melanocytes, melanin-synthesis, and related signaling pathway
TRANSCRIPT

HAL Id: tel-01823030https://tel.archives-ouvertes.fr/tel-01823030
Submitted on 25 Jun 2018
HAL is a multi-disciplinary open accessarchive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come fromteaching and research institutions in France orabroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, estdestinée au dépôt et à la diffusion de documentsscientifiques de niveau recherche, publiés ou non,émanant des établissements d’enseignement et derecherche français ou étrangers, des laboratoirespublics ou privés.
Melanocytes, melanin-synthesis, and related signalingpathway
yinjuan Wang
To cite this version:yinjuan Wang. Melanocytes, melanin-synthesis, and related signaling pathway. Dermatology. Uni-versité Bourgogne Franche-Comté, 2017. English. �NNT : 2017UBFCE005�. �tel-01823030�

1
UNIVERSITE BOURGOGNE- FRANCHE COMTE ECOLE DOCTORALE « ENVIRONNEMENTS-SANTE »
Année 2017
THESE
Pour obtenir le grade de
Docteur de l’Université de Franche-Comté
Spécialité : Sciences de la Vie et de la Santé
Présentée et soutenue publiquement
Le 12 Sept 2017
Par
Yinjuan Wang
Melanocyte,melaninsynthesisandrelatedsignalingpathway
Thesis director: Professor Philippe Humbert
Li LI, Professor, University of Sichuan, China Reviewer
Xiuli WANG, Professor, University of Tongji, China Reviewer
Philippe HUMBERT, Professor, University of Bourgogne Franche-Comté, France Thesis Director
Li HE, Professor, University of Kunming Medical, China Examinator
Jean-Yves BERTHON, Director, Greentech Biotechnical Company, France Examinator
Céline VIENNET, Research engineer, University of Bourgogne Franche-Comté, France Examinator

2
Acknowledgments
ForProf.PhilippeHUMBERT
ThankyouforbelievingmeandgivingmethepossibilitytodomyPhDthesisinyourlab.
Thankyouforsupportingmesomuchchancestolearningandtraininginpast3-year.This
3-yearismeaningfulforcareerandmakemehavemoreconfidencetochoosethefuturelife
Ilove.
ForProf.LiHE
YouareoneofprofessorsIadmiremost.Youaremyprofessorbothinacademicandinlife.
Thankyouforyoualwaysbelievingmeandgivingmefreedomandspacetofinishmywork
individually.Yourencouragementandtrustsupportmestrongertofaceanydifficult.
ForCélineVIENNET
Thankyouforstayingwithmeinpast3-year,weexperiencedlotsofdifficultandchallenge
together.Youspenttoomuchspiritandtimeinconductingmythesisandhelpingme,always
bepatientandbepolite.Thanksalotforallyourhelpandwarmsmile.
ForMr.Jean-YvesBERTHON,
Thank you for being my co-director and providing me the precious chance to widemy
academicvisionandpresentmythesis inSPIM.Thankyouforthefinancialsupport from
GreentechBiotechologyCompany.
ForProf.LiLIandProf.Xiu-liWANG
Thankyouforbeingthereviewersofmythesis.Itismygreathonorthatyouwouldliketo
bethejuriesinmydefense,yourcomingwillsupportmemoreconfidencetodefenseand
finishmythesis.

3
ForProf.MURET,Sophie,Marion,andGwen
Thankyouforjoiningmythesisandgivingmeprecioussuggestions.ThankstoMarionfor
allthetechnicalsupport.
Forcolleagues,
ThankstoThomasforgivingmestatisticssupportinginmythesis,kindnessandalltheother
help.ThankstoFerialforgivingmeonemonthclinicaltrainingandfollowinghelpduring
work.ThankstoAdelineforhelpingmetofinishmyclinicalresearchandalltheotherhelp.
ThankstoRaneshaforprovidingmetheprecioussampleformyexperiment.Thankstoall
thehelpfromAgnès,Ahmed,Céline,Vanessa,Aurélie,Isabelle.
Formyfriends,
ThankstoYoussef,Vaheide,Som,Phaeng,Delphine,Victoria,Faty,thefriendshipfromallof
yougivemespiritandconfidencetoovercomethedifficulties.
Formyparents
Lastbutnotleast,Iwouldliketoexpressmysincerelygratefultomyparents.HowluckyI
amtohavemyopen-mindedparentswhoalwaysrespectmyidea,believeme,encourageme.
Thanksforunderstandingandsupportingmetoachievemydreaminpast3years.Thanks
forkeepinghealth.

4
Contents
Acknowledgments................................................................................................................................2
Contents...................................................................................................................................................4
ListofFigures.........................................................................................................................................8
ListofTables.......................................................................................................................................13
ListofAbbreviations........................................................................................................................14
GeneralIntroduction........................................................................................................................17
Abstract.................................................................................................................................................19
ChapterI...............................................................................................................................................22
LiteratureReview..............................................................................................................................22
I. Structureoftheskin...........................................................................................................................23
I.IEpidermis.........................................................................................................................................................24
I.IIDermis...............................................................................................................................................................25
I.IIISubcutaneoustissue...................................................................................................................................25
II. SkinPigmentation...............................................................................................................................25
II.IMelanocyteanditscharacteristics.......................................................................................................25
II.IIMelanogenesisandrelatedenzymesandmodulators................................................................26
II.IIIMainsignalingpathwaysregulatingmelanogenesisinmelanocyte....................................30
II.IVRoleofepidermalkeratinocytesinmelanogenesis....................................................................33
II.VRoleofdermalfibroblastsinmelanogenesis.................................................................................35
III. DKKsfamily...........................................................................................................................................38

5
III.IStructure........................................................................................................................................................38
III.IIFunctions.......................................................................................................................................................39
IV.DKK1........................................................................................................................................................40
IV.IDKK1-LRP5/6complex..........................................................................................................................40
IV.IIDKK1skinexpression.............................................................................................................................40
IV.IIIDKK1effectsonkeratinocytes...........................................................................................................40
IV.IVDKK1effectsonmelanocytes..............................................................................................................44
IV.VRoleofDKK1invitiligo..........................................................................................................................45
IV.VIRoleofDKK1inmelanoma..................................................................................................................46
V. TGF-bfamily..........................................................................................................................................47
V.ITGF-bandrelatedsignalingpathway..............................................................................................47
V.IITGF-bsandfibroblasts...........................................................................................................................48
V.IIIp38-MAPKsignalingpathwayactivationbyTGF-b..................................................................49
VI. Conclusion.........................................................................................................................................50
Article1:Preciseroleofdermalfibroblastsonmelanocytepigmentation........................................50
ChapterII..............................................................................................................................................84
Developmentandvalidationofasimplemethodfortheextractionofhumanskin
melanocytes.........................................................................................................................................84
Article2:Developmentandvalidationofasimplemethodfortheextractionofhumanskin
melanocytes....................................................................................................................................................87
ChapterIII..........................................................................................................................................107
InvolvementofDickkopf-relatedprotein1inmelanogenesis:focusonsolarlentigo
lesion...................................................................................................................................................107

6
I. AimsoftheStudy.....................................................................................................................108
II. Hypothesis.................................................................................................................................108
III. MethodsandMaterials.....................................................................................................111
III.I.PartI-Solarlentigobiopsies.................................................................................................................111
III.I.I.Collectionofskinbiopsies...............................................................................................................................111
III.I.II.Histology.................................................................................................................................................................111
III.I.II.IFixationandsectioning...........................................................................................................................111
III.I.II.IIMasson'strichromestainingforcollagenfibers...........................................................................111
III.I.II.IIIMasson'sFontanastainingformelanin...........................................................................................112
III.I.II.IV.Immunostainingforβ-catenin............................................................................................................112
III.I.III.Cellculture.............................................................................................................................................................112
III.I.III.I.Extractionoffibroblasts.........................................................................................................................112
III.I.III.II.Establishmentofskinfibroblastcultures......................................................................................112
III.I.IV.Enzyme-linkedimmunosorbentassay(ELISA)....................................................................................113
III.I.V.Biuretproteinassay..........................................................................................................................................113
III.I.VI.Real-timequantitativereversetranscription-PCR..............................................................................113
III.I.VII.ConditionedmediumfromFLandFNinculturedhumanmelanocytes...................................114
III.I.VIII.MTTassay............................................................................................................................................................115
III.I.IX.ImmunostainingforMITF.............................................................................................................................116
III.I.X.Melaninassay......................................................................................................................................................116
III.II.3Dbiologicalmodel.................................................................................................................................117
III.II.I.Fibroblasts-populatedtensecollagenlatticepreparation(FPCL)................................................117
III.II.II.Repeated-UVAirradiation.............................................................................................................................118
III.III.ELISA.............................................................................................................................................................119
III.IV.Statisticalanalysis....................................................................................................................................120
IV. Results....................................................................................................................................120

7
IV.I.ComparativeanalysesofstaininginSLversusSNbiopsies...................................................120
IV.II.ExpressionandsecretionofDKK1fromFLandFL..................................................................121
IV.III.SecretionofTGF-β1fromFLandFL...............................................................................................123
IV.IV.EffectsofconditionedmediumfromFLandFNonnormalmelanocytecultures.......124
IV.IV.I.Melanocyteviability.........................................................................................................................................124
IV.IV.II.Melanocytecharacteristics............................................................................................................................126
IV.V.SecretionofDKK1afterirradiationofnormalfibroblastsembeddedin3Dcollagen
gel......................................................................................................................................................................................129
V. Discussion-Perspectives.......................................................................................................129
ListofPublications&Communications...................................................................................133
Publications.................................................................................................................................................134
Posters...........................................................................................................................................................135
Article3:Assessmentoftheefficacyofanewcomplexanti-sensitiveskincream.............138
References.........................................................................................................................................163

8
ListofFigures
Figure1.Structureoftheskin(SourcefromPearsonEducation)...............................................23
Figure2.Structureofepidermis(SourcefromHumanAnatomyLibrary).............................24
Figure3.Fourstagesofmelanosome:stageI,premelanosome,sphericalformcontaining
dense spot and few filaments; stage II, premelanosome, ellipsoidal form containing
organized, structured fibrillar matrix; stage III, the beginning of the melanin
production;stageIV,beingfullofmelanin99...................................................................................27
Figure4.Melaninsynthesisinmelanocytes100.................................................................................29
Figure5.Canonicalandnon-canonicalWntsignalingpathways.a.CanonicalWnt/β-
catenin signaling pathway, the ternary complex of Wnt-Lrp5/6-Fzd interacts with
Axin/APC/GSK-3toactiveordegradetheaccumulationofβ-catenin.b.Non-canonical
Wnt/Ca2+signalingpathwaythroughinteractionofWntligandswithFzdreceptorscan
lead to an increase in intracellular calcium level, and involves activation of PLC
(phospholipase C). c. Non-canonicalWnt/PCP (Planar Cell Polarity) pathway, it’s is
characterizedbyanasymmetricdistributionofFzd,CELSR,PkandVANGL2,resulting
inthepolarizationofthecell16...............................................................................................................32
Figure6.Melanintransferfrommelanocytetokeratinocytebymelanosome.Different
formbetweenlightskinanddarkskin21...........................................................................................34

9
Figure 7. Schematic representation of relationships between fibroblasts and
melanocytes.................................................................................................................................................37
Figure 8. The different structure of DKKs family. DKK1, 2 and 4 have similar gene
sequence;DKK3andDKKL1havesamehomologyofsgy-domain34....................................38
Figure9.DKK1andDKK2interactwithWnt/β-cateninsignalingpathway.(a)Wnt,Fz,
and LRP6 form a complex to active the signaling pathway by increasing β-catenin
accumulation; (b)DKK1bindingwithLRP6 prevents the formationof complexand
inhibitsthesignaling38...............................................................................................................................39
Figure10.TheeffectsofDKK1 frompalmoplantar fibroblastsonbothmelanocytes
and keratinocytes. DKK1 binds to LRP5/6 against Wnt signaling pathway,
downregulatesMITFinmelanocytes,andatthemeanwhile,downregulatesthetransfer
proteinPAR-2inkeratinocytesanddecreasesthemelanintakenbykeratinocytes.On
theotherhand,DKK1enhancesthethicknessoftheskinbyupregulatingkeratin9in
keratinocytes31..............................................................................................................................................42
Figure11.ReconstructedskinmodelwithrhDKK1treatment(left)showedsignificant
lesspigmentationthanthecontrolgroup(right)31..............................................................44
Figure12.TGF-bfamilysignalingbytypeIIandIreceptors,andsmadproteins56......48

10
Figure 13. Hypothesis of the study: Structural damages, pro-inflammatory cytokines,
immune-modulatoryfactors(SASP)causedbyexternalstimulatedfactors(UVrays,air
pollution, environmental toxins) induce high secretion of TGF-b by fibroblasts and
inflammatorycells (Tcells,macrophages).Thisprocess inducesadownregulationof
DKK1geneexpressionandadecreaseofDKK1secretionbyfibroblasts.Lowexpression
andsecretionofDKK1leadstotheactivationofWnt/b-cateninsignalingpathway,and
resultsinincreasedmelaninsynthesisbymelanocytes..........................................................110
Figure14.TenseFPCLembeddedbyanylonmeshring...........................................................118
Figure15.Shematicrepresentationoftheirradiationprotocol..........................................119
Figure16.UVA-irradiationoftenseFPCL..........................................................................................119
Figure17.a)Melanin(HC,Masson-Fontanastaining)andb)β-catenin(IHC,DABstaining)
inSLandSNtissues.InSL,levelsofmelanin(a-1)andβ-catenin(b-1)↗comparedto
SN(a-2,b-2respectively).InSL,thebasallayerisdisrupted,causingmelaninreleasein
papillarydermis(a-2).MagnificationX20....................................................................................121
Figure18.RelativegeneexpressionofDKK1assessedbyreal-timequantitativeRT-PCR.
Toanalyzetherelativechangesingeneexpressionfromthereal-timePCRexperiments,
thecomparative2-△△Ctmethodwasused.The△CtvalueforFLandFNwascalculated
usingtheequation:Ct(DKK1)-Ct(reference,Abl).Thefoldchangeingeneexpression
ofDKK1wasfinallyobtainedfromtheformula2-△△Ct,wherethe△△Ctvaluewasthe

11
differencebetween△Ct(FL)and△Ct(FN)values.Therelativegeneexpressionisset
to1 forFNsamples.DKK1expression↘ inFLcomparedtoFN.Themean±SDwas
determinedfrom9independentexperiments,eachperformedinduplicate.***p<0,001.
...........................................................................................................................................................................122
Figure19.DKK1secretion into themediumbyELISA.DKK1secretion isnormalized to
proteincontentasmeasuredusingBiuretassay.Themean±SDwasdeterminedfrom9
independentexperiments,eachperformedinduplicate.DKK1proteinlevels↘inthe
mediumofFLcomparedtoFNbutnosignificantdifferencewasdetectedbetweenboth
groups............................................................................................................................................................123
Figure20.TGF-β1secretionintothemediumbyELISA.TGF-β1secretionisnormalizedto
cellnumber.Themean±SDwasdetermined from9 independentexperiments,each
performedinduplicate.TGF-β1levels↗inthemediumofFLcomparedtoFN.**p<0,01.
...........................................................................................................................................................................124
Figure21.Phasecontrast imagesofmelanocytescultures ina)normalconditions,b)FL
conditionedmediumandc)FNconditionedmedium.Arrowpointstomelanin.Normal
epidermal melanocytes exhibit a typical dendritic morphology and produce more
melaninwithFL-conditionedmediumtreatment(15%v/v).MagnificationX20.......127
Figure22.Immunofluorescenceimagesofmelanocytesculturesina)normalconditions,b)
FLconditionedmediumandc)FNconditionedmedium.Normalepidermalmelanocytes

12
exhibit a typical dendritic morphology and express MITF marker with FL or FN-
conditionedmediumtreatment(15%v/v).MagnificationX20...........................................128
Figure23.DKK1secretionintothemediumbyELISA.TGF-β1secretionisnormalizedto
cellnumber.Themean±SDwasdetermined from3 independentexperiments,each
performedinduplicate.DKK1levels↘inthemediumafterUVirradiation.***p<0,001.
...........................................................................................................................................................................129

13
ListofTables
Table1.Comparisonofthreeclassicmelanocyteextractionmethods.........................................86
Table2.PrimersandTaqmanprobesusedforqRT-PCR................................................................114
Table3.Preparationofdifferentconcentrationsoffibroblast-conditionedmedium.........115
Table4.OpticaldensitymeasurementsforMTTassayofviabilityofmelanocytescultured
withdifferent concentrationsof fibroblast-conditionedmedium.Themean±SDwas
determined from 3 independent experiments, each performed 4 times. Fibroblasts-
conditionedmediumfrombothFLandFNinconcentrationrangingfrom0to15%v/v
showednocytotoxiceffectonmelanocytes..................................................................................125

14
ListofAbbreviations
AMH:Anti-MüllerianHormone
APC:AdenomatousPolyposisColi
bFGF:basicFibroblastGrowthFactor
BMK1:BigMAPKinase1
BMPs:BoneMorphogeneticProteins
cAMP:cyclicAdenosineMonophosphate
CREB:cAMPResponsiveElementBindingprotein
DCT:DOPAChromeTautomerase
DHI:5,6-dihydroxyindole
DHICA:DHI-2-carboxylicAcid
DKK:Dickkopf
DMEM:Dulbecco’sModifiedEagle’sMedium
DMSO:DimethylSulfoxide
DOPA:L-3,4-dihydroxyphenylalanine
Dsh:Dishevelled
ERKs:ExtracellularSignal-regulatedKinases
FBS:FetalBovineSerum
FPCL:Fibroblast-populatedCollagenLattice
GDF:GrowthandDifferentiationFactors
Gsk-3b:GlycogenSynthaseKinase-3𝛽
HGF:HepatocyteGrowthFactor

15
JNK/SAPK:c-junN-terminalkinases
KGF:KeratinocyteGgrowthFactor
LDLR:Low-densityLipoproteinReceptor
LEF:LymphoidEnhancer-bindingFactor
MAPK:Mitogen-activatedProteinKinase
MATP:MyoacitveTeradecaPeptide
MTT:3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazoliumbromide
MITF:Microphthalmia-associatedTranscriptionFactor
NT-3:Neurotrophin-3
NRG-1:Neuregulin-1
PAR-2:Protease-activatedReceptor-2
PBS:Phosphate-bufferedSaline
PI3K/Akt:phosphatidylinositol3′-kinase/Akt
PKA:ProteinKinaseA
P/S:Penicillin/Streptomycin
R-Smads:Receptor-activatedSmad
rhDKK1:recombinantHumanDKK1
ROS:ReactiveOxygenSpecies
Sema7a:Semaphorin7a
SCF:StemCellFactor
SFRP:SecretedFrizzled-relatedProteins
TGF-b:TransformingGrowthFactor-beta
TRP-1:Tyrosinase-relatedProtein-1

16
TYR:Tyrosinase
WIf-1:WntInhibitoryFactor1

17
GeneralIntroduction
Our skin reflects our general health and well-being. It plays the role of interface
between our body and the outside environment, and protective barrier against external
aggressors.Thereforemanypeoplepayspecialattentiontotheirskin.Amongskindiseases,
pigmentation disorders are now a very important area of research in cosmetology and
dermopharmacy. Melanin plays key roles in determining human skin pigmentation.
Melanocytes are cells that possess the unique capacity to synthesize melanin within
melanosomes. This thesis focuses on the study of skin pigmentation in different views:
melanocyte,melaninsynthesis,andrelatedsignalingpathway.
The first chapter introduces the process of melanogenesis and the factors that regulate
melanin production in melanocytes. Transcription factors specifically expressed by
melanocytes and keratinocytes-derived paracrine factors regulate the functions of
melanocytes.Today,thereisgrowingscientificevidencesuggestingthatfibroblastssecrete
factors that are involved in the regulation of skin pigmentation. Therefore, a literature
reviewwaswritten todescribe therecent findingsonmelanogenic factorssecreted from
fibroblasts.
The second chapter describes and validates an extraction method of human skin
melanocytesbeing simple, effectiveandapplicable to smaller skin samples, andavoiding
animal reagents. This method would be suitable for establishment of optimal primary
melanocyteculturesforclinicalapplicationandresearch.
Lastly, the third chapter explores the roleof fibroblast-derivedparacrine factorDKK1 in
melanogenesiswith a focus on solar lentigo lesion. An hypothesis proposes that TGF-β1

18
mediatesdevelopmentofsolarlentigobyreducingDKK1expressioninfibroblaststhrough
thep38MAPkinasepathway,whichleadstoanactivationoftheWnt/b-cateninsignaling
cascade. The investigation is conducted with primary cultures of fibroblasts from solar
lentigo,anddermalequivalentmodelsexposedtoUVradiation.

19
Abstract
Thisthesisfocusesonthestudyofskinpigmentationindifferentviews:melanocyte,
melaninsynthesis,andrelatedsignalingpathway.
The first part of the experimental work was to develop a new extraction method of
melanocytes. Cell extraction is an inevitable and critical step in the development and
productionofAdvancedTherapyMedicinalProducts (ATMP) as for the establishmentof
primarycellbank.Thetechniquesdescribedintheliteratureareusuallybasedontrypsin,
aloneorincombinationwithdispase.Suchenzymesareusedinordertoseparatedermis
from epidermis and subsequently provide a suspension of epidermal cells. The
implementation of these protocols is often operator-dependent, not suitable for small
samplesandrequiresanimalderivedproducts(notcompatiblewithclinicalapproach).The
objectiveofthisworkwastodefineandvalidateanepidermalcellextractionmethodbeing
the simplest, the most effective and applicable to smaller samples and avoiding animal
reagents.Anewproduct(TrypLE,Lifescience),animal-freeproduct,hasbeentestedonskin
biopsies.Theextractionefficiencywasjudgedonthefollowingcriteria:separationepidermis
/dermis,culturedmelanocytes,functionalityoftheextractedcells.Resultsshowedtheease
of separation between the dermis and epidermis on the one hand, and between the
epidermalcellsontheotherhand.Aminimumsizeofskinsamplewasdefinedtoallowthe
extraction of functional melanocytes. In conclusion, this optimal method opens new
perspectives for establishment optimal melanocyte cell lines suitable for cutaneous
pathophysiologyresearchandproductionofATMP.

20
Thesecondpartoftheexperimentalworkwastoinvestigatethemechanismunderlyingthe
lesion of solar lentigo by exploring the role of Dickkopf-related protein 1 (DKK1) in
hyperpigmentation. Recent studies reported that high DKK1 expression by dermal
fibroblastsiscloselyrelatedtohypopigmentedskin,aspalmoplantarskinandvitiligolesion.
DKK1encodedbyDKK1geneisanantagonisticinhibitoroftheWntsignalingpathway,by
isolating the LRP5/6 co-receptor and preventing the Frizzeled-Wnt-LRP5/6 complex
formation.Wnt/bcateninsignalingregulatesthetranscriptionofmelanocyte-specificgenes
likeMITF, a gene involved inmelanin synthesis.Wehypothesized that TGF-β1mediates
developmentofsolarlentigobyreducingDKK1expressioninfibroblaststhroughthep38
MAPkinasepathway,whichleadstoanactivationoftheWnt/b-cateninsignalingcascade.In
vitromodelsthatmimictheinvivoenvironmentwereused:fibroblastsisolatedfromsolar
lentigoandperi-lesionalbiopsies,andnormalfibroblastsembeddedin3Dcollagengeland
exposedtorepeateddosesofUVA.Q-PCRandELISAtechniquesshowedthatfibroblastsfrom
solar lentigo and fibroblasts irradiated to UVA, express low level of DKK1.
Immunohistochemicalstudiesrevealedastrongstainingofbcateninandmelanininsolar
lentigo skin, which indicates an activation of Wnt/b catenin signaling and melanocyte
function.Ourpreviousdatademonstratedthesenescent-likephenotypeoffibroblastsfrom
solar lentigo with particularly a high secretion of TGF-β1, suggesting its role in the
development of the lesion. Fibroblasts should respond tomany inflammatorymediators
releasedduringhyperpigmentarydisorderbyincreasingTGF-β1secretion.TGF-β1isknown
tosuppresstheexpressionofDKK1inap38-dependentmanner.Therefore,ourhypothesis
confirmedthatTGF-β1regulatesdevelopmentofsolarlentigobyreducingDKK1expression

21
infibroblaststhroughthep38MAPkinasepathway.Thisprocessmayresultinanuneven
distributionofactivemelanocytes,withareasofhyperpigmentationintheskin.

22
ChapterI
LiteratureReview
23
I. Structureoftheskin
Skinasoneofthelargestorganofhumanbodybecomesthemostimportantbarrierbetween
humanbodyandexternalenvironment.Theaverageareaof theskin is to1.8m2,and its
average sickness is to 1.2 mm. It is responsible of three main functions: protection
(mechanical trauma, UV radiation, microorganisms, infection), regulation (temperature,
fluid,immunity,synthesisofvitaminD)andsensation(pressure/pain,heat/cold)1.
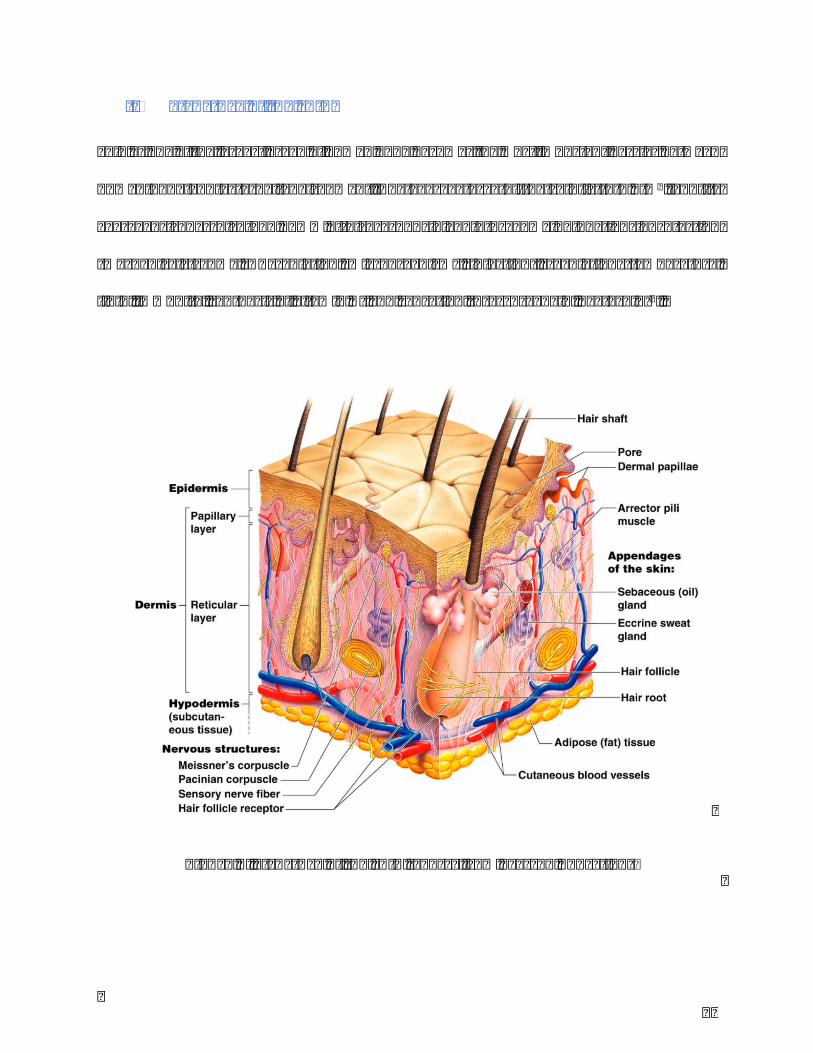
Figure1.Structureoftheskin(SourcefromPearsonEducation)
24
Theskiniscomposedbythreemainlayers:theepidermis,thedermisandthesubcutaneous
tissue(Figure1).
I.IEpidermis
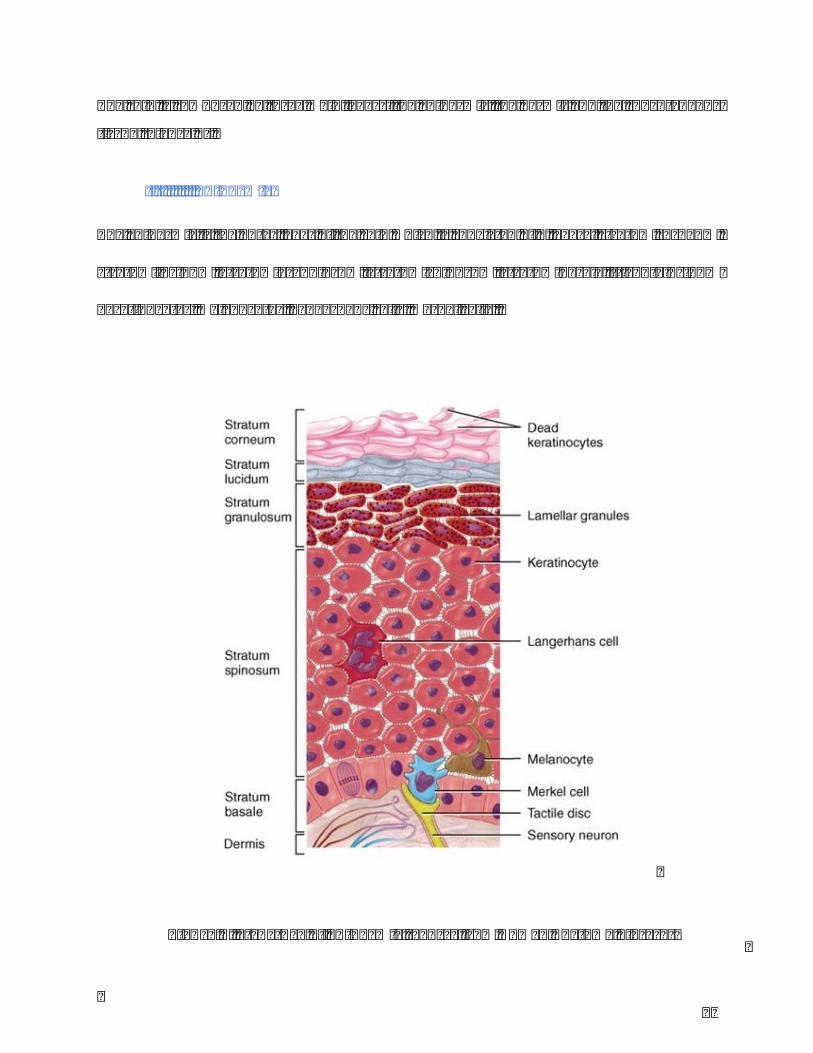
Theepidermisistheouterlayeroftheskinwhichisconsistedof5layers:stratumcorneum,
stratumlucidum,stratumgranulosum,stratumspinosum,stratumbasale.Itcontains95%
keratinocytes,melanocytes,Langerhanscells,Merkelcells.
Figure2.Structureofepidermis(SourcefromHumanAnatomyLibrary)

25
Keratinocytesoriginateandproliferatesfromdeepestlayeroftheepidermisupdatedtothe
surfacelayerstepbystep.Melanocytes,whichlocatedinbasallayerofepidermis,secrete
melanintoprotecttheskinfromUVradiation.Langerhanscell,areresponsibletotheskin
immunesystem.Merkelcellsdisplaythefeatureofsensoryreceptorcells(Figure2).
I.IIDermis
Thedermisisconnectedtotheepidermisthroughabasementmembrane.Itisdividedin2
layers:thesuperficialareaknownaspapillaryregion,anddeeperandthickerareaknownas
reticularregion.Itsskeletonstructureismainlyconstitutedbycollagenandelastinfibers,
extrafibrillarmatrix.Inaddition,therearealsohairfollicles,sweatglands,sebaceousglands,
lymphandbloodvessels.Fibroblastsaretheprincipaldermalcells.
I.IIISubcutaneoustissue
Subcutaneous tissue also known as hypodermis, consists of loose connective tissue, fat,
largerbloodvesselsandnerves.
II. SkinPigmentation
II.IMelanocyteanditscharacteristics
Melanocytesderivefromneural-crestcellslocatedonthebasallayeroftheepidermis,the
uvea of the eyes, inner ear, nervous system, heart, bones and so on. In epidermis, each
melanocyte connects with 30-40 associated keratinocytes to form a unit as “Epidermal
MelaninUnit”.Theproportionofmelanocytestokeratinocytesintheepidermalbasallayer
is1:10.1200melanocytespermm2areapproximatelycontainedinhumanskin.However,

26
the number of melanocytes in the skin of palms and soles are fivefold lower than
melanocytesnumberatotherpartoftheskin2.Italsodecreasesby10-20%perdecadesafter
30yearsold.Withsenescence,melanocytestrendtobelargerandmoredendritic,andto
havelesstyrosinaseactivity3.Melanocyteisnottheonlycellwhichcouldproducemelanin
inhumanbody,othercellssuchascellsofpigmentedepitheliumofretina,epitheliaofiris
andciliarybodyoftheeye,someneurons,adipocytescouldalsosynthesizemelanin4.
II.IIMelanogenesisandrelatedenzymesandmodulators
In melanocytes, melanin production takes place in cytoplasmic organelles named
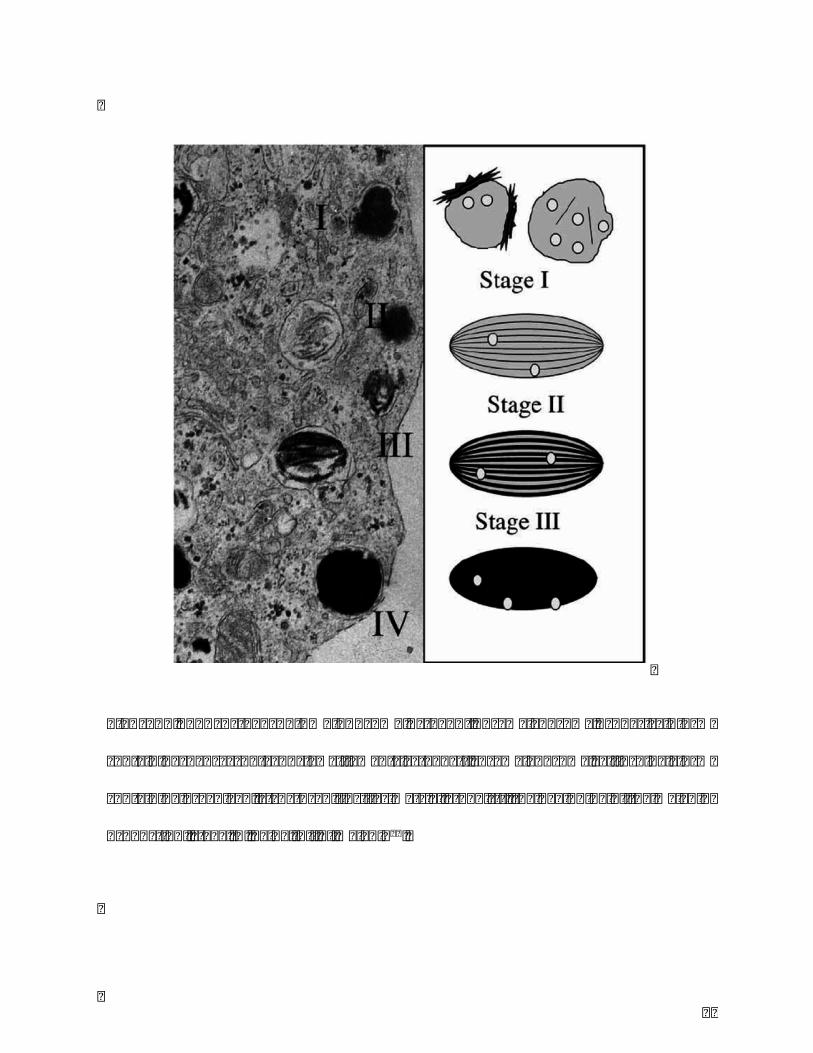
melanosomes. Itexperiences fourstages tobemature:stage I,premelanosome,spherical
form containingdense spot and few filaments; stage II, premelanosome, ellipsoidal form
containing organized, structured fibrillar matrix; stage III, the beginning of the melanin
production;stageIV,beingfullofmelanin5(Figure3).
27
Figure 3. Four stages of melanosome: stage I, premelanosome, spherical form
containing dense spot and few filaments; stage II, premelanosome, ellipsoidal form
containingorganized,structuredfibrillarmatrix;stageIII,thebeginningofthemelanin
production;stageIV,beingfullofmelanin99.

