co-expression of a cellobiose phosphorylase and lactose permease enables intracellular cellobiose...
TRANSCRIPT

APPLIED MICROBIAL AND CELL PHYSIOLOGY
Co-expression of a cellobiose phosphorylase and lactosepermease enables intracellular cellobiose utilisationby Saccharomyces cerevisiae
Christa J. Sadie & Shaunita H. Rose & Riaan den Haan &
Willem H. van Zyl
Received: 17 November 2010 /Revised: 29 January 2011 /Accepted: 1 February 2011 /Published online: 20 February 2011# Springer-Verlag 2011
Abstract The cellobiose phosphorylase (cepA) gene fromClostridium stercorarium was cloned and successfullyexpressed under transcriptional control of the phosphoglyc-erate kinase gene (PGK1) promoter and terminator inSaccharomyces cerevisiae Y294. The recombinant CepAenzyme showed optimal activity at 60 °C and pH 5 anddisplayed a Km value of 92.85 mM and 1.69 mM oncellobiose and pNPG, respectively. A codon-optimisedsynthetic cepA gene was also expressed; however, it didnot enhance cellobiose utilisation. Transport of cellobiosewas subsequently facilitated through the heterologousexpression of the lac12 of Kluyveromyces lactis. Strainsco-producing the heterologous CepA and Lac12 were ableto grow on cellobiose as sole carbon source. This is the firstreport of successful intracellular utilisation of cellobiose byS. cerevisiae producing a cellobiose phosphorylase and ofcellobiose transport into S. cerevisiae via the K. lactis lac12encoded permease.
Keywords Cellobiose phosphorylase . Lactose permease .
Disaccharide transport . Cellobiose utilisation
Introduction
A worldwide decline in crude oil production has initiatedthe exploration of alternative energy sources such asbioethanol that is already extensively used as a partial
gasoline replacement (Sun and Cheng 2002). Optimisationof yeasts for efficient cellulose degradation could helpdecrease the high cost associated with the current enzymatichydrolysis and fermentation processes for producing bioetha-nol from lignocellulosic feedstocks (Lynd et al. 2002; Lynd etal. 2005). Developing Saccharomyces cerevisiae for cellulosedegradation requires the successful expression of cellulases,including the three key enzymes: exoglucanase (such ascellobiohydrolases), endoglucanase and β-glucosidase (vanRensburg et al. 1998). The employment of a single organismfor degradation of cellulose as well as the fermentation of theresulting hexoses (known as consolidated bioprocessing) canresult in a significant reduction in costs by simplifying theproduction process (Lynd et al. 1999).
In most cellulolytic organisms,β-glucosidases are essentialfor the efficient utilisation of cellobiose, the main product ofcellobiohydrolase activity and longer cello-oligosaccharides(Gonde et al. 1984; Lynd et al. 2002). β-Glucosidasesconvert cellobiose and cello-oligosaccharides to glucose thatcan be metabolised by the host organism. However,Clostridium species also produce cellobiose phosphorylasesthat hydrolyse cellobiose and simultaneously phosphorylateone of the glucose molecules with an inorganic phosphategroup yielding one glucose molecule and one glucose-1-phosphate molecule that are both further metabolisedthrough glycolysis (Tanaka et al. 1994). Since the oneglucose molecule is already phosphorylated prior to enteringthe glycolytic pathway, the expression of a cellobiosephosphorylase in yeast should theoretically be energeticallyadvantageous compared to the expression of a β-glucosidase(Fujita et al. 2002; Reichenbecher et al. 1997; Skory et al.1996). In organisms that make use of cellobiose phosphor-ylases, the rate of phosphorolysis greatly exceeds the rate ofhydrolysis, indicating the possible metabolic advantage ofthis reaction (Zhang and Lynd 2004).
C. J. Sadie : S. H. Rose :R. den Haan :W. H. van Zyl (*)Department of Microbiology, University of Stellenbosch,De Beer Street,Stellenbosch 7600, South Africae-mail: [email protected]
Appl Microbiol Biotechnol (2011) 90:1373–1380DOI 10.1007/s00253-011-3164-z

Cellobiose phosphorylase is an intracellular enzyme thatrequires the transport of the cellobiose molecule into thecell (Alexander 1961). S. cerevisiae is known to transportthe disaccharides maltose and sucrose across the cellmembrane, both via the maltose proton symport system(van der Rest et al. 1995; Stambuk et al. 2000), but isunable to import cellobiose. Although several yeast species(including the yeasts Clavispora lusitaniae and Pichiaguillermondii) have the ability to transport cellobiose acrossthe plasma membrane, the transport mechanism has not yetbeen resolved (Freer and Greene 1990; Freer 1991). Theyeast Kluyveromyces lactis produces an intracellularβ-glucosidase, implying that the yeast also has the abilityto transport cellobiose into the cell (Tingle and Halvorson1971; Tingle and Halvorson 1972). K. lactis produces alactose permease encoded by lac12 to facilitate uptake ofthe disaccharide lactose for intracellular utilisation. Heterolo-gous expression of the lac12 gene in S. cerevisiae enabled therecombinant strain to grow on lactose (Sreekrishna andDickson 1985). Here, we report for the first time the cloningand successful expression of a cellobiose phosphorylase gene(cepA) from Clostridium stercorarium in S. cerevisiae. Wealso report the use of the K. lactis lactose permease (Lac12)for cellobiose transport in S. cerevisiae facilitating theintracellular utilisation of cellobiose.
