an updated organic classification of tyrosinase inhibitors on melanin biosynthesis
TRANSCRIPT

Send Orders for Reprints to [email protected]
4 Current Organic Chemistry, 2015, 19, 4-18
An Updated Organic Classification of Tyrosinase Inhibitors on Melanin Biosynthesis
Chung-Yi Chen1, Li-Ching Lin
2, Wen-Feng Yang
2, Jennifer Bordon
3 and Hui-Min D. Wang
2,4,*
1School of Medicine and Health Sciences, Fooyin University, Kaohsiung County 831, Taiwan, ROC;
2Department of
Fragrance and Cosmetic Science, Kaohsiung Medical University, Kaohsiung 807, Taiwan, ROC; 3Department of Psy-
chology, San Diego State University, California 92182, USA; 4Graduate Institute of Natural Products, Kaohsiung
Medical University, Kaohsiung 807, Taiwan, ROC
Abstract: Tyrosinase (EC 1.14.18.1) is known to be the rate-limiting enzyme responsible for catalyzing the first two steps
in the synthesis of melanin pigments that determine the coloring of hair, skin and eyes. Tyrosinase catalyzes two- se-
quence distinct bio-reactions in pigment productions: L-tyrosine to L-dopa (hydroxylation); and L-dopa to dopaquinone
(oxidation). The inhibition of tyrosinase has been established as one of the major strategies to regulate the production of melanin and the
inhibitors are used to treat hyper-pigmentation and other unwanted effects from melanogenesis. There are now a variety of medicaments
inhibiting melanin synthesis to treat excess melanin production or hyper-pigmentation of human skin, including the usage of natural plant
extracts or chemically synthesized compounds. Recently, due to an increasing emphasis on healthy concepts, attention has been drawn to
the usage of naturally produced crude materials and drugs in cosmetics and pharmaceutical applications. In this review, we demonstrate
melanogenic inhibitory factors such as flavonoids, stilbenes, simple phenolics, furans, triterpenes, alkaloids, and other synthetic com-
pounds. The structure-activity relationships are classified in terms of their inhibition of tyrosinase functions and copper chelating activity.
Keywords: Alkaloids, flavonoids, furans, melanin, tyrosinase inhibitor, stilbenes, simple phenolics, triterpenes.
BASIC ELEMENTS OF THE HUMAN SKIN
The human skin is a multi-faceted organ consisting of three ba-
sic layers: underlying hypodermis, dermis, and the epidermis, all of
them are separated by the basal membrane. The horny layer of the
bottom epidermis plays a photo-protective role by reflecting sub-
stantial amount of solar radiation [1, 2] The skin color of human is
determined by melanin, a biopolymer material produced by mela-
nocytes, which are special dendritic pigment cells that reside mostly
below or between the basal cells of the epidermis [3, 4]. Melano-
somes are organelles found in animal cells, and are the cellular site
of synthesis, storage and transport of pigment, synthesizing pigment
from an amino acid, L-tyrosine, via a series of biochemical conver-
sions. The significant enzyme to these transformations is the pres-
ence of tyrosinase, responsible for catalyzing the reaction of L-
tyrosine to melanin [5]. The darkening of the skin or epidermis
changes owing to melanin. Melanin takes several biochemical ap-
pearances; the color changeability of each form depends on the
degrees of oxidation. There are three basic types of melanin:
eumelanin, pheomelanin, and neuromelanin. The major type is
eumelanin, and is formed in brown and black subtypes. Pheomela-
nin is a cysteine-containing reddish-brown benzothiazine unit
polymer mostly responsible for red freckles and hair. Pheomelanin
and eumelanin, both resulting from L-dopa, are the principal cate-
gories of melanin found within human skin [3, 6]. Neuromelanin is
found in the brain, though its meaning remains unclear.
HUMAN MELANIN BIOSYNTHESIS RELATED PROTEINS
Tyrosinase is a widely used attractive target enzyme for many
business purposes [7]: in environmental applications, it has been an
*Address correspondence to this author at the Department of Fragrance and Cosmetic
Science; Graduate Institute of Natural Products, Kaohsiung Medical University, 100,
Shih-Chuan 1st Road, San-Ming District, Kaohsiung 80708, Taiwan (ROC); Tel: 886-7-3121101 ext 2804; Mobil: 886-935753718; Fax: 886-7-3136059;
E-mail: [email protected]
important component in the detoxification of contaminated soils
and phenol-containing water waste [8-10], the formation of bio-
polymers with viscoelastic functional abilities [11], the enhance-
ment of the fragrance, texture, and taste of food by the polymeriza-
tion of phenol analogues, and by the cross-linking of protein-
polysaccharide and protein-protein. Tyrosinases are bound in a
great range of cells and tissues, including bacteria, fungi, plants,
insects, and other mammalians. Tyrosinases are bio-functional on
various multiplicities of purposes: browning in plants, skin melano-
genesis in mammals, differentiation of reproductive organs, the host
defense system in arthropods, and spore developments in fungi [12-
15]. Tyrosinase could be activated by exposing to ultraviolet (UV)
light, and intervene inductively in several intermediate stages of
human skin pigment formation. Tyrosinase (EC 1.14.18.1, mono-
phenol, L-dopa: oxygen oxidoreductase) is a bi-functional copper-
containing enzyme used in molecular oxygen to react with two
distinct reactions in melanin synthesis: the hydroxylation of mono-
phenols (L-tyrosine) to the matching o-diphenols (L-dopa, mono-
phenolase) and the following oxidation to o-quinones (dopaqui-
none, catecholase) [3, 16-20]. After spontaneous reactions from
dopaquinone to dopachrome, dopachrome tautomerase (TRP-
2/DCT, tyrosinase-related protein-2) proceeds with the transforma-
tion of dopachrome to 5,6-dihydroxyindole-2-carboxylic acid
(DHICA). Afterwards, DHICA is catalyzed to indole–quinone–
carboxylic acid by DHICA oxidase (TRP-1, tyrosinase-related pro-
tein-1) [21]. Tyrosinase-related proteins, TRP-2 and TRP-1, react in
the end of eumelanin bio-conversion steps that manage the type of
eumelanin synthesized. Even though, three enzymes are known to
be implicated in melanogenesis bio-production in human, ty-
rosinase is mostly known to it regulate the quantity of pigment syn-
thesized, while other two enzymes merely adjust the type of pig-
ment productions [22]. Besides the role in melanogenesis, ty-
rosinase protein family may control the biological properties of
melanocytes to some degree in some ways [5, 23].
1875-5348/15 $58.00+.00 © 2015 Bentham Science Publishers

An Updated Organic Classification of Tyrosinase Inhibitors on Melanin Biosynthesis Current Organic Chemistry, 2015, Vol. 19, No. 1 5
MULTIPLE APPROACHES TO INHIBIT TYROSINASE ACTIVITY INCLUDING NATURAL OR SYNTHETIC AGENTS
Several reports have been published to find out the molecular and enzymatic characteristics of tyrosinase from each different resource. The literature reports reveal much difference due to the sources, especially in catalytic mechanisms [24-29]. Although mammalian and fungal tyrosinases both utilize the same substrate and produce a dark, insoluble pigment, they are distinguished as two different types of enzymes [30-32]. For example, the fungal enzyme is capable of hydroxylating estradiol of the 2-hydroxy compound and oxidizing the latter to the quinone, whereas the mammalian enzyme does not have this ability. Studies using mush-room tyrosinase have been emphasized that several aspects were significantly different from mammalian tyrosinase [33, 34]. A number of agents possessing the ability to suppress mushroom ty-rosinases did not show equivalent consequences with mammalian type enzymes [35-37].
There are several famous melanogenesis suppressors, such as kojic acid, PTU, arbutin, or hydroquinone, currently being devel-oped worldwide as cosmetic additives. But, the melanogenesis sup-pressors might stimulate human skin tumor proliferation via a high level concentration or frequent use [38-40]. There are specific dose limitations in certain countries for their acceptable applications of PTU, arbutin, or others as whitening agents in cosmetic compo-nents. Since the relative laws concerning concentration doses in each country have been strictly regulated, in order to apply these constituents as ingredients within cosmetic stuffs, manufacturers have to follow the policies.
Tyrosinase activity depends on the binding and functioning of two copper ions at the active site. Chelating copper suppressed the functions of recombinant tyrosinase, for example, by phen-ylthiourea [41], 5-hydroxy-2-(hydroxymethyl)-4-pyrone, also termed as kojic acid [42], dithiothreitol [43], by bathocuproein di-sulphonate [44], or ellagic acid [45]. Another non-competitive way of suppressing enzyme function is through a decrease in its phos-phorylation. Inhibition of tyrosinase phosphorylation could reduce tyrosinase activity via a selective protein kinase C inhibitor, bisin-dolylmaleimide [46]. Some better tolerated skin lightening sub-stances currently being used originate from natural sources, e.g.,liquorice extract (from liquorice root), ascorbic acid (vitamin C from citrus fruits) and their derivatives, as well as kojic acid (from carbohydrate fermentation medium under the effect of certain bac-teria, Aspergillus oryzae).
Maeda et al. reported that arbutin (from the leaves of the com-mon bearberry, Uvae ursi), a naturally occurring �-D-gluco-pyranoside of hydroquinone, is effective in the topical treatment of various cutaneous hyperpigmentations characterized by hyperactive melanocyte function [47]. Melanin production of cultured human melanocytes was observed to be inhibited significantly by arbutin at noncytotoxic concentrations. The IC50 values were 5.7 �� 10-3 or 1.89 ��10-2 M (maybe toxic) using L-tyrosine or L-dopa as the sub-strate. The enzyme kinetic examinations and the mechanisms for inhibition of tyrosinase ensured the reversibility of arbutin as a competitive suppressor. The bio-function whitening of arbutin in human being is involved in inhibition of the melanosome enzyme action, rather than the inhibition of the synthesis and expression of tyrosinase [47].
Earlier reports highlighted that azelaic acid and hydroquinone, both being potential remedies for symptoms of hyper-pigmentations
[48, 49], are competitive suppressors of the activity on tyrosinase, but showed cytotoxicity influences in human melanocytes [50-53]. Arbutin also competitively inhibits mushroom tyrosinase activity, but at high concentration, is cytotoxic to cultured melanocytes [47].
In this review, we classify extracted plant metabolites and some synthetic compounds in terms of their inhibitory mechanisms of tyrosinase function.
FLAVONOIDS AS TYROSINASE INHIBITORS
In plants, these compounds play an important role in self-protection against UV radiation, pathogens and herbivores. There is great potential to discover new compounds for medicinal and cos-metic additives. Flavonoids have been established to have anti-tumoral, anti-platelet, anti-ischemic, anti-allergic and anti-inflammatory effects [54]. Flavonoids have an effect on the target enzyme’s function in several diverse mechanisms. For example, they work as cofactors without inhibiting monophenolase activity, or conversely inhibit monophenolase activity without acting as a cofactor. In general, flavonoids are a group of naturally occurring antioxidants and metal chelators to copper at the tyrosinase active site forming the copper-flavonoid complexes [55, 56].
Matsuda et al. [57] used a 50% ethanolic extract obtained from the dried leaves and bark of the Myrica rubra to inhibit melanin biosynthesis in vitro. Results revealed that the extracts suppressed the tyrosinase bio-function through the conversion of L-dopa to dopachrome. Furthermore, the extracts inhibited the production of melanin from dopachrome by auto-oxidation and showed superox-ide dismutase (SOD)-like activity. Compounds, such as quercetin (1), myricetin (2) and myricetin 3-O-rhamnoside (3), were shown to have an inhibitory effect on tyrosinase activity (Fig. 1). At 50 �g/mL dose, it was reported that quercetin (1) has the best inhibi-tory effect with a decreased activity of 68%; meanwhile, com-pounds (2) and (3) both had a decrease of only 21%. Nonetheless, the demonstrated inhibitory effects on tyrosinase activity from these compounds imply their potential use in whitening agents.
Shimizu et al. [58] examined tyrosinase activities on the crude methanol extracts from the heartwood of 23 Papua New Guinean wood species. It was demonstrated that the Artocarpus incisus(Moraceae) extract showed tyrosinase inhibitory activity with com-parative strength to that of kojic acid. The extracts suppressed pig-ment biosynthesis of both cultured B16-F10 melanoma cells with no sign of cytotoxicity or skin irritation on brown guinea pig backs. They further isolated seven functional components, including a novel constitute, which has been identified as 6-(3"-methyl-1"-butenyl)-5,7,2',4'-tetrahydroxyflavone, named isoartocarpesin. Other six functional components were (+)-dihydromorin (4), chlo-rophorin (45, Fig. 4), (+)-norartocarpanone (5), 4-prenyloxy-resveratrol (46), artocarbene (47) and artocarpesin (48) (Fig. 1).According to the data, these compounds may be responsible for the melanin biosynthesis inhibitory effects due to the inhibition of ty-rosinase. Following this finding, several other flavonoids were found; stilbenes and related 4-substituted resorcinols either obtained from A. incisus, other plants, or synthesized [59]. Following this research, they isolated the prenylated flavonol, artocarpin (10),against the UV-induced hyperpigmentation of guinea pig skin [60]. The testing guinea pig skins treated with (10) were significantly whiter than those of the base-solution-treated skins. The positive control groups, kojic acid and arbutin, appeared to produce only a slightly lightening effect, and this effect was not significantly dif-ferent from that of the non-treated guinea pigs.