28
Theprocessofmelanin formationstartswiththerate limitingenzyme, tyrosinase.
TyrosineishydroxylatedtoL-3,4-dihydroxyphenylalanine(DOPA)bytyrosinase(TYR),then
rapidlyoxidizedtoDOPAquinone.Withcysteineandglutathione,cysteinylDOPAisformed
thenoxidizedandpolymerizedtopheomelaninwhichpresentsyellow-redsolublemelanin.
Intheabsenceofcysteineandglutathione,DOPAquinoneiscyclizatedtoDOPAchrome,then
transferedto5,6-dihydroxyindole(DHI)whichisoxidizedtothedark-brown-blacknamed
as DHI-melanin. On the other way, DOPAchrome transformes to DHI-2-carboxylic acid
(DHICA)byDOPAchrometautomerase(TYRP2/DCT),thenTYRP1catalyzesDHICAtoform
lighter brown color DHOCA-melanin. Both DHICA-melanin and DHI-melanin belonge to
Eumelaninwhichtypeplaysmoreimportantroleintheskincoloranddiversityofethnic
groups. Comparing to pheomelanin, eumelanin ismore effective in photoprotectionwith
higherresistancetodegradation,andabilitytoneutralizereactiveoxygenspecies(ROS)4,6
(Figure4).
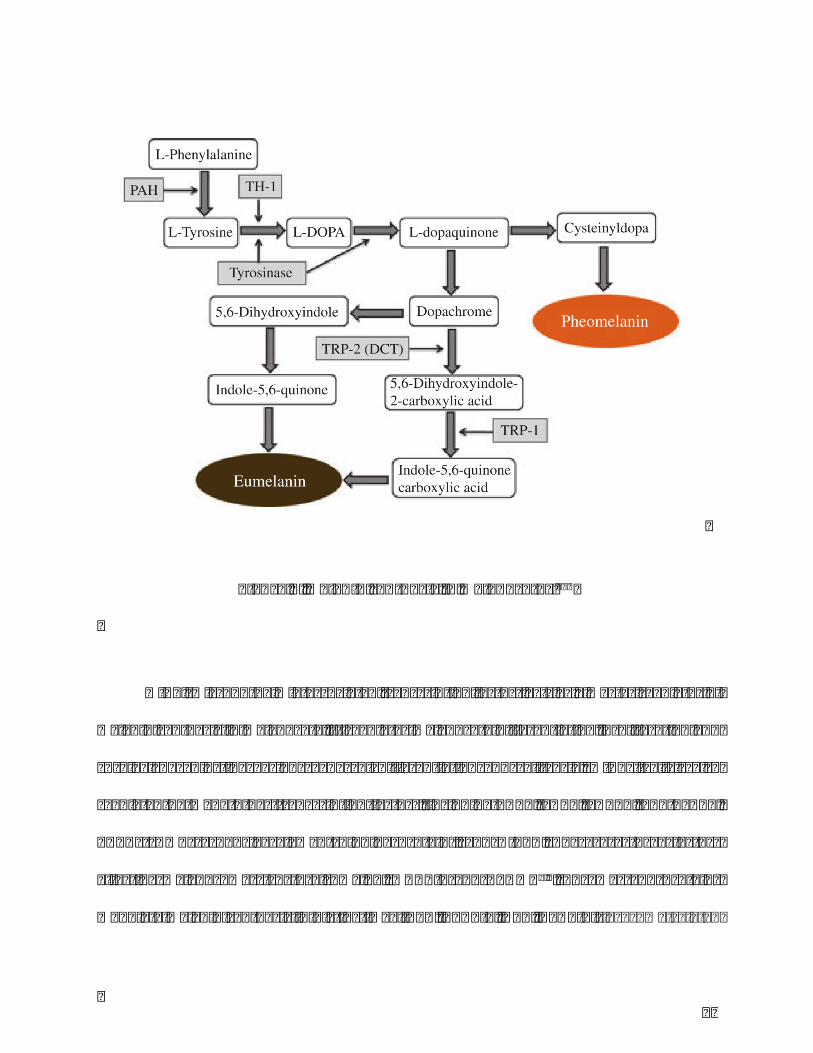
29
MITF(Microphthalmia-associatedtranscriptionfactor)actsasamasterregulatorof
melaninsynthesisinmelanocyte.Itregulatesmelanocytedifferentiation,proliferationand
survival by adjusting various genes of differentiation and cell-cycle. MITF itself is also
regulatedbymanyother transcription factors, includingPAX3, SOX9, SOX10,LEF-1/TCF,
CREB (cAMP responsive element binding protein), and DICER. There are three factors
affecting melanosome structure: Pmel17, MART-1 and GPNMB7,8. Enzymes and proteins
modulatemelaninsynthesisindistinctway:TYR,TYRP-1,DCT,BLOC-1(lysosome-related
Figure4.Melaninsynthesisinmelanocytes100

30
organelle complexe), OA1 (melanosomal G-protein-coupled receptor), P (membrane
transporters),andSLC45A2(implicatedinthecontrolofmelanosomeosmolarity)1.Some
proteinsinvolvemelanosometraffickingortransport:microtubules,F-actin,kinesin,dynein,
Rab27a,melanophilin,myosinVa,RILP5,9.
II.IIIMainsignalingpathwaysregulatingmelanogenesisinmelanocyte
a) MAPK/ERK (mitogen-activated protein kinases/extracellular signal-regulated
kinases) signaling is crucial to the proliferation and differentiation of melanocytes. The
kinases MEK and ERK in MAPK signaling pathway involve the activation of melanocyte
receptorsvialigandbindingtotheirextracellulardomain(eg,receptortyrosinekinasec-Kit).
Withbindingtotheirreceptors,ligandactivatescomplexmechanisms(Ras-Raf-MEK-ERK)
that lead to up-regulateMITF. Themutation ofMAPKs’ inhibitor such as BRAF affecting
MAPKstransductiongivesapotentialformelanoma10.TheMAPKsignalingpathwaybythe
synergistic action of factors derived from melanocytes and keratinocytes induces ERK
activation,whichtriggersmelanocyteproliferation11,12.
b) Wnt signaling pathway is involved in numerous developmental events during
embryogenesisandisessentialtokeeptissuehomeostasis.Italsoplayssignificantrolein
celldifferentiation.Thereare2typesofWntsignalingpathway:canonicalpathwayandnon-
canonicalpathway. Incanonical signalingpathway, the factorsconnectwithFrizzledand
WntcoreceptorLPR(Lowdensitylipoproteinreceptor-relatedprotein)5,6andregulate𝛽-
catenin by interactions with Axin/APC/GSK-3. In non-canonical signaling pathway, the
signal binding to Frizzled/LRP receptor transduces a signal to Ca2+or PCP (planar cell

31
polarity) without the participant of𝛽 -catenin. At the result, the accumulated𝛽 -catenin
interactswithTCF(Tcell factor)andregulates transcription.Thereare fiveextracellular
Wntantagonists:sFRP,WIF1,xenopuscerberus,DKKfamilyofsecretedproteins13,14(Figure
5).
Inmelanocytes,Wnt/β-cateninsignalingisanimportantpathwayinpigmentationprocess
andmelanocytesdifferentiation.ActivationofWnt/β-cateninsignalingoccursuponbinding
ofWnttofrizzledreceptorsandLRP5/6.Signalsaretransducedthroughtheinhibitionof
glycogensynthasekinase-3β(GSK-3β)activity,leadingtostabilizationandtransportofβ-
catenin into the nucleus that regulates transcription of MITF through interactions with
lymphoid enhancer-binding factor (LEF). Wnt signaling is modulated by secreted and
transmembraneWntinhibitorsandactivators13,15.
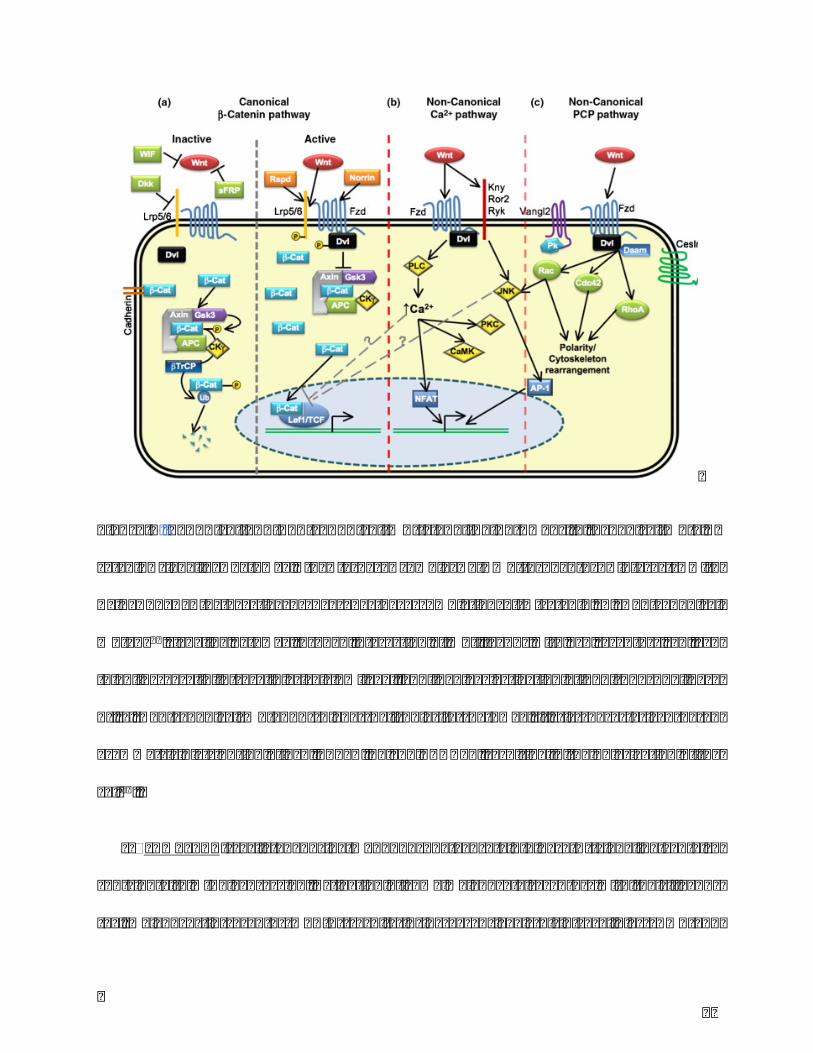
32
Figure5.Canonicalandnon-canonicalWntsignalingpathways.a.CanonicalWnt/𝛽-
catenin signaling pathway, the ternary complex of Wnt-Lrp5/6-Fzd interacts with
Axin/APC/GSK-3 to active or degrade the accumulation of𝛽 -catenin. b. Non-canonical
Wnt/Ca2+signalingpathwaythroughinteractionofWntligandswithFzdreceptorscanlead
toanincreaseinintracellularcalciumlevel,andinvolvesactivationofPLC(phospholipase
C). c. Non-canonicalWnt/PCP (Planar Cell Polarity) pathway, it’s is characterized by an
asymmetricdistributionofFzd,CELSR,PkandVANGL2,resultinginthepolarizationofthe
cell16.
c) cAMP/PKA(cyclicadenosinemonophosphate/proteinkinaseA) signalingcanalso
contributetoMITFexpression.Activationofsomemelanocytereceptorswiththeirligands
(eg, melanocortin receptorMCR-1) results in increased levels of intracellular cAMP and

33
activationofPKA.PKAphosphorylatesCREBwhichactsasatranscriptionfactorofMITF.It
has also been reported that activation of PKC (ProteinKinase C) can be associatedwith
cAMP-dependent pathway17. Various intrinsic and extrinsic factors affect melanogenesis
throughsignaltransductionpathways.Theyexerttheiractionsdirectlyonmelanocytesor
indirectlyviamediatorsproducedbysurroundingskincells12.
d) PI3K/Akt(phosphatidylinositol3′-kinase/Akt)signalingpathwayplaysacriticalrole
inproliferationandapoptosisofmelanocytethroughtheinhibitionofBAD(pro-apoptotic
memberoftheBcl-2family),aswellasadjustingcellcyclebyinactivatingGSK3andthen
activatingcyclinD1.ItalsocouldcooperatewithRAS-RAF-MEK-ERKpathwaytoeffecton
melanocyte activity. Because of the significant role of PI3K/Akt in melanoma by the
apoptosis resistance and cell proliferation, the utilization of its inhibitor was widely
developedintheclinicaltreatment18–20.
II.IVRoleofepidermalkeratinocytesinmelanogenesis
Melanosomeistheonlyorganellewhichcouldloadandtransmitmelanininthebasallayer
frommelanocytetokeratinocyte.Apartfromthetypeofmelanin,themelaninreleaseand
transferareanothersignificantfactorsinfluencingskincolor.Inlightskin,melanosomesare
released from melanocyte to keratinocyte in cluster form while in dark skin, they are
distributedindividually21(Figure6).
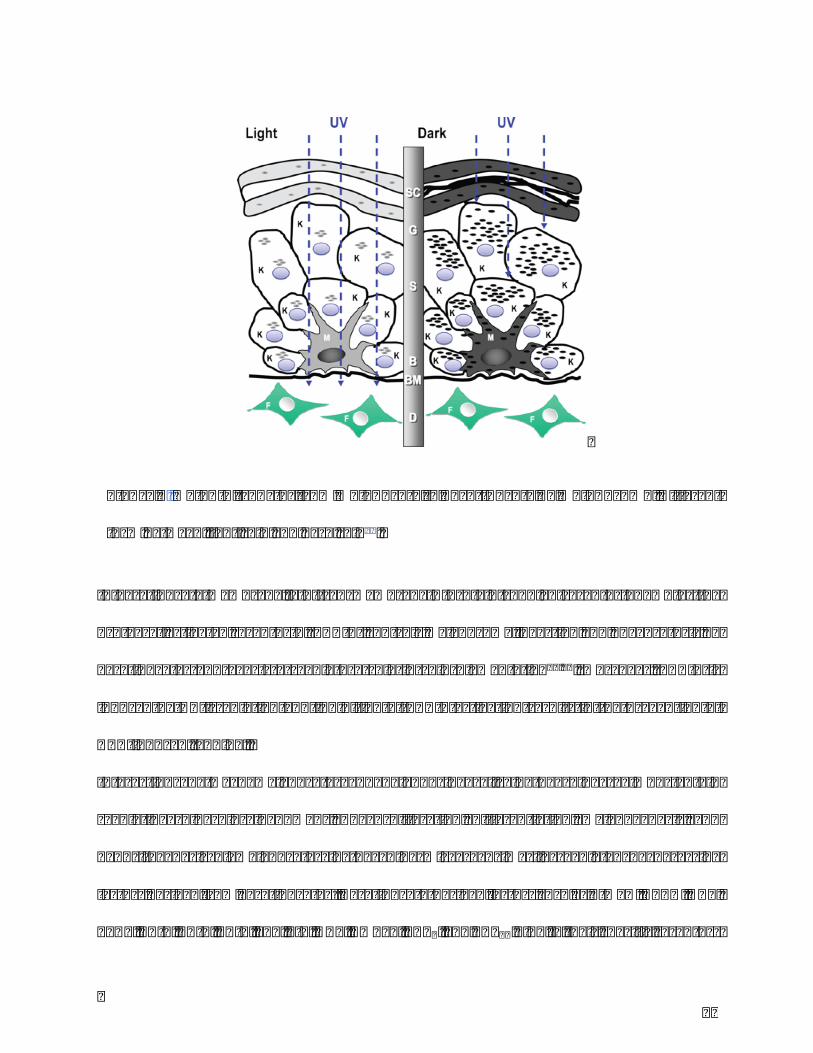
34
Inkeratinocytemembrane,a7-transmembraneG-protein-coupledreceptorknownas the
protease-activatedreceptor-2(PAR-2)controlsmelanosomeingestionandphagocytosisby
keratinocytes and exerts a regulatory role in skin pigmentation22,23. Moreover, PAR-2 is
inducedbyUV irradiationand inhibitionofPAR-2activation results in thepreventionof
UVB-inducedtanning.
Interactions between melanocytes and neighboring cells in the skin are important in
regulating skin color in humans. The proliferation, differentiation, melanogenesis, and
dendritogenesis of melanocytes in the epidermis are primarily regulated by paracrine
factorsderivedfromkeratinocytes.Keratinocyte-derivedfactorssuchasα-MSH,ACTH,NGF,
bFGF,ET-1,ET-2,ET-3,SCF,LIF,HGF,GMCSF,PGE2,andPGF2αbindtotheirspecificreceptors
Figure6.Melanintransferfrommelanocytetokeratinocytebymelanosome.Different
formbetweenlightskinanddarkskin21.

35
(e.g.,Mc1r,NGFR,FGFR-1,FGFR-2,ETBR,Kit,gp130,LIFRα,c-Met,GMCSFR,EP1,EP2,and
EP3)onthemembraneofmelanocytesandstimulatedifferentsignalingpathways,PKA,PKC,
andMAPK.
II.VRoleofdermalfibroblastsinmelanogenesis
Increasingevidencehasunderlinedthecontributionofdermalcomponentsintheregulation
ofpigmentation.Morerecently,dermalfibroblastsweredemonstratedtoexertaregulatory
roleonpigmentationthroughthesecretionofsolublefactors.
Factors secreted from fibroblasts bind to membrane receptors of melanocytes, active
differentsignalingpathwaysandmodulatemelanocytefunctionssuchasmelaninsynthesis.
Stemcell factor(SCF),hepatocytegrowthfactor(HGF),keratinocytegrowthfactor(KGF),
basic fibroblast growth factor (bFGF), neuregulin-1 (NRG-1), neurotrophin-3 (NT-3) and
semaphorin7a (Sema7a) target CREB promotor via MAPK signaling pathway and active
expressionof theMITF24–30.DKK1, an antagonist ofWnt/β-catenin signalingpathwayby
disruptingtheWnt-inducedFrizzle-Lrp5/6complex,preventstheWnt-inducedstabilization
of β-catenin and the binding to the LEF-1 transcription factor31. Secreted frizzle-related
protein (sFRP) activesWnt/β-catenin signaling pathway by binding to Frizzled receptor
whereasWntinhibitoryfactor-1inhibitsit32,33.WiththeadjustedpromotorofLEF-1,CREB
orPAX3,MITFcontrolstheactivityofTyrosinase,Tyrosinase-relatedprotein-1(TRP-1),
dopachrometautomerase (DCT)and therebymodulatesmelaninsynthesis.NRG-1,Nerve
growthfactor(NGF)andNT-3regulatePI3K/Aktsignalingpathway,andSema7aactivesFAK
andLIMKII,FAP-αandCCNwhichareassociatedwithmelanocyteproliferation,adhesion
andmigration5(Figure7).

36
Theroleofdermalfibroblastsinsolarlentigo(SL)lesionhasbeenrecentlydemonstrated.SL
is a common pigmentation disorder presented as aging spot on the exposure skin. It is
characterizedbyhyperpigmentedmacules,andthenumberofthespotsisrelatedtotheage
andskinphototype.ThehistologicalcharacterizationofSL isdefinedbyahighermelanin
depositioninthebasallayer,elongatedepidermalridgesandlargemelanosomalcomplexes.
Differentialgene-profilinganalysesbetweenSLandnormalskinbiopsiesrevealedthatSL
tissuesaremainlycomposedofactivatedmelanocytes34.Therearesomefibroblast-derived
paracrine factors such as HGF (Hepatocyte growth Factor), KGF (Keratinocyte Growth
Factor),SCF(StemCellfactor)involvedinSLlesionformation69,35.Immuno-staininganalyses
ofsomegrowth factorsandsecretedproteins in theupperdermisofSLbiopsiesstrongly
suggestthatdermalfibroblastscontributetofunctionallydysregulatingtheepidermalcells36.
Moreover, the treatmentwithKGFandKGFassociatedwithIL-1 induceshyperpigmented
lesionsinvivo37.
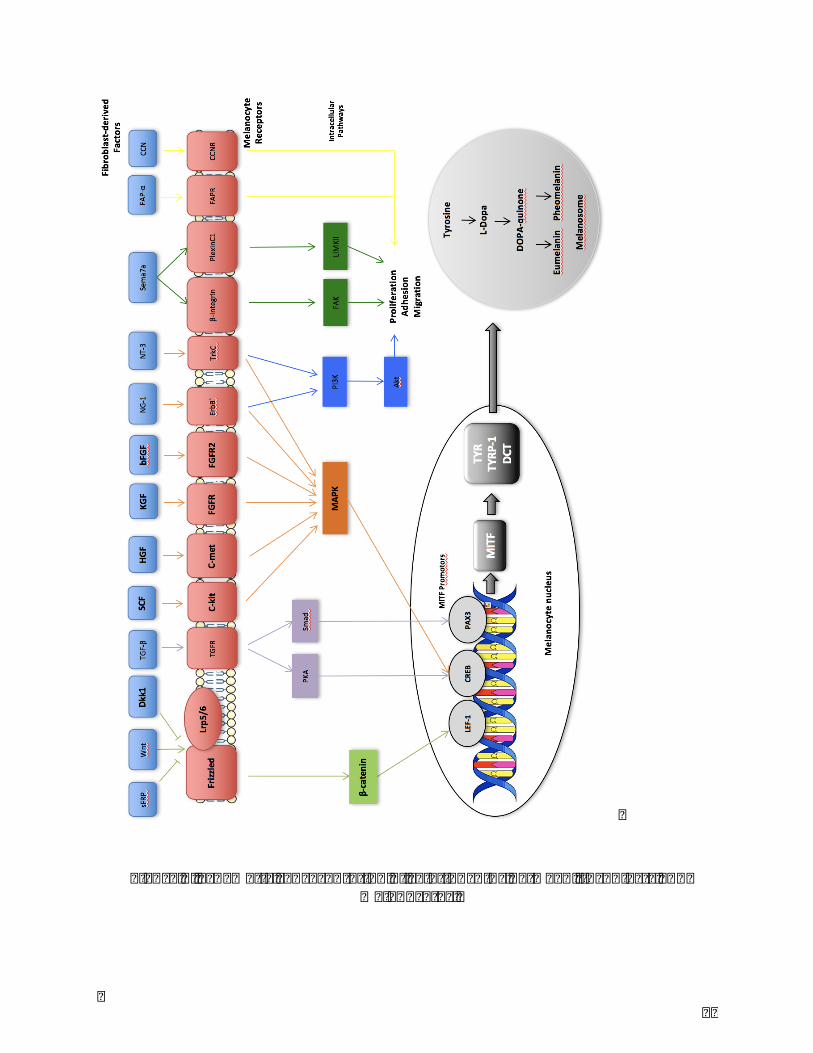
37
Figure7.Schematicrepresentationofrelationshipsbetweenfibroblastsandmelanocytes.

38
III. DKKsfamily
III.IStructure
Inmiceandhuman, thereweremultipleDKK(Dickkopf)genes.The familyofDKKgenes
includesDKK1,DKK2,DKK3,DKK4andDKKL1(Dickkopf-likeprotein1,alsocalledsoggy)
thatencodeDKKsecretedproteins.TheseDKKproteinsantagonizeWnt/𝛽-cateninsignaling
pathway by internalization withWnt coreceptor Lrp5 and 6, and by affinity ligands for
Kremen1and2transmembraneproteins.DKK1,DKK2andDKK4were identified tohave
similarsequence,4locatschromosome4/5/8/10paralogygroup,mappedto10q11,4q25
and8p11respectively.AtthemeanwhileDKK3andDKKL1showedthesamehomologyand
arelocatedonanotherparalogygroupmappedto11p15.338(Figure8).
Figure8.Thedifferent structureofDKKs family.DKK1,2and4have similargene
sequence;DKK3andDKKL1havesamehomologyofsgy-domain34.
39
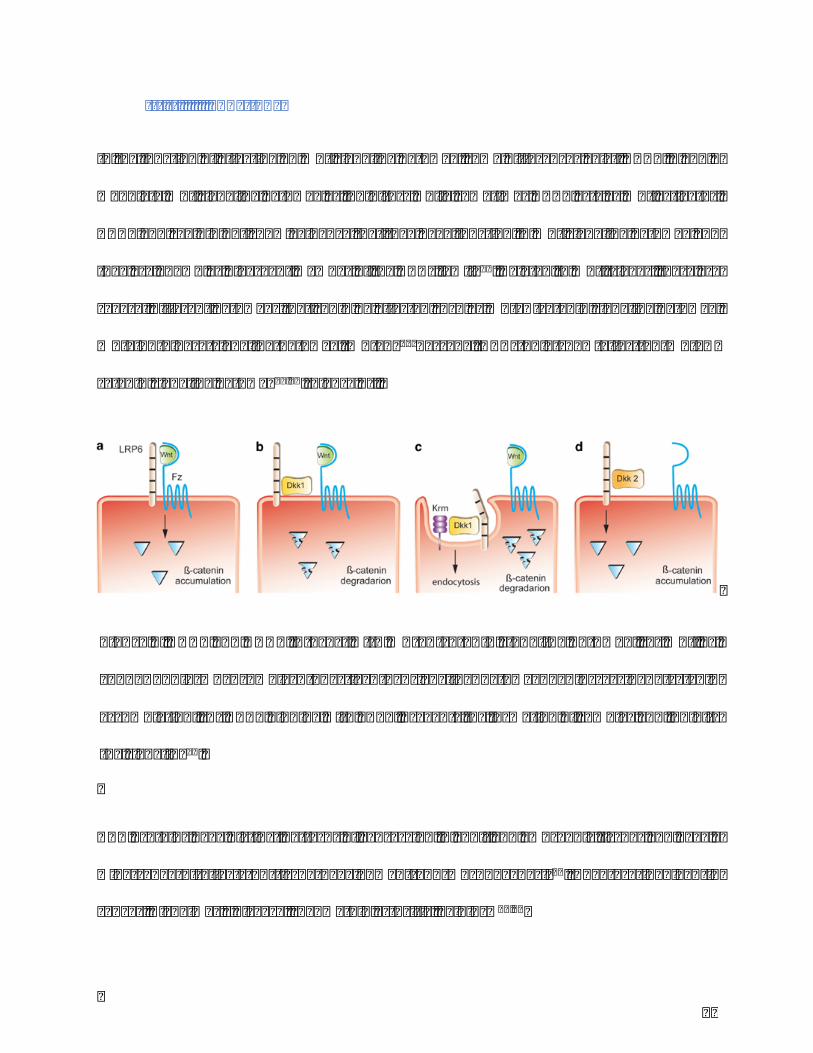
III.IIFunctions
InthefunctionofeffectingonWntsignalingpathway,somedifferencesexist.DKK1,2and4
modulateWntsignalingpathwayasinhibitorswhilesometimesDKK2actsasWntactivator.
DKK3andsgydidnotshowevidencethattheyhavefunctioninWntsignalingpathwayand
theyareknownasdivergentmemberoftheDKKsfamily39.RelatedtoWnttrigger,thereare
severaldifferentpathwaysthatcouldbeeffectedsuchasWnt/𝛽-cateninsignalingpathway,
Wnt/Junkinase signalingpathway,Wnt/Ca2+Cascade.DKKsonly showeffect onWnt/𝛽-
cateninsignalingpathway38,40(Figure9).
DKKproteinshavedistinctpatternsofexpressioninadultandembryonictissuesandhavea
wide range of effects on tissue development andmorphogenesis41. They are involved in
cancer,Alzheimer’sdisease,rheumatoidarthritis,keloids…41,42
Figure9.DKK1andDKK2interactwithWnt/β-cateninsignalingpathway.(a)Wnt,Fz,
and LRP6 form a complex to active the signaling pathway by increasing β-catenin
accumulation;(b)DKK1bindingwithLRP6preventstheformationofcomplexandinhibits
thesignaling38.

40
IV. DKK1
IV.IDKK1-LRP5/6complex
DKK1,as the foundingmemberof the family,was identifiedasa secretedprotein that is
required for head formation duringXenopous embryogenesis. InWnt signaling pathway,
DKK1 is the only antagonist that interacts with co-receptors known as LRP5 and LRP6,
contrarytoWntinhibitoryfactor-1andsecretedfrizzled-relatedproteinwhichdirectlybind
with Wnt43. LRP5 and LRP6 are specific coreceptors for DKK1. DKK1 binding to LRP6
interrupts the complex withWnt-Fz and causes the accumulation of𝛽 -catenin, thereby
blockstheWnt/𝛽-cateninsignalingpathway.
ItexistsanothertypeofDKK1coreceptorsknownasKremen1andKremen2,asingle-pass
transmembraneproteins.
IV.IIDKK1skinexpression
IthasrecentlybedemonstratedthatDKK1secretedbyfibroblastsinthedermiselicitsthe
hypopigmentedphenotypeofpalmoplantarskinduetosuppressionofmelanocytefunction
andgrowthviatheregulationoftwoimportantsignalingfactors,MITFandβ-catenin.2The
levels of DKK1 in palmoplantar dermal fibroblasts (at mRNA and protein levels) are
physiologicallyhigher than thoseobserved innon-palmoplantardermal fibroblasts.Thus
DKK1 has a role on skin pigmentation and thickness by inducing the palmoplantar
phenotype.
IV.IIIDKK1effectsonkeratinocytes
In previous study, transfected-DKK1 keratinocytes showed a stimulation of growth and

41
densityandaninhibitionofmelaninuptaken(lowexpressionofPAR-2)44.Inaddition,the
thicknessofareconstructedskinmodelwithkeratinocytestreatedbyrhDKK1(recombinant
humanDKK1)wassignificantly thicker 31,44.Therefore,DKK1regulatespigmentationnot
only on melanin synthesis and melanocyte activity but also on melanin uptaken by
keratinocytes(Figure10).
42
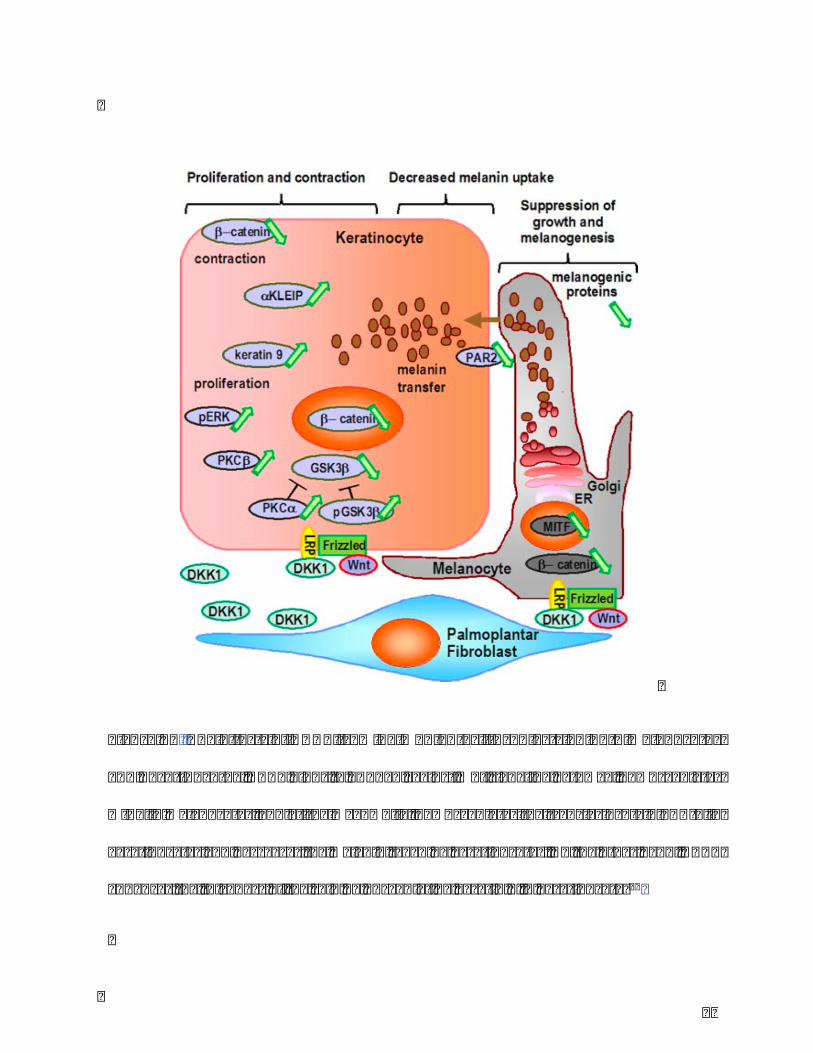
Figure10.TheeffectsofDKK1frompalmoplantarfibroblastsonbothmelanocytes
andkeratinocytes.DKK1bindstoLRP5/6againstWntsignalingpathway,downregulates
MITFinmelanocytes,andatthemeanwhile,downregulatesthetransferproteinPAR-2in
keratinocytesanddecreasesthemelanintakenbykeratinocytes.Ontheotherhand,DKK1
enhancesthethicknessoftheskinbyupregulatingkeratin9inkeratinocytes31

43
In rhDKK1 treated-keratinocytes, upregulation of a-KLEIP and keratin 9 and
downregulationofTulp3wereobserved.Keratin9 isspecificallyexpressedinsuprabasal
palmoplantarepidermisandisonlyobservedinacrosyringiainnonpalmoplantarepidermis.
Highexpressionofkeratin9inrhDKK1treated-keratinocyteconfirmsthatDKK1undirectly
regulates the thicknessofpalmoplantar skin45.aKLEIP is aprotein that is related to cell
adhesion and cytokinesis. High expression of aKLEIP shows that DKK1 regulates
proliferationandcelldensityofkeratinocytesbyincreasingcelladhesionandcytokinesis31.
Tulp3 is a protein belong to Tubby-like protein family which involves cell apoptosis.
DownregulationofTulp3suggeststhatDKK1regulateskeratinocytenumberbydecreasing
cellapoptosis31.
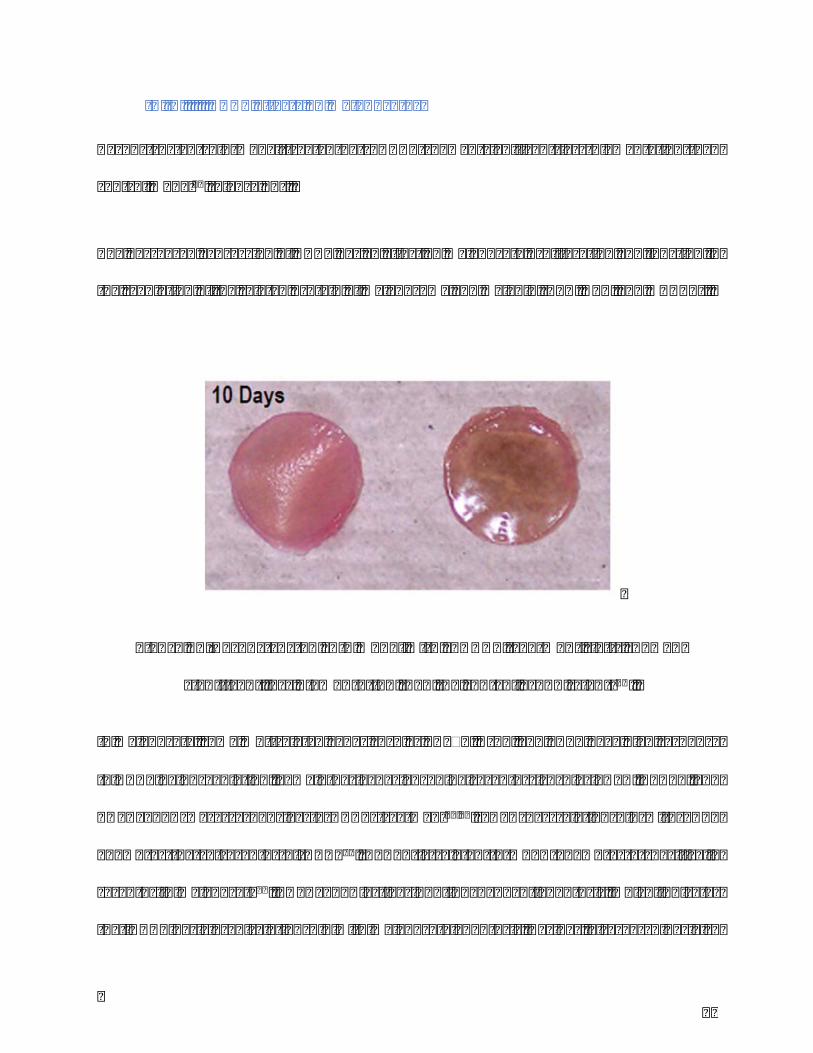
44
IV.IVDKK1effectsonmelanocytes
Reconstructed skinmodel treated by rhDKK1 showed significant less pigmentation than
controlmodel44(Figure11).
ThedecreasedexpressionofDKK1hasaneffectonmelanocyteproliferationandfunctionin
theregulationoftherelatedproteinofmelanosomeandmelanin(TYR,DCT,andMART1).
Inmelanocyte,someWnt-relatedgenessuchasPKCb1,Krn1,andLRP6havehighresponse
toDKK1byupregulation.SomeothergenesencodingreceptorssuchasLDLR,GPR51,and
TNFRSF10A were responsive to DKK1 treatment39,46. LDLR has relationship with LRP6
known as the co-receptor of DKK147. GPR51 is related toMC1R known as a specifically
receptorinmelanocyte48.TNFRSF10Aactivesp53-independentapoptosis,whichindicates
thatDKK1hasclosedrelationshipwithmelanocyteapoptosis.Besides,aresearchhasalso
Figure11.ReconstructedskinmodelwithrhDKK1treatment(left)showed
significantlesspigmentationthanthecontrolgroup(right)31.

