanism [27]. UDPGT
Fig. 2. Loss of microsomal thiol content elicited by Fe3+/ascorbate: effect of mela-tonin. Microsomes were pre-incubated for 15 min at 25 ◦C with either 0.1 or 1 mMmelatonin prior to incubation with the Fe3+/ascorbate for 30 min at 37 ◦C with con-stant agitation (grey bars). Another set of microsomes were incubated with the
10 M.E. Letelier et al. / Chemico-Biolo
.7. CYP450 monooxygenase spectrum
CYP450 monooxygenase spectrum was obtained according tomura and Sato [31]. This method uses the ability of the carbononoxide to coordinate with the heme group of the monooxyge-
ase; the resulting conjugate has a peak of maximum absorbancet 450 nm (ε450 = 91 mM−1 cm−1). Microsomes (1 mg protein/mL)ere pre-incubated with increasing concentrations of melatonin
up to 5 �M) for different lengths of time (up to 120 min) at 37 ◦Cn 4 mM MgCl2 and 50 mM phosphate buffer, pH 7.4. Afterwards,eaction mixtures (blank and sample) were supplemented withmM sodium dithionite and the 400–500 nm spectrum was gener-ted following addition of carbon monoxide to the sample in a UV3nicam UV–VIS spectrophotometer.
.8. p-Nitroanisole O-demethylation
The O-demethylating activity of CYP450 system was determinedsing p-nitroanisole as substrate, according to Letelier et al. [28].icrosomes (2 mg protein/mL) were pre-incubated with 5 �M ormM melatonin for 15 min at 37 ◦C in 4 mM MgCl2 and 50 mMhosphate buffer, pH 7.4. Afterwards, O-demethylating activityas assayed incubating these samples with 0.5 mM p-nitroanisole,mM G-6-P, 0.6 mM NADP and 5 units/mL G-6-P dehydrogenase.lanks contained all reagents but G-6-P dehydrogenase. All mix-ures were incubated for 20 min at 37 ◦C. Reactions were stoppedy the addition of TCA to 5% (w/v) final concentration; samplesere then centrifuged at 10,000 × g for 10 min in a Suprafuge 22eraeus centrifuge and supernatants were collected. To develop
he colorimetric reaction, NaOH was added to supernatants to final.5 M. Absorbance of samples was then determined at 410 nm inUV3 Unicam UV–VIS spectrophotometer, using their respective
lanks as reference. To calculate the O-demethylating activityf the CYP450 system, a standard curve of p-nitrophenol wasenerated. Reaction rates were determined in conditions whereroduct formation was linearly dependent to time and proteinoncentration.
.9. Spectral properties of melatonin
Melatonin (5 �M) was incubated in the presence or absence of00 �M FeCl3 and 1 mM sodium ascorbate in 50 mM phosphateuffer, pH 7.4, for 10 min at 25 ◦C. In addition, some samples wereurther incubated with 5 �M l-histidine for 10 min at 25 ◦C. Then,00–400 nm absorbance spectra were generated using a UV3 Uni-am UV–VIS spectrophotometer.
.10. Statistical analysis
Data presented correspond to the mean of at least four indepen-ent experiments ± SEM. Statistical significance (ANOVA, followedy post hoc Bonferroni) and regression analyses were performedsing GraphPad Prism 5.0. Differences were considered as signifi-ant when p < 0.05.
. Results
.1. Effect of melatonin on the oxidative damage to microsomalipids and protein-thiols promoted by Fe3+/ascorbate
As shown in Fig. 1, melatonin inhibited the microsomal lipid
eroxidation promoted by Fe3+/ascorbate in a linear-dependentanner, with a half-maximal effect at ∼0.5 mM. Furthermore,mM melatonin completely prevented this phenomenon; in con-rast, 1 mM l-cysteine or GSH prevented lipid peroxidation elicitedy Fe3+/ascorbate in 56 ± 2.3% and 89 ± 3.0%, respectively. On the
at 37 C with constant agitation. TBARS were then assayed as described in Section 2.Data correspond to the mean of at least 4 independent experiments ± SD. The IC50
value was obtained by interpolation of the linear regression of the data.
other hand, melatonin prevented the loss (50%) of microsomal thiolcontent elicited by Fe3+/ascorbate, in a concentration-dependentmanner (Fig. 2). Remarkably, 1 mM melatonin partially reversedthe loss of microsomal thiol content promoted by Fe3+/ascorbate.