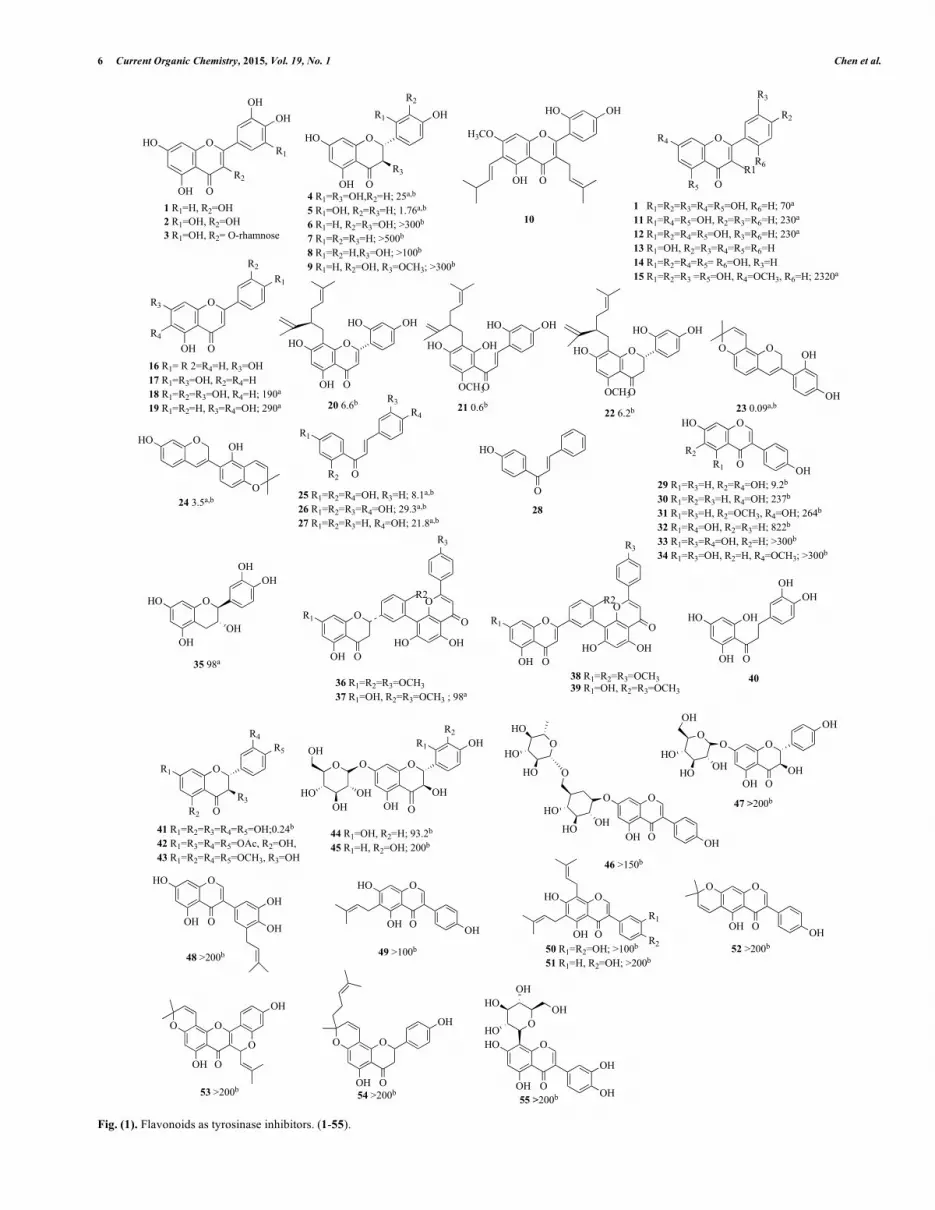
6 Current Organic Chemistry, 2015, Vol. 19, No. 1 Chen et al.
1 R1=H, R2=OH2 R1=OH, R2=OH3 R1=OH, R2= O-rhamnose
HO O
OH
OH O
OH
R2
R1
OHO
OH
OH
O
R1
R3
4 R1=R3=OH,R2=H; 25a,b
5 R1=OH, R2=R3=H; 1.76a,b
6 R1=H, R2=R3=OH; >300b
7 R1=R2=R3=H; >500b
8 R1=R2=H,R3=OH; >100b
9 R1=H, R2=OH, R3=OCH3; >300b
H3CO O
OH
OH O
HO
10
R4 O
R2
R5 O
R3
R1R6
1 R1=R2=R3=R4=R5=OH, R6=H; 70a
11 R1=R4=R5=OH, R2=R3=R6=H; 230a
12 R1=R2=R4=R5=OH, R3=R6=H; 230a
13 R1=OH, R2=R3=R4=R5=R6=H14 R1=R2=R4=R5= R6=OH, R3=H15 R1=R2=R3 =R5=OH, R4=OCH3, R6=H; 2320a
R2
R3 O
R1
OH O
R2
R4
16 R1= R 2=R4=H, R3=OH17 R1=R3=OH, R2=R4=H18 R1=R2=R3=OH, R4=H; 190a
19 R1=R2=H, R3=R4=OH; 290a
HO O
OHHO
OH O
20 6.6b
HO OH
OHHO
OCH3O
21 0.6b
HO O
OHHO
OCH3O
22 6.2b
O OH
OH
O
23 0.09a,b
HO O OH
O24 3.5a,b
R4
OR2
R1
R3
O
HO
25 R1=R2=R4=OH, R3=H; 8.1a,b
26 R1=R2=R3=R4=OH; 29.3a,b
27 R1=R2=R3=H, R4=OH; 21.8a,b28
HO O
OR2
OHR1
29 R1=R3=H, R2=R4=OH; 9.2b
30 R1=R2=R3=H, R4=OH; 237b
31 R1=R3=H, R2=OCH3, R4=OH; 264b
32 R1=R4=OH, R2=R3=H; 822b
33 R1=R3=R4=OH, R2=H; >300b
34 R1=R3=OH, R2=H, R4=OCH3; >300b
OHO
OH
OHOH
OH
35 98a
OO
R3
OHHOOH
R1
O
O
R2
36 R1=R2=R3=OCH337 R1=OH, R2=R3=OCH3 ; 98a
OO
R3
OHHOOH
R1
O
O
R2
38 R1=R2=R3=OCH339 R1=OH, R2=R3=OCH3
OHHO
OH
OHOH
O
40
O
OR3
R4R5
R1
R2
41 R1=R2=R3=R4=R5=OH;0.24b
42 R1=R3=R4=R5=OAc, R2=OH,43 R1=R2=R4=R5=OCH3, R3=OH
O
OH
O
HO OH
OHO
OHOH
OHR1
R2
44 R1=OH, R2=H; 93.2b
45 R1=H, R2=OH; 200b
O
OOH OH
O
OHHO
HO
O
O
HO
HO
HO
46 >150b
O
O
O
OH
OHO
HO OHHO
OH
OH
47 >200b
O
OOH
HO
OH
OH
48 >200b
O
OOH
HO
OH
49 >100b
O
OOH
HO
R2
R1
50 R1=R2=OH; >100b
51 R1=H, R2=OH; >200b
O
OH O
O
OH52 >200b
53 >200b
O
O
OH O
O
OH
O
OOH
OH
O
54 >200b
O
O
OH
HO
OOH
OH
HOOH
HO
OH
55 >200b
O
Fig. (1). Flavonoids as tyrosinase inhibitors. (1-55).

An Updated Organic Classification of Tyrosinase Inhibitors on Melanin Biosynthesis Current Organic Chemistry, 2015, Vol. 19, No. 1 7
Kubo et al. [61] also reported (1) as the principal tyrosinase in-
hibitor from the dried flower of Heterotheca inuloides Cass (Com-positae). This ordinary flavonol was also purified as the main ty-rosinase suppressor from just picked flowers of Trixis michuacana var longifolia (D. Dow) C. Anderson (Compositae), recognized as “huipate floreado” in Mexico. Later, they further assayed two groups of compounds for tyrosinase inhibition. Tyrosinase inhibi-tory activity of flavonols (competitive inhibition), (1), galangin (11), kaempferol (12, ID50, 0.23 �M), 3-hydroxyflavon (15), morin (16) and rhamnetin (17) were observed from their ability to chelate coppers (Fig. 1). On the contrary, the related flavones, chrysin (16), apigenin (17), luteolin (18) and baicalein (19) did not chelate cop-per in this enzyme. In addition, a pre-incubation examination of tyrosinase with the presence of 0.07 �M of (1), or with the absence of L-dopa enhanced the inhibitory influence from 45% to 77%. To compare with (1), its 3-O-glycosides, isoquercitrin, quercetin 3-O-glucoside, rutin or quercetin 3-O-rutinoside, were discovered to act neither as substrates nor inhibitors [16, 44]. These flavonols affect tyrosinase function by means of various mechanisms. For example, (1) played the role as a cofactor and did not suppress monopheno-lase activity. In contrast, (11) suppressed monophenolase bio-purpose and did not act as a cofactor. Moreover, Kaempferol (12) neither acted as a cofactor nor suppressed monophenolase action. Nevertheless, these three above mentioned flavonols did not have the ability to restrain diphenolase bio-movements by chelating cop-per.
Kim et al. [62] displayed the ethanol extract and dichloro-methane fraction from Sophora flavescens to have significant inhi-bition against mushroom tyrosinase. From the dichloromethane fraction, three known prenylated flavonoids, sophoraflavanone G (20), kuraridin (21) and kurarinone (22), were isolated (Fig. 1). Compared with kojic acid (IC50 = 20.5 �M), these compounds in-hibited more potent tyrosinase inhibitory activity with IC50 values of 6.6, 0.6, and 6.2 �M for (20-22), respectively.
Nerya et al. [63] discovered the licorice extract with the inhibi-tion ability to tyrosinase function, which was better than expected due to the amount of glabridin (23) within the extracts (Fig. 1). Amongst these pigmentation-whitening compounds, 1,4-dihydro-quinone is one of the most broadly approved [6]. It had been dem-onstrated as a poor substrate for tyrosinase, competing for L-dopa oxidation in the reaction site, and also causing reversible inhibition of cellular metabolism by affecting both DNA and RNA synthesis. Therefore, these results suggest that 1,4-dihydroquinone had a potential melanocyte cytotoxicity component with a history of inducing mutations.
Yokota et al. [64] presented advanced results that erythema and UV-B-induced pigmentation were repressed by topical uses of 0.5% (23) (Fig. 1). The major disadvantage of (23) is its low derma-penetration capability and formulation instabilities. An isoflavene, glabrene (24), isolated from licorice roots [65], had two separated hydroxyl groups at the 2' and 7 positions, a 2,2-dimethyl-�-pyran ring fused to the B ring, and a double bond between carbon atoms 3 and 4 in the C ring, which gave maximal conjugation of the double bonds of this target compound. The conjugation increased the steadiness of the phenoxyl radical which was produced after the donation of an electron, resulting in amplified antioxidant achieve-ments [65, 66]. Inhibitory consequences pointed out that (24) and isoliquiritigenin (2',4',4-trihydroxychalcone, 25) within the licorice extracts blocked both mono- and di-phenolase tyrosinase bio-functions [65]. When L-tyrosine was applied as a substrate, the influences of (24) and (25) on inhibitory activities were dose-
dependent in melanocytes, and IC50 values for both compounds were 3.5 and 8.1 �M, respectively. This was the earliest study to show that (24) and (25) exerted different levels of suppression in tyrosinase-dependent melanogenesis.
Nerya et al. [67] also tested nine mono-, di-, tri-, and tetra-hydroxychalcones as suppressors of mono- and di-phenolase, pre-senting that the major aspect in their efficiency was the position of the hydroxyl groups located in both aromatic rings, with a notewor-thy action to a 4-substituted B ring, more than the substituted A ring. Neither the number of hydroxyls nor the presence of a catechol moiety on ring B was associated with the rising potency of tyrosinase suppression. Isoliquiritigenin (25), butein (26) and 4-hydroxychalcone (27) decreased tyrosinase activity and reduced the lag period of enzyme monophenolase function (Fig. 1). How-ever, 4’-hydroxychalcone (28) showed no inhibition properties. Butein (26) exhibited competitive inhibition while (25) and (27) exhibited semi-competitive inhibition. The work established that chalcones were potential de-pigmentary components, exemplified in their dual outcomes of reduction and anti-oxidative actions.
Chang et al. [68] found 6,7,4�-trihydroxyisoflavone (29), identi-fied as a potent tyrosinase inhibitor (Fig. 1). This was reported to have reduced mushroom tyrosinase activity with an IC50 value of 9.2 �M, stronger than the well-established anti-tyrosinase agent of kojic acid (IC50 = 54.4 �M). When L-tyrosine was applied as the substrate, enzyme kinetic examinations indicated that (29) was a competitive suppressor to tyrosinase. It was suggested that hydroxyl groups at the C-6 and C-7 positions of the isoflavone skeleton might play an important role in tyrosinase inhibitory activity. The biosynthesis precursors and analogs, including daidzein (30), gly-citein (31), and genistein (32), showed little effects in anti-tyrosinase.
Cheng et al. [69] isolated a new biflavonoid, 2,3-dihydro-4’,4’-di-O-methyl- amentoflavone (37), and five known compounds: (18), (-)-catechin (35), 2,3-dihydrosciadopitysin (36), sciadopitysin (38) and isoginkgetin (39), separated from Podocarpus macrophyl-lus var. macrophyllus (Podocarpaceae) (Fig. 1). These compounds were evaluated by their ability to restrain cellular tyrosinase activity and their melanin inhibitory activity in human epidermal melano-cytes (HEMn). When the inhibitor’s concentration was 100 �M, (35), (37) and (39) showed little toxicity in HEMn cells (over 80% viability). In the melanin synthesis, (37) showed a potent anti-tyrosinase effect with IC50 = 0.098 �M in HEMn.
Lin et al. [70] isolated thirteen components from Formosan ap-ples, which were then identified and assessed for their free radical scavenging properties and cellular tyrosinase inhibitory abilities in HEMn. Within the purified compounds, 3-hydroxyphloretin (40) and catechol (79, Fig. 5) were potentially the most interesting (Fig. 1). Both (40) and (79) decreased tyrosinase functions in HEMn cells. To compare with the positive group, arbutin, (40) and (79) were more potential at around fifty folds. In addition, (40) made marginal diminution in cellular proliferation (about 20%, at 100 �M) and had a marginal decrease in melanin amount (about 20%). (79) influenced the intensities of related gene expressions and TRP-2 protein at 50 and 100 �M [70]. Therefore, the decrease in melanin production by (34) and (79) might not be recognized to direct ty-rosinase suppression, but the cellular cytotoxicity. Moreover, these two efficient agents reacted to the di-copper active domain area of tyrosinase, according to molecular docking analysis. Decreasing of tyrosinase function by above agents perhaps due to the competitive inhibition from the suppressors binding into the tyrosinase active site [71].

8 Current Organic Chemistry, 2015, Vol. 19, No. 1 Chen et al.
In the study by Mitsuo Miyazawa and Naotaka Tamura [72], (2R,3R)-+-taxifolin(41, 42, 43) was purified via activity-guided fractionations from the sprouts of Polygonum hydropiper L. (Beni-tade) (Fig. 1). The inhibition efficiency of mushroom tyrosinase was reported to be higher than arbutin and was equal to kojic, with an ID50 of 0.24 �M.
