bioinformatic and enzymatic characterization of the mapeg superfamily
TRANSCRIPT

Bioinformatic and enzymatic characterizationof the MAPEG superfamilyAnders Bresell1,*, Rolf Weinander2,*, Gerd Lundqvist3, Haider Raza3, Miyuki Shimoji3,Tie-Hua Sun3, Lennart Balk5, Ronney Wiklund6, Jan Eriksson6, Christer Jansson6, Bengt Persson1,4,Per-Johan Jakobsson2 and Ralf Morgenstern3
1 IFM Bioinformatics, Linkoping University, Sweden
2 Department of Medicine, Division of Rheumatology Unit, Karolinska Institutet, Stockholm, Sweden
3 Institute of Environmental Medicine Karolinska Institutet, Stockholm, Sweden
4 Centre for Genomics and Bioinformatics, Karolinska Institutet, Stockholm, Sweden
5 Stockholm Marine Research Centre, University of Stockholm, Sweden
6 Department of Plant Biology & Forestry Genetics, Swedish Agricultural University, Uppsala, Sweden
Keywords
MAPEG; microsomal glutathione
transferase; prostaglandin; leukotriene
Correspondence
R. Morgenstern, Institute of Environmental
Medicine, Karolinska Institutet, S-171 77
Stockholm, Sweden
Fax: +46 8 343849
Tel: +46 8 5248 7574
E-mail: [email protected]
*Both authors contributed equally to this
work
(Received 15 November 2004, revised 27
January 2005, accepted 3 February 2005)
doi:10.1111/j.1742-4658.2005.04596.x
The membrane associated proteins in eicosanoid and glutathione metabo-
lism (MAPEG) superfamily includes structurally related membrane proteins
with diverse functions of widespread origin. A total of 136 proteins belong-
ing to the MAPEG superfamily were found in database and genome
screenings. The members were found in prokaryotes and eukaryotes, but
not in any archaeal organism. Multiple sequence alignments and calcula-
tions of evolutionary trees revealed a clear subdivision of the eukaryotic
MAPEG members, corresponding to the six families of microsomal gluta-
thione transferases (MGST) 1, 2 and 3, leukotriene C4 synthase (LTC4),
5-lipoxygenase activating protein (FLAP), and prostaglandin E synthase.
Prokaryotes contain at least two distinct potential ancestral subfamilies, of
which one is unique, whereas the other most closely resembles enzymes that
belong to the MGST2 ⁄FLAP ⁄LTC4 synthase families. The insect members
are most similar to MGST1 ⁄prostaglandin E synthase. With the new data
available, we observe that fish enzymes are present in all six families, show-
ing an early origin for MAPEG family differentiation. Thus, the evolution-
ary origins and relationships of the MAPEG superfamily can be defined,
including distinct sequence patterns characteristic for each of the sub-
families. We have further investigated and functionally characterized repre-
sentative gene products from Escherichia coli, Synechocystis sp., Arabidopsis
thaliana and Drosophila melanogaster, and the fish liver enzyme, purified
from pike (Esox lucius). Protein overexpression and enzyme activity ana-
lysis demonstrated that all proteins catalyzed the conjugation of 1-chloro-
2,4-dinitrobenzene with reduced glutathione. The E. coli protein displayed
glutathione transferase activity of 0.11 lmolÆmin)1Æmg)1 in the membrane
fraction from bacteria overexpressing the protein. Partial purification of
the Synechocystis sp. protein yielded an enzyme of the expected molecular
mass and an N-terminal amino acid sequence that was at least 50%
pure, with a specific activity towards 1-chloro-2,4-dinitrobenzene of
11 lmolÆmin)1Æmg)1. Yeast microsomes expressing the Arabidopsis enzyme
Abbreviations
BSA, bovine serum albumin; CDNB, 1-chloro-2,4-dinitrobenzene; DEAE, diethylaminoethyl; FLAP, 5-lipoxygenase activating protein; LT,
leukotriene; MGST, microsomal glutathione transferase; PG, prostaglandin; PGES, prostaglandin E synthase; GST, glutathione S-transferase;
GPx, glutathione peroxidase; CuOOH, cumene hydroperoxide.
1688 FEBS Journal 272 (2005) 1688–1703 ª 2005 FEBS

Microsomal glutathione transferases (MGSTs) repre-
sent a recently recognized superfamily of enzymes
involved in detoxification, but also in specific biosyn-
thetic pathways of arachidonic acid metabolism. The
superfamily was termed the membrane associated
proteins in eicosanoid and glutathione metabolism
(MAPEG) and consists of proteins from mammals,
plants, fungi and bacteria [1]. The six members in
humans include 5-lipoxygenase activating protein
(FLAP) and leukotriene (LT) C4 synthase, which are
both involved in leukotriene biosynthesis [2,3];
MGST1, MGST2 and MGST3, which all are gluta-
thione transferases as well as glutathione dependent
peroxidases [4–7]; and finally, prostaglandin (PG) E
synthase (PGES), earlier referred to as MGST1-L1 [8].
PGES catalyzes the formation of PGE2 from PGH2,
which in turn is generated from arachidonic acid by
the prostaglandin endoperoxide synthase systems.
PGES has also been referred to as p53 induced gene
12 (PIG12) because the gene expression was found to
increase extensively following p53 expression [9]. The
relationships and other functional aspects of the
MAPEG enzymes have been reviewed [10].
Two groups of bacteria, purple bacteria and cyano-
bacteria, have been found to produce and maintain
significant levels of glutathione [11] and, interestingly,
also contain MAPEG members [1]. Glutathione was
observed in various species within the two groups,
among those in Escherichia coli, one of the most well
characterized species of purple bacteria [11]. The func-
tion of glutathione metabolism in bacteria may be pro-
tection against xenobiotics and ⁄or oxidative stress but
also as part of specific biosynthetic pathways [12].
Cyanobacteria produce oxygen by photosynthesis and
purple bacteria can use oxygen as a terminal electron
acceptor. Glutathione production in bacteria is thus
closely associated with those bacteria that generate or
utilize oxygen in specific biochemical pathways indica-
ting that glutathione metabolism originated in bacteria
at the time when an oxygen-containing atmosphere
developed on earth [11,12].
A low level of glutathione S-transferase (GST) activ-
ity has been demonstrated in E. coli but not in cyano-
bacteria [11]. Cytosolic GSTs have been identified in
various strains of bacteria [12] and in a few studies,
including those on Proteus mirabilis and E. coli, cyto-
solic GSTs have been purified and further character-
ized [13–15]. The three-dimensional structure of the
P. mirabilis cytosolic GST has also been determined
[16]. In Synechocystis sp. a gene homologous to cyto-
solic GST exists but has not been characterized further
[17]. In general, the enzymes involved in glutathione
metabolism in prokaryotes have not been so exten-
sively studied and therefore less is known about their
properties as compared to the corresponding proteins
in eukaryotes. Microsomal GST activity has not been
demonstrated in any prokaryotic organism.
Expressed sequence tag (EST) clones with open
reading frames (ORFs) similar to MAPEG proteins
have been found in E. coli, Synechocystis sp. and
Vibrio cholerae [1]. The Synechocystis sp. ORF dis-
played sequence similarity to the MAPEG subfamily
consisting of FLAP, LTC4 synthase and MGST2, and
also to the MGST3 subfamily but it could not be sig-
nificantly grouped to any of those two subfamilies,
whereas the E. coli and V. cholerae sequences form a
separate group [1]. Nothing is known, however, about
the enzymatic properties of any prokaryotic MAPEG
protein.
As the number of sequenced bacterial genomes has
increased considerably during recent years, we de-
signed this study to search further for MAPEG pro-
teins and functionally characterize representative gene
products. Database searches revealed various new gene
products, in some cases coexisting, with homologies to
the two MAPEG subfamilies (described above and in
[1]). We investigated representative gene products from
the E. coli and Synechocystis sp. bacteria further, to
gain insight into the function of these proteins and the
evolution of the MAPEG superfamily. Cloning and
overexpression demonstrated that both are membrane-
bound glutathione transferases.
showed an activity of 0.02 lmolÆmin)1Æmg)1, whereas the Drosophila
enzyme expressed in E. coli was highly active at 3.6 lmolÆmin)1Æmg)1. The
purified pike enzyme is the most active MGST described so far with a spe-
cific activity of 285 lmolÆmin)1Æmg)1. Drosophila and pike enzymes also
displayed glutathione peroxidase activity towards cumene hydroperoxide
(0.4 and 2.2 lmolÆmin)1Æmg)1, respectively). Glutathione transferase activity
can thus be regarded as a common denominator for a majority of MAPEG
members throughout the kingdoms of life whereas glutathione peroxidase
activity occurs in representatives from the MGST1, 2 and 3 and PGES sub-
families.
A. Bresell et al. Characterization of MAPEG members
FEBS Journal 272 (2005) 1688–1703 ª 2005 FEBS 1689

