bioinformatic and biochemical studies on the phylogenetic variability of proenkephalin-derived...
TRANSCRIPT

BPO
EAa
Ab
e
ArtpTmpsw(o(appbvpPpsatnilqtI
Kol
MYieakeP
*EAM
Neuroscience 165 (2010) 542–552
0d
IOINFORMATIC AND BIOCHEMICAL STUDIES ON THEHYLOGENETIC VARIABILITY OF PROENKEPHALIN-DERIVED
CTAPEPTIDESokievd1owai1(rtcmdspflYdp1in
tecdcebmJgOaaithalua
. BOJNIK,a F. BABOS,b A. MAGYAR,b A. BORSODIa
ND S. BENYHEa*
Institute of Biochemistry, Biological Research Centre, Hungariancademy of Sciences, 6726 Szeged, Temesvari krt 62, Hungary
Research Group of Peptide Chemistry, Hungarian Academy of Sci-nces and Eötvös Lorand University, Budapest, Hungary
bstract—Leu- and Met-enkephalins are proenkephalin-de-ived endogenous pentapeptides with opioid (morphine) ac-ivity. Among the seven enkephalin units found in the humanroenkephalin (PENK), the fourth copy being an octapeptide:yr-Gly-Gly-Phe-Met-Arg-Gly-Leu (HsYGGFMRGL). Bioinfor-atic analysis of the available PENK sequences revealed theresence of other octapeptide orthologues in these precur-or polypeptides. Four types of the elongated Met-enkephalinse identified by searching protein databases, XlYGGFMRGY
three frog species and platypus), GgYGGFMRSV (chicken andne fish species), HpYGGFMNGF (shark) and MmYGGFMRSL
mouse and two lungfish species) were chemically synthesizednd studied in receptor binding and G-protein activation assayserformed on rat brain membranes. All peptides have also beenrepared containing oxidized methionine (M(O)). The overallinding and signalling profile of the novel octapeptides re-ealed moderate opioid agonist activities and a rank order ofotencies for the mu�delta��kappa receptor binding sites.eptides with the oxidized M(O) residue were found to be lessotent in both receptor binding and G-protein stimulationtudies. Phylogenetic neuropeptide libraries, defined here ascollection of mutationally different species variants of or-
hologous and paralogous peptide sequences, represent theatural molecular diversity of the neuropeptides. Such librar-
es can provide a wide range of structural information estab-ishing comparative functional analyses. Since DNA se-uencing data are rapidly increasing, more development inhe natural peptide library approach is expected. © 2010BRO. Published by Elsevier Ltd. All rights reserved.
ey words: binding assay, chemical biodiversity, enkephalin,pioid receptor, PENK evolution, phylogenetic neuropeptide
ibrary.
et- and Leu-enkephalin (Tyr-Gly-Gly-Phe-Met/Leu;GGFM/L) were the first natural opioid peptides to be
dentified (Hughes et al., 1975). Like almost all endog-nous opioid peptides, the enkephalins are cleft fromlarger molecular mass precursor polypeptide, proen-
ephalin or proenkephalin-A (PENK), by posttranslationalnzymatic processing (Noda et al., 1982). MammalianENK contains seven enkephalin repeats, among them
Corresponding author. Tel: �36-62-432-099; fax: �36-62-433-432.-mail address: [email protected] (S. Benyhe).
ibbreviation: DOP, delta opioid peptide; KOP, kappa opioid peptide;OP, mu opioid peptide; PENK, proenkephalin.
306-4522/10 $ - see front matter © 2010 IBRO. Published by Elsevier Ltd. All rightoi:10.1016/j.neuroscience.2009.10.008
542
nly one is Leu-enkephalin, all others belong to Met-en-ephalin. Beside the pentapeptide enkephalin units, PENKs also source for two C-terminally extended, Met-nkephalin containing oligopeptides. The PENK-deri-ed heptapeptide Met-enkephalin-Arg6-Phe7 (YGGFMRF),isplaying opioid and non-opioid properties (Benyhe et al.,997), represents the C-terminal tail of the precursor. Thectapeptide, Met-enkephalin-Arg6-Gly7-Leu8 (YGGFMRGL),as discovered in chromaffin granule cells of the bovinedrenal medulla (Kilpatrick et al., 1981), and later found
n brains of the mouse, rat and guinea pig (Ikeda et al.,982; Zamir et al., 1985), as well as in the human brainPittius et al., 1983; Coveñas et al., 2004). The octapeptideegion in PENK stands at the fourth position (IVth) amonghe opioid core sequences and is encompassed by theanonical Lys-Arg (KR) basic amino acid repeats deter-ining the recognition sites for the processing endopepti-ases (Noda et al., 1982). Injection of YGGFMRGL into thepinal subarachnoid space in chronically catheterized ratsroduced a naloxone-reversible suppression of the tail-ick response (Iadarola et al., 1986). Altered level of theGGFMRGL has been reported for several neurologicalisorders, such as epilepsy (Lason et al., 1992), schizo-hrenia (Iadarola et al., 1991), Parkinson’s (Baronti et al.,991) and Huntington disease (Seizinger et al., 1986),
ndicating the physiological significance of this endoge-ous opioid peptide.
Mutations of nucleotide bases in the coding regions ofhe DNA lead to changes in amino acid sequence, thusxplain the differences observed at the amino acid level ofertain neuropeptides in various animal species. The moreistantly related two species are, the more substitutionsan be found in one and the same neuropeptide. Differ-nce in the amino acid sequence may cause difference iniological function, but usually, the orthologous familyembers seem to produce the same effect (Holmgren and
ensen, 2001). The region carrying the biological activity isenerally the most conserved segment in neuropeptides.ur analysis of up to 55 different PENK sequences avail-ble in major protein databases (PubMed Protein, NCBInd SwissProt) showed that the pentapeptide units are
ndeed well conserved, whereas the C-termini of the ex-ended enkephalins in the (IVth) octapeptide and (VIIth)eptapeptide regions, respectively, are rather variablemong the species studied (Bojnik and Benyhe, unpub-
ished). Thus, we have collected altogether twelve individ-al PENK octapeptide sequences possessing mutationalmino acid replacements. Four novel peptides, represent-
ng one third of the bioinformatically known orthologouss reserved.

scbtpm
C
[omdeH[ttITpbdS
O
AbkcTccwtlMwBSwt
A
IsRgasmfcm
R
Cg2laHs
mahTtltputuwcmzarafri
R
ATBimotaaNsldbppm1wGgw7ctiuvmRr
[
RiTc(�tmfw
E. Bojnik et al. / Neuroscience 165 (2010) 542–552 543
tructures and eleven animal species, have been chemi-ally synthesized and studied by the means of functionaliochemistry. The aim of the present study was, therefore,
o characterize these peptide family members, and com-are them with the most abundant, although not exclusiveammalian octapeptide structure YGGFMRGL.
EXPERIMENTAL PROCEDURES
hemicals and radioligands
3H]Naloxone (28 Ci/mmol), [3H]DAMGO ((D-Ala2,NMePhe4,Gly5-l)enkephalin; 41 Ci/mmol), [3H]Tyr1,Ile5,6deltorphin-2 (48 Ci/mol) were radiolabeled in the Isotope Laboratory of BRC (Ra-iolab), Szeged as described previously (Tóth et al., 1982; Nevint al., 1994). [3H]U-69,593 (55 Ci/mmol) and unlabelled U-69,593that is (�)-(5�,7�,8�)-N-Methyl-N-[7-(1-pyrrolidinyl)-1-oxaspiro-
4.5]dec-8-yl] were purchased from Amersham Biosciences andhe Upjohn Co. Radiolabeled GTP�S, guanosine-5-[�-35S]-riphosphate (1204 Ci/mmol) was purchased from the Isotopenstitute Ltd. (Budapest, Hungary). EDTA, polyethylenimine (PEI),ris(hydroxymethyl)amino-methane (Tris), guanosine 5’-diphos-hate (GDP), unlabeled GTP�S, and the peptidase inhibitorsestatin and phosphoramidon were purchased from Sigma-Al-rich (St. Louis, MO, USA). Captopril was obtained from Thequibb Institute for Medical Research (Princeton, NJ, USA).
pioid peptides
ll natural peptides were chemically synthesized (not purified fromiological sources). Mammalian Met-enkephalin and Met-en-ephalin-Arg-Gly-Leu (YGGFMRGL) used in this study was pur-hased from Bachem Feinbiochimica, Bubendorf, Switzerland.he other endogenous octapeptides, as XlYGGFMRGY (Africanlawed frog, Xenopus laevis), YGGFMRSV (Florida gar, fish spe-ies), YGGFMNGF (bullhead shark) and YGGFMRSL (lungfish)as prepared at the Research Group for Peptide Chemistry of
he Hungarian Academy of Sciences, Budapest, Hungary, by so-id phase peptide synthesis with Fmoc/tBu strategy (SYRO,
ultiSyntech automated synthesizer, 2-Cl-trityl resin). Peptidesere purified by analytical RP-HPLC (Knauer, Synergi MAX-RP,erlin, Germany) and preparative FPLC (Pharmacia, Uppsala,veden; Vydac C18 column, Hesperia, CA, USA). The structuresere confirmed by mass spectrometry (Bruker Esquire 3000� ion
rap machine).
