cht1, an ig superfamily molecule required for t cell differentiation
TRANSCRIPT

Seediscussions,stats,andauthorprofilesforthispublicationat:https://www.researchgate.net/publication/13068379
ChT1,anIgsuperfamilymoleculerequiredforTcelldifferentiation
ARTICLEinTHEJOURNALOFIMMUNOLOGY·JUNE1999
ImpactFactor:4.92·Source:PubMed
CITATIONS
17
READS
24
8AUTHORS,INCLUDING:
BeatAImhof
UniversityofGeneva
219PUBLICATIONS10,012CITATIONS
SEEPROFILE
RichardBoyd
MonashUniversity(Australia)
209PUBLICATIONS7,705CITATIONS
SEEPROFILE
AnnPChidgey
MonashUniversity(Australia)
66PUBLICATIONS1,915CITATIONS
SEEPROFILE
OlliVainio
UniversityofOulu
196PUBLICATIONS3,289CITATIONS
SEEPROFILE
Availablefrom:AnnPChidgey
Retrievedon:04February2016

of June 13, 2013.This information is current as
for T Cell DifferentiationChT1, an Ig Superfamily Molecule Required
Olli VainioAndrew Bean, Dominique Dunon, Thomas W. F. Göbel and Kaisa Katevuo, Beat A. Imhof, Richard Boyd, Ann Chidgey,
http://www.jimmunol.org/content/162/10/56851999; 162:5685-5694; ;J Immunol
Referenceshttp://www.jimmunol.org/content/162/10/5685.full#ref-list-1
, 16 of which you can access for free at: cites 52 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 1999 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

ChT1, an Ig Superfamily Molecule Required for T CellDifferentiation 1,2
Kaisa Katevuo,3* Beat A. Imhof,† Richard Boyd,‡ Ann Chidgey,‡ Andrew Bean,‡
Dominique Dunon,§ Thomas W. F. Gobel,¶ and Olli Vainio*
The thymus is colonized by circulating progenitor cells that differentiate into mature T cells under the influence of the thymicmicroenvironment. We report here the cloning and function of the avian thymocyte Ag ChT1, a member of the Ig superfamily withone V-like and one C2-like domain. ChT1-positive embryonic bone marrow cells coexpressing c-kit give rise to mature T cells uponintrathymic cell transfer. ChT1-specific Ab inhibits T cell differentiation in embryonic thymic organ cultures and in thymocyteprecursor cocultures on stromal cells. Thus, we provide clear evidence that ChT1 is a novel Ag on early T cell progenitors thatplays an important role in the early stages of T cell development. The Journal of Immunology,1999, 162: 5685–5694.
T cell differentiation follows immediately after colonizationof the thymus by pluripotential hemopoietic progenitorcells and is driven by the interaction of cell surface mol-
ecules and cytokines. Most immature cells found in the thymusmay not yet be committed to the T lineage, since they can stilldifferentiate into T, B, NK, and dendritic cells (1–5). The ensuingdifferentiation pathways are complex, and although multiple stagesof development can be identified, it is a continuous process ratherthan a series of precise shifts. Each level of maturation is definednot by unique stage-specific markers but by a phenotypic profilebased on a constellation of membrane molecules (1, 6–9). Eventhe earliest intrathymic precursors lacking TCR, CD4 and CD8,which represent,5% of the total thymocyte pool, are divisibleinto at least four subsets based on differential expression of CD117(c-kit), CD44 (Pgp-1), and CD25 (IL-2Ra) (2, 8, 10). None ofthese markers, however, is T-lineage specific. In fact, no cell sur-face molecule has yet been identified that exclusively distinguishesT-lineage cells from the earliest precursors.
Although many studies have been performed in mice, thechicken offers an exceptional model for analyzing the events ofstem cell differentiation and T cell development because of theeasy access to embryos at exact stages. A further advantage of thechicken is that T and B cells develop in separate tissues, thymus
and bursa, respectively. Lymphoid precursors colonize the embry-onic thymus in three distinct waves, beginning on embryonic days(E)4 6.5, 12, and 18 (11–14). It has been shown that T cell pro-genitors seeding the thymus during the second period of coloni-zation on E12 stem from the embryonic bone marrow (15) andexpress the receptor-type tyrosine kinase c-kit and adhesion mol-ecules HEMCAM and BEN (16, 17). These three marker mole-cules are useful in defining T cell progenitors and immature thy-mocytes, but their distribution is again not restricted to the Tlineage. We therefore searched for new molecules that would al-low further characterization of hemopoietic progenitors and lead tobetter definition of discrete stages of thymocyte differentiation oc-curring before TCR expression. ChT1 is a T cell Ag that has beenshown to be expressed on the surface of thymocytes by E10. Inyoung chickens about 90% of the thymocytes and a subpopulationof peripheral lymphocytes, representing the recent thymic emi-grants, are ChT1 positive (18–20). Expression in the peripherydeclines with age and also in correlation with partial thymectomy,indicating that ChT1 can be used as an accurate marker for study-ing thymic function (20).
In the present work we cloned ChT1 cDNA from two thymuscDNA libraries. The Ag was identified as a member of the Igsuperfamily (IgSF) with two extracellular Ig domains. We alsoshowed that ChT1 is a close homologue of CTX, a recently clonedXenopusAg. Intrathymic cell transfer of ChT11c-kit1 bone mar-row cells into congenic animals defined a T cell progenitor pop-ulation in embryonic bone marrow. In embryonic thymus organcultures (ETOC) and in in vitro thymic stromal cell and thymocyteprecursor cocultures the differentiation of T cells was inhibited atthe CD32CD42CD82 stage. These data provide distinct evidencethat ChT1 is a novel IgSF molecule that has a functional role in thedifferentiation of T cells from embryonic bone marrow precursors.
Materials and MethodsAnimals
H.B2 and H.B15 chickens and chicken embryos from the colonies at theDepartment of Medical Microbiology, Turku University (Turku, Finland),two ov-Ag congenic H.B19 strains at the Basel Institute for Immunology
*Turku Immunology Center, Department of Medical Microbiology, University ofTurku, Turku, Finland;†Department of Pathology, Centre Medical Universitaire, Uni-versity of Geneva, Geneva, Switzerland;‡Department of Pathology and Immunology,Monash University Medical School, Prahran, Melbourne, Victoria, Australia;§Unitede Recherche Associee-Centre National de la Recherche Scientifique 1135, Univer-sity of Pierre and Marie Curie, Paris, France; and¶Basel Institute for Immunology,Basel, Switzerland
Received for publication June 22, 1998. Accepted for publication February 26, 1999.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisementin accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by the Turku University Foundation, the Academy ofFinland, the Human Frontier Science Program (RG 366/96), the Swiss National Sci-ence Foundation (Grant 31-49241.96), the National Health and Medical ResearchCouncil of Australia, and the Australian Chicken Meat Research and DevelopmentCouncil. The Basel Institute for Immunology was founded and is fully supported byF. Hoffmann-La Roche (Basel, Switzerland).2 The following cDNA sequences have been submitted to the EMBL database withaccession numbers Y14063 and Y14064.3 Address correspondence and reprint requests to Dr. Kaisa Katevuo, Turku Immu-nology Center, Department of Medical Microbiology, University of Turku, Kiinamyl-lynkatu 13, 20520 Turku, Finland. E-mail address: [email protected]
4 Abbreviations used in this paper: E, day of embryonic development; IgSF, Ig superfamily; ETOC, embryonic thymus organ culture; CU20, CU32, and CU36, Marek’sdisease virus-transformed MDCC-CU20, MDCC-CU32, and MDCC-CU36 cell lines,respectively; RP13, reticuloendothelial virus-transformed REVCC-RP13 chicken cellline.
Copyright © 1999 by The American Association of Immunologists 0022-1767/99/$02.00
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

(Gipf-Oberfrick, Switzerland), and the Australorp3 White Leghorn hy-brids at the Research Poultry Farm (Research, Australia) were used. Thetwo congenic lines, H.B19ov1 and H.B19ov2, differ in their expression ofthe ov-Ag on thymocytes and T cells. The ov-Ag is recognized by mAb11A9 (15, 21).
Abs and flow cytometry
Abs detecting the ChT1 Ag were T10A6 (IgG1) (19), CT1 (IgG1) and CT1a(IgG3) (18), RR5-89 (IgG2b), cF3c210 (IgM), MUI-83 (IgG1) (22), and2-1 (IgG1). Other Abs were kit2c75 (IgG2a) against c-kit (16), 11A9 (IgM)against ov-Ag, 2-6 (IgG1) and 2-35 (IgG2b) against CD4 (23), 11-39(IgG1), 3-298 (IgG2b) against CD8a (24), MUI-78 (IgG2a) against MHCclass II (22, 25), 10-2.16 (IgG1) against mouse I-Ak (26), MUI-36 (IgG2a),MUI-53 (IgM), MUI-54 (IgM), MUI-70 (IgM), MUI-71 (IgM), andMUI-82 (IgG1). TCR1 (IgG1) againstgdTCR, TCR2 (IgG1) againstab1TCR, TCR3 (IgG1) againstab2TCR, and CT3 (IgG1) against CD3were purchased from Southern Biotechnology Associates (Birmingham,AL). Abs were used as ascites, hybridoma supernatant, or purified Ab. Abpurification and conjugation to biotin (biotin-N-hydroxysuccinimidester,NHS-D-biotin, Calbiochem, La Jolla, CA) and PE (R-PE, MolecularProbes, Leiden, The Netherlands) were performed using standard proce-dures.