To understand the evolutionary relationships better
on a more global scale we also cloned and expressed
(or purified) MAPEG representatives from plant,
insect and fish. Together with earlier data on the frog
enzyme [18] these data define glutathione transferase
activity as a central property of MAPEG members
from a wide range of organisms and suggest ancestral
MAPEG members.
Results
MAPEG members from complete genomes
Over 130 MAPEG members were retrieved from
sequence databases and completed genomes, of which
less than half (56) were previously known members
according to the PF01124 entry in Pfam release 11 [19].
Multiple sequence alignments and hydrophobicity plots
were calculated (for a full alignment see supplementary
Fig. 1). Even though several members are distantly
related, all exhibit the typical MAPEG properties of
150 residue subunits with four hydrophobic regions,
compatible with four transmembrane regions [20,21].
Using information from completed genomes, we
have traced the evolutionary relationships of the
MAPEG members. The general relationships are
depicted in Fig. 1. MGST1, PGES and insect forms
have a common branch, compatible with their overlap-
ping substrate-specificities [22]. Likewise, MGST2,
FLAP and LTC4 synthase also show somewhat closer
relationships, indicating properties in common.
MGST3 forms a separate branch. The bacterial E. coli
and Synechocystis variants are found on separate bran-
ches. A detailed dendrogram is shown in Fig. 2.
The bacterial forms show distant relationships and
their exact grouping is not significant at all sites, as
indicated from their low bootstrap values (no asterisks
in Fig. 2). Furthermore, the bacterial forms are present
at three sites in the dendrogram. However, the group-
ing of the families MGST1, MGST2, MGST3, PGES,
FLAP and LTC4 synthase is significant. In a dendro-
gram without the bacterial forms, the grouping of these
families becomes even more evident (not shown).
Among the MAPEG sequences from fish, we find
members from all six branches (MGST1, MGST2,
MGST3, PGES, FLAP and LTC4 synthase), suggest-
ing that the origin of these forms dates back to before
the occurrence of vertebrates, i.e. more than 500 mya.
This dates the differentiation of the MAPEG forms
back to the late Cambrian multiplicity of eukaryotic
species. Notably, in the screenings we have not found
any members from the archaea kingdom, indicating
that the enzymatic activities of the MAPEG family are
not present in these species or that these activities are
catalysed by other enzymes. The absence of MAPEG
members in archaea is certainly consistent with the
lack of GSH in these organisms.
Cloning, expression and characterization
of selected MAPEG members
MGST homologues from Synechocystis and E. coli
After identifying MAPEG members in several bacterial
strains, the E. coli and Synechocystis sp. proteins were
Fig. 1. Schematic evolutionary tree of the MAPEG superfamily. The
evolutionary tree shows the relationships between the six MAPEG
families and three further groups (Insect, E.coliMGST cluster and
SynMGST cluster). A major subgrouping is visible with MGST1,
PGES and Insect in the upper part of the tree and the remaining
families ⁄ groups in the lower part. In the lower part, MGST2, FLAP
and LTC4 synthase have a close relationship, as judged by the
short branches between these enzymes.
Fig. 2. Detailed dendrogram of the MAPEG superfamily. The tree shows all presently known MAPEG forms, excluding species variants
which differ at only a single position. In the tree, the six families are clearly distinguished. The prokaryotic forms are found at three sites –
the E. coli cluster, the Synechocystis cluster, and the group of remaining forms, denoted Bacteria. Two further groups are marked, denoted
Insects and Waterliving. The branch lengths are proportional to the number of residue differences, with the scale bar indicating a 5% amino
acid difference. The fish forms, having representatives for all six MAPEG families, are marked with a fish symbol. Accession numbers refer
to the databases Uniprot, NCBI or ENSEMBL.
Characterization of MAPEG members A. Bresell et al.
1690 FEBS Journal 272 (2005) 1688–1703 ª 2005 FEBS

A. Bresell et al. Characterization of MAPEG members
FEBS Journal 272 (2005) 1688–1703 ª 2005 FEBS 1691

selected for functional characterization of bacterial
MGST homologues. These homologues represent two
different groups of prokaryotic MAPEG members found.
The E. coli ORF, which we refer to as E.coliMGST,
encodes a 141 amino acid residue polypeptide with a cal-
culated molecular mass of 16.2 kDa. The Synechocystis
sp. ORF (from strain PCC6803 [23]) encodes a 137 resi-
due polypeptide with a predicted molecular mass of
15.4 kDa, which we refer to as SynMGST.
The ORFs encoding E.coliMGST and SynMGST
were amplified by PCR, the products cloned into an
expression vector and the DNA sequences were verified
against the EMBL database entries. Following hetero-
logous expression in E. coli, the membrane fractions
were assayed for enzyme activities. The membrane
fraction from cells overexpressing E.coliMGST cata-
lyzed the conjugation of 1-chloro-2,4-dinitrobenzene
(CDNB) with reduced glutathione with a specific
activity of 0.11 lmolÆmin)1Æmg)1. When a shorter con-
struct beginning from the alternative translation start
site of the E.coliMGST was expressed no activity was
detected. Incubation with N-ethylmaleimide (which
activates mammalian MGST1) did not affect the
activity of E.coliMGST. Membranes from cells over-
expressing the SynMGST also displayed glutathione
transferase activity. The glutathione conjugating activ-
ity towards CDNB was 1.7 lmolÆmin)1Æmg)1 for the
SynMGST membrane fraction. Neither LTC4 synthase
activity, nor any glutathione-dependent peroxidase
activity (towards cumene hydroperoxide or 5-hydrope-
roxy-eicosatetraenoic acid) could be observed in any of
the fractions. No activity could be detected with these
enzymes towards 1,2-epoxy-3-para-nitrophenoxypro-
pane or trans-phenylbut-3-en-2-one as substrates (sum-
marised in Table 1).
Partial purification of SynMGST
To characterize bacterial MGSTs further we concen-
trated on SynMGST. RT-PCR was used to confirm
that SynMGST is indeed expressed in the cyanobac-
teria (Fig. 3).
Having established gene expression of SynMGST in
the cyanobacteria and a functional overexpression of
recombinant protein in E. coli we made an attempt to
purify the protein for further characterization. Bacterial
membranes isolated from cells overexpressing recom-
binant SynMGST were solubilized in Triton X-100.
The recombinant SynMGST was also enzymatically
active upon detergent solubilization and the CDNB
conjugating activity was used to monitor subsequent
purification steps. The SynMGST is basic (the cal-
culated isoelectric point being 9.9) and could therefore
be expected to yield a purified product using meth-
ods developed for MGST1 [24]. However, although
the enzyme behaved in a predictable manner upon
hydroxyapatite batch chromatography, in cation
exchange chromatography the enzyme was recovered in
the flow-through fractions. Diethylaminoethyl (DEAE)
columns, likewise, did not retain the enzyme. Because
cation and anion exchange chromatography, in concert,
did retain most of the contaminating proteins, a parti-
ally purified protein was nevertheless recovered. In fact,
SDS ⁄PAGE (Fig. 4) shows that the protein is nearly
homogeneous. Furthermore, N-terminal amino acid
Table 1. Comparison of glutathione transferase and peroxidase
activity of MAPEG members expressed ⁄ purified from prokaryotes,
plant, nonmammalian and mammalian species. ND, not detectable.
Species
CDNB activity
(lmolÆmin)1Æmg)1)
CuOOH GPx
activity
(lmolÆmin)1Æmg)1)
Activity of purified enzyme
Human MGST1 [71,72] 1.9 0.04
Rat MGST1 [43] 2 0.08
Xenopus laevis, frog [18] 210 2.1
Esox lucius, pike 290 2.2
Synechocystis sp. 11 ND
(partially purified)
Activity in membrane fraction
after heterologous expression
Drosophila melanogaster 3.6 0.4
Arabidopsis thaliana 0.02 n.d.
Synechocystis sp. 1.7 n.d.
Escherichia coli 0.11 n.d.
800
1 2 3 4
400
200100
Fig. 3. RT-PCR. To demonstrate that the SynMGST gene was
expressed in Synechocystis 6803, total RNA was isolated and
amplified by PCR with SynMGST-specific primers, in the presence
(lane 4) or absence (lane 3) of reverse transcriptase. PCR amplifica-
tion from isolated total DNA, using the same primers (lane 2)
served as a positive control. Sizes in bp, deduced from a 100 bp
ladder (lane 1) are indicated.
Characterization of MAPEG members A. Bresell et al.
1692 FEBS Journal 272 (2005) 1688–1703 ª 2005 FEBS