Zong-Ping Zheng et al. [73] screened effective extracts to mushroom tyrosinase and discovered that the twigs of Cudrania tricuspidata had a potent inhibition of IC50=113.5 �g/mL. They further isolated 28 compounds, and three among them, trans-dihydromorin (4), oxyresveratrol (58) and steppogenin (5), show-cased IC50=21.54, 2.85 and 2.52 �M, respectively (Fig. 1). Oxyres-veratrol (58) and protocatechuic acid (89) belonged to stilbenes. According to the structure–activity relationship (SAR), they pointed out the hydroxyl group substitutions at 2 and 4 positions of aro-matic ring from phenolic compounds might play a vital role in in-hibiting tyrosinase activity.
STILBENES AS TYROSINASE INHIBITORS
Iida et al. [74] tested thirty-three crude drug extracts to examine tyrosinase inhibitory activities. Among them, the acetone extract from the rhizomes of Rheum officinale Baillon showed the strongest inactivity. Tyrosinase suppressive activity-guided fractionation and chemical analysis led to the purification of two potent components, 3,4',5-trihydroxystilbene -4'-O-b-D-(2"-O-galloyl)glucopyranoside (56) and 3,4',5-trihydroxystilbene-4'-O-b-D- (6"-O-galloyl)gluco-pyranoside (57) (Fig. 2). The inhibitory effects of (56) and (57) were: IC50 = 6.71 �M and 14.7 �M (L-tyrosine); IC50 = 24.6 �M and 82.3 �M (L-dopa). The inhibitory effects of 56 and 57 were IC50 = 6.71 �M (14.7 �M L-tyrosine) and IC50 = 24.6 �M (82.3 �M
L-dopa) respectively. Both compounds illustrated competitive inhi-bition mechanisms against tyrosinase and played a role in the inhi-bition of melanin biosynthesis.
Shin et al. [75] studied a naturally occurring compound found in Morus alba, L. oxyresveratrol (2,3�,4,5�-tetrahydroxystilbene (58)), which exhibited potent suppressive influences on L-dopa oxidase function of tyrosinase (Fig. 2). Component (58) with con-centration ranging from 0.3 to 5 �M exhibited potent and dose-dependent inhibitory properties on the target enzyme activities (25 to 84%), where IC50 was approximately 1 �M. When L-dopa was used as the reaction substrate, oxyresveratrol (58) appeared to re-duce the L-dopa oxidase action in a non-competitive behavior (Ki = 9.1 � 10�7 M), and demonstrated a 150-fold more potentially sup-pressive outcome than (3,4�,5-trihydroxystilbene) (59, resveratrol). With greater number of hydroxy groups within the hydroxystil-benes being methylated to methoxy groups (60-63), the lesser re-strained effect in tyrosinase role was revealed. Consequences indi-cated that both positions and numbers of hydroxy groups within (58) seemed play a critical role in the application to the suppressive effects on L-dopa oxidase reaction on mushroom tyrosinase. Kim et al. [76], continuing these works, confirmed that (58) had a 32-fold stronger inhibition than kojic acid. Studies on the inhibitory kinetic and mechanism of mushroom tyrosinase exhibited a reversi-bility of (58) as a noncompetitive inhibitor with L-tyrosine as the substrate. The interaction between (58) and mushroom tyrosinase
displayed a high affinity reflected in a Ki value of 3.2-4.2 � 107 M. It did not affect the promoter activity of the tyrosinase gene in mur-ine melanoma B16 at 10 and 100 �M, respectively. Therefore, de-pigmentation worked in the course of reversible suppression of the enzyme function rather than through the inhibition of the synthesis and expression of tyrosinase. Hydroxystilbene compounds of (59),
OR2O
HOHO
OR1
OOH
OHOH
OH
OH
CO
OH
OH
OH
CO
56 R1=
57 R1=H R2=
R2=H; 6.71b, 24.6a
; 14.7b, 82.3a
R3
R1
R6
R8
58 R1=R3=R6=R8=OH, R2=R4=R7=H; 1a
59 R1=R2=R4=R5=R7=H, R3=R6=R8=OH; 155a
60 R1=R2=R4=R5=R7=H, R3=OCH3, R6=R8=OH; 252a
61 R1=R2=R4=R5=R7=H, R3=R6=OCH3, R8=OH ; 490a
62 R1=R2=R4=R5=R7=H, R3=R6=R8=OCH3; >500a
63 R1=R2=R4=R5=R7=H, R3=R6=OH, R8=OGlc; >500a
64 R1=R3=R5=R6=H, R2=R4=R7=OH, R8=OCH3; >100b
65 R1=R3=R5=R6=H, R2=R4=OH, R7=OGlc, R8=OCH3; 40.1b, >100e
70 R1=OH, R2=R3=R4=R5=R6=R7=R8=H; >200.0a,e
71 R2=OH, R1=R3=R4=R5=R6=R7=R8=H; >200.0a,e
72 R1=R2=OH, R3=R4=R5=R6=R7=R8=H; >200.0a,e
73 R2=R3=OH, R1=R4=R5=R6=R7=R8=H; >200.0a,e
74 R2=R4=OH, R1=R3=R5=R6=R7=R8=H; >200.0a,e
75 R2=R6=OH, R1=R3=R4=R5=R7=R8=H; >200.0a,e
76 R2=R3=R6=OH, R1=R4=R5=R7=R8=H; 87.7a,e
77 R2=R3=R6=R7=OH, R1=R4=R5=R8=H; 29.1a,e
78 R1=R3=R6=R8=OCH3, R2=R4=R5=R7=H; >100a
79 R1=OCH3, R2=R4=R5=R7=H, R3=R6=R8=OH; >100a
80 R1=R6=OCH3, R2=R4=R5=R7=H, R3=R8=OH; >100a
81 R1=R8=OH, R2=R4=R5=R7=H, R3=R6=OCH3; >100a
83 R1=R7=R9=OH, R2=R4=R5=R6=R8=H, R3=OXyl84 R1=R3=OXyl, R2=R4=R5=R6=R8=H, R7=R9=OH85 R1=R7=R9=OH, R3=OGlc, R2=R4=R5=R6=R8=H86 R1=R7=R9=OH, R3=OCel, R2=R4=R5=R6=R8=H87 R1=R3=R7=R9=OH, R2=R4=R5=R6=R8=H88 R1=OXyl, R2=R4=R5=R6=R8=H, R3=R7=R9=OH
HO
OH
OH
OH
CH3
R
66 R=CH3; 0.26b
67 R=
CH3
CH3 ; 0.66b
R2
R4
R5
R7
HO
OH
OH
OCH3
CH3 O
OH
HO OH
68 2.45b
69 2.45a,b
HO
OH
OH
OH
82 1.6a
OHHO
OH
O
89 >500b
Fig. (2). Tyrosinase inhibitors obtained from Stilbenes. (56-89).

An Updated Organic Classification of Tyrosinase Inhibitors on Melanin Biosynthesis Current Organic Chemistry, 2015, Vol. 19, No. 1 9
3,5-dihydroxy-4'-methoxystilbene and rhapontigenin (64) presented higher than 50% suppression with 100 �M in mushroom tyrosinase assay platform, but other methylated or glycosylated hydroxystil-benes of 3,4'-dimethoxy-5-hydroxystilbene, trimethylresveratrol,
piceid and rhaponticin (60-63, 65) did not reduce notably. None of
these hydroxystilbene components, excluding (58), revealed higher than 50% reduction with 100 �M in L-tyrosine oxidation on murine tyrosinase assay.
Ohguchi et al. [77] synthesized several kinds of hydroxystil-bene derivatives, which were examined in suppressive results against murine tyrosinase assay examinations (70-77) (Fig. 2). The suppressive powers of the hydroxy-trans-stilbene derivatives were outstandingly prominent through increasing the number of phenolic hydroxy substituents. Methylated hydroxy-trans-stilbene did not possess inhibition ability; besides, hydroxy-cis-stilbene or hydro-genated hydroxystilbene possessed almost no suppression ability compared to hydroxy-trans-stilbene. SAR illustrated that phenolic hydroxy functional groups and trans-olefin structures of the parent stilbene scaffold confirmed the contribution to the reduction po-tency of hydroxystilbene in tyrosinase function.
Likhitwitayawuid et al. [78] prepared seven stilbene deriva-tives, including trans-2,4,3’,5’-tetramethoxystilbene (78), trans-2-methoxy-4,3',5'-trihydroxystilbene (79), trans-2,3'-dimethoxy-4,5'-dihydroxystilbene (80), trans-4,3'-dimethoxy-2,5'-dihydroxystil-bene (81), trans-2,4,3',5'-tetramethoxystilbene (82) and cis-2,4,3',5'-tetramethoxystilbene, 2,4,3',5'-tetrahydroxybibenzyl, and 2,4,3',5'-tetramethoxybibenzyl (Fig. 2). Compound (82) is a hydrogenation compound of (78), which showed a higher potential of tyrosinase functional reduction than the parent type, with no toxicity. The enzyme kinetic work depicted that (82) was a reversible and mixed-type inhibitor to mushroom tyrosinase when L-dopa was the sub-strate. Examination of the values of Ki suggested that (82) had a moderate affinity to tyrosinase than (78). A tetra-O-methylated analog of (78), (81), with cis-configuration, was poor to suppres-sion in tyrosinase, but confirmed extremely high cytotoxicities to the human tumor cells BC, KB and NCI-H187; Compound (81) presented potency comparable to that of anti-tumor agents, ellip-ticine and doxorubicin. Results from tyrosinase suppression and cytotoxicity of compounds (78-82) indicated that O-methylation on (78) destroyed anti-tyrosinase activity but generated cytotoxicity.
Reiko et al. [79] synthesized six bibenzyl glycosides derivatives (83-87) all that showed significant tyrosinase inhibitory activity
(Fig. 2). Bibenzyl xyloside (88) is a considerably potent tyrosinase inhibitor with an IC50 value of 0.43 �M. In general, all six bibenzyl glycosides derivatives were demonstrated to be more potent ty-rosinase inhibitors than kojic acid. As the bibenzyl glycosides in-cluded bulky groups, results in the research pointed out that there might be a hydrophilic cavity as the catalytic centre for tyrosinase.
SIMPLE PHENOLIC COMPOUNDS AS TYROSINASE IN-HIBITORS
Kubo et al. [61] isolated anacardic acids, 2-methylcardols, and cardols from various parts of cashew (Anacardium occidentale, Anacardiaceae) fruit, which have been found to exhibit tyrosinase inhibitory activity. Kinetic studies with the two principally active compounds, 5-[8(Z),11(Z),14-pentadecatrienyl]resorcinol (90) and 6-[8(Z),11(Z),14-pentadecatrienyl] salicylic acid (93) have indi-cated that both of these phenolic compounds exhibited competitive inhibition of the oxidation of L-dopa by mushroom tyrosinase (Fig. 3). The other compounds with different unsaturated bonds (90-92;93-96; 97-99) listed in Figure 5, demonstrated weaker inhibition properties.
Maeda et al. [47] isolated the famous compound, arbutin (100), from the fresh fruit of California buckeye, Aesculus californica (Spach) Nutt. Hippocastanaceae has been widely used as a whiten-ing agent in cosmetic ingredients (Fig. 3). In the study conducted by Funayama et al. [32], compound (100) was reported to show a dose-dependent inhibitory effect on the oxidation of L-dopa cata-lyzed by mushroom tyrosinase with IC50 of 10 �M, and this inhibi-tion was described as competitive inhibition. It also revealed the differences in the inhibitory effects between the mushroom and human tyrosinase. It was shown that the effect of mushroom inhibi-tion was half as effective as the human type. Similarly, Hori et al. [80] found that (100) itself was oxidized as a monophenol substrate at an extremely slow rate, and this oxidation was accelerated as soon as catalytic amounts (0.01 �M) of L-dopa became available as a cofactor. They synthesized a series of arbutin analogs (102-104). The result observed was supported by monitoring oxygen consump-tion, and the depigmenting mechanism of (100) reported if a cofac-tor is not available in the melanocytes. The combination with L-ascorbic acid is a useful application, particularly when oxygen is limited.
Kim et al. [81] purchased 4,4�-dihydroxybiphenyl (105) from a chemical Company to investigate the in vitro inhibition of ty-
RHO
OH
HO
COOH
R HO RR OH
OH
OH
OHHO
R OH
HO
OHOH
HO
OHOH
OH
90 R= C15:3; 40a
91 R= C15:2; 50a
92 R= C15:1; 80a
93 R= C15:3; 1100a
94 R= C15:2; 6000a
95 R= C15:1; 7500a
96 R= C15:0; 4200a
97 R= C15:3 98 R= C15:2 99 R= C15:1,
OOH
HOO OH
OH
100 R=
Mushroom: 10000a
Human: 5700b,18900a 101 0.396b £gg/ml
OOH
HOO OH
OH100 R=
102 R= OH103 R= OCH3104 R= CO2CH2CH2CH3
105 1.91b 106 0.07b107 22d
Fig. (3). Simple phenolic compounds as Tyrosinase inhibitors. ( 90-107).

10 Current Organic Chemistry, 2015, Vol. 19, No. 1 Chen et al.
rosinase, and in vivo melanin biosynthesis was assessed in B16 cells (Fig. 3). The results showed that (105) exhibited strong anti-tyrosinase activity with IC50=1.91 M. Enzyme kinetic investiga-tions of tyrosinase suppression discovered that (105) proved to be a competitive agent (Ki = 4.0�10�4 M at 2.5 M; and Ki = 2.1�10�5 M at 5 M). Furthermore, results from melanogenesis indicated that the production contents of melanin were evidently inhibited by (105). Ha et al. [82] synthesized 4-(6-hydroxy-2-naphthyl)-1,3-bezendiol (106), a novel series of hydroxyl substituted phenyl naph-thalenes, as the isostere derivatives of (58). Compound (106) re-duced mushroom tyrosinase with the value of IC50, 0.07 M, which was shown to be more potent than the kojic acid (IC50 = 38.24). Enzyme kinetic evaluations of inhibitory mechanisms on tyrosinase suppression described that (106) was a competitive suppressor (Ki = 4.78�10-9 M at 0.125 M and Ki = 6.21 � 10-9 M at 0.25 M). Moreover, tyrosinase activity and melanin production were sup-pressed by (106) in B16 F10 cells. The consequences indicated that (106) reduced melanogenesis through the decrease in tyrosinase function.