nimals
nbred Wistar rats (250–300 g body weight) and guinea pigs (R9train) were housed in the local animal house of the Biologicalesearch Center (BRC, Szeged, Hungary). Animals were kept inroups of four, allowed free access to standard food and tap waternd maintained on a 12:12 h light/dark cycle until the time ofacrifice. Animals were handled according to the European Com-unities Council Directives (86/609/ECC) and the Hungarian Act
or the Protection of Animals in Research (XXVIII.tv. 32. §). Ac-ordingly, the number of animals and their suffering were mini-ized.
at brain membrane preparations
rude membrane fractions from brains of Wistar rats and R9uinea pigs were prepared as described earlier (Bojnik et al.,009). Animals were decapitated and the brains without cerebel-
um were quickly removed and washed several times to removeny unwanted blood or tissue particles with chilled 50 mM Tris–Cl (pH 7.4) buffer. The brains were blotted dry, weighed and
uspended in 5 vol/wt of the original brain tissue with ice cold 50 tM Tris–HCl (pH 7.4) buffer. The brains were than homogenizedt 1000� rpm with an electrically driven Braun Teflon-glass rota-omogenizer at 4 °C, using 10 to 15 strokes of the homogenizer.he final volume of the homogenate was made up to 30 vol/wt of
he brain and filtered through four layers of gauze to remove anyarger aggregates. After centrifugation with a Sorvall RC5C cen-rifuge at 40,000�g (18,000� rpm) for 20 min at 4 °C, the resultingellet was resuspended in fresh buffer (30 volumes/weight) bysing a vortex. The suspension was incubated for 30 min at 37 °Co remove any endogenous opioids. Centrifugation was repeatednder the same conditions as described above, and final pelletas resuspended in 5 vol of 50 mM Tris–HCl (pH 7.4) bufferontaining 0.32 M sucrose to give a final concentration of 3–4g/ml protein. The presence of sucrose is necessary for stabili-
ation of proteins for storage. The membranes were kept in 5 mlliquots at �70 °C until use. The binding activity of the proteinemained stable for at least 2 months. Membranes were thawednd resuspended in 50 mM Tris–HCl (pH 7.4) buffer and centri-uged at 40,000�g for 20 min at 4 °C to remove the sucrose. Theesulting pellets were taken up in appropriate fresh buffer andmmediately used in binding assays.
eceptor binding assays
ll binding assays were performed at 25 °C for 30 min in 50 mMris–HCl buffer (pH 7.4) in a final volume of 1 ml, containing 1 mgSA and 0.2–0.4 mg/ml membrane protein. Samples were made
n disposable plastic assay tubes (Sarstedt Co., Nümbrecht, Ger-any). Rat brain membranes were incubated with the generalpioid agonist radioligand [3H]naloxone (0.8–1.2 nM), or with
ype-specific radioprobes, such as the selective MOP receptorgonist [3H]DAMGO (0.9–1.2 nM), the DOP receptor selectivegonist [3H]Tyr1,Ile5,6deltorphin-2 (Tyr-D-Ala-Phe-Glu-Ile-Ile-Gly-H2; 0.8–1.3 nM), or with 0.4–0.7 nM of the KOP receptor-pecific agonist [3H]U-69,593 in the presence of unlabeled testigands (their concentrations ranged from 10�5 to 10�11 M). Con-itions for incubations are given in the figure legends. Non-specificinding was determined in the presence of 10 �M naloxone. Threeeptidase inhibitors, 1 �M captopril, 1 �M bestatin and 1 �Mhosphoramidon were included in the assay buffer to preventethabolic inactivation of the octapeptides (Hiranuma et al.,997). Reaction was terminated and bound and free radioligandsere separated by rapid filtration under vacuum through WhatmanF/C (radiolabeled peptides) or GF/B (radiolabeled alkaloids)lass fiber filters by using Brandel M24R Cell Harvester. Filtersere washed three times with 5 ml ice cold 50 mM Tris–HCl (pH.4) buffer. After filtration and separation procedure had beenompleted, fiber-disks were dried under an infrared lamp, andhen removed from the filter-sheet by tweezers. Each disk wasnserted into UltimaGold™ environment-friendly, non-volatile, tol-ene-free scintillation cocktail, and placed into individual sampleials (transparent glass, Packard). Bound radioactivity was deter-ined in Packard Tricarb 2300TR liquid scintillation analyzer.eceptor binding experiments were performed in duplicate and
epeated at least three times.
35S]GTP�S binding assays
at brain membrane fractions (�10 �g of protein/sample) werencubated at 30 °C for 60 min in Tris–EGTA buffer (50 mMris–HCl, 1 mM EGTA, 3 mM MgCl2, 100 mM NaCl, pH 7.4)ontaining [35S]GTP�S (0.05 nM) and increasing concentrations10�9 to 10�5 M) of the compounds tested in the presence of 30M GDP in a final volume of 1 ml. Total binding was measured in
he absence of test compound, non-specific binding was deter-ined in the presence of 10 �M unlabeled GTP�S and subtracted
rom total binding to calculate the specific binding. The reactionas started by addition of [35S]GTP�S and terminated by filtrating
he samples through Whatman GF/B glass fiber filters. Filters

w7r(to
D
RtEGSpwfictvoc
ardwG
Bh
AssdTmmp
M
t(cnptbtqtalcpTte(A
tcotpolpttli((tnacabta
R
OeeoaDtmeTpMar[a
X
wt
p[Naatf
H
tsat
K
E. Bojnik et al. / Neuroscience 165 (2010) 542–552544
ere washed three times with ice cold 50 mM Tris–HCl buffer (pH.4) using Brandel M24R Cell Harvester, then dried, and boundadioactivity was detected in UltimaGold™ scintillation cocktailPackard). Agonist-induced receptor-mediated G-protein stimula-ion is given as percentage over the specific [35S]GTP�S bindingbserved in the absence of receptor ligands (basal activity).
ata analysis
adioligand binding experiments were performed in duplicate andhe [35S]GTP�S binding assays were carried out in triplicate.xperimental data were analyzed and graphically processed byraphPad Prism (version 4.00 for Windows, GraphPad Software,an Diego, CA, USA, www.graphpad.com) research softwareackage with standard office computers. Displacement curvesere fitted by non-linear regression using the one-site competitiontting option with no ligand depletion. The equilibrium inhibitiononstant (Ki value) was calculated from the IC50 values accordingo the built-in Cheng-Prusoff equation module. Whereas the IC50
alue for a compound may vary between experiments dependingn radioligand concentration, the Ki is an absolute value. It isalculated from the IC50 using the Cheng-Prusoff equation:
Ki�IC50 ⁄ (1�([L] ⁄ Kd))
Where (L) is the concentration of free radioligand used in thessay and Kd is the dissociation constant of the radioligand for theeceptor. G-protein stimulation data were analyzed by the sigmoidose-response curve fit option of Prism. Statistical comparisonsere made by the built-in statistics modul of GraphPadPrism andraphPad InStat 3.0 software packages.