45
foundthatDKK1upregulatesGadd45bandinducesmelanocyteapoptosisbyp38mitogen-
activated protein kinase pathway49. These studies suggest that DKK1 is involved in the
inhibitionofmelanocytegrowth.
In terms ofmelanosomal proteins, DKK1plays a significant role directly or indirectly in
upregulation of some expression proteins such as SVF3B, caveolin, syntaxin 5A, and
melanophilin. DKK1 also could directly influences tyrosinase by upregulating the critical
proteinmyoacitveteradecapeptide(MATP)inmelanocyte.Thereby,wecouldalsosuggest
thatDKK1haseffectonmelanocytedifferentiation39.
DKK1alsoaffectstheHOXrelatedgenesthatplayaroleinthegrowthanddifferentiationof
melanocytes,byupregulatingacidfibroblastgrowthfactor-likeprotein50.
IV.VRoleofDKK1invitiligo
Vitiligoisahypopigmentationdisorderinducedbydysfunctionordeficiencyofmelanocytes.
Theetiologyofvitiligoisstillunclearanditcouldco-existwithotherautoimmunediseases.
Mostresearcheshaveconcentratedontheabnormalityofmelanocytesandkeratinocytes
ratherthantheabnormalityoffibroblasts.Animbalanceofkeratinocyte-derivedcytokines
andadysregulationofinteractionswiththeirrespectivereceptorhavebeendemonstrated.
Thereisonlyonestudy,basedonfifteenskinbiopsies,ontherelationshipbetweenDKK1
andvitiligo.The immunohistochemistrystainingrevealed that theexpressionofDKK1in
dermisofvitiligoissignificantlyhigherthaninnon-lesionaldermis.ThemRNAlevelofDKK1
in fibroblasts from vitiligo was also higher than fibroblast from non-lesion51. At the

46
meanwhile,theexpressionofPAR-2indermisofvitiligowassignificantlowerthaninnon-
lesion.ThesefindingsindicatethatDKK1invitiligonotonlydecreasesmelanocyteactivity
butalsomelaninuptakenbykeratinocyte.Inaddition,thelowerexpressionofb-cateninin
vitiligo epidermis compared to non-lesional part confirmed that DKK1 causes
hypopigmentationbyWnt/b-cateninsignalingpathwayaspreviousstudies51–55.
IV.VIRoleofDKK1inmelanoma
Melanomaisthemostserioustypeofhumanskincancerthatdevelopsfrommelanocytes
andoccursanywhereonthebodyexposedtodirectsunlight.Themainreasoniscausedby
excessiveUVexposure thatdamagesDNA inmelanocytes.Melanoma implicatesnotonly
malignantmelanocytes,butalsoaheterogeneousmixofgeneticallystablenon-cancercells,
including fibroblasts, endothelial and inflammatory cells. DKK1 was proved to be a
diagnostic/prognosticserumbiomarkerindifferenthumancancers56.Recently,DKK1was
foundtobelinkedtomelanomadevelopment.ThelevelofDKK1inbloodserumofmelanoma
patient is significantly higher than those in healthy control. In vitro, DKK1 inhibits the
invasion of melanoma cells40. In B16F10 melanoma induced by viral-mediated tumor
transfection, DKK1 was found to inhibit melanoma development by decreasing tumor
angiogenesisandvascularperfusion57.

47
V. TGF-bfamily
V.ITGF-bandrelatedsignalingpathway
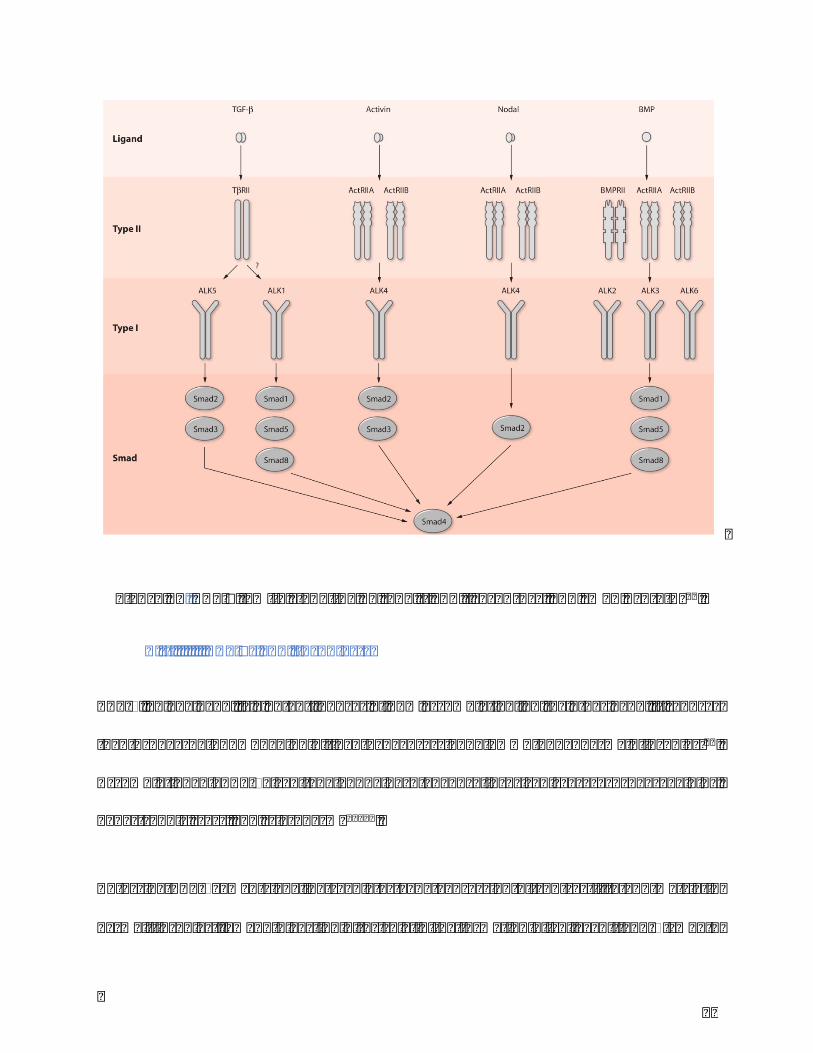
TGF-bsuperfamilycomprisesnearly30growthanddifferentiationfactorsincludingBMPs
(Bone morphogenetic proteins), GDFs (Growth and differentiation factors), AMH (anti-
müllerianhormone),Activin,NodalandTGF-bs(Figure12).Thesignalingisconductedby
transmembranereceptorstypeIandtypeII.Inresponsetoligandbinding,typeIIreceptors
(BMPRII,ActRIIA,ActRIIB,TβRII (1-3)) catalyze thephosphorylationofType I receptors
(ALK1-7). Activated type I receptors phosphorylate downstream Smads (Smad1, Smad2,
Smad3,Smad5,andSmad8),activatingthemtotransducethesignaltothenucleus.Italso
exists non-smad signaling pathway in stem cells. TGF-b signaling pathway is involved in
embryonic development, wound healing, angiogenesis by adjusting cell proliferation,
differentiation,andapoptosis56,58.
48
Figure12.TGF-bfamilysignalingbytypeIIandIreceptors,andsmadproteins56.
V.IITGF-bsandfibroblasts
TGF-b,ononehand,isapotentinducerofECMaccumulation;ontheotherhand,itreduces
the turnover of ECM by inhibiting the expression of MMP-1 by dermal fibroblast59.
Accumulation of TGF-b is often found in cutaneous fibrosis diseases such as keloids,
hypertrophicscars,andscleroderma60–62.
AutocrineTGF-b/Smad signaling regulates gene expressionof contractility andmatrix in
dermalfibroblast.Smad7playsinhibitorroleintheformationofcollagenIanda-smooth

49
muscleactin63.TGF-b1ishighlyexpressedbothinmRNAandproteinlevelsinhypertrophic
andscartissuederivedfibroblastscomparedtonormalfibroblasts,indicatingthepotential
ofTGF-b1intheformationhypertrophicscar64,65.
V.IIIp38-MAPKsignalingpathwayactivationbyTGF-b
MAPKsignalingpathwaysareidentifiedbyfourdifferentgroupsincludingERKs,JNK/SAPK,
ERK/BMK1andP38,whichgeneratediversecellularresponsessuchascellproliferation,
differentiation, migration and apoptosis. p38-MAPK is activated by a variety of cellular
stresses including inflammatory cytokines, growth factors, UV… Activated p38-MAPK
phosphorylates and activates transcription factors which are involved in DNA damage
response,inflammation,proliferation,apoptosis,cellularregulation.
TGF-bregulatesviap38/MAPKsignalingpathwaytheexpressionofcollagenase-3(MMP-13)
ingingivalfibroblasts66.
TGF-b is akey regulatorof fibroblast activation thatdrives the synthesisof extracellular
matrixinwoundhealingandfibroticdiseases61,64,66,67.TheroleofWntsignalingpathwayin
fibrosisaswellasscarringmakesaconsideringaboutthecrosstalkbetweentwosignaling
pathways.Infibroticfibroblast,TGF-bstimulatescanonicalWntsignalinginap38/MAPK
mannerbydownregulatingDKK168.

50
VI. Conclusion
Review published in Journal of Dermatological Science
http://dx.doi.org/10.1016/j.jdermsci.2017.06.018
Article1:Preciseroleofdermalfibroblastsonmelanocytepigmentation
YinjuanWangM.D.a,CélineViennetPh.D.a*,SophieRobinPh.D.b,Jean-YvesBerthon
Ph.D.c,LiHeM.D.,Ph.D.d*,PhilippeHumbertM.D.,Ph.D.a,e
aEngineeringandCutaneousBiologyLaboratory,UMR1098,UniversityofBourgogneFranche-
Comté,Besançon,France; bBioexigenceS.A.R.L,Besançon,France; cGREENTECHSA,Biopôle
Clermont Limagne, Saint Beauzire, France; dDepartment of Dermatology, First Affiliated
Hospital of Kunming Medical University, Kunming, China; eDepartment of Dermatology,
UniversityHospital,Besançon,France
*Correspondingauthors:CélineViennet,LiHe
E-mailaddresses:[email protected]/[email protected]
Abstract
Dermalfibroblastsaretraditionallyrecognizedassynthesizing,remodelinganddepositing
collagen and extracellularmatrix, the structural framework for tissues, helping to bring
thicknessand firmness to theskin.However, theroleof fibroblastsonskinpigmentation
arouses concern recently. More is known about the interactions between epidermal
melanocytesandkeratinocytes.
This review highlights the importance of fibroblast-derived melanogenic paracrine

51
mediatorsintheregulationofmelanocyteactivities.Fibroblastsactonmelanocytesdirectly
and indirectly throughneighboring cells by secreting a largenumber of cytokines (SCF),
proteins(DKK1,sFRP,Sema7a,CCN,FAP-α)andgrowthfactors(KGF,HGF,bFGF,NT-3,NRG-
1, TGF-β) which bind to receptors and modulate intracellular signaling cascades
(MAPK/ERK,cAMP/PKA,Wnt/-catenin,PI3K/Akt)relatedtomelanocytefunctions.These
factorsinfluencethegrowth,thepigmentationofmelanocytesviatheexpressionofmelanin-
producing enzymes and melanosome transfer, as well as their dendricity, mobility and
adhesive properties. Thus, fibroblasts are implicated in both skin physiological and
pathologicalpigmentation.Inordertoinvestigatetheircontribution,variousinvitromodels
havebeendeveloped,basedoncellularsenescence.UVexposure,amajorfactorimplicated
inpigmentarydisorders,mayaffectthesecretorycrosstalkbetweendermalandepithelial
cells.Therefore,identificationoftheinteractionsbetweenfibroblastsandmelanocytescould
providenovelinsightsnotonlyforthedevelopmentofmelanogenicagentsintheclinicaland
cosmetic fields, but also for a better understanding of the melanocyte biology and
melanogenesisregulation.
Highlights
• Cutaneouspigmentationisregulatedbyacomplexdermal-epidermalnetwork.
• Fibroblastsinteractwithmelanocytesviathesynthesisofbiochemicalfactors
• Fibroblasts-derivedfactorsbindtomelanocytereceptorsandmodulateintracellular
melanogenicpathways.
• The importance of fibroblasts-derived factors is demonstratedby skinpigmentary
changes.

52
1.Introduction
Dermalfibroblastsaretraditionallyrecognizedassynthesizing,remodelinganddepositing
collagenandnon-collagenextracellularmatrix(ECM),thestructuralframeworkfortissues,
helpingtobringthicknessandfirmnesstotheskin.Theycommunicatewitheachotherand
neighboring cellsby secretinga largenumberof cytokines andgrowth factors, playinga
crucialroleinskinphysiology.Intermofpigmentation,fibroblastsexhibitagreatdynamic
in theepidermalmelanogenesis, andparticipateactively in thesignal cross-talkbetween
melanocytes and keratinocytes.Most of previous research studied the regulation of skin
pigmentationbyfocusingonbothmelanocyteswhichsynthesizemelanin,andneighboring
keratinocyteswhichreceiveanddistributethepigmentinupperlayersoftheskin.Interest
infibroblastshasincreasedinrecentyearsduetotheirabilitytosecretemelanogenicfactors.
Thisreviewoutlinestheroleofdermalfibroblastsinconstitutivepigmentationandinthe
developmentofpigmentarydisorders.
2.Correlationbetweenfibroblastandmelanocytepigmentation
Studiesincreasinglyelucidatedthesignificantroleoffibroblastinpigmentation.Photoaged
fibroblasts in reconstructed skin model stimulate pigmentation, including both melanin
production andmelanogenic gene expression, compared to unexposed fibroblasts[1]. In
addition, Salducci et al. observed an increase of melanocytes number in reconstructed
epidermis cultured with conditioned media of UVA-treated fibroblasts[2]. However,
fibroblasts limit thespontaneouspigmentationofmelanocytes in3D- reconstructedwith
cells frompatient of phototype I and II, suggesting a role of fibroblasts in the control of
pigmentation[3].Murineandhumanfibroblastsdidnotsecretethesamemelanogenicand

53
mitogenicmelanocyte factors, resulting in adifferent effect onpigmentation. It hasbeen
shown that fetal fibroblasts in reconstructed skin model induce dramatic increase of
pigmentation compared to adult fibroblast, resulting in the elevation of melanogenic
mediators fromfetal fibroblasts.Tsuchiyamaetal. foundthatmultilineage-differentiating
stress-enduringcells,distinctstemcellsamonghumanfibroblasts,couldbereprogrammed
intomelanocytes.TheseMuse-derivedmelanocytesresideinthebasallayerofepidermisin
3D-skinmodel,andacquiremelanocyticfunctions.Thistechniqueshouldpermittreatment
of vitiligo by autologous transplantation[4].Moreover, Yang et al. successfully converted
mouse and human fibroblasts to functional melanocytes by combination of several
transcriptional factors, Microphthalmia- associated transcription factor (MITF), paired
domain and homeodomain-containing transcription factor 3 (PAX3) and SRY-related
transcriptionfactor10(SOX10)[5].Theseinduced-melanocytesproducemelanosomesand
melanin,delivermelanintokeratinocytesin3D-skinmodeland,evengeneratepigmentation
invivo.Theymayprovideanewefficientwaytotreatmelanogenicdysfunctions.Therefore,
allthesefindingsconfirmanimportantcross-talkingbetweenfibroblastsandmelanocytes
in pigmentation. Specifically, these are fibroblast-derived secreted factors which are
involvedinthefibroblastinteractionswithmelanocytes.
3.Signalingpathwaysoffibroblast-derivedfactorsinmelanocytes
Asknown, thepigmentmelanin includingdifferent types,pheomelaninandeumelanin, is
producedbymelanocytesinacomplexprocesscalledmelanogenesis:melaninsynthesisin
melanocytes,melanin transport frommelanocytes to keratinocytes bymelanosome, and
melanin distribution in epidermis. All factors related to this process can affect melanin

54
synthesis, including structural proteins of melanosome (Pmel17, MART-1, GPNMB),
enzymes required for melanin synthesis (tyrosinase (TYR), tyrosinase-related protein-1
(TYRP-1) and dopachrome tautomerase (DCT)), and proteins necessary formelanosome
transport and distribution (Rab27A, myosin Va, Slac2-a/melanophilin). MITF plays a
significantroleinmelanogenesisandmelanocytesdifferentiation,dendricity,proliferation
andapoptosis. Specifically,MITF regulates theexpressionofmelanogenicenzymes (TYR,
TYRP-1andDCT),melanosomalmatrix(Pmel17,Rab27)andanti-apoptoticproteins(bcl-
2).Signalingpathwaysplayakeyroleinrelayingextracellularsignalsfromfactorbindingto
cellmembranereceptortocellnucleusviaacascadeofphosphorylationevents.Fourcrucial
intracellular signaling pathways regulate melanocyte functions, and three of them are
associatedwiththeexpressionandfunctionofMITF[6,7].
3.1MAPK/ERK
MAPK/ERK (mitogen-activated protein kinases/ extracellular signal-regulated kinases)
signaling is essential to the proliferation anddifferentiation ofmelanocytes. The kinases
MEKandERKinMAPKsignal transductionpathway involvetheactivationofmelanocyte
receptorsvia ligandbindingtotheirextracellulardomain(eg,receptortyrosinekinasec-
Kit)[8].With binding to their receptors, ligand activates complexmechanisms (Ras-Raf-
MEK-ERK)thatleadtoup-regulateMITF[9,10].AmutationinthegenethatencodestheRAF
kinaseBRAF leads to constitutive activation of downstream signaling in theMAP kinase
pathway[11,12].
3.2Wnt/β-catenin

55
Wnt/β-catenin signaling is another important pathway in pigmentation process and
melanocytes differentiation, also for melanocyte stem cell[13–15]. Activation of Wnt/β-
cateninsignalingoccursuponbindingofWnttofrizzledreceptorsandlipoproteinreceptor-
relatedprotein5and6(LRP5/6).Signalsaretransducedthroughtheinhibitionofglycogen
synthasekinase-3β(GSK-3β)activity,leadingtostabilizationandtransportofβ-catenininto
thenucleus,where itregulatestranscriptionofMITFthrough interactionswith lymphoid
enhancer-bindingfactor(LEF).Wntsignalingismodulatedbysecretedandtransmembrane
Wntinhibitorsandactivators[16].
3.3cAMP/PKA
cAMP/PKA (cyclic adenosine monophosphate/protein kinase A) signaling can also
contributetoMITFexpression.Activationofsomemelanocytereceptorswiththeirligands
(eg, melanocortin receptorMCR-1) results in increased levels of intracellular cAMP and
activation of PKA[17]. PKA phosphorylates cAMP responsive element binding protein
(CREB)whichactsasatranscriptionfactorofMITF.Ithasalsobeenreportedthatactivation
ofPKCcanbeassociatedwithcAMP-dependentpathway[18].Variousintrinsicandextrinsic
factors affect melanogenesis through this signal transduction pathway. They exert their
actionsdirectlyonmelanocytesorindirectlyviamediatorsproducedbysurroundingskin
cells[18,19].
3.4PI3K/Akt
PI3K/Akt (phosphatidylinositol 3′-kinase/Akt) signaling pathway plays a critical role in
melanocyteproliferationandapoptosis throughthecellcycleregulationwithGSK-3and

56
proteincyclinD1[20,21],andthecontroloftheproapoptoticproteinBAD[22,23].Itcould
alsocooperatewithRas-Raf-MEK-ERKsignalingcascadeonregulatingmelanocyteactivity.
4.Fibroblast-derivedfactorsinvolvedinmelanocyteactivity
Fibroblasts release melanogenic factors which act both directly and indirectly on
melanocytes. Numerousmediators secreted from fibroblasts play significant roles in the
processof skinpigmentation throughdifferent signalingpathways. Someare involved in
down-regulation(DKK1),modulation(sFRP)andinduction(KGF,NRG-1)ofpigmentation,
someareinvolvedininductionofproliferationandsurvival(SCF,bFGF,NT-3,Sema7a,TGF-
β,CNN,FAP-α),andsomeareuniversalcontributor(HGF)(Table.1).
4.1Inhibitingfactor
-DKK1
Dickkopf(DKK)familycomprises4members(DKK1-4),andencodessecretedproteinsthat
antagonize Wnt signaling by inhibiting Wnt coreceptors Lrp5 and 6. It takes part in
numerousprocessesasboneformation,Alzheimer’sdisease,eyedevelopmentandalsoskin
pigmentation. DKK1 is produced by fibroblasts in skin and its regulatory role in
melanogenesiswas firstly described in 2004. Yamaguchi et al. reported thatmelanocyte
densityinpalmoplantarhumanskinwasfivetimeslowerthanthatinnonpalmoplantarpart.
FibroblastsexpresshighlylevelsofDKK1mRNAonthepalmsandsoles,andhighlylevels
mRNAofDKK-3innonpalmoplantararea[24].Infurtherstudy,Yamaguchietal.provedthat
DKK1hasaninhibitoryeffectonMITFexpressionwhichresultsmainlyfromthedecreased
activity of GSK-3β and β-catenin[25]. In addition, DKK1 up-regulates the expression of

57
myoactivetetradecapeptide(MATP)whichreduceTYRactivity[26].Therefore,DKK1acts
on melanocytes by suppressing proliferation and melanin production. These combined
effectsexplainthelowerpigmentationobservedonthepalmsandsoles.
4.2Modulatingfactor
-sFRP
Secreted frizzle-related protein (sFRP) family consists of 5 secreted proteins in humans
(sFRP1-5) thatmodulateWnt signaling by bindingWnt proteins and Frizzled receptors.
Early studies found that sFRPbinding toWntprevented theactivationofWnt receptors,
leading to the initial classificationof sFRPs asWnt signaling inhibitors[27-28].However,
subsequentfindinghassuggestedthatsFRP2functionsasamelanogenicstimulatorthrough
Wnt/β-catenin signaling, but the precisemechanismneeds to be clarified[29]. Kimet al.
highlighted a certain paracrine role of fibroblast-derived sFRP2 in pigmentation. A co-
culture experiment with sFRP2 over/downexpressed fibroblasts demonstrated that
fibroblast-derivedsFRP2increasedpigmentationinnormalhumanmelanocytes.Thereby,
the termofWnt-signalingmodulator is preferentially attributed to sFRP.Wnt inhibitory
factor-1(WIF-1)belongingtosFRPfamilyisalsoanagonistofWntsignalingpathway.Itwas
shown that WIF-1 increases pigmentation in melanocytes co- cultured with WIF-1
overexpressedfibroblasts[30].
4.3Activatingfactors-KGF
The keratinocyte growth factor (KGF) derived from fibroblasts participates in
melanogenesis process by inducing melanosome transfer. Interleukin-1 α (IL-1α), an

58
inflammatory mediator produced by keratinocytes after UVB exposure, stimulates
fibroblaststogenerateKGF.InsynergywithcAMP,transferrin,ET-1andbFGF,KGFincreases
differentiation,cellbodyexpansion,dendritesextensionandmelanosometransfer[31]. In
addition, KGF alone or in synergywith IL-1α and bFGF, inducesmelanin deposition and
elongatedreteridges[32].
-NGR-1
Neuregulin-1(NRG-1),anervegrowthfactorrelatedtothedifferentiationandmigrationof
neurons, is expressed differently among skin phototypes, and visibly increases skin
pigmentation[33]. A higher level of NRG-1 is expressed in 3D-skin equivalents included
fibroblastsfromtypeVI(darkskin).Inaddition,theamountandsizeofmelanocytes,aswell
asthicknessofdendritesareincreased[34].NRG-1bindsspecificallytoERBB3andERBB4
receptorsandactivatesPI3KandMAPKsignalingpathwaysinmelanocytes[35].
-SCF
Thecytokinestemcellfactor(SCF)issecretedconstitutivelybyfibroblasts.Thesolubleform
secreted from fibroblasts binds to the c-kit receptor of melanocytes and activates the
MAPK/ERK signaling pathway. SCF increases proliferation and differentiation of
melanocytes with or without factors produced by keratinocytes, as cAMP, ET-1 and
bFGF[36]. However the absence of SCF is correlated to a dysfunction of melanocyte
proliferation.ThesignalingSCF/c-kitisnecessarytotheviabilityofmelanocytes.Theuseof
anantibodyneutralizingc-kit(ACK2)inducesapoptosisofmurinemelanocytes[37].
-bFGF

59
Thebasicfibroblastgrowthfactor(bFGF,FGF2),amemberofthefibroblastgrowthfactor
family,issynthesizedbyfibroblastsandactsinaparacrinemanneronmelanocytesviaits
transmembrane receptor FGFR2 and the intracellular signaling MAPK pathway. bFGF is
mitogenicandmelanogenicformelanocytes[38].
-NT-3
Neurotrophin-3 (NT-3) belongs to a family of nerve growth factors, synthesized by
fibroblasts.Thesefactorshavebeenextensivelystudiedfortheirroleinthedevelopmentof
neuronsandneuralcrest-derivedcellssuchasmelanocytes.NT-3canlinkeachTrkreceptor
includingTrk-A,Trk-BandTrk-C,butmainlyplaysabiologicalfunctionbybindingtoTrk-C.
It modulates intracellular signal transduction through MAPK and PI3K- Akt pathways,
regulatingmelanocytedifferentiationandsurvivalrespectively[39].
-Sema7a
Semaphorin7a(Sema7a)fromsemaphorinfamily,alargeclassofsecretedandmembrane
anchoredproteins that is involved innumerousbiologicalprocesses, stimulatesdendrite
outgrowth frommelanocytes. Sema7a is a paracrine and UV irradiation-inducible ligand
expressedbyfibroblasts[40].PlexinC1andβ1-integrinsreceptorsareligandsforSema7a,
and signaling by these receptors has opposing effects on Sema7a-induced dendrite
formation.Sema7ainducesfocalFAKandMAPKactivationviaβ1-integrin,andstimulates
melanocyte spreading and dendricity in human melanocytes. It regulates negatively
melanocytedendricityviathereceptorPlexinC1.
-TGF-β

60
TheTransforminggrowthfactor-β(TGFβ)familyregulatesamultitudeofcellularprocesses,
including cell survival, proliferation and apoptosis[41]. It is secreted from various cells
includingfibroblasts.AfterbindingtoitstypeIandIIcellsurfacereceptors,TGF-βactivates
smadssignalingcascades.TGFβsignalinghasbeenshowntoexhibitarepressiveeffecton
bothmelanocytedifferentiationandmelanogenesisviadownregulationofMITFandPAX3,
and to influence quiescence of melanocyte stem cells[42–44]. It is interesting that both
TGFβ1 and TGFβ2 are upregulated by PAX3, and PAX3 itself is repressed by TGFβ1,
suggesting a negative feedback mechanism. On the other hand, TGFβ reduces CREB-
dependent transcriptionofMITFby repressionof PKA[45]. In termsof paracrine action,
TGF-βisapotentinhibitorofHGFsecretionfromfibroblasts.
-CCN
TheCCNfamilyisagroupofmultifunctionalsecretedproteinsdesignatedCCN1toCCN6.
CCN proteins regulate crucial biological processes by connecting cell surface and ECM.
Although they appear not to have specific high-affinity receptors, they signal through
integrins andproteoglycans. CCN2andCCN5aremostly expressed in thedermis[46,47].
CCN1isincreasinglyassociatedwithagegrowth[48].Aroleinskinpigmentationhasbeen
recently discovered. UV radiation upregulates CCN1-2 whereas CCN3-6 are
downregulated[49].
-FAP-α
Fibroblastactivationprotein-α(FAP-α),amemberofserineproteasefamily,isselectively
expressed in fibroblasts. It plays an important role on tumor spreading and is highly

61
expressedafterUVRtreatment[50].Thisprocesscouldbeupregulatedbyplateletderived
growthfactor-BB(PDGF-BB),TGF-β1,signalingproteinWnt5areleasedfrommelanocytes
andplasminogenactivatorfrommelanomacells[51,52].
-HGF
The hepatocyte growth factor (HGF, also known as scatter factor), highly expressed by
fibroblasts,bindstomelanocytereceptorc-METandtriggerstheMAPKandthePI3K-Akt
signaling pathways, modulating melanocyte proliferation, migration, and
melanogenesis[53,54]. The MAPK activates the ribosomal S6 kinase (RSK) family and
improves the phosphorylation of CREB protein. The secretion of HGF is stimulated by
keratinocytecytokines,IL-1αandtumornecrosisfactor-α(TNF-α)[55].
5.Fibroblastsinthedevelopmentofpigmentarydisorders
Alterationinmelaninproductionisthefundamentalchangeinpigmentarydisordersofthe
skin, which can be caused by defects of melanocytes, keratinocytes and fibroblasts.
Pigmentaryanomaliescanbeclassifiedaccordingwhetherthepigmentmelaninisincreased
(lentigo,melasma,café-au-laitmacules)ordecreased(vitiligo),withlocalizedorgeneralized
distribution(Table.2).
Theearlierscientiststhoughtthatkeratinocytesplaytheleadingrolesinpigmentation,and
fibroblastshavequitefewinfluenceonpigmentation.Forexample,membrane-boundSCF
derived from keratinocytes is more likely to increase the melanin production and
proliferation of melanocytes rather than soluble SCF derived from fibroblasts[56].
Fibroblastsareinvolvedinpathologicalpigmentation,byanalteredexpressionofvarious

62
factors. Levels of HGF, SCF and KGF in dermis of café-au-lait macules and freckles are
significantlyhigherthantheminnon-hyperpigmenteddermis[57].Thetanningresponseto
UVradiationexposureismediatedbyaspectrumoflocallyproducedcytokinesandgrowth
factors[58].Mutationsingenesencodingtheseregulatorsmodifytheirexpressionand/or
functionality, leading to altered signaling pathways, modified skin phenotypes, and
developmentofbenignlesionsortumors[59].Tobetterunderstandthedermalinfluenceon
skinpigmentation,thisreviewfocusesonsomecommonpigmentaryanomaliesthatinvolve
fibroblasts-derivedfactors.
5.1Melasma
Melasmaisacommonacquiredhyperpigmentationdisorder,occurredonforehead,cheeks
andmandible.ItaffectsappearanceandhashighlyincidenceinLatino,AsianandDark-skin
women.Thenumberofmelanocytesinmelasmalesionisincreasinghowevertheactivityof
melanin synthesis enhanced. The pathogenesis and etiology of melasma have not been
clearlyidentified,however,thepreviousresearchesidentifiedthehormonalfactors,family
history,sunexposureandcosmeticsasthefourmaintriggeringandaggravatingfactorsfor
melasmadevelopment[60].SomecytokinesinepidermissuchasSCF,PGE2,ET-1arehighly
expressed in epidermis of melasma lesion[61]. The paracrine linkage between dermal
fibroblasts and melanocytes also played an important role in the mechanism of
hyperpigmentation in melasma. UV-repeated radiations stimulate directly or indirectly,
throughkeratinocyte-derivedcytokines,thesecretionofsolubleSCFbydermalfibroblasts.
UVAradiationsinduceweakereffectonSCFsecretionthanthoseofUVB.Kangetal.found
thatSCFexpressioninthedermisofmelasmalesionissignificantlyincreased,itsreceptorc-

63
kitonmelanocytesisupregulatedindermisofmelasmaaswellbyimmunohistochemical
staining and RT-PCR[61]. Inmelasma, CCN proteins could be regulated by some factors
relatedtopigmentarydiseasessuchasFGF2,ET,estrogenanditsreceptors,progesterone
and its receptors. Fibroblasts extracted frommelasmaexpressmoreNGF-β compared to
fibroblastsfromperilesionalskin.Recently,researchersshowedthatexpressionofWIF-1is
significantly reduced in melasma lesion. Downregulation of WIF-1 in fibroblasts was
reportedtostimulatetyrosinaseexpressionandmelanosometransfer[62].
5.2SolarLentigo
Solar lentigo (SL) is a common pigmentation disorder presented as aging spot on the
exposureskin.Itischaracterizedbyhyperpigmentedmacules,andthenumberofthespots
isrelatedtotheageandskinphototype.ThehistologicalcharacterizationofSLisdefinedby
a higher melanin deposition in the basal layer, elongated epidermal ridges and large
melanosomalcomplexes.Differentialgene-profilinganalysesbetweenSLandnormalskin
biopsies revealed that SL tissues are mainly composed of activated melanocytes[63].
Overexpression ofHGF has been consistently proved in lentigo, and its receptor, c-MET,
highlyexpressedinmetastaticmelanoma[64].Therearesomefibroblast-derivedparacrine
factorssuchasHGF,KGF,SCFinvolvedinSLlesionformation[65].Immuno-staininganalyses
ofsomegrowthfactorsandsecretedproteinsintheupperdermisofSLbiopsiesstrongly
suggest that dermal fibroblasts contribute to functionally dysregulating the epidermal
cells[66].KGFsynergywithIL-1networkdescribedbeforeeffectwasprovedinsolarlentigo
aswell[67].KGFshowsalsoefficientlyabilitytopromotetheproductionandsecretionof
SCFinkeratinocytes[65].