For one-, two-, and three-color immunofluorescence, cell suspensionsfrom thymus, spleen, PBL, and bone marrow cells were prepared as de-scribed previously (16, 27). Cells (13 106) were incubated with primarymAb followed by incubation with FITC-conjugated anti-mouse Ig (SilenusLaboratories, Hawthorn, Australia) or anti-mouse Ig isotype-specific Abs(Southern Biotechnology Associates). Thereafter, the cells were incubatedwith secondary mAb, which were used as hybridoma culture supernatantsor PE conjugates. The unconjugated mAb were detected with PE-conju-gated anti-mouse Ig isotype-specific Abs (Southern Biotechnology Asso-ciates). For three-color analysis cells were blocked with 1% normal mouseserum and further incubated with biotinylated tertiary mAb followed bystreptavidin Tri-color reagent (Caltag, South San Francisco, CA). Aftereach step the cells were washed twice with PBS containing 2% FCS and0.01% NaN3. Immunofluorescence analysis was performed with a FACS-can instrument (Becton Dickinson, Mountain View, CA).
Intrathymic injection and differentiation of embryonic bonemarrow cells
Intrathymic injections were performed as described previously (16, 28). Inbrief, bone marrow cells from congenic ov1 E13.5 donor animals of theH.B19 strain were flushed from the cavity of the femurs and tibiae, washedtwice in PBS, counted, and adjusted to the required cell concentration (15).Before receiving the bone marrow cells, the recipients, 14-day-old ov2
congenic H.B19 chicks, were irradiated with 600 rad (Gamma Cell irra-diator, Atomic Energy of Canada, Ottawa, Canada). For the intrathymicinjection the recipients were anesthetized with an injection of 0.4 ml ofketamine (Imalgene 500, Rhone Merieux, France) diluted 1/10 in PBS i.m.followed by a short inhalation of halotane (Hoechst, Frankfurt, Germany).The donor bone marrow cells were injected into two upper lobes on eachside through a dorsal incision in the skin. Ten microliters of cell suspension(1000 or 100 cells/thymus lobe) in PBS was injected. Donor-derived cellsfrom the injected thymus lobes were identified by immunofluorescence 2wk after chimera construction with the anti-ov mAb 11A9.
Immunoprecipitation and peptide mapping
The lactoperoxidase-catalyzed method (29) was used to surface label 53107 thymocytes (viability,.95%) with 1 mCi of Na125I (Amersham, Ay-lesbury, U.K.). The cells were lysed in 150 mM NaCl, 50 mM Tris-HCl(pH 7.4), 5 mM EDTA, 0.1% NaN3, 1 mM Pefabloc (Boehringer Mann-heim, Indianapolis, IN), 1mg/ml leupeptin, 1mg/ml pepstatin A, 100mg/ml trypsin inhibitor, 20 mMe-amino-n-caproic-acid, 2mg/ml antipain,20 mM iodoacetamide (all reagents from Sigma, St. Louis, IL), and 1%Nonidet P-40 (Calbiochem, La Jolla, CA) for 45 min at 4°C. Insolubleparticles were removed by centrifugation (10,0003 g, 30 min, 4°C). Thelysate was then immunoprecipitated using a solid phase method (30). Theabsorbed molecules were eluted with Laemmli sample buffer for 45 min at37°C or with 0.5% SDS/0.1 M 2-ME for 20 min at 80°C for glycolyticenzyme digestions.N-linked carbohydrates were removed by incubatingthe samples in the presence of 50 U/mlN-glycanase at 37°C overnight.Sialic acids were digested with 3 U/ml neuraminidase at 37°C for 2 h aftersetting the pH to 6.3 with 1 M CH3COOH; after this treatment theO-linkedsugars were removed by incubation with 82 mU/ml ofO-glycanase (allenzymes from Genzyme, Boston, MA) at 37°C overnight. Undigested con-trols for each enzyme were treated similarly but without adding the en-
zyme. For electrophoresis, 63 Laemmli sample buffer was added, andsamples were boiled for 5 min. Reduced conditions were obtained by add-ing 5% 2-ME before boiling. Electrophoresis was performed on 5–15%gradient SDS-PAGE.
Peptide mapping by proteolysis with endoproteinase Glu-C (proteaseV8, Boehringer Mannheim) in SDS was performed basically as describedby Cleveland (31). Briefly, after exposing the immunoprecipitates to anautoradiography film (Kodak X-OMAT AR, Eastman Kodak, Rochester,NY) the bands were cut out from the dried gel with a sharp scalpel. The gelslices were rehydrated in a sample buffer with 1 mM EDTA for 20 min,placed in the sample wells of a second 5–15% SDS gel, and overlaid with10 ml of 20% and 10ml of 10% glycerol in sample buffer and finally with1 mg (in H2O) of the protease. Electrophoresis was started, and after thesamples had reached the stacking gel the run was stopped, and the gel wasincubated at 37°C for 60 min to allow protein digestion to proceed. There-after electrophoresis was completed.
Ag purification
The ChT1 Ag was purified from chicken thymus Nonidet P-40 lysates byan immunoaffinity column as described previously (32). Samples were runon a 12.5% SDS-PAGE, and the proteins were visualized by silver staining.The relatively pure protein was sequenced directly on a 475A Protein Se-quencer (Applied Biosystems, Foster City, CA). Half the material wasdigested with endoproteinase Lys-C (Boehringer Mannheim) at a ratio of1/100 at 37°C overnight, and the resulting fragments were separated byreverse phase HPLC and analyzed by microsequencing to obtain internalpeptide sequences.
cDNA cloning and sequencing
The COS cell expression screening method was used to clone the cDNAencoding for ChT1 Ag from two different chicken cDNA libraries. Thethymus cDNA library in pCDM8 vector from an adult RPRL Line 0 animalwas a gift from Dr. J. R. Young. The other library was constructed frommRNA of strain H.B19 E13 thymus into vector pcDNA3 (16). The trans-fection and staining were performed as described by Tregaskes and Young(33). Positively stained cells were picked up from the slide with a Drum-mond sequencing pipette (Drummond Scientific, Broomall, PA). Proteinswere precipitated, after which the DNA was extracted. The plasmid DNAcontaining the desired cDNA insert was then introduced toEscherichia coli(MC1061/p3 or TOP10F9) by electroporation. For sequencing of thecDNA, the plasmid DNA was extracted using Qiagen spin columns (Qia-gen, Chatsworth, CA) and was sequenced with either dideoxy chain ter-mination (Sequenase version 2.0, U.S. Biochemical Corp., Cleveland, OH)or automated cycle sequencing (ABI PRISM Dye Terminator Cycle Se-quencing Ready Reaction Kit, Perkin-Elmer, Norwalk, CT) with an ABI373A DNA Sequencer (Applied Biosystems). The cDNA sequences havebeen submitted to the EMBL database with accession numbers Y14063 andY14064. Sequence data analysis and sequence comparisons were per-formed using the Wisconsin University software package GCG (GeneticsComputer Group, Madison, WI), Lasergene (DNAStar, Madison, WI), andBlast (National Center for Biotechnology Information, Bethesda, MD).
Analysis of mRNA expression
Northern blot was performed with mRNA isolated from H.B2 chickenthymus, spleen, liver, bursa, ileum, colon, brain, lung, kidney, testis, andoviduct as well as with Marek’s disease virus-transformed MDCC-CU32(CU32), MDCC-CU36 (CU36) and reticuloendothelial virus-transformedREVCC-RP13 (RP13) chicken cell lines using RNeasy and OligotexmRNA Spin column kits (Qiagen). After electrophoretic separation on a1.2% formaldehyde agarose gel the samples were transferred overnight toa nylon membrane (Hybond-N1, Amersham). The membrane was prehy-bridized in 1% SDS, 53 Denhardt’s solution, 23 SSPE, 10% dextransulfate, and 20mg/ml ssDNA at 60°C for 6 h. For hybridization the mem-brane was incubated at 60°C overnight in fresh hybridization solution withtotal cDNA of ChT1 or a PCR product that encodes for a 1030-bp part ofthe chickenb-actin cDNA (34) as probes. These were labeled with[a-32P]dCTP (3000 Ci/mM; Amersham) using a Rediprime labeling kit(Amersham). Membrane was washed in 23 SSC/1% SDS at 60°C for 30min followed by washing in 0.53SSC/1% SDS at 60°C for 15 min. Sig-nals were visualized by autoradiography.