3.2. Effect of melatonin on the oxidative activation ofUDP-glucuronyltransferase promoted by Fe3+/ascorbate
Fe3+/ascorbate system as above prior to incubation with either 0.1 or 1 mM mela-tonin as above (black bars). Thiol content of preparations was then titrated withDTNB as detailed in Section 2. Data represent the % of residual thiol content con-sidering 100% (hatched bar) as the value obtained in the absence of Fe3+/ascorbate.Data correspond to the mean of at least 4 independent experiments ± SD. *p < 0.05compared to Control (hatched bar). ns: values not significantly different (p < 0.05).
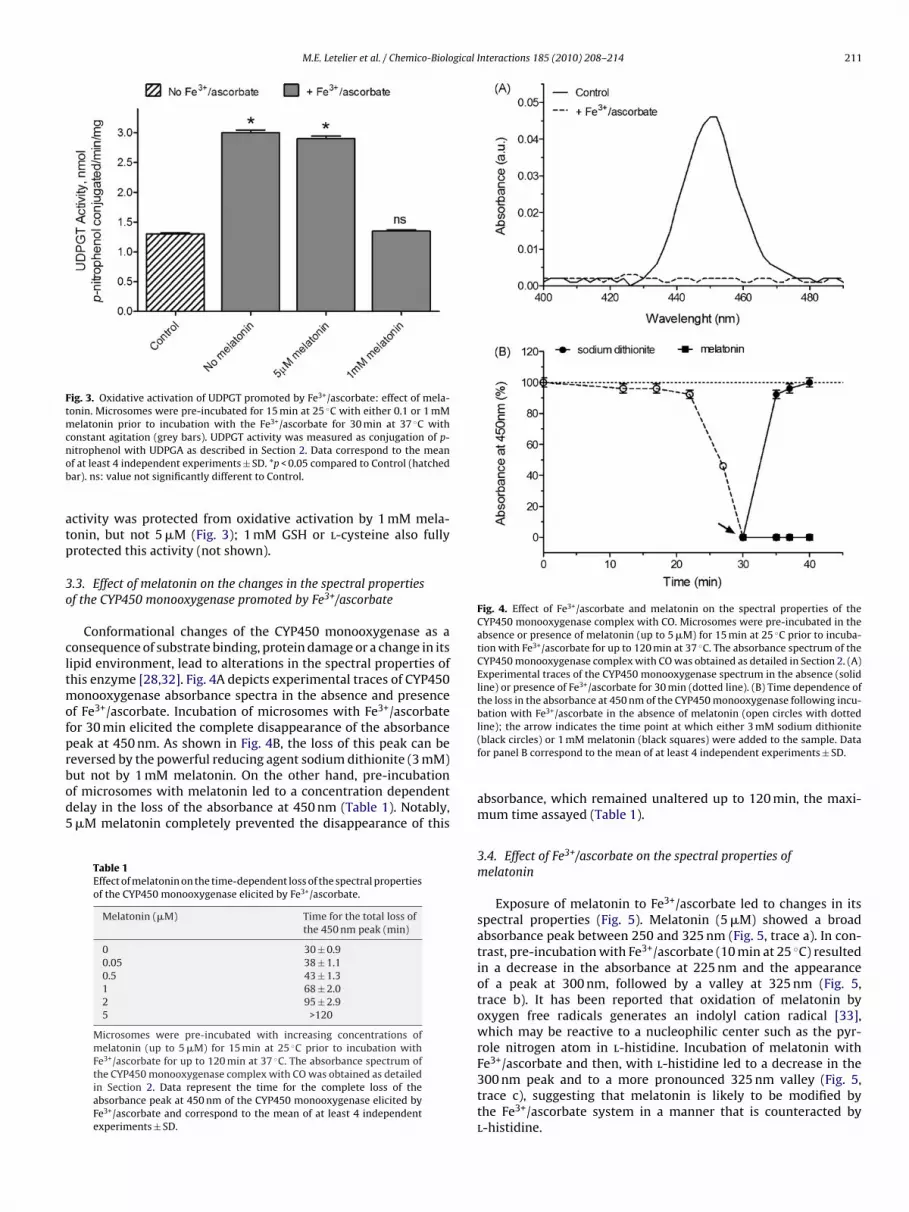
M.E. Letelier et al. / Chemico-Biological Interactions 185 (2010) 208–214 211
Fig. 3. Oxidative activation of UDPGT promoted by Fe3+/ascorbate: effect of mela-tonin. Microsomes were pre-incubated for 15 min at 25 ◦C with either 0.1 or 1 mMmelatonin prior to incubation with the Fe3+/ascorbate for 30 min at 37 ◦C withconstant agitation (grey bars). UDPGT activity was measured as conjugation of p-nob
atp
3o
cltmofprbod5
Fig. 4. Effect of Fe3+/ascorbate and melatonin on the spectral properties of theCYP450 monooxygenase complex with CO. Microsomes were pre-incubated in theabsence or presence of melatonin (up to 5 �M) for 15 min at 25 ◦C prior to incuba-tion with Fe3+/ascorbate for up to 120 min at 37 ◦C. The absorbance spectrum of theCYP450 monooxygenase complex with CO was obtained as detailed in Section 2. (A)Experimental traces of the CYP450 monooxygenase spectrum in the absence (solidline) or presence of Fe3+/ascorbate for 30 min (dotted line). (B) Time dependence ofthe loss in the absorbance at 450 nm of the CYP450 monooxygenase following incu-bation with Fe3+/ascorbate in the absence of melatonin (open circles with dotted
itrophenol with UDPGA as described in Section 2. Data correspond to the meanf at least 4 independent experiments ± SD. *p < 0.05 compared to Control (hatchedar). ns: value not significantly different to Control.