64
5.3Vitiligo
Vitiligoisahypopigmentationdisorderinducedbydysfunctionordeficiencyofmelanocytes.
Theetiologyofvitiligoisstillunclear.Mostresearcheshaveconcentratedontheabnormality
ofmelanocytesandkeratinocytesratherthantheabnormalityoffibroblasts.Lowexpression
of SCF receptor c-kit in melanocyte at the edge of the lesion may indicate that
downregulationofSCF/c-kit/MAPKsignalingpathwaycontributestodysfunctionandloss
ofmelanocytes[56].ItwasreportedthatDKK1secretedfromfibroblastishighlyexpressed
invitiligolesionand,β-cateninandPAR-2expressioninmelanocyteissignificantlylowerin
vitiligo lesion compared to non-lesion skin[68-69]. The dysregulation of Wnt signaling
pathway in vitiligo lesion could prevent melanocyte differentiation. The occurrence of
vitiligocouldalsobeassociatedwithsomeimmunesystemdisorders[70]suchasthyroid
disease, it has effectively been proved that DKK1 could increase the expression level of
thyrotrophic embryonic factor and mitochondrial ribosomal proteins[71]. Furthermore,
attachment of melanocytes to collagen IV is mediated through collagen-receptor DDR1
(Discoidindomainreceptor1)whichisunderthecontrolofCCN3.ThedysfunctionofCCN3
andDDR1 interaction in vitiligo lesion causesweakly adhesion ofmelanocytes, and this
processinteractswithTGF-βandCCN2[72].
5.4Melanoma
Melanomaisthemostserioustypeofhumanskincancerthatdevelopsfrommelanocytes
andoccursanywhereonthebodyespeciallyinareasofthebodythatareexposedtodirect
sunlight. The main reason is caused by excessive UV exposure that damages DNA in
melanocytes. Melanoma implicates not only malignant melanocytes, but also a

65
heterogeneousmixofgeneticallystablenon-cancercells,includingfibroblasts,endothelial
and inflammatory cells. Infiltrated and surrounding stromal fibroblasts are recruited,
perpetually activated through paracrine factors released from melanoma cells and
transdifferentiatedintocancer-associatedfibroblasts(CAFs).OnemajorsourceofCAFsin
melanoma is resident normal skin fibroblasts. CAFs exhibit both morphological and
functional differences compared to normal fibroblasts. They acquire properties of
myofibroblasts, produce various growth factors, cytokines, and ECM proteins. Therefore
CAFsparticipateinthegrowthandinvasionofthetumorcellsbypromotingangiogenesis,
inflammation and metastasis[73]. However, the exact mechanisms of how normal skin
fibroblastsinteractwithmalignantmelanomacellsandsubsequentlytransformtoCAFs,are
poorlyunderstood[74].Melanomacellsanddermalfibroblastscommunicatethroughoutthe
tumorigenicprocess,includingexpressionofnumerouschemokines(IL-6,IL-8,CXCL1−3,
CXCL8,CCL5,CCL2...)andchemokine-receptors (CXCR4,CXCR2,CXCR3,CCR7andCCR10
and CXCL1...), and deregulation of signaling pathways (MAPK - PI3K/AKT, Wnt/β-
catenin)[75–77]. CAFs secretion of HGF results in activation of the c-MET receptor and
signaling pathways[78]. Expression of SCF, bFGF and KGF is involved in melanoma
genesis[79,80].UVexposureinducesupregulationofFAP-αinfibroblasts,andcontributes
tomigrationandinvasionofmelanomacells[52].HGFreceptor,c-MET,highlyexpressedin
metastaticmelanomapathwaymaintainsthemelanocytesurvivalbysuppressingthepro-
apoptoticBadprotein and increasing the anti-apoptoticBcl-xl[38].Nuclear Factor-kappa
(NF-kB), a transcription factor involved in the immune response, has been shown to be
upregulated inmelanoma throughderegulations inupstreamsignalingpathwayssuchas
Ras/Raf,PI3K/Akt.Therefore, targeting stromal fibroblasts and inhibiting stromalNF-kB

66
signaling could be a possible treatment formelanoma [35,50,81,82]. NT-3mediates cell
invasionandECMdegradationinmetastaticmelanoma[81].Additionally,developmentand
progressionofmelanomaareassociatedwithderegulationofWntsignalingandlossofDKK-
1 secretedby fibroblasts [83–85].DKK-1 isknown tobe regulatedby the canonicalWnt
pathway.Innormalmelanocytes,WntpathwayisactivatedbysecretionofWnt3a.However,
inmalignantmelanocytes,Wnt5awhich is an inhibitor of DKK-1, is expressed. It is also
suggestedthatphenotypicchangesofmelanocytesinmelanomatumorareassociatedwith
senescence,particularlyinthedermalcompartment.Thesecretionoffactorsfromsenescent
fibroblasts,suchasMMP-3,IL-6,TGF-,altersepithelialdifferentiation,promotesendothelial
cellmotilityandstimulatescancercellgrowthandtumorigenesis[86].Inaddition,senescent
fibroblasts accumulate during aging. Secreted sFRP of aged dermal fibroblasts drive
melanoma promoting both angiogenesis and metastasis[87]. Besides, loss of Sema7a
receptorPlexinC1mayinducemelanomainvasionandmetastasis,andthereforePlexinC1
isknownasapotentialtumorsuppressorformelanomaprogression[88].Downregulation
ofCCN3inmelanomacellscontributestotheirinvasivephenotype[89].
5.5Dermatofibroma
Dermatofibroma(DF),alsoknownasfibroushistiocytoma,isabenigncommoncutaneous
nodule of fibroblast-like cells with unknown etiology. Histologically, DF lesion is
characterizedbyahyperpigmentationintheoverlyingepidermiswithacanthosisandwith
an increased number of melanocytes. Proliferating fibroblast-like cells and histiocytes
accumulate in the dermis and are surrounded by mature collagen and by increased
capillaries. They secrete elevated gene and protein levels of SCF andHGF cytokines and

67
stimulate melanocytes located in the adjacent epidermis, which results in the
hyperpigmentationoftheoverlyingskin[50].ThealteredexpressionofSCFandHGFinDF
lesionscanbeassociatedwiththetumorcellproliferationandinductionofDF.SCForHGF
derivedfromthefibroblastictumormayfunctionasamitogenformelanocytes[90].SCFis
alsoknownasmastcellgrowthfactorandthisexplainswhyaccumulationanddegranulation
ofmastcellsareobservedwithmelanocyteactivationinDFlesions.However,furtherstudies
arerequiredtoidentifythefactorsthatstimulateexclusivelyinabundancethesecretionof
SCFandHGF.
6.Conclusion
Cutaneouspigmentationisregulatedbymelanogenicfactors,locallysynthesizedintheskin
bybothkeratinocytesandfibroblasts,orproducedbydistanttissuesandtransportedtothe
skin by the circulation. Many of those factors regulate constitutive and induced
pigmentation. In this review, the contribution of dermal fibroblasts to the regulation of
melanocyteactivities isdemonstrated.Fibroblasts interactwithmelanocytesdirectlyand
indirectlythroughkeratinocytes.Theyreleasenumerousbiochemicalfactorsthatmodulate
thepigmentedstatusofmelanocytesbyactivatingsignalingcascades,geneexpressionand
enzymeactivity.Levelsof fibroblast-derivedmelanogenicparacrinemediatorsdependon
intrinsicandextrinsicfactorssuchasskintype,genetic,sunexposition.Increasedresearches
on the importance role of fibroblasts in melanocyte function under physiological or
pathologicalconditionsprovidetheoryevidencesfortreatmentofpigmentarydisorders.
Financialsupport

68
ThisworkwasfinanciallysupportedbyProgramforInnovativeResearchTeaminUniversity
ofMinistryofEducationofChina(GrandNo.IRT13067)andTheFundofYunnanProvince
ChineseAcademyofSciencesCooperation(GrandeNo.2014IB008)(China)andGreentech
SA(France).
Conflictsofinterest
Theauthorshavenoconflictofinteresttodeclare.
References
[1]C.Duval,C.Cohen,C.Chagnoleau,V.Flouret,E.Bourreau,etal.,Keyregulatoryrole
ofdermalfibroblastsinpigmentationasdemonstratedusingareconstructedskinmodel:
Impactofphoto-aging,PLoSOne.9(2014)e114182.
[2] M. Salducci, N. André, C. Guéré,M.Martin, R. Fitoussi, et al., Factors secreted by
irradiatedagedfibroblastsinducesolarlentigoinpigmentedreconstructedepidermis,
PigmentCellMelanomaRes.27(2014)502–504.
[3] S.J.Hedley,C.Layton,M.Heaton,K.H.Chakrabarty,R.aDawson,etal.,Fibroblasts
playaregulatoryroleinthecontrolofpigmentationinreconstructedhumanskinfrom
skintypesIandII,PigmentCellRes.15(2002)49–56.
[4] K. Tsuchiyama, S. Wakao, Y. Kuroda, F. Ogura, M. Nojima, et al., Functional
melanocytes are readily reprogrammable from multilineage-differentiating stress-
enduring(muse)cells,distinctstemcellsinhumanfibroblasts,J.Invest.Dermatol.133
(2013)2425–35.

69
[5] R.Yang,Y.Zheng,L.Li, S.Liu,M.Burrows,etal.,Direct conversionofmouseand
humanfibroblaststofunctionalmelanocytesbydefinedfactors,Nat.Commun.5(2014)
5807.
[6]H.R.Widlund,D.E.Fisher,Microphthalamia-associatedtranscriptionfactor:acritical
regulatorofpigmentcelldevelopmentandsurvival,Oncogene.22(2003)3035–3041.
[7]S.A.N.DMello,G.J.Finlay,B.C.Baguley,M.E.Askarian-Amiri,Signalingpathwaysin
melanogenesis,Int.J.Mol.Sci.17(2016)E1114.
[8]T.J.Hemesath,E.R.Price,C.Takemoto,T.Badalian,D.E.Fisher,MAPkinaselinksthe
transcription factor Microphthalmia to c-Kit signalling in melanocytes, Nature. 391
(1998)298–301.
[9]J.Yan,S.Roy,A.Apolloni,A.Lane,J.F.Hancock,Rasisoformsvaryintheirabilityto
activateRaf-1andphosphoinositide3-kinase,J.Biol.Chem.273(1998)24052–6.
[10]R.Buscà,P.Abbe,F.Mantoux,E.Aberdam,C.Peyssonnaux,etal.,Rasmediatesthe
cAMP-dependent activation of extracellular signal-regulated kinases (ERKs) in
melanocytes,EMBOJ.19(2000)2900–10.
[11] D.B. Solit, L.A. Garraway, C.A. Pratilas, A. Sawai, G. Getz, et al., BRAF mutation
predictssensitivitytoMEKinhibition,Nature.439(2006)358–362.
[12]A.A.Marusiak,Z.C.Edwards,W.Hugo,E.W.Trotter,M.R.Girotti,etal.,Mixedlineage
kinasesactivateMEKindependentlyofRAFtomediateresistancetoRAFinhibitors,Nat.

70
Commun.5(2014)195–201.
[13]L.Vibert,G.Aquino,I.Gehring,T.Subkankulova,T.F.Schilling,etal.,Anongoingrole
forWntsignalingindifferentiatingmelanocytesinvivo,PigmentCellMelanomaRes.30
(2017)219–232.
[14] H. Guo, Y. Xing, Y. Liu, Y. Luo, F. Deng, et al.,Wnt/β-catenin signaling pathway
activatesmelanocytestemcellsinvitroandinvivo,J.Dermatol.Sci.83(2016)45–51.
[15]X.Lim,R.Nusse,Wntsignalinginskindevelopment,homeostasis,anddisease,Cold
SpringHarb.Perspect.Biol.5(2013)a008029.
[16] C.-M. Cruciat, C. Niehrs, Secreted and Transmembrane Wnt Inhibitors and
Activators,ColdSpringHarb.Perspect.Biol.5(2013)a015081.
[17]W.-R.Lee,S.-C.Shen,P.-R.Wu,C.-L.Chou,Y.-H.Shih,etal.,CSE1LLinkscAMP/PKA
andRas/ERKpathwaysandregulatestheexpressionsandphosphorylationsofERK1/2,
CREB,andMITFinmelanomacells,Mol.Carcinog.55(2016)1542–1552.
[18] M.S. Feschenko,E. Stevenson,K.J. Sweadner, InteractionofproteinkinaseC and
cAMP-dependent pathways in the phosphorylation of theNa,K-ATPase, J. Biol. Chem.
275(2000)34693–34700.
[19]E.Roh,C.-Y.Yun,J.YoungYun,D.Park,N.DooKim,etal.,cAMP-BindingSiteofPKA
asaMolecularTargetofBisabolangeloneagainstMelanocyte-SpecificHyperpigmented
Disorder,J.Invest.Dermatol.133(2013)1072–1079.

71
[20]P.Blume-Jensen,R.Janknecht,T.Hunter,Thekitreceptorpromotescellsurvivalvia
activation of PI 3-kinase and subsequent Akt-mediated phosphorylation of Bad on
Ser136,Curr.Biol.8(1998)779–782.
[21]S.R.Datta,H.Dudek,T.Xu,S.Masters,F.Haian,etal.,AktphosphorylationofBAD
couplessurvivalsignalstothecell-intrinsicdeathmachinery,Cell.91(1997)231–241.
[22] M. Cheung, A. Sharma, S. V. Madhunapantula, G.P. Robertson, Akt3 andmutant
V600EB-Rafcooperatetopromoteearlymelanomadevelopment,CancerRes.68(2008)
3429–3439.
[23]L.C.W.Vredeveld,P.A.Possik,M.A.Smit,K.Meissl,C.Michaloglou,etal.,Abrogation
of BRAFV600E-induced senescence by PI3K pathway activation contributes to
melanomagenesis,GenesDev.26(2012)1055–1069.
[24] Y. Yamaguchi, S. Itami, H. Watabe, K.I. Yasumoto, Z.A. Abdel-Malek, et al.,
Mesenchymal-epithelial interactions in theskin: Increasedexpressionofdickkopf1by
palmoplantarfibroblastsinhibitsmelanocytegrowthanddifferentiation,J.CellBiol.165
(2004)275–285.
[25]Y.Yamaguchi,T.Passeron,H.Watabe,K.Yasumoto,F.Rouzaud,etal.,Theeffectsof
dickkopf 1 on gene expression and Wnt signaling by melanocytes: mechanisms
underlyingitssuppressionofmelanocytefunctionandproliferation,J.Invest.Dermatol.
127(2007)1217–25.

72
[26] G.-E. Costin, J.C. Valencia, W.D. Vieira, M.L. Lamoreux, V.J. Hearing, Tyrosinase
processing and intracellular trafficking is disrupted in mouse primary melanocytes
carrying the underwhite (uw)mutation. Amodel for oculocutaneous albinism (OCA)
type4,J.CellSci.116(2003)3203–3212.
[27]S.Dennis,M.Aikawa,W.Szeto,P.A.D’Amore,J.Papkoff,Asecretedfrizzledrelated
protein,FrzA,selectivelyassociateswithwnt-1proteinandregulateswnt-1signaling,
J.Cell.Sci112(1999)3815-3820.
[28]Y.Kawano,R.Kypta,SecretedantagonistsoftheWntsignallingpathway,J.CellSci.
116(2003)2627-2634.
[29]M.Kim,J.H.Han,J.-H.Kim,T.J.Park,H.Y.Kang,SecretedFrizzled-RelatedProtein2
(sFRP2) Functions as a Melanogenic Stimulator; the Role of sFRP2 in UV-Induced
HyperpigmentaryDisorders,J.Invest.Dermatol.136(2016)236–44.
[30]T.J.Park,M.Kim,H.Kim,S.Y.Park,K.C.Park,etal.,Wntinhibitoryfactor(WIF)-1
promotes melanogenesis in normal melanocytes, Pigment Cell Melanoma Res. 27
(2013)72-81.
[31]T.Hirobe,K.Hasegawa,R.Furuya,R.Fujiwara,K.Sato,Effectsoffibroblast-derived
factors on the proliferation and differentiation of human melanocytes in culture, J.
Dermatol.Sci.71(2013)45–57.
[32]N.Chen,Y.Hu,W.H.Li,M.Eisinger,M.Seiberg,etal.,Theroleofkeratinocytegrowth

73
factorinmelanogenesis:Apossiblemechanismfortheinitiationofsolarlentigines,Exp.
Dermatol.19(2010)865–872.
[33]T.B.Fitzpatrick,Thevalidityandpracticalityofsun-reactiveskintypesIthroughVI,
Arch.Dermatol.124(1988)869–871.
[34] W. Choi, R.Wolber,W. Gerwat, T.Mann, J. Batzer, et al., The fibroblast-derived
paracrinefactorneuregulin-1hasanovelroleinregulatingtheconstitutivecolorand
melanocytefunctioninhumanskin,J.CellSci.123(2010)3102–3111.
[35] R.M. Esper, M.S. Pankonin, J.A. Loeb, Neuregulins: Versatile growth and
differentiation factors innervous systemdevelopmentandhumandisease,BrainRes.
Rev.51(2006)161–175.
[36]G.Imokawa,Y.Yada,N.Morisaki,M.Kimura,Biologicalcharacterizationofhuman
fibroblast-derivedmitogenic factors for humanmelanocytes, Biochem. J. 330 ( Pt 3
(1998)1235–9.
[37] G.-E. Costin, V.J.Hearing,Human skin pigmentation:melanocytesmodulate skin
colorinresponsetostress,FASEBJ.21(2007)976–94.
[38]V.B.Swope,E.E.Medrano,D.Smalara,Z.A.Abdel-Malek,Long-termproliferationof
human melanocytes is supported by the physiologic mitogens alpha-melanotropin,
endothelin-1,andbasicfibroblastgrowthfactor,Exp.CellRes.217(1995)453–459.
[39] M.Yaar,M.S.Eller,P.DiBenedetto,W.R.Reenstra,S.Zhai,etal.,Thetrkfamilyof

74
receptorsmediatesnervegrowthfactorandneurotrophin-3effects inmelanocytes, J.
Clin.Invest.94(1994)1550–1562.
[40] G. a Scott, L. aMcClelland, A.F. Fricke, Semaphorin 7a promotes spreading and
dendricity in human melanocytes through beta1-integrins, J. Invest. Dermatol. 128
(2008)151–161.
[41]R.L.Elliott,G.C.Blobe,Roleoftransforminggrowthfactorbetainhumancancer,J.
Clin.Oncol.23(2005)2078–2093.
[42]M.Martínez-Esparza,C.Jiménez-Cervantes,F.Beermann,P.Aparicio,J.A.Lozano,et
al., Transforming growth factor-β1 inhibits basal melanogenesis in B16/F10mouse
melanoma cells by increasing the rate of degradation of tyrosinase and tyrosinase-
relatedprotein-1,J.Biol.Chem.272(1997)3967–3972.
[43] G.Yang,Y.Li,E.K.Nishimura,H.Xin,A.Zhou,etal., InhibitionofPAX3byTGF-β
ModulatesMelanocyteViability,Mol.Cell.32(2008)554–563.
[44]Y.Yamaguchi,V.J.Hearing,Physiologicalfactorsthatregulateskinpigmentation,35
(2009)193–199.
[45] M.J. Pierrat, V.Marsaud, A.Mauviel, D. Javelaud, Expression ofmicrophthalmia-
associated transcription factor (MITF),which is critical formelanoma progression, is
inhibited by both transcription factor GLI2 and transforming growth factor-β, J. Biol.
Chem.287(2012)17996–18004.

75
[46]M.Brassie,C.Pain,CCN3andCCN5,NewFactorsAssociatedwithSkinPigmentation,
J.Pigment.Disord.3(2016)239.
[47]L.Rittié,B.Perbal,J.J.Castellot,J.S.Orringer,J.J.Voorhees,etal.,Spatial-temporal
modulation of CCN proteins during wound healing in human skin in vivo, in: J. Cell
Commun.Signal.,2011:pp.69–80J.
[48] T.Quan,T.He,Y. Shao,L.Lin,S.Kang,etal.,Elevatedcysteine-rich61mediates
aberrantcollagenhomeostasisinchronologicallyagedandphotoagedhumanskin,Am.J.
Pathol.169(2006)482–90.
[49]T.Quan,S.Shin,Z.Qin,G.J.Fisher,ExpressionofCCNfamilyofgenesinhumanskin
invivoandalterationsbysolar-simulatedultravioletirradiation,J.CellCommun.Signal.
3(2009)19–23.
[50] D. Ruiter, T. Bogenrieder, D. Elder, M. Herlyn, Melanoma-stroma interactions:
Structuralandfunctionalaspects,LancetOncol.3(2002)35–43.
[51]M.A.Huber,N.Kraut,J.E.Park,R.D.Schubert,W.J.Rettig,etal.,Fibroblastactivation
protein: Differential expression and serine protease activity in reactive stromal
fibroblastsofmelanocyticskintumors,J.Invest.Dermatol.120(2003)182–188.
[52]P.Wäster,I.Rosdahl,B.F.Gilmore,O.Seifert,Ultravioletexposureofmelanomacells
induces fibroblast activation protein-α in fibroblasts: Implications for melanoma
invasion,Int.J.Oncol.39(2011)193–202.

76
[53]R.Halaban,TheRegulationofNormalMelanocyteProliferation,PigmentCellRes.
13(2000)4–14.
[54] K. Matsumoto, H. Tajima, T. Nakamura, Hepatocyte growth factor is a potent
stimulator of human melanocyte DNA synthesis and growth, Biochem. Biophys. Res.
Commun.176(1991)45–51.
[55]M.Okazaki,K.Yoshimura,Y.Suzuki,G.Uchida,Y.Kitano,etal.,Themechanismof
epidermalhyperpigmentationincafe-au-laitmaculesofneurofibromatosistype1(von
Recklinghausen’sdisease)maybeassociatedwithdermal fibroblast-derivedstemcell
factorandhepatocytegrowthfactor,BrJDermatol.148(2003)689–697.
[56]G.Imokawa,AutocrineandParacrineRegulationofMelanocytesinHumanSkinand
inPigmentaryDisorders,PigmentCellRes.17(2004)96–110.
[57]G.Cardinali,D.Kovacs,M.DelGiglio,C.Cota,N.Aspite,etal.,Akindredwithfamilial
progressivehyperpigmentation-likedisorder:Implicationoffibroblast-derivedgrowth
factorsinpigmentation,Eur.J.Dermatology.19(2009)469–473.
[58]J.D’Orazio,S.Jarrett,A.Amaro-Ortiz,T.Scott,UVradiationandtheskin,Int.J.Mol.
Sci.14(2013)12222–12248.
[59] M.Picardo,G.Cardinali,TheGeneticDeterminationofSkinPigmentation:KITLG
andtheKITLG/c-KitPathwayasKeyPlayersintheOnsetofHumanFamilialPigmentary
Diseases,J.Invest.Dermatol.131(2011)1182–1185.

77
[60]A.Y.Lee,Recentprogressinmelasmapathogenesis,PigmentCellMelanomaRes.28
(2015)648–660.
[61]H.Y.Kang,J.S.Hwang,J.Y.Lee,J.H.Ahn,J.Y.Kim,etal.,Thedermalstemcellfactor
andc-kitareoverexpressedinmelasma,Br.J.Dermatol.154(2006)1094–1099.
[62] J.-Y. Kim, T.-R. Lee, A.-Y. Lee, Reduced WIF-1 Expression Stimulates Skin
HyperpigmentationinPatientswithMelasma,J.Invest.Dermatol.133(2012)191–200.
[63]H.Aoki,O.Moro,H.Tagami,J.Kishimoto,Geneexpressionprofilinganalysisofsolar
lentigoinrelationtoimmunohistochemicalcharacteristics,Br.J.Dermatol.156(2007)
1214–1223.
[64] H. Takayama, W.J. LaRochelle, R. Sharp, T. Otsuka, P. Kriebel, et al., Diverse
tumorigenesis associated with aberrant development in mice overexpressing
hepatocytegrowthfactor/scatterfactor,Proc.Natl.Acad.Sci.U.S.A.94(1997)701–6.
[65]D.Kovacs,G.Cardinali,N.Aspite,C.Cota,F.Luzi,etal.,Roleoffibroblast-derived
growth factors in regulating hyperpigmentation of solar lentigo, Br. J. Dermatol. 163
(2010)1020–1027.
[66] E.Bastonini,D.Kovacs,M.Picardo,Skinpigmentationandpigmentarydisorders:
Focusonepidermal/dermalcross-talk,Ann.Dermatol.28(2016)279–289.
[67]N.Chen,Y.Hu,W.H.Li,M.Eisinger,M.Seiberg,etal.,Theroleofkeratinocytegrowth
factorinmelanogenesis:Apossiblemechanismfortheinitiationofsolarlentigines,Exp.

78
Dermatol.19(2010)865–872.
[68]C.Regazzetti,F.Joly,C.Marty,M.Rivier,B.Mehul,etal.,TranscriptionalAnalysisof
Vitiligo Skin Reveals the Alteration of WNT Pathway: A Promising Target for
RepigmentingVitiligoPatients,J.Invest.Dermatol.135(2015)3105–3114.
[69]S.H.Oh,J.Y.Kim,M.R.Kim,J.E.Do,J.Y.Shin,etal.,DKK1ishighlyexpressedinthe
dermisofvitiligolesion:IsthereassociationbetweenDKK1andvitiligo?,J.Dermatol.
Sci.66(2012)163–165.
[71]P.E.Grimes,Newinsightsandnewtherapiesinvitiligo,293(2005)730.
[72]A.S.Ricard,C.Pain,A.Daubos,K.Ezzedine,I.Lamrissi-Garcia,etal.,StudyofCCN3
(NOV) and DDR1 in normalmelanocytes and vitiligo skin, Exp. Dermatol. 21 (2012)
411–416.
[70] A. Rojas-Villarraga, J. Amaya-Amaya, A. Rodriguez-Rodriguez, R.D. Mantilla, J.M.
Anaya,Introducingpolyautoimmunity:secondaryautoimmunediseasesnolongerexist,
AutoimmuneDis.2012(2012)254319.
[73] L. Zhou, K. Yang, T. Andl, R. RandallWickett, Y. Zhang, Perspective of targeting
cancer-associatedfibroblastsinmelanoma,J.Cancer.6(2015)717–726.
[74] M. Augsten, Cancer-associated fibroblasts as another polarized cell type of the
tumormicroenvironment,Front.Oncol.4(2014)62.

79
[75]L.Zhou,K.Yang,R.RandallWickett,Y.Zhang,Dermalfibroblastsinducecellcycle
arrestandblockepithelial-mesenchymaltransitiontoinhibittheearlystagemelanoma
development,CancerMed.5(2016)1566–1579.
[76] P.Zigrino,R.Nischt,C.Mauch,Thedisintegrin-likeandcysteine-richdomainsof
ADAM-9mediateinteractionsbetweenmelanomacellsandfibroblasts,J.Biol.Chem.286
(2011)6801–6807.
[77]M.Tiago,E.M.deOliveira,C.A.Brohem,P.C.Pennacchi,R.D.Paes,etal.,Fibroblasts
protectmelanomacellsfromthecytotoxiceffectsofdoxorubicin,TissueEng.PartA.20
(2014)2412–21.
[78] R. Straussman, T.Morikawa, K. Shee,M. Barzily-Rokni, Z.R. Qian, et al., Tumour
micro-environment elicits innate resistance toRAF inhibitors throughHGF secretion,
Nature.487(2012)500–4.
[79]F.Belleudi,G.Cardinali,D.Kovacs,M.Picardo,M.R.Torrisi,KGFPromotesParacrine
ActivationoftheSCF/c-KITAxisfromHumanKeratinocytestoMelanomaCells,Transl.
Oncol.3(2010)80–90.
[80]K.A.Giehl,U.Nägele,M.Volkenandt,C.Berking,Proteinexpressionofmelanocyte
growthfactors(bFGF,SCF)andtheirreceptors(FGFR-1,c-kit)inneviandmelanoma,J.
Cutan.Pathol.34(2007)7–14.
[81] P.F. Innominato,L.Libbrecht, J.J.vandenOord,Expressionofneurotrophinsand
theirreceptorsinpigmentcelllesionsoftheskin,J.Pathol.194(2001)95–100.

80
[82]N.Erez,M.Truitt,P.Olson,D.Hanahan,Cancer-AssociatedFibroblastsAreActivated
in Incipient Neoplasia to Orchestrate Tumor-Promoting Inflammation in an NF-??B-
DependentManner,CancerCell.17(2010)135–147.
[83]J.Chen,H.Li,H.Chen,D.Hu,Q.Xing,etal.,Dickkopf-1inhibitstheinvasiveactivity
ofmelanomacells,Clin.Exp.Dermatol.37(2012)404–410.
[84]H.Park,H.Y.Jung,H.J.Choi,D.Y.Kim,J.Y.Yoo,etal.,DistinctrolesofDKK1andDKK2
intumorangiogenesis,Angiogenesis.17(2014)221–234.
[85] S.Kuphal,S.Lodermeyer,F.Bataille,M.Schuierer,B.Hoang,etal.,Expressionof
Dickkopfgenesisstronglyreducedinmalignantmelanoma,Oncogene.25(2006)5027–
5036.
[86]A.Krtolica,S.Parrinello,S.Lockett,P.Y.Desprez,J.Campisi,Senescentfibroblasts
promoteepithelialcellgrowthandtumorigenesis:alinkbetweencancerandaging,Proc.
Natl.Acad.Sci.U.S.A.98(2001)12072–7.
[87]A.Kaur,M.R.Webster,K.Marchbank,R.Behera,A.Ndoye,etal.,sFRP2intheaged
microenvironment drives melanoma metastasis and therapy resistance, Nature. 532
(2016)250–4.
[88] G. a Scott, L. a McClelland, A.F. Fricke, A. Fender, Plexin C1, a receptor for
semaphorin 7a, inactivates cofilin and is a potential tumor suppressor formelanoma
progression,J.Invest.Dermatol.129(2009)954–963.

81
[89] M. Fukunaga-Kalabis, G. Martinez, S.M. Telson, Z.-J. Liu, K. Balint, et al.,
Downregulation of CCN3 expression as a potential mechanism for melanoma
progression,Oncogene.27(2008)2552–2560.
[90] T.Yamamoto, I.Katayama,K.Nishioka. Impairedexpressionof stemcell factor in
dermatofibromafibroblasts.ActaDermVenereol76(1996)257-9.
AuthorBiography
Yinjuan Wang is a Ph.D. candidate from the Laboratory of Engineering and Cutaneous
Biology, UMR 1098, Bourgogne Franche-Comte University, Besancon, France. Her thesis,
supervisedbyProfessorsPhilippeHumbertandHeLi,isconcernedwithskinpigmentation.
She receivedMedicalDoctor on2011andDermatologicalMaster on2014, fromSichuan
UniversityandKunmingMedicalUniversityofChina, respectively.Her research interests
involvemelaninsynthesisbymelanocytesanditspathways,inparticulartheepidemiology
andphysiopathologyofhyperpigmentedskinlesionsincludingmelasmaandlentigo.
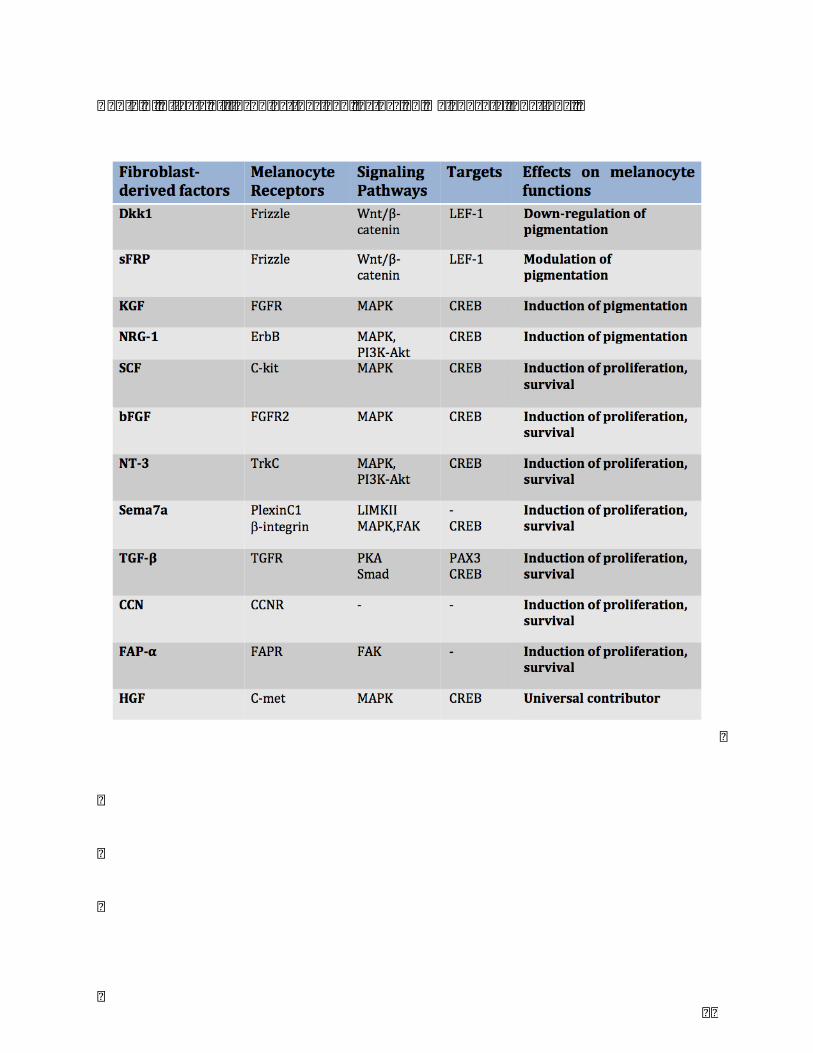
82
Table.1.Effectsoffibroblast-derivedfactorsonmelanocytefunctions.
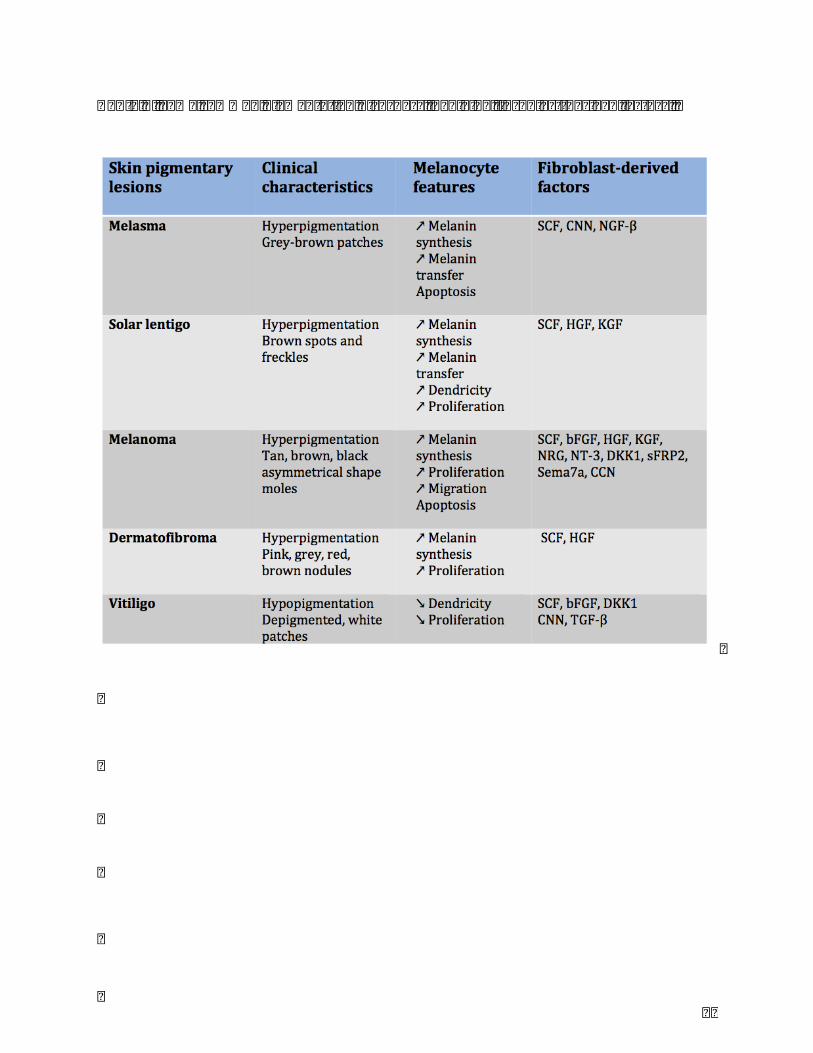
83
Table.2.Somecommonpigmentationdisordersinvolvingfibroblast-derivedfactors.
84
ChapterII
Developmentandvalidationofasimplemethodforthe
extractionofhumanskinmelanocytes