Materials and methods
Microbial strains and plasmids
The relevant genotypes and corresponding sources of themicrobial strains, as well as the plasmids used in this study aresummarised in Table 1. Genomic DNA of C. stercorariumwas obtained from the Institute for Microbiology, TechnicalUniversity Munich, Germany (Reichenbecher et al. 1997).
Media and culture conditions
Recombinant plasmids were amplified in Escherichia coliXL1-Blue and cultivated at 37 °C in Luria-Bertani liquidmedium (Sambrook et al. 1989). Ampicillin (100 μg mL−1)was added for selection of recombinant bacteria. S. cerevisiaetransformants were initially plated onto synthetic complete(SC−URA or SC−TRP) medium (3.4 gL-1 yeast nitrogen base(Difco) containing amino acid supplements, but lacking uraciland/or tryptophan, 5 gL−1 (NH4)2SO4, 20 gL−1 glucose).Liquid cultivation took place at 30 °C on an orbital shaker at200 rpm in double strength SC−URA medium containingeither 10.52 gL−1 glucose or 10 gL−1 cellobiose (molarequivalent of glucose) as carbohydrate source. The growthmedium was buffered with 20 mM sodium phosphate buffer(pH 6.5). All solid media contained 20 gL−1 agar.
DNA manipulation and plasmid construction
Standard protocols were followed for DNA manipulations(Sambrook et al. 1989). Restriction endonucleases, Klenowfragment and T4 DNA ligase were purchased from Rocheand used as recommended by the supplier. Restrictionendonuclease-digested DNA was recovered from agarosegels by the method of Tautz and Renz (1983).
The cellobiose phosphorylase encoding gene (cepA) wascloned from C. stercorarium genomic DNA and the lactosepermease encoding gene (lac12) was isolated from K. lactisgenomic DNA using PCR amplification (Table 2). Thesequence-specific primers were designed based on the cepAand lac12 sequences obtained from Genbank (accessionnumbers U56242 and XO6997). Polymerase chain reac-tions were carried out using the Perkin-Elmer GeneAmp®PCR System 2400 (The Perkin-Elmer Corporation), andTaq DNA polymerase (Roche Molecular Biochemicals)was applied according to the supplier’s specifications. Theresulting 2,436-bp cepA open reading frame was digestedwith BglII and XhoI and ligated into the corresponding sitesof plasmid pJC1 (Crous et al. 1995), resulting in yCEPA(Fig. 1). The nucleotide sequences of the cepA gene werecodon-optimised for expression in S. cerevisiae using aproprietary gene design software from Geneart (Regens-berg, Germany) based on the amino acid sequence of thegene (accession number U56242). Unwanted sequencerepeats, polyA sites, splice sites, potential secondary RNAstructures and unwanted restriction endonucleases siteswere avoided while optimal GC content and codon usagewere prioritised. The complete gene sequences including5′ EcoRI and 3′ XhoI restriction endonuclease sites weresent for de novo gene synthesis by Geneart. The syntheticgene (now called syncepA) was excised from the plasmidprovided by digesting with EcoRI and XhoI and cloned intothe corresponding sites of pJC1, resulting in ySynCEPA(Fig. 1). The yeast expression plasmid yBBH1 is similar tothe plasmid yENO1 that was previously described (DenHaan et al. 2007), but contains an additional 150-bp spacerregion upstream and 220 bp downstream of the ENO1promoter/terminator cassette. The cassette was designed toenable the consecutive cloning of additional cassettes intothe BamHI and BglII sites making use of the compatibilityof the restriction sites. The lac12 PCR product was digestedwith EcoRV and XhoI and cloned into the EcoRI (bluntended with Klenow treatment) and XhoI sites of yBBH1,generating yBBH1-Lac12. The ENO1P-lac12-ENO1T cassettewas retrieved as a BamHI and BglII fragment from yBBH1-Lac12 and cloned into the BamHI site of YIplac204 andyCEPA, generating yLac12 and yCEPA+lac(epi) (Fig. 1).Sequence verification of all constructs was done aftersequence determination by the dideoxy chain terminationmethod, using an ABI PRISM™ 3100 Genetic Analyzer.
1374 Appl Microbiol Biotechnol (2011) 90:1373–1380

Sequence analysis utilised mainly the PC-based DNAMAN(version 4.1) package from Lynnon BioSoft and the BLASTprogramme at the National Center for BiotechnologyInformation (www.ncbi.nih.gov/BLAST/).