RESULTS
ioinformatic studies on the phylogeneticeterogeneity of PENK-related octapeptides
n analysis of public protein databases (NCBI Protein, En-embl) has shown significant diversity in the amino acidequences of proenkephalin-derived octapeptides. Fourteenistinct orthologues have been identified among 55 species.he ubiquitous HsYGGFMRGL sequence is present in 26ammals including the human (Homo sapiens), althoughore other mammalian octapeptides exist, for exam-le, OaYGGFMRGY (platypus, Ornithorhyncus anatinus),
dYGGFMRSY (rainforest opossum, Monodelphis domes-ica), FcYGGFMRAL (cat, Felis catus) and CfYGGFMRALdog, Canis familiaris), MmYGGFMRSL (mouse, Mus mus-ulus), OcYGGFMRGV (European rabbit, Oryctolagus cu-iculus) and OpYGGFMRGV (American pika, Ochotonarinceps). Of the other vertebrate animals tested, includingen fish species, seven amphibians, four reptiles and twoirds, even more variability in the structure of the octapep-ide is observed (data not shown). Four different se-uences have been selected for the chemical synthesis:he “mouse sequence” MmYGGFMRSL (1) is uniquemong mammals, but is also present in the Australian
ungfish (Neoceratodus forsteri; NfYGGFMRSL) and Afri-an lungfish (Protopterus annectens; PaYGGFMRSL)roenkephalins (Sollars et al., 2000; Dores et al., 2000).he double substituted HpYGGFMNGF (2) sequence dis-
inctively found in a cartilaginous fish species (Komorowskit al., 2004, unpublished observations), the bullhead sharkHeterodontus portusjacksoni), is the only one in that the
rg6 residue is replaced (R¡N). Moreover, the shark oc- zapeptide ends with an aromatic phenylalanine (Phe8) sidehain as a consequence of L¡F substitution. The structuref OaYGGFMRGY (3) represent another orthologous pep-
ide bearing aromatic (Tyr8) C-terminus due to L¡Y re-lacement. In addition to the platypus, this sequence alsoccurs in two pipide frog species, the tetraploid Xenopus
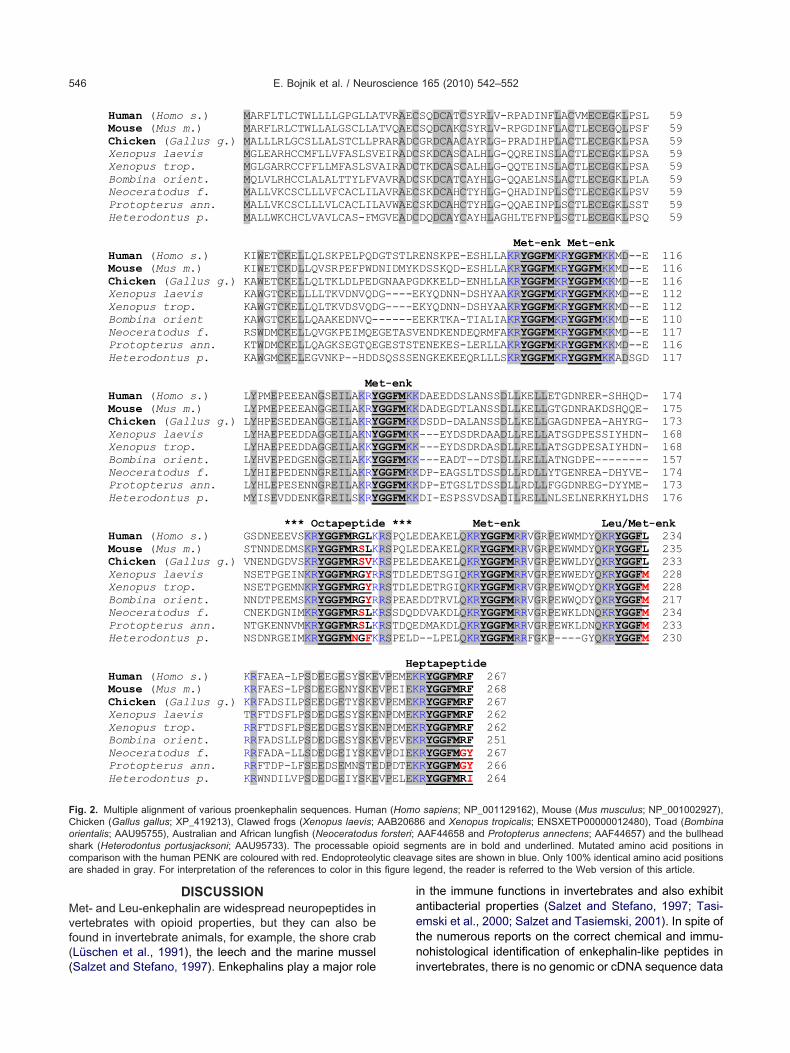
aevis (Martens and Herbert, 1994) and the diploid Xeno-us tropicalis, and in the fire-bellied toad, Bombina orien-alis (Dores et al., 2001). The GgYGGFMRSL (4) octapep-ide is included in the chicken (Gallus gallus) proenkepha-in (prediction from genomic sequence, NCBI), moreovern the incompletely sequenced PENK of the florida garLepisosteus platyrhynchus; LpYGGFMRSL) fish speciesKhalap et al., 2005). The natural occurrence of the oc-apeptides studied here in detail are shown in a phyloge-etic tree illustrating the evolutionary positions of their hostnimal species (Fig. 1A). A dendrogram indicating thealculated bootstrap values (MEGA: Molecular Evolution-ry Genetics Analysis; http://www.megasoftware.net) andranch lengths is also shown (Fig. 1B). All full-size propep-ides carrying various octapeptide components can well beligned to the human PENK sequence (Fig. 2).
eceptor binding experiments
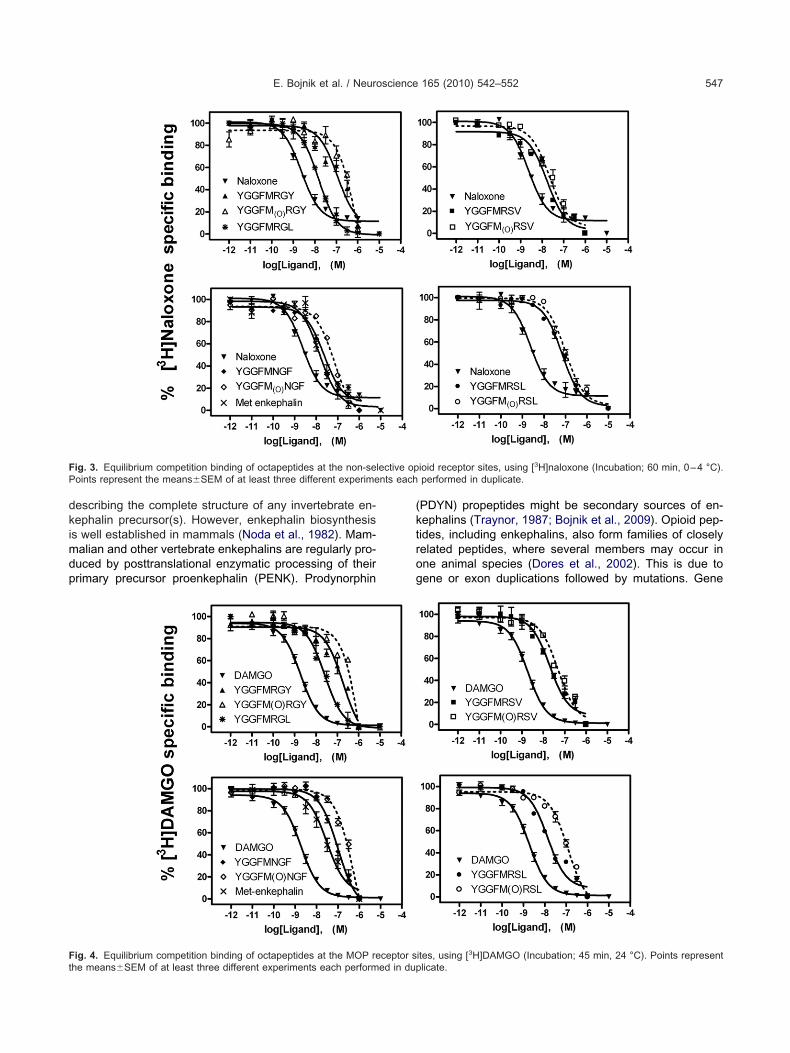
pioid receptor binding affinities of the elongated Met-nkephalin sequences were studied in [3H]naloxone het-rologous competition binding assays. Naloxone is an opi-id antagonist with high affinity, capable of interacting withll types of opioid receptors, including �- or MOP, �- orOP and �- or KOP receptors. Affinities of the octapep-
ides in [3H]naloxone equilibrium binding assays wereodest, while the unlabelled homologous ligand naloxonexhibited the highest affinity in these experiments (Fig. 3).he frog octapeptide YGGFMRGY was the weakest com-etitor among the four sequences in this assay. Using aOP receptor specific peptide radioprobe, [3H]DAMGO,ll of the novel sequences could effectively displace theadioligand in concentration dependent manner. In the3H]DAMGO competition assays MmYGGFMRSL (mouse)nd GgYGGFMRSV (chicken) were the most potent, while
lYGGFMRGY (clawed frog) and HpYGGFMNGF (shark)ere weaker in competing reversibly with the MOP recep-
or binding sites (Fig. 4).DOP receptor binding and selectivity of the com-
ounds were measured with a radiolabelled peptide,3H]Tyr1,Ile5,6deltorphin-2 (Tyr-D-Ala-Phe-Glu-Ile-Ile-Gly-H2). This synthetic, disubstituted deltorphin derivative ishighly specific agonist for the DOP receptors (Nevin et
l., 1994). In our DOP receptor assays the novel octapep-ides concentration dependently displaced the radioproberom the opioid �-sites. Again, XlYGGFMRGY (frog) and
pYGGFMNGF (shark) peptides having aromatic C-erminal side chains were slightly weaker, whereas theerine replaced orthologues GgYGGFMRSV (chicken)nd MmYGGFMRSL (mouse) were more effective in
erms of DOP receptor binding (Fig. 5).Octapeptide analogues were further studied at the
OP receptor sites labelled by a tritiated nonpeptide ben-
eneacetamide analogue, U-69,593 which is among the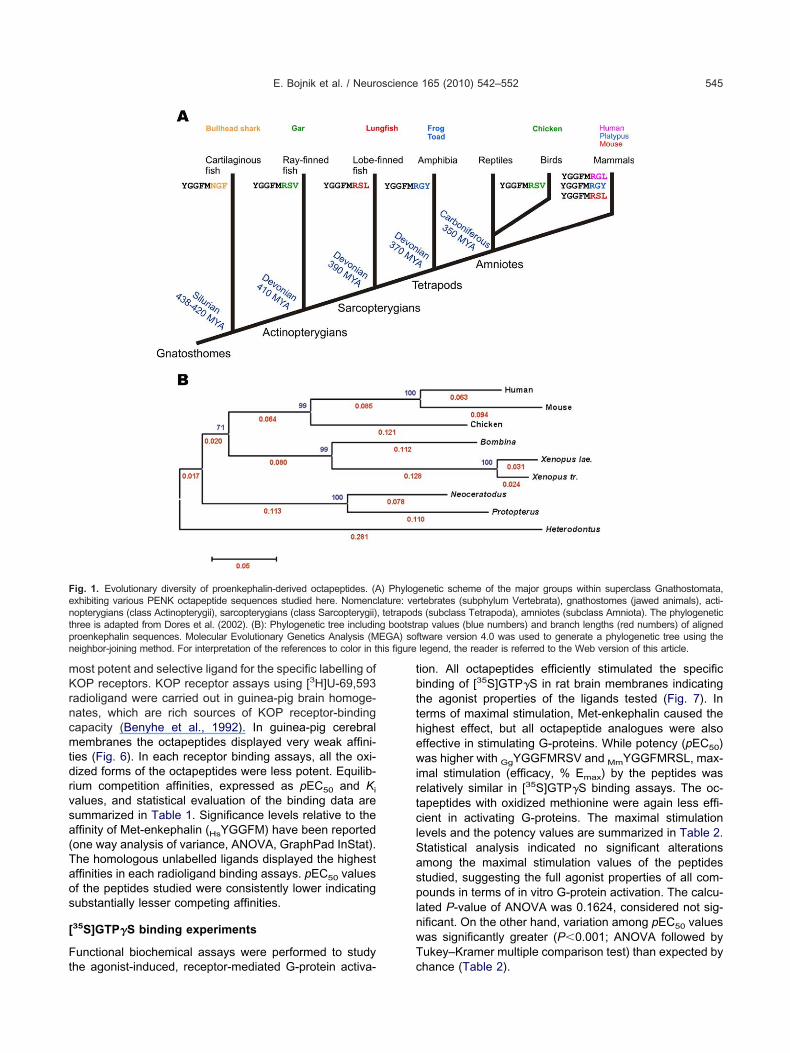
mKrncmtdrvsa(Taos
[
Ft
tbtthewirtclSasplnwT
Fentp EGA) son his figure
E. Bojnik et al. / Neuroscience 165 (2010) 542–552 545
ost potent and selective ligand for the specific labelling ofOP receptors. KOP receptor assays using [3H]U-69,593