OR1
R2
108 R1=CHO, R2=CH2-CH2-OH; 980a,b
109 R1=CHO, R2=H; 350a,b
110 R1=CHO, R2=CH3; 760a,b
111 R1=COOH, R2=H; ND Fig. (4). Tyrosinase inhibitors obtained from Furans. (108-111).
FURANS AS TYROSINASE INHIBITORS
Sharma et al. [83] extracted 5-(hydroxymethyl)-2-furfural, (108), by methanolic phase of Dictyophora indusiata (Fig. 4). The function of (108) demonstrated inhibition in a dose-dependent manner of the oxidation of L-dopa in mushroom tyrosinase assay, and the value of ID50 was measured as 0.98 M. Enzyme kinetic mechanism works exposed (108) as a non-competitive suppressor in the L-dopa oxidation. They applied other commercially available chemistry-scaffold similar derivatives of (108), such as furfural (109), 5-methylfurfural (110) and furoic acid (111) to gain more
insight into the SAR. Lineweaver-Burk plots of (109) and (110)suggested a non-competitive suppression model with respect to L-dopa. The status of instability of Schiff’s base is the major cause of lower suppression demonstrated from (108) and (110). Remarkably, (111) possessed no suppression, which might be due to the capacity to form Schiff’s base with the functional amino group within ty-rosinase. Thus, the simplicity of (109) structure showed a higher suppressive potential than other analogs.
TRITERPENES AS TYROSINASE INHIBITORS
Um et al. [84] synthesized several compounds, such as glycyr-rhetinic acid (112) and examined the in vitro mushroom tyrosinase functions (Fig. 5). In these compounds, (112) derivatives, (113-115) suppressed L-dopa oxidation of tyrosinase; the parental (112) and other analogs including commercial products, such as stearyl gly-cyrrhetinate were inactive. SAR evaluation indicated that both ke-tone and piperidine functional groups were important to the inhibi-tion of (112) analogs in tyrosinase function. It was also revealed that the suppression was conducted in a time-dependent manner, pointing out that these derivatives were slow-binding suppressors as described by other agents like m-coumaric acid and L-mimosine. The widely applied pigment-lightening compounds in cosmetic ingredients, kojic acid and vitamin C, powerfully suppressed ty-rosinase function for 10 and 20 min intervals, respectively. But it was demonstrated that the function harshly decreased as time passed by, suggesting that these agents were not quite stable during the extended reaction. Within the triterpene analogs, (114) was the most powerful one with the IC50 value of 35 M at 120-min reac-tion. IC50 values of analogs (113) and (115) were 70 and 100 M, respectively. In addition, time-course experiments indicated that (114) was much more stable than kojic acid and vitamin C .
Meade et al. [85] isolated a melanogenesis suppressor from Ar-nica montana L. (Compositae)’s flowers, and through 1D, 2D-NMR and liquid chromatography/high-resolution mass spectrome-try (LC-HR-MS) to identify it as a taraxastene-type triterpene (3�,16�-di- hydroxy-21�-hydroperoxy-20(30)-taraxastene (116) (Fig. 5). In melanoma cultured B16 cells, (116) at a dose of 0.53 M totally suppressed melanin production. This was one of the most potential inhibitors among the recognized natural plant sup-pressors of melanogenesis in cell based assays, being 50 times
R
HO
X
HO
OH
COOH
112 X=O, R=COOH; 500a
113 X=O, R=
NHNOC
114 X=O, R= HNOC NNH
O
115 X=H2, R= HNOC NNH
O
116 0.04e
; 70a
; 35a
; 100a
Fig. (5). Triterpenes as Tyrosinase inhibitors. (112-116).
An Updated Organic Classification of Tyrosinase Inhibitors on Melanin Biosynthesis Current Organic Chemistry, 2015, Vol. 19, No. 1 11
higher than 4-methoxyphenol, which was applied as a skin whiten-ing compound. It was considered that the mechanism was related to the down-regulation of melanocyte-type isoform of mi-crophthalmia-associated transcription factor (MITF-M, a transcrip-tional factor), which made a reduction of tyrosinase and related genes and proteins. The mRNA expression of these melanogenic proteins, TRP-1, TRP-2 and PMEL17, was inhibited in the presence of (116) and these inhibitions were statistically significant.
ALKALOIDS AS TYROSINASE INHIBITORS
Shaheen et al. [86] obtained lycoctonine-type norditerpenoid alkaloid, swatinine, along with four known norditerpenoid alka-loids: delphatine, lappaconitine (117), puberanine (118), and N-acetylsepaconitine, which were isolated from the aerial parts of Aconitum laeve Royle (Fig. 6). Only (117) and (118) (IC50 = 93.33 M and 205.21 M) exhibited mild inhibition against tyrosinase. Puberanine (118) also had anti-inflammatory activity. Regarding anti-oxidant activities, only swatinine (54.1%) and delphatine (55.4%) scavenged DPPH radicals.
AMIDES AS TYROSINASE INHIBITORS
Seon-Yeong Kwak [87] conjugated hydroxycinnamic acid ana-logs with phenylalanyl hydroxamic acid and prolyl hydroxamic acid for a series of tyrosinase inhibitors (119-129) (Fig. 7). Pheny-lalanyl hydroxamic acid showed 5–20% higher tyrosinase inhibition activity than prolyl hydroxamic acid. Cinnamic acid analogs dem-onstrated potent mushroom tyrosinase inhibition might be from the substrate similarity, also effective in Mel-Ab cells in melanogene-sis. Cinnamic acid analogs, possibly due to similarities with the tyrosinase substrate, demonstrated potent mushroom tyrosinase inhibition. Cinnamic acid was effective in inhibiting Mel-Ab cell melanogenesis.
MISCELLANEOUS COMPOUNDS AS TYROSINASE IN-HIBITORS
Morisaki et al. [88] synthesized hybrid L-ascorbic acid deriva-tives (vitamin C analogs) with the active biologically substances: �-tocopherol (vitamin E) and 5-hydroxy-2-hydroxymethyl-b-
Fig. (6). Tyrosinase inhibitors obtained from Alkaloids. (117-118).
HO
HO
NH
OHN
O
OH
HO
HO
N
O
O
NHHO
NH
O
R3
R2
R1HN
O
OH
120 CA:R1=R2=OH, R3=H; 4.9a
121 pCoA: R1=R3=H, R2=OH122 FA: R1=OCH3, R2=OH, R3=H123 SA: R1=R3=OCH3, R2=OH
R1
R2
R3
N
O
O
NHHO
125 CA:R1=R2=OH, R3=H126 pCoA: R1=R3=H, R2=OH127 FA: R1=OCH3, R2=OH, R3=H128 SA: R1=R3=OCH3, R2=OH
H2N
O
NH
O
O
O
OH
OCu (II)
Cu (II)129 4.2a
119124
Fig. (7). Amides as Tyrosinase inhibitors. (119-129).
OCH3OH
OH
O
N
OCH3OCH3
HN
O
Ac
OCH3OH
OHN
OCH3OCH3
OH
OHOCH3HO
117 93.33a118 205.21a

12 Current Organic Chemistry, 2015, Vol. 19, No. 1 Chen et al.
pyrone (kojic acid) linked to the C-2 or C-3 hydroxyl group. They discovered that as a hydrophilic analog, 2-O-(5-hydroxy-4H-pyran-4-one-2-methyl)-L-ascorbic acid (130), showed superior inhibitory activities and thermal stabilities to suppress tyrosinase-catalyzed pigment production and free radical synthesis compared to vitamin C, its conventional analogs (such as the 2-O-octadecylascorbic acid, 2-phosphate 6-stearate and 2.6-dipalmitate), arbutin, kojic acid, and vitamin E. It was obvious that (130) had both biochemical abilities of kojic acid and vitamin C, and performed in a synergistic way (Fig. 8). Hydroxyl functional groups at C-5 position of the kojic acid moiety domain, and C-3 position of the vitamin C moiety area biochemical properties biochemical properties. It was thought that the functional group moiety of kojic acid in (130) counterbalanced the attenuation of the inhibitory action due to the shielding of the biochemical vital C-2 hydroxyl group of the vitamin C moiety do-main. Moreover, the thermal stabilities were drastically enhanced not only in vitamin C but also in kojic acid. A lipophilic analog, 3-O-[(alpha-tocopheryloxy)-2-hydroxypropyl]-L-ascorbic acid (131), was far more stable than vitamin C and the typical lipophilic analogs. (131) displayed almost the same suppressive properties to inhibit melanogenesis and free radical productions as typical lipo-philic analogs, even though these biochemical properties of (131) were lower than those of vitamin C.
Lee et al. [89] investigated the in vitro influences of the 85% methanol extract of Morus alba dried leaves in melanogenesis, which was closely related to inhibit the hyperpigmentation. These extracts reduced the function of tyrosinase that transferred L-dopa to dopachrome in the biological developments of pigment. Mulber-roside F (132, moracin M-6, 3'-di-O-b-D-glucopyranoside), ac-quired from the extracts after the activity-guided fractionation, ex-posed suppressive influences on tyrosinase function and in melano-genesis on melan-a cells (Fig. 8). (132) presented the value of IC50 = 50.29 mg/ml with kojic acid as the positive control (IC50 = 51.30 mg/ml) in the tyrosinase reduction capacity platform assay. Cellular tyrosinase function in the (132) treated HM3KO cell line also re-duced in a manner correlating to mushroom type enzyme. Mammal-ian tyrosinase function was inhibited by (132) to 50% at a dose of 68.3 mg/ml, and was much higher than mushroom tyrosinase in the value of IC50.
Masamoto et al. [90] provided a tyrosinase inhibitor, esculetin (133), from the seeds of Euphorbia lathyris L. by bioassay-guided fractionation and purification, using silica gel column chromatogra-phy (Fig. 8). The IC50 value of esculetin in the mushroom ty-rosinase activity test was 43 M. The kinetic study indicated that esculetin exhibited competitive inhibition against the oxidation of L-dopa. The SAR among five (133) analogs (coumarin (134), um-belliferone (135), esculin (136), and scopoletin (137) from Aldrich Chemical Co.) suggested that hydroxyl groups at the C6 and C7 positions of the coumarin skeleton played an important role in the expression of tyrosinase inhibitory activity.
Kanehira et al. [91] produced kinobeon A (138) from the me-dicinal plant cell cultures, safflower (Carthamus tinctorius L., Compositae) (Fig. 8). Mushroom tyrosinase activity was inhibited in a concentration-dependent manner when treated with (138) using L-tyrosine or L-dopa as reaction substrates. The values of IC50 were 22 M (substrate: L-tyrosine) and 27 M (L-dopa), respec-tively. In addition, suppression in human melanoma HMVII cellular tyrosinase function increased with the increasing treated doses of (138) applying L-dopa as the reaction substrate to show the value of IC50, 2.5 M. It was also found that Kinobeon A (138) was a high potential competitive suppressor than L-ascorbic acid, arbutin
or kojic acid to both mushroom and human tyrosinase as verified in Lineweaver-Burk method.
Kim et al. [62] synthesized a stable derivative of kojic acid, 5-[(3-aminopropyl)phosphinooxy]-2-(hydroxymethyl)-4H-pyran-4-one (Kojyl-APPA, 139) (Fig. 8). (139) showed tyrosinase inhibition effect (30%) in situ, but not in vitro. (139) inhibited tyrosinase ac-tivity significantly in 24 h after treatment in normal human melano-cytes, suggesting that (139) was not a direct inhibitor of tyrosinase, but instead, converted to a potential inhibitor enzymatically in cells. In addition, (139) decreased melanin content to 75% of control in melanoma cells and decreased neomelanin synthesis to 43% of control in normal human melanocytes. Its permeation through skin was stronger by approximately 8 times than the strength of kojic acid.
Jun et al. [92] synthesized a series of hydroxychalcones. It was shown that 2',4',6'-trihydroxychalcone (140), 2,2',3,4',6'-pentahydroxychalcone (141), 2',3,4,4',5,6'-hexahydroxychalcone (142), 2',4',6'-trihydroxy- 3,4-dimethoxychalcone (143) and 2,2',4,4',6'-pentahydroxychalcone (144) were the most potent ty-rosinase inhibitors. They discovered that (144) was a tyrosinase competitive suppressor of a Ki value of 3.1 M. Compounds (140) and (141) had IC50 values of 193 M and 200 M respectively, whereas (144) had an IC50 value of 1 M.
In a study by Ghani et al. [93] a series of each of the categories were synthesized; 1,3,4-thiadiazole-2(3H)-thiones (146-151), 1,3,4-oxadiazole-2(3H)-thiones (145), 4-amino-1,2,4-triazole-5(4H)-thiones (152) , and substituted hydrazides (Fig. 8). Compounds in (146-151) series were found to be the most potent inhibitors; more specifically, (S1) compound (154-156) (158-166). In the series of (152), compound (153) was the most potent tyrosinase inhibitor. In series of (145), (157) was the most potent inhibitor due to the 4 benzyloxyphenyl group on the oxadiazole ring. Compounds from the hydrazide series were moderate tyrosinase inhibitors.
Wang et al. [94] isolated (-)-N-Formylanonnaine (167, IC50 = 74.3 mg/ml) from the leaves of Michelia alba, which exhibited tyrosinase activity through binding with the predicted tyrosinase model by chelating two copper ions present in the active site (Fig. 8). In Wang et al. (-)-N-Formylanonnaine (167, IC50 = 74.3 mg/ml), isolated from leaves of Michelia alba, exhibited tyrosinase activity of which might be due to its chelation with the two copper ions in the tyrosinase active site. On the other hand, (167) also demon-strated good antioxidant ability in multiple assay platforms.
In Wang et al. [95], Linderanolide B (168) and Subamolide A (169) (IC50 = 138.2 ± 4.8 M and 205.2 ± 3.5 M) were extracted from Cinnamomum subavenium and were found to be potential tyrosinase inhibitors (Fig. 8). Their properties were revealed not only in the mushroom tyrosinase but also in the human melanocyte and zebrafish. Inhibitory properties were present not only in mush-room tyrosinase, but also in the human melanocyte and zebrafish. The article showed that melanin-reducing effects might be due to the copper ion bindings in the tyrosinase reaction domain area, as signified by the molecular model docking.
Emami et al. [96] synthesized a series of 2(3H)-thiazole thiones (170) and one of the compounds from the series, 3-methyl-4-phenyl-2(3H)-thiazole thione (171), which proved to be a sufficient tyrosinase inhibitor as there was 66% inhibition at 30 g/mL (Fig. 8).