Yeast transformation and growth on cellobiose
DNA transformation of S. cerevisiae was performed usingthe lithium acetate dimethyl sulfoxide method described byHill et al. (1991). Transformants were tested for thepresence of the relevant genes by means of PCR (notshown). A strain containing the pJC1 plasmid, S. cerevisiaeY294[pJC1], acted as reference strain. Growth of the
recombinant yeast was measured as dry cell weight(DCW) determined as described by Den Haan et al.(2007). All growth curves and dry weight estimations weredone in triplicate. Yeast strains were inoculated to aconcentration of 5×105 cells per mL in SC−URA mediumcontaining cellobiose as sole carbohydrate source. Growthwas monitored over a period of 114 h. Samples of thesupernatant were taken at regular intervals, and thecellobiose, glucose and ethanol content was determinedwith high performance liquid chromatography. Sampleswere separated on a Phenomenex Rezex column (RHM,monosaccharide H+8%, 300×7.8 mm) with guard column(security guard KJO-4282) using a Finnigan Surveyor LC
Table 1 Microbial strains and plasmids used in this study
Strain or plasmid Relevant genotype Reference
Strains:
E. coli XL1-Blue MRF’ endA1 supE44 thi-1 recA1 gyrA96 relA1 lac[F’.proAB lacq ZΔM15 Tn10 (tet)]
Stratagene
K. lactis GG399 Wild type NEBiolabs
S. cerevisiae Y294 α leu2-3,112 ura3-52 his3 trp1-289 ATCC 201160
S. cerevisiae Y294[pJC1] ura3 URA3 PGK1P-PGK1T This study
S. cerevisiae Y294[yCEPA] ura3 URA3 PGK1P-cepA-PGK1T This study
S. cerevisiae Y294[ySynCEPA] ura3 URA3 PGK1P-syncepA-PGK1T This study
S. cerevisiae Y294[yLac12] ura3 URA3 PGK1P-PGK1T This studytrp1 TPR1 ENO1P-lac12-ENO1T
S. cerevisiae Y294[yCEPA+lac(epi)] ura3 URA3 PGK1P-cepA-PGK1T; This studyENO1P-lac12-ENO1T
S. cerevisiae Y294[yCEPA+lac(int)] ura3 URA3 PGK1P-cepA-PGK1T; This studytrp1 TRP1 ENO1P-lac12-ENO1T
S. cerevisiae Y294[ySynCEPA+lac(int)] ura3 URA3 PGK1P-syncepA-PGK1T; This studytrp1 TRP1 ENO1P-lac12-ENO1T
Plasmids:
pJC1 bla URA3 PGK1P-PGK1T Crous et al. 1995
yCEPA bla URA3 PGK1P-cepA-PGK1T This study
ySynCEPA bla URA3 PGK1P-syncepA-PGK1T This study
yENO1 bla URA3 ENO1P-ENO1T Den Haan et al. 2007
yBBH1 bla URA3 ENO1P-ENO1T This study
YIplac 204 bla TRP1 ATCC 87591
yBBH1-Lac12 bla URA3 ENO1P-lac12-ENO1T This study
yCEPA-lac bla URA3 PGK1P-cepA-PGK1T; ENO1P-lac12-ENO1T This study
yLac12 bla TRP1 ENO1P-lac12-ENO1T This study
Primer name Sequence (5′→3′) Source
cepA
cepA-L GACTAGATCTATGAAGTTCGGTTATTTTGAC Genbank U56242cepA-R CAGTCTCGAGCAGCCCATTATAACAATTACT
Lac12
Lac12-L ACTCGAGGGTTAAACAGATTCTGC Genbank XO6997Lac12-R CGATATCAAATGGCAGATCATTCGA
Table 2 PCR primers used forgene isolation and identificationin this study
Restriction sites are underlined
Appl Microbiol Biotechnol (2011) 90:1373–1380 1375

pump (Thermo Scientific) and were detected with a FinniganSurveyor RI detector (Thermo Scientific). The mobile phaseusedwas 5mMH2SO4 at a flow rate of 0.7 mL/min at 65 °C.
Isolation of recombinant CepA
Whole yeast cells were prepared by centrifuging earlystationary phase cultures (OD600=0.6) at 2.795×g for10 min. The cell pellets were washed twice with ice-colddistilled water and resuspended in citric acid/phosphatebuffer (0.05 M, pH 5.0). Intracellular proteins were isolatedusing the glass bead method described by La Grange et al.(1996). β-Mercaptoethanol was excluded from all of thesolutions or buffers, and the supernatant containing theproteins was used without precipitation. The pellet (withglass beads) remaining after extraction of the intracellularproteins was subsequently treated with 1% Triton X-100,mixed for 30 s and centrifuged for 10 min at 11,269×g. Theresulting supernatant (containing the solubilised membranebound proteins) was kept for protein analysis. Proteinconcentrations were determined using the Bradford proteinassay method (Biolab) with bovine serum albumin as standard(Bradford 1976). Isolated proteins were stored at −20 °C.Cells were also lysed by treatment with 20 U of Zymolyase™
per gramme cells. Cell lysis was followed microscopicallyand the yeast membrane suspension was centrifuged at1,006×g for 15 min and resuspended in distilled water.
Enzyme assays
Activity of cellobiose phosphorylase was determined usingthe chromogenic substrate p-nitrophenyl-β-D-glucopyranoside
(pNPG, Sigma). Different cell fractions (depending on theexperiment) were appropriately diluted and incubated with5 mM pNPG in 50 mM citrate–phosphate buffer (pH 5)60 °C for 15 min. Equal volumes of sodium carbonate (1 M)were added to the reaction mixture to raise the pH andterminate the enzyme reaction. Release of the p-nitrophenolgroup was measured as absorbance at 405 nm. To determineoptimal conditions for the recombinant CepA the assayreaction was performed with a range of different citrate–phosphate buffers (pH 3.5–7) at a temperature of 60 °C orwith citrate–phosphate buffer pH 5 at a range of temperatures(30 to 70 °C). One unit of activity (U) was defined as theamount of enzyme catalysing the release of 1 μmol p-nitrophenol per minute.