85
Theonlycellsthatproducemelaninaremelanocyteswhichareusefulforstudying
manypigmentarydisordersaswellasmechanismsregulatingmelanogenesis,andforcell
transplantationintreatmentofvitiligo.
Therearevariousmethodstoisolatemelanocytesfromskin,whicharegenerallybasedon
two-steps enzyme digestion (by dispase and trypsin) process (Table 1)76–78. Although
previouslydevelopedprotocolsaresatisfactoryintermsofcellyieldandviability,theyuse
enzymesofanimalorigin.Inthischapter,weintroducedasimpleandoptimizedmethodfor
extractionofmelanocytesfromlowsurfaceofhumanskinsample,whichshouldbeeffective
forfunctionalandmolecularstudiesandalsoformedicalapplicationsuchcellgrafting.We
compare the new protocol with a classic melanocyte extraction method. This work is
summarizedinthefollowingarticlesubmittedinCytotechnologyjournal.
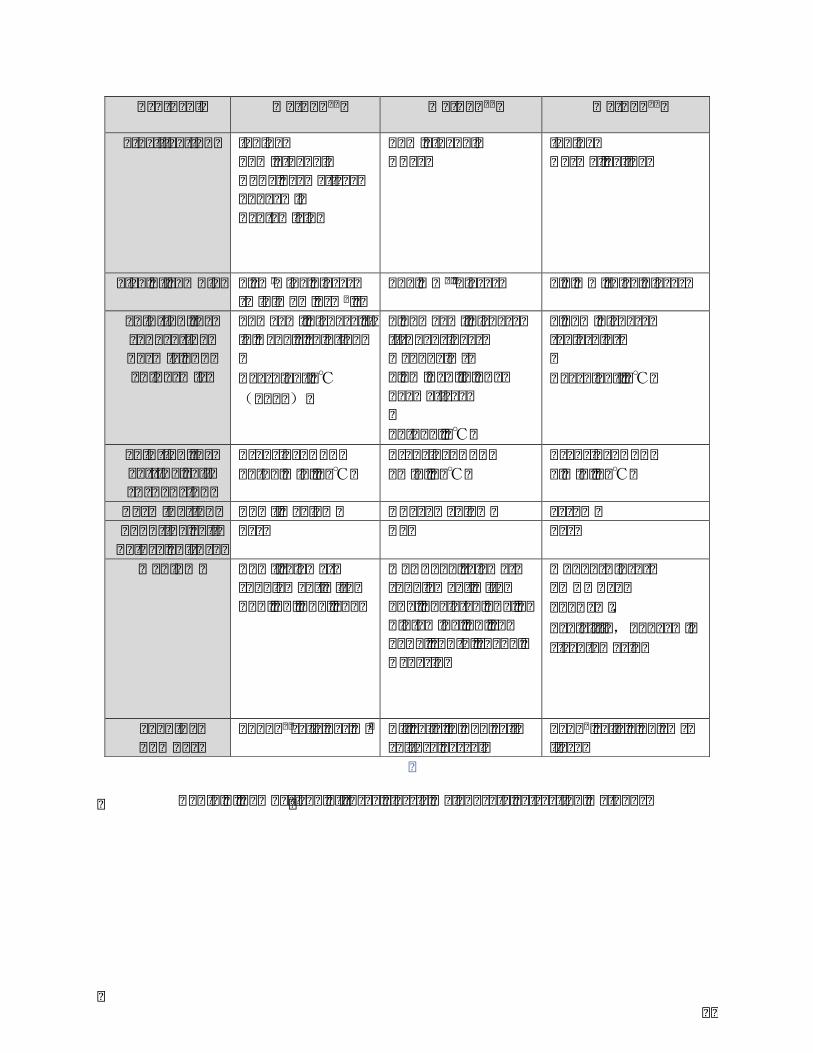
86
Protocol Method77 Method78 Method76
Sterilization Iodine70%ethanolDPBSCa\Mg-free10μg/mlGentamicin
70%ethanolHBSS
IodineNormalSaline
Sizeofsample 1cmwidepieces(minimum1cm2)
5×5mm2pieces 0.5mmthickpieces
Solutionforseparatingdermisandepidermis
10mg/mldispaseIIinHank’ssolutionOvernight4℃(≥18h)
0.48mg/mldispaseII(BoehringerMannheim)0.1%BSAinPBSCa/Mg-free18-24h,4℃
0.25%dispase(Solarbio)Overnight,4℃
Solutionforgettingcellsuspension
Trypsin/EDTA10-15min,37℃
Trypsin/EDTA5min,37℃
Trypsin/EDTA5min,37℃
Termination RPMIMedium HBSS+Medium SerumCountingcellbeforeculture
Yes No Yes
Medium RPMI(Sigma)supplementwithTPA,CT,ET1,SCF
MCDB153(Sigma)supplementwithFBS,ChelatedFBS,L-Glutamine,CT,rh-bFGF,rET-3,rhSCF,Heparin
MC254(Gibco)1%HMGS2100U/ml,Penicillin,50μg/mlstreptomycin
Seedingnumber
15×104cellsperml AllcellsinT25cell-culturevessel
5x105cells/25cm²flask
Table1.Comparisonofthreeclassicmelanocyteextractionmethods

87
Article2:Developmentandvalidationofasimplemethodfortheextractionofhuman
skinmelanocytes
YinjuanWanga,CélineVienneta*,GwenaëlRolina,f,MarionTissota,PatriceMureta,Sophie
Robinb,Jean-YvesBerthonc,LiHed*,PhilippeHumberta,e
aEngineeringandCutaneousBiologyLaboratory,UMR1098,UniversityofBourgogneFranche-
Comté,Besançon,France
bBioexigenceS.A.R.L,Besançon,France
cGREENTECHSA,BiopôleClermontLimagne,SaintBeauzire,France
dDepartmentofDermatology,FirstAffiliatedHospitalofKunmingMedicalUniversity,Kunming,
China
eDepartmentofDermatology,UniversityHospital,Besançon,France
fClinicalInvestigationCenter,InsermCICB1431,UniversityHospital,Besançon,France
*Correspondingauthors:CélineViennet,LiHe
E-mailaddressforcorrespondance:[email protected]@126.com
Acknowledgments: this work was financially supported by Program for Innovative
ResearchTeaminUniversityofMinistryofEducationofChina(GrandNo.IRT13067),the
FundofYunnanProvinceChineseAcademyofSciencesCooperation(GrandNo.2014IB008)
andGreentechSA(France).
Conflictofinterests:theauthorshavenoconflictofinteresttodeclare.

88
Abstract
Primarymelanocytesincultureareusefulmodelsforstudyingepidermalpigmentationand
efficacyofmelanogenic compounds,ordevelopingadvanced therapymedicinalproducts.
Cellextractionisaninevitableandcriticalstepintheestablishmentofcellcultures.Many
enzymatic methods for extracting and growing cells derived from human skin, such as
melanocytes, are described in literature. They are usually based on 2 enzymatic steps,
Trypsin in combination with Dispase, in order to separate dermis from epidermis and
subsequently provide a suspension of epidermal cells. The objective of thisworkwas to
develop and validate an extraction method of human skin melanocytes being simple,
effectiveandapplicable tosmallerskinsamples,andavoidinganimalreagents.TrypLETM
productwastestedonverylimitedsizeofhumanskin,equivalentofmultiple3-mmpunch
biopsies,andwascomparedtoTrypsin/Dispaseenzymes.Functionalityofextractedcells
was evaluated by analysis of proliferation, morphology and melanin production. In
comparisonwith Trypsin/Dispase incubationmethod, themain advantages of TrypLETM
incubationmethodwere theeasierof separationbetweendermisandepidermisand the
lower contamination by keratinocytes after extraction. Both protocols preserved
morphological and biological characteristics of melanocytes. The minimum size of skin
samplethatallowedtheextractionoffunctionalcellswas6x3-mmpunchbiopsies(e.g.,42
mm2)whatever themethod used. In conclusion, this newprocedure based onTrypLETM
incubationwouldbesuitableforestablishmentofoptimalprimarymelanocytesculturesfor
clinicalapplicationsandresearch.

89
Introduction
In human skin,melanocytes are specialized cells of the epidermis and hair follicles that
producethepigmentmelanin.Innormalepidermis,theyconstituteapproximately3to7%
of the cells, contactingwith severalneighboringkeratinocytesand forminganepidermal
melaninunit.Melanocytesarelocalizedinthebasallayeroftheepidermisatthejunctionof
thedermis,andtheirdendritesallowmelanintransfertokeratinocytes.Establishmentand
characterization of melanocyte cultures are important for understanding and treating
pigment-relateddisorders,ordevelopinginvitropigmentedskinmodels(Guerraetal.2003;
Duvaletal.2003;Yoonetal.2003;Vermaetal.2015;Kumaretal.2012).Althoughanumber
of cell lines originated fromhealthy and lesional skin are used for studying cellular and
molecularbiology, testing cosmetic and chemicalproducts, primary culture remains as a
more reliablemodel thatmimics in vivo skin. Severalmethods for the isolation and the
cultureofmelanocytesfromhumanskinhavebeendescribed(Hsuetal.2005;Godwinetal.
2014;Tangetal.2014;Nielsenetal.1984;Gilchrestetal.1984;Yaranietal.2013).Thefirst
human melanocyte culture from skin was published by Hu et al. (1957). Unlike basal
keratinocytes that are proliferative and capable of regenerating the epidermal layers,
melanocyte cultureshave lowproliferationpotential and short life span.Toobtainhigh-
densityprimaryculturesofmelanocytes,differentselectivemediaaresupplementedwith
mitogens as Phorbol 12-myristate 13-acetate (PMA), endothelins, growth factors (basic
fibroblastgrowthfactor,stemcellfactor,hepatocytegrowthfactorandnervegrowthfactor)
(Eisinger andMarko 1982; Halaban et al. 1987; Imokawa et al. 1998; Yada et al. 1991;
Peacocke et al. 1988). 5-fluorouracil (5-FU) is utilized to isolate pure populations of
melanocytes free from keratinocyte (Takuo et al. 1983). Isolation of melanocytes from

90
human skin explants is usually performed in a two-step protocol, dispase incubation to
separate the dermal and epidermal layers, following by trypsin treatment to release the
epidermalcells(Tangetal.2014).Althoughtrypsinisutilizedforavarietyofcellculture
applications,itisderivedfromanimalsandmaycontaintoxicsubstances.Thereforemostof
thetechniquesdescribedintheliteraturearealimitingfactorinusingcellsforclinic.
Thisworkdescribesanewmethodforisolationofmelanocytes,basedonone-stepenzyme
incubationusingtheTrypLETM,ananimalorigin-freeandrecombinantenzyme.Wereported
thenumberofcellsisolatedfromdifferentlimitedsizesofskintissueandcharacterizedthe
cellpopulation.Inaddition,wecomparedournovelprocedurewithanestablishedtechnique
(Godwinetal.2014),studyingthefunctionalityoftheextractedmelanocytes.
MaterialsandMethods
Collectionofskinbiopsies
Skinsampleswereobtainedfromdiscardedskinfollowingabdominoplastywithinformed
donor consent (5 patients, 43±17 years, skin types II-IV). The hypodermis layer was
removed.Subsequently,skinbiopsiesof3mmindiameterwererealizedandputfor10min
in phosphate-buffered saline (PBS) containing 100 IU/ml penicillin and 100 μg/ml
streptomycin (PS) (all from Pan-Biotech, Dutscher, France). In total, 45 biopsies were
collectedperskinand6distinctgroupswererealizedaccordingtothenumberofbiopsies
(5to10).
Dissociationofepidermisfromdermisandseparationofepidermalcells
Method I: simpleenzymaticdigestion (TrypLETM) (Figure.1)Biopsieswere incubated in
TrypLETMenzyme1X(GIBCOÒTrypLETMSelectTMCTSTM,ThermoFisherScientific,France)at
roomtemperatureovernight(17h).Duringthetreatment,theepidermallayerwaseasily

91
separated fromthedermis.After incubation,alldermiswerediscarded, thenmelanocyte
culture medium was added and epidermal cells were disaggregated mechanically by
repeatedpipettingupanddown.Mixtureswerefilteredthrough70μmcellstrainers,and
centrifugedfor5minat1,100rpm.Cellpelletswereresuspendedinmelanocytemedium
which consists ofM254 culturemedium supplementedwith humanmelanocytes growth
supplement (HMGS, Life Technologies, ThermoFisher Scientific, France), 100 IU/ ml
penicillinand100μg/mlstreptomycin.Epidermalcellswerecounted,placedinto25cm2
tissuecultureflasksatacelldensityof2to4x104cells/cm2,andkeptat37°Cinahumidified
atmospherewith5%CO2.
MethodII:double-enzymaticdigestion(dispaseandtrypsin)Acontrolgroupwassetupby
incubating biopsies in 10 mg/ml dispase II (GIBCOÒ, ThermoFisher Scientific, France)
overnight(17h)at4°C.Theepidermiswasseparatedfromthedermisusingfineforceps.
Isolatedepidermal layerswereput in0.05%trypsin/0.02%EDTAsolution (Pan-Biotech,
Dutscher,France)at37̊Cfor10min.
After incubation, melanocyte culture medium was added and epidermal cells were
disaggregatedmechanicallybyrepeatedpipettingupanddown.Thefollowingstepswere
similarasTrypLETMmethod.
Primarymelanocytecultures
Culture medium was firstly changed after 48 h, then every 3 days. When melanocytes
reached 70% confluency, they were harvested by 3 min treatment at 37°C with 0.05%
trypsin/0.02%EDTAsolution.Thecellsuspensionwasneutralizedwithfetalbovineserum
(Pan-Biotech,Dutscher,France)andcentrifugedfor5minat1,100rpm.Thecellpelletwas
resuspended in melanocyte medium. Viable cell counts were performed before seeding

92
melanocytesat16x103cells/cm2 intonewculture flask foranotherroundof incubation.
ImagesofcellcultureswereacquiredusinganOlympusIX50invertedmicroscope.
Melanocyteculturesatpassage2wereusedforthefollowingexperiments.
Melanocyteproliferationassay
Melanocyteswereseededatadensityof6x104cells/wellin12-wellplates.After1,2,3and
4 days of culture, cellswere harvested by trypsinization and counted using trypan blue
exclusion.
Measurementofmelanincontent
Melanocyteswere plated at 400000 cells on 25 cm2 flask and grown for 5 days to 70%
confluence.TheywerewashedwithPBS, collectedbyscrapingandpelleted. Intracellular
melaninwasextractedbysolubilizingfrozencellpelletsin1NNaOHcontaining10%DMSO
for 2 h at 90 °C. The samples were then centrifuged at 1100 rpm for 10 min and the
supernatantswere transferredto fresh tubes.Themelanincontent in thesesupernatants
wasmeasuredspectrophotometricallyatanabsorbanceof420nmagainstastandardcurve
ofknownconcentrationsofsyntheticmelanin(0-50μg/mL,Sigma-Aldrich,France).
TheproteinlevelincellpelletwasdeterminedusingtheBradfordassay.Theintracellular
melanincontentwasadjustedbytheamountofproteininthesamesample.
Immunocytochemistry
Melanocytesweregrownin8-wellchamberslides(NuncLab-TekChamberSlides,Dutscher,
France)atadensityof1×104cells/well.After24h,cellswerefixedin3%paraformaldehyde
in PBS, then rinsed in PBS and permeabilised for 15min in 0.1% Triton X-100 in PBS.
Nonspecificbindingsiteswereblockedatroomtemperaturebya10minincubationin1%
glycineinPBSfollowedbya1hincubationin3%bovineserumalbumin(BSA)solutionin

93
PBScontaining10%normalgoatserumand0.1%TritonX100.Cellswereincubatedat4°C
overnight with polyclonal rabbit anti-human microphthalmia-associated transcription
factor (MITF) at 1:1000 dilution, then 1 h at room temperature with goat anti-rabbit
antibodyconjugatedwithrhodaminediluted1:2000.Theyweredouble-stainedwithFITC-
conjugatedphalloidinfor15minatroomtemperature(allantibodiesfromSigma-Aldrich,
France).PBSwith1%BSAand0.1%TritonX-100wasusedforwashesbetweenincubations
andantibodydilutions.SamplesweremountedonglassslideswithFluoromountTMmedium
andobservedunderaZeissAxioskop40fluorescencemicroscope.
StatisticalAnalysis
Allvalueswereexpressedasmean±SE.Statisticalanalysesbetweenskinbiopsygroups
wereperformedusingANOVAwithSigmaStat.Differenceswereconsideredsignificantifthe
Pvaluewas<0.05.
Resultsanddiscussion
Primary cultures of human melanocytes are appropriate models to study the
pathomechanismsofpigmentation,thegeneticdisordersofmelanocytedevelopmentorthe
efficacy of melanogenic compounds. Unfortunately, the clinical size and the anatomical
locationofmelanocyticlesionsoftenpreventawideexcisionofskinand,thusmakedifficult
the establishment of melanocyte cultures. Many methods for extracting and growing
melanocytesaredescribed(Hsuetal.2005;Godwinetal.2014;Tangetal.2014;Nielsenet
al.1984;Gilchrestetal.1984;Yaranietal.2013).Theyareusuallybasedon2enzymatic
steps, dispase in combination with trypsin, to separate dermis from epidermis and
subsequently provide a suspension of epidermal cells. The objective of thisworkwas to
developandvalidateamelanocyteextractionmethodbeingthesimplest,themosteffective

94
andapplicabletosmallskinsamples.
Enzymaticdissociationofhumanskintissue
Melanocyteswereisolatedfromverylimitedsizeofhumanskin,equivalentofmultiple3-
mmpunchbiopsies(notexceeding70mm2intotalsurfacearea).Weusedone-stepenzyme
incubation with TrypLETM, an animal origin free and recombinant enzyme, for isolating
epidermis from dermis and release epidermal cells. We compared our method with a
standarddissociationprotocolwhichisperformedintwo-stepenzymeincubation.Asshown
inFigure.2,theincubationinTrypLETMresultedinatotalseparationoftheepidermisfrom
thedermis.Theincubationwithdispasewaslesseffectivewithapartialseparationofthe2
skinlayersontheoutsideedgeofbiopsies.Epidermalsheetswerepeeledfromthedermis
with forceps. Following TrypLETM treatment, the intact epidermal layer was easily
dissociatedmechanicallyintoacellsuspension.Withthestandardprotocol,thedigestionof
epidermisrequiredasecondenzymeincubationusingtrypsin,supportedbyamechanical
action.
Inaddition,contrarytothetraditionalmethod,theadvantageofourmethodistheabsence
of animal derived enzyme, a major condition for the production of advanced therapy
medicinal products. Kormos et al. (2016) described the use of the Mel-mix medium
supplementedwithhuman serum forobtainingpuremelanocyte cultures.Therefore, the
combinationofdifferent techniquesoffersvaluableopportunity inculturedmelanocytes-
basedtransplantationtherapies.
Growthpatternofextractedcells
As Normand and Karasek (1995) who focused on keratinocytes, endothelial cells and
fibroblasts,ourmethodestablishedmelanocyteculturesfromsmallamountoftissue.There

95
wasnosignificantdifferenceinnumberofcellextractedamonggroupsof6to10biopsies,
andwhatever themethodused (Figure. 3). Surfaceof skin tissue less thanor equal to5
biopsies(e.g.,35mm2)didn’tallowsurvivalandproliferationofextractedcells (datanot
shown).Extractedcellsattachedtothecultureflaskwithin48handmelanocytesinitiateda
dendritic appearance (Figure. 4). After 10 days of incubation, it was observed that
melanocytes were 40-50% confluent and displayed typical dendritic morphology. Cells
became highly proliferative. In case of TrypLETMmethod, we obtained a relatively pure
primarymelanocyteculture.Incontrast,primaryculturesobtainedfromdispaseandtrypsin
incubations were contaminated with keratinocytes, suggesting a significant increase of
adhesionandgrowthofextractedkeratinocytescomparedtoTrypLETM.Incubationfor17h
at4°CtoseparateepidermisfromdermiswithTrypLETMmethodgavebetterresultswith
less keratinocytes. However in subsequent passages these clusters of keratinocytes
disappeared, inducing the decrease of cell number between extraction day and end of
passage 1. Extracted cells reached 70% confluency around 14 days and they were
subculturedata1:2splitratioapproximatelyevery10to14days.Thereisnodifferencein
thecellyieldofearlypassagemelanocytesusingbothmethodsandfromskinsampleswith
morethan42andlessthan70mm2oftissue.
FunctionalityofmelanocytesPrimarymelanocytesinculturemighttobeoptimaltoexamine
invivopathomechanisms, toassessbiologicaleffectofcompoundsortotreatpigmentary
lesions. Previous work showed that non-enzymatic tissue incubation method for cell
isolationpreservesmolecularmarkers(Xinyanetal.2014).Prolongedpassagesofcultured
melanocytes tend to lose their pigmentation (Virador et al. 1999). Due to the limited
proliferativecapacityofprimarymelanocytesandthevariabilityofphenotypesgenerated,

96
it’s necessary to limitmelanocyte passage number. In the presentwork,we studied the
functionality of melanocytes cultured at passage 2. Several melanocytic markers can be
utilized to identify typically melanocytes, including MITF. MITF is a cell-type specific
regulator that is required for thedevelopmentandsurvivalofpigmentscells. Itactsasa
masterregulatorofmelaninproduction(Shietal.2016).Weanalyzedcellmorphologyand
moreprecisely thedynamic cytoskeletal networkby filamentous actin (F actin) labeling.
Resultsobtainedwereequivalentwhateverthenumberof3-mmpunchbiopsies.Onlythe
data for10x3-mmpunchbiopsiesareshown.Similargrowthratewereobservedamong
TrypLETM andDispase/Trypsinmethods (Figure. 5). Therewas also no difference in the
intracellular melanin content of melanocytes (Figure. 6). Immunocytochemical staining
confirmed that cells extracted by both methods positively expressed MITF and F actin
(Figure.7).Itwas concluded that TrypLETM andDispase/Trypsinmethods are of similar
efficacy for preserving cell biological characteristics. Melanocytes extracted with our
protocolandculturedwithclassicalconditionsexhibitednormalfunctionalitywithrespect
todendriticmorphology,pigmentproductionandgrowthkinetic.
Conclusion
Wereportinthisstudyanovelandsimpleextractionmethodofhumanskinmelanocytes
basedontheuseofTrypLETM.Thisprotocolprovidedpurepopulationsofmelanocyteafter
the extraction process and preserved cell morphology and functional characteristics. It
maximizedthenumberofmelanocytesobtainedfromsmallskinsurface(greaterthan42
mm2or6x3-mmpunchbiopsies),andthereforegeneratedsufficientcellstostartaprimary
culture.Inaddition,ourprocedurehadthesignificantadvantagetouseananimal-freecell
culturereagentandcouldbeapplicabletothedevelopmentofadvancedcelltherapyproduct

97
indicatedforthetreatmentofpigmentarydisorders.
References
- DuvalC,SchmidtR,RegnierM,FacyV,AsselineauD,BernerdF(2003)Theuseof
reconstructed human skin to evaluate UV-induced modifications and sunscreen
efficacy.ExpDermatol12:64
- EisingerM,MarkoO(1982)Selectiveproliferationofnormalhumanmelanocytesin
vitro in the presence of phorbol ester and cholera toxin. Proc Natl Acad Sci USA
79:2018–2022
- Gilchrest BA, Vrabel MA, Flynn E, Szabo G (1984) Selective cultivation of human
melanocytesfromnewbornandadultepidermis.JInvestDermatol83:370-376
- GodwinLS,CastleJT,KohliJS,GoffPS,CairneyCJ,KeithWN,SviderskayaEV,Bennett
DC(2014)Isolation,culture,andtransfectionofmelanocytes.CurrProtocCellBiol
3;63:1.8.1-20
- Guerra L, Primavera G, Raskovic D et al (2003) Erbium:YAG laser and cultured
epidermisinthesurgicaltherapyofstablevitiligo.ArchDermatol139:1303-1310
- HalabanR,GhoshS,BairdA(1987)bFGF is theputativenaturalgrowthfactor for
humanmelanocytes.InVitroCellDevBiol2347–52
- HsuMY,LiL,HerlynM(2005)Cultivationofnormalhumanepidermalmelanocytes
intheabsenceofphorbolesters.MethodsMolMed107:13-28
- HuF,StariccoR.J,PinkusH,FosnaughR(1957)Humanmelanocytesintissueculture.
JInvestDermatol28:15-32

98
- Imokawa G, Yada Y, Morisaki N, Kimura M (1998) Biological characterization of
humanfibroblast-derivedmitogenicfactorsforhumanmelanocytes.BiochemJ330:
1235–1239
- KormosB,BelsoN,BebesAetal(2016)Invitrodedifferentiationofmelanocytesfrom
adultepidermis.PloSOne6:e17197
- Kumar R, Parsad D, Kanwar A, Kaul D (2012) Development of melanocyte-
keratinocyte co-culture model for controls and vitiligo to assess regulators of
pigmentationandmelanocytes.IndianJDermatolVenereolLeprol78:599–604
- NielsenHI,DonP(1984)Cultureofnormaladulthumanmelanocytes.BrJDermatol
110:569–580
- NormandJ,KarasekMA(1995)Amethodfortheisolationandserialpropagationof
keratinocytes,endothelialcellsandfibroblastsfromasinglepunchbiopsyofhuman
skin.InvitroCellDevBiolAnimal31:447-455
- PeacockeM,YaarM,MansurCP,ChaoMV,GilchrestBA(1988) Inductionofnerve
growth factor receptors on cultured humanmelanocytes. Proc Natl Acad Sci USA
85:5282–5286
- ShiZ,JiK,YangSetal(2016)JunzhenZhangJ,JianboYao,ChangshengDong,Ruiwen
Fan.Biologicalcharacteristicsofmouseskinmelanocytes.Tissueandcell48:114-120
- TangJ,LiQ,ChengB,JingL(2014)Primarycultureofhumanfaceskinmelanocytes
forthestudyofhyperpigmentation.Cytotechnology66:891-8
- Tang X, Richardson WJ, Fitch RD et al (2014) A new non-enzymatic method for
isolating human intervertebral disc cells preserves the phenotype of nucleus
pulposuscells.Cytotechnology66:979-986

99
- Tsuji T, Karasek M (1983) A procedure for the isolation of primary cultures of
melanocytesfromnewbornandadulthumanskin.JInvestDermatol81:179-80
- VermaG,VarkhandeSR,KarHK,RaniR(2015)Evaluationofrepigmentationwith
culturedmelanocytetransplantation(CMT)comparedwithnon-culturedepidermal
celltransplantationinvitiligoat12thweekrevealsbetterrepigmentationwithCMT.
JInvestDermatol135:2533-5
- ViradorVM,KobayashiN,MatsunagaJ,HearingVJ(1999)Astandardizedprotocolfor
assessingregulatorsofpigmentation.AnalyticalBiochemistry270:207-219
- YadaY,HiguchiK,ImokawaG(1991)Effectsofendothelinsonsignaltransduction
andproliferationinhumanmelanocytes.BiolChem266:18352–18357
- YaraniR,MansouriK,Mohammadi-MotlaghHR,BakhtiariM,MostafaieA(2013)New
procedureforepidermalcellisolationusingkiwifruitactinidin,andimprovedculture
ofmelanocytesinthepresenceofleukaemiainhibitoryfactorandforskolin.CellProlif
46:348-
- YoonTJ,LeiTC,YamaguchiY,BatzerJ,WolberR,HearingVJ(2003)Reconstituted3-
dimensionalhumanskinofvariousethnicoriginsasaninvitromodelforstudiesof
pigmentation.AnalBiochem318:260-269

100
Figure.1Shematicdiagramfortheisolationofmelanocytesfromsmallamountofhuman
skin,viatheTrypLETMmethod.Theinitialseedingdensityforinitiatingtheprimaryculture
is2to4×104cellsper25cm2tissuecultureflask.Generally,onesuchflaskissetupfroman
initialyieldof4to10×105cellsisolatedfromabout42to70mm2pieceofskin.
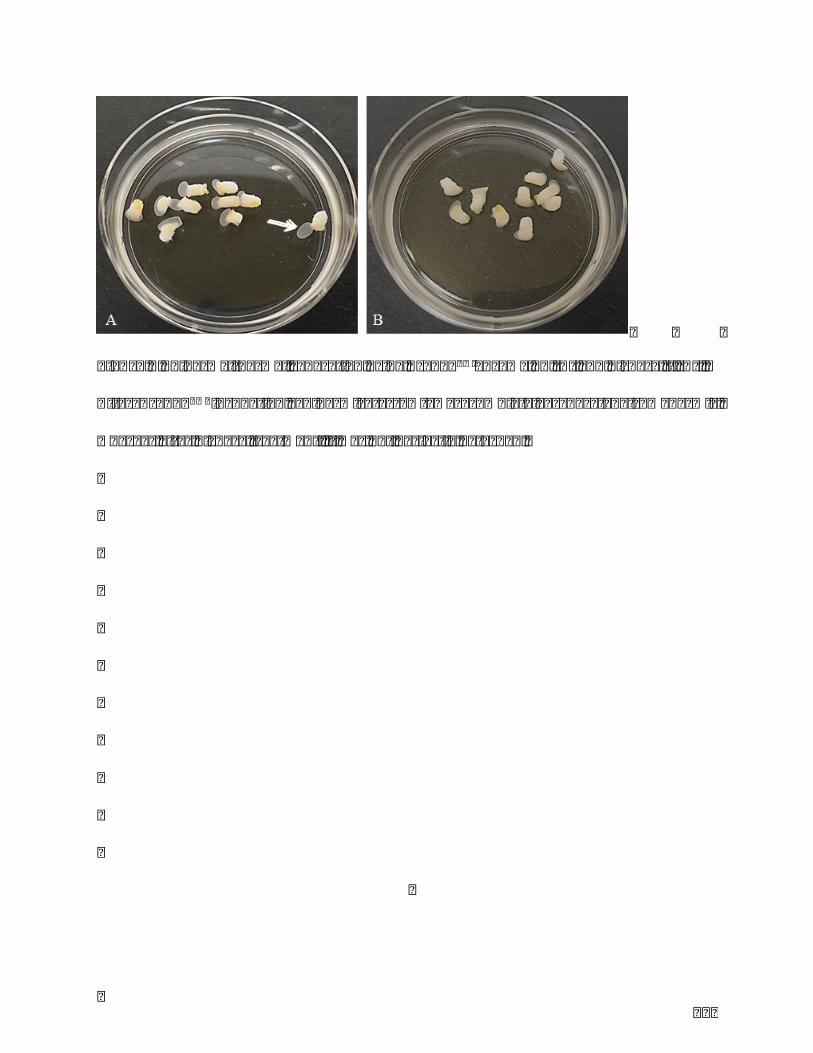
101
Figure.2Epidermal-dermalseparationusingTrypLETMenzyme1X(A)anddispaseII(B).
After TrypLETM incubation, epidermis (arrow) was completely separated from dermis,
whereasafterdispasetreatment,itwasnotperfectlydetached.
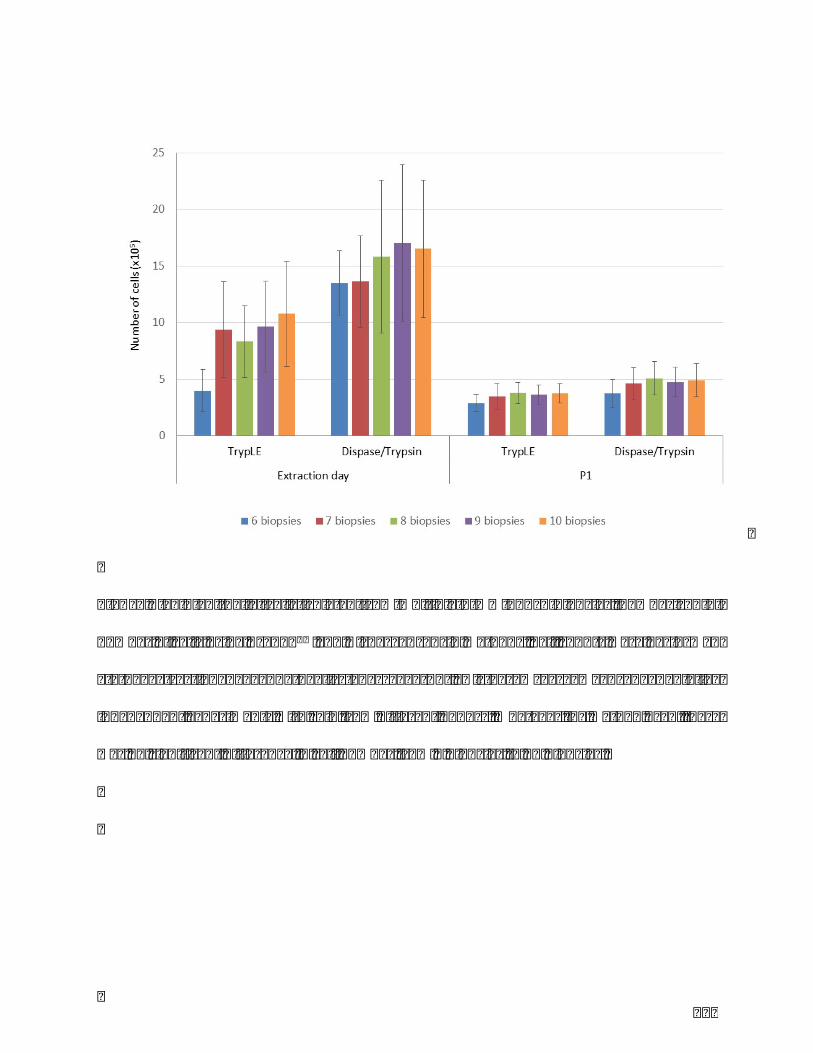
102
Figure.3Skinepithelialcells isolatedfrommultiple3-mmpunchbiopsies.Comparisonof
numberofcellsusingTrypLETMandDispase/Trypsinmethods.Cellcountswereperformed
at the extraction day and the first passage (P1). Histograms are means ± SE of five
independentexperimentswithskinfromdifferentdonors.Whateverthemethodused,there
wasnosignificantdifferenceincellnumberfrom6biopsiesto10biopsies.