sequence analysis of the predominant band displaying
the correct molecular mass, purified from the gel,
yielded the expected sequence. The partially purified
protein constitutes a major part of the preparation and
therefore the specific activities measured will be close to
those of the pure enzyme.
The enzyme is more active than its mammalian
counterparts and expressed extremely well. Assuming
that the protein was at least 50% pure, the purification
factor (12-fold) indicates that expressed SynMGST
constituted about 8% of the E. coli membrane asso-
ciated proteins. The specific activity of the partially
purified enzyme with 1-chloro-2,4-dinitrobenzene was
11 ± 0.4 lmolÆmin)1Æmg)1 (mean ± SD, n ¼ 3). The
activity was not affected by incubation with the sulf-
hydryl reagent N-ethylmaleimide in contrast to mam-
malian MGST1, which is activated several-fold by this
reagent.
MGST3 from Arabidopsis
When plant MGST3 was cloned and overexpressed
in a yeast expression system, the yeast microsomes
displayed a low glutathione transferase activity with
CDNB (0.02 lmolÆmin)1Æmg)1) that was not activa-
ted ⁄ inhibited by N-ethylmaleimide. Glutathione peroxi-
dase activity was not altered compared to that in
microsomes from yeast expressing the pYeDP60 vector
only (the negative control).
MGST1/PGES-like enzyme from Drosophila
The MGST from Drosophila was cloned and over-
expressed in E. coli where the isolated membrane
fraction displayed a high glutathione transferase acti-
vity (3.6 lmolÆmin)1Æmg)1) and glutathione peroxidase
activity (0.4 lmolÆmin)1Æmg)1). Addition of 1% (v ⁄ v)Triton X-100 to the membrane fraction resulted in a
slight increase in activity, whereas N-ethylmaleimide
had no effect on enzyme activity. The enzyme did not
display PGES activity.
MGST1/PGES-like enzyme from pike
MGST was successfully purified to apparent homogen-
eity (Fig. 4) from pike liver using protocols developed
for the rat enzyme. The N-terminal sequence of the
purified pike enzyme was determined using Edman de-
gradation. Sequence comparisons reveal that the pike
form purified is closely related to the MGST1 ⁄PGES
branch (Fig. 5). Of the N-terminal 47 residues, 22–28
residues are identical to fish MGST1 sequences, while
only 2–12 residues are identical to the fish sequences of
other MAPEG families.
The enzymatic properties of the pike MGST1-like
enzyme were extensively characterised (Table 1) dem-
onstrating that the protein has the highest glutathione
transferase activity of any MAPEG member detected
so far. As the enzyme displays similar substrate speci-
ficity to MGST1, including glutathione peroxidase
activity, the assignment to the MGST1 ⁄PGES sub-
family appears well founded.
Sequence patterns of the MAPEG members
For the MGST1–3, FLAP, LTC4 synthase, PGES and
Insect family clusters we generated sequence patterns,
shown in Table 2. These patterns are all 100%
unambiguous when scanned against Swiss-Prot and
TrEMBL, i.e. no nonmembers are ranked higher than
SynMGST
MGST MGST1RatPike
1mg/lane 1mg/lane
MGST1Rat
75 ng 25 ng kDa
Mrmarkers
kDa
Mrmarkers
45
31
21.5
14.4
10
2015
150 ng
Fig. 4. SDS ⁄ PAGE analysis of purified
SynMGST and pike MGST. The protein was
fractionated on SDS ⁄ PAGE (15%) and
visualized by silver staining. Major proteins
were detected that comigrated with purified
RatMGST1 (17 kDa).
A. Bresell et al. Characterization of MAPEG members
FEBS Journal 272 (2005) 1688–1703 ª 2005 FEBS 1693

the lowest ranked true member. These patterns are
more specific then the existing PROSITE pattern
PS01297 (FLAP ⁄GST2 ⁄LTC4S: G-x(3)-F-E-R-V-[FY]-
x-A-[NQ]-x-N-C) [25]. The patterns are selected based
on conserved regions in the sequence. Notably, the
PGES pattern is located at the beginning of loop one
and for FLAP it is located in the third hydrophobic seg-
ment. All of the remaining patterns are located at the
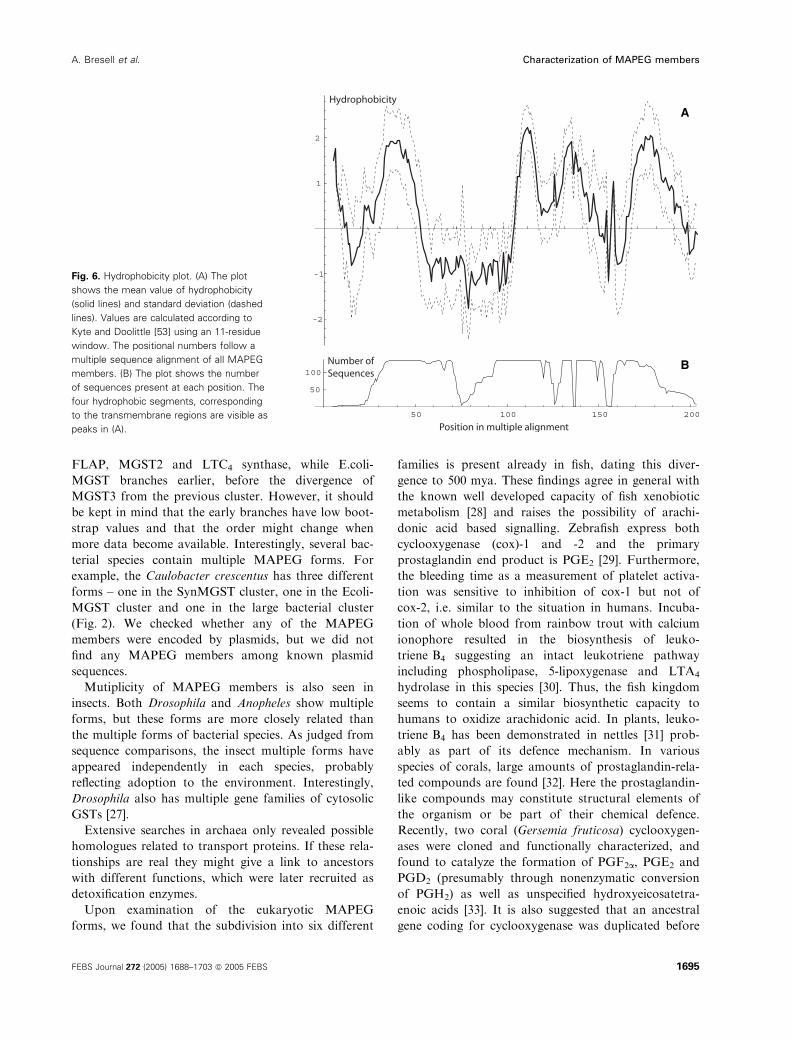
end of first loop (Fig. 6). Both the first and third loop
are located on the cytosolic side of the membrane and
are regions earlier postulated to host the active site
[21,26]. Furthermore, the patterns of the two very sim-
ilar families of PGES (earlier denoted MGST1-like) and
MGST1 do not overlap, even though they both are
located in the first loop.
For the classical FERV pattern, which is a part of
PS01297, we note that it is still included in the two
new and more specific patterns of MGST2 and LTC4
synthase. The last member of PS01297 is FLAP for
which the novel pattern is located in the third loop.
The reason for the similarity and location of these pat-
terns could be a result of short evolutionary time
rather than gain of new features as FLAP, MGST2
and LTC4 synthase have been detected only in higher
eukaryotes to date. However, all patterns in Table 2
will be useful in genome characterizations and func-
tional annotations.
Discussion
The MAPEG family
We have characterized the MAPEG family and found
the eukaryotic forms to consist of six families, while
the prokaryotic forms are clustered at two sites or
more, depending upon whether the E. coli cluster (top)
and the bacterial cluster (bottom) are separated or not
(Fig. 2). The SynMGST branches with the cluster of
Fig. 5. Alignment of pike MGST1 with homologous forms. The N-terminal fragment of pike MGST1 is multiply aligned with other MAPEG
fish members. Positions identical between the pike form and any other fish form are shown in bold. It can be seen that most of the bold
amino acid residues are found within the MGST1 family, supporting evidence for the pike form to belong here. A dendrogram is shown to
the left of the alignment, calculated from the aligned sequences.
Table 2. Sequence patterns for the different MAPEG families.
Family Pattern Position
FLAP P-A-A-F-A-G-x(0,1)-L-x(0,1)-Y-L-x(2)-R-Q-K-Y-F-V-G-Y 123
LTC4 synthase G-P-P-E-F-[DE]-R-[IV]-[FY]-R-A-Q-[AV]-N-[CS]-[ST]-E-Y-F-P 66
MGST1 E-R-V-R-R-[ACG]-H-x-N-D-[IL]-E-N-[IV]-[IV]-P-F-[FLV]-[AGV]-I 92
MGST2 V-[ST]-G-[APS]-[LP]-[DE]-F-[DE]-R-x-F-R-A-x(0,1)-Q-x(0,1)-N-[CNS]-[ALV]-E 63
MGST3 F-N-C-[AIV]-Q-R-[AGS]-H-[AQ]-[NQ]-x(2)-E-x(2,3)-P 90
PGES M-Y-[AIV]-[IV]-A-[IV]-I-T-G-Q-[IMV]-R-L-R-[KR]-K-A-x-A-N 47
Insect D-P-x-V-E-R-V-R-R-A-H-x-N-D-x-E-N-I-L-P 87
Characterization of MAPEG members A. Bresell et al.
1694 FEBS Journal 272 (2005) 1688–1703 ª 2005 FEBS
FLAP, MGST2 and LTC4 synthase, while E.coli-
MGST branches earlier, before the divergence of
MGST3 from the previous cluster. However, it should
be kept in mind that the early branches have low boot-
strap values and that the order might change when
more data become available. Interestingly, several bac-
terial species contain multiple MAPEG forms. For
example, the Caulobacter crescentus has three different
forms – one in the SynMGST cluster, one in the Ecoli-
MGST cluster and one in the large bacterial cluster
(Fig. 2). We checked whether any of the MAPEG
members were encoded by plasmids, but we did not
find any MAPEG members among known plasmid
sequences.
Mutiplicity of MAPEG members is also seen in
insects. Both Drosophila and Anopheles show multiple
forms, but these forms are more closely related than
the multiple forms of bacterial species. As judged from
sequence comparisons, the insect multiple forms have
appeared independently in each species, probably
reflecting adoption to the environment. Interestingly,
Drosophila also has multiple gene families of cytosolic
GSTs [27].
Extensive searches in archaea only revealed possible
homologues related to transport proteins. If these rela-
tionships are real they might give a link to ancestors
with different functions, which were later recruited as
detoxification enzymes.
Upon examination of the eukaryotic MAPEG
forms, we found that the subdivision into six different
families is present already in fish, dating this diver-
gence to 500 mya. These findings agree in general with
the known well developed capacity of fish xenobiotic
metabolism [28] and raises the possibility of arachi-
donic acid based signalling. Zebrafish express both
cyclooxygenase (cox)-1 and -2 and the primary
prostaglandin end product is PGE2 [29]. Furthermore,
the bleeding time as a measurement of platelet activa-
tion was sensitive to inhibition of cox-1 but not of
cox-2, i.e. similar to the situation in humans. Incuba-
tion of whole blood from rainbow trout with calcium
ionophore resulted in the biosynthesis of leuko-
triene B4 suggesting an intact leukotriene pathway
including phospholipase, 5-lipoxygenase and LTA4
hydrolase in this species [30]. Thus, the fish kingdom
seems to contain a similar biosynthetic capacity to
humans to oxidize arachidonic acid. In plants, leuko-
triene B4 has been demonstrated in nettles [31] prob-
ably as part of its defence mechanism. In various
species of corals, large amounts of prostaglandin-rela-
ted compounds are found [32]. Here the prostaglandin-
like compounds may constitute structural elements of
the organism or be part of their chemical defence.
Recently, two coral (Gersemia fruticosa) cyclooxygen-
ases were cloned and functionally characterized, and
found to catalyze the formation of PGF2a, PGE2 and
PGD2 (presumably through nonenzymatic conversion
of PGH2) as well as unspecified hydroxyeicosatetra-
enoic acids [33]. It is also suggested that an ancestral
gene coding for cyclooxygenase was duplicated before
A
B
Fig. 6. Hydrophobicity plot. (A) The plot
shows the mean value of hydrophobicity
(solid lines) and standard deviation (dashed
lines). Values are calculated according to
Kyte and Doolittle [53] using an 11-residue
window. The positional numbers follow a
multiple sequence alignment of all MAPEG
members. (B) The plot shows the number
of sequences present at each position. The
four hydrophobic segments, corresponding
to the transmembrane regions are visible as
peaks in (A).
A. Bresell et al. Characterization of MAPEG members
FEBS Journal 272 (2005) 1688–1703 ª 2005 FEBS 1695