To study the mRNA expression of ChT1 further by RT-PCR, 0.1mg oftotal RNA from spleen, bursa, liver, small intestine, skeletal muscle, andMarek’s disease virus-transformed MDCC-CU20 (CU20), CU36, andRP13 cells was used in 50ml of RT-PCR reaction (Titan One Tube RT-PCR Kit, Boehringer Mannheim, Mannheim, Germany). The SMARTPCR cDNA Library Construction Kit (Clontech, Palo Alto, CA) was usedaccording to the manufacturer’s instructions to prepare cDNA from E13
5686 ChT1, A NEW MARKER IN T CELL DIFFERENTIATION
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from
bone marrow mRNA for analysis of ChT1 expression in the bone marrow.One microliter of amplified cDNA was used for PCR in a 50-ml reaction,and the PCR conditions were as follows: 96°C for 2 min; then 35 cycles of96°C for 30 s, 52°C for 30 s, and 72°C for 1 min; and a final extension at72°C for 5 min. ChT1-specific oligonucleotides were V59-GTG ACC GTTCCT GAG AAG- and C39-GTT GCT GGC TAT GCA TCG-, which am-plify a 572-bp part of the extracellular region of ChT1.b-Actin was usedas a positive control, and H2O was used as a negative control in PCRreactions. After PCR, 15ml of each sample was run on a 1.2% agarose gel,transferred to a nylon membrane, and hybridized at 65°C overnight in 23SSC/1% SDS, 10% dextran sulfate, 0.5% pyrophosphate, 0.5 mg/mlssDNA, and 0.05% low fat powdered milk. Oligonucleotides, V39-CAGCAC ATT GAC AAT CAC TG- and 4611-TAC CAC AAT GTA CCCTGG C- (35), labeled with [g-32P]ATP (10 mCi/ml; Amersham) by T4kinase (Life Technologies, Gaithersburg, MD) were used as probes forChT1 andb-actin, respectively. Washes were performed at 65°C in 23SSC. Radioactive signal was exposed to film.
Embryonic thymus organ culture
For ETOC, thymus lobes were isolated on E10 and cultured for 6 days aspreviously described (36) in medium that contained purified ChT1 in aconcentration of 100mg/ml specific mAb MUI-83 (dialyzed against RPMI1640 culture medium). Control cultures lacked mAb, contained an irrele-vant, isotype-matched control mAb (10-2.16) that was not reactive withchicken tissues, or contained other chicken thymus- or stromal cell-specificmAb (MUI-36, MUI-53, MUI-54, MUI-70, MUI-71, MUI-78, MUI-82).The results were identical for all control conditions; hence, only the onewith isotype-matched control mAb has been included herein. The fre-quency and cell yield of the various thymocyte subsets present in controlvs mAb-treated ETOC were statistically compared using unpaired Stu-dent’s t test (p,p , 0.05; ‡,p # 0.01; §,p # 0.001).
In vitro thymic stromal cell and thymocyte precursor coculture
In vitro thymic stromal cell and thymocyte precursor coculture was per-formed as described previously (37). E13 T cell precursors were obtainedfrom ov1 congenic chicks. The thymocyte suspensions were labeled with
anti-CD4 and anti-CD8 mAb and then sorted for CD42CD82 lymphoidcells on a FACStar Plus (Becton Dickinson). The sorted CD42CD82 cellswere prepared at a concentration of 3.43 106 cells/ml. Where relevant,purified anti-ChT1 (MUI-83) and anti-c-kit (kit2c75) Abs were added at aconcentration of 100mg/ml. Several control mAbs that were reactive witheither chicken thymocytes or stromal cells were also used. Stromal cellsfrom adult chickens (ov2) were prepared by enzymatic digestion of lym-phocyte-depleted thymi using 0.15% collagenase/0.1% DNase (BoehringerMannheim). Stromal cells were enriched by elutriation and resuspended ata concentration of 6.73 105 cells/ml (37, 38). T cell precursors weremixed with the stromal cells at a ratio of 5:1 and cocultured as hangingdrops in inverted Terasaki plates at 40°C in 10% CO2. Cells were harvestedon day 5 and were stained with anti-ov mAb 11A9 followed by stainingwith anti-CD4 and anti-CD8 mAb. Cells were analyzed by flow cytometryusing a FACScan (Becton Dickinson).
ResultsTissue distribution and ontogeny of ChT1
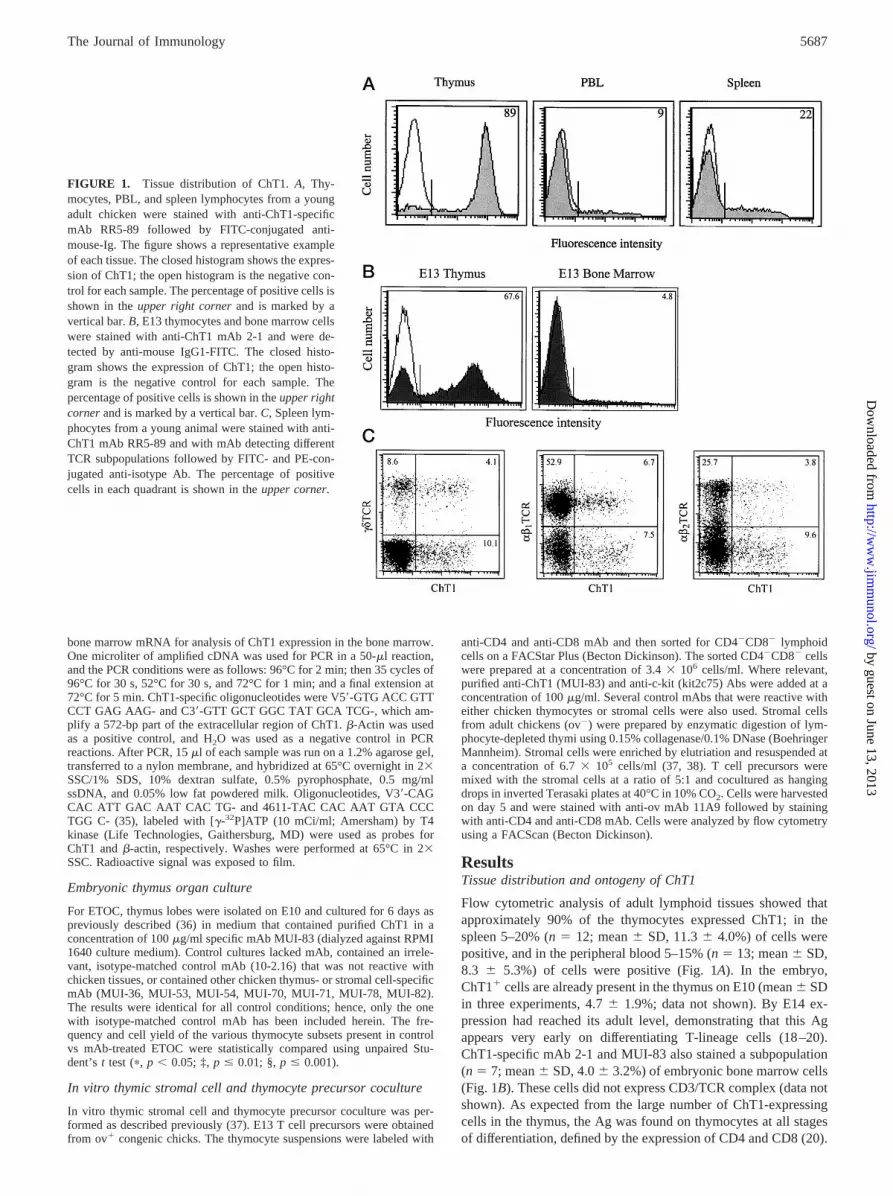
Flow cytometric analysis of adult lymphoid tissues showed thatapproximately 90% of the thymocytes expressed ChT1; in thespleen 5–20% (n5 12; mean6 SD, 11.36 4.0%) of cells werepositive, and in the peripheral blood 5–15% (n 5 13; mean6 SD,8.3 6 5.3%) of cells were positive (Fig. 1A). In the embryo,ChT11 cells are already present in the thymus on E10 (mean6 SDin three experiments, 4.76 1.9%; data not shown). By E14 ex-pression had reached its adult level, demonstrating that this Agappears very early on differentiating T-lineage cells (18–20).ChT1-specific mAb 2-1 and MUI-83 also stained a subpopulation(n 5 7; mean6 SD, 4.06 3.2%) of embryonic bone marrow cells(Fig. 1B). These cells did not express CD3/TCR complex (data notshown). As expected from the large number of ChT1-expressingcells in the thymus, the Ag was found on thymocytes at all stagesof differentiation, defined by the expression of CD4 and CD8 (20).