103
Figure.4Growthpatternofcellsextractedfrom10punchbiopsiesofskin,byTrypLETM(A,
C) and Dispase/Trypsin (B, C), at 2 days (A, B) and 10 days (C, D) after cell isolation.
Extracted cells attached to the culture flaskwithin 2 days andmelanocytes initiated a
dendriticappearance.After10daysofincubation,melanocytesdisplayedtypicaldendritic
morphology.WithTrypLETMmethod,relativelypuremelanocytecultureswereobtained,
whereas with Dispase/Trypsin method, melanocyte cultures were contaminated with
keratinocytes.Bar100µm
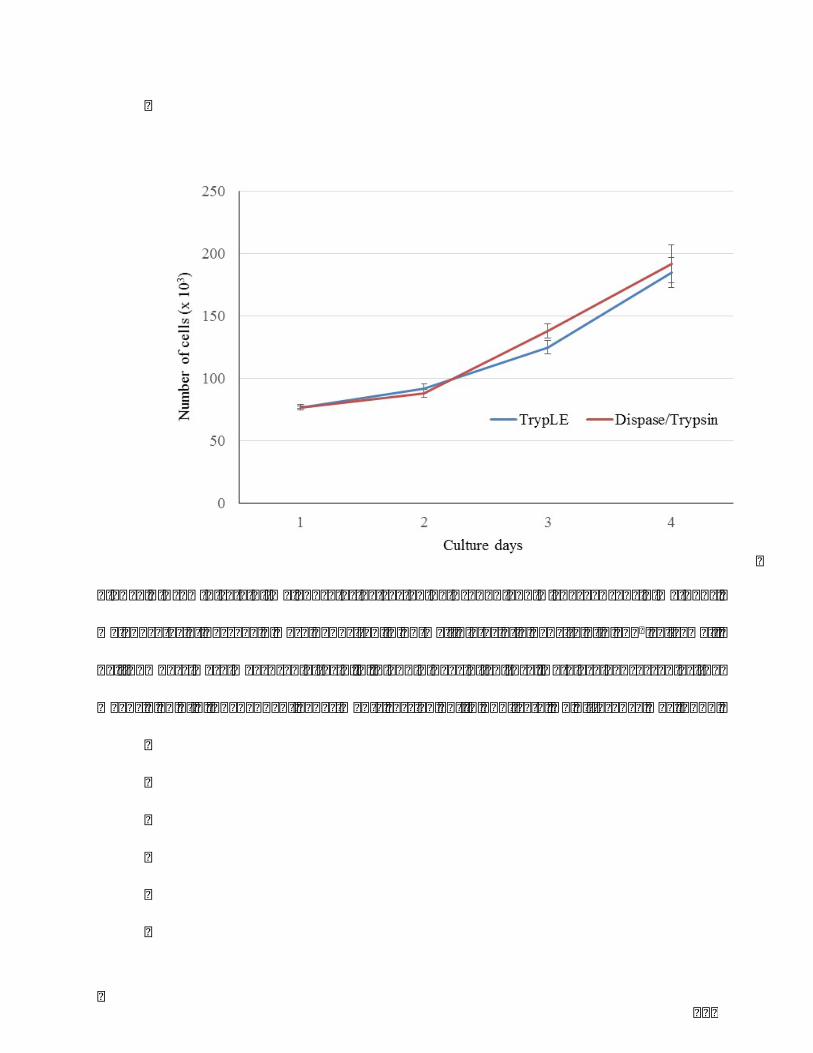
104
Figure.5GrowthrateofmelanocytesextractedbyTrypLEandDispase/Trypsinmethods.
Melanocytesatpassage2weredepositedin12-wellplatesatadensityof6x104cells/well.
Cellnumbersweremeasuredafter1,2,3and4daysofculture.Dataareexpressedasthe
means±SEof5independentexperimentscarriedoutinduplicate.Nodifferencewasfound.
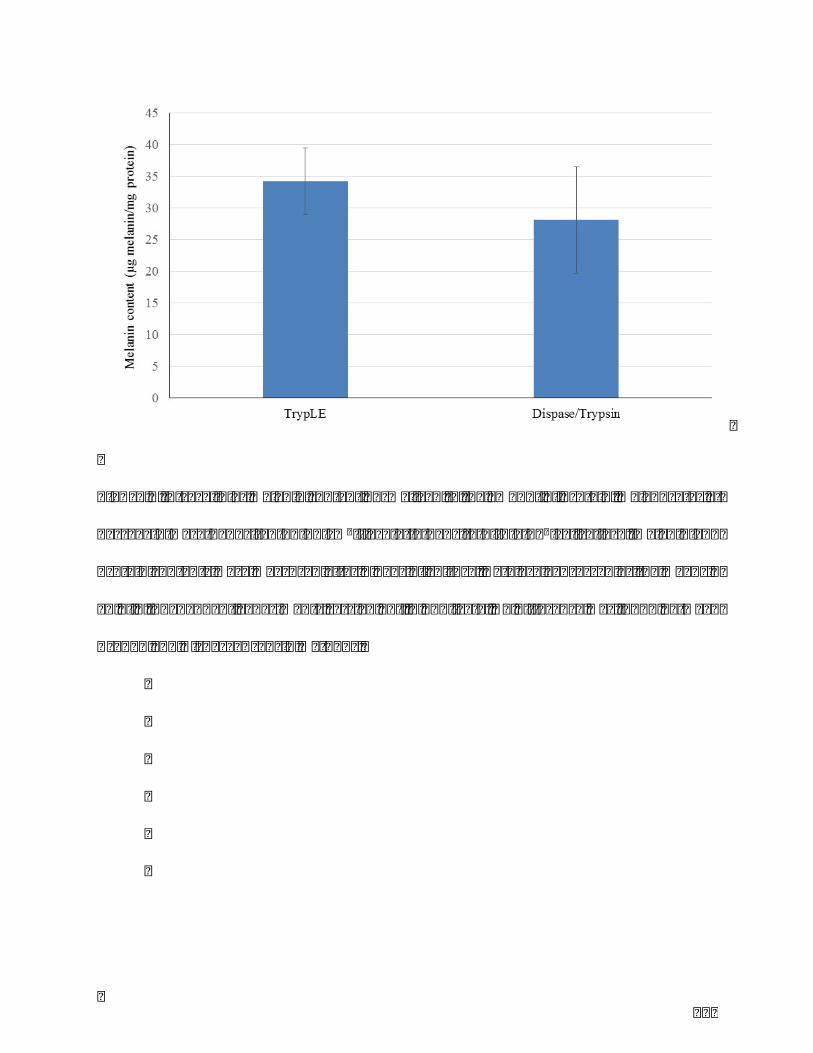
105
Figure.6Intracellularmelanincontentnormalizedtotheamountofprotein.Melanocytesat
passage2weredepositedon25cm2flaskatadensityof4x105cells/flask.Melaninand
proteincontentsweremeasuredafter5daysofculture.Dataareexpressedasthemeans±
SEof5independentexperimentscarriedoutinduplicate.Nodifferencewasfoundbetween
TrypLEandDispase/Trypsinmethods.
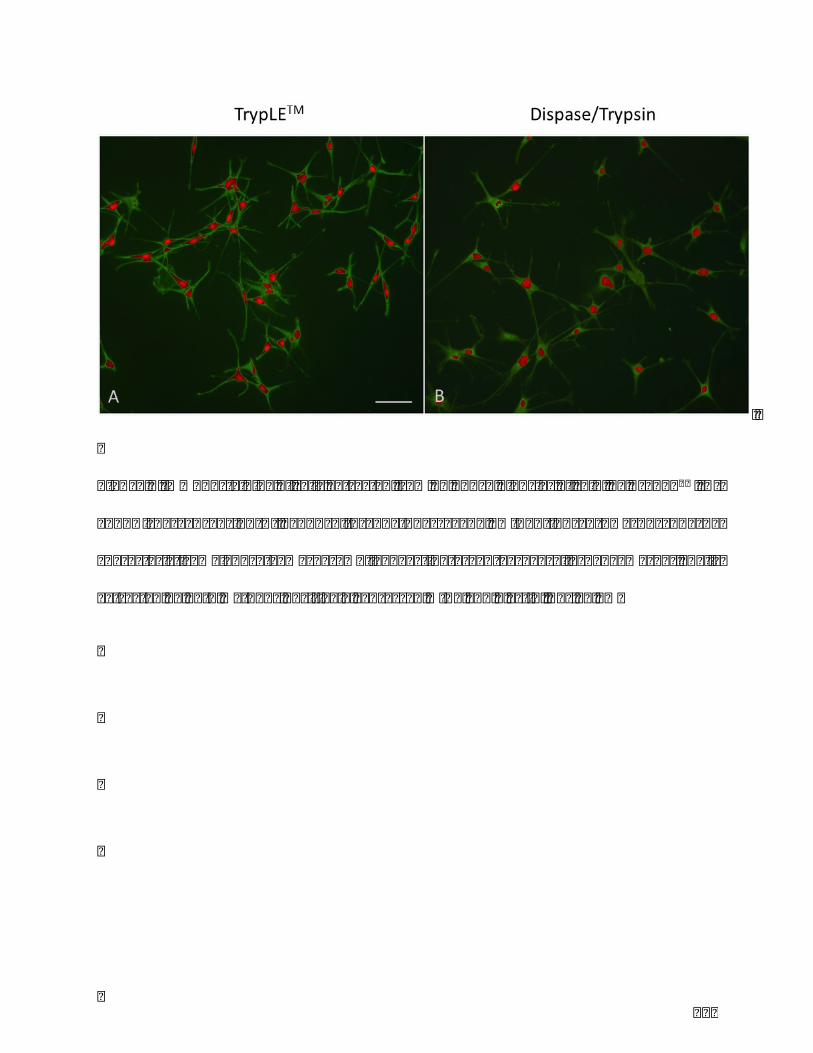
106
Figure.7Immunostainingofcellsextractedfrom10punchbiopsiesofskin,byTrypLETM(A)
and Dispase/Trypsin (B), and cultured at passage 2. MITF (in red) was used as a
characteristic melanocyte marker while F actin (green) as a cell shape marker. Cells
extractedbybothmethodspositivelyexpressedMITFandFactin.Bar50µm

107
ChapterIII
InvolvementofDickkopf-relatedprotein1in
melanogenesis:focusonsolarlentigolesion

108
I. AimsoftheStudy
Recent studies reported that high DKK1 expression by dermal fibroblasts is closely
relatedtohypopigmentedskin,aspalmoplantarskinandvitiligolesion.DKK1encodedby
DKK1geneisanantagonisticinhibitoroftheWntsignalingpathway,byisolatingtheLRP5/6
co-receptorandpreventingtheFrizzeled-Wnt-LRP5/6complex formation.Wnt/bcatenin
signalingregulatesthetranscriptionofmelanocyte-specificgeneslikeMITF,ageneinvolved
inmelaninsynthesis.
The aims of the study were firstly, to determine the cellular level of DKK1 on
hyperpigmented skin including solar lentigo; secondly to conduct a comprehensive
investigationofthemechanismsunderlyingthislesion72.
Invitromodelsthatmimictheinvivoenvironmentwereused:fibroblastsisolatedfromsolar
lentigoandperi-lesionalbiopsies,andnormalfibroblastsembeddedin3Dcollagengeland
exposedtorepeateddosesofUVA.
II. Hypothesis
TGF-β1mediatesdevelopmentofsolarlentigobyreducingDKK1expressioninfibroblasts
through thep38-MAPkinasepathway,which leads toanactivationof theWnt/b-catenin
signalingcascade.Thisprocessmayresultinanunevendistributionofactivemelanocytes,
withareasofhyperpigmentationintheskin(Figure13).
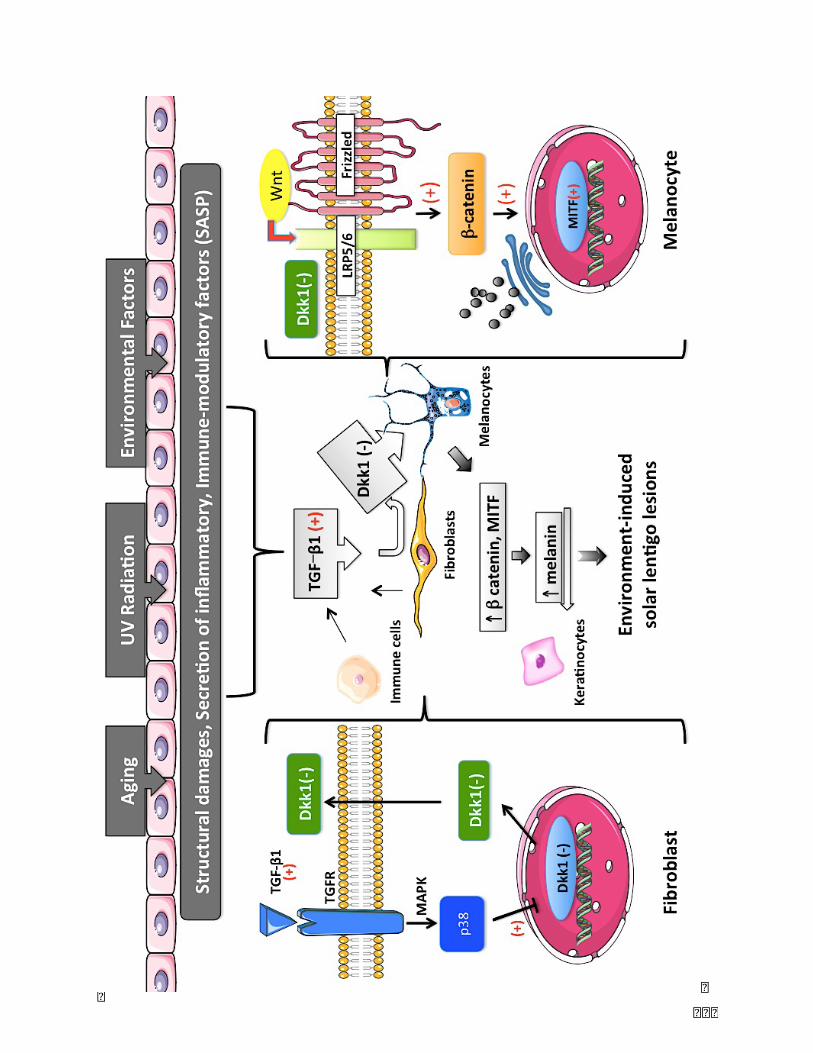
109

110
Figure 13. Hypothesis of the study: Structural damages, pro-inflammatory cytokines,
immune-modulatory factors (SASP) caused by external stimulated factors (UV rays, air
pollution, environmental toxins) induce high secretion of TGF-b by fibroblasts and
inflammatorycells(Tcells,macrophages).ThisprocessinducesadownregulationofDKK1
gene expression and a decrease of DKK1 secretion by fibroblasts. Low expression and
secretionofDKK1leadstotheactivationofWnt/b-cateninsignalingpathway,andresultsin
increasedmelaninsynthesisbymelanocytes.

111
III. MethodsandMaterials
III.I.PartI-Solarlentigobiopsies
III.I.I.Collectionofskinbiopsies
Twobiopsiesof3-mmpunches(onefromsolarlentigolesionalskin(SL),onefromadjacent
normal skin (SN)) were obtained from nine female volunteers on the back of hands
(Caucasianfemales,67–89yearsofage)afterwritteninformedconsent(Universityhospital
ofBesançon,DepartmentofDermatology;Pr.PHumbert).
III.I.II.Histology
III.I.II.IFixationandsectioning
SLandSNbiopsieswerefixedinPFAsolution(3%inPBS),dehydratedthroughagraded
seriesofalcoholandembeddedinparaffin.Sections7µminthicknesswereobtainedusing
arotarymicrotome(Leitz1512;Leica).Theywereclearedinxylenerehydratedinsolutions
ofalcoholandbrieflywashedindistilledwater.
III.I.II.IIMasson'strichromestainingforcollagenfibers
ThesectionswerestainedinHarrishematoxylinsolution,rinsedindistilledwater,stained
with0.1%eosinsolution,treatedwith1%aceticacid-water,washedagainanddifferentiated
in1%phosphomolybdicacid.Finally,thesectionswerestainedwith0.5%lightgreenand
rinsedindistilledwaterbefore1%aceticacid-water.Thestainedsectionsweremounted
withFaramountaqueousmountingmedium,andobservedusingaZeissAxioskop40light
microscope.

112
III.I.II.IIIMasson'sFontanastainingformelanin
The sections were stained with Fontana-Masson kit following the manufacturer’s
instructions(Diapath,Italy).ThestainedsectionsweremountedwithFaramountaqueous
mountingmedium,andobservedusingaZeissAxioskop40lightmicroscope.
III.I.II.IV.Immunostainingforβ-catenin
Slideswereheatedat95 °C for20min in10 mM citratebuffer pH 6. Sampleswere then
incubatedovernightat4°Cwiththeprimaryantibodiesat1:50(MAB13291-SP,R&Dsystem,
USA).Subsequently,theywereincubatedfor1hwithanti-mouseHRP-DAB(CTS002,R&D
system,USA)followingmanufacturer’sprocedure.Thestainedsectionsweremountedwith
Faramount aqueous mounting medium, and observed using a Zeiss Axioskop 40 light
microscope.
III.I.III.Cellculture
III.I.III.I.Extractionoffibroblasts
All cell culture products were purchased from PAN Biotech, Germany. Primary human
fibroblasts from SL (FL) and from SN (FN) were obtained from the biopsies by explant
culture,andcellculturesweremaintainedinDulbecco’sModifiedEagle’sMedium(DMEM)
supplementedwith10%fetalbovineserum(FBS)and1%Penicillin/Streptomycin(P/S,100
units/mLpenicillin,100µg/mLstreptomycin)inahumidified5%CO2atmosphereat37℃.
III.I.III.II.Establishmentofskinfibroblastcultures
FLandFNweremaintaineduntilthemonolayerculturesreached80%confluencyandwere
passagedusing0.25%trypsin-EDTAsolution.Cellswereusedforexperimentsatpassages
2-7.FLandFSwereseededat400,000cellsin25cm2cultureflasks(F25)with3mLfull
DMEMmedium.

113
III.I.IV.Enzyme-linkedimmunosorbentassay(ELISA)
After3daysofculture,supernatantswerecollectedwith10%(v/v)anti-proteasesolution
(P1860,Sigma-Aldrich,France)andstoredat-80°C.ELISAanalyzesforDKK1(kitEHDKK1,
ThermoScientific,USA)andTGF-b1(kitBMS2065,Invitrogen,Australia)wereperformed
accordingtothemanufacturer’sinstructions.Duplicateanalyseswereperformedforeach
sample. Dkk1 concentration in the supernatantswas normalized to the concentration of
intracellular protein (which corresponds directly to the cell number) determined by the
Biuretmethod.TGF-b1concentrationinthesupernatantswasnormalizedtothecellnumber.
III.I.V.Biuretproteinassay
After3daysofculture,cellswerescrapped,lysedwithNaOH0.1NandmixedwithBiuret
reagent(theBiuretreaction(Cu2++e-→Cu+)usesbicinchoninicacidanditisbasedonthe
measurementoftheformationofpurple-coloredcomplexofbicinchoninicacidandCu+)73.
Mixtureswerecentrifugated,andabsorbanceofsupernatantswasdeterminedat620nm.
Bovineserumalbuminwasusedasstandard.Duplicateanalyseswereperformedforeach
sample.
III.I.VI.Real-timequantitativereversetranscription-PCR
After3daysofculture,cellpelletswerecollectedwith0.25%trypsin-EDTAsolution,lysed
inRLTbuffer(Qiagen,France)with1%b-mercaptoethanolandstoredat-80°C.TotalRNA
wasextractedusingtheRNeasyMiniKit(Qiagen,France)accordingtothemanufacturer’s
protocol. Next, cDNA was performed using the High Capacity RNA-to-cDNA kit
(Thermofisher, Applied Biosystems, France). TaqmanÒ Gene Expression Assay kits
(Thermofisher,AppliedBiosystems,France)wereusedtoassessDKK1(Hs00183740_m1)
andAbl(Abelson,Dm01843160_g1)geneexpressions.Ablwasusedashousekeepinggene.

114
ThePCRreactionmixturecomprised2μLofcDNAsample,10μLofTaqManuniversalPCR
mastermix,1μLofeachofTaqMangeneexpressionassayandRNase-freewater.Reactions
wereperformedandmonitoredusing theBioradCFX96Real-TimePCRSystem.Thermal
cyclingconditionswereinitiatedwith2minat50°C,followedby10minat95°C,andthen
40cyclesof95°Cfor10s(denaturation)and60°Cfor1min(annealing/extension).
Forquantification, geneexpression levelswerecalculatedby the comparativeCtmethod
(alsoknownasthe2-DDCt).Duplicatereactionspersamplewererun.
Table2.PrimersandTaqmanprobesusedforqRT-PCR
III.I.VII.ConditionedmediumfromFLandFNinculturedhumanmelanocytes
Primary melanocytes cultures were prepared from adult human abdominal skin.
Melanocytes(passage2-3)wereseededin24-wellplatesatadensityof40,000cells/mLand
culturedfor48hinmelanocyte medium which consists of M254 culture medium supplemented
with human melanocytes growth supplement (HMGS, Life Technologies, ThermoFisher
Scientific, France), 100 IU/mL penicillin and 100 µg/mL streptomycin. The fibroblast-

115
conditionedmediumwas obtainedby culturing FL andFS for4days at density 150,000
cells/well in full DMEM medium. It was then concentrated by lyophilization. Each
lyophilisatewas added to fullmediumM254 and themixturewas added onmelanocyte
cultures(Table3).
Table3.Preparationofdifferentconcentrationsoffibroblast-conditionedmedium
III.I.VIII.MTTassay
Melanocyteswereculturedin96-wellplatesataninitialdensityof5×103cellsperwellfor
24h,andthentreatedwithseveralconcentrationsoffibroblast-conditionedmedium.After
48 h, MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, 0,5 mg/mL,
Sigma-Aldrich,France)wasaddedtoeachwellandthecellswereincubatedat37°Cfor4h.
The medium was removed and dimethyl sulfoxide (DMSO) was added to dissolve the
Concentrationof
fibroblast-conditionedmedium
VolumeofmelanocytefullmediumM254
Volumeoflyophilisate(eachlyophilisateismixedin1.2mLoffullmediumM254)
0% 120µL 0µL
10% 108µL 12µL
15% 82µL 18µL
20% 96µL 24µL
25% 90µL 30µL
50% 60µL 60µL
75% 30µL 90µL
100% 0µL 120µL

116
formazancrystals.Theabsorbancewasmeasuredat571nmwithaspectrophotometer.The
experimentswereperformedinquadruplicate.
III.I.IX.ImmunostainingforMITF
MelanocyteswereculturedinLab-Tek®chamberataninitialdensityof10,000cellsperwell
for24h,andthentreatedwithfibroblast-conditionedmedium(15%v/v).After48h,cells
were fixed for15minwith3%PFA inPBS,washed inPBS andpermeabilizedwith cold
acetonefor10minat-20ºC.Afterwashing,nonspecificbindingsiteswereblockedby10
minincubationin1%glycineinPBSfollowedby1hincubationinPBScontaining3%BSA,
10%serumand0.1%TritonX100.Then,sampleswereincubatedovernightwithantiMITF
primaryantibody(rabbitAcManti-MITF,HPA003259,Sigma-Aldrich)diluted1:200inPBS
containing1%BSAand0.1%TritonX100.Afterwashing inPBSwith1%BSAand0.1%
Triton X100, the secondary antibody (goat anti-rabbit IgG-rhodamine or TRITC, 18772,
Sigma-Aldrich)diluted1:200inPBScontaining1%BSAand0.1%TritonX100,wasapplied
for1h.AfterwashinginPBS,sampleswereincubated15mininrhodamine-conjugatedFITC
(Fluorescein isothiocyanate, P5282, Sigma-Aldrich) for F-actin staining (2.5 µg/mL).
Samples were thenmounted in Dako fluorescent medium, observed under fluorescence
microscopy(Axioskop40,Zeiss).
III.I.X.Melaninassay
Melanocytes were cultured at 400,000 cells on 25 cm2 flask for 24 h, and treated with
fibroblast-conditionedmedium(15%v/v).After48h,theywerewashedwithPBS,collected
by scraping and pelleted. Intracellularmelanin was extracted by solubilizing frozen cell
pellets in 1 N NaOH containing 10% DMSO for 2 h at 90 °C. The samples were then
centrifugedat1100rpmfor10minandthesupernatantsweretransferredtofreshtubes.

117
Theintracellularmelanincontentinsupernatantswasmeasuredspectrophotometricallyat
anabsorbanceof420nmagainst a standardcurveofknownconcentrationsof synthetic
melanin (0-50µg/mL, Sigma-Aldrich, France).Experimentswereperformed induplicate.
Resultswereexpressedinµgmelanin/mL.
III.II.3Dbiologicalmodel
III.II.I.Fibroblasts-populatedtensecollagenlatticepreparation(FPCL)
Primary fibroblast cultures were prepared from adult human facial skin (passage 4-6).
Fibroblastswerebroughttoaconcentrationof8x105cells/mLinfullDMEMmedium.They
wereembeddedinthree-dimensionalhydratedcollagengels74,75.Eachcollagenlatticewas
preparedbymixing,inthefollowingorder:3mLof1.76XconcentratedDMEM(containing
0.5%NaHCO3,0.1NNaOH,15%FBS),1.5mLofrattailtypeIcollagensolution(2mg/mL,
InstitutdeBiotechnologiesJacquesBoy,France)and0.5mLoffibroblastsuspension.The
mixturewasquicklypouredintoa16X60-mmpetri-dishcontainingasterilizednylonmesh
ring(SefarNitex03-150/50,HeidenSwitzerland)(Figure14).Afterpolymerizationat37°C
for 1 h, 3 mL of full DMEM mediumwereadded.
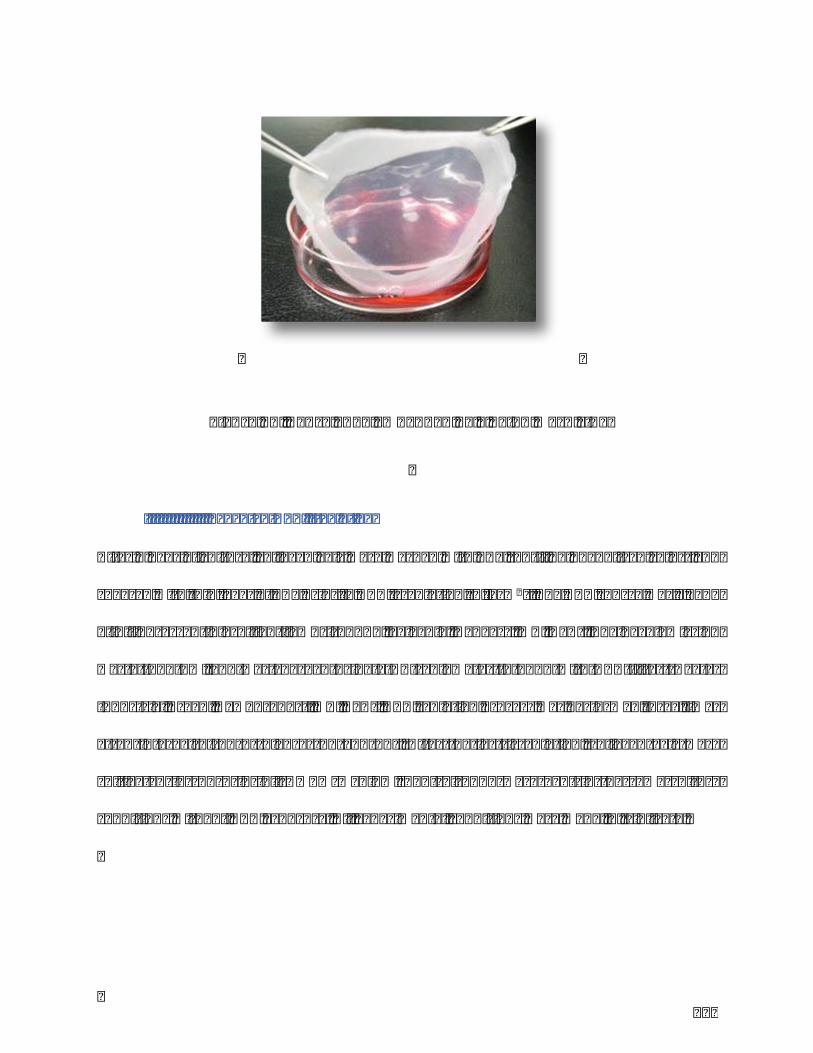
118
Figure14.TenseFPCLembeddedbyanylonmeshring
III.II.II.Repeated-UVAirradiation
After5daysofculture,collagengelswerewashedwithPBS(untilnophenol-redcolor)and
coveredwiththinlayerofPBSpriortoUVAirradiation(6J/cm2).TheUVAsourcewasabank
of8fluorescentblacklightlamps(F15T8,Sylvania,Danvers,MA,USA).Thepeakemission
wasat370nm,andwascheckedusingaUVradiometer IL-1700withUVA filter (Dexter
IndustrialGreen,Newburyport,MA,USA).UVAradiationprocesswasperformedthreetimes
at an interval of 48 h (Figures 15 and 16). After the last irradiation, collagen gelswere
cultivated for 24 h in full DMEMmedium. Control FPCL were kept in the same culture
conditionswithoutUVAexposure.Allexperimentalconditionsweremadeintriplicate.
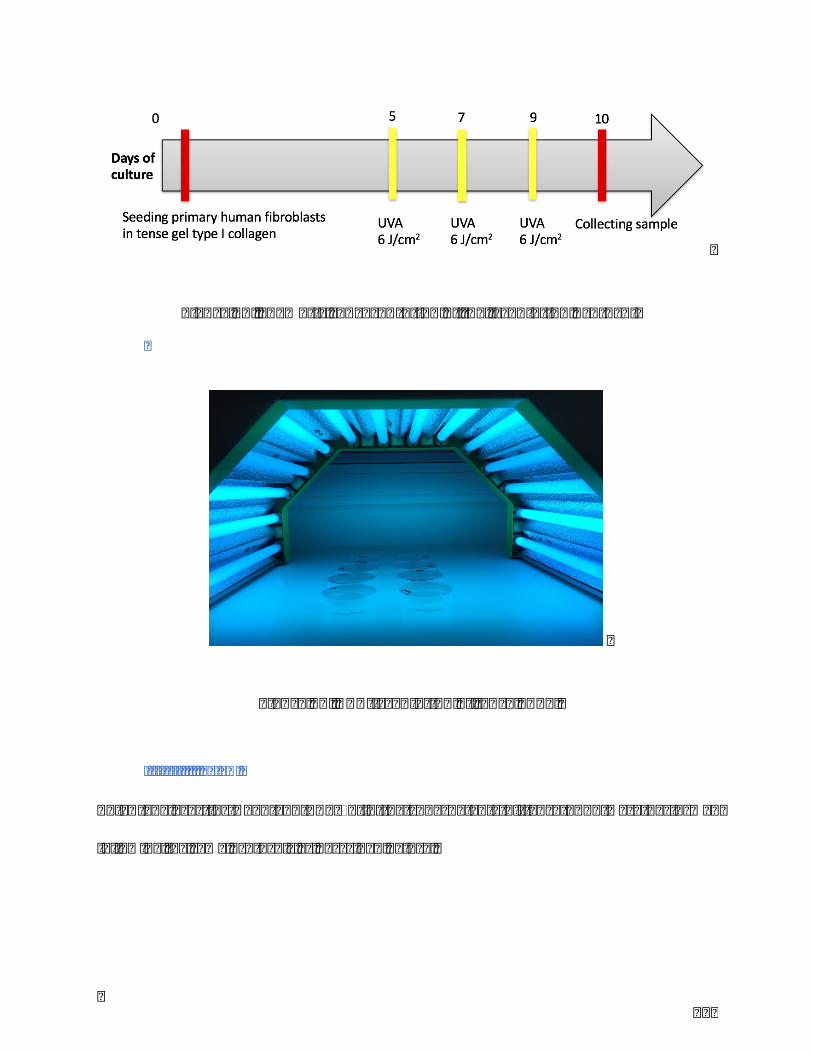
119
Figure15.Shematicrepresentationoftheirradiationprotocol
Figure16.UVA-irradiationoftenseFPCL.
III.III.ELISA
ELISAanalyzes forDkk1andTGF-b1 in the supernatants of tenseFPCLwereperformed
followingthesameprotocolsasdescribedbefore.