the divergence of the modern cyclooxygenase-1 and -2.
It would be interesting to know at what time during
development the MAPEG proteins (specifically PGE
synthase and FLAP ⁄LTC4 synthase) were associated
with the cyclooxygenase and lipoxygenase protein
families, respectively. At the introduction of these
MAPEG proteins a more specialized level of product
control must have occurred, allowing for the specific
metabolism of the products derived from cyclooxygen-
ases and lipoxygenases into the end products known
today.
Structural implications
Now that over 100 different MAPEG forms are avail-
able, a limited number of conserved residues have
appeared. Two of these, Glu81 and Arg114 (human
MGST1 positional numbering), are found in the puta-
tive transmembrane segments 2 and 3, respectively.
According to electron crystallographic structure deter-
mination of MGST1 [34] and LTC4 synthase [35] and
hydrophobicity properties, the MAPEG forms all
appear to contain four transmembrane regions. MGST1,
LTC4 synthase and PGES [22] are all trimeric
proteins. At the tight border between transmembrane
region 2 and 3, some of the sequences have a Gly-Pro
sequence, typical of a sharp bend. Interestingly, the
almost strictly conserved charged residues mentioned
above are both spaced by exactly 15 residues from the
Gly-Pro bend, strengthening a role for structural
charge interactions. In addition, Asn78 is conserved in
almost all MAPEG members. This residue faces the
cytosol, positioned just before the second transmem-
brane segment, and is probably involved at the active
site. In fact, mutation of these residues in MGST1
seriously affects activity (unpublished observations).
Mutation of the residue corresponding to Arg114
(Arg110) in human mPGES-1 also abolishes activity
[36]. Similarly Arg130, facing the cytosol and adjacent
to the fourth transmembrane segment, is conserved in
nearly all members. The sequence patterns diagnostic
for the PGES and FLAP families are both found
in regions facing the cytosol, thus implying that they
represent family specific regions of the active site
and ⁄or substrate-binding areas.
Observations on the proteins
E.coliMGST and SynMGST represent the first charac-
terized prokaryotic members of the MAPEG super-
family. It was therefore of strong interest to determine
their catalytic properties. Both enzymes efficiently cata-
lyze a glutathione transferase reaction and conse-
quently may be involved in detoxification. In contrast
to human MGSTs 1, 2 and 3, no glutathione peroxi-
dase activity could be detected. Our results thus
demonstrate that both of these highly divergent pro-
karyotic MAPEG members indeed are microsomal
glutathione transferases.
SynMGST, MGST2, and LTC4 synthase to some
extent, align with a postulated lipid binding site of
FLAP (amino acids 48–61) [37–39]. In addition, Syn-
MGST contains conserved arginine and tyrosine resi-
dues implicated in LTC4 synthase activity [40].
However no such activity could be detected, logically
coinciding with the fact that 5-lipoxygenase (forming
the substrate) as well as other lipoxygenases are found
later in evolution [41]. However, recently a 15-lipoxy-
genase was characterized as a secretable enzyme
in Pseudomonas aeruginosa [42] and is, to the best of
our knowledge, the first example of a lipoxygenase in
bacteria.
The cyanobacteria, Synechocystis spp., represent an
interesting model system for further studies of the bio-
logical functions of SynMGST. Knock out experi-
ments, as well as studies of the effects caused by
environmental factors such as light and oxygen on
SynMGST gene expression, will provide important
information about the biological function. Moreover,
if the MGSTs represent common bacterial components
involved in glutathione metabolism mediating cell sur-
vival, they may constitute possible targets for the
development of novel antibiotics.
N-ethylmaleimide, activity and activation
Mammalian MGST1 is activated by sulfhydryl rea-
gents and its relatively modest activity towards CDNB
is increased by 20-fold (from 3 lmolÆmin)1Æmg)1 to
60 lmolÆmin)1Æmg)1) [43]. An MGST has been purified
from Xenopus laevis that was extremely active
(200 lmol min)1Æmg)1) but on the other hand very sen-
sitive to sulfhydryl reagents [44]. The pike enzyme is
also inactivated by N-ethylmaleimide (not shown).
Synechocystis, Arabidopsis and Drosophila MGSTs
appear to represent a third category, namely enzymes
that are insensitive to sulfhydryl reagents. In the case
of Synechocystis and Drosophila enzymes, this is
accounted for by the fact that no cysteine residues are
present and probably explains why SynMGST is an
exceptionally stable protein (in our experience). The
catalytically active form of E.coliMGST contains
three cysteine residues but was not activated by
N-ethylmaleimide. Instead a slight inhibition of the
activity towards CDNB was observed. Apparently,
none of the cysteines is situated at an accessible posi-
Characterization of MAPEG members A. Bresell et al.
1696 FEBS Journal 272 (2005) 1688–1703 ª 2005 FEBS