ctivity was protected from oxidative activation by 1 mM mela-onin, but not 5 �M (Fig. 3); 1 mM GSH or l-cysteine also fullyrotected this activity (not shown).
.3. Effect of melatonin on the changes in the spectral propertiesf the CYP450 monooxygenase promoted by Fe3+/ascorbate
Conformational changes of the CYP450 monooxygenase as aonsequence of substrate binding, protein damage or a change in itsipid environment, lead to alterations in the spectral properties ofhis enzyme [28,32]. Fig. 4A depicts experimental traces of CYP450
onooxygenase absorbance spectra in the absence and presencef Fe3+/ascorbate. Incubation of microsomes with Fe3+/ascorbateor 30 min elicited the complete disappearance of the absorbanceeak at 450 nm. As shown in Fig. 4B, the loss of this peak can be
eversed by the powerful reducing agent sodium dithionite (3 mM)ut not by 1 mM melatonin. On the other hand, pre-incubationf microsomes with melatonin led to a concentration dependentelay in the loss of the absorbance at 450 nm (Table 1). Notably,�M melatonin completely prevented the disappearance of thisTable 1Effect of melatonin on the time-dependent loss of the spectral propertiesof the CYP450 monooxygenase elicited by Fe3+/ascorbate.
Melatonin (�M) Time for the total loss ofthe 450 nm peak (min)
0 30 ± 0.90.05 38 ± 1.10.5 43 ± 1.31 68 ± 2.02 95 ± 2.95 >120
Microsomes were pre-incubated with increasing concentrations ofmelatonin (up to 5 �M) for 15 min at 25 ◦C prior to incubation withFe3+/ascorbate for up to 120 min at 37 ◦C. The absorbance spectrum ofthe CYP450 monooxygenase complex with CO was obtained as detailedin Section 2. Data represent the time for the complete loss of theabsorbance peak at 450 nm of the CYP450 monooxygenase elicited byFe3+/ascorbate and correspond to the mean of at least 4 independentexperiments ± SD.
line); the arrow indicates the time point at which either 3 mM sodium dithionite(black circles) or 1 mM melatonin (black squares) were added to the sample. Datafor panel B correspond to the mean of at least 4 independent experiments ± SD.
absorbance, which remained unaltered up to 120 min, the maxi-mum time assayed (Table 1).