Results
All S. cerevisiae Y294 transformants were subsequentlytested for cellobiose phosphorylase activity and their abilityto grow on solid SC−URA medium containing cellobiose assole carbon source (Fig. 1b). Only S. cerevisiae Y294[yCEPA+lac(int)], S. cerevisiae Y294[yCEPA+lac(epi)] andS. cerevisiae Y294[ySynCEPA+lac(int)] were able to growby utilising the cellobiose. This indicated that import ofcellobiose into the yeast cells producing CepA by the Lac12permease was a prerequisite for growth on this dimeric sugar.
Enzyme localisation, optima and kinetics
No activity could be detected in the growth medium ofstrains expressing the cepA or syncepA indicating that the
URA3
yCEPA, ySynCEPA
bla
PGK1P
ori
EcoRI
XhoI
PGK1T
2 micron
cepA/syncepABamHI
BglII
URA3
yCEPA-lac
bla
PGK1P
ori
PGK1T
2 micron
cepAENO1P
lac12ENO1T
URA3
yBBH1-lac12
bla
ENO1P
ori
XhoI
ENO1T
2 micron
lac12BamHI
BglII
EcoRI
TRP1
yLac12
bla
ENO1P
ori
XhoI
ENO1T
lac12BamHI
EcoRI
BglII
[yCEPA+lac (int)]
[yCEPA+lac(epi)]
[ysynCEPA+lac(int)]
[pJC1]
[yLac12]
[yCEPA]
[ySynCEPA]
a b c
d e
Fig. 1 a–d Plasmid maps ofvectors used in this studyincluding the orientation ofcloned genes and relevantrestriction enzyme positions.e The different strains cultivatedon cellobiose as sole carbohy-drate source revealed growthfor S. cerevisiae Y294[yCEPA+lac(int)], S. cerevisiae Y294[yCEPA+lac(epi)] andS. cerevisiae Y294[ySynCEPA+lac(int)] after 72 h ofincubation at 30 °C
1376 Appl Microbiol Biotechnol (2011) 90:1373–1380

enzyme was not secreted. Cell-associated activity could bemeasured with whole cells incubated with pNPG in thesestrains. When yeast cell walls were removed with Zymolyase,no hydrolytic activity could be detected in the periplasmicfraction. The intracellular and cell membrane protein fractionsdisplayed hydrolytic activity towards pNPG (data not shown)and were subsequently used for the characterization of therecombinant CepA enzyme. The control strain S. cerevisiaeY294[pJC1] did not show activity on pNPG in any of thefractions.
The optimum pH for the recombinant CepA protein wasestimated to be at pH 5 (Fig. 2a). Strains S. cerevisiae Y294[yCEPA] and Y294[ySynCEPA] produced similar levels ofactivity, 2.45±0.39 U/mg DCW, at optimal pH andtemperature conditions. The optimum temperature of thehydrolytic activity of the CepAwas estimated to be at 60 °C(Fig. 2b). At 30 °C, the optimum temperature for yeastgrowth, the enzyme exhibited only 14% of its optimalactivity at 60 °C, while only 53% of the maximum activitystill remained at 65 °C.
The recombinant enzyme’s affinity constant and maxi-mum velocity was estimated using Michaelis-Mentenkinetics. The enzyme activities on different cellobiose andpNPG concentrations were determined and used to draw aLineweaver–Burke plot (Fig. 2c). For pNPG, the Km of theenzyme was determined to be 1.69 mM and the Vmax was33.89 Umin−1 (Fig. 2d). For cellobiose, the Km was363.81 mM and the Vmax 3.41 Umin−1. These differences
in affinity for cellobiose and pNPG were also observed forother cellobiose phosphorylases (Reichenbecher et al. 1997;Yernool et al. 2000; Rajashekhara et al. 2002).