tion that is critical for enzyme activity of the
E.coliMGST. In conclusion, sulfhydryl reagent activa-
tion ⁄ inactivation cannot be used as a criterion to iden-
tify MAPEG MGST1 members as the activation has
been detected so far only with mammalian MGST1.
Also, the closest relative of MGST1, PGES, is inacti-
vated by N-ethylmaleimide [22] as well as LTC4 syn-
thase [45]. It is evident that cysteine is not involved in
the catalytic mechanism of several MAPEG members,
but could well be relevant for PGES and LTC4 syn-
thase, which harbour cysteines at unique positions.
Conclusion
We have identified several new MAPEG proteins by
sequence homologies with proteins in various databases.
The mammalian members can be traced back 500 mya
as all six families can be found in fish, consistent with a
role in eicosanoid signalling. The gene products
from two representative bacterial strains, E. coli and
Synechocystis sp. were cloned and overexpressed in
E. coli. In addition, plant, insect and fish MAPEG mem-
bers were characterized. As a common denominator,
most MAPEG members catalyze glutathione conjuga-
ting activity towards CDNB or a specific substrate such
as LTC4, some with remarkable efficiency. The enzymes
represent early MAPEG members in their respective
phylogenetic classes and thus create a defined basis for
understanding this superfamily.
Experimental procedures
Materials
Oligonucleotides were synthesized by KEBO, (Stockholm,
Sweden). Pfu DNA polymerase was purchased from Strata-
gene (La Jolla, CA, USA). pGEM T-vector was from
Promega (Madison, WI, USA). Gel extraction and plasmid
isolation kits were from Qiagen (Hilden, Germany). DNA
sequencing kit (ABI PRISM Dye Terminator Cycle Sequen-
cing Ready Reaction Kit) was obtained from Perkin-Elmer
(Boston, MA, USA). Hydroxyapatite (Bio-Gel HTP) was
from Bio-Rad (Hercules, CA, USA).
Sequence comparisons
In the search for new members of the MAPEG superfamily
a set of representative members were selected as seeds. The
seeds were the human member proteins of MGST1-3 (Uni-
prot-Swissprot identifiers P10620, Q99735 and O14880);
FLAP (P20292); LTC4 synthase (Q16873) and PGES
(O14684). Two bacterial members, SynMGST (P73795) and
E.coliMGST (P64515), were additionally selected to com-
plement the six human forms. The eight seeds were used as
query sequences in the search for homologues using fasta
[46] against Swissprot release 41.24 [47], TrEMBL release
24.13 [47] and 138 completely sequenced genomes. Further
screenings were performed against the NCBI non-redund-
ant protein database using psi-blast [48]. Finally, to fetch
unverified translations of MAPEG members the NCBI EST
database (excluding human and mouse) [49] was searched
using tblastn [48]. The resulting nucleotide sequences from
the EST search were translated using getorf from the
emboss package [50]. The open reading frames were filtered
by a minimum size of 100 amino acid residues and flanked
by start and stop codons. These homology searches resulted
in nearly 1000 redundant amino acid sequences which were
followed by an extensive work of manual filtering to obtain
a non-redundant set of sequences by removing duplicates
and non-EST supported alternative splicings.
Multiple sequence alignments and dendrograms
To study the relationships between the new members of the
superfamily we calculated multiple alignments using clu-
stalw [51] on the resulting sequences from the homology
searches. Dendrograms were obtained using neighbor-join-
ing method in the clustalw package and protpars from
the phylip package [52]. An unrooted tree was generated
based on the complete set of sequences of all superfamily
members. To also visualise the more general relationships
of the families included in MAPEG an unrooted consensus
tree was produced. The consensus sequences of the families
of MGST1–3, FLAP, LTC4 synthase, PGES, SynMGST
cluster, E.coliMGST cluster and Insect cluster were gener-
ated by the cons program from the emboss package. A
hydrophobicity plot was generated to verify the structural
similarities of the proteins. It was based on the multiple
sequence alignment of the complete superfamily and calcu-
lated according to Kyte and Doolittle [53] using a window
of 11 residues.
Pattern detection
To characterize the MAPEG families further we extracted
patterns compatible to the PROSITE database [25,54].
These patterns are helpful in annotation of new sequences
and model the unique motifs of a family. The patterns were
generated by the program pratt version 2.1 [55,56]. pratt
was run on sequences from each of the MGST1-3, FLAP,
LTC4 synthase, PGES and Insect families by setting the
maximal pattern length parameter to 20. The best ranked
patterns of each family, shown in Table 2, were selected
and tested for unambiguousness by performing a scan
against Swiss-Prot and TrEMBL with the program fuzz-
pro from the emboss package. The degree of unambiguous-
ness was defined as the fraction of member ranked higher
than the first occurring non-member.
A. Bresell et al. Characterization of MAPEG members
FEBS Journal 272 (2005) 1688–1703 ª 2005 FEBS 1697

Determination of SynMGST mRNA expression
RT-PCR analysis was performed in a PCR buffer contain-
ing 1 mm dNTPs, 1.5 mm MgCl2, 1 lm of the appropriate
primer and 50 ng template (final volume 50 lL). Two
20-mer synthetic oligonucleotides, 5¢-GGGCGGGCTCGG
GCTAAATA-3¢ (sense primer) and 5¢-GGTTGGATCTC
GGTAAATGG-3¢ (antisense primer) (DNA technology,
ApS, Aarhus, Denmark), were designed to anneal within
the ORF of SynMGST. The antisense primer, complement-
ary to the coding strand, was annealed to the SynMGST
mRNA by incubating the mixture at 70 �C for 2 min, fol-
lowed by a temperature shift to 42 �C at a speed of
2 �CÆmin)1. After an additional 30 min at 42 �C, 2.5 units
of AMV reverse transcriptase (Pharmacia Biotech, Uppsala,
Sweden) was added, and primer extension reactions were
carried out for 30 min. Finally, the AMV reverse transcrip-
tase was heat inactivated at 95 �C and the extension prod-
uct was PCR amplified by the addition of sense primer and
2.5 units of Taq polymerase (Pharmacia Biotech). The tem-
perature cycles were 95 �C, 1 min, 55 �C, 1 min and 72 �C1 min, repeated 25 times. The PCR product was run on a
1% agarose gel with a 100 bp ladder and visualized with
ethidium bromide. As positive and negative controls, DNA
and RNA were used as templates, respectively, without the
addition of reverse transcriptase. Preparations of Synecho-
cystis 6803 DNA and RNA were performed as described
previously [57].
Isolation and cloning of the SynMGST
and E.coliMGST
The coding sequence for the SynMGST, corresponding to
the complementary strand of the nucleotide sequence from
89 254 to 89 667 in the Synechocystis genome, accession
number D90909, was amplified from genomic DNA by
PCR. Oligonucleotide primers were constructed to incor-
porate suitable restriction sites (NdeI-HindIII) into the
5¢- and 3¢ ends of the product. Sense primer, 5¢-GAGA
GAGGATCCATATGACAAAAACCGAGTTAC-3¢, NdeI
site; antisense primer, 5¢-GAGAGAAAGCTTCAAAACT
GGGACAGTTG-3¢, HindIII site.
The same method was used to amplify the coding
sequence for the E.coliMGST, corresponding to the nucleo-
tide sequence 10 655–11 080 in the E. coli genome with a
GTG start. Sense primer, 5¢-GAGAGACATATGCCA
TCGGCCATTTTAAAG-3¢; antisense primer, 5¢-GAGA
GAAAGCTTCTAACGCAGGGAGAAAAC-3¢. An alter-
native start site would be the in-frame ATG, 30 nucleotides
downstream of the GTG. This coding region was amplified
using the sense primer 5¢-GAGAGACATATGGTAAGC
GCGCTGTACGCC-3¢.PCR was performed with 0.2 mm dNTPs, 2 mm MgCl2,
0.25 lm of the respective primer, about 0.1 pmol of tem-
plate and 0.5 U of Pfu polymerase. The temperature cycles
were 30 s at 94 �C, 1 min at 40 �C and 2 min at 72 �C,repeated 10 times, followed by 30 s at 94 �C, 1 min at
50 �C and 2 min at 72 �C, repeated 20 times. Finally, the
reaction was terminated by a 7 min extension at 72 �C. ThePCR product was isolated by agarose gel electrophoresis,
purified from the gel and digested with NdeI and HindIII.
The resulting product was gel purified and ligated into the
bacterial expression vector pSP19T7LT as described previ-
ously [58]. Ligated plasmids were transformed into E. coli
XL-1 Blue as described previously [59]. Plasmids were isola-
ted from a number of clones and cleaved with NdeI and
HindIII followed by agarose gel electrophoresis to verify
the size of the inserts. Selected inserts were sequenced on
an Applied Biosystems (Foster City, CA, USA) 373A auto-
mated DNA sequencer using a dye terminator cycle sequen-
cing kit.
The expression construct containing the correct coding
sequence for both the SynMGST and E.coliMGST was
transformed into E. coli BL21 (DE3) (that harboured the
plasmid pLys SL [60]) using the same protocol. Glycerol
stocks were prepared and stored frozen at )70 �C for sub-
sequent use as starting material for the expression experi-
ments.
Overexpression of SynMGST and E.coliMGST
and preparation of membrane fraction
Small aliquots (1–2 lL) of bacterial glycerol stock were
grown in 2· YT medium overnight at 37 �C. The cultures
were diluted 1 : 100 into 2 L of Terrific Broth medium
containing ampicillin (75 lgÆmL)1) and chloramphenicol
(10 lgÆmL)1) in a 5 L flask placed in a thermostated water
bath. The culture was oxygenated by air bubbling and
grown until the D600 was 0.4–1.2. Expression was then
induced by the addition of 0.4 mm isopropyl thio-b-d-gal-actoside, the temperature was switched to 30 �C and the
culture allowed to grow for another 4 h. Thereafter, cells
were pelleted and resuspended in 200 mL 15 mm Tris ⁄HCl,
pH 8.0, 0.25 m sucrose, 0.1 mm EDTA, 1 mm glutathione
(TSEG buffer). Lysozyme was added to a final concentra-
tion of 0.2 mgÆmL)1, and the mixture was gently stirred for
30 min at 4 �C. The resulting spheroplasts were pelleted
(8000 g, 10 min), resuspended in 200 mL TSEG and lysed
by four 30 s sonication pulses from an MSE Soniprep
150 sonifier (Beckenham, Kent, UK) at 40–60% of maxi-
mum power. Magnesium chloride was added to a final con-
centration of 6 mm and DNA and RNA were hydrolyzed
by incubation with DNaseI (4 lgÆmL)1) and RNase A
(4 lgÆmL)1) for 30 min at 4 �C with gentle stirring. Cell
debris was removed by centrifugation at 5000 g for 10 min.
The supernatant was then centrifuged at 180 000 g for 2 h
and the membrane pellets were suspended in 10 mm
potassium phosphate, pH 7.0, 20% (v ⁄ v) glycerol, 0.1 mm
EDTA, 1 mm glutathione.
Characterization of MAPEG members A. Bresell et al.
1698 FEBS Journal 272 (2005) 1688–1703 ª 2005 FEBS