FIGURE 1. Tissue distribution of ChT1.A, Thy-mocytes, PBL, and spleen lymphocytes from a youngadult chicken were stained with anti-ChT1-specificmAb RR5-89 followed by FITC-conjugated anti-mouse-Ig. The figure shows a representative exampleof each tissue. The closed histogram shows the expres-sion of ChT1; the open histogram is the negative con-trol for each sample. The percentage of positive cells isshown in theupper right cornerand is marked by avertical bar.B, E13 thymocytes and bone marrow cellswere stained with anti-ChT1 mAb 2-1 and were de-tected by anti-mouse IgG1-FITC. The closed histo-gram shows the expression of ChT1; the open histo-gram is the negative control for each sample. Thepercentage of positive cells is shown in theupper rightcornerand is marked by a vertical bar.C, Spleen lym-phocytes from a young animal were stained with anti-ChT1 mAb RR5-89 and with mAb detecting differentTCR subpopulations followed by FITC- and PE-con-jugated anti-isotype Ab. The percentage of positivecells in each quadrant is shown in theupper corner.
5687The Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

ChT11 thymocytes and peripheral T cells includedgd T cells aswell as both Vb1- and Vb2-expressingab T cells (Fig. 1C). To-gether these results show that ChT1 is highly expressed on mostthymocytes and prethymically on a subpopulation of embryonicbone marrow cells as well as on a subset of peripheral T cells.
A ChT1-positive population of embryonic bone marrow cellscontains T cell progenitors
The E13.5 bone marrow cells can be divided into ChT11c-kit1
and ChT12c-kit1 populations (Fig. 2A). To determine whetherChT1 is expressed on cells that have a capacity to differentiate intoT cells in vivo, we sorted these two cell populations from E13.5bone marrow of congenic H.B19ov1 animals and injected the cellsintrathymically into 14-day-old H.B19ov2 recipients. Thymus re-
constitution was measured by flow cytometry with the ov-alloan-tigen-specific mAb 11A9 at 2 wk after injection. A clear thymusreconstitution was obtained by 1000 or even 100 ChT11 c-kit1
cells with mean chimerism of 13.7 and 6.8%, respectively. In con-trast, the injection of ChT12c-kit1 cells resulted in only 2.9 and0.9% chimerism, respectively (Table I and Fig. 2B). The chimericcells were analyzed for their capacity for thymus reconstitution bythe surface expression of CD4, CD8, and TCR. The majority of thedonor cells expressed both CD4 and CD8, and only a small frac-tion of the injected cells had remained at the CD42CD82 stage.The bone marrow progenitors also developed into CD4 and CD8single-positive thymocytes (Fig. 2C). Differentiated thymocytesexpressed bothgd- andab-TCR, indicating that the ChT11c-kit1
embryonic bone marrow precursors have a capacity to maturealong all pathways of T cell development (data not shown).
ChT1 cDNA cloning and protein characterization
To clone the cDNA encoding the ChT1 Ag, we screened COS cellstransiently transfected with an adult RPRL Line 0 thymus cDNAlibrary with various mAb. Clone p10.6 (1114 bp) was isolated withmAb T10A6. The specificity of other mAb (e.g., 2-1, RR5-89,MUI-83, CT1, CT1a) for ChT1 was confirmed by their reactivitywith p10.6-transfected COS cells. Another independent clone,pc210 (1087 bp), was isolated with mAb cF3c210 from a cDNAlibrary made from E13 H.B19 thymus mRNA. The complete nu-cleotide sequence from both strands of these two clones was thendetermined (Fig. 3). The cDNA clones p10.6 and pc210 consist ofa 28-bp and an 8-bp, respectively, 59 untranslated region, a1008-bp open reading frame encoding a 21-aa leader peptide, a212-aa extracellular region, a 24-aa transmembrane region, and a78-aa cytoplasmic part followed by a 78-bp and a 71-bp 39 un-translated region that does not contain a poly(A) tail. The codingregion sequences of the two clones are identical except for onenucleotide difference in the transmembrane region at position 765,where an A is replaced by a C in thepc210 clone. This changes acharged aspartic acid residue to an aliphatic alanine residue in thededuced protein sequence. The predicted molecular mass of ChT1is 36.5 kDa.
To further confirm the specificity of mAb and to clarify the startof the mature polypeptide, the ChT1 protein was purified fromthymus lysates by an immunoaffinity column using mAb RR5-89.N-terminal microsequencing of the purified protein of 46 kDa re-sulted in a 30-residue sequence, VVVTVPEKTVNVKTGGx-ATLLxTYTSSQPL, which completely matched the predictedamino acid sequence. Internal peptides were obtained by partialdigestion with an endoproteinase Lys-C. These peptides were se-quenced, and two sequences, residues 87–96 (DRITAATSPG) and
FIGURE 2. Thymus reconstitution capacity of ChT11c-kit1 embryonicbone marrow cells.A, E13.5 bone marrow cells from ov1 strain werestained with anti-ChT1 and anti-c-kit mAb. The cells were sorted intoChT11c-kit1 (region 1) and ChT12c-kit1 (region 2) populations for in-trathymic injections into ov2 congenic chickens.B, Two weeks after theintrathymic injection of E13.5 bone marrow cells, the donor cells in therecipient thymi were analyzed by staining with anti-ov-specific mAb 11A9.The histogram shows a representative example of the reconstitution capac-ity of 1000 ChT11c-kit1 cells from three experiments. The horizontal barmarks the ov1 population.C, The ov1 cells from B were gated and ana-lyzed for CD4 and CD8 expression to study the T cell differentiation of theinjected cells.
Table I. Thymus reconstitution capacity of ChT11c-kit1 and ChT12c-kit1 cellsa
Injected CellPopulation
No. of Injected Cells/Thymus Lobe
ov1 Cells/Animal(%)
Mean6 SD(%)
ChT11c-kit1 1000 13.8, 14.7, 12.7 13.76 1.0100 4.6, 9.4, 6.5 6.86 2.4
ChT12c-kit1 1000 1.1, 2.0, 2.9 2.06 0.9100 0.7, 1.1, 1.0 0.96 0.2
a E13.5 ov1 bone marrow cells were sorted into two populations (ChT11 c-kit1
and ChT12c-kit1) and then injected into two upper thymus lobes of the recipient ov2
14-day-old animals. The chimerism was analyzed 2 wk later by flow cytometry afterstaining thymocytes with anti-ov mAb.
5688 ChT1, A NEW MARKER IN T CELL DIFFERENTIATION
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

residues 130–150 (xVIVNVLVKPSKPFxKIEGTP), were assem-bled. Both internal sequences showed 100% identity to the de-duced amino acid sequence and spanned the first half of the ex-tracellular region.
The anti-ChT1-specific mAb immunoprecipitated four bandsfrom a surface-labeled thymocyte lysate as analyzed by SDS-PAGE: a major band of 63 kDa and its putative dimer of 138 kDaas well as minor bands of 46 and 121 kDa, which are probablydegradation products of the monomer and putative dimer bands,respectively (Fig. 4). A similar pattern was present under reducing,nonreducing, and mild detergent (digitonin) conditions (data notshown). The material that was precipitated with mAb RR5-89 wasdigested with V8 protease, and SDS-PAGE analysis of the diges-tion products revealed a similar peptide pattern for each band (datanot shown). This suggests that the complex immunoprecipitationpattern is composed of a single polypeptide. The glycosylation ofChT1 was studied by removingN- andO-linked carbohydrates.
After treatment withN-glycanase, the core protein of the majorband measured about 46 kDa. Additional treatments with neur-aminidase andO-glycanase did not change the migration of theprotein (data not shown).