3.4. Effect of Fe3+/ascorbate on the spectral properties ofmelatonin
Exposure of melatonin to Fe3+/ascorbate led to changes in itsspectral properties (Fig. 5). Melatonin (5 �M) showed a broadabsorbance peak between 250 and 325 nm (Fig. 5, trace a). In con-trast, pre-incubation with Fe3+/ascorbate (10 min at 25 ◦C) resultedin a decrease in the absorbance at 225 nm and the appearanceof a peak at 300 nm, followed by a valley at 325 nm (Fig. 5,trace b). It has been reported that oxidation of melatonin byoxygen free radicals generates an indolyl cation radical [33],which may be reactive to a nucleophilic center such as the pyr-role nitrogen atom in l-histidine. Incubation of melatonin withFe3+/ascorbate and then, with l-histidine led to a decrease in the
300 nm peak and to a more pronounced 325 nm valley (Fig. 5,trace c), suggesting that melatonin is likely to be modified bythe Fe3+/ascorbate system in a manner that is counteracted byl-histidine.
212 M.E. Letelier et al. / Chemico-Biological Interactions 185 (2010) 208–214
F 3+
t(F
3O
eoicwit
Ftfemvp(Dc
Fig. 7. Effect of DEPC and Fe3+/ascorbate-modified melatonin on the inhibitionof the O-demethylating activity of the CYP450 system elicited by Fe3+/ascorbate.Microsomes were pre-incubated with or without 2 �M DEPC for 5 min at 25 ◦C.Samples were then incubated with melatonin (15 min at 25 ◦C) prior to assayingO-demethylation of p-nitroanisole in the presence of Fe3+/ascorbate, according toSection 2 (grey bars). In parallel experiments, samples were incubated with mela-tonin previously treated with Fe3+/ascorbate (15 min at 25 ◦C) prior to determining
ig. 5. Effect of Fe /ascorbate on the spectral properties of melatonin. Representa-ive traces of the absorbance spectra of melatonin (5 �M), incubated in the absencea, continuous line) or presence of Fe3+/ascorbate (b, dotted line) or a mixture ofe3+/ascorbate and 5 �M l-histidine (c, segmented line) for 10 min at 25 ◦C.
.5. Effect of melatonin on the inhibition of p-nitroanisole-demethylation induced by Fe3+/ascorbate
Incubation of microsomes with Fe3+/ascorbate inhibited thisnzymatic reaction in approximately 46% (Fig. 6). Pre-incubationf microsomes with 5 �M melatonin completely prevented thisnhibition; the same effect was elicited by either 1 mM GSH or l-
ysteine (not shown). In contrast, pre-incubation of microsomesith 1 mM melatonin, promoted an approximately 75% decreasen the O-demethylating activity (Fig. 6). Melatonin, previouslyreated with Fe3+/ascorbate, prevented the inhibition of the p-
ig. 6. Effect of Fe3+/ascorbate and melatonin on the O-demethylating activity ofhe CYP450 system. Microsomes were pre-incubated with 5 �M or 1 mM melatoninor 15 min at 25 ◦C prior to assaying O-demethylation of p-nitroanisole in the pres-nce of Fe3+/ascorbate (grey bars), as detailed in Section 2. In parallel experiments,icrosomes were treated, for 15 min at 25 ◦C, with melatonin (5 �M or 1 mM) pre-
iously incubated with Fe3+/ascorbate, prior to determining O-demethylation of-nitroanisole without further addition of Fe3+ or ascorbate (black bars). Controlhatched bar) samples were incubated in the absence of melatonin or Fe3+/ascorbate.ata correspond to the mean of at least 4 independent experiments ± SD. *p < 0.05ompared to Control. ns: values not significantly different (p < 0.05).
O-demethylation of p-nitroanisole (black bars). Hatched bars indicate samples incu-
bated in the absence of Fe3+/ascorbate. Control: enzymatic activity in the absenceof Fe3+/ascorbate, DEPC or melatonin. Data correspond to the mean of at least 4independent experiments ± SD. *p < 0.05 compared to Control. ns: values not signif-icantly different (p < 0.05).nitroanisole O-demethylation elicited by this oxidative system tothe same extent as untreated melatonin (Fig. 6). Furthermore, mela-tonin previously treated with Fe3+/ascorbate in the presence ofequimolar amounts of l-histidine failed to elicit this protectiveeffect.