Purification of SynMGST
Solubilisation and partial purification of SynMGST was
performed at 4 �C as follows. Membranes were solubilised
by the addition of an equal volume of 10 mm potassium
phosphate, pH 7.0, 20% (v ⁄ v) glycerol, 0.1 mm EDTA,
1 mm glutathione, 10% (v ⁄ v) Triton X-100 and 15 min
incubation on ice. Insoluble particles were removed by
centrifugation (100 000 g, 20 min). Solubilized membranes
were adsorbed to 10 g of hydroxyapatite equilibrated with
10 mm potassium phosphate, pH 7.0, 20% (v ⁄ v) glycerol,
0.1 mm EDTA, 1 mm glutathione, 1% (v ⁄ v) Triton X-100
(hereafter referred to as buffer A), for 15 min. Hydroxyapa-
tite elution was performed by a batch procedure where the
hydroxyapatite was pelleted by a low speed centrifugation
pulse and washed with two volumes of buffer A, followed
by one volume of 50 mm potassium phosphate in buffer A.
SynMGST was eluted with 0.4 m potassium phosphate in
buffer A and desalted by dialysis for 20 h against two chan-
ges of 60 volumes of buffer A.
Cation-exchange chromatography was performed on a
HiTrap SP, 5 mL (Pharmacia Biotech) column equilibrated
with buffer A. The eluate from the hydroxyapatite chroma-
tography step was loaded on the column and elution of
SynMGST was monitored by enzyme activity measure-
ments (see below). As no activity was retained on the col-
umn, the flow-through was applied onto a DEAE-Sephadex
A-25 (Pharmacia Biotech) anion exchanger equilibrated
with buffer A. As no enzyme activity was retained on either
ion exchanger, the protein content of the flow-through from
the DEAE-Sephadex was examined by SDS ⁄PAGE. A pre-
dominant protein band, comigrating with rat MGST1, was
observed. This band was also observed in fractions from
the other purification steps. The band was cut out from the
gel followed by elution of the protein and the N-terminal
amino acid sequence was determined using an Applied Bio-
systems 477A instrument with on-line analysis of the phe-
nylthiohydantoin amino acids.
Cloning and overexpression of Drosophila
and Arabidopsis MAPEG members
D. melanogaster
The (EST) clone (GenBank AF111426, Mgst1 gene) was
purchased from Invitrogen (Stockholm, Sweden). This
clone encodes Drosophila MGST-like protein. Primers were
designed to introduce a NdeI restriction site immediately
upstream of initiation codon and a HindIII site immediately
downstream of the stop codon. Forward primer, 5¢-gagacatATGGCCAGCCCCGTGGAACT-3¢; reverse primer
5¢-cccaagcttTCAGAAGGCGGCCGAG-3¢ (lower case indi-
cates CLAMP and restriction site). Drosophila MGST was
amplified using PCR (94 �C for 2 min, followed by 35
cycles of 45 s at 94 �C, 30 s at 50 �C and 3 min at 72 �C,
with a final extension of 10 min at 72 �C) with Pfu DNA
polymerase. The PCR product was subcloned into pGEM-T
Easy vector for sequencing, using a PerkinElmer 373 auto-
mated sequencer (Applied Biosystems) using T7 and Sp6
primers. The EST clone showed an additional nucleotide
sequence GAAGA (position 29–33) and C (position 35) as
compared to GenBank AF111246 [61]. In the EST clone,
the asparagine at position 42 of GenBank AF111246was
replaced with a lysine. The cDNA was digested with NdeI
and HindIII and inserted between these sites in the
pSP19T7LT vector. The protein was expressed in E. coli
BL21(DE3) by a slightly modified method compared to
SynMGST. Protein induction with 0.8 mm isopropyl thio-
b-d-galactoside was carried out at 30 �C. After low speed
centrifugation (4000 g) the resulting supernatant was centri-
fuged at 105 000 g for 60 min. The membrane fraction was
resuspended in 0.05 m potassium phosphate buffer (pH 7.4)
containing 0.25 m sucrose and 0.3 mm EDTA. The freshly
prepared membrane fraction was assayed for GST and
glutathione peroxidase (GPx) activities. The protein concen-
tration was measured by the Lowry method using BSA as a
standard [62].
A. thaliana
A plant cDNA displaying sequence homology to human
MGST3 was cloned from an Arabidopsis thaliana cDNA
library from immature green siliques [63]. Primers for the
complete cDNA were designed as follows: forward primer
1, 5¢-ATGGCGGCGATTACAGAATT-3¢, reverse primer1,
5¢-TCAAGCAAGGATCAGAGTGA-3¢. The sequence was
obtained from the TIGR database (At1g65820 68408.m06848
putative microsomal glutathione S-transferase). The same
PCR method was used as described for Drosophila and then
Taq DNA polymerase was added to the PCR mix. Five
additional cycles were performed to add A overhangs to
the linear fragment. The PCR product was subcloned into
the pGEM-T Easy vector for sequence analysis. No muta-
tion was detected. To confirm whether the putative MGST3
is expressed in A. thaliana, northern blot analysis using the
cDNA was carried out in seedlings (data not shown). To
examine the catalytic activity the A. thaliana MGST3 was
expressed in W(R) Saccharomyces cerevisiae strain WAT11
[64]. Reformatting and cloning the MGST3 into the expres-
sion vector pYeDP60 was performed by PCR amplification
using Pfu DNA polymerase. The specific primers to intro-
duce a BamHI site (indicated in bold) immediately
upstream of the initiation codon (start of uppercase) were
designed as follows: forward primer 2, 5¢-cgggatccATGG
CGGCGATTACAGAATTTC-3¢. To obtain the cDNA with
suitable restriction sites, forward primer 2 and T7 primer
were used for PCR with Pfu DNA polymerase as the pGEM-
T Easy vector has an EcoRI site downstream of the stop
codon of MGST3. The PCR product was digested with
A. Bresell et al. Characterization of MAPEG members
FEBS Journal 272 (2005) 1688–1703 ª 2005 FEBS 1699

BamHI and EcoRI and then the reformatted BamHI-EcoRI
fragment was subcloned into BamHI and EcoRI sites of the
pYeDP60 vector. The nucleotide sequence of the cDNA was
determined with a PerkinElmer model 373 automated
sequencer using forward primer 1 and reverse primer 1. No
mutation was detected. The transformed yeast cells were
cultured and the microsomes were prepared by differential
centrifugation. The microsomes were washed with 0.15 m
Tris ⁄HCl buffer (pH 8.0) twice by ultracentrifugation. The
microsomal membranes were resuspended in 0.05 m potas-
sium phosphate buffer containing 0.25 m sucrose and
0.3 mm EDTA. The freshly prepared suspension was assayed
for GST and GPx activities within 4 h. Microsomal proteins
were quantified by the method of Lowry et al. [63].
Protein analysis of pike MGST
Pike liver MGST was purified essentially as was the rat
enzyme [43]. The enzyme displayed a single band upon
SDS ⁄PAGE analysis. The following procedure was used to
prepare samples for amino acid sequence determination.
For removing Triton X-100 from the enzyme solution, the
microsomal GST solution (0.9 mgÆmL)1) was dialysed
against 0.4 m Tris ⁄HCl, pH 8.0, 2 mm EDTA, 8 m urea.
Triton X-100 in the solution was determined by measuring
the absorbance at 275 nm. When Triton X-100 was
removed, the protein was dialysed against H2O for 48 h.
The amino acid sequence of the intact protein was deter-
mined by automated Edman degradation using an Applied
Biosystems 477A instrument with on-line detection of
amino acid phenylthiohydantion derivatives.
Enzyme assays
Enzyme activity with 1-chloro-2,4-dinitrobenzene (CDNB)
and 1,2-epoxy-3-para-nitrophenoxypropane (0.5 mm) was
assayed in 0.1 m potassium phosphate, pH 6.5, containing
1%, 0.1% or 0% (v ⁄ v) Triton X-100 at 30 �C as indicated
[65]. Activity with trans-phenylbut-3-en-2-one was measured
according to [66]. When activity was determined in bacterial
membranes and crude fractions, the concentration of Triton
X-100 was increased to 1% in order to avoid an increase in
turbidity that has been observed with bacterial membrane
fractions. GPx activity towards cumene hydroperoxide or
phospholipid hydroperoxide was determined in a coupled
assay system as described previously [67]. LTC4 synthase
activity as well as glutathione-dependent peroxidase activity
towards 5-hydroperoxy-eicosatetraenoic acid was measured
as described previously [5,7].
Gel electrophoresis and protein determination
SDS ⁄PAGE was performed according to Laemmli [68] in
15% polyacrylamide gels. Purified rat MGST1 and Rain-
bow molecular weight markers (Amersham, Little Chalfont,
Buckinghamshire, UK) were used as standards. Protein
bands were visualized by staining with Coomassie Brilliant
Blue R-250 or silver staining as described [69]. Protein was
determined by the method of Peterson [70] with BSA as
standard.
Acknowledgements
We thank Jan–Ove Jarrhed and the National Super-
computer Centre in Linkoping, Sweden, for valuable
support on the computer side. Per L. Petterson is
gratefully acknowledged for performing PGE synthase
measurements. Technical assistance by Gudrun Tibbe-
lin is gratefully acknowledged. Financial support from
the Swedish Research Council (13x-12564, 13x-12573),
the Swedish Cancer Society, the Swedish National
Board for Laboratory Animals, the Swedish Society of
Medicine, the Magnus Bergvall, Harald Jeansson, the
Research Committee, FMHS, UAE University (HR
was on a supported leave from UAE University, Al
Ain, UAE) and Carl Tryggers foundations, Karolinska
Institutet and Linkoping University is gratefully
acknowledged.
References
1 Jakobsson PJ, Morgenstern R, Mancini J, Ford-
Hutchinson A & Persson B (1999) Common structural
features of MAPEG – A widespread superfamily of
membrane associated proteins with highly divergent
functions in eicosanoid and glutathione metabolism.
Protein Sci 8, 689–692.
2 Samuelsson B (1983) Leukotrienes: mediators of imme-
diate hypersensitivity reactions and inflammation.
Science 220, 568–575.
3 Ford-Hutchinson AW, Gresser M & Young RN (1994)
5-Lipoxygenase. Annu Rev Biochem 63, 383–417.
4 Weinander R, Ekstrom L, Raza H, Lundqvist G, Lindk-
vist B, Sun T-H, Hebert H, Schmidt-Krey I & Morgen-
stern R (1996) Microsomal glutathione transferase
dimensions. In Glutathione S-Transferases: Structure, Fun-
ction and Clinical Implications (Vermeulen NPE, Mulder
GJ, Nieuwenhuyse H, Peter, WHM & van Bladeren PJ,
eds), pp. 49–56. Taylor & Francis Ltd, London.
5 Jakobsson P-J, Mancini JA & Ford-Hutchinson AW
(1996) Identification and characterization of a novel
human microsomal glutathione S-transferase with leuko-
triene C4 synthase activity and significant sequence
identity to 5-lipoxygenase activating protein and leuko-
triene C4 synthase. J Biol Chem 271, 22203–22210.
6 Scoggan KA, Jakobsson PJ & Fordhutchinson AW
(1997) Production of leukotriene C-4 in different human
Characterization of MAPEG members A. Bresell et al.
1700 FEBS Journal 272 (2005) 1688–1703 ª 2005 FEBS