adioligand were carried out in guinea-pig brain homoge-ates, which are rich sources of KOP receptor-bindingapacity (Benyhe et al., 1992). In guinea-pig cerebralembranes the octapeptides displayed very weak affini-
ies (Fig. 6). In each receptor binding assays, all the oxi-ized forms of the octapeptides were less potent. Equilib-ium competition affinities, expressed as pEC50 and Ki
alues, and statistical evaluation of the binding data areummarized in Table 1. Significance levels relative to theffinity of Met-enkephalin (HsYGGFM) have been reportedone way analysis of variance, ANOVA, GraphPad InStat).he homologous unlabelled ligands displayed the highestffinities in each radioligand binding assays. pEC50 valuesf the peptides studied were consistently lower indicatingubstantially lesser competing affinities.
35S]GTP�S binding experiments
unctional biochemical assays were performed to study
ig. 1. Evolutionary diversity of proenkephalin-derived octapeptides. (Axhibiting various PENK octapeptide sequences studied here. Nomencopterygians (class Actinopterygii), sarcopterygians (class Sarcopterygii)
hree is adapted from Dores et al. (2002). (B): Phylogenetic tree includinroenkephalin sequences. Molecular Evolutionary Genetics Analysis (Meighbor-joining method. For interpretation of the references to color in t
he agonist-induced, receptor-mediated G-protein activa- c
ion. All octapeptides efficiently stimulated the specificinding of [35S]GTP�S in rat brain membranes indicating
he agonist properties of the ligands tested (Fig. 7). Inerms of maximal stimulation, Met-enkephalin caused theighest effect, but all octapeptide analogues were alsoffective in stimulating G-proteins. While potency (pEC50)as higher with GgYGGFMRSV and MmYGGFMRSL, max-
mal stimulation (efficacy, % Emax) by the peptides waselatively similar in [35S]GTP�S binding assays. The oc-apeptides with oxidized methionine were again less effi-ient in activating G-proteins. The maximal stimulation
evels and the potency values are summarized in Table 2.tatistical analysis indicated no significant alterationsmong the maximal stimulation values of the peptidestudied, suggesting the full agonist properties of all com-ounds in terms of in vitro G-protein activation. The calcu-
ated P-value of ANOVA was 0.1624, considered not sig-ificant. On the other hand, variation among pEC50 valuesas significantly greater (P�0.001; ANOVA followed byukey–Kramer multiple comparison test) than expected by
enetic scheme of the major groups within superclass Gnathostomata,rtebrates (subphylum Vertebrata), gnathostomes (jawed animals), acti-s (subclass Tetrapoda), amniotes (subclass Amniota). The phylogeneticap values (blue numbers) and branch lengths (red numbers) of alignedftware version 4.0 was used to generate a phylogenetic tree using thelegend, the reader is referred to the Web version of this article.
) Phyloglature: ve, tetrapodg bootstr
hance (Table 2).
Mvf((
iaetn
FCosca figure le
E. Bojnik et al. / Neuroscience 165 (2010) 542–552546
DISCUSSIONet- and Leu-enkephalin are widespread neuropeptides in
ertebrates with opioid properties, but they can also beound in invertebrate animals, for example, the shore crabLüschen et al., 1991), the leech and the marine mussel
Human (Homo s.) MARFLTLCTWLLLLGPGLLAMouse (Mus m.) MARFLRLCTWLLALGSCLLAChicken (Gallus g.) MALLLRLGCSLLALSTCLLPXenopus laevis MGLEARHCCMFLLVFASLSVXenopus trop. MGLGARRCCFFLLMFASLSVBombina orient. MQLVLRHCCLALALTTYLFVNeoceratodus f. MALLVKCSCLLLVFCACLILProtopterus ann. MALLVKCSCLLLVLCACLILHeterodontus p. MALLWKCHCLVAVLCAS-FM
Human (Homo s.) KIWETCKELLQLSKPELPQDMouse (Mus m.) KIWETCKDLLQVSRPEFPWDChicken (Gallus g.) KAWETCKELLQLTKLDLPEDXenopus laevis KAWGTCKELLLLTKVDNVQDXenopus trop. KAWGTCKELLQLTKVDSVQDBombina orient KAWGTCKELLQAAKEDNVQ-Neoceratodus f. RSWDMCKELLQVGKPEIMQEProtopterus ann. KTWDMCKELLQAGKSEGTQEHeterodontus p. KAWGMCKELEGVNKP--HDD
MeHuman (Homo s.) LYPMEPEEEANGSEILAKRYMouse (Mus m.) LYPMEPEEEANGGEILAKRYChicken (Gallus g.) LYHPESEDEANGGEILAKRYXenopus laevis LYHAEPEEDDAGGEILAKNYXenopus trop. LYHAEPEEDDAGGEILAKKYBombina orient. LYHVEPEDGENGGEILAKKYNeoceratodus f. LYHIEPEDENNGREILAKRYProtopterus ann. LYHLEPESENNGREILAKRYHeterodontus p. MYISEVDDENKGREILSKRY
*** OctapeptidHuman (Homo s.) GSDNEEEVSKRYGGFMRGLKMouse (Mus m.) STNNDEDMSKRYGGFMRSLKChicken (Gallus g.) VNENDGDVSKRYGGFMRSVKXenopus laevis NSETPGEINKRYGGFMRGYRXenopus trop. NSETPGEMNKRYGGFMRGYRBombina orient. NNDTPEEMSKRYGGFMRGYRNeoceratodus f. CNEKDGNIMKRYGGFMRSLKProtopterus ann. NTGKENNVMKRYGGFMRSLKHeterodontus p. NSDNRGEIMKRYGGFMNGFK
Human (Homo s.) KRFAEA-LPSDEEGESYSKEMouse (Mus m.) KRFAES-LPSDEEGENYSKEChicken (Gallus g.) KRFADSILPSEEDGETYSKEXenopus laevis TRFTDSFLPSDEDGESYSKEXenopus trop. RRFTDSFLPSEEDGESYSKEBombina orient. RRFADSLLPSDEDGESYSKENeoceratodus f. RRFADA-LLSDEDGEIYSKEProtopterus ann. RRFTDP-LFSEEDSEMNSTEHeterodontus p. KRWNDILVPSDEDGEIYSKE