Growth of recombinant strains on cellobiose
All yeast strains were cultivated in double strength SC−URA
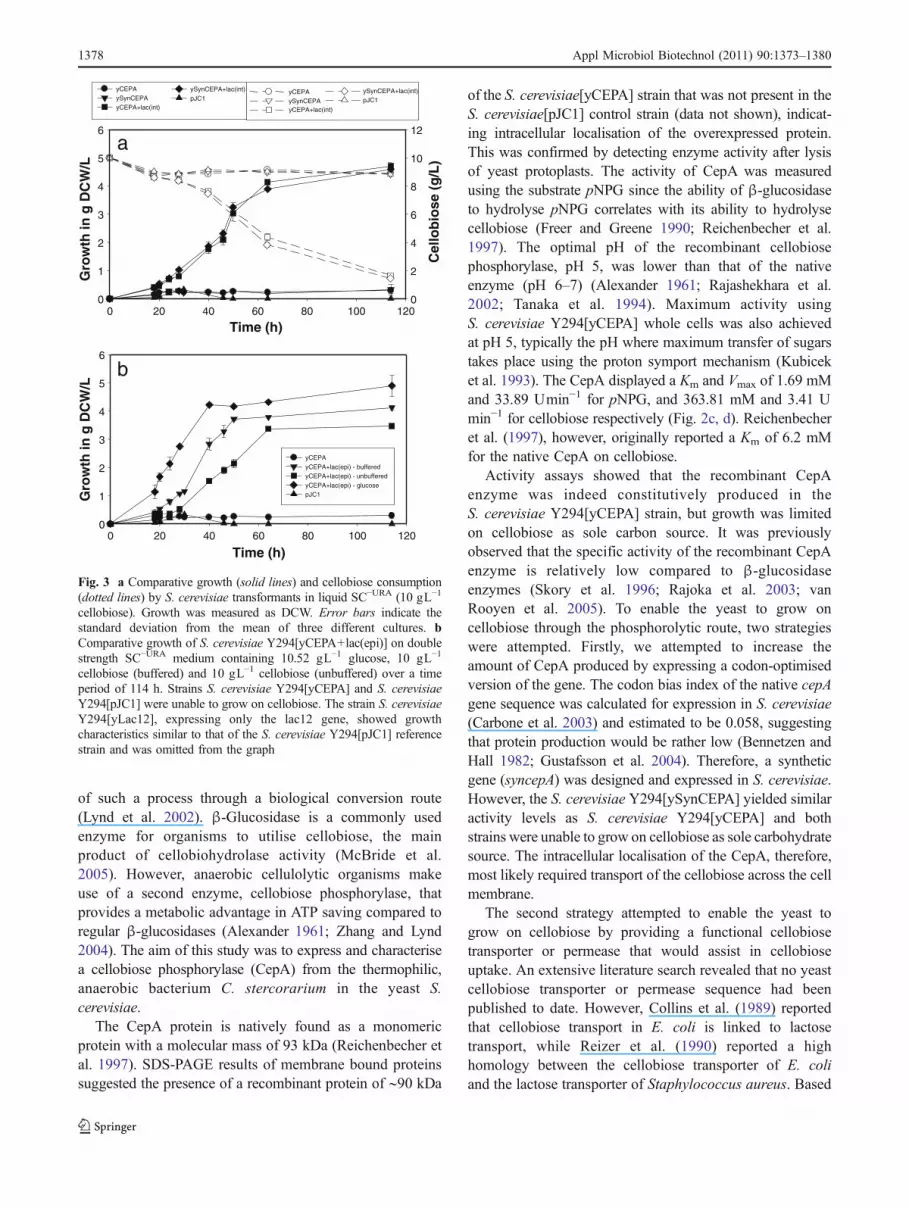
medium containing 10 gL−1 cellobiose and 20 mM sodiumphosphate buffer (pH 6.5). Growth and cellobiose con-sumption was monitored for a period of 114 h (Fig. 3a).Strains S. cerevisiae Y294[ySynCEPA+lac(int)] and S.cerevisiae Y294[yCEPA+lac(int)] were able to grow oncellobiose and utilise more than 80% of the cellobiosepresent in the medium (Fig. 3a). Strains S. cerevisiae Y294[pJC1], S. cerevisiae Y294[yLac12], S. cerevisiae Y294[ySynCEPA] and S. cerevisiae Y294[yCEPA] were unableto utilise the cellobiose. The addition of sodium phosphatebuffer to the medium enhanced growth of the S. cerevisiaeY294[yCEPA+lac(epi)] strain (Fig. 3b).
Discussion
Cellulose is a viable and potentially cost-effective substratefor the production of bioethanol as substitute for fossil fuels(Kosaric et al. 2001; Lynd et al. 2005). Using organismsthat are efficient in utilising this substrate and subsequentfermentation to ethanol is critical for the commercialisation
a
pH3 4 5 6 7 8
Rel
ativ
e ac
tivi
ty (
%)
0
20
40
60
80
100
120b
Temperature (oC)20 30 40 50 60 70 80
Rel
ativ
e ac
tivi
ty (
%)
0
20
40
60
80
100
120
cy = 106.56x + 0.2929
-5
0
5
10
15
20
25
-0.05 0.05 0.15 0.25
1/[Cellobiose]
1/V
o
dy = 0.05x + 0.0295
-0.02
-0.01
0
0.01
0.02
0.03
0.04
0.05
-0.8 -0.4 0.2
1/V
o
1/[pNPG]
-0.2-0.6
Fig. 2 The activity of S. cer-evisiae Y294[yCEPA] wholecells on pNPG at different a pHvalues and b temperatures.Lineweaver–Burke plots fordetermination of Km and Vmax
values of the recombinant CepAfor c cellobiose and d pNPG.Cell membrane fractions wereincubated with different con-centrations (5 to 50 mM) of eachsubstrate. The initial velocity ofthe enzyme at pH 5 and 60 °Cwith each substrate was calcu-lated and the reciprocal valuesof the substrate concentrationplotted against the inverseof the initial velocity of each
Appl Microbiol Biotechnol (2011) 90:1373–1380 1377
of such a process through a biological conversion route(Lynd et al. 2002). β-Glucosidase is a commonly usedenzyme for organisms to utilise cellobiose, the mainproduct of cellobiohydrolase activity (McBride et al.2005). However, anaerobic cellulolytic organisms makeuse of a second enzyme, cellobiose phosphorylase, thatprovides a metabolic advantage in ATP saving compared toregular β-glucosidases (Alexander 1961; Zhang and Lynd2004). The aim of this study was to express and characterisea cellobiose phosphorylase (CepA) from the thermophilic,anaerobic bacterium C. stercorarium in the yeast S.cerevisiae.