tissues is attributable to distinct membrane bound bio-
synthetic enzymes. J Biol Chem 272, 10182–10187.
7 Jakobsson P-J, Mancini JA, Riendeau D & Ford-
Hutchinson AW (1997) Identification and characteriza-
tion of a novel microsomal enzyme with glutathione-
dependent transferase and peroxidase activities. J Biol
Chem 272, 22934–22939.
8 Jakobsson PJ, Thoren S, Morgenstern R & Samuelsson
B (1999) Identification of human prostaglandin E
synthase: a microsomal, glutathione-dependent, induci-
ble enzyme, constituting a potential novel drug target.
Proc Natl Acad Sci USA 96, 7220–7225.
9 Polyak K, Xia Y, Zweier JL, Kinzler KW & Vogelstein
B (1997) A model for p53-induced apoptosis. Nature
389, 300–305.
10 Jakobsson PJ, Morgenstern R, Mancini J, Ford-Hut-
chinson A & Persson B (2000) Membrane-associated
proteins in eicosanoid and glutathione metabolism
(MAPEG). A widespread protein superfamily. Am J
Respir Crit Care Med 161, S20–S24.
11 Fahey RC & Sundquist AR (1991) Evolution of glu-
tathione metabolism. Adv Enzymol Rel Areas Mol Biol
64, 1–53.
12 Vuilleumier S (1997) Bacterial glutathione S-transferases:
what are they good for? J Bacteriol 179, 1431–1441.
13 Di Ilio C, Aceto A, Piccolomini R, Allocati N, Faraone
A, Cellini L, Ravagnan G & Federici G (1988) Purifica-
tion and characterization of three forms of glutathione
transferase from Proteus mirabilis. Biochem J 255, 971–975.
14 Iizuka M, Inoue Y, Murata K & Kimura A (1989) Puri-
fication and some properties of glutathione S-transferase
from Escherichia coli B. J Bacteriol 171, 6039–6042.
15 Nishida M, Kong KH, Inoue H & Takahashi K (1994)
Molecular cloning and site-directed mutagenesis of glu-
tathione S-transferase from Escherichia coli. The con-
served tyrosyl residue near the N terminus is not
essential for catalysis. J Biol Chem 269, 32536–32541.
16 Rossjohn J, Polekhina G, Feil SC, Allocati N, Masulli
M, De Illio C & Parker MW (1998) A mixed disulfide
bond in bacterial glutathione transferase: functional and
evolutionary implications. Structure 6, 721–734.
17 Kaneko T, Tanaka A, Sato S, Kotani H, Sazuka T,
Miyajima N, Sugiura M & Tabata S (1995) Sequence
analysis of the genome of the unicellular cyanobacter-
ium Synechocystis sp. strain PCC6803. I. Sequence
features in the 1 Mb region from map positions 64% to
92% of the genome. DNA Res 2 (153–66), 191–198.
18 Sun TH, Ling X, Persson B & Morgenstern R (1998) A
highly active microsomal glutathione transferase from
frog (Xenopus laevis) liver that is not activated by
N-ethylmaleimide. Biochem Biophys Res Commun 246,
466–469.
19 Bateman A, Birney E, Durbin R, Eddy SR, Howe KL
& Sonnhammer EL (2000) The Pfam protein families
database. Nucleic Acids Res 28, 263–266.
20 Holm PJ, Morgenstern R & Hebert H (2002) The 3-D
structure of microsomal glutathione transferase 1 at 6 A
resolution as determined by electron crystallography of
p22 (1), 2 (1) crystals. Biochim Biophys Acta 1594, 276–
285.
21 Schmidt-Krey I, Mitsuoka K, Hirai T, Murata K,
Cheng Y, Fujiyoshi Y, Morgenstern R & Hebert H
(2000) The three-dimensional map of microsomal glu-
tathione transferase 1 at 6 A resolution. EMBO J 19,
6311–6316.
22 Thoren S, Weinander R, Saha S, Jegerschold C,
Pettersson PL, Samuelsson B, Hebert H, Hamberg M,
Morgenstern R & Jakobsson PJ (2003) Human
microsomal prostaglandin E synthase-1: purification,
functional characterization, and projection structure
determination. J Biol Chem 278, 22199–22209.
23 Kaneko T, Sato S, Kotani H, Tanaka A, Asamizu E,
Nakamura Y, Miyajima N, Hirosawa M, Sugiura M,
Sasamoto S et al. (1996) Sequence analysis of the gen-
ome of the unicellular cyanobacterium Synechocystis sp.
strain PCC6803. II. Sequence determination of the
entire genome and assignment of potential protein-cod-
ing regions. DNA Res 3, 109–136.
24 Morgenstern R, Guthenberg C & DePierre JW (1982)
Microsomal glutathione transferase. Purification, initial
characterization and demonstration that it is not identi-
cal to the cytosolic glutathione transferases A, B and C.
Eur J Biochem 128, 243–248.
25 Hulo N, Sigrist CJ, Le Saux V, Langendijk-Genevaux
PS, Bordoli L, Gattiker A, De Castro E, Bucher P &
Bairoch A (2004) Recent improvements to the PRO-
SITE database, Nucleic Acids Res 32 Database issue,
D134–137.
26 Busenlehner LS, Codreanu SG, Holm PJ, Bhakat P,
Hebert H, Morgenstern R & Armstrong RN (2004)
Stress sensor triggers conformational response of the
integral membrane protein microsomal glutathione
transferase 1. Biochemistry 43, 11145–11152.
27 Toung YP, Hsieh TS & Tu CP (1993) The glutathione
S-transferase D genes. A divergently organized, intron-
less gene family in Drosophila melanogaster. J Biol Chem
268, 9737–9746.
28 Balk L, Meijer J, Bergstrand A, Astrom A, Morgenstern
R, Seidegard J & DePierre JW (1982) Preparation and
characterization of subcellular fractions from the liver
of the northern pike, Esox lucius. Biochem Pharmacol
31, 1491–1500.
29 Grosser T, Yusuff S, Cheskis E, Pack MA & FitzGerald
GA (2002) Developmental expression of functional
cyclooxygenases in zebrafish. Proc Natl Acad Sci USA
99, 8418–8423.
30 Pettitt T, Rowley A & Barrow S (1989) Synthesis of leu-
kotriene B and other conjugated triene lipoxygenase
products by blood cells of the rainbow trout, Salmo
gairdneri. Biochim Biophys Acta 1003, 1–8.
A. Bresell et al. Characterization of MAPEG members
FEBS Journal 272 (2005) 1688–1703 ª 2005 FEBS 1701