Analysis of the mRNA expression of ChT1
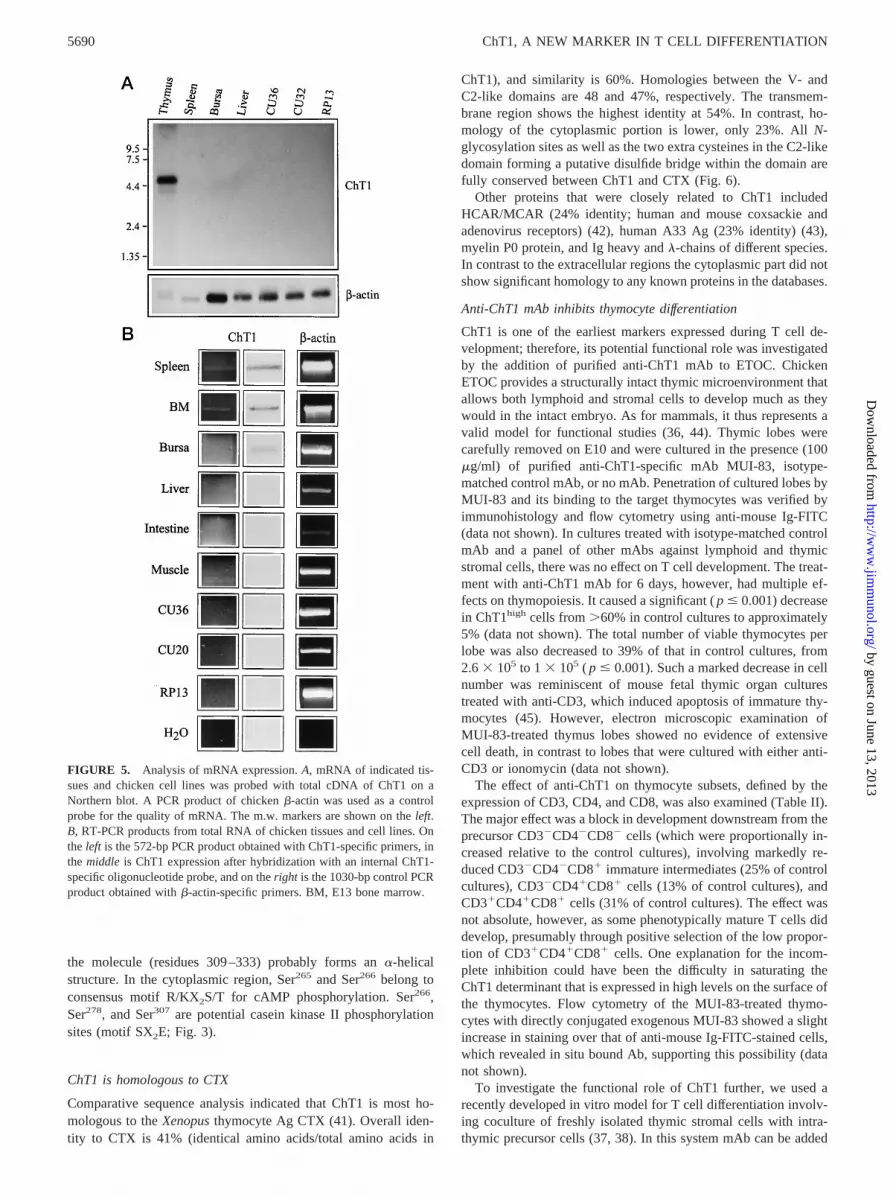
Northern blot analysis with a probe containing the total cDNA ofChT1 revealed two mRNA species, one major band of 4.7 kb anda fainter band of 4.0 kb from the thymus, but no ChT1-specificmRNA was detected in any other organ or cell line tested (Fig. 5Aand data not shown).
Using a more sensitive RT-PCR method we were also able toshow ChT1 mRNA in 4-wk-old chick spleen and E13 bone mar-row (Fig. 5B). After hybridization of the PCR products with aChT1-specific oligonucleotide probe, bursa, liver, and small intes-tine also showed a very low amount of ChT1 mRNA, most likelyindicating the presence of a few ChT11 T-cells in these tissues(Fig. 5B).
ChT1 is an IgSF member consisting of two Ig domains
Structural analysis of the deduced ChT1 protein sequence identi-fies the molecule as a member of the IgSF consisting of two Igdomains: an N-terminal V-like domain and a membrane-proximalC2-like domain. Both represent typical Ig domains, as all cysteinesforming intradomain disulfide bridges are conserved in the B andF b strands, and the tryptophans were conserved at positions 58and 173 in the C strands. The cysteines at positions 144 and 221are located in the A and G strands in the C2-like domain, so thatduring protein tertiary structure formation they may come closeenough to form an additional intradomain disulfide bridge. At theend of the V-like domain the sequence AGQSQKSVIVNVLV re-sembles a J-like segment characterized by features of a diglycinebulge (39, 40) (Fig. 3). However, the J-like segment in ChT1 ismodified so that the second glycine is replaced by a serine. Theextracellular part of ChT1 contains four possibleN-glycosylationsites at aa positions 38, 97, 199, and 218 (Fig. 6). The putativehydrophobic transmembrane region is followed by a long cyto-plasmic tail rich in glutamic acids. The carboxyl-terminal end of
FIGURE 3. The cDNA sequence of ChT1. The figure shows the cDNAsequence of clone p10.6. Numbers on theleft correspond to the position ofthe first nucleotide, and the numbers onright correspond to the position ofthe last amino acid in each line. The beginning and end of Ig domains areindicated by, and ., respectively. The cysteines and tryptophans thatcharacterize the Ig domains are in bold. The transmembrane region is un-derlined. The difference in clone pc210 is marked in the transmembraneregion.
FIGURE 4. Immunoprecipitation of ChT1. ChT1 was immunoprecipi-tated with mAb RR5-89 from the lysate of surface iodinated thymocytes.The precipitated proteins were run on 5–15% SDS-PAGE, which was ex-posed to an autoradiography film. The m.w. markers are shown on theleft.
5689The Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from
the molecule (residues 309–333) probably forms ana-helicalstructure. In the cytoplasmic region, Ser265 and Ser266 belong toconsensus motif R/KX2S/T for cAMP phosphorylation. Ser266,Ser278, and Ser307 are potential casein kinase II phosphorylationsites (motif SX2E; Fig. 3).
ChT1 is homologous to CTX
Comparative sequence analysis indicated that ChT1 is most ho-mologous to theXenopusthymocyte Ag CTX (41). Overall iden-tity to CTX is 41% (identical amino acids/total amino acids in
ChT1), and similarity is 60%. Homologies between the V- andC2-like domains are 48 and 47%, respectively. The transmem-brane region shows the highest identity at 54%. In contrast, ho-mology of the cytoplasmic portion is lower, only 23%. AllN-glycosylation sites as well as the two extra cysteines in the C2-likedomain forming a putative disulfide bridge within the domain arefully conserved between ChT1 and CTX (Fig. 6).
Other proteins that were closely related to ChT1 includedHCAR/MCAR (24% identity; human and mouse coxsackie andadenovirus receptors) (42), human A33 Ag (23% identity) (43),myelin P0 protein, and Ig heavy andl-chains of different species.In contrast to the extracellular regions the cytoplasmic part did notshow significant homology to any known proteins in the databases.
Anti-ChT1 mAb inhibits thymocyte differentiation
ChT1 is one of the earliest markers expressed during T cell de-velopment; therefore, its potential functional role was investigatedby the addition of purified anti-ChT1 mAb to ETOC. ChickenETOC provides a structurally intact thymic microenvironment thatallows both lymphoid and stromal cells to develop much as theywould in the intact embryo. As for mammals, it thus represents avalid model for functional studies (36, 44). Thymic lobes werecarefully removed on E10 and were cultured in the presence (100mg/ml) of purified anti-ChT1-specific mAb MUI-83, isotype-matched control mAb, or no mAb. Penetration of cultured lobes byMUI-83 and its binding to the target thymocytes was verified byimmunohistology and flow cytometry using anti-mouse Ig-FITC(data not shown). In cultures treated with isotype-matched controlmAb and a panel of other mAbs against lymphoid and thymicstromal cells, there was no effect on T cell development. The treat-ment with anti-ChT1 mAb for 6 days, however, had multiple ef-fects on thymopoiesis. It caused a significant (p # 0.001) decreasein ChT1high cells from.60% in control cultures to approximately5% (data not shown). The total number of viable thymocytes perlobe was also decreased to 39% of that in control cultures, from2.63 105 to 1 3 105 ( p # 0.001). Such a marked decrease in cellnumber was reminiscent of mouse fetal thymic organ culturestreated with anti-CD3, which induced apoptosis of immature thy-mocytes (45). However, electron microscopic examination ofMUI-83-treated thymus lobes showed no evidence of extensivecell death, in contrast to lobes that were cultured with either anti-CD3 or ionomycin (data not shown).
The effect of anti-ChT1 on thymocyte subsets, defined by theexpression of CD3, CD4, and CD8, was also examined (Table II).The major effect was a block in development downstream from theprecursor CD32CD42CD82 cells (which were proportionally in-creased relative to the control cultures), involving markedly re-duced CD32CD42CD81 immature intermediates (25% of controlcultures), CD32CD41CD81 cells (13% of control cultures), andCD31CD41CD81 cells (31% of control cultures). The effect wasnot absolute, however, as some phenotypically mature T cells diddevelop, presumably through positive selection of the low propor-tion of CD31CD41CD81 cells. One explanation for the incom-plete inhibition could have been the difficulty in saturating theChT1 determinant that is expressed in high levels on the surface ofthe thymocytes. Flow cytometry of the MUI-83-treated thymo-cytes with directly conjugated exogenous MUI-83 showed a slightincrease in staining over that of anti-mouse Ig-FITC-stained cells,which revealed in situ bound Ab, supporting this possibility (datanot shown).
To investigate the functional role of ChT1 further, we used arecently developed in vitro model for T cell differentiation involv-ing coculture of freshly isolated thymic stromal cells with intra-thymic precursor cells (37, 38). In this system mAb can be added
FIGURE 5. Analysis of mRNA expression.A, mRNA of indicated tis-sues and chicken cell lines was probed with total cDNA of ChT1 on aNorthern blot. A PCR product of chickenb-actin was used as a controlprobe for the quality of mRNA. The m.w. markers are shown on theleft.B, RT-PCR products from total RNA of chicken tissues and cell lines. Onthe left is the 572-bp PCR product obtained with ChT1-specific primers, inthe middle is ChT1 expression after hybridization with an internal ChT1-specific oligonucleotide probe, and on theright is the 1030-bp control PCRproduct obtained withb-actin-specific primers. BM, E13 bone marrow.