The CepA protein is natively found as a monomericprotein with a molecular mass of 93 kDa (Reichenbecher etal. 1997). SDS-PAGE results of membrane bound proteinssuggested the presence of a recombinant protein of ∼90 kDa
of the S. cerevisiae[yCEPA] strain that was not present in theS. cerevisiae[pJC1] control strain (data not shown), indicat-ing intracellular localisation of the overexpressed protein.This was confirmed by detecting enzyme activity after lysisof yeast protoplasts. The activity of CepA was measuredusing the substrate pNPG since the ability of β-glucosidaseto hydrolyse pNPG correlates with its ability to hydrolysecellobiose (Freer and Greene 1990; Reichenbecher et al.1997). The optimal pH of the recombinant cellobiosephosphorylase, pH 5, was lower than that of the nativeenzyme (pH 6–7) (Alexander 1961; Rajashekhara et al.2002; Tanaka et al. 1994). Maximum activity usingS. cerevisiae Y294[yCEPA] whole cells was also achievedat pH 5, typically the pH where maximum transfer of sugarstakes place using the proton symport mechanism (Kubiceket al. 1993). The CepA displayed a Km and Vmax of 1.69 mMand 33.89 Umin−1 for pNPG, and 363.81 mM and 3.41 Umin−1 for cellobiose respectively (Fig. 2c, d). Reichenbecheret al. (1997), however, originally reported a Km of 6.2 mMfor the native CepA on cellobiose.
Activity assays showed that the recombinant CepAenzyme was indeed constitutively produced in theS. cerevisiae Y294[yCEPA] strain, but growth was limitedon cellobiose as sole carbon source. It was previouslyobserved that the specific activity of the recombinant CepAenzyme is relatively low compared to β-glucosidaseenzymes (Skory et al. 1996; Rajoka et al. 2003; vanRooyen et al. 2005). To enable the yeast to grow oncellobiose through the phosphorolytic route, two strategieswere attempted. Firstly, we attempted to increase theamount of CepA produced by expressing a codon-optimisedversion of the gene. The codon bias index of the native cepAgene sequence was calculated for expression in S. cerevisiae(Carbone et al. 2003) and estimated to be 0.058, suggestingthat protein production would be rather low (Bennetzen andHall 1982; Gustafsson et al. 2004). Therefore, a syntheticgene (syncepA) was designed and expressed in S. cerevisiae.However, the S. cerevisiae Y294[ySynCEPA] yielded similaractivity levels as S. cerevisiae Y294[yCEPA] and bothstrains were unable to grow on cellobiose as sole carbohydratesource. The intracellular localisation of the CepA, therefore,most likely required transport of the cellobiose across the cellmembrane.
The second strategy attempted to enable the yeast togrow on cellobiose by providing a functional cellobiosetransporter or permease that would assist in cellobioseuptake. An extensive literature search revealed that no yeastcellobiose transporter or permease sequence had beenpublished to date. However, Collins et al. (1989) reportedthat cellobiose transport in E. coli is linked to lactosetransport, while Reizer et al. (1990) reported a highhomology between the cellobiose transporter of E. coliand the lactose transporter of Staphylococcus aureus. Based
Cel
lob
iose
(g
/L)
0
2
4
6
8
10
12
Time (h)0 20 40 60 80 100 120
Gro
wth
in g
DC
W/L
0
1
2
3
4
5
6
yCEPAySynCEPAyCEPA+lac(int)
ySynCEPA+lac(int)pJC1
yCEPAySynCEPAyCEPA+lac(int)
ySynCEPA+lac(int)pJC1
Time (h)0 20 40 60 80 100 120
Gro
wth
in g
DC
W/L
0
1
2
3
4
5
6
yCEPAyCEPA+lac(epi) - bufferedyCEPA+lac(epi) - unbufferedyCEPA+lac(epi) - glucosepJC1
a
b
Fig. 3 a Comparative growth (solid lines) and cellobiose consumption(dotted lines) by S. cerevisiae transformants in liquid SC−URA (10 gL−1
cellobiose). Growth was measured as DCW. Error bars indicate thestandard deviation from the mean of three different cultures. bComparative growth of S. cerevisiae Y294[yCEPA+lac(epi)] on doublestrength SC−URA medium containing 10.52 gL−1 glucose, 10 gL−1
cellobiose (buffered) and 10 gL−1 cellobiose (unbuffered) over a timeperiod of 114 h. Strains S. cerevisiae Y294[yCEPA] and S. cerevisiaeY294[pJC1] were unable to grow on cellobiose. The strain S. cerevisiaeY294[yLac12], expressing only the lac12 gene, showed growthcharacteristics similar to that of the S. cerevisiae Y294[pJC1] referencestrain and was omitted from the graph
1378 Appl Microbiol Biotechnol (2011) 90:1373–1380

on these findings we speculated that at least some lactosepermeases or transporters can transport more than one typeof disaccharide. The yeast K. lactis is able to transport andutilise cellobiose intracellularly, but the cellobiose transporterhad not yet been identified. We, therefore, postulated that theK. lactis lactose permease (Lac12) might be able to transportboth cellobiose and lactose in K. lactis.