31 Czarnetzki BM, Thiele T & Rosenbach T (1990) Immu-
noreactive leukotrienes in nettle plants (Urtica urens).
Int Arch Allergy Appl Immunol 91, 43–46.
32 Bundy GL (1985) Nonmammalian sources of eicosa-
noids. Adv Prostaglandin Thromboxane Leukot Res 14,
229–262.
33 Jarving R, Jarving I, Kurg R, Brash AR & Samel N
(2004) On the evolutionary origin of cyclooxygenase
(COX) isozymes: Characterisation of marine inverte-
brate Cox genes points to independent duplication
events in vertebrate and invertebrate lineages. J Biol
Chem 279, 13624–13633.
34 Hebert H, Schmidt-Krey I & Morgenstern R (1995) The
projection structure of microsomal glutathione transfer-
ase. EMBO J 14, 3864–3869.
35 Schmidt-Krey I, Kanaoka Y, Mills DJ, Irikura D,
Haase W, Lam BK, Austen KF & Kuhlbrandt W
(2004) Human leukotriene C(4) synthase at 4.5 A
resolution in projection. Structure 12, 2009–2014.
36 Murakami M, Naraba H, Tanioka T, Semmyo N,
Nakatani Y, Kojima F, Ikeda T, Fueki M, Ueno A,
Oh S & Kudo I (2000) Regulation of prostaglandin E2
biosynthesis by inducible membrane- associated prosta-
glandin E2 synthase that acts in concert with cyclooxy-
genase-2. J Biol Chem 275, 32783–32792.
37 Vickers PJ, Adam M, Charleson S, Coppolino MG,
Evans JF & Mancini JA (1992) Identification of amino
acid residues of 5-lipoxygenase-activating protein essen-
tial for the binding of leukotriene biosynthesis inhibi-
tors. Mol Pharmacol 42, 94–102.
38 Mancini JA, Abramovitz M, Cox ME, Wong E, Charle-
son S, Perrier H, Wang Z, Prasit P & Vickers PJ (1993)
5-lipoxygenase-activating protein is an arachidonate
binding protein. FEBS Lett 318, 277–281.
39 Mancini JA, Coppolino MG, Klassen JH, Charleson S &
Vickers PJ (1994) The binding of leukotriene biosynthesis
inhibitors to site-directed mutants of human 5-lipoxygenase-
activating protein. Life Sci 54, PL137–142.
40 Lam BK, Penrose JF, Xu KY, Baldasaro MH &
Austen KF (1997) Site-directed mutagenesis of human
leukotriene C-4 synthase. J Biol Chem 272, 13923–
13928.
41 Toh H, Yokoyama C, Tanabe T, Yoshimoto T &
Yamamoto S (1992) Molecular evolution of cyclooxy-
genase and lipoxygenase. Prostaglandins 44, 291–315.
42 Vance RE, Hong S, Gronert K, Serhan CN & Meka-
lanos JJ (2004) The opportunistic pathogen Pseudo-
monas aeruginosa carries a secretable arachidonate
15-lipoxygenase. Proc Natl Acad Sci USA 101, 2135–
2139.
43 Morgenstern R & DePierre JW (1983) Microsomal glu-
tathione transferase, purification in unactivated form
and further characterization of the activation process,
substrate specificity and amino acid composition. Eur J
Biochem 134, 591–597.
44 Sun J, Chen Y, Li M & Ge Z (1998) Role of antioxi-
dant enzymes on ionizing radiation resistance. Free
Radic Biol Med 24, 586–593.
45 Nicholson DW, Ali A, Klemba MW, Munday NA,
Zamboni RJ & Ford-Hutchinson AW (1992) Human
leukotriene C4 synthase expression in dimethyl sulfoxide-
differentiated U937 cells. J Biol Chem 267, 17849–17857.
46 Pearson WR & Lipman DJ (1988) Improved tools for
biological sequence comparison. Proc Natl Acad Sci
USA 85, 2444–2448.
47 Boeckmann B, Bairoch A, Apweiler R, Blatter MC,
Estreicher A, Gasteiger E, Martin MJ, Michoud K,
O’Donovan C, Phan I et al. (2003) The SWISS-PROT
protein knowledgebase and its (Suppl.)TrEMBL in
2003. Nucleic Acids Res 31, 365–370.
48 Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang
Z, Miller W & Lipman DJ (1997) Gapped BLAST and
PSI-BLAST: a new generation of protein database
search programs. Nucleic Acids Res 25, 3389–3402.
49 Boguski MS, Lowe TM & Tolstoshev CM (1993)
dbEST – database for ‘expressed sequence tags’. Nat
Genet 4, 332–333.
50 Rice P, Longden I & Bleasby A (2000) EMBOSS: the
European molecular biology open software suite. Trends
Genet 16, 276–277.
51 Thompson JD, Higgins DG & Gibson TJ (1994) CLUS-
TAL W: improving the sensitivity of progressive multi-
ple sequence alignment through sequence weighting,
position-specific gap penalties and weight matrix choice.
Nucleic Acids Res 22, 4673–4680.
52 Felsenstein J (1989) Phylip – Phylogeny Inference Pack-
age Version 3.2. Cladistics, 5, 164–166.
53 Kyte J & Doolittle RF (1982) A simple method for dis-
playing the hydropathic character of a protein. J Mol
Biol 157, 105–132.
54 Sigrist CJ, Cerutti L, Hulo N, Gattiker A, Falquet L,
Pagni M, Bairoch A & Bucher P (2002) PROSITE: a
documented database using patterns and profiles as
motif descriptors. Brief Bioinform 3, 265–274.
55 Jonassen I (1997) Efficient discovery of conserved pat-
terns using a pattern graph. Comput Appl Biosci 13,
509–522.
56 Jonassen I, Collins JF & Higgins DG (1995) Finding
flexible patterns in unaligned protein sequences. Protein
Sci 4, 1587–1595.
57 Jansson C, Salih G, Eriksson J, Wiklund R & Ghebre-
medhin H (1998) Methods Enzymol 297, 166–182.
58 Weinander R, Mosialou E, Dejong J, Tu CPD, Dypbukt
J, Bergman T, Barnes HJ, Hoog JO & Morgenstern R
(1995) Heterologous expression of rat liver microsomal
glutathione transferase in simian cos cells and Escheri-
chia coli. Biochem J 311, 861–866.
59 Inoue H, Nojima H & Okayama H (1990) High effi-
ciency transformation of Escherichia coli with plasmids.
Gene 96, 23–28.
Characterization of MAPEG members A. Bresell et al.
1702 FEBS Journal 272 (2005) 1688–1703 ª 2005 FEBS

60 Studier FW (1991) Use of bacteriophage T7 lysozyme
to improve an inducible T7 expression system. J Mol
Biol 219, 37–44.
61 Toba G, Ohsako T, Miyata N, Ohtsuka T, Seong KH &
Aigaki T (1999) The gene search system. A method for
efficient detection and rapid molecular identification of
genes in Drosophila melanogaster. Genetics 151, 725–737.
62 Lowry OH, Rosebrough NJ, Farr AL & Randall RJ,
(1951) Protein measurement with the Folin phenol
reagent. J Biol Chem 193, 265–275.
63 Benveniste I, Tijet N, Adas F, Philipps G, Salaun JP &
Durst F (1998) CYP86A1 from Arabidopsis thaliana
encodes a cytochrome P450-dependent fatty acid
omega-hydroxylase. Biochem Biophys Res Commun 243,
688–693.
64 Helvig C, Tijet N, Benveniste I, Pinot F, Salaun JP &
Durst F (2002) Selective covalent labeling with radiola-
beled suicide substrates for isolating P450s. Methods
Enzymol 357, 352–359.
65 Habig WH, Pabst MJ & Jakoby WB (1974) Glutathione
S-transferases. The first enzymatic step in mercapturic
acid formation. J Biol Chem 249, 7130–7139.
66 Keen JH, Habig WH & Jakoby WB (1976) Mechanism
for the several activities of the glutathione S-trans-
ferases. J Biol Chem 251, 6183–6188.
67 Wendel A (1981) Glutathione peroxidase. Methods Enz-
ymol 77, 325–333.
68 Laemmli UK (1970) Cleavage of structural proteins
during the assembly of the head of bacteriophage T4.
Nature 227, 680–685.
69 Oakley BR, Kirsch DR & Morris NR (1980) A simpli-
fied ultrasensitive silver stain for detecting proteins in
polyacrylamide gels. Anal Biochem 105, 361–363.
70 Peterson GL (1977) A simplification of the protein assay
method of Lowry et al. which is more generally applic-
able. Anal Biochem 83, 346–356.
71 McLellan LI, Wolf CR & Hayes JD (1989) Human
microsomal glutathione S-transferase, Its involvment in
the conjugation of hexachloro-1.3-butadiene with glu-
tathione. Biochem J 258, 87–93.
72 Mosialou E, Andersson C, Lundqvist G, Andersson G,
Bergman T, Jornvall H & Morgenstern R (1993) Human
liver microsomal glutathione transferase – substrate
specificity and important protein sites. FEBS Lett 315,
77–80.
Supplementary material
The following material is available from http://www.blackwellpublishing.com/products/journals/suppmat/EJB/EJB4596/EJB4596sm.htm
Fig. S1. Full alignment of all MAPEG members.
A. Bresell et al. Characterization of MAPEG members
FEBS Journal 272 (2005) 1688–1703 ª 2005 FEBS 1703