3.6. Comparison between the effect of Fe3+/ascorbate-modifiedmelatonin and DEPC on the inhibition of p-nitroanisoleO-demethylation induced by Fe3+/ascorbate
Diethylpyrocarbonate (DEPC) is a classical histidine modify-ing agent. Incubation of microsomes with 2 �M DEPC (in theabsence of melatonin or Fe3+/ascorbate) led to a ∼15% decreasein p-nitroanisole O-demethylating activity (Fig. 7); a similar inhibi-tion was observed with Fe3+/ascorbate-modified melatonin (5 �M).DEPC (2 �M) also decreased the absorbance at 450 nm of theCYP450 monooxygenase in approximately 50% (not shown). Onthe other hand, pre-incubation of microsomes with DEPC (inthe absence of melatonin) partially prevented the loss of p-nitroanisole O-demethylating activity elicited by Fe3+/ascorbate(Fig. 7). Following treatment of microsomes with 2 �M DEPC,Fe3+/ascorbate-modified melatonin was unable to further protectthis activity.
4. Discussion
We focused the present work on the antioxidant properties of
melatonin. We proposed that this function could have a special rele-vance in the hepatic endoplasmic reticulum, which is a sub-cellularorganelle involved in melatonin biotransformation and is also oneof the main cellular sources of reactive oxygen species (ROS). To thisend, we studied the protective effects of melatonin on the oxidative
gical
di
rltwmaToe
mmfaigtmTr
RiaFsortwmpRutioosFfi
abskmwdCa[aTcramCwwim
[
[
[
[
M.E. Letelier et al. / Chemico-Biolo
amage promoted by Fe3+/ascorbate on the biomolecules occurringn rat liver microsomes [27,29].
Our results show that mM concentrations of melatonin wereequired to completely prevent oxidative damage to microsomalipids (Fig. 1), microsomal thiol groups (Fig. 2), and oxidative activa-ion of UDPGT (Fig. 3). Noteworthy, melatonin antioxidant activityas more effective than that of l-cysteine and GSH to protecticrosomal lipids, confirming data previously described with other
ntioxidants, including vitamin E and mannitol [12–14,16,33].hese data are consistent with the idea that high concentrationsf melatonin display its widely described oxygen free radical scav-nging properties [12–14,16,33].
Although 5 �M melatonin failed to prevent oxidative damage toicrosomal lipids and thiol groups, it completely protected CYP450onooxygenase and CYP450 system O-demethylating activity
rom the oxidative damage elicited by Fe3+/ascorbate (Figs. 4 and 6nd Table 1). Higher concentrations of melatonin (1 mM), however,nhibited the O-demethylating activity of the CYP450 system to areater extent than that of Fe3+/ascorbate alone (Fig. 6). In addi-ion to the hydroxylation in position 6, it has been reported that
elatonin can be O-demethylated by the CYP450 system [18,19].herefore, the observed inhibition by 1 mM melatonin may be aeflection of competitive inhibition.
The antioxidant mechanism by which melatonin scavengesOS involves the non-enzymatic formation of a nitrogen-centered
ndolyl cation radical [34]. Substantial changes in the melatoninbsorbance spectrum following incubation of 5 �M melatonin withe3+/ascorbate (Fig. 5) suggest the modification of the hormonetructure. If this modification is oxidative in nature, then formationf a melatonin cation radical may occur; this type of electrophilicadicals is reactive to nucleophilic centers in molecules, such ashe pyrrole nitrogen atom occurring in l-histidine. In agreementith this possibility, addition of this amino acid to Fe3+/ascorbate-odified melatonin counteracted the changes in its spectral
roperties (Fig. 5). This evidence suggests that, upon reaction withOS (generated by the Fe3+/ascorbate system), melatonin is likely tondergo non-enzymatic oxidation yielding a cation radical reactiveo l-histidine. This postulate is supported by l-histidine prevent-ng the protective activity of Fe3+/ascorbate-modified melatoninn the CYP450 system O-demethylating activity (Fig. 6). Formationf a melatonin cation radical may also explain the partial rever-ion of the oxidative loss of microsomal thiol content elicited bye3+/ascorbate (Fig. 2), possibly through homolytic rupture of disul-de bonds, followed by stabilization by proton ions.