120
III.IV.Statisticalanalysis
All quantitative data are expressed as means ± SD. Paired Student’s test (SPSS 24.0) was used to
compare groups. p < 0.05 was considered significant. (* p<0.05, ** p<0.01, *** p<0.001)
IV. Results
IV.I.ComparativeanalysesofstaininginSLversusSNbiopsies
Stainingtechniquessuchashistochemistry(HC)andimmunohistochemistry(IHC)provide
useful informationregarding theskinstructure, the localizationandrelativeamountofa
moleculeinskin.
SLandSNtissuesdisplayeddifferenthistological features.SLshoweda thinepidermis,a
disorganized basementmembrane and elongated rete ridges collapsing into the dermis.
Masson-Fontanastainingdemonstratedhighermelanindeposition intheepidermalbasal
layer of SL compared to SN. The disruption of epidermal basal layer in SL caused some
melaninreleaseintothedermis.β-cateninstainingpresentedmorepositivestaininginSL
comparedtoSN(Figure17).Altogether,histologicalanalysisvalidatedtheSLoriginofour
invitrofibroblasticmodel.
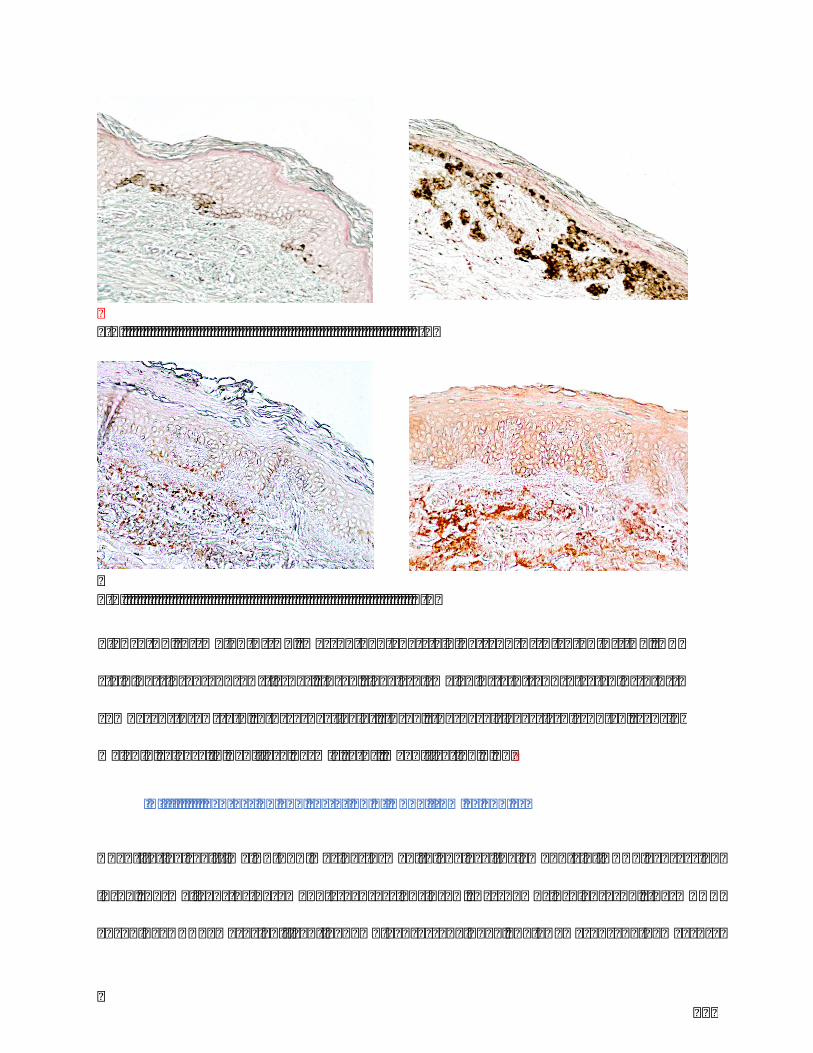
121
a-1a-2
b-1b-2
IV.II.ExpressionandsecretionofDKK1fromFLandFL
Quantitativereal-timeRT-PCRwasperformedtoanalyzetheamountsofDKK1expression
in FL, normalized to the amounts expression in FN. As shown in Figure 18, the mRNA
encoding DKK1 was significantly down-expressed in FL. ELISA was used to measure
Figure17. a)Melanin (HC,Masson-Fontana staining) andb) β-catenin (IHC, DAB
staining) in SL and SN tissues. In SL, levels of melanin (a-1) and β-catenin (b-1) ↗
compared to SN (a-2, b-2 respectively). In SL, the basal layer is disrupted, causing
melaninreleaseinpapillarydermis(a-2).MagnificationX20.
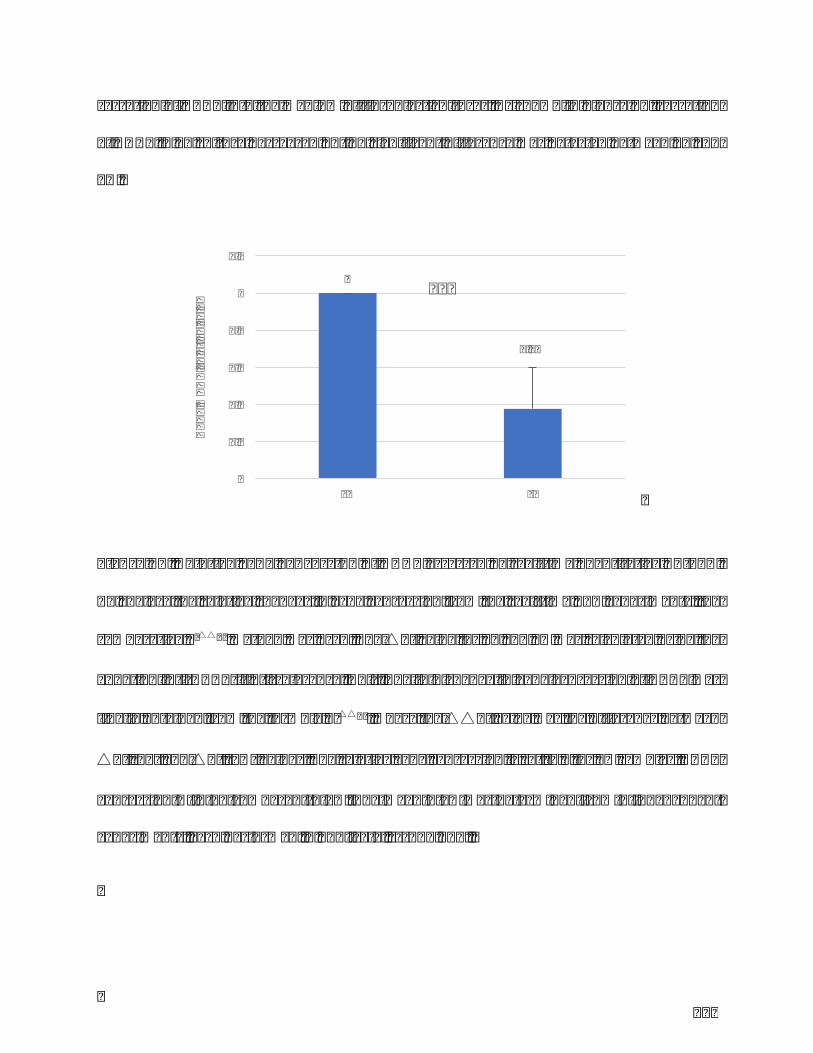
122
secretionofDKK1intothemediumoffibroblastcultures.AsshowninFigure19,secretion
ofDDK1inFLculturedecreasedbutnosignificantdifferencewasdetectedbetweenFLand
FN.
Figure18.RelativegeneexpressionofDKK1assessedbyreal-timequantitativeRT-PCR.
Toanalyzetherelativechangesingeneexpressionfromthereal-timePCRexperiments,the
comparative2-△△Ctmethodwasused.The△CtvalueforFLandFNwascalculatedusingthe
equation:Ct(DKK1)-Ct(reference,Abl).ThefoldchangeingeneexpressionofDKK1was
finallyobtainedfromtheformula2-△△Ct,wherethe△△Ctvaluewasthedifferencebetween
△Ct(FL)and△Ct(FN)values.Therelativegeneexpressionissetto1forFNsamples.DKK1
expression↘inFLcomparedtoFN.Themean±SDwasdeterminedfrom9 independent
experiments,eachperformedinduplicate.***p<0,001.
1
0,38
0
0.2
0.4
0.6
0.8
1
1.2
FN FL
DKK1
mRN
Arelativ
evalue
***
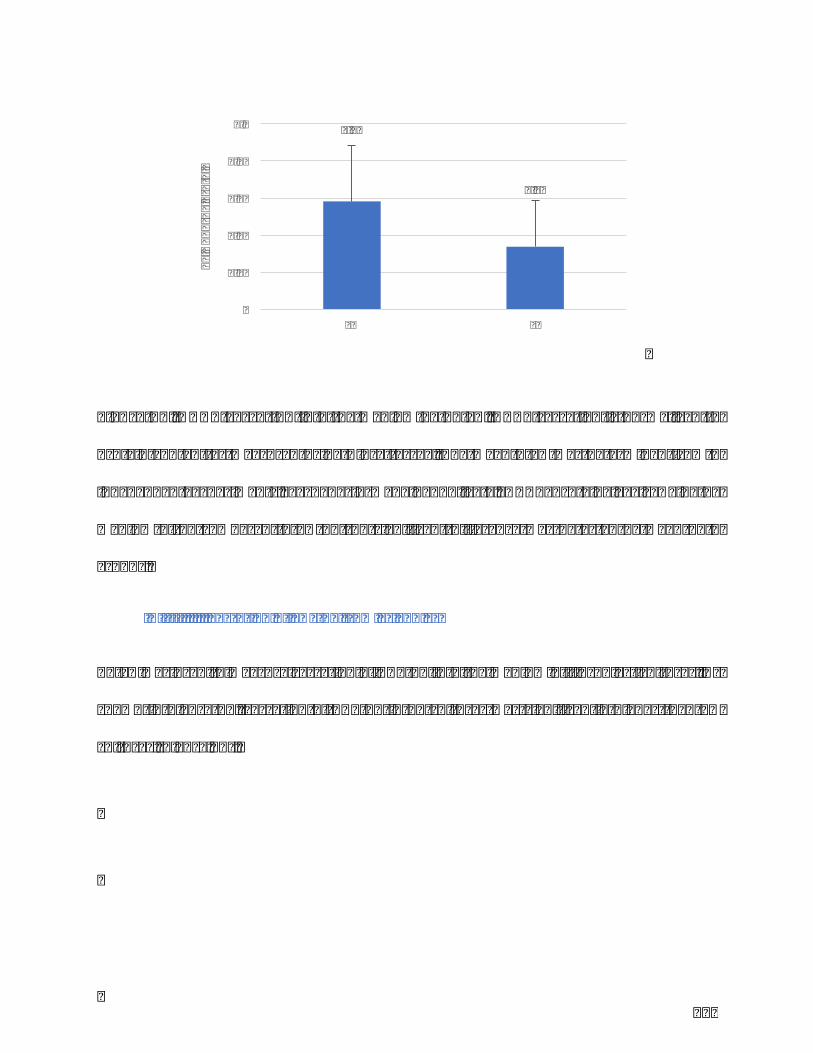
123
Figure19.DKK1secretion into themediumbyELISA.DKK1secretion isnormalized to
protein content asmeasuredusingBiuret assay.Themean± SDwasdetermined from9
independent experiments, each performed in duplicate. DKK1 protein levels ↘ in the
medium of FL compared to FN but no significant differencewas detected between both
groups.
IV.III.SecretionofTGF-β1fromFLandFL
ELISAwasusedtomeasuresecretionofTGF-β1intothemediumoffibroblastcultures.As
shown in Figure 23, secretion of TGF-β1 in FL cultureswas significantly higher than FN
cultures(Figure20).
0.06
0.03
0
0.02
0.04
0.06
0.08
0.1
FN FL
pgDKK
1/pgprotein
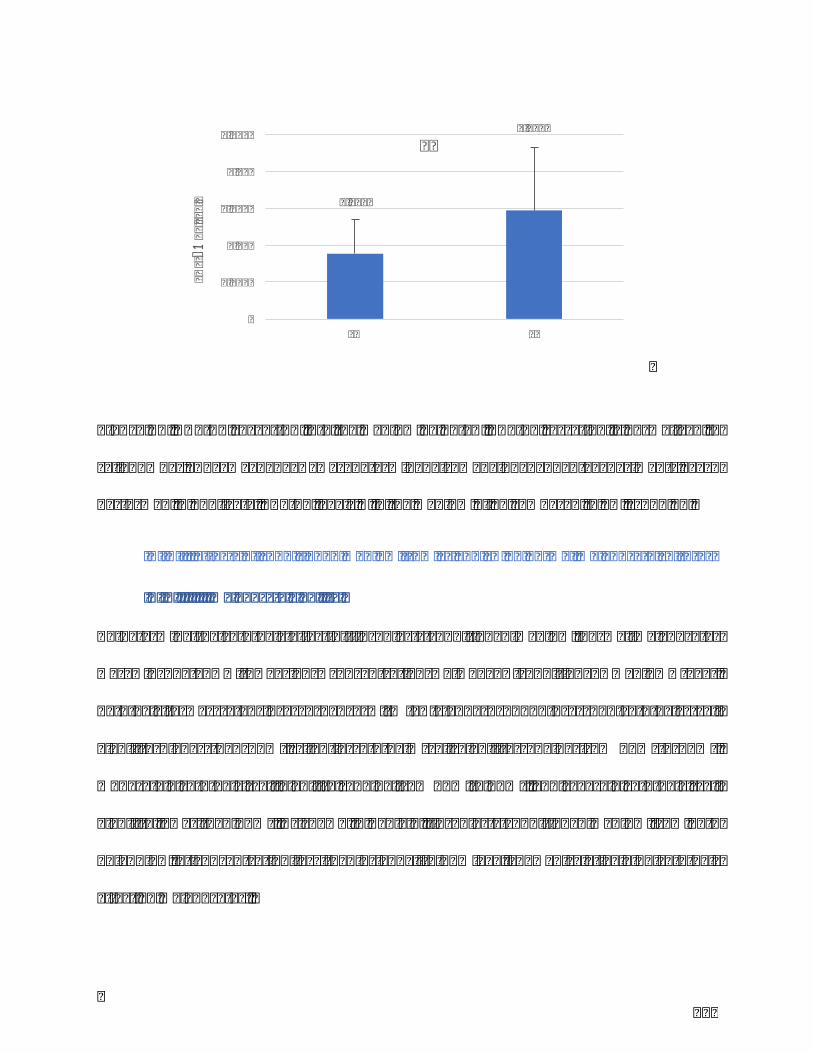
124
Figure20.TGF-β1secretionintothemediumbyELISA.TGF-β1secretionisnormalizedto
cell number. The mean ± SD was determined from 9 independent experiments, each
performedinduplicate.TGF-β1levels↗inthemediumofFLcomparedtoFN.**p<0,01.
IV.IV.EffectsofconditionedmediumfromFLandFNonnormalmelanocytecultures
IV.IV.I.Melanocyteviability
Todeterminethecytotoxiceffectof fibroblasts-conditionedmedium,normalmelanocytes
were incubated with various concentrations of FL/FN-conditioned medium (v/v).
Cytotoxicity was rated in accordance with ISO-standard 10993-5 as non-cytotoxic (cell
viability higher than 75%), slightly cytotoxic (cell viability ranging from 50% to 75%),
moderatelycytotoxic(cellviabilityrangingfrom 25%to50%),andseverelycytotoxic(cell
viabilitylowerthan25%).AsshowninTable4,fibroblasts-conditionedmediumfromboth
FLandFN,atconcentrationlessthanorequalto15%v/v,showedrelativelynocytotoxic
effectonmelanocytes.
0.0009
0.0015
0
0.0005
0.001
0.0015
0.002
0.0025
FN FL
TGF-β1
pg/cell
**
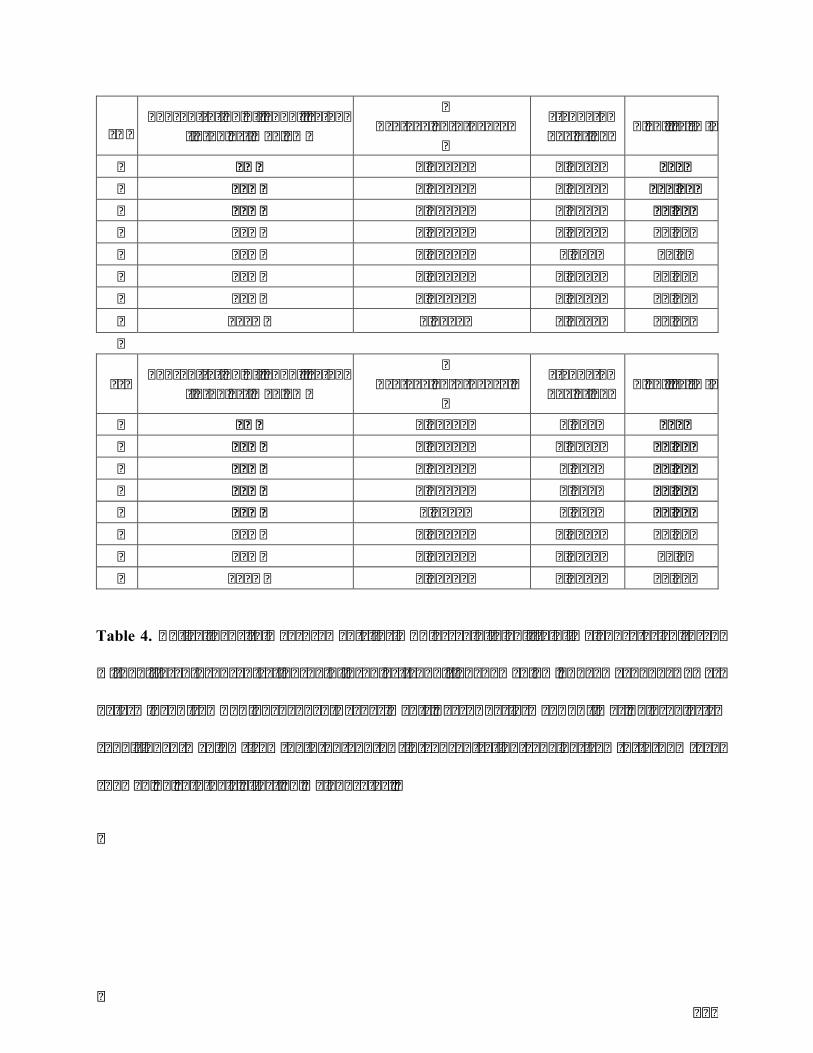
125
FNConcentrationoflyophilizated
fibroblastmedium
Averageabsorbance
Standarddeviation Viability(%)
0% 0.61583 0.0848 100 10% 0.61623 0.1564 100.06 15% 0.52647 0.0766 85.49 20% 0.30455 0.0617 49.45 25% 0.38493 0.022 62.5 50% 0.28323 0.0429 45.99 75% 0.25637 0.0325 41.63 100% 0.2873 0.0485 46.65
FL Concentrationoflyophilizatedfibroblastmedium
Averageabsorbance
Standarddeviation Viability(%)
0% 0.46867 0.101 100 10% 0.42543 0.0626 90.78 15% 0.42123 0.533 89.88 20% 0.38343 0.322 81.81 25% 0.3582 0.082 76.43 50% 0.27873 0.0326 59.47 75% 0.27087 0.0353 57.8 100% 0.21635 0.0624 46.16
Table 4. OpticaldensitymeasurementsforMTTassayofviabilityofmelanocytescultured
with different concentrations of fibroblast-conditioned medium. The mean ± SD was
determined from 3 independent experiments, each performed 4 times. Fibroblasts-
conditionedmedium from both FL and FN in concentration ranging from 0 to 15% v/v
showednocytotoxiceffectonmelanocytes.

126
IV.IV.II.Melanocytecharacteristics
In accordance with MTT results, the concentration of 15% v/v was chosen to evaluate
melanocytemorphologyandMITFexpression.
Following FL-conditioned medium treatment, melanocytes showed increased dendritic
morphology and intracellular melanin (Figure 21). These findings were supported by
stainingofMITFandF-actin,usedrespectivelyasacharacteristicmelanocytemarkeranda
cellshapemarker,revealingapositiveMITFexpressionandaclassicaldendriticmorphology
(Figure22).
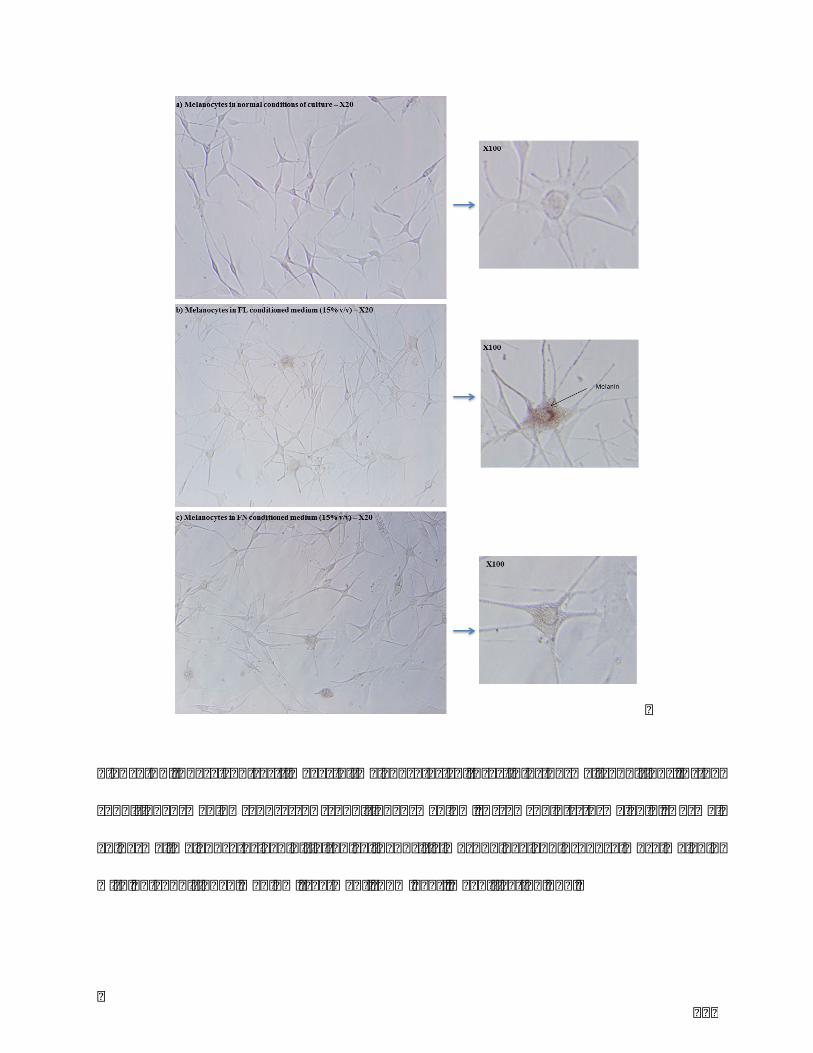
127
Figure21.Phasecontrast imagesofmelanocytescultures ina)normalconditions,b)FL
conditioned medium and c) FN conditioned medium. Arrow points to melanin. Normal
epidermalmelanocytesexhibitatypicaldendriticmorphologyandproducemoremelanin
withFL-conditionedmediumtreatment(15%v/v).MagnificationX20.
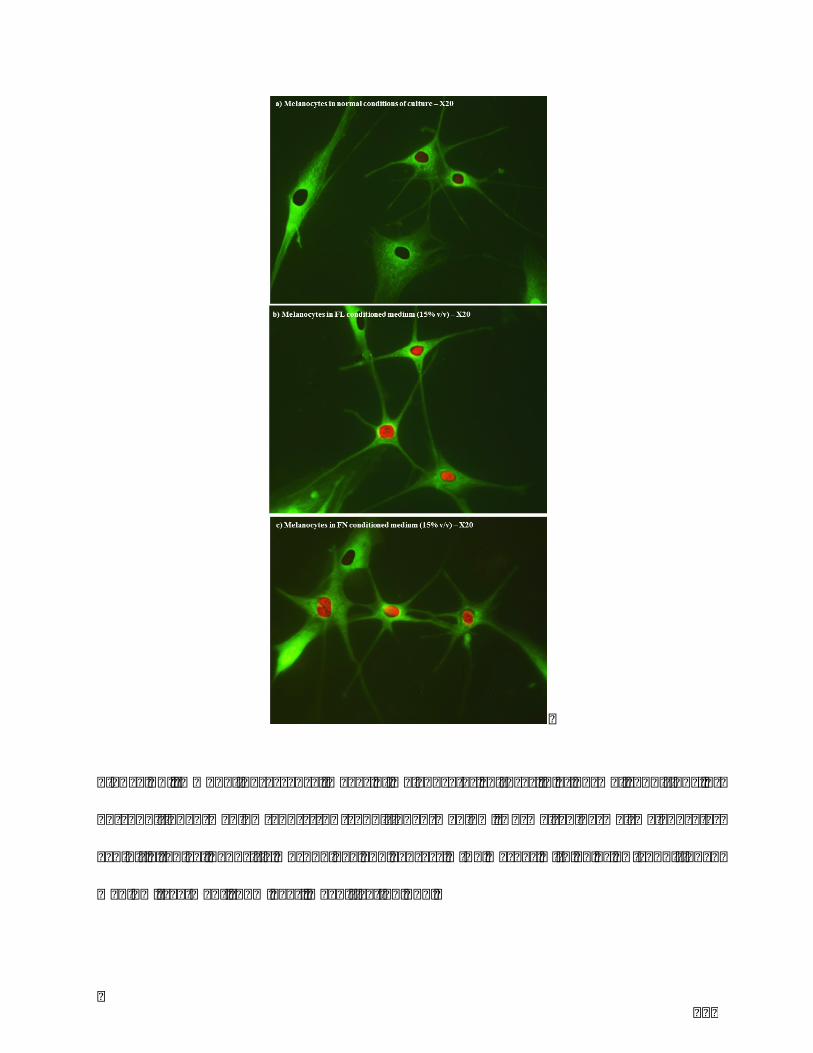
128
Figure22.Immunofluorescenceimagesofmelanocytesculturesina)normalconditions,b)
FL conditionedmedium and c) FN conditionedmedium. Normal epidermalmelanocytes
exhibitatypicaldendriticmorphologyandexpressMITFmarkerwithFLorFN-conditioned
mediumtreatment(15%v/v).MagnificationX20.
129
IV.V.SecretionofDKK1afterirradiationofnormalfibroblastsembeddedin3Dcollagen
gel
Under6J/cm2repeatedUVA-treatment,facialfibroblastssecretedsignificantlylessDKK1
thanfibroblastwithoutUVAtreatment(Figure23).
Figure23.DKK1secretionintothemediumbyELISA.TGF-β1secretionisnormalizedto
cell number. The mean ± SD was determined from 3 independent experiments, each
performedinduplicate.DKK1levels↘inthemediumafterUVirradiation.***p<0,001.
V. Discussion-Perspectives
Theagingprocessinhumanskinresultsnotonlyinelasticityloss,dryandthinskinbut
also in benign lesion development such as solar lentigo (SL). Recent studies have
demonstratedthatfibroblastsareactivatedbyUVexposuretoreleasemelanogenicgrowth
factors (HGF, KGF, SCF) which act on melanocytes both directly and indirectly through
0.27
0.08
0
0.05
0.1
0.15
0.2
0.25
0.3
Non-UVA UVA
DKK1
pg/cell
***

130
keratinocytesand,thereforemaycontributetothehyperpigmentationofSL.Theprevious
research in our lab has identified cellular senescence-like features of fibroblasts derived
fromSL76.Otherfactorsareknowntobeinvolvedinskinpigmentation,suchasDKK1which
regulateshypopigmentationinvitiligoandpalmoplantarskin,andinhibitscellinvasionand
angiogenesisinmelanomadevelopment31,40,54,77,78.
ThisstudyinvestigatesanewpossibleroleofDKK1inhyperpigmentation,byusingsolar
lentigo(SL)andnormaladjacent(SN)skinbiopsiesfromthesamepatient,whichprovidesa
more precision evidence for the research. Our experimental data indicate a reduced
expression and secretion of DKK1 in fibroblasts from solar lentigo (FL) compared to
fibroblasts from normal adjacent skin (FN). As previous studies described, DKK1 is an
antagonisticinhibitoroftheWntsignalingpathway,byisolatingtheLRP5/6co-receptorand
preventing the Frizzeled-Wnt-LRP5/6 complex formation both in melanocyte and
keratinocyte31,77. The down-expression of DKK1 in FL compared to FN highlights the
potentialroleofDKK1inthehyperpigmentationprocess.ThisdecreaseofDKK1fromFL
maybecausedbydifferentendogenousandexogenousfactors,suchaspro-inflammatory
cytokines.
It was found that TGF-b1 stimulates canonical Wnt signaling in p38-MAPK by
downregulatingDKK1infibroticfibroblast.Weaimedtoexplorethehypothesisthathigh
levels of TGF-b1 may downregulate DKK1 by p38-MAPK signaling pathway in SL. We
reportedthatthesecretionofTGF-b1fromFLwashigherthanfromFN.
Tofurtherverifythehypothesis,wetriedtomimicinvitrotheenvironmentofsolarlentigo
andreproducetheskinphotoagingprocessunderpro-inflammatoryconditionsviaTGF-b1.
WehavechosenthemodelofUVA-irradiated3Dtensecollagengel. In3Dtensecollagen

131
lattices,fibroblastsundergomechanicalstress,differentiateinmyofibroblastsandsecrete
TGF-b1.Besides,UVA-irradiationwassetuptoreproduceUVexposurewhichisthemost
exogenousfactorinducingsolarlentigo36.ExperimentaldataindicatedthatUVAtreatment
couldstimulateTGF-b1secretionandinhibitDKK1expressionbyfibroblastsembeddedin
collagenmatrix.Toverifymorepreciselyourhypothesis,futureexperimentswillfocuson
theinhibitionofthep38MAPkinasepathway.
DKK1regulatesmelanocytefunctionviatheregulationoftwoimportantsignalingfactors,
MITFandβ-catenin.AdecreaseofDKK1expressioninducesanactivationofWnt/β-catenin
signalingassociatedwithanincreaseofmelaninandβ-catenin,whichwasconfirmedbyour
histologicalobservations.
In addition, undefined conditioned medium of fibroblast derived from SL can stimulate
melanin synthesis inmelanocytes. These results confirmed thatmelanogenic factors are
secretedbyfibroblasts.
ThereforeallthesedatarevealalinkbetweenTGF-β1,DKK1andtheWnt/βcateninpathway:
UVirradiationÞ↗TGF-β1Þ↘DKK1ÞupregulationofWnt/βcatenincascadeÞ↗MITF
Þ↗melaninÞSLlesion.
This process may result in an uneven distribution of active melanocytes, with areas of
hyperpigmentationintheskin.
In the future, it would be a good practice to do co-cultures of melanocyte and
keratinocyteinfibroblast-conditionedmediumtofurtherexploretheinfluenceofDKK1in
hyperpigmentation both in melanin synthesis in melanocyte and melanin taken by
keratinocyte.

132
Up-regulationofmelanocytefunctionscausedbyagingandUVirradiationisinvolvedinthe
development of solar lentigo lesions or other age-related hyperpigmentation disorders.
Cosmeticcompaniesusuallydevelopanti-photoagingproductsbyfocusingonmelanocyte
and keratinocyte functions. This study brings new approach to prevent and treat solar
lentigo lesions, by controlling the expression and secretion of several fibroblast-derived
solublefactors.

133
ListofPublications&Communications

134
Publications
• YJ.Wang, C. Viennet, S. Robin,M. Tissot, P.Muret, L.He, JY. Berthon, P.Humbert.
InvolvementofDickkopf-1inmelanogenesis:focusonsolarlentigo.(Inpreparation)
• YinjuanWang,Céline.Viennet,LiHe,PhilippeHumbert.Assessmentoftheefficacy
of a new complex anti-sensitive skin cream. (Original Article. Submitted to Journal of
CosmeticDermatology)
• YinjuanWang,CélineViennet,GwenaëlRolin,MarionTissot,PatriceMuret,Sophie
Robin, Jean-Yves Berthon, Li He, Philippe Humbert.Development and validation of a
simple method for the extraction of human skin melanocytes. (Original Article.
SubmittedtoCytotechnology)
• YJ. Wang, C. Viennet, S. Robin, JY. Berthon, L. He, P. Humbert. Precious role of
fibroblast on melanocyte pigmentation. Journal of Dermatology Science.
http://dx.doi.org/10.1016/j.jdermsci.2017.06.018

135
Posters
§ InvolvementofDickkopf-relatedprotein1inmelanogenesis:focusonsolar
lentigolesion
SkinPhysiologyInternationalMeeting,Vichy,France,2016
§ Developmentandvalidationofasimpleandcommonmethodforthe
extractionofepidermalcells
• CongrèsAnnuelDeRechercheDermatologique,Lille,France,2015
• EuropeanTissueRepairSociety,Copenhagen,Danmark,2016
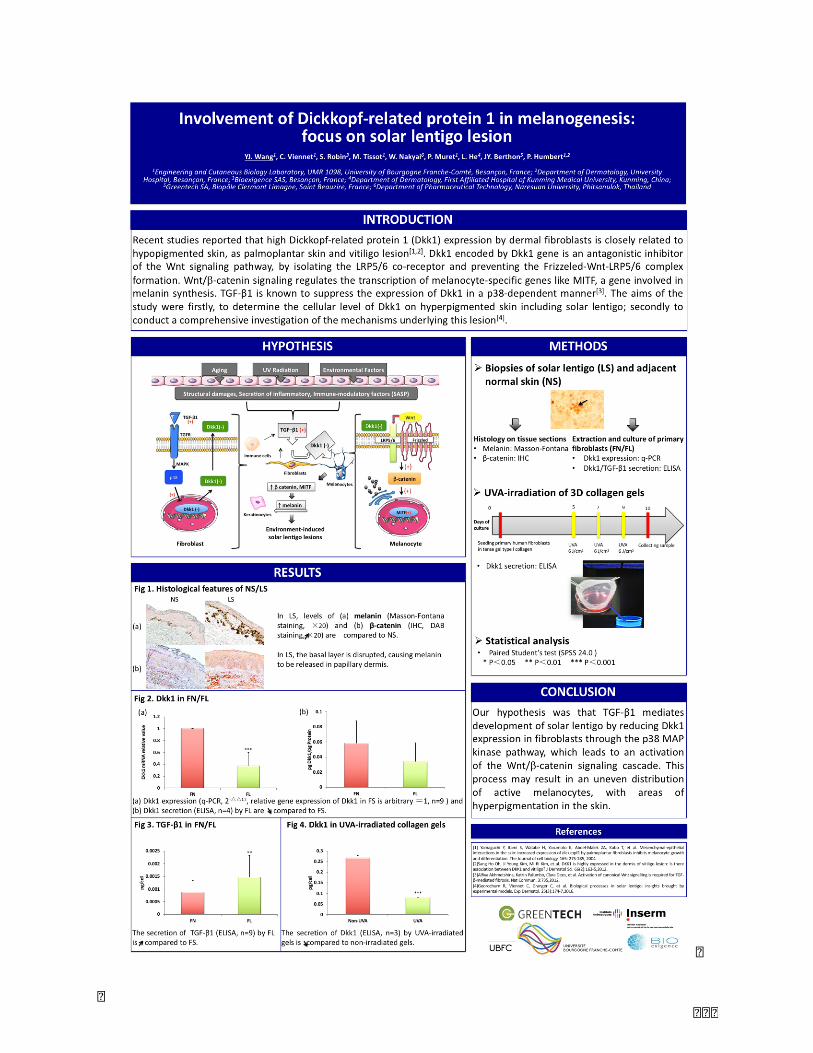
136

137

138
Article3:Assessmentoftheefficacyofanewcomplexanti-sensitiveskincream
YinjuanWang, MD,1 Céline Viennet, PhD,1 Adeline Jeudy, PhD,2 Ferial Fanian, MD,
PhD,2LiHeMD,PhD,3*PhilippeHumbert,MD,PhD,1,2*
1Engineering and Cutaneous Biology Laboratory, UMR 1098, University of Bourgogne
Franche-Comté,Besançon,France
2Research and Studies Center on the Integument (CERT), Department of Dermatology,
UniversityHospital,Besançon,France
3Department of Dermatology, First Affiliated Hospital of Kunming Medical University,
Kunming,China
*Correspondingco-authors:PrPhilippeHumbertandPrLiHe
E-mail addresses for correspondence: [email protected] or
Disclosureandconflictsofinterests:theauthorshavenoconflictofinteresttodeclare.

139
Abstract
Background Sensitive skin is frequently complaint in dermatology consultation with
cutaneous manifestations such as stinging, redness, dryness and burning sensation that
affectthequalityoflife.Itspathogenesisismainlyrelatedtodysfunctionofneurosensory,
skinbarrier,andalsoimmuneactivity.Thetreatmentisgenerallybasedoncontinuousand
topicaltherapybynon-irritatingcomplex.
Objective:Toevaluate theanti-sensitive functionof anewcomplex creamcomposedby
Yunnan Portulaca olreacea extracted, Prinsepia utilis oil, beta-glucan, and sodium
hyaluronateextractedfrommushroom.
MethodsArandomizeddouble-blindandself-controlstudywasconductedon20selected
volunteerswithsensitiveskin.Subjectsappliedthetestcreamtoonesideoftheface,andthe
controlcream(toleranceextremecream)totheothersideoftheface,twicedailyover28
days.Evaluationswereperformedatbaselineandat28days.Expertclinicalgradingoffacial
skinincludingdryness,roughness,desquamationanderythemawasassessed.Subjectself-
assessment questionnaires, digital photography and noninvasive bioinstrumentation of
hydration, transepidermalwater loss, lipid index, skin texture andwettabilitywere also
includedinthestudy.
ResultsProductswerewelltolerated.Forallparametersstudied,nosignificantdifference
wasobservedbetweentestandcontrolcreams.Resultsshowedthattestcreamprovideda
statisticallysignificantimprovementinclinicalgradingscoresfordryness,roughnessand

140
erythemaat28dayscomparedtobaseline.Inaddition,statisticallysignificantimprovement
of skin hydration and texture parameters (eg, smoothness and roughness) were
demonstrated. Volunteers’questionnaire revealed self-perceived benefits consistent with
expertvisualgrading.
ConclusionThisstudyconfirmedtheeffectivenessandtoleranceofthenewcomplexcream
insubjectswithsensitiveskin.Thetestcreamcouldserveaasadailycaremoisturizerfor
face.
Keywords:skin,clinicaltrial,sensitive,hydration,cream

141
Introduction
Sensitive skin is a frequent complaint indermatology consultation. It isdefinedby some
clinicalmanifestations includingstinging, redness,burning,pruritus, tingling thathappen
mainly on the face but also on other areas such as scalp and hands.1 The potential
mechanisms of sensitive skin involve skin neurosensory dysfunction,2,3 neurogenic
inflammation,4epidermalbarrierdisruption(increasingtransepidermalwaterloss)5-7and
immunecellsactivity(transientreceptorpotentialchannels).Variousconditionsinducethe
symptomsofsensitiveskin,suchasenvironmentalfactors(e.g.UVradiation,heat,cold,hash
winds, pollution), lifestyle factors (e.g. cosmetic usage, diet and alcohol), and even
psychological factors (e.g. stress) underlying dermatological diseases as atopic
dermatitis.3,8,9Thediagnosisofsensitiveskincouldbeassistedbystingingtests,occlusion
tests,behind-the-kneetests,washingandexaggeratedimmersiontests,totheevaluationof
itching and quantitative sensory testing (QST).10 In Europe, approximately 39% of the
populationarebotheredbysensitiveskin,andfemaleskinisdeclaredmoresensitivethan
maleskin.ApparentlysensitiveandverysensitiveskinshowedhighincidenceinFranceand
Italy.11 In response to this prevalence, the effective treatment of sensitive skin becomes
challenging.Continuousandtropiccareplaysasignificantroleintreatmentofsensitiveskin
whether in daily care or in the combination of drug treatment.Mildness, UV protection,
hydrationarethefundamentaldemandingfortropicdailycareofsensitiveskin,andthere
areseveralcosmeticsandcosmeceuticalsforsensitiveskinaccordingtothisprinciple.