ig. 2. Multiple alignment of various proenkephalin sequences. Humahicken (Gallus gallus; XP_419213), Clawed frogs (Xenopus laevis;rientalis; AAU95755), Australian and African lungfish (Neoceratodushark (Heterodontus portusjacksoni; AAU95733). The processable oomparison with the human PENK are coloured with red. Endoproteolyre shaded in gray. For interpretation of the references to color in this
Salzet and Stefano, 1997). Enkephalins play a major role i
n the immune functions in invertebrates and also exhibitntibacterial properties (Salzet and Stefano, 1997; Tasi-mski et al., 2000; Salzet and Tasiemski, 2001). In spite ofhe numerous reports on the correct chemical and immu-ohistological identification of enkephalin-like peptides in
SQDCATCSYRLV-RPADINFLACVMECEGKLPSL 59SQDCAKCSYRLV-RPGDINFLACTLECEGQLPSF 59GRDCAACAYRLG-PRADIHPLACTLECEGKLPSA 59SKDCASCALHLG-QQREINSLACTLECEGKLPSA 59TKDCASCALHLG-QQTEINSLACTLECEGKLPSA 59SKDCATCAYHLG-QQAELNSLACTLECEGKLPLA 59SKDCAHCTYHLG-QHADINPLSCTLECEGKLPSV 59SKDCAHCTYHLG-QQAEINPLSCTLECEGKLSST 59DQDCAYCAYHLAGHLTEFNPLSCTLECEGKLPSQ 59
Met-enk Met-enkENSKPE-ESHLLAKRYGGFMKRYGGFMKKMD--E 116DSSKQD-ESHLLAKRYGGFMKRYGGFMKKMD--E 116DKKELD-ENHLLAKRYGGFMKRYGGFMKKMD--E 116KYQDNN-DSHYAAKRYGGFMKRYGGFMKKMD--E 112KYQDNN-DSHYAAKRYGGFMKRYGGFMKKMD--E 112EKRTKA-TIALIAKRYGGFMKRYGGFMKKMD--E 110ENDKENDEQRMFAKRYGGFMKRYGGFMKKMD--E 117ENEKES-LERLLAKRYGGFMKRYGGFMKKMD--E 116NGKEKEEQRLLLSKRYGGFMKRYGGFMKKADSGD 117
DAEEDDSLANSSDLLKELLETGDNRER-SHHQD- 174DADEGDTLANSSDLLKELLGTGDNRAKDSHQQE- 175DSDD-DALANSSDLLKELLGAGDNPEA-AHYRG- 173---EYDSDRDAADLLRELLATSGDPESSIYHDN- 168---EYDSDRDASDLLRELLATSGDPESAIYHDN- 168---EADT--DTSDLLRELLATNGDPE-------- 157DP-EAGSLTDSSDLLRDLLYTGENREA-DHYVE- 174DP-ETGSLTDSSDLLRDLLFGGDNREG-DYYME- 173DI-ESPSSVDSADILRELLNLSELNERKHYLDHS 176
Met-enk Leu/Met-enkDEAKELQKRYGGFMRRVGRPEWWMDYQKRYGGFL 234DEAKELQKRYGGFMRRVGRPEWWMDYQKRYGGFL 235DEAKELQKRYGGFMRRVGRPEWWLDYQKRYGGFL 233DETSGIQKRYGGFMRRVGRPEWWEDYQKRYGGFM 228DETRGIQKRYGGFMRRVGRPEWWQDYQKRYGGFM 228DDTRVLQKRYGGFMRRVGRPEWWQDYQKRYGGFM 217DVAKDLQKRYGGFMRRVGRPEWKLDNQKRYGGFM 234DMAKDLQKRYGGFMRRVGRPEWKLDNQKRYGGFM 233--LPELQKRYGGFMRRFGKP----GYQKRYGGFM 230
ptapeptideRYGGFMRF 267RYGGFMRF 268RYGGFMRF 267RYGGFMRF 262RYGGFMRF 262RYGGFMRF 251RYGGFMGY 267RYGGFMGY 266RYGGFMRI 264
sapiens; NP_001129162), Mouse (Mus musculus; NP_001002927),6 and Xenopus tropicalis; ENSXETP00000012480), Toad (BombinaAAF44658 and Protopterus annectens; AAF44657) and the bullheadments are in bold and underlined. Mutated amino acid positions in
age sites are shown in blue. Only 100% identical amino acid positionsgend, the reader is referred to the Web version of this article.
TVRAECTVQAECRARADCEIRADCAIRADCAVRADCAVRAECAVWAECGVEADC
GTSTLRNIDMYKGNAAPGG----EG----E-----EGETASVGESTSTSQSSSE
t-enkGGFMKKGGFMKKGGFMKKGGFMKKGGFMKKGGFMKKGGFMKKGGFMKKGGFMKKe ***RSPQLERSPQLERSPELERSTDLERSTDLERSPEAERSSDQDRSTDQERSPELD
HeVPEMEKVPEIEKVPEMEKNPDMEKNPDMEKVPEVEKVPDIEKDPDTEKVPELEK
n (HomoAAB2068forsteri;pioid segtic cleav
nvertebrates, there is no genomic or cDNA sequence data
dkimdp
(ktrog
F lective opP nts each
Ft
E. Bojnik et al. / Neuroscience 165 (2010) 542–552 547
escribing the complete structure of any invertebrate en-ephalin precursor(s). However, enkephalin biosynthesiss well established in mammals (Noda et al., 1982). Mam-
alian and other vertebrate enkephalins are regularly pro-uced by posttranslational enzymatic processing of theirrimary precursor proenkephalin (PENK). Prodynorphin
ig. 3. Equilibrium competition binding of octapeptides at the non-seoints represent the means�SEM of at least three different experime
ig. 4. Equilibrium competition binding of octapeptides at the MOP receptor she means�SEM of at least three different experiments each performed in dup
PDYN) propeptides might be secondary sources of en-ephalins (Traynor, 1987; Bojnik et al., 2009). Opioid pep-ides, including enkephalins, also form families of closelyelated peptides, where several members may occur inne animal species (Dores et al., 2002). This is due toene or exon duplications followed by mutations. Gene
ioid receptor sites, using [3H]naloxone (Incubation; 60 min, 0–4 °C).performed in duplicate.
3
ites, using [ H]DAMGO (Incubation; 45 min, 24 °C). Points representlicate.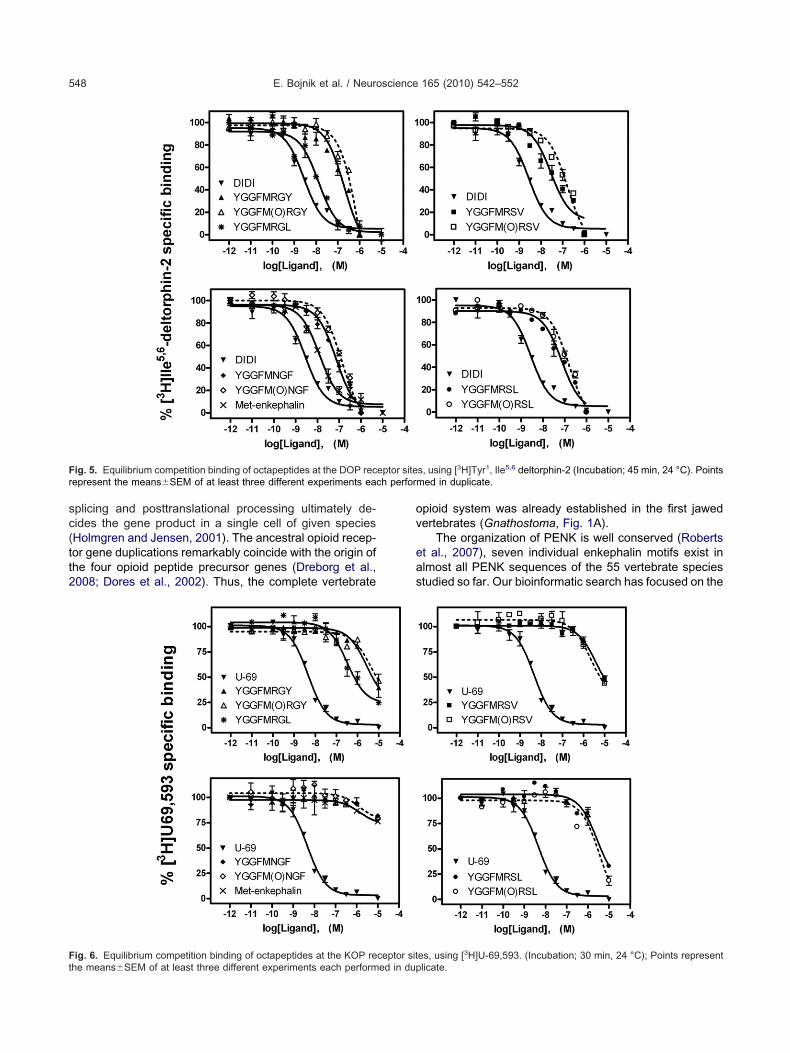
sc(tt2
ov
eas
F eptor siter h perfor
Ft
E. Bojnik et al. / Neuroscience 165 (2010) 542–552548
plicing and posttranslational processing ultimately de-ides the gene product in a single cell of given speciesHolmgren and Jensen, 2001). The ancestral opioid recep-or gene duplications remarkably coincide with the origin ofhe four opioid peptide precursor genes (Dreborg et al.,008; Dores et al., 2002). Thus, the complete vertebrate
ig. 5. Equilibrium competition binding of octapeptides at the DOP recepresent the means�SEM of at least three different experiments eac
ig. 6. Equilibrium competition binding of octapeptides at the KOP receptor sithe means�SEM of at least three different experiments each performed in dup
pioid system was already established in the first jawedertebrates (Gnathostoma, Fig. 1A).