A covalent bond between a non-axial histidine side chain (H117)nd the heme 2-vinyl group of hemoglobin has been shown toe important for hemoglobin structure [35]. This bond seems totabilize the ferric protein against denaturation and to affect theinetics of cyanide association [35–37]. Because cytochrome P450onooxygenase is also a heme protein, it may form a similar bondith a histidine residue in an equivalent position; this type of histi-ine ligation has been recently reported to contribute to a bacterialYP450 monooxygenase crystal structure [38]. Noteworthy, thective site of the CYP450 monooxygenase is close to the heme group39,40], highlighting the possibility that this type of histidine lig-tion may be critical for substrate binding in the monooxygenase.hus, it may be possible that the putative melatonin cation radi-al generated under oxidative conditions may bind to this histidineesidue, preventing oxidative damage to the active site. Our datare in agreement with these postulates; in fact, Fe3+/ascorbate-odified melatonin protected the O-demethylating activity of the
YP450 system from oxidative damage (Fig. 7). This protectionas completely abolished when equimolar amounts of l-histidineere added to Fe3+/ascorbate-modified melatonin, possibly reflect-
ng the competition between the histidine residue in the CYP450onooxygenase and free histidine (Fig. 7).
[
[
Interactions 185 (2010) 208–214 213
In order to strengthen our hypothesis, we used the classical his-tidine residue modifying agent diethylpyrocarbonate (DEPC). Wefound that 2 �M DEPC decreased the 450 nm absorbance of theCYP450 monooxygenase in ∼50% and inhibited the CYP450 sys-tem O-demethylating activity in ∼15% (Fig. 7). This further supportsthe idea that a histidine residue in CYP450 monooxygenase maybe close to its substrate binding site. Moreover, DEPC itself pro-tected the O-demethylating activity of the CYP450 system from theoxidative damage elicited by Fe3+/ascorbate, mimicking the effectof Fe3+/ascorbate-modified melatonin (Figs. 6 and 7). The fact thatFe3+/ascorbate-modified melatonin was unable to protect the activ-ity of the CYP450 system further than the protection provided byDEPC, strongly suggests that both agents are likely targeting thesame site. If this postulate is correct, the mechanism underlyingthe antioxidant activity of �M concentrations of melatonin wouldinvolve: (1) the oxidation of this hormone, leading to the generationof a melatonin cation radical; and (2) direct binding of this radi-cal to nucleophilic centers in proteins, such as those occurring inhistidine residues. This mechanism would be more efficient to pro-tect specific proteins from oxidative damage than membrane lipids,as long as they bear nucleophilic centers critical to their structureand/or function, which is the case of the CYP450 monooxygenase.This mechanism does not exclude the more classical oxygen freeradical scavenging activity of melatonin, which was observed athigher concentrations. More studies are currently being conductedto provide structural insights to the novel antioxidant mechanismof melatonin proposed in this work.
Conflict of interest
The authors declare that there are no conflicts of interest.
References
[1] K. Ackermann, J.H. Stehle, Melatonin synthesis in the human pineal gland:advantages, implications, and difficulties, Chronobiol. Int. 23 (1–2) (2006)369–379.
[2] S. Ganguly, S.L. Coon, D.C. Klein, Control of melatonin synthesis in the mam-malian pineal gland: the critical role of serotonin acetylation, Cell Tissue Res.309 (1) (2002) 127–137.
[3] V. Simonneaux, C. Ribelayga, Generation of the melatonin endocrine messagein mammals: a review of the complex regulation of melatonin synthesis bynorepinephrine, peptides, and other pineal transmitters, Pharmacol. Rev. 55(2) (2003) 325–395.
[4] P. Delagrange, J. Atkinson, J.A. Boutin, L. Casteilla, D. Lesieur, R. Misslin, S. Pel-lissier, L. Penicaud, P. Renard, Therapeutic perspectives for melatonin agonistsand antagonists, J. Neuroendocrinol. 15 (4) (2003) 442–448.
[5] S.R. Pandi-Perumal, N. Zisapel, V. Srinivasan, D.P. Cardinali, Melatonin and sleepin aging population, Exp. Gerontol. 40 (12) (2005) 911–925.
[6] J. Wagner, M.L. Wagner, W.A. Hening, Beyond benzodiazepines: alternativepharmacologic agents for the treatment of insomnia, Ann. Pharmacother. 32(6) (1998) 680–691.
[7] D. Healy, J.M. Waterhouse, The circadian system and the therapeutics of theaffective disorders, Pharmacol. Ther. 65 (2) (1995) 241–263.