5690 ChT1, A NEW MARKER IN T CELL DIFFERENTIATION
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

directly to the precursor cells from the very onset of coculture,saturating the determinants before contact with the stroma andhence differentiation induction. Sorted CD42CD82 E13 thymo-cytes from ov-alloantigen-positive embryos and isolated adult thy-mic stromal cells (ov2) were cocultured for 4 days in the presenceof anti-ChT1, anti-c-kit, no mAb, or a variety of control mAb tothymic stromal or lymphoid molecules. After 4 days the cultureswere harvested, the total cell yield was counted, and the expressionof CD4 and CD8 was analyzed on ov1 thymocytes by flow cy-tometry. Treatment with anti-ChT1 was comparable to that withanti-c-kit, so that in both cases the frequency of CD41CD81 cellswas decreased from 33.7% in control cultures to approximately 3.5and 3.1%, respectively (Fig. 7). A substantial proportion (38.4 and54.8%, respectively) of double-negative cells partially differenti-ated to CD42CD81, presumably immature thymocytes in the pres-ence of both anti-ChT1 and anti-c-kit mAb, but no CD41CD82
cells were detectable. The total cell recovery slightly increased incocultures with either anti-ChT1 or anti-c-kit as a result of mostcells staying at an undifferentiated CD42CD82 or CD42CD81
stage (Table III). There was no alteration numerically or propor-tionally in the control cocultures. These in vitro data emphasize thefunctional role of ChT1 during the proximal stages of thymocytedifferentiation.
DiscussionSearching for molecules involved in early T cell differentiation, weisolated a cDNA encoding the avian thymocyte Ag ChT1, a mem-ber of the IgSF with two extracellular domains, V and C2. Theexpression of ChT1 on peripheral T cells defines recent thymicemigrants (20). We show here that the ChT11c-kit1 subpopulationof embryonic bone marrow cells is enriched for T cell progenitors,as assayed by intrathymic cell transfer to congenic animals. Anti-ChT1 mAb also blocks T cell differentiation in vitro in thymicorgan cultures and thymocyte precursor cultures on stromal cells.These results demonstrate clearly that ChT1 plays an importantrole in the early stages of T cell development.
FIGURE 6. The alignment of ChT1 and CTXamino acid sequences. Identical amino acids areboxed in black, and the cysteines forming the do-main-specific disulfide bridges are marked withgray boxes. ConservedN-glycosylation sites aremarked with an asterisk. The leader peptide ismarked with a dashed line, and the transmembraneregion is indicated with a solid line. The aminoacids are aligned according to the Clustal method.
Table II. Effect of anti-ChT1 treatment on ETOCa
ThymocytePopulation
Isotype Control ETOCAnti-ChT1-Treated ETOC,
100 mg/ml
Anti-ChT1-Treated Cells/ControlCells (%)
Positive cells(%)
Cell numberb
(3104)Positive cells
(%)Cell numberb
(3104)
CD32CD42CD82 10.46 2.7 1.56 0.6 19.36 5.9‡ 1.06 0.5 66CD32CD41CD82 1.46 1.3 0.26 0.2 1.56 0.3 0.16 0.02 50CD32CD42CD81 20.26 5.0 3.16 1.8 14.56 1.6* 0.86 0.3* 25CD32CD41CD81 19.06 6.2 2.96 1.4 7.96 1.8‡ 0.46 0.2§ 13CD31CD41CD81 14.16 6.0 2.26 1.3 12.46 3.9 0.76 0.4* 31CD31CD41CD82 1.26 0.7 0.26 0.1 1.26 0.2 0.16 0.03 50CD31CD42CD81 15.56 4.3 2.36 1.0 23.66 5.8* 1.36 0.5 56CD31CD42CD82 23.26 4.9 3.36 0.8 19.76 5.9 1.16 0.5§ 33
a E10 thymus lobes were isolated and cultured for 6 days in the presence of anti-ChT1 mAb or an isotype-matched control mAb 10-2.16 (anti-mouse I-Ak). After culture thelymphocytes were isolated, and recovered cells were counted, stained, and analyzed for the expression of CD3, CD4, and CD8 by flow cytometry. Results are expressed as mean6SD of six experiments. Unpaired Student’st test was used to test for significance: *p , 0.05,‡ p , 0.01,§ p , 0.001.
b All subsets were defined by discrete nonoverlapping gates; cells outside of these gates, including CD4lowCD8low apoptosing cells (present to some degree in all cultures),were excluded.
5691The Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

The earliest intrathymic T cell precursors in the mouse havebeen defined as CD32CD42CD82CD1171CD441CD252 cells inwhich the TCR loci are still in germline configuration (8, 46). Inaddition, several other markers, e.g., Sca-1/2, HSA, Thy-1, andIL-7R, have been used to characterize early T cell precursors eitherin the thymus or fetal bone marrow (1, 3, 47, 48). However, noneof these markers is T-lineage specific. Since ChT1 is the earliestT-lineage marker to be defined to date, being present on thymo-cytes on E10 of embryogenesis, well before CD4, CD8, and theCD3/TCR complex, and since its expression is developmentallyregulated, we explored the functional role of ChT1 in embryonicthymocyte differentiation. In ETOC the treatment with anti-ChT1-specific mAb resulted in a significant decrease in total cell yield.The Ab treatment most significantly perturbed the differentiationstep from the CD42CD82 to the CD41CD81 stage. The immatureCD32 thymocyte subpopulations were affected to a much largerextent than the CD31 populations. ChT1 mAb also inhibited T celldifferentiation in in vitro coculture of precursors with thymic stro-mal cells. Thus, these in vitro assays convincingly show that ChT1has an important function in the early stages of T cell differenti-ation. A similar effect on T cell development has been reported inreaggregate fetal thymic organ cultures with an anti-E-cadherin-specific mAb that blocks homotypic E-cadherin interactions be-tween T cell precursors and thymic stromal cells (49).
ChT1 was found to be a close homologue to a recently clonedXenopusthymocyte Ag, CTX (41, 50). The structures of these twomolecules are highly conserved, as both are composed of two Igdomains with fully conserved putativeN-linked glycosylationsites. In addition to the conserved features of their Ig domain struc-tures, ChT1 and CTX share other interesting characteristics. First,there is a J-like segment with a diglycine bulge at the end of the Vdomain (39). Most molecules with a J-like segment bind their li-
gands as dimers and are thought to serve as receptors (39, 40).Although in ChT1 the second glycine of the diglycine bulge isreplaced by a serine residue, similar modification has been ob-served in other molecules, such as human CD28 (51), mouse B cellspecific glycoprotein B29 (52), andXenopuss locus light chain(40, 53), without affecting the dimerization function of the motif.Even though CTX does not form dimers spontaneously, it can beinduced to dimerize by cross-linking (54). Indeed, earlier reportsof ChT1 or of an Ag that most likely is ChT1 have proposed thatChT1 is able to form spontaneous homodimers (18, 55), and ourresults in this study support this idea. Because the migration of allbands was increased equally afterN-deglycosylation, we suggestthat the bands are composed of similar proteins and carbohydrates.However, as we were not able to dissociate the proposed dimers byreduction there are other possibilities, in addition to dimerization,to interpret the immunoprecipitation pattern obtained. It may bethat the subunits of the protein complex are linked by forces thatare not breakable by the reducing agent used. Cross-reactivity ofanti-ChT1 mAb with an unrelated protein is still another possibil-ity to produce this result, but since we have used many differentanti-ChT1 mAb with identical results this seems to be an unlikelysolution. In addition to diglycine bulge, the ChT1 sequence con-tains an extra cysteine residue, approximately in the B/C loop ofthe C2-like domain, which could mediate dimerization by formingan intermolecular disulfide bridge. Thus, there is a possibility thatChT1 functions as a lymphocyte-specific receptor during T celldifferentiation.
The second common feature with CTX is that the C2-like do-main of the ChT1 harbors a conserved extra disulfide bridge,which has been found in the same position in other molecules thatare obviously ChT1/CTX relatives, e.g., A33 (43) and HCAR/MCAR (42). Third, the intracellular tail is highly polar, containing
FIGURE 7. T cell differentiation in in vitro thymocyte precursor and stromal cell coculture. E13 thymocyte precursors (sorted CD42CD82 populationfrom the ov1 congenic strain) were cultured for 4 days with adult thymus stromal cells (from the ov2 strain). Anti-ChT1 mAb MUI-83 and anti-c-kit mAbkit2c75 were added to the cultures. After culture the number of ov1 thymocytes was counted, then cells were stained with anti-CD4 and anti-CD8 mAbto analyze T cell differentiation during culture. The percentage of positive cells in each quadrant is shown in theupper corner. The effect of anti-ChT1 mAbis compared with T cell differentiation when no Ab or anti-c-kit mAb was added to the culture. Results shown are representative of three separateexperiments.