142
Inthisarticle,weevaluatedananti-sensitivemoisturizingtolerance-extremecreamthatis
an innovative formulationmainly composedbyChinaYunnanPortulacaolreaceaextract,
Prinsepia utilis oil, and beta-glucan, sodium hyaluronate extracted from mushroom.
Portulaca oleracea has anti-inflammatory and anti-allergy effects, it can relieve skin
irritationandenhanceskintolerance.12Prinsepiautilispromotesthesynthesisofceramide
bystratumcorneumcells,andthereforecaninducerepairoftheskinbarrier.13Mushroom
beta-glucanstimulatesproliferationofkeratinocytesand,thuspromoteshealingofdamaged
skin.14Weperformedamonocentric,double-blinded,controlledandrandomizedstudyina
panelof20volunteersusingclinicalandinstrumentalevaluations.
MaterialandMethods
Studydesign
Thisrandomizeddouble-blindcomparativestudywascarriedoutattheCERT(Researchand
StudyCenteronIntegument)locatedinthedepartmentofDermatology,UniversityHospital
of Besançon, France. The study protocol was approved by the ethics committee of the
institute.Thisstudywasconductedinaccordancewiththeethicalprinciplesderivedfrom
the Declaration of Helsinki with Good Clinical Practices (ICH E6) and the law of local
regulation clinical studies, and was considered as a non-interventional study (directive
2001/20/CE,decreedated27April2006).The finalprotocol and the relateddocuments
werereviewedbytheCPPEst2ofFranche-Comté,Besançon,aswellasANSM,Paris,France.

143
StudySubjects
20femalevolunteersaged37.1±10.6yearsoldwithFitzpatrickskinphototypesI–IVand
dryandsensitiveskin,participatedinthisstudyafterhavinggiventheirwritteninformed
consent.Theywereselectedbasedondermatologist’smedicalexamination,measurement
of skin hydration (facial hydration index < 60, Corneometerâ CM825, Courage-Khazaka
Electronic)andsensitivity(lacticacidstiningtest).Exclusioncriteriawerethepresenceof
any skin-related pathologies and abnormalities (eczema, psoriasis…), allergies,
hypersensitivity to the testedproduct, acuteand/or chronic inflammationor infectionof
facial skin, an exposure of sunlight or artificial UV rays within 15 days, pregnancy and
nursing.Subjectswereadvisedtoavoidtheapplicationofothersimilarproductduringthe
wholestudy.
Testproduct
The test product is an anti-sensitive moisturizing tolerance-extreme cream which is
manufacturedbyBeitainiBio-technologicalCo.,Ltd.(China).Atoleranceextremecreamwas
used as control. Both products were provided by Beitaini Bio-technological Co.,Ltd.
IngredientsoftestandcontrolcreamswerelistedinTable1.
Treatment
Volunteersservedastheirowncontrolswithonesideofthefacereceivingthetestcream
andtheothersideofthefacereceivingthecontrolcream.Evaluationswereperformedat

144
baseline(T0)and28days(T1)aftertwicedailyapplication.Volunteersstayedinacontrolled
room (temperature 22±2°C, humidity 50±10%) for at least 20 minutes before each
evaluation.Theywereevaluatedinthestandardskinsituation(lastfacewashingthenight
before visit, without any cosmetic, water and makeup application until the measures).
Measurements were recorded using a flexible transparent plastic mask. This allowed
measurements to be done in exactly the same place at each visit (Thermocell-Test®
professionalkit,Monaderm).
Evaluation
Clinicalscores
Visualclinicalscoreswererealizedincludingdryness,roughness,desquamation,erythema
witha5-pointscale(0=none,1=slight,2=mild,3=moderate,4=severe).
Self-assessment
Subjectscompletedaself-assessmentquestionnaireatT0andT1(Table2).
Photographies
AtT0 andT1, digital imageswere takenusing a standardizedphotography system (SSA,
Courage-KhazakaElectronic)connectedtoaCanonPowerShotA620camera. Allfrontaland
profilephotographswerestandardizedforlightandposition.

145
Bioinstrumentation
Transepidermalwaterloss(TEWL)
The skinbarrier functionof the stratum corneumwasmeasuredbyVapometer® (Delfin
Technologies).Thesensormonitorinclosedcylindricalchambermeasurestheincreasing
relative humidity (RH) of evaporated water from the skin covered by chamber. The
evaporation rate value was expressed in g /m²/ h. One assessment was performed on
externalsideofeachcheekbone.
Skinhydration
Skin hydration was determined on each cheekbone by the measurement of the electric
capacitance(i.e.thecapacityoftheintercellularwatertoconductelectrons)ofthestratum
corneum using a Corneometerâ CM825 (Courage-Khazaka Electronic). The mean value
(hydrateindex,HI)ofthreeassessmentswascalculated.
Lipidindex
Quantitativeanalysisof thesebumwasmeasuredon foreheadbyaSebumeter®SM815
(Courage-KhazakaElectronic).Valueswereexpressedinmg/cm2.Themeanvalueofthree
assessmentswascalculated.

146
Skintexture
Visioscanâ VC98 (Courage-Khazaka Electronic) was used for assessing skin texture and
roughness.15 It takes pictures of the skin surface under standardized homogenous ring-
shaped UVA illumination. Evaluation of images was carried out according texture
parameters: energy reflectingoverall skin condition (NRJ), skin smoothness (Sesm), skin
roughness(Ser),andarithmeticmeanroughness(R5).Themeanvalueofthreeacquisitions
oneachtemplewascalculated.
Skinsurfacewettability
Themeasurementof the contact angle formedbetween the skin surface andwater is an
indicatorofitslipophilicorhydrophilicproperty,thusthecontentinlipids.Thesystemis
composedofasurgicalmicroscope(WildM650Heerbrugg®,Leica)withamirrororiented
at45°totheskinsurface.Adropofdistilledwaterwasdepositedontheskinsurfaceand
thecontactanglewasmeasuredusingsoftwarebasedonthegeometricaldimensionsofthe
drop.16Themeanvalueofthreemeasuresonforeheadwascalculated.
Skincolor
TheMinolta400ChromaMeterisacolorimeterthatcombinesasource-filter-photodetector
andaunitthatdirectlycomputesthechromaticdimensionsofthecolour.Themeanvalueof
threemeasuresoneachcheekbonewascalculated.

147
Sensitivity
Stingingtestisnowacceptedasaroutinelytestandisusedfortherecruitmentofsubjects
with sensitive skin. Itmeasures the skin reactivity particularly in the face. It consists of
applyinga10%lacticacidatthenasolabialfoldcomparedwithphysiologicalserumapplied
simultaneously to the other side. Subjects assessed stinging based to a 4-point scale
(0=nothing,1=slighting tingling,2=moderate tingling,3= intense tingling)at10s,2.5min
and5min.
Statisticalanalysis
The statistical analysis was conducted using Statistica Version 7.1 and Graphpad Instat
Version3.06.DistributionofthedatawasassessedusingtheShapiro-Wilktest.Statistical
comparisonsofthedatawerecarriedoutwiththeAnovaorWilcoxontest(dependingofthe
normalityofthedistribution).Dataarepresentedasmean±standarderrorofmean(SEM).
Differenceswithp<0.05wereconsideredtobestatisticallysignificant.
Results
Test and control creams were well tolerated. For all parameters studied, no significant
differencewas observed between test and control creams. Only data for test cream are
shown.
Highresolutiondigitalimageswerecapturedforallsubjectsatbaselineandafter28

148
daysoftreatment.RepresentativeexampleofonesubjectisshowninFigure1.Visualgrading
analysis demonstrated that after test cream application, a significant improvement of
dryness, erythema, and roughness was observed (Figure 2). At T0, 85% of volunteers
exhibitedslightandmild skindrynesswhereasatT1after test cream treatment,90%of
volunteers had none and slight skin dryness (P<0.0001). 25% of volunteers showed
moderateskinroughnessbeforetreatment,whichpopulationdisappearedaftertreatment.
AtT1,roughnessscoresdecreasedwithonlynonetomildlevels(P=0.0107).AtT0,nearly
50%of volunteers exhibitedmild erythemawhereas atT1, thispercentagedecreased to
14%.Inadditionafter28daysoftreatment,10%and60%ofvolunteershadrespectivelyno
andslighterythema(P=0.0210).However,forskindesquamation,nosignificantdifference
wasnotedbetweenT0andT1.
Thehydrationindexofskinwasscoringfrom48.7±9.5atbaselineT0to57.2±10.6at
T1 after test creamapplication,which reflected a significant increaseof the skin surface
hydration(P=0.0017)(Figure3).
Skintextureparametersincludingroughness(Ser)andsmoothness(Sesm)showed
significantdecline(respectivelyP=0.0333andP=0.0111)aftertestcreamtreatment,andthe
textureparameterofenergyshowedsignificantincrease(P=0.0045)(Figure4).
However,nosignificanteffectwasobservedforskinbarrierfunction(TEWL),sebum
recovery(lipidindex),wettability,colorandstingingtest(datanotshown).
Resultsoftheself-assessmentquestionnaireshowedthattestcreamwassignificantly

149
favourable for 3 questions (Figure5). Subjects considered their facial skin less tightness
(p=0.0169), more hydrated (p=0.0004) and comfortable (p=0.0127) after test cream
application.
Questionnairerevealedself-perceivedbenefitsconsistentwithexpertvisualgrading.
Discussion
Thetreatmentofsensitiveskinischallengingandgenerallybasedoncontinuousandtopical
application of anti-sensitive moisturizing tolerance extreme product that improves skin
features associated with itching, stinging, dryness, tightness, burning or pain.10,17–19 To
maintainahealthyskin,itisrecommendedtohydrateandprotectit.Inthisstudy,ananti-
sensitivemoisturizingtolerance-extremecreamfromBeitainiBio-technologicalCo.,Ltd.in
China,wasfoundtobeaneffectivecosmeticcreamtoimproveskinhydrationandtexture.
Itseffectwassimilarasthecontrolcream.Theclinicalassessmentofqualityoffacialskin
usingvisualscoringsystem,theinstrumentalassessmentandtheself-assessmentbysubject
using questionnaire reported that after 28 days of twice daily application of test cream,
dryness,smoothness,roughnessanderythemaofskinweresignificantlyimproved.These
interesting skin benefits are due to the components of the innovative formulation. The
improvement of skin texture is likely contributedbyPrinsepia utilis oil, beta-glucan and
sodiumhyaluronate.
Prinsepiautilis isagenusoftrees intheRosaceaeandmainlydistributedinChina, India,
BangladeshandFarmosa. InChina,Prinsepiautilis is rich in theareaabove2000meters

150
altitudeofYunnanprovincewithpollution-free.Itskerneliscomposedofaround40%oil
whichcontainsamajorityofunsaturatedfattyacidandvitamin.Inpreviousstudy,Prinsepia
utilisoilenhancedthesynthesisofceramideincultureofkeratinocytes.Innudemicemodel,
its topical application during 7 days significantly decreased TEWL, increased epidermal
hydration and skin surface lipids.20,21 Therefore, Prinsepia utilis oil seems to be an ideal
activeingredientforrecoverytheskinbarrierinanti-sensitiveformulation.
Beta-glucansareglucosepolymers,extracted fromcellwallsofnumerous fungal species.
Theyarefoundtocontributetowoundhealing,depressingimmunesystem,andareeven
recommendedfortreatmentofatopicdermatitis.22–25Sodiumhyaluronateisthesodiumsalt
ofhyaluronicacidwhich isacomponentof thedermalextracellularmatrix. Itshydration
propertymakes it an excellentmoisturizing ingredient. ConcerningPortulacaoleracea, it
belongstoPortulacaceaefamilyandisdistributedinmainlywarm-climatearea.Thereare
many studies on its functions (anti-inflammatory,26,27 anti-oxydant,27 anti-ulcerogenic,28
anti-bacterial,29 anti-cancer,30 and inwound healing31,32). In addition, Portulaca oleracea
extracts could prevent the apoptosis of keratinocyte and fibroblast cultures and has an
assistant function in reversing the keratinocyte differentiation and skin barrier
dysfunction.33,34 In this study, theclinical scoringoferythemaafter test creamtreatment
firmlyprovedtheanti-sensitivefunctionofPortulacaoleraceaintheinnovativeformulation.
Conclusion
The combination of different actives in this new anti-sensitive moisturizing tolerance-
extreme cream improved skin hydration and texture after 28 days of treatment. This

151
complexcreamcouldgivefurtherassistancetothecontinuoustreatmentofsensitiveskin.
Acknowledgments
ThisworkwasfinanciallysupportedbyProgramforInnovativeResearchTeaminUniversity
ofMinistryofEducationofChina(GrandNo.IRT13067)andtheFundofYunnanProvince
ChineseAcademyofSciencesCooperation(GrandeNo.2014IB008)(China).
We would like to thank Céline Thiebaut, Aurélie Durai and Vanessa Ecarnot for their technical
help.
Conflictsofinterest
Theauthorshavenoconflictofinteresttodeclare.

152
References
1 Misery L, Sibaud V, Ambronati M, et al. Sensitive scalp: does this condition exist? An
epidemiologicalstudy. Contact Dermatitis. 2008;58:234–238 .
2 Misery, L. Les nerfs a fleur de peau. Int J Cosmet Sci. 2002;24:111–116.
3 Sugiura H, Omoto M, Hirota Y, Danno K, Uehara M. Density and fine structure of
peripheral nerves in various skin lesions of atopic dermatitis. Arch. Dermatol. Res.
1997;289: 125–131.
4 Reilly DM, Parslew R, Sharpe GR, Powell S, Green MR. Inflammatory mediators in
normal, sensitive and diseased skin types. Acta Derm. Venereol. 2000;80: 171–4.
5 Goffin V, Piérard-Franchimont C, Piérard GE. Sensitive skin and stratum
corneumreactivity to household cleaning products. Contact Dermatitis 1996;34: 81–85.
6 Roussaki-Schulze AV, Zafiriou E, Nikoulis D, et al. Objective biophysical findings in
patients with sensitive skin. Drugs Exp. Clin. Res. 2005;31: 17–24.
7 Cua AB, Wilhelm K P, Maibach HI. Cutaneous sodium lauryl sulphate irritation potential:
age and regional variability. Br. J. Dermatol. 1990;123: 607–13.
8 Misery L, Myon E, Martin N, et al. Sensitive skin: psychological effects and seasonal
changes. J. Eur. Acad. Dermatol Venereol. 2007;21: 620-680.
9 Ständer S, Schneider SW, Weishaupt C, Luger TA, Misery L. Putative neuronal
mechanisms of sensitive skin. Exp. Dermatol. 2009;18:417–23.
10 Misery L, Loser K, Ständer S. Sensitive skin. Journal of the European Academy of
Dermatology and Venereology. 2016;30: 2–8.

153
11 Misery L, Boussetta S, Nocera T, Perez-Cullell N, Taieb C. Sensitive skin in Europe. J.
Eur. Acad. Dermatology Venereol. 2009;23: 376–381.
12 PANG Q, He L, etal. Research in anti-inflammation function by various extracted method
and concentration of Portulaca oleracea. Dermatology and Venereology. 2012;34:318-
322.
13 TU Y, Gu H, etal. Effects of Prinsepia utilis Royle oil on the synthesis of ceramide and
expression of ceramidase. Chinese Journal of Dermatology. 2012;45:718-722.
14 Van den Berg LM1, Zijlstra-Willems EM1, Richters CD2, Ulrich MM3, Geijtenbeek TB4.
Dectin-1 activation induces proliferation and migration of human keratinocytes
enhancing wound re-epithelialization. Cell Immunol. 2014;289:49-54.
15 Fanian F, Mac-Mary S, Jeudy A, et al. Efficacy of micronutrient supplementation on skin
aging and seasonal variation: A randomized, placebo-controlled, double-blind study. Clin
Interv Aging. 2013;8:1527-1537.
16 Fotoh C1, Elkhyat A, Mac S, Sainthillier JM, Humbert P. Cutaneous differences between
Black, African or Caribbean Mixed-race and Caucasian women: biometrological approach
of the hydrolipidic film. Skin Res Technol. 2008;14:327-35.
17 Isoda K, Seki T, Inoue Y, et al. Efficacy of the combined use of a facial cleanser and
moisturizers for the care of mild acne patients with sensitive skin. J. Dermatol. 2015;42:
181–188.
18 RichtersR1,FalconeD,UzunbajakavaN,etal.Whatissensitiveskin?Asystematic
literaturereviewofobjectivemeasurements.SkinPharmacologyandPhysiology,
2015;28:75–83.

154
19 Greive K, Heinicke I, Barnes T, Adams D. Evaluation of a topical treatment for the relief
of sensitive skin. Clin. Cosmet. Investig. Dermatol. 2015;8: 405.
20 Hu L, Man H, Elias P. M, Man M-Q. Herbal medicines that benefit epidermal permeability
barrier function. Dermatologica Sin. 2015;33: 90–95.
21 Tu Y, Gu H, Li N, Pang Q, He L. Effects of Prinsepia utilis Royle oil on the synthesis of
ceramide and expression of ceramidase. Chin J Dermatol 2012:718–722.
22 Majtán J, Kumar P, Koller J, Dragúnová J, Gabriz J. Induction of metalloproteinase 9
secretion from human keratinocytes by pleuran (beta-glucan from Pleurotus ostreatus). Z.
Naturforsch. C. 2009;64: 597–600.
23 Bobovcak M, Kuniakova R, Gabriz J, Majtan J. Effect of Pleuran (beta-glucan from
Pleurotus ostreatus) supplementation on cellular immune response after intensive exercise
in elite athletes. Appl. Physiol. Nutr. Metab. Appl. Nutr. Metab. 2010;35: 755–762.
24 Jesenak M, Urbancek S, Majtan J, Banovcin P, Hercogova J. β -Glucan-based cream
(containing pleuran isolated from pleurotus ostreatus ) in supportive treatment of mild-to-
moderate atopic dermatitis. J. Dermatolog. Treat. 2016:27; 351–354.
25 HauCS1,TadaY,ShibataS,etal.HighCalcium,ATP,andPoly(I:C)Augmentthe
ImmuneResponsetoβ-GlucaninNormalHumanEpidermalKeratinocytes.J.Invest.
Dermatol.2011;131:2255–2262.
26 LeeAS,KimJS,LeeYJ,KangDG,LeeHS.Anti-TNF-αactivityofPortulacaoleraceain
vascularendothelialcells.Int.J.Mol.Sci.2013;13;5628–5644.
27 YangX,YanY,LiJ,TangZetal.ProtectiveeffectsofethanolextractfromPortulaca
oleraceaLondextransulphatesodium-inducedmiceulcerativecolitisinvolving
anti-inflammatoryandantioxidant.Am.J.Transl.Res.2016;8;2138–2148.

155
28 Karimi G, Hosseinzadeh H, Ettehad N. Evaluation of the gastric antiulcerogenic effects of
Portulaca oleracea L. extracts in mice. Phyther. Res. 2014;18: 484–487.
29 Lei X, Li J, Liu B, Zhang N, Liu H. Separation and Identification of Four New Compounds
with Antibacterial Activity from Portulaca oleracea L. Molecules 2015;20: 16375–87.
30 Al-SheddiES,FarshoriNN,Al-OqailMM,MusarratJ,Al-KhedhairyAA,Siddiqui
MA.PortulacaoleraceaSeedOilExertsCytotoxicEffectsonHumanLiverCancer
(HepG2)andHumanLungCancer(A-549)CellLines.AsianPac.J.CancerPrev.
2015;16:3383–3387.
31 Rashed, A. Simple evaluation of the wound healing activity of a crude extract of Portulaca
oleracea L. (growing in Jordan) in Mus musculus JVI-1. J. Ethnopharmacol. 2003;88: 131–
136.
32 ZhouYX,XinHL,RahmanK,WangSJ,PengC,ZhangH.PortulacaoleraceaL.:Areview
ofphytochemistryandpharmacologicaleffects.BiomedRes.Int.2015:925631.
33 Lee S, Kim K, Park C, Lee J-S, Kim Y-H. Portulaca oleracea extracts protect human
keratinocytes and fibroblasts from UV-induced apoptosis. Exp. Dermatol. 2014;23: 13–17.
34 Zhao H, Li S, Luo F et al. Portulaca oleracea L. aids calcipotriol in reversing keratinocyte
differentiation and skin barrier dysfunction in psoriasis through inhibition of the nuclear
factor kappa B signaling pathway. Exp. Ther. Med. 2015;9: 303–310.

156
Table 1. List of ingredients of test cream
Test cream: Anti-sensitive moisturizing tolerance-extreme cream
Aqua, Glycerin, Pentylene glycol, butyrospermum parkII (shea butter), tridecyl,
trimellitate, dimethicone, glyceryl stearate, prinsepia utilis oil, sodium hyaluronate,
portulaca oleracea extract, beta-glucan, acrylates/c10-30 alkyl acrylate crosspolumer,
xanthan gum, butylene glycol, aminomethyl propano
Control cream: Tolerance extreme cream
Thermal spring water, glycerin, mineral oil (paraffinum liquidum), squalene, carthamus
tinctorius (Safflower), seed oil (carthamus tinctorius seed oil), cyclomethicone, glyceryl
stearate, sodium carbomer, titanium dioxide

157
Table 2. Self-assessment questionnaire designed for subjects
Does your skin reddens easily? (0 = never / 9 = very easy)
What is the intensity of your rash? (0 = no redness / 9 = very intense redness)
Is your skin reacts with emotions, stress? (0 = never / 9 = very often)
Is your skin reacts it with changes in temperature? (0 = never / 9 = very often)
Is your skin does presents persistent redness? (0 = never / 9 = very often)
Do you feel tingling? (0 = never / 9 = very often)
Do you feel tightness? (0 = never / 9 = very often)
Do you feel itchy? (0 = never / 9 = very often)
Do you feel your skin hydrated? (0 = not hydrated, dry skin / 9 = very hydrated)
Have you got desquamation? (0 = no desquamation / 9 = significant desquamation)
Do you feel the skin of your face comfortable? (0 = Very Comfortable / 9 = Uncomfortable)
Is your skin looks rough? (0 = no rough / 9 = very rough)
Is your skin soft? (0 = no soft / 9 = very soft)
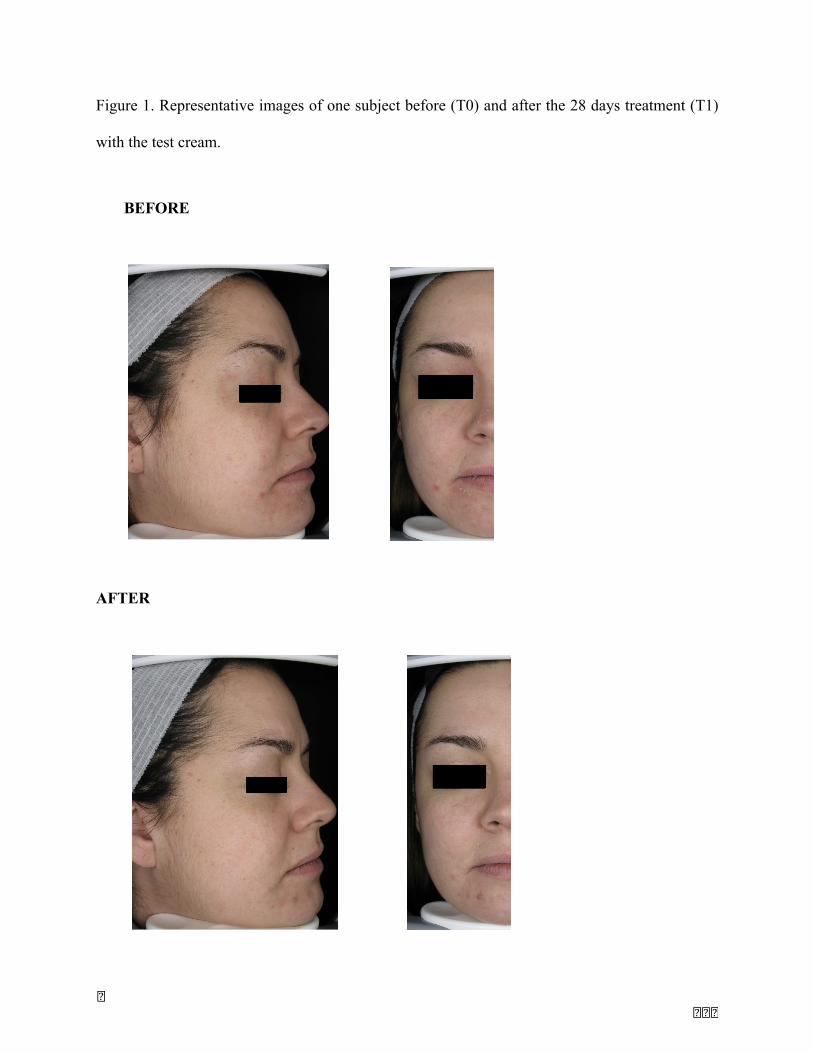
158
Figure 1. Representative images of one subject before (T0) and after the 28 days treatment (T1)
with the test cream.
BEFORE
AFTER
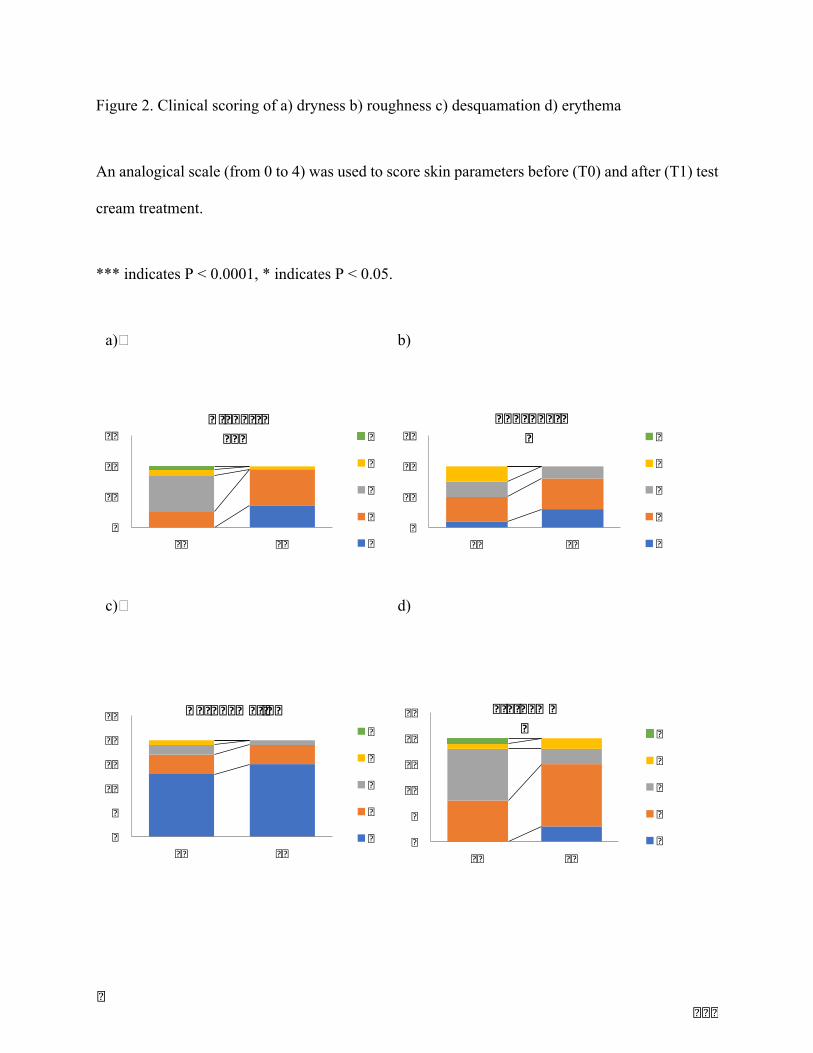
159
Figure 2. Clinical scoring of a) dryness b) roughness c) desquamation d) erythema
An analogical scale (from 0 to 4) was used to score skin parameters before (T0) and after (T1) test
cream treatment.
*** indicates P < 0.0001, * indicates P < 0.05.
a) b)
c) d)
0
10
20
30
T0 T1
Dryness*** 4
3
2
1
00
10
20
30
T0 T1
Roughness* 4
3
2
1
0
0
5
10
15
20
25
T0 T1
Desquamation4
3
2
1
0 0
5
10
15
20
25
T0 T1
Erythema* 4
3
2
1
0
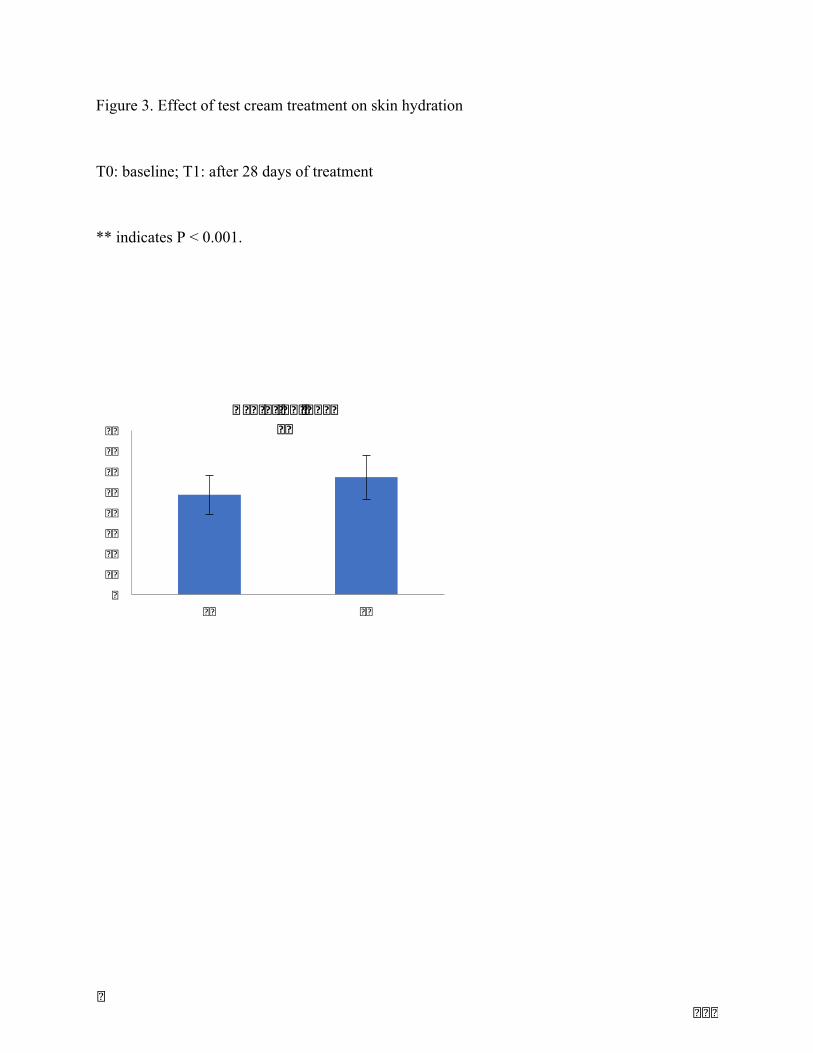
160
Figure 3. Effect of test cream treatment on skin hydration
T0: baseline; T1: after 28 days of treatment
** indicates P < 0.001.
01020304050607080
T0 T1
Hydrationindex**
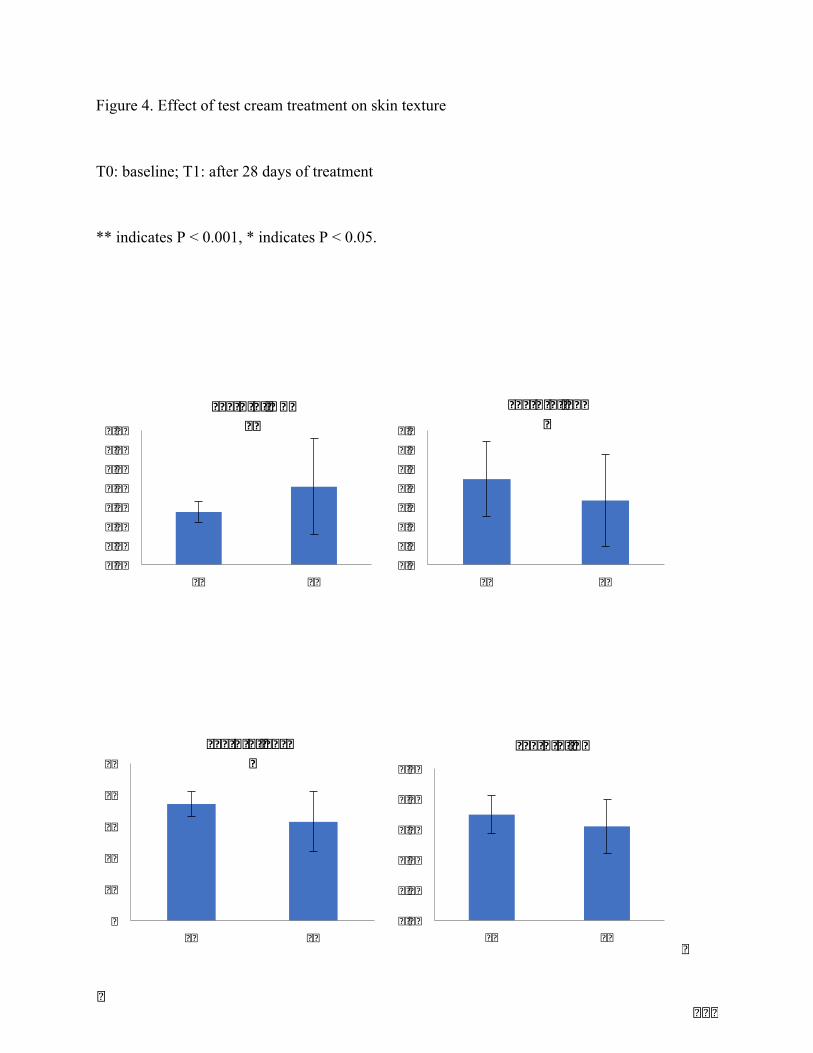
161
Figure 4. Effect of test cream treatment on skin texture
T0: baseline; T1: after 28 days of treatment
** indicates P < 0.001, * indicates P < 0.05.
0.000.020.040.060.080.100.120.14
T0 T1
Texture:NRJ**
0.00.51.01.52.02.53.03.5
T0 T1
Texture:Ser*
0
10
20
30
40
50
T0 T1
Texture:Sesm*
0.00
0.02
0.04
0.06
0.08
0.10
T0 T1
Texture:R5
162
Figure 5. Significant results of self-assessment questionnaire
T0: baseline; T1: after 28 days of treatment
*** indicates P < 0.0001, ** indicates P < 0.001.
0
2
4
6
8
T0 T1
Do you feel your skin hydrated?
***
0
2
4
6
8
T0 T1
Do you feel the skin of your face comfortable?
**
02468
10
T0 T1
Do you feel tightness?
**

163
References
1. Bilousova, G. et al. Melanocytes and Their Diseases. (2014).doi:10.1101/cshperspect.a017046
2. Yamaguchi, Y. et al. Mesenchymal-epithelial interactions in the skin: Increasedexpressionofdickkopf1bypalmoplantarfibroblastsinhibitsmelanocytegrowthanddifferentiation.J.CellBiol.165,275–285(2004).
3. Bennett, D. C. Human melanocyte senescence and melanoma susceptibility