[8] J.Z. Wang, Z.F. Wang, Role of melatonin in Alzheimer-like neurodegeneration,Acta Pharmacol. Sin. 27 (1) (2006) 41–49.
[9] B. Bruguerolle, N. Simon, Biologic rhythms and Parkinson’s disease: achronopharmacologic approach to considering fluctuations in function, Clin.Neuropharmacol. 25 (4) (2002) 194–201.
10] J. Borjigin, X. Li, S.H. Snyder, The pineal gland and melatonin: molecular andpharmacologic regulation, Annu. Rev. Pharmacol. Toxicol. 39 (1999) 53–65.
11] D.X. Tan, L.C. Manchester, M.P. Terron, L.J. Flores, R.J. Reiter, One molecule, manyderivatives: a never-ending interaction of melatonin with reactive oxygen andnitrogen species? J. Pineal Res. 42 (1) (2007) 28–42.
12] R. Hardeland, Antioxidative protection by melatonin: multiplicity of mecha-nisms from radical detoxification to radical avoidance, Endocrine 27 (2) (2005)119–130.
13] R.J. Reiter, D.X. Tan, M.D. Maldonado, Melatonin as an antioxidant: physiologyversus pharmacology, J. Pineal Res. 39 (2) (2005) 215–216.
14] R.J. Reiter, D.X. Tan, J.C. Mayo, R.M. Sainz, J. Leon, Z. Czarnocki, Melatonin as anantioxidant: biochemical mechanisms and pathophysiological implications inhumans, Acta Biochim. Pol. 50 (4) (2003) 1129–1146.
15] F. Zhao, Z.Q. Liu, D. Wu, Antioxidative effect of melatonin on DNA and erythro-cytes against free-radical-induced oxidation, Chem. Phys. Lipids 151 (2) (2008)77–84.

2 gical
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
14 M.E. Letelier et al. / Chemico-Biolo
16] C. Pieri, M. Marra, F. Moroni, R. Recchioni, F. Marcheselli, Melatonin: a per-oxyl radical scavenger more effective than vitamin E, Life Sci. 55 (15) (1994)271–276.
17] C. Rodriguez, J.C. Mayo, R.M. Sainz, I. Antolin, F. Herrera, V. Martin, R.J. Reiter,Regulation of antioxidant enzymes: a significant role for melatonin, J. PinealRes. 36 (1) (2004) 1–9.
18] I. Semak, E. Korik, M. Antonova, J. Wortsman, A. Slominski, Metabolism of mela-tonin by cytochrome P450s in rat liver mitochondria and microsomes, J. PinealRes. 45 (4) (2008) 515–523.
19] X. Ma, J.R. Idle, K.W. Krausz, F.J. Gonzalez, Metabolism of melatonin by humancytochromes P450, Drug Metab. Dispos. 33 (4) (2005) 489–494.
20] S.S. Singer, J.E. Hagedorn, D.M. Smith, J.L. Williams, The enzymatic basis forthe rat liver 6-hydroxymelatonin sulfotransferase activity, J. Pineal Res. 18 (1)(1995) 49–55.
21] S.C. Bondy, S. Naderi, Contribution of hepatic cytochrome P450 systems tothe generation of reactive oxygen species, Biochem. Pharmacol. 48 (1) (1994)155–159.
22] S. Puntarulo, A.I. Cederbaum, Production of reactive oxygen species by micro-somes enriched in specific human cytochrome P450 enzymes, Free Radic. Biol.Med. 24 (7–8) (1998) 1324–1330.
23] R.C. Zangar, D.R. Davydov, S. Verma, Mechanisms that regulate production ofreactive oxygen species by cytochrome P450, Toxicol. Appl. Pharmacol. 199 (3)(2004) 316–331.
24] M.E. Letelier, F. Lagos, M. Faundez, D. Miranda, M. Montoya, P. Aracena-Parks,V. Gonzalez-Lira, Copper modifies liver microsomal UDP-glucuronyltransferaseactivity through different and opposite mechanisms, Chem. Biol. Interact. 167(1) (2007) 1–11.
25] M.E. Letelier, A.M. Lepe, M. Faundez, J. Salazar, R. Marin, P. Aracena, H.Speisky, Possible mechanisms underlying copper-induced damage in biolog-ical membranes leading to cellular toxicity, Chem. Biol. Interact. 151 (2) (2005)71–82.