The organization of PENK is well conserved (Robertst al., 2007), seven individual enkephalin motifs exist inlmost all PENK sequences of the 55 vertebrate speciestudied so far. Our bioinformatic search has focused on the
s, using [3H]Tyr1, Ile5,6 deltorphin-2 (Incubation; 45 min, 24 °C). Pointsmed in duplicate.
3
es, using [ H]U-69,593. (Incubation; 30 min, 24 °C); Points representlicate.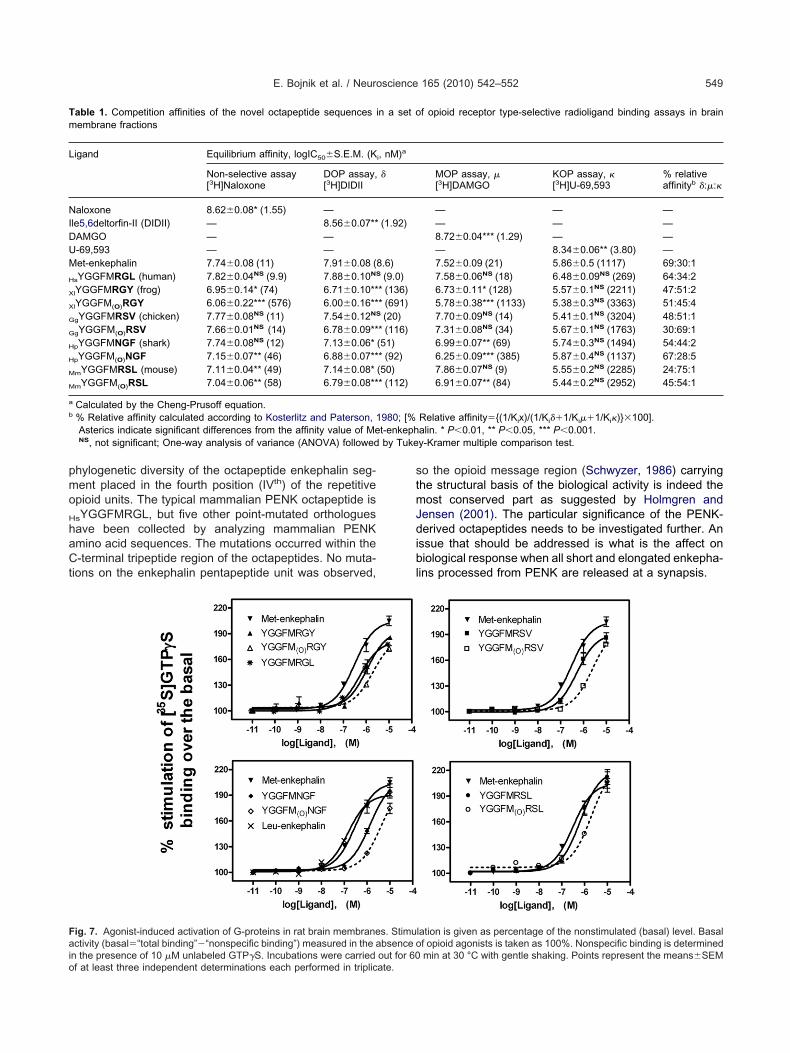
pmo
H
haCt
stmJdibl
Tm
L
NIDUM
H
X
X
G
G
H
H
M
M
a
b
by Tuke
Faio
E. Bojnik et al. / Neuroscience 165 (2010) 542–552 549
hylogenetic diversity of the octapeptide enkephalin seg-ent placed in the fourth position (IVth) of the repetitivepioid units. The typical mammalian PENK octapeptide is
sYGGFMRGL, but five other point-mutated orthologuesave been collected by analyzing mammalian PENKmino acid sequences. The mutations occurred within the-terminal tripeptide region of the octapeptides. No muta-
ions on the enkephalin pentapeptide unit was observed,
able 1. Competition affinities of the novel octapeptide sequences iembrane fractions
igand Equilibrium affinity, logIC50�S.E.M. (K
Non-selective assay[3H]Naloxone
DOP assay,[3H]DIDII
aloxone 8.62�0.08* (1.55) —le5,6deltorfin-II (DIDII) — 8.56�0.07**AMGO — —-69,593 — —et-enkephalin 7.74�0.08 (11) 7.91�0.08 (
sYGGFMRGL (human) 7.82�0.04NS (9.9) 7.88�0.10NS
lYGGFMRGY (frog) 6.95�0.14* (74) 6.71�0.10**
lYGGFM(O)RGY 6.06�0.22*** (576) 6.00�0.16**
gYGGFMRSV (chicken) 7.77�0.08NS (11) 7.54�0.12NS
gYGGFM(O)RSV 7.66�0.01NS (14) 6.78�0.09**
pYGGFMNGF (shark) 7.74�0.08NS (12) 7.13�0.06*
pYGGFM(O)NGF 7.15�0.07** (46) 6.88�0.07**
mYGGFMRSL (mouse) 7.11�0.04** (49) 7.14�0.08*
mYGGFM(O)RSL 7.04�0.06** (58) 6.79�0.08**
Calculated by the Cheng-Prusoff equation.% Relative affinity calculated according to Kosterlitz and Paterson, 1Asterics indicate significant differences from the affinity value of MeNS, not significant; One-way analysis of variance (ANOVA) followed
ig. 7. Agonist-induced activation of G-proteins in rat brain membranectivity (basal�“total binding”�“nonspecific binding”) measured in the a
n the presence of 10 �M unlabeled GTP�S. Incubations were carried out for 60f at least three independent determinations each performed in triplicate.
o the opioid message region (Schwyzer, 1986) carryinghe structural basis of the biological activity is indeed theost conserved part as suggested by Holmgren and
ensen (2001). The particular significance of the PENK-erived octapeptides needs to be investigated further. An
ssue that should be addressed is what is the affect oniological response when all short and elongated enkepha-
ins processed from PENK are released at a synapsis.
f opioid receptor type-selective radioligand binding assays in brain
MOP assay, �[3H]DAMGO
KOP assay, �[3H]U-69,593
% relativeaffinityb �:�:�
— — —— — —8.72�0.04*** (1.29) — —— 8.34�0.06** (3.80) —7.52�0.09 (21) 5.86�0.5 (1117) 69:30:17.58�0.06NS (18) 6.48�0.09NS (269) 64:34:26.73�0.11* (128) 5.57�0.1NS (2211) 47:51:25.78�0.38*** (1133) 5.38�0.3NS (3363) 51:45:47.70�0.09NS (14) 5.41�0.1NS (3204) 48:51:17.31�0.08NS (34) 5.67�0.1NS (1763) 30:69:16.99�0.07** (69) 5.74�0.3NS (1494) 54:44:26.25�0.09*** (385) 5.87�0.4NS (1137) 67:28:57.86�0.07NS (9) 5.55�0.2NS (2285) 24:75:16.91�0.07** (84) 5.44�0.2NS (2952) 45:54:1
Relative affinity�{(1/Kix)/(1/Ki��1/Ki��1/Ki�)}�100].alin. * P�0.01, ** P�0.05, *** P�0.001.y-Kramer multiple comparison test.