Table III. In vitro coculture of embryonic CD42CD82 ov1 thymocytes on ov2 stromal cellsa
mAb (100mg/ml)
Number (3104) of Recovered ov1 Cells per 106 Input Day 13 Embryonic ov1 Thymocytes
Total cell recovery CD42CD82 CD41CD81 CD42CD81 CD41CD82
MUI-83 (anti-ChT1) 17.3 10.0 0.6 6.6 0kit2c75 (anti-c-kit) 22.4 9.3 0.7 12.2 0MUI-82 (antithymic stroma) 19.8 3.9 8.5 5.0 2.2— 15.0 4.8 5.1 3.6 1.5
a E13 ov1 thymocytes were sorted for CD42CD82 cells and prepared at a concentration of 3.43 106 cells/ml. Adult ov2 stromal cells were prepared by enzymatic digestionof lymphocyte-depleted thymi. Stromal cells were enriched by elutriation and resuspended at a concentration of 6.73 105 cells/ml. Two cell populations were mixed at a ratioof 5:1 and cocultured in the presence of anti-ChT1, anti-c-kit, control Ab or no mAb as hanging drops in inverted Terasaki plates at 40°C, 10% CO2 for 4 days. Results for MUI-82are representative of 50 control Abs reactive to thymic stromal cells and lymphoid markers which included isotype-specific controls.
5692 ChT1, A NEW MARKER IN T CELL DIFFERENTIATION
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

several glutamic acid residues. The carboxyl-terminal end of bothChT1 and CTX can be predicted to form ana-helical structurewhere the negatively charged amino acids would gather close toone side of the helix and thus possibly interact with other polarintracellular molecules. A similar cytoplasmic tail, rich in glutamicacids, has been found in A33 and HCAR/MCAR. However, eventhough ChT1 and CTX are clearly homologues, their cytoplasmicregions are quite different. There are several potential serine phos-phorylation sites in the ChT1 sequence, none of which is presentin CTX. Thus, in addition to a possible extracellular receptor func-tion, ChT1 may function in signal transduction. Determiningwhether ChT1, CTX, and other products of the ChT1/CTX familyof genes mediate similar functions awaits additional experimentsin different species.
In addition, to being a T-lineage marker in embryonic thymus,ChT1 expression together with c-kit expression define a prethymicT cell precursor population in embryonic bone marrow. Afteradoptive intrathymic cell transfer, the ChT11c-kit1 cell popula-tion gave rise togd as well as CD41 and CD81 matureab T cells.Recently, the existence of a common lymphoid progenitor popu-lation, Lin2IL-7R1Thy-12Sca-1lowc-kitlow, has been described inthe mouse (9). Whether the ChT11c-kit1 bone marrow subpopu-lation contains T-lineage-restricted, common lymphoid, or multi-potent hemopoietic progenitors remains to be clarified. In vivoChT1 interaction with its putative ligand might result in a main-tenance or viability signal that would allow thymic selectionevents to occur. Interestingly, CTX has been shown to mediate acellular growth inhibition signal inXenopusT cell tumors in agree-ment with the idea that ChT1/CTX is involved in the regulation ofcell proliferation (54). According to our data from ETOC and pre-cursor coculture with thymic stromal cells, we suggest that ChT1is required for thymocyte precursors to develop to the CD41CD81
stage; again, this could operate at the level of cell proliferation. Itmay be that when the cells have matured to the CD31CD41CD81
stage, ChT1 would no longer be crucial for T cell differentiation.However, ChT1 expression would continue for some time after thecell has emigrated from the thymus, thus marking the recent thy-mic emigrants (20). We hypothesize that the signal received fromChT1 Ag interaction with its ligand is necessary for developingthymocytes to proceed to the stage of positive (and negative) se-lection. The results presented in this study demonstrate that ChT1Ag is an early T-lineage-specific marker on embryonic thymus andbone marrow cells and emphasize its importance in the first phasesof T cell development.
AcknowledgmentsWe thank John Young for the RPRL line 0 thymus cDNA library, RiittaKoskinen for her help with cDNA cloning, Natalie Davidson for ETOCsubset analysis, Tatsuya Uchida for E13 bone marrow cDNA, ElisabethHoussaint for the mAb T10A6, and Max D. Cooper and Chen-lo H. Chenfor mAb CT1 and CT1a. Ann Sofie Hakulinen, Raija Raulimo, Jean-PierreDangy, and Barbara Ecabert provided expert technical assistance, andDavid Avila performed protein sequencing. We also thank Louis Du Pas-quier, Isabelle Chretien, Olli Lassila, and Jan Salomonsen for their criticalreading and improvement of the manuscript.
References1. Shortman, K., and L. Wu. 1996. Early T lymphocyte progenitors.Annu. Rev.
Immunol. 14:29.2. Kisielow, P., and H. von Boehmer. 1995. Development and selection of T cells:
facts and puzzles.Adv. Immunol. 58:87.3. Zuniga-Pflucker, J. C., and M. J. Lenardo. 1996. Regulation of thymocyte de-
velopment from immature progenitors.Curr. Opin. Immunol. 8:215.4. Matsuzaki, Y., J. Gyotoku, M. Ogawa, S. Nishikawa, Y. Katsura, G. Gachelin,
and H. Nakauchi. 1993. Characterization of c-kit positive intrathymic stem cellsthat are restricted to lymphoid differentiation.J. Exp. Med. 178:1283.
5. Galy, A., M. Travis, D. Cen, and B. Chen. 1995. Human T, B, natural killer, anddendritic cells arise from a common bone marrow progenitor cell subset.Immu-nity 3:459.
6. Levelt, C. N., and K. Eichmann. 1995. Receptors and signals in early thymicselection.Immunity 3:667.
7. Fehling, H. J., and H. von Boehmer. 1997. Earlyab T cell development in thethymus of normal and genetically altered mice.Curr. Opin. Immunol. 9:263.
8. Godfrey, D. I., and A. Zlotnik. 1993. Control points in early T-cell development.Immunol. Today 14:547.
9. Kondo, M., I. L. Weissman, and K. Akashi. 1997. Identification of clonogeniccommon lymphoid progenitors in mouse bone marrow.Cell 91:661.
10. Wu, L., R. Scollay, M. Egerton, M. Pearse, G. J. Spangrude, and K. Shortman.1991. CD4 expressed on earliest T-lineage precursor cells in the adult murinethymus.Nature 349:71.
11. Jotereau, F. V., and N. M. Le Douarin. 1982. Demonstration of a cyclic renewalof the lymphocyte precursor cells in the quail thymus during embryonic andperinatal life.J. Immunol. 129:1869.
12. Le Douarin, N. M., F. V. Jotereau, E. Houssaint, and J. P. Thiery. 1984. Primarylymphoid organ ontogeny in birds. InChimeras in Developmental Biology.N. M. Le Douarin and A. McLaren, eds. Academic Press, London, p. 179.
13. Coltey, M., F. V. Jotereau, and N. M. Le Douarin. 1987. Evidence for cyclicrenewal of lymphocyte precursor cells in the embryonic chick thymus.Cell Dif-fer. 22:71.
14. Dunon, D., D. Courtois, O. Vainio, A. Six, C. H. Chen, M. D. Cooper, J.-P.Dangy, and B. A. Imhof. 1997. Ontogeny of the immune system:gd andab Tcells migrate from thymus to the periphery in alternating waves.J. Exp. Med.186:977.
15. Dunon, D., J. Kaufman, J. Salomonsen, K. Skjoedt, O. Vainio, J. P. Thiery, andB. A. Imhof. 1990. T cell precursor migration towardsb2-microglobulin is in-volved in thymus colonization of chicken embryos.EMBO J. 9:3315.
16. Vainio, O., D. Dunon, F. Aissi, J.-P. Dangy, K. M. McNagny, and B. A. Imhof.1996. HEMCAM, an adhesion molecule expressed by c-kit1 hemopoietic pro-genitors.J. Cell Biol. 135:1655.
17. Corbel, C., O. Pourquie, F. Cormier, P. Vaigot, and N. M. Le Douarin. 1996.BEN/SC1/DM-GRASP, a homophilic adhesion molecule, is required for in vitromyeloid colony formation by avian hemopoietic progenitors.Proc. Natl. Acad.Sci. USA 93:2844.
18. Chen, C.-l. H., T. C. Chanh, and M. D. Cooper. 1984. Chicken thymocyte-spe-cific antigen identified by monoclonal antibodies: ontogeny, tissue distributionand biochemical characterization.Eur. J. Immunol. 14:385.
19. Houssaint, E., E. Dietz, and F. V. Jotereau. 1985. Tissue distribution and onto-genic appearance of a chicken T lymphocyte differentiation marker.Eur. J. Im-munol. 15:305.