26] M.E. Letelier, M. Martinez, V. Gonzalez-Lira, M. Faundez, P. Aracena-Parks, Inhi-bition of cytosolic glutathione S-transferase activity from rat liver by copper,
Chem. Biol. Interact. 164 (1–2) (2006) 39–48.27] M.E. Letelier, A. Pimentel, P. Pino, A.M. Lepe, M. Faundez, P. Aracena, H. Speisky,Microsomal UDP-glucuronyltransferase in rat liver: oxidative activation, BasicClin. Pharmacol. Toxicol. 96 (6) (2005) 480–486.
28] M.E. Letelier, M. Faúndez, J. Jara-Sandoval, A. Molina-Berríos, J. Cortés-Troncoso, P. Aracena-Parks, R. Marín-Catalán, Mechanisms underlying the
[
Interactions 185 (2010) 208–214
inhibition of the cytochrome P450 system by copper ions, J. Appl. Toxicol. 29(8) (2009) 675–702.
29] M.E. Letelier, A. Molina-Berrios, J. Cortes-Troncoso, J. Jara-Sandoval, A. Müller,P. Aracena-Parks, Comparative effects of superoxide anion and hydrogen per-oxide on microsomal and cytosolic glutathione S-transferase activities of ratliver, Biol. Trace Elem. Res. 134 (2) (2010) 203–211.
30] O.H. Lowry, N.J. Rosebrough, A.L. Farr, R.J. Randall, Protein measurement withthe Folin phenol reagent, J. Biol. Chem. 193 (1) (1951) 265–275.
31] T. Omura, R. Sato, The carbon monoxide-binding pigment of liver microsomes.I. Evidence for its hemoprotein nature, J. Biol. Chem. 239 (1964) 2370–2378.
32] T. Omura, R. Sato, The carbon monoxide-binding pigment of liver micro-somes. II. Solubilization, purification, and properties, J. Biol. Chem. 239 (1964)2379–2385.
33] B. Poeggeler, S. Thuermann, A. Dose, M. Schoenke, S. Burkhardt, R. Hardeland,Melatonin’s unique radical scavenging properties—roles of its functional sub-stituents as revealed by a comparison with its structural analogs, J. Pineal Res.33 (1) (2002) 20–30.
34] A. Crespo, A.G. Turjanski, D.A. Estrin, Electronic spectra of indolyl radicals: atime-dependent DFT study, Chem. Phys. Lett. 365 (1–2) (2002) 15–21.
35] J.A. Hoy, B.J. Smagghe, P. Halder, M.S. Hargrove, Covalent heme attachment inSynechocystis hemoglobin is required to prevent ferrous heme dissociation,Protein Sci. 16 (2) (2007) 250–260.
36] L. Tesoriere, M. Allegra, D. D’Arpa, D. Butera, M.A. Livrea, Reaction of mela-tonin with hemoglobin-derived oxoferryl radicals and inhibition of thehydroperoxide-induced hemoglobin denaturation in red blood cells, J. PinealRes. 31 (2) (2001) 114–119.
37] L. Tesoriere, G. Avellone, L. Ceraulo, D. D’Arpa, M. Allegra, M.A. Livrea, Oxidationof melatonin by oxoferryl hemoglobin: a mechanistic study, Free Radic. Res. 35(6) (2001) 633–642.
38] L.-H. Xu, S. Fushinobu, H. Ikeda, T. Wakagi, H. Shoun, Crystal structures ofcytochrome P450 105P1 from Streptomyces avermitilis: conformational flexi-bility and histidine ligation state, J. Bacteriol. 191 (4) (2009) 1211–1219.
39] T.L. Chen, T.H. Ueng, S.H. Chen, P.H. Lee, S.Z. Fan, C.C. Liu, Human cytochrome
P450 mono-oxygenase system is suppressed by propofol, Br. J. Anaesth. 74 (5)(1995) 558–562.40] D.H. Sherman, S. Li, L.V. Yermalitskaya, Y. Kim, J.A. Smith, M.R. Waterman, L.M.Podust, The structural basis for substrate anchoring, active site selectivity, andproduct formation by P450 PikC from Streptomyces venezuelae, J. Biol. Chem.281 (36) (2006) 26289–26297.