lation is given as percentage of the nonstimulated (basal) level. Basalf opioid agonists is taken as 100%. Nonspecific binding is determined
n a set o
i, nM)a
�
(1.92)
8.6)(9.0)
* (136)* (691)(20)
* (116)(51)* (92)(50)* (112)
980; [%t-enkeph
s. Stimubsence o
min at 30 °C with gentle shaking. Points represent the means�SEM

Aafo(psAat
M
bbsmtBpqmtsttct(aogYdoPr
fm
scqaY(LptcctVowimai(r
X
hpaaoarliciwrtsL
dpaccb(bte1tMtGf
Tm
P
ML
H
X
X
G
G
H
H
M
M
M
bs
E. Bojnik et al. / Neuroscience 165 (2010) 542–552550
Cat proenkephalin contains mutationally variant (Gly7¡la7) FcYGGFMRAL octapeptide as was prognosticated longgo (Chaminade et al., 1996). This sequence has also been
ound in the dog PENK. The recent false positive detectionf immunoreactive YGGFMRGL in the cat brainstemBelda et al., 2003) is likely due to a cross reaction of theolyclonal rabbit antisera, raised against the conjugate ofynthetic YGGFMRGL and human serum albumin, with thela7 containing cat octapeptide. Further variability existsmong the mammalian octapeptides since four other mo-ifs OaYGGFMRGY (platypus), MdYGGFMRSY (opossum),
mYGGFMRSL (mouse) and OcYGGFMRGV (rabbit) cane distinguished. The octapeptide pattern in lower verte-rates is more diverse exhibiting the same and differentequences related to the mammalian ones. Thus, theouse octapeptide MmYGGFMRSL is also present in the
wo lungfish species (Fig. 1). The amphibian Xenopus andombina octapeptides have the same structure as in thelatypus. However, the most abundant mammalian se-uence HsYGGFMRGL is apparently absent in the non-ammalian vertebrate taxa. Instead, data collected from
wo birds, four reptiles, seven amphibians and ten fishpecies reveal the occurrence of at least seven such motifshose are not found in the mammalian PENKs. The oc-apeptide structure, for instance, is GgYGGFMRSV in thehicken and also in the Florida gar fish species. An iden-ical sequence is evidently present in one of the zebrafishDanio rerio) PENKs, although here the C-terminal cleav-ge site is missed (Gonzalez Nuñez et al., 2003). Since thether, paralogous copy of the zebra fish PENK raised byene or genom duplication (Taylor et al., 2001) containsGGFM sequence directly followed by the RR endopepti-ase recognition motif in the aligned position, no maturedctapeptide enkephalin is expected in this species. TheENK octapeptide motifs are flanked by basic dipeptide
able 2. G-protein activation by opioid peptides in rat brainembranes
eptides PotencylogEC50�S.E.M(EC50, nM)
Efficacy (Emax), %stimulation�S.E.Mover the basal
et-enkephalin 6.51�0.08 (303) 205�4eu-enkephalin 6.83�0.07NS (144) 191�3
sYGGFMRGL (human) 6.22�0.07NS (594) 182�3
lYGGFMRGY (frog) 6.03�0.12NS (993) 194�6
lYGGFM(O)RGY 5.7�0.08* (1990) 186�5
gYGGFMRSV (chicken) 6.28�0.09NS (515) 191�4
gYGGFM(O)RSV 5.6�0.60** (2465) 197�4
pYGGFMNGF (shark) 5.87�0.08NS (1335) 207�5
pYGGFM(O)NGF 5.4�0.10*** (3958) 203�8
mYGGFMRSL (mouse) 6.18�0.08NS (653) 220�6
mYGGFM(O)RSL 5.7�0.12* (1979) 226�10
Asterics indicate significant differences from the potency value ofet-enkephalin. * P�0.05, ** P�0.01, *** P�0.001.NS, not significant; One-way analysis of variance (ANOVA) followed
y Tukey-Kramer multiple comparison test. %Emax values were nottatistically different.
epeats (KR or RR) in all other species, permitting the e
ormation of the mature oligopeptide by sequential enzy-atic cleavage.
Four of the 14 different octapeptide enkephalins foundo far in protein sequence databases have been chemi-ally synthesized and studied along with the human se-uence. Three novel structures are rather ubiquitous, as theyre ranged in different animals, while the fourth sequence ofGGFMNGF is solely present in a cartilaginous fish species
Fig. 1). From chemical aspects the Leu8¡Tyr8 (frog) andeu8¡Phe8 (shark) mutations are consistent with an ali-hatic to aromatic change. The Arg6¡Asn6 replacement inhe shark PENK is accompanied by the loss of the positivelyharged residue. The lungfish octapeptide NfYGGFMRSLontains a single Gly7¡Ser7 rather neutral mutation, whilehe GgYGGFMRSV peptide holds the Gly7-Leu8¡Ser7-al8 duplicate point mutations. The chemical degradationf the peptides in the presence of rat brain membranesas prevented by a combination of specific enzyme inhib-
tors as suggested by Hiranuma et al. (1997). A similarixture of amastatin, captopril and phosphoramidon wasble to greatly increase the antinociception of i.c.v. admin-
stered YGGFMRGL in rats (Kanai et al., 2002). The serineSer7) containing peptides displayed higher affinities inadioligand binding assays, while the frog sequence
lYGGFMRGY, also present in the platypus PENK, seems toave lesser affinity. The opioid receptor-type selectivityattern (Table 1, last column) supports that endogenousgonists are not by necessity the most specific ligands forparticular receptor. The binding characteristics of the
ctapeptides revealed moderate opioid agonist activitiesnd a rank order of potencies for the DOP�MOP��KOPeceptor binding sites. Predictably all compounds were theess potent in the KOP receptor selective assays compar-ng with the affinities observed in the DOP and MOP re-eptor sensitive experiments. The poor kappa binding data
ndicates that at physiological concentrations these ligandsould not be important if they make contact with the KOP
eceptor. In the G-protein activation assays all octapep-ides displayed full agonist properties attaining maximaltimulation levels (Emax) as high as in the case of Met- oreu-enkephalin (Table 2).
The oxidation of the Met5 residue considerably re-uced the affinity of the enkephalins by shifting the com-etition curves to the right (Figs. 3–6). Methionine is a keymino acid that has numerous roles in essential vital pro-esses. Moreover, methionine oxidation might be biologi-ally important during conditions of oxidative stress (Bo-rowski et al., 2008). Methionine oxidation in enkephalinsTurkall et al., 1982) generally results in the decrease iniological response, although enhanced antinociception ofhe oxidized enkephalin analogue (D-Ala2,Met5sulfoxide)nkephalin-amide was also reported (Kiritsroy et al.,983). The potency of Leu-enkephalin used as control washe highest in the [35S]GTP�S assays (Table 2), whereaset-enkephalin and the C-terminally extended octapep-
ides effectively stimulated the binding of [35S]GTP�S to
i/o proteins in rat brain membranes. Consistent with theirull agonist effect, the maximal stimulation values (% E ,
maxfficacy) of the octapeptides, even those containing oxi-

dt
Stetas(tsbsecatosrgpp
A2NOFsFa(
B
B
B
B
B
B
C
C
D
D
D
D
G
H
H
H
I
I
I
K
K
K
K
K
L
L
M
E. Bojnik et al. / Neuroscience 165 (2010) 542–552 551
ized Met5 side chain, were around two times higher overhe basal activity.
CONCLUSION
ummarizing the results, ligand binding and G-protein ac-ivating profiles of four novel, PENK-derived octapeptidenkephalin orthologues were described and compared to
hose of the well known Met-enkephalin-Arg6-Gly7-Leu8
nd the pentapeptide Met-enkephalin. Among the fourtructures tested, the “chicken type” peptide variant
GgYGGFMRSV) exhibited the highest affinities in recep-or binding assays. Importantly, these new endogenousequences represent further illustration of the chemicaliodiversity observed within the opioid peptide family. Re-earch on the non-conventional neuropeptide structures isssential from both phylogenetic and chemical-biochemi-al viewpoints. One significance of these mutationally vari-ble sequences is that they represent a natural “combina-orial” peptide library emerged by the evolution. The vari-us peptides evolved by gene mutations and naturalelection offer template sequences for structure-activityelationship studies. Inasmuch as the availability of theenome sequencing data grows rapidly, an increasing im-act of the phylogenetic and bioinformatic studies on ex-erimental biology is expected.
cknowledgments—This work was supported by a DNT-RET-004 grant (“Delalfoldi Neurobiologiai Tudaskozpont”) from theational Office for Research and Development (NKTH) and by anTKA CK-78566 grant from the National Scientific Researchund, Budapest, Hungary. E.B. is applying for the conditionalupport to young scientists provided by the Dr. Rollin D. Hotchkissoundation. Appreciation would be due to that organization if thepplicant is awarded. E.B. would like to thank to Valeria BiscottiForli, Italy) for her continuous support.