Han et al. [97] synthesized a novel compound, MHY384 (172), and the results demonstrated its ability to act as a tyrosinase inhibi-tor (Fig. 8). Results indicated that MHY384 was a more potent
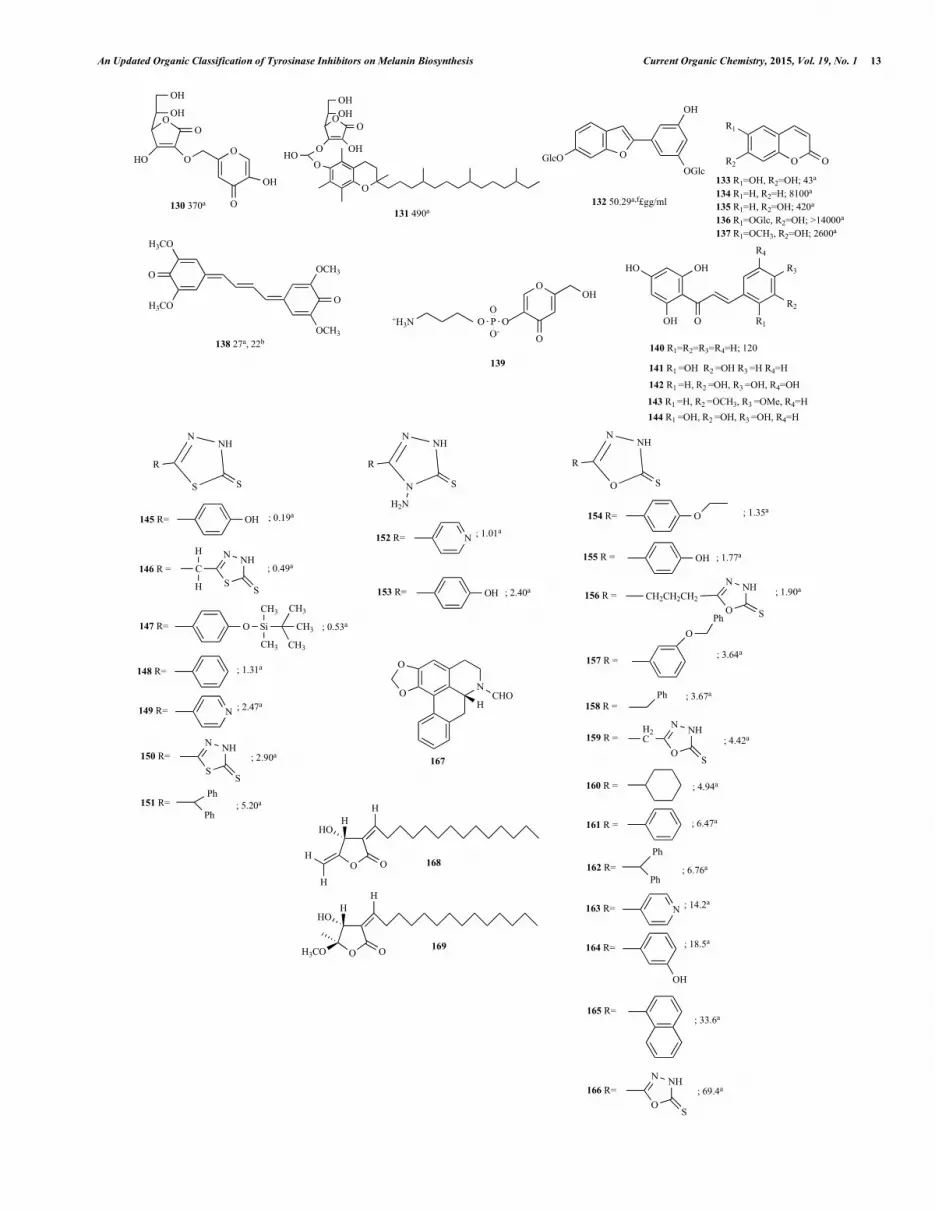
An Updated Organic Classification of Tyrosinase Inhibitors on Melanin Biosynthesis Current Organic Chemistry, 2015, Vol. 19, No. 1 13
OOH
OH
OHO
O
O
O
OH
OOHOH
OHO
O
OHO
O
OGlcOOGlc
OH
O OR2
R1
O
O
OCH3
OCH3
H3CO
H3CO
130 370a131 490a
132 50.29a,f£gg/ml
133 R1=OH, R2=OH; 43a
134 R1=H, R2=H; 8100a
135 R1=H, R2=OH; 420a
136 R1=OGlc, R2=OH; >14000a
137 R1=OCH3, R2=OH; 2600a
138 27a, 22b
O
OHHO
OH
140 R1=R2=R3=R4=H; 120
R2
R1
R3
R4
141 R1 =OH R2 =OH R3 =H R4=H
142 R1 =H, R2 =OH, R3 =OH, R4=OH
143 R1 =H, R2 =OCH3, R3 =OMe, R4=H144 R1 =OH, R2 =OH, R3 =OH, R4=H
S
NHN
S
R
145 R=
146 R =
N
NHN
S
R
H2N
152 R=
O
NHN
S
R
154 R=
155 R =
O
O NCHO
H
O
H
OH
H
HHO
O
H
O
HHO
H3CO
167
168
169
OH
C
H
H
S
NHN
S
147 R= O Si
CH3
CH3 CH3
CH3
CH3
148 R=
149 R= N
150 R=S
NHN
S
151 R=Ph
Ph
; 0.19a
; 0.49a
; 0.53a
; 1.31a
; 2.47a
; 2.90a
; 5.20a
N ; 1.01a
OH ; 2.40a153 R=
O
OH ; 1.77a
; 1.35a
156 R = CH2CH2CH2O
NHN
S
157 R =
OPh
158 R =Ph
159 R =H2C
O
NHN
S
160 R =
161 R =
162 R=Ph
Ph; 6.76a
163 R= N ; 14.2a
164 R= ; 18.5a
OH
165 R=
166 R=O
NHN
S
; 69.4a
; 33.6a
; 6.47a
; 4.94a
; 4.42a
; 3.67a
; 3.64a
; 1.90a
OOH
O
PO
OOO-
+H3N
139
14 Current Organic Chemistry, 2015, Vol. 19, No. 1 Chen et al.
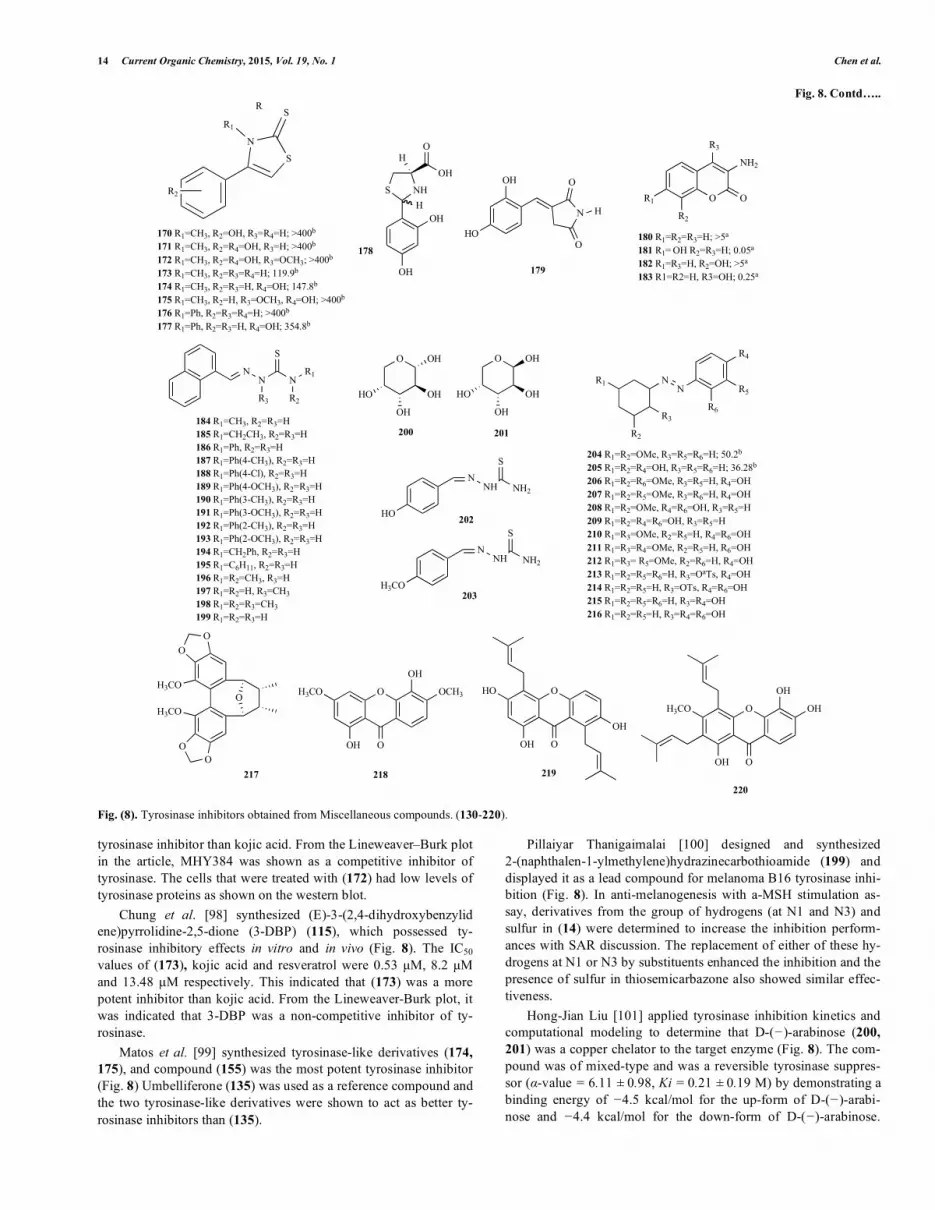
Fig. 8. Contd…..
S
N
SR1
R2
R
178
OH
NHS
O
OHH
H
OH
N
O
O
H
OH
HO
179
O
NH2
O
180 R1=R2=R3=H; >5a
181 R1= OH R2=R3=H; 0.05a
182 R1=R3=H, R2=OH; >5a
183 R1=R2=H, R3=OH; 0.25a
R1
R2
N
R3
R2
R1N
R6
R5
R4
204 R1=R2=OMe, R3=R5=R6=H; 50.2b
205 R1=R2=R4=OH, R3=R5=R6=H; 36.28b
206 R1=R2=R6=OMe, R3=R5=H, R4=OH207 R1=R2=R5=OMe, R3=R6=H, R4=OH208 R1=R2=OMe, R4=R6=OH, R3=R5=H209 R1=R2=R4=R6=OH, R3=R5=H210 R1=R3=OMe, R2=R5=H, R4=R6=OH211 R1=R3=R4=OMe, R2=R5=H, R6=OH212 R1=R3= R5=OMe, R2=R6=H, R4=OH213 R1=R2=R5=R6=H, R3=OaTs, R4=OH214 R1=R2=R5=H, R3=OTs, R4=R6=OH215 R1=R2=R5=R6=H, R3=R4=OH216 R1=R2=R5=H, R3=R4=R6=OH
NN
R3
N
S
R1
R2
184 R1=CH3, R2=R3=H185 R1=CH2CH3, R2=R3=H186 R1=Ph, R2=R3=H187 R1=Ph(4-CH3), R2=R3=H188 R1=Ph(4-Cl), R2=R3=H189 R1=Ph(4-OCH3), R2=R3=H190 R1=Ph(3-CH3), R2=R3=H191 R1=Ph(3-OCH3), R2=R3=H192 R1=Ph(2-CH3), R2=R3=H193 R1=Ph(2-OCH3), R2=R3=H194 R1=CH2Ph, R2=R3=H195 R1=C6H11, R2=R3=H196 R1=R2=CH3, R3=H197 R1=R2=H, R3=CH3198 R1=R2=R3=CH3199 R1=R2=R3=H
O
OH
OH
HO
OH O OH
OH
OH
HO
HO
NNH NH2
S
H3CO
NNH NH2
S
O
OH O
H3CO
OH
OCH3
200 201
202
203
218
170 R1=CH3, R2=OH, R3=R4=H; >400b
171 R1=CH3, R2=R4=OH, R3=H; >400b
172 R1=CH3, R2=R4=OH, R3=OCH3; >400b
173 R1=CH3, R2=R3=R4=H; 119.9b
174 R1=CH3, R2=R3=H, R4=OH; 147.8b
175 R1=CH3, R2=H, R3=OCH3, R4=OH; >400b
176 R1=Ph, R2=R3=R4=H; >400b
177 R1=Ph, R2=R3=H, R4=OH; 354.8b
R3
O
OH O
HO
OH
O
OH O
H3CO
OH
OH
219
220
OO
OO
H3CO
H3COO
217
Fig. (8). Tyrosinase inhibitors obtained from Miscellaneous compounds. (130-220).
tyrosinase inhibitor than kojic acid. From the Lineweaver–Burk plot in the article, MHY384 was shown as a competitive inhibitor of tyrosinase. The cells that were treated with (172) had low levels of tyrosinase proteins as shown on the western blot.
Chung et al. [98] synthesized (E)-3-(2,4-dihydroxybenzylid ene)pyrrolidine-2,5-dione (3-DBP) (115), which possessed ty-rosinase inhibitory effects in vitro and in vivo (Fig. 8). The IC50 values of (173), kojic acid and resveratrol were 0.53 �M, 8.2 �M and 13.48 �M respectively. This indicated that (173) was a more potent inhibitor than kojic acid. From the Lineweaver-Burk plot, it was indicated that 3-DBP was a non-competitive inhibitor of ty-rosinase.
Matos et al. [99] synthesized tyrosinase-like derivatives (174, 175), and compound (155) was the most potent tyrosinase inhibitor (Fig. 8) Umbelliferone (135) was used as a reference compound and the two tyrosinase-like derivatives were shown to act as better ty-rosinase inhibitors than (135).
Pillaiyar Thanigaimalai [100] designed and synthesized 2-(naphthalen-1-ylmethylene)hydrazinecarbothioamide (199) and displayed it as a lead compound for melanoma B16 tyrosinase inhi-bition (Fig. 8). In anti-melanogenesis with a-MSH stimulation as-say, derivatives from the group of hydrogens (at N1 and N3) and sulfur in (14) were determined to increase the inhibition perform-ances with SAR discussion. The replacement of either of these hy-drogens at N1 or N3 by substituents enhanced the inhibition and the presence of sulfur in thiosemicarbazone also showed similar effec-tiveness.