The lac12 gene of K. lactis was cloned and expressed incombination with the cepA and syncepA genes. All yeaststrains were tested for their ability to utilise cellobiose insolid and liquid media. Only the strains expressing both thelac12 permease and the cepA or syncepA (S. cerevisiaeY294[yCEPA+lac(int)], S. cerevisiae Y294[yCEPA+lac(epi)] and S. cerevisiae Y294[ySynCEPA+lac(int)]) wereable to utilise the cellobiose displaying similar growthprofiles (Fig. 3a, b). Multicopy expression of the lac12 inS. cerevisiae Y294[yCEPA+lac(epi)] did therefore notresult in a significantly improved growth rate implying thatsingle copy integration of the lac12 was sufficient forgrowth on cellobiose. The addition of sodium phosphatebuffer enhanced growth of S. cerevisiae Y294[yCEPA+lac(epi)], probably by supplementing the insufficient inorganicphosphate in the synthetic medium (Fig. 3b). Cellobioseutilising strains were unable to utilise all the cellobiosepresent in the medium which can be attributed to the highKm of the CepA or the permease. A high Km would alsoexplain why the S. cerevisiae Y294[yCEPA+lac(epi)] strainperformed better in medium containing glucose than incellobiose (Fig. 3b).
S. cerevisiae is currently being used for bioethanolproduction with extracellular degradation of celluloseresulting in the release of glucose into the medium,increasing the possibility of contamination. Intracellularcellobiose utilisation can therefore be applied to minimisethe risk of contamination by keeping the extracellularglucose concentration to a minimum. Furthermore, strainswere recently engineered to coferment mixtures of xyloseand cellobiose, using an intracellular β-glucosidase tohydrolyse cellobiose (Ha et al. 2011). It was shown thatintracellular hydrolysis of cellobiose minimised glucoserepression of xylose fermentation allowing co-consumptionof cellobiose and xylose that improved ethanol yields.Recently, Galazka et al. (2010) were the first to reportcellodextrin transport across the cell membrane ofS. cerevisiae using the transport system of the cellulolyticfungus, Neurospora crassa. This led to the efficient growthof a recombinant strain also producing an intracellularβ-glucosidase on cellodextrins up to cellotetraose. Howev-er, our study confirms that the Lac12 of K. lactis was ableto transport cellobiose when expressed in S. cerevisiae andmost probably also in K. lactis. To our knowledge, this isthe first report of the functional expression of a cellobiosephosphorylase in S. cerevisiae and the first report of
cellobiose transport enabled by a disaccharide permease ofyeast origin.
Acknowledgements The authors would like to thank the NationalResearch Foundation (NRF) for financial support.
References
Alexander JK (1961) Characteristics of cellobiose phosphorylase. JBacteriol 81:903–910
Bennetzen JL, Hall BD (1982) Codon selection in yeast. J Biol Chem257:3026–3031
Bradford MM (1976) A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein-dye binding. Anal Biochem 72:248–254
Carbone A, Zinovyev A, Kepes F (2003) Codon adaptation index as ameasure of dominating codon bias. Bioinformatics 19:2005–2015
Collins JC, Permuth SF, Brooker RJ (1989) Isolation and character-ization of lactose permease mutants with an enhanced recognitionof maltose and diminished recognition of cellobiose. J Biol Chem264:14698–14703
Crous JM, Pretorius IS, van Zyl WH (1995) Cloning and expressionof an Aspergillus kawachii endo-1, 4-β-xylanase gene inSaccharomyces cerevisiae. Curr Genet 28:467–473
Den Haan R, McBride JE, La Grange DC, Lynd LR, Van Zyl WH(2007) Functional expression of cellobiohydrolases in Saccharo-myces cerevisiae towards one-step conversion of cellulose toethanol. Enzyme Microb Technol 40:1291–1299
Freer SN (1991) Fermentation and aerobic metabolism of cellodex-trins by yeasts. Appl Environ Microbiol 57:655–659
Freer SN, Greene RV (1990) Transport of glucose and cellobiose byCandida wickerhamii and Clavispora lusitaniae. J Biol Chem265:12864–12868
Fujita Y, Takahashi S, Ueda M, Tanaka A, Okada H, Morikawa Y,Kawaguchi T, Arai M, Fukuda H, Kondo A (2002) Direct andefficient production of ethanol from cellulosic material with ayeast strain displaying cellulolytic enzymes. Appl EnvironMicrobiol 68:5136–5141
Galazka JM, Tian C, Beeson WT, Martinez B, Glass NL, Cate JH(2010) Cellodextrin transport in yeast for improved biofuelproduction. Science 330:84–86
Gonde P, Blondin B, Leclerc M, Ratomahenina R, Arnaud A, Galzy P(1984) Fermentation of cellodextrins by different yeast strains.Appl Environ Microbiol 48:265–269
Gustafsson CS, Govindarajan S, Minshull J (2004) Codon bias andheterologous protein expression. Trends Biotechnol 22:346–353