REFERENCES
aronti F, Conant KE, Giuffra M, Davis TL, Brughitta G, Iadarola MJ,Berrettini WH, Chase TN, Mouradian MM (1991) Opioid peptides inParkinson’s disease: effects of dopamine repletion. Brain Res560:92–96.
elda M, Coveñas R, Narváez JA, Aguirre JA, Gonzalez-Barón S(2003) An immunocytochemical mapping of methionine-enkepha-lin-Arg6-Gly7-Leu8 in the cat brainstem. Anat Embryol 206:399–408.
enyhe S, Szücs M, Borsodi A, Wollemann M (1992) Species differ-ences in the stereoselectivity of kappa opioid binding sites for[3H]U-69593 and [3H]ethylketocyclazocine. Life Sci 51:1647–1655.
enyhe S, Farkas J, Tóth G, Wollemann M (1997) Met5-enkephalin-Arg6-Phe7, an endogenous neuropeptide, binds to multiple opioidand nonopioid sites in rat brain. J Neurosci Res 48:249–258.
obrowski K, Houee-Levin C, Marciniak B (2008) Stabilization andreactions of sulfur radical cations: relevance to one-electron oxi-dation of methionine in peptides and proteins. Chimia 62:728–734.
ojnik E, Magyar A, Tóth G, Bajusz S, Borsodi A, Benyhe S (2009)Binding studies of novel, non-mammalian enkephalins, structurespredicted from frog and lungfish brain cDNA sequences. Neuro-science 158:867–874.
haminade M, Chelot E, DeCarvalho LP, Bochet P, Rossier J (1996)Cat proenkephalin-A does not contain the opioid octapeptide. Neu-
rochem Int 28:155–160.oveñas R, Martín F, Salinas P, Rivada E, Smith V, Aguilar LA,Díaz-Cabiale Z, Narváez JA, Tramu G (2004) An immunocyto-chemical mapping of methionine-enkephalin-Arg6-Gly7-Leu8 in thehuman brainstem. Neuroscience 128:843–859.
ores RM, Costantino D, Walnutt J, Danielson PB, Lecaude S (2001)Analyzing the radiation of the proenkephalin gene in tetrapods:cloning of a Bombina orientalis peoenkephalin cDNA. Peptides22:2021–2025.
ores RM, Lee J, Sollars C, Danielson P, Lihrmann I, Vallarino M,Vaudry H (2000) In the African lungfish Met-enkephalin and Leu-enkephalin are derived from separate genes: cloning of a proen-kephalin cDNA. Neuroendocrinology 72:224–230.
ores RM, Lecaude S, Bauer D, Danielson PB (2002) Analyzing theevolution of the opioid/orphanin gene family. Mass Spectrom Rev21:1–24.
reborg S, Sundström G, Larsson TA, Dan Larhammar D (2008)Evolution of vertebrate opioid receptors. Proc Natl Acad Sci U S A105:15487–15492.
onzalez Nuñez V, Gonzalez Sarmiento R, Rodriguez RE (2003)Characterization of zebrafish proenkephalin reveals novel opioidsequences. Brain Res Mol Brain Res 114:31–39.
iranuma T, Iwao K, Kitamura K, Matsumiya T, Oka T (1997) Almostcomplete protection from [Met5]-enkephalin-Arg6-Gly7-Leu8 (Met-enk-RGL) hydrolysis in membrane preparations by the combina-tion of amastatin, captopril and phosphoramidon. J Pharmacol ExpTher 281:769–774.
olmgren S, Jensen J (2001) Evolution of vertebrate neuropeptides.Brain Res Bull 55:723–735.
ughes J, Smith TH, Kosterlitz HW, Fothergill LA, Morgan BA, MorrisHR (1975) Identification of two related pentapetides from the brainwith potent opiate agonist activity. Nature 258:577–579.
adarola MJ, Tang J, Costa E, Yang HY (1986) Analgesic activity andrelease of [MET5]enkephalin-Arg6-Gly7-Leu8 from rat spinal cord invivo. Eur J Pharmacol 121:39–48.
adarola MJ, Ofri D, Kleinman JE (1991) Enkephalin, dynorphin andsubstance P in postmortem substantia nigra from normals andschizophrenic patients. Life Sci 48:1919–1930.
keda Y, Nakao K, Yoshimasa T, Yanaihara N, Numa S, Imura H(1982) Existence of Met-enkephalin-Arg6-Gly7-Leu8 with Met-en-kephalin, Leu-enkephalin and Met-enkephalin-Arg6-Phe7 in thebrain of guinea pig, rat and golden hamster. Biochem Biophys ResCommun 107:656–662.
anai M, Takahashi S, Kosaka K, Iwao K, Kobayashi K, Oka T (2002)[Met5]enkephalin-Arg-Gly-Leu-induced antinociception is greatlyincreased by peptidase inhibitors. Eur J Pharmacol 453:53–58.
halap A, Bagrosky B, Lecaude S, Youson J, Danielson P, Dores RM(2005) Trends in the evolution of the proenkephalin and prodynor-phin genes in gnathostomes. Ann NY Acad Sci 1040:22–37.
ilpatrick DL, Jones BN, Kojima K, Udenfriend S (1981) Identificationof the octapeptide [Met]enkephalin-Arg6-Gly7-Leu8 in extracts ofbovine adrenal medulla. Biochem Biophys Res Commun 103:698–705.
iritsyroy JA, Chan SK, Iwamoto ET (1983) Methionine oxidationenhances opioid activity of an enkephalin analog. Life Sci 32:889–893.
osterlitz HW, Paterson SJ (1980) Characterization of opioid receptormediated antinociception in nervous tissues. Proc R Soc Lond B210:113–122.
ason W, Przewłocka B, Van Luijtelaar EL, Coenen AM, Przewłocki R(1992) Endogenous opioid peptides in brain and pituitary of ratswith absence epilepsy. Neuropeptides 21:147–152.
üschen W, Buck F, Willig A, Jaros PP (1991) Isolation, sequenceanalysis, and physiological properties of enkephalins in the ner-vous tissue of the shore crab Carcinus maenas L. Proc Natl AcadSci U S A 88:8671–8675.
artens GJ, Herbert E (1994) Polymorphism and absence of Leu-enkephalin sequences in proenkephalin genes in Xenopus laevis.
Nature 310:251–254.
N
N
P
R
S
S
S
S
S
T
T
T
T
T
Z
E. Bojnik et al. / Neuroscience 165 (2010) 542–552552
evin ST, Kabasakal L, ötvös F, Tóth G, Borsodi A (1994) Bindingcharacteristics of the novel highly selective delta agonist,[3H]Ile5,6deltorphin-II. Neuropeptides 26:261–265.
oda M, Furutani Y, Takahashi H, Toyosato M, Hirose T, Inayama S,Nakanishi S, Numa S (1982) Cloning and sequence analysis ofcDNA for bovine adrenal proenkephalin. Nature 295:202–206.
ittius CW, Seizinger BR, Mehraein P, Pasi A, Herz A (1983) Proen-kephalin-A-derived peptides are present in human brain. Life Sci1:41–44.
oberts E, Shoureshi P, Kozak K, Szynskie L, Baron A, Lecaude S,Dores RM (2007) Tracking the evolution of the proenkephalin genein tetrapods. Gen Comp Endocrinol 153:189–197.
alzet M, Stefano GB (1997) Invertebrate proenkephalin: � opioidbinding sites in leech ganglia and immunocytes. Brain Res 768:224–232.
alzet M, Tasiemski A (2001) Involvement of proenkephalin-derivedpeptides in immunity. Dev Comp Immunol 25:177–185.
chwyzer R (1986) Molecular mechanism of opioid receptor selection.Biochemistry 25:6335–6342.
eizinger BR, Liebisch DC, Kish SJ, Arendt RM, Hornykiewicz O, HerzA (1986) Opioid peptides in Huntington’s disease: alterations in
prodynorphin and proenkephalin system. Brain Res 378:405–408.ollars C, Danielson P, Joss JM, Dores RM (2000) Deciphering theorigin of Met-enkephalin and Leu-enkephalin in lobe-finned fish:cloning of Australian lungfish proenkephalin. Brain Res 874:131–136.
asiemski A, Verger-Bocquet M, Cadet M, Goumon Y, Metz-BoutigueM-H, Aunis D, Stefano GB, Salzet M (2000) Proenkephalin A-de-rived peptides in invertebrate innate immune processes. Mol BrainRes 76:237–252.
aylor JS, van de Peer Y, Braasch I, Meyer A (2001) Comparativegenomics provides evidence for an ancient genome duplicationevents in fish. Philos Trans R Soc Lond B Biol Sci 356:1661–1679.
óth G, Krámer M, Sirokmán F, Borsodi A, Rónai A (1982). Prepara-tion of [7,8,19:20-3H]naloxone of high specific activity. J LabelComp Radiopharm 19:1021–1030.
raynor JR (1987) Prodynorphin as a source of [Leu] enkephalin.Trends Pharmacol Sci 8:47–48.
urkall RM, Denison RC, Tsan MF (1982) Degradation and oxidationof methionine enkephalin by human-neutrophils. J Lab Clin Med99:418–427.
amir N, Palkovits M, Brownstein M (1985) Distribution of immunore-active Met-enkephalin-Arg6-Gly7-Leu8 and Leu-enkephalin in dis-
crete regions of the rat brain. Brain Res 326:1–8.(Accepted 3 October 2009)(Available online 9 October 2009)