Hong-Jian Liu [101] applied tyrosinase inhibition kinetics and computational modeling to determine that D-(�)-arabinose (200, 201) was a copper chelator to the target enzyme (Fig. 8). The com-pound was of mixed-type and was a reversible tyrosinase suppres-sor (�-value = 6.11 ± 0.98, Ki = 0.21 ± 0.19 M) by demonstrating a binding energy of �4.5 kcal/mol for the up-form of D-(�)-arabi-nose and �4.4 kcal/mol for the down-form of D-(�)-arabinose.

An Updated Organic Classification of Tyrosinase Inhibitors on Melanin Biosynthesis Current Organic Chemistry, 2015, Vol. 19, No. 1 15
Compounds (200) and (201) were the two chemistry forms of D-(–)-arabinose down-form and up-form, and were different in one hydroxyl group conformation. Assessments from intrinsic fluores-cence determined an induction of tertiary tyrosinase change (bind-ing constant K = 1.58±0.02M�1, binding number n = 1.49±0.06).
Liang-Hua Chen [102] synthesized the combination compounds of benzaldehyde and thiosemicarbazone analog, hydroxy- and methoxysubstituted benzaldehyde thiosemicarbazone, HBT (202) and MBT (203) to inhibit B16 mouse melanoma mushroom ty-rosinase and free-cell tyrosinase (Fig. 8). HBT and MBT reduced the steady state of the monophenolase activity (IC50 = 0.76 and 7.0 �M), and both compounds inhibited diphenolase activity dose-dependently (IC50 = 3.80 and 2.62 �M, respectively). Their inhibi-tory mechanisms were reversible and mixed-type, and the inhibition constants of HBT (KI = 2.82 �M; KIS = 6.79 �M) and MBT (KI = 1.47 �M; KIS = 15.10 �M) were identified from kinetics analyses.
Yu Min Song et al. [103] applied a modified Curtius rear-rangement and diazotization followed by a coupling reaction with various phenolic compounds to form azo-resveratrol and azo-oxyresveratrol analogs (Fig. 8). In mushroom tyrosinase inhibition reactions, 4-hydroxyphenyl analogs suppressed the target enzyme more efficiently than the other derivatives including 2,4-dihydroxyphenyl, 2-methoxy-4-hydroxyphenyl and 2-hydroxy-4-methoxyphenyl. Compounds (204) and azo-resveratrol (205) at 50 �M demonstrated 56.25% and 72.75%, respectively. In further study, azo-resveratrol (205) presented an IC50 value of 36.28, while resveratrol’s IC50 was 26.63.
Sung Jin Bae et al. [104] synthesized a series of mushroom ty-rosinase inhibitors, an azo-resveratrol analog, (E)-2-((2,4-dihydroxyphenyl)diazenyl)phenyl 4-methylbenzenesulfonate (213), to be the most effective (Fig. 8). Compound (213) demonstrated competitive inhibition via Lineweaver-Burk plots with an IC50 of 17.85 �M (kojic acid, IC50 = 49.08 �M), as further confirmed by the docking results. Compound (213) demonstrated competitive inhibition and an IC50 of 17.85 �M (kojic acid, IC50 = 49.08 �M) via Lineweaver-Burk plots, as further confirmed by the docking results. They also found these compounds inhibited B16F10 cellular tyrosinase and melanin content with cytotoxicity-free properties.
Myeong-Jin Goh et al. [105] purified a lignan, Kadsuralignan F (217) from Kadsura coccinea, which presented tyrosinase protein down-regulation to inhibit murine melanocyte cellular melanin synthesis in a dose dependent manner (Fig. 8). Interestingly, they found out that Kadsuralignan F did not inhibit tyrosinase activity from mushroom or melan-A cells, and other melanogenic mRNA gene expressions, including Trp-1 or Trp-2.
Zong-Ping Zheng et al. [73] screened the effective extracts to mushroom tyrosinase and discovered that the twigs of Cudrania tricuspidata had a potent inhibition of IC50=113.5 �g/mL. In addi-tion to flavonoids (Fig. 2), three compounds were also isolated: 1,5-dihydroxy-3,6-dimethoxy-xanthen-9-one (218), cudraxanthone H (219) and Dulxanthone-B (220) from the twigs of C. tricuspidata (Fig. 8). Compounds (218-220) demonstrated 2 to 4-fold weaker tyrosinase inhibition activities than that of kojic acid.
MIXTURE EXTRACTS FROM PLANTS AS TYROSINASE INHIBITORS
There are some literature reports about the extracts from natural plants, which inhibit tyrosinase. Matsuda et al. [106] collected from the herbal garden of Kinki University in Japan, get thirty-eight plant dried leaves with 50% ethanolic extracts to investigate the in vitro effects on melanin biosynthesis. Of the thirty-eight extracts, Prunus
yedoensis, P. zippeliana, P. amygdalus, P. persica, P. armeniaca, Thea sinensis and Chaenomeles sinensis demonstrated potential tyrosinase suppression. These herbal extracts obtained from P. ye-doensis and P. zippeliana leaves among the Prunus plants de-creased the melanin amount via auto-oxidation. The suppressive influences of P. zippeliana extracts in melanogenesis were discov-ered from the mouse melanoma B16 cell model.
Matsuda et al. [107] examined melanogenesis from the ob-tained 50% ethanol extracts of dried leaves of 6 Arctostaphylos plants (A. patula, A. viscida, A. canescens, A. columbiana, A. ne-vadensis and A. uva-ursi) (Ericaceae). Two potential extracts among them in A. patula and A. viscida presented suppression to tyrosinase. These functional extracts not only reduced the pigment synthesis amount from dopachrome through auto-oxidation, but had superoxide dismutase (SOD)-like inhibitory actions, and demon-strated a reasonable absorbance in the UV-B area.
Lee et al. [108] evaluated and screened one hundred plant natu-ral extracts to clarify the skin lightening influences via in vitro sup-pression of L-tyrosinase and L-dopa auto-oxidation assay platform. Some herbal natural extracts including Chaenomeles speciosa, Dryopteris crassirhizoma, Gastrodia ellata, Glycyrrhiza glabra, Morus alba, Myristica fragrans, Rheum palmatum and Sophora japonica presented suppressive capacities in mushroom tyrosinase function. Herbal extracts such as Bupleurum falcatum, Caragana sinica, Morus alba and Tussilago farfara demonstrated L-dopa auto-oxidation inhibition.
Kim et al. [62] evaluated the influences in Radix Ginseng (RG) and Radix Trichosanthis (RT) on the melanoma B16 cells in the melanin production. These cells were incubated for two days at concentrations ranging from 1 to 50 mg/ml with RT; at concentra-tions of 10-1000 mg/ml with RG; at various doses (10-1000 mg/ml) with RG; and at 25 mg/ml with RT. Incubation with RT only dose-dependently inhibited tyrosinase function and melanin production compared to the vehicle control group, and drastically suppressed the proliferation in B16 cells. RG at the testing conditions displayed no considerable alteration in tyrosinase function. Incubation with RT in the presence of the testing conditions of RG inhibited ty-rosinase function and melanin amount, similar to the consequence with RT alone, but somewhat augmented proliferation in B16 cells. In addition, the protein expression level of tyrosinase was notably reduced at treatments of 25 mg/ml RT only and with the combina-tion of RG at 100 mg/ml.
An et al. [109] separated the polyphenols of green teas and irra-diated them at 40 kGy by g-ray. The tyrosinase inhibition influ-ences presented 45.2% and 42.9% consequences at a 100 ppm level with the irradiated or non-the irradiated polyphenols, respectively. Two superior than 90% proliferation decreases in dermal cancer SK-MEL-2 and G361 cells were verified in both polyphenols with the irradiated or non-irradiated. In the anti-wrinkle influence test, the collagenase suppression was obvious in the irradiated sample group (65.3%) than that of the non-irradiated control group (56.8%) at 200 ppm of the experimental conditions. The biosynthesis of collagen rates in the irradiated and the non-irradiated polyphenols applying human fibroblasts was 19.4% and 16.3%, respectively. Thus, the green tea polyphenol irradiation had no change, and even augmented its anti-cancer effects, skin-whitening and anti-wrinkle in the human skin.
Wang et al. [110] examined 25 traditional Chinese herbal medi-cine extracts (100 mg/mL) to test tyrosinase inhibition and cytotox-icity on HEMn; 12 extracts exhibited low cytotoxicity. Four natural plans, Pharbitis nil, Sophora japonica, Spatholobus suberectus, and

16 Current Organic Chemistry, 2015, Vol. 19, No. 1 Chen et al.
Morus alba (currently in use as cosmetic additives), revealed effec-tive suppressive effectiveness in tyrosinase, and the values of IC50 were 24.9, 95.6, 83.9, and 78.3 �g/mL, respectively. Pigment sup-pression was not evident in a dose-dependent manner. Two species of nature herbal extracts, Sophora japonica and Spatholobus suberectus proved to be better antioxidants and had better phenolic amounts (IC50: 14.46 �g/mL and DPPH: 1.95 �g/mL, hydroxyl radical; IC50: 10.51 �g/mL and DPPH, 4.36 �g/mL, hydroxyl radi-cal; 255 and 189 mg of gallic acid/g extract, respectively).
Rout et al. [111] isolated a polysaccharide fraction from pome-granate at 4 �g/ml dose, which reduced DPPH and ABTS+ radical abilities in 69% and 88%, respectively. At 10 �g/ml dose, the sup-pression of mushroom tyrosinase was 43%. In addition, the capacity of polysaccharide fraction from pomegranate for the free radical scavenging was assessed through electron spin resonance spectro-photometer and DPPH dot blot assay. Moreover, anti-glycation aptitude of polysaccharide fraction from pomegranate was exam-ined using BSA, which suppressed the synthsis of advanced glyca-tion end-products in 28% and reduced the production of fructosa-mine in the BSA/Glucose model.
Li et al. [112] investigated the Xanthium strumarium L stem ex-tract and found it to inhibit melanogenesis through down-regulation of tyrosinase through GSK3�.
In Zheng et al. [113], oxyresveratrol was extracted from Morus australis besides many new compounds such as austraone A, mora-cenin D, sanggenon T, and kuwanon O. These new compounds showed more effective tyrosinase inhibitory abilities than kojic acid. It was found that Morus australis root extract was a good natural source of tyrosinase inhibitors.
CONCLUSION
Tyrosinase is activated with exposure to UV light, leading to pigment formation. In this article, we classified (natural and synthetic) tyrosinase suppressive factors in terms of the suppression of tyrosinase bio-functions from natural plants or synthetic com-pounds, which included the types of compounds and their sources with characteristics as flavonoids, stilbenes, simple phenolics, furans, triterpenes, alkaloids and other synthetic compounds. The structure-activity relationships have been classified in terms of their inhibition of tyrosinase functions and copper chelating activity.
CONFLICT OF INTEREST
The authors confirm that this article content has no conflict of interest.
ACKNOWLEDGEMENTS
The work was supported by the National Science Council under the Grant nos. NSC99-2221-E-037-006-MY3, NSC102-2622-E-037-002, NSC102-2221-E-037-005, NSC102-2622-E-011-001-CC2 and MOST103-2221-E-037 -004. We thank the project of Center for Stem Cell Research, Kaohsiung Medical University, Kaohsiung, Taiwan, KMU-TP103G00 and KMU-TP103G02-05.
REFERENCES
[1] Fleming, H.; Condon, R.; Peterson, G.; Guck, I.; Prescott, E.; Chatfield, K.; Duff, M. Role of biotin-containing membranes and nuclear distribution in differentiating human endometrial cells. J. Cell. Biochem., 1998, 71(3), 400-415.
[2] Obinata, K.; Maruyama, T.; Hayashi, M.; Watanabe, T.; Nittono, H. Effect of taurine on the fatty liver of children with simple obesity. Adv. Exp. Med.Biol., 1996, 403, 607-613.
[3] Tripathi, R.K.; Hearing, V.J.; Urabe, K.; Aroca, P.; Spritz, R.A. Mutational mapping of the catalytic activities of human tyrosinase. J. Biol. Chem., 1992, 267(33), 23707-23712.
[4] Lugassy, C.; Kleinman, H.; Vernon, S.; Welch, D.; Barnhill, R. C16 laminin peptide increases angiotropic extravascular migration of human melanoma cells in a shell-less chick chorioallantoic membrane assay. Br. J. Dermatol.,2007, 157(4), 780-782.
[5] Sturm, R.A.; Box, N.F.; Ramsay, M. Human pigmentation genetics: The difference is only skin deep. BioEssays, 1998, 20(9), 712-721.
[6] Hermanns, J.F.; Petit, L.; Martalo, O.; Pierard-Franchimont, C.; Cauwenbergh, G.; Pierard, G.E. Unraveling the patterns of subclinical pheomelanin-enriched facial hyperpigmentation: Effect of depigmenting agents. Dermatol., 2000, 201(2), 118-122.
[7] Haq, I.; Ali, S.; Qadeer, M.A.; Iqbal, J. Inducive effect of cresoquinone on microbiological transformation of L-tyrosine to 3,4 dihydroxy phenyl L-alanine by Aspergillus oryzae NG-11(P1). Appl. Microbiol. Biotechnol.,2003, 60(6), 696-699.
[8] Ikehata, K.; Nicell, J.A. Color and toxicity removal following tyrosinase-catalyzed oxidation of phenols. Biotechnol. Prog., 2000, 16(4), 533-540.
[9] Montiel, A.M.; Fernandez, F.J.; Marcial, J.; Soriano, J.; Barrios-Gonzalez, J.; Tomasini, A. A fungal phenoloxidase (tyrosinase) involved in pentachlorophenol degradation. Biotechnol. Lett., 2004, 26(17), 1353-1357.
[10] Ensuncho, L.; Alvarez-Cuenca, M.; Legge, R.L. Removal of aqueous phenol using immobilized enzymes in a bench scale and pilot scale three-phase fluidized bed reactor. Bioprocess Biosyst. Eng., 2005, 27(3), 185-191.