Ha SJ, Galazka JM, Rin Kim S, Choi JH, Yang X, Seo JH, Glass NL,Cate JH, Jin YS (2011) Engineered Saccharomyces cerevisiaecapable of simultaneous cellobiose and xylose fermentation. ProcNatl Acad Sci USA 108:504–509
Hill J, Donald KA, Griffiths DE, Donald G (1991) DMSO-enhancedwhole cell yeast transformation. Nucleic Acids Res 19:5791
Kosaric N, Pieper HJ, Vardar-Sukan F (2001) The biotechnologyof ethanol. Classical and future applications. Wiley-VCH,Weinheim
Kubicek CP, Messner R, Gruber F, Mandels M, Kubicek-Pranz EM(1993) Triggering of cellulase biosynthesis by cellulose inTrichoderma reesei. Involvement of a constitutive, sophorose-inducible, glucose-inhibited β-diglucoside permease. J BiolChem 268:19364–19368
la Grange DC, Pretorius IS, van Zyl WH (1996) Expression of aTrichoderma reesei β-xylanase gene (XYN2) in Saccharomycescerevisiae. Appl Environ Microbiol 62:1036–1044
Appl Microbiol Biotechnol (2011) 90:1373–1380 1379

Lynd LR, Wyman CE, Gerngross TU (1999) Biocommodity engi-neering. Biotechnol Prog 15:777–793
Lynd LR, Weimer PJ, van Zyl WH, Pretorius IS (2002) Microbialcellulose utilization: fundamentals and biotechnology. MicrobiolMol Biol Rev 66:506–577
Lynd LR, van Zyl WH, McBride JE, Laser M (2005) Consolidatedbioprocessing of cellulosic biomass: an update. Curr OpinBiotechnol 16:577–583
McBride JE, Zietsman JJ, van Zyl WH, Lynd LR (2005) Utilization ofcellobiose by recombinant β-glucosidase-expressing strains ofSaccharomyces cerevisiae: characterization and evaluation of thesufficiency of expression. Enzyme Microb Technol 37:93–101
Rajashekhara E, Kitaoka M, Kim YK, Hayashi K (2002) Character-ization of a cellobiose phosphorylase from a hyperthermophiliceubacterium, Thermotoga maritima MSB8. Biosci BiotechnolBiochem 66:2578–2586
Rajoka MI, Shaukat F, Ghauri MT, Shahid R (2003) Kinetics of β-glucosidase production by Saccharomyces cerevisiae recombinantsharboring heterologous bgl genes. Biotechnol Lett 25:945–948
Reichenbecher M, Lottspeich F, Bronnenmeier K (1997) Purificationand properties of a cellobiose phosphorylase (CepA) and acellodextrin phosphorylase (CepB) from the cellulolytic thermo-phile Clostridium stercorarium. Eur J Biochem 247:262–267
Reizer J, Reizer A, Saier MH Jr (1990) The cellobiose permease ofEscherichia coli consists of three proteins and is homologous tothe lactose permease of Staphylococcus aureus. Res Microbiol141:1061–1067
Sambrook J, Fritsch EF,Maniatis T (1989)Molecular cloning: a laboratorymanual. Cold Spring Harbor Laboratory, Cold Spring Harbor
Skory CD, Freer SN, Bothast RJ (1996) Expression and secretion ofthe Candida wickerhamii extracellular β-glucosidase gene, bglB,in Saccharomyces cerevisiae. Curr Genet 30:417–422
Sreekrishna K, Dickson RC (1985) Construction of strains ofSaccharomyces cerevisiae that grow on lactose. Proc Natl AcadSci USA 82:7909–7913
Stambuk BU, Batista AS, de Araujo PS (2000) Kinetics of activesucrose transport in Saccharomyces cerevisiae. J Biosci Bioeng89:212–214
Sun Y, Cheng J (2002) Hydrolysis of lignocellulosic materials forethanol production: a review. Bioresour Technol 83:1–11
Tanaka K, Kawaguchi T, Imada Y, Ooi T, Arai M (1994) Purificationand properties of cellobiose phosphorylase from Clostridiumthermocellum. J Ferment Bioeng 79:212–216
Tautz D, Renz M (1983) An optimized freeze–squeeze method for therecovery of DNA fragments from agarose gels. Anal Biochem132:14–19
Tingle M, Halvorson HO (1971) A comparison of β-glucanase and β-glucosidase in Saccharomyces lactis. Biochim Biophys Acta250:165–171
Tingle M, Halvorson HO (1972) Biochemical and genetic character-ization of 3 β-glucosidase mutants in Saccharomyces lactis. JBacteriol 110:196–201
van der Rest ME, Kamminga AH, Nakano A, Anraku Y, Poolman B,Konings WN (1995) The plasma membrane of Saccharomycescerevisiae: structure, function, and biogenesis. Microbiol Rev59:304–322
van Rensburg P, van Zyl WH, Pretorius IS (1998) Engineering yeastfor efficient cellulose degradation. Yeast 14:67–76
van Rooyen R, Hahn-Hägerdal B, la Grange DC, van Zyl WH (2005)Construction of cellobiose-growing and fermenting Saccharomycescerevisiae strains. J Biotechnol 120:284–295
Yernool DA, McCarthy JK, Eveleigh DE, Bok JD (2000) Cloning andcharacterization of the glucooligosaccharide catabolic pathwayβ-glucan glucohydrolase and cellobiose phosphorylase in themarine hyperthermophile Thermotoga neapolitana. J Bacteriol182:5172–5179
Zhang YH, Lynd LR (2004) Kinetics and relative importance ofphosphorolytic and hydrolytic cleavage of cellodextrins andcellobiose in cell extracts of Clostridium thermocellum. ApplEnviron Microbiol 70:1563–1569
1380 Appl Microbiol Biotechnol (2011) 90:1373–1380