20. Kong, F.-k., C.-l. H. Chen, and M. D. Cooper. 1998. Thymic function can beaccurately monitored by the level of recent T cell emigrants in the circulation.Immunity 8:97.
21. Houssaint, E., A. Mansikka, and O. Vainio. 1991. Early separation of B and Tlymphocyte precursors in chick embryo.J. Exp. Med. 174:397.
22. Boyd, R. L., T. J. Wilson, A. G. Bean, H. A. Ward, and M. E. Gershwin. 1992.Phenotypic characterization of chicken thymic stromal elements.Dev. Immunol.2:51.
23. Luhtala, M., J. Salomonsen, Y. Hirota, T. Onodera, P. Toivanen, and O. Vainio.1993. Analysis of chicken CD4 by monoclonal antibodies indicates evolutionaryconservation between avian and mammalian species.Hybridoma 12:633.
24. Luhtala, M., R. Koskinen, P. Toivanen, and O. Vainio. 1995. Characterization ofchicken CD8-specific monoclonal antibodies recognizing novel epitopes.Scand.J. Immunol. 42:171.
25. Wilson, T. J., and R. L. Boyd. 1991. The ontogeny of chicken bursal stromal cellsdefined by monoclonal antibodies.Dev. Immunol. 1:31.
26. Oi, V. T., P. P. Jones, J. W. Goding, L. A. Herzenberg, and L. A. Herzenberg.1978. Properties of monoclonal antibodies to mouse Ig allotypes, H-2 and Iaantigens.Curr. Top. Microbiol. Immunol. 81:115.
27. Katevuo, K., and O. Vainio. 1996. Thymocyte emigration in the chicken: anover-representation of CD41 cells over CD81 in the periphery.Immunology89:419.
28. Dunon, D., O. Vainio, and B. A. Imhof. 1997. Lymphocyte migration in vivo: thechicken embryo model. InImmunology Methods Manual,Vol. 3. I. Lefkovits, ed.Academic Press, London, p. 1345.
29. Jurgens, J. B., L. A. Gartland, L. Du Pasquier, J. D. Horton, T. W. F. Gobel, andM. D. Cooper. 1995. Identification of a candidate CD5 homologue in the am-phibianXenopus laevis.J. Immunol. 155:4218.
30. Tamura, G. S., M. O. Dailey, W. M. Gallatin, M. S. McGrath, I. L. Weissman,and E. A. Pillemer. 1984. Isolation of molecules recognized by monoclonal an-tibodies and antisera: the solid phase immunoisolation technique.Anal. Biochem.136:458.
31. Cleveland, D. W., S. G. Fischer, M. W. Kirschner, and U. K. Laemmli. 1977.Peptide mapping by limited proteolysis in sodium dodecyl sulfate and analysis bygel electrophoresis.J. Biol. Chem. 252:1102.
32. Schneider, C., R. A. Newman, D. R. Sutherland, U. Asser, and M. F. Greaves.1982. A one-step purification of membrane proteins using a high efficiency im-munomatrix.J. Biol. Chem. 257:10766.
33. Tregaskes, C. A., and J. R. Young. 1997. Cloning of chicken lymphocyte markercDNAs from eukaryotic expression libraries. InImmunology Methods Manual,Vol. 4. I. Lefkovits, ed. Academic Press, London, p. 2295.
34. Liippo, J., and O. Lassila. 1997. Avian Ikaros gene is expressed early in embry-ogenesis.Eur. J. Immunol. 27:1853.
5693The Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

35. Dunon, D., J. Schwager, J. P. Dangy, and B. A. Imhof. 1995. Ontogeny of TCRVb1 expression revealed novel invariant alternative transcripts.J. Immunol. 154:1256.
36. Davidson, N. J., C.-l. H. Chen, and R. L. Boyd. 1992. Kinetics of chicken em-bryonic thymocyte developmentin ovo and in organ culture.Eur. J. Immunol.22:1429.
37. Lahoud, M., D. Vremec, R. L. Boyd, and K. Shortman. 1993. Characterization ofthymic nurse-cell lymphocytes, using an improved procedure for nurse-cell iso-lation. Dev. Immunol. 3:103.
38. Chidgey, A. P., H. Pircher, H. R. MacDonald, and R. Boyd. 1998. An adultthymic stromal cell suspension model for in vitro positive selection.Dev. Immu-nol. 6:157.
39. Carayannopoulos, L., and J. D. Capra. 1993. Immunoglobulins, structure andfunction. InFundamental Immunology. W. E. Paul, ed. Raven Press, New York,p. 283.
40. Du Pasquier, L., and I. Chretien. 1996. CTX, a new lymphocyte receptor inXenopusand the early evolution of Ig domains.Res. Immunol. 147:218.
41. Chretien, I., J. Robert, A. Marcuz, J. A. Garcia-Sanz, M. Courtet, andL. Du Pasquier. 1996. CTX, a novel molecule specifically expressed on the sur-face of cortical thymocytes inXenopus. Eur. J. Immunol. 26:780.
42. Tomko, R. P., R. Xu, and L. Philipson. 1997. HCAR and MCAR: the human andmouse cellular receptors for subgroup C adenoviruses and group B coxsackievi-ruses.Proc. Natl. Acad. Sci. USA 94:3352.
43. Heath, J. K., S. J. White, C. N. Johnstone, B. Catimel, R. J. Simpson,R. L. Moritz, G.-F. Tu, H. Ji, R. H. Whitehead, L. C. Groenen, et al. 1997. Thehuman A33 antigen is a transmembrane glycoprotein and a novel member of theimmunoglobulin superfamily.Proc. Natl. Acad. Sci. USA 94:469.
44. Davidson, N. J., A. G. D. Bean, and R. L. Boyd. 1991. Chicken fetal thymusorgan culture: a model for T cell development within the thymic microenviron-ment. In Lymphatic Tissues and In Vivo Immune Responses. S. Ezine,S. Berrih-Aknin, and B. A. Imhof, eds. Marcel Dekker, New York, p. 89.
45. Smith, C. A., G. T. Williams, R. Kingston, E. J. Jenkinson, and J. J. T. Owen.1989. Antibodies to CD3/T-cell receptor complex induce death by apoptosis inimmature T cells in thymic cultures.Nature 337:181.
46. Petrie, H. T., F. Livak, D. Burtrum, and S. Mazel. 1995. T cell receptor generecombination patterns and mechanisms: cell death, rescue, and T cell produc-tion. J. Exp. Med. 182:121.
47. Hozumi, K., M. Kondo, H. Nozaki, A. Kobori, T. Nishimura, S. Nishikawa,K. Sugamura, and S. Habu. 1994. Implication of the commong chain of the IL-7receptor in intrathymic development of pro-T cells.Int. Immunol. 6:1451.
48. Peschon, J. J., P. J. Morrissey, K. H. Grabstein, F. J. Ramsdell, E. Maraskovsky,B. C. Gliniak, L. S. Park, S. F. Ziegler, D. E. Williams, C. B. Ware, et al. 1994.Early lymphocyte expansion is severely impaired in interleukin 7 receptor-defi-cient mice.J. Exp. Med. 180:1955.
49. Muller, K. M., C. J. Luedecker, M. C. Udey, and A. G. Farr. 1997. Involvementof E-cadherin in thymus organogenesis and thymocyte maturation.Immunity6:257.
50. Chretien, I., A. Marcuz, M. Courtet, K. Katevuo, O. Vainio, J. K. Heath,S. J. White, and L. Du Pasquier. 1998. CTX, aXenopusthymocyte receptor,defines a molecular family conserved throughout vertebrates.Eur. J. Immunol.28:4094.
51. Aruffo, A., and B. Seed. 1987. Molecular cloning of a CD28 cDNA by a high-efficiency COS cell expression system.Proc. Natl. Acad. Sci. USA 84:8573.
52. Hermanson, G. G., D. Eisenberg, P. W. Kincade, and R. Wall. 1988. B29: amember of the immunoglobulin gene superfamily exclusively expressed on B-lineage cells.Proc. Natl. Acad. Sci. USA 85:6890.
53. Schwager, J., N. Burckert, M. Schwager, and M. Wilson. 1991. Evolution ofimmunoglobulin light chain genes: analysis ofXenopusIgL isotypes and theircontribution to antibody diversity.EMBO J. 10:505.
54. Robert, J., I. Chretien, C. Guiet, and L. Du Pasquier. 1997. Cross-linking CTX,a novel thymocyte-specific molecule, inhibits the growth of lymphoid tumor cellsin Xenopus. Mol. Immunol. 34:133.
55. Pink, J. R., B. Fedecka Bruner, M. Coltey, B. M. Peault, and N. M. Le Douarin.1981. Biochemical characterization of avian T lymphocyte-specific antigens.Eur.J. Immunol. 11:517.
5694 ChT1, A NEW MARKER IN T CELL DIFFERENTIATION
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from