[11] Chen, T.; Embree, H.D.; Wu, L.Q.; Payne, G.F. In vitro protein–polysaccharide conjugation: Tyrosinase-catalyzed conjugation of gelatin and chitosan. Biopolymers, 2002, 64(6), 292-302.
[12] Claus, H.; Decker, H. Bacterial tyrosinases. Syst. Appl. Microbiol., 2006, 29(1), 3-14.
[13] Sugumaran, M. Comparative biochemistry of eumelanogenesis and the protective roles of phenoloxidase and melanin in insects. Pigm. Cell Res.,2002, 15(1), 2-9.
[14] Westerhof, W. The discovery of the human melanocyte. Pigm. Cell Res.,2006, 19(3), 183-193.
[15] Wang, N.; Hebert, D.N. Tyrosinase maturation through the mammalian secretory pathway: Bringing color to life. Pigm. Cell Res., 2006, 19(1), 3-18.
[16] Furumura, M.; Solano, F.; Matsunaga, N.; Sakai, C.; Spritz, R.A.; Hearing, V.J. Metal ligand-binding specificities of the tyrosinase-related proteins. Biochem. Biophys. Res. Commun., 1998, 242(3), 579-585.
[17] Branza-Nichita, N.; Petrescu, A.J.; Dwek, R.A.; Wormald, M.R.; Platt, F.M.; Petrescu, S.M. Tyrosinase folding and copper loading in vivo: A crucial role for calnexin and �-glucosidase II. Biochem. Biophys. Res. Commun., 1999, 261(3), 720-725.
[18] Halaouli, S.; Record, E.; Casalot, L.; Hamdi, M.; Sigoillot, J.C.; Asther, M.; Lomascolo, A. Cloning and characterization of a tyrosinase gene from the white-rot fungus Pycnoporus sanguineus, and overproduction of the recombinant protein in Aspergillus niger. Appl. Microbiol. Biotechnol., 2006, 70(5), 580-589.
[19] Seo, S.Y.; Sharma, V.K.; Sharma, N. Mushroom tyrosinase: Recent prospects. J. Agric. Food Chem., 2003, 51(10), 2837-2853.
[20] Jolivet, P.; Queiroz-Claret, C.; Bergeron, E.; Meunier, J.C. Characterization of an exocellular protein phosphatase with dual substrate specificity from the yeast Yarrowia lipolytica. Int. J. Biochem. Cell Biol., 1998, 30(7), 783-796.
[21] Aroca, P.; Garcia-Borron, J.C.; Solano, F.; Lozano, J.A. Regulation of mammalian melanogenesis. I: Partial purification and characterization of a dopachrome converting factor: dopachrome tautomerase. Biochim. Biophys.Scta, 1990, 1035(3), 266-275.
[22] Kobayashi, T.; Urabe, K.; Winder, A.; Jimenez-Cervantes, C.; Imokawa, G.; Brewington, T.; Solano, F.; Garcia-Borron, J.C.; Hearing, V.J. Tyrosinase related protein 1 (TRP1) functions as a DHICA oxidase in melanin biosynthesis. EMBO J., 1994, 13(24), 5818-5825.
[23] Hirobe, T. Role of keratinocyte-derived factors involved in regulating the proliferation and differentiation of mammalian epidermal melanocytes. Pigm. Cell Res., 2005, 18(1), 2-12.
[24] Lerch, K. Neurospora tyrosinase: Molecular weight, copper content and spectral properties. FEBS Lett., 1976, 69(1), 157-160.
[25] Halaouli, S.; Asther, M.; Kruus, K.; Guo, L.; Hamdi, M.; Sigoillot, J.C.; Asther, M.; Lomascolo, A. Characterization of a new tyrosinase from Pycnoporus species with high potential for food technological applications. J. Appl. Microbiol., 2005, 98(2), 332-343.
[26] Naraoka, T.; Uchisawa, H.; Mori, H.; Matsue, H.; Chiba, S.; Kimura, A. Purification, characterization and molecular cloning of tyrosinase from the cephalopod mollusk, Illex argentinus. Eur. J. Biochem., 2003, 270(19), 4026-4038.
[27] Selinheimo, E.; Saloheimo, M.; Ahola, E.; Westerholm-Parvinen, A.; Kalkkinen, N.; Buchert, J.; Kruus, K. Production and characterization of a secreted, C-terminally processed tyrosinase from the filamentous fungus Trichoderma reesei. FEBS J., 2006, 273(18), 4322-4335.
[28] Ito, M.; Oda, K. An organic solvent resistant tyrosinase from Streptomyces sp. REN-21: Purification and characterization. Biosci. Biotechnol. Biochem.,2000, 64(2), 261-267.

An Updated Organic Classification of Tyrosinase Inhibitors on Melanin Biosynthesis Current Organic Chemistry, 2015, Vol. 19, No. 1 17
[29] Olivares, C.; Solano, F. New insights into the active site structure and catalytic mechanism of tyrosinase and its related proteins. Pigm. Cell.Melanoma Res., 2009, 22(6), 750-760.
[30] Galindo, J.D.; Martinez, J.H.; Lopez-Ballester, J.A.; Penafiel, R.; Solano, F.; Lozano, J.A. The effect of polyamines on tyrosinase activity. Biochem. Int.,1987, 15(6), 1151-1158.
[31] Jacobsohn, G.M.; Jacobsohn, M.K. Incorporation and binding of estrogens into melanin: Comparison of mushroom and mammalian tyrosinases. Biochim. Biophys. Acta., (BBA) - General Subjects, 1992, 1116(2), 173-182.
[32] Funayama, M.; Arakawa, H.; Yamamoto, R.; Nishino, T.; Shin, T.; Murao, S. Effects of alpha- and beta-arbutin on activity of tyrosinases from mushroom and mouse melanoma. Bioscience, Biotechnol. Biochem., 1995,59(1), 143-144.
[33] Jaenicke, E.; Decker, H. Functional changes in the family of type 3 copper proteins during evolution. Chem. Bio. Chem., 2004, 5(2), 163-169.
[34] Gerdemann, C.; Eicken, C.; Krebs, B. The crystal structure of catechol oxidase: new insight into the function of type-3 copper proteins. Acc. Chem.Res., 2002, 35(3), 183-191.
[35] Volbeda, A.; Hol, W.G.J. Crystal structure of hexameric haemocyanin from Panulirus interruptus refined at 3.2 Å resolution. J. Mol. Biol., 1989, 209(2), 249-279.
[36] Solomon, E.I.; Sundaram, U.M.; Machonkin, T.E. Multicopper oxidases and oxygenases. Chem. Rev., 1996, 96(7), 2563-2606.
[37] Ros-Martinez, J.R.; Rodriguez-Lopez, J.N.; Castellanos, R.V.; Garcia-Canovas, F. Discrimination between two kinetic mechanisms for the monophenolase activity of tyrosinase. Biochem. J., 1993, 294(Pt 2), 621-622.
[38] Cheng, S.L.; Huang Liu, R.; Sheu, J.N.; Chen, S.T.; Sinchaikul, S.; Tsay, G.J. Toxicogenomics of kojic acid on gene expression profiling of a375 human malignant melanoma cells. Biol. Pharm. Bull., 2006, 29(4), 655-669.
[39] Burdock, G.A.; Soni, M.G.; Carabin, I.G. Evaluation of health aspects of kojic acid in food. Regul. Toxicol. Pharmacol., 2001, 33(1), 80-101.
[40] Higa, Y.; Ohkubo, A.; Kitajima, S.; Moriyasu, M.; Kariya, K. Effects of kojic acid on thyroidal functions in rats by single-dose administration and in cultured rat thyroid cells (FRTL-5 cells). J. Toxicol. Sci., 2002, 27(5), 423-431.
[41] Qiu, L.; Zhang, M.; Sturm, R.A.; Gardiner, B.; Tonks, I.; Kay, G.; Parsons, P.G. Inhibition of melanin synthesis by cystamine in human melanoma cells. J. Invest. Dermatol., 2000, 114(1), 21-27.
[42] Mishima, Y.; Hatta, S.; Ohyama, Y.; Inazu, M. Induction of melanogenesis suppression: Cellular pharmacology and mode of differential action. Pigment cell research / sponsored by the European Society for Pigment Cell Research and the International Pigment Cell Society, 1988, 1(6), 367-374.
[43] Naish-Byfield, S.; Cooksey, C.J.; Riley, P.A. Oxidation of monohydric phenol substrates by tyrosinase: Effect of dithiothreitol on kinetics. Biochem.J., 1994, 304(Pt 1), 155-162.
[44] Petris, M.J.; Strausak, D.; Mercer, J.F. The Menkes copper transporter is required for the activation of tyrosinase. Hum. Mol. Genet., 2000, 9(19), 2845-2851.
[45] Shimogaki, H.; Tanaka, Y.; Tamai, H.; Masuda, M. In vitro and in vivoevaluation of ellagic acid on melanogenesis inhibition. Int. J. Cosmet. Sci.,2000, 22(4), 291-303.
[46] Park, H.S.; Lee, U.H.; Choi, J.C.; Chun, D.K. Mucinous eccrine nevus. J.Dermatol., 2004, 31(8), 687-689.
[47] Maeda, K.; Fukuda, M., Arbutin: Mechanism of its depigmenting action in human melanocyte culture. J. Pharmacol. Exper. Therap., 1996, 276(2), 765-769.
[48] Arndt, K.A.; Fitzpatrick, T.B. Topical use of hydroquinone as a depigmenting agent. JAMA : J. Am. Med. Assoc., 1965, 194(9), 965-967.
[49] Grimes, P.E. Melasma. Etiologic and therapeutic considerations. Arch.Dermatol., 1995, 131(12), 1453-1457.
[50] Jimbow, K.; Obata, H.; Pathak, M.A.; Fitzpatrick, T.B. Mechanism of depigmentation by hydroquinone. J. Invest. Dermatol., 1974, 62(4), 436-449.
[51] Nazzaro-Porro, M.; Passi, S. Identification of tyrosinase inhibitors in cultures of Pityrosporum. J. Invest. Dermatol., 1978, 71(3), 205-208.
[52] Nazzaro-Porro, M.; Passi, S.; Balus, L.; Breathnach, A.; Martin, B.; Morpurgo, G. Effect of dicarboxylic acids on lentigo maligna. J. Invest.Dermatol., 1979, 72(6), 296-305.
[53] Palumbo, A.; Solano, F.; Misuraca, G.; Aroca, P.; Garcia Borron, J.C.; Lozano, J.A.; Prota, G. Comparative action of dopachrome tautomerase and metal ions on the rearrangement of dopachrome. Biochim. Biophys. Acta.,(BBA) - General Subjects, 1991, 1115(1), 1-5.
[54] Benavente-Garcia, O.; Castillo, J. Update on uses and properties of citrus flavonoids: new findings in anticancer, cardiovascular, and anti-inflammatory activity. J. Aagric. Food Chem., 2008, 56(15), 6185-6205.
[55] van Acker, S.A.; van den Berg, D.J.; Tromp, M.N.; Griffioen, D.H.; van Bennekom, W.P.; van der Vijgh, W.J.; Bast, A. Structural aspects of antioxidant activity of flavonoids. Free Radical Biol. Med., 1996, 20(3), 331-342.
[56] Jacob, V.; Hagai, T.; Soliman, K. Structure-activity relationships of flavonoids. Curr. Org. Chem., 2011, 15(15), 2641-2657.
[57] Matsuda, H.; Higashino, M.; Chen, W.; Tosa, H.; Iinuma, M.; Kubo, M. Studies of cuticle drugs from natural sources. III. Inhibitory effect of Myrica rubra on melanin biosynthesis. Biol. Pharm. Bull., 1995, 18(8), 1148-1150.
[58] Shimizu, K.; Kondo, R.; Sakai, K.; Lee, S.H.; Sato, H. The inhibitory components from Artocarpus incisus on melanin biosynthesis. Planta Med.,1998, 64(5), 408-412.
[59] Shimizu, K.; Kondo, R.; Sakai, K. Inhibition of tyrosinase by flavonoids, stilbenes and related 4-substituted resorcinols: structure-activity investigations. Planta med., 2000, 66(1), 11-15.
[60] Shimizu, K.; Kondo, R.; Sakai, K.; Takeda, N.; Nagahata, T. The skin-lightening effects of artocarpin on UVB-induced pigmentation. Planta Med.,2002, 68(1), 79-81.
[61] Kubo, I.; Kinst-Hori, I.; Yokokawa, Y. Tyrosinase inhibitors from Anacardium occidentale fruits. J. Nat. Prod., 1994, 57(4), 545-551.
[62] Kim, D.S.; Hwang, E.S.; Lee, J.E.; Kim, S.Y.; Kwon, S.B.; Park, K.C. Sphingosine-1-phosphate decreases melanin synthesis via sustained ERK activation and subsequent MITF degradation. J. Cell Sci., 2003, 116(Pt 9), 1699-1706.
[63] Nerya, O.; Vaya, J.; Musa, R.; Izrael, S.; Ben-Arie, R.; Tamir, S. Glabrene and isoliquiritigenin as tyrosinase inhibitors from licorice roots. J. Agric.Food Chem., 2003, 51(5), 1201-1207.
[64] Yokota, M.; Saido, T.C.; Miyaji, K.; Tani, E.; Kawashima, S.; Suzuki, K. Stimulation of protein-tyrosine phosphorylation in gerbil hippocampus after global forebrain ischemia. Neurosci. Lett., 1994, 168(1-2), 69-72.
[65] Vaya, J.; Belinky, P.A.; Aviram, M. Antioxidant constituents from licorice roots: Isolation, structure elucidation and antioxidative capacity toward LDL oxidation. Free Radical Biol. Med., 1997, 23(2), 302-313.
[66] Belinky, P.A.; Aviram, M.; Mahmood, S.; Vaya, J. Structural aspects of the inhibitory effect of glabridin on LDL oxidation. Free Radical Biol. Med.,1998, 24(9), 1419-1429.
[67] Nerya, O.; Musa, R.; Khatib, S.; Tamir, S.; Vaya, J. Chalcones as potent tyrosinase inhibitors: The effect of hydroxyl positions and numbers. Phytochem., 2004, 65(10), 1389-1395.
[68] Chang, T.S.; Ding, H.Y.; Lin, H.C. Identifying 6,7,4'-trihydroxyisoflavone as a potent tyrosinase inhibitor. Biosci. Biotechnol. Biochem., 2005, 69(10), 1999-2001.
[69] Cheng, K.T.; Hsu, F.L.; Chen, S.H.; Hsieh, P.K.; Huang, H.S.; Lee, C.K.; Lee, M.H. New constituent from Podocarpus macrophyllus var. macrophyllus shows anti-tyrosinase effect and regulates tyrosinase-related proteins and mRNA in human epidermal melanocytes. Chem. Pharm. Bull.,2007, 55(5), 757-761.
[70] Lin, Y.P.; Hsu, F.L.; Chen, C.S.; Chern, J.W.; Lee, M.H. Constituents from the Formosan apple reduce tyrosinase activity in human epidermal melanocytes. Phytochemistry., 2007, 68(8), 1189-1199.
[71] Khatib, S.; Nerya, O.; Musa, R.; Shmuel, M.; Tamir, S.; Vaya, J. Chalcones as potent tyrosinase inhibitors: the importance of a 2,4-substituted resorcinol moiety. Bioorg. Med. Chem., 2005, 13(2), 433-441.
[72] Miyazawa, M.; Tamura, N. Inhibitory compound of tyrosinase activity from the sprout of Polygonum hydropiper L. (Benitade). Biol. Pharm. Bull., 2007,30(3), 595-597.
[73] Zheng, Z.P.; Tan, H.Y.; Chen, J.; Wang, M. Characterization of tyrosinase inhibitors in the twigs of Cudrania tricuspidata and their structure-activity relationship study. Fitoterapia., 2013, 84(0), 242-247.
[74] Iida, K.; Hase, K.; Shimomura, K.; Sudo, S.; Kadota, S.; Namba, T. Potent inhibitors of tyrosinase activity and melanin biosynthesis from Rheum officinale. Planta Med., 1995, 61(5), 425-428.
[75] Shin, N.H.; Ryu, S.Y.; Choi, E.J.; Kang, S.H.; Chang, I.L.M.; Min, K.R.; Kim, Y. Oxyresveratrol as the potent inhibitor on dopa oxidase activity of mushroom tyrosinase. Biochem. Biophys. Res. Commun., 1998, 243(3), 801-803.
[76] Kim, D.S.; Kim, S.Y.; Chung, J.H.; Kim, K.H.; Eun, H.C.; Park, K.C. Delayed ERK activation by ceramide reduces melanin synthesis in human melanocytes. Cell Signalling, 2002, 14(9), 779-785.
[77] Ohguchi, K.; Tanaka, T.; Kido, T.; Baba, K.; Iinuma, M.; Matsumoto, K.; Akao, Y.; Nozawa, Y. Effects of hydroxystilbene derivatives on tyrosinase activity. Biochem. Biophys. Res. Commun., 2003, 307(4), 861-863.
[78] Likhitwitayawuid, K.; Sornsute, A.; Sritularak, B.; Ploypradith, P. Chemical transformations of oxyresveratrol (trans-2,4,3',5'-tetrahydroxystilbene) into a potent tyrosinase inhibitor and a strong cytotoxic agent. Bioorg. Med. Chem.Lett., 2006, 16(21), 5650-5653.
[79] Tajima, R.; Oozeki, H.; Muraoka, S.; Tanaka, S.; Motegi, Y.; Nihei, H.; Yamada, Y.; Masuoka, N.; Nihei, K.-i. Synthesis and evaluation of bibenzyl glycosides as potent tyrosinase inhibitors. Eur. J. Med. Chem., 2011, 46(4), 1374-1381.
[80] Hori, I.; Nihei, K.-i.; Kubo, I. Structural criteria for depigmenting mechanism of arbutin. Phytotherapy Res., 2004, 18(6), 475-479.
[81] Kim, Y.J.; Uyama, H. Tyrosinase inhibitors from natural and synthetic sources: structure, inhibition mechanism and perspective for the future. Cell.Mol. Life Sci. CMLS., 2005, 62(15), 1707-1723.
[82] Ha, Y.M.; Chung, S.W.; Song, S.; Lee, H.; Suh, H.; Chung, H.Y. 4-(6-Hydroxy-2-naphthyl)-1,3-bezendiol: A potent, new tyrosinase inhibitor. Biol.Pharml bul., 2007, 30(9), 1711-1715.
[83] Sharma, V.K.; Choi, J.; Sharma, N.; Choi, M.; Seo, S.-Y. In vitro anti-tyrosinase activity of 5-(hydroxymethyl)-2-furfural isolated from Dictyophora indusiata. Phytotherapy Res., 2004, 18(10), 841-844.
[84] Um, S.J.; Park, M.S.; Park, S.H.; Han, H.S.; Kwon, Y.J.; Sin, H.S. Synthesis of new glycyrrhetinic acid (GA) derivatives and their effects on tyrosinase activity. Bioor. Med. Chem., 2003, 11(24), 5345-5352.
[85] Maeda, K.; Naitou, T.; Umishio, K.; Fukuhara, T.; Motoyama, A., A novel melanin inhibitor: Hydroperoxy traxastane-type triterpene from flowers of Arnica montana. Biol. Pharm. Bull., 2007, 30(5), 873-879.

18 Current Organic Chemistry, 2015, Vol. 19, No. 1 Chen et al.
[86] Shaheen, F.; Ahmad, M.; Khan, M.T.; Jalil, S.; Ejaz, A.; Sultankhodjaev, M.N.; Arfan, M.; Choudhary, M.I.; Atta ur, R. Alkaloids of Aconitum laeve and their anti-inflammatory antioxidant and tyrosinase inhibition activities. Phytochemistry, 2005, 66(8), 935-940.
[87] Kwak, S.Y.; Yang, J.K.; Choi, H.R.; Park, K.C.; Kim, Y.B.; Lee, Y.S. Synthesis and dual biological effects of hydroxycinnamoyl phenylalanyl/prolyl hydroxamic acid derivatives as tyrosinase inhibitor and antioxidant. Bioorg. Med. Chem. Lett., 2013, 23(4), 1136-1142.
[88] Morisaki, K.; Ozaki, S. Design of novel hybrid vitamin C derivatives: Thermal stability and biological activity. Chem. Pharm. Bull., 1996, 44(9), 1647-1655.
[89] Lee, S.H.; Choi, S.Y.; Kim, H.; Hwang, J.S.; Lee, B.G.; Gao, J.J.; Kim, S.Y. Mulberroside F isolated from the leaves of Morus alba inhibits melanin biosynthesis. Biol. Pharm. Bull., 2002, 25(8), 1045-1048.
[90] Masamoto, Y.; Ando, H.; Murata, Y.; Shimoishi, Y.; Tada, M.; Takahata, K. Mushroom tyrosinase inhibitory activity of esculetin isolated from seeds of Euphorbia lathyris L. Biosci. Biotechnol. Biochem., 2003, 67(3), 631-634.
[91] Kanehira, T.; Takekoshi, S.; Nagata, H.; Osamura, R.Y.; Homma, T. Kinobeon A as a potent tyrosinase inhibitor from cell culture of safflower: In vitro comparisons of kinobeon A with other putative inhibitors. Planta Med.,2003, 69(5), 457-459.
[92] Jun, N.; Hong, G.; Jun, K. Synthesis and evaluation of 2',4',6'-trihydroxychalcones as a new class of tyrosinase inhibitors. Bioorg. Med.Chem., 2007, 15(6), 2396-2402.
[93] Ghani, U.; Ullah, N. New potent inhibitors of tyrosinase: Novel clues to binding of 1,3,4-thiadiazole-2(3H)-thiones, 1,3,4-oxadiazole-2(3H)-thiones, 4-amino-1,2,4-triazole-5(4H)-thiones, and substituted hydrazides to the dicopper active site. Bioorg. Med. Chem., 2010, 18(11), 4042-4048.
[94] Wang, H.M.; Chen, C.Y.; Chen, C.Y.; Ho, M.L.; Chou, Y.T.; Chang, H.C.; Lee, C.H.; Wang, C.Z.; Chu, I.M. (�)-N-Formylanonaine from Michelia alba as a human tyrosinase inhibitor and antioxidant. Bioorg. Med. Chem., 2010,18(14), 5241-5247.
[95] Wang, H.M.; Chiu, C.C.; Wu, P.F.; Chen, C.Y. Subamolide E from cinnamomum subavenium induces sub-G1 cell-cycle arrest and caspase-dependent apoptosis and reduces the migration ability of human melanoma cells. J. Agric. Food Chem., 2011, 59(15), 8187-8192.
[96] Emami, S.; Hosseinimehr, S.J.; Shahrbandi, K.; Enayati, A.A.; Esmaeeli, Z. Synthesis and evaluation of 2(3H)-thiazole thiones as tyrosinase inhibitors. Archiv der Pharmazie, 2012, 345(8), 629-637.
[97] Han, Y.K.; Park, Y.J.; Ha, Y.M.; Park, D.; Lee, J.Y.; Lee, N.; Yoon, J.H.; Moon, H.R.; Chung, H.Y. Characterization of a novel tyrosinase inhibitor, (2RS,4R)-2-(2,4-dihydroxyphenyl)thiazolidine-4-carboxylic acid (MHY 384). Biochim. Biophys. Acta (BBA) - Gen. Sub., 2012, 1820(4), 542-549.
[98] Chung, K.W.; Park, Y.J.; Choi, Y.J.; Park, M.H.; Ha, Y.M.; Uehara, Y.; Yoon, J.H.; Chun, P.; Moon, H.R.; Chung, H.Y. Evaluation of in vitro and in vivo anti-melanogenic activity of a newly synthesized strong tyrosinase inhibitor (E)-3-(2,4 dihydroxybenzylidene)pyrrolidine-2,5-dione (3-DBP). Biochim. Biophys. Acta., (BBA) - Gen. Sub., 2012, 1820(7), 962-969.
[99] Matos, M.J.; Santana, L.; Uriarte, E.; Serra, S.; Corda, M.; Fadda, M.B.; Era, B.; Fais, A. Tyrosine-like condensed derivatives as tyrosinase inhibitors. J.Pharm. Pharmacol., 2012, 64(5), 742-746.
[100] Thanigaimalai, P.; Rao, E.V.; Lee, K.C.; Sharma, V.K.; Roh, E.; Kim, Y.; Jung, S.H. Structure–activity relationship of naphthaldehydethiosemicar-bazones in melanogenesis inhibition. Bioorg. Med. Chem. Lett., 2012, 22(2), 886-889.
[101] Liu, H.J.; Ji, S.; Fan, Y.Q.; Yan, L.; Yang, J.M.; Zhou, H.M.; Lee, J.; Wang, Y.L. The effect of D-(-)-arabinose on Tyrosinase: An integrated study using computational simulation and inhibition kinetics. Enzyme Res., 2012, 2012,731427.
[102] Chen, L.H.; Hu, Y.H.; Song, W.; Song, K.K.; Liu, X.; Jia, Y.L.; Zhuang, J.X.; Chen, Q.X. Synthesis and antityrosinase mechanism of benzaldehyde thiosemicarbazones: Novel tyrosinase inhibitors. J. Agric. Food Chem.,2012, 60(6), 1542-1547.
[103] Song, Y.M.; Ha, Y.M.; Kim, J.A.; Chung, K.W.; Uehara, Y.; Lee, K.J.; Chun, P.; Byun, Y.; Chung, H.Y.; Moon, H.R. Synthesis of novel azo-resveratrol, azo-oxyresveratrol and their derivatives as potent tyrosinase inhibitors. Bioorg. Med. Chem. Lett., 2012, 22(24), 7451-7455.
[104] Bae, S.J.; Ha, Y.M.; Kim, J.A.; Park, J.Y.; Ha, T.K.; Park, D.; Chun, P.; Park, N.H.; Moon, H.R.; Chung, H.Y. A novel synthesized tyrosinase inhibitor: (E)-2-((2,4-dihydroxyphenyl)diazenyl)phenyl 4-methylbenzenesul-fonate as an azo-resveratrol analog. Biosci. Biotechnol.Biochem., 2013,77(1), 65-72.
[105] Goh, M.J.; Lee, H.K.; Cheng, L.; Kong, D.Y.; Yeon, J.H.; He, Q.Q.; Cho, J.C.; Na, Y.J. Depigmentation effect of kadsuralignan f on melan-a murine melanocytes and human skin equivalents. Int. J. Mol. Sci., 2013, 14(1), 1655-1666.
[106] Matsuda, H.; Nakamura, S.; Kubo, M. Studies of cuticle drugs from natural sources. II. Inhibitory effects of Prunus plants on melanin biosynthesis. Biol.Pharm. Bull., 1994, 17(10), 1417-1420.
[107] Matsuda, H.; Higashino, M.; Nakai, Y.; Iinuma, M.; Kubo, M.; Lang, F.A. Studies of cuticle drugs from natural sources. IV. Inhibitory effects of some Arctostaphylos plants on melanin biosynthesis. Biolog. Pharm. Bull., 1996,19(1), 153-156.
[108] Lee, K.T.; Kim, B.J.; Kim, J.H.; Heo, M.Y.; Kim, H.P. Biological screening of 100 plant extracts for cosmetic use (I): Inhibitory activities of tyrosinase and DOPA auto-oxidation. Int. J. Cosmet. Sci., 1997, 19(6), 291-298.
[109] An, B.J.; Kwak, J.H.; Son, J.H.; Park, J.M.; Lee, J.Y.; Park, T.S.; Kim, S.Y.; Kim, Y.S.; Jo, C.; Byun, M.W. Physiological activity of irradiated green tea polyphenol on the human skin. Am. J. Chin. Med., 2005, 33(4), 535-546.
[110] Wang, K.J.; Zhang, Y.J.; Yang, C.R. Antioxidant phenolic compounds from rhizomes of Polygonum paleaceum. J. Ethnopharmacol., 2005, 96(3), 483-487.
[111] Rout, S.; Banerjee, R. Free radical scavenging, anti-glycation and tyrosinase inhibition properties of a polysaccharide fraction isolated from the rind from Punica granatum. Bioresour. Technol., 2007, 98(16), 3159-3163.
[112] Li, H.; Min, Y.S.; Park, K.C.; Kim, D.S. Inhibition of melanogenesis by Xanthium strumarium L. Biosci. Biotechnol. Biochem., 2012, 76(4), 767-771.
[113] Zheng, Z.P.; Tan, H.Y.; Wang, M. Tyrosinase inhibition constituents from the roots of Morus australis. Fitoterapia, 2012, 83(6), 1008-1013.
Received: February 19, 2014 Revised: August 20, 2014 Accepted: November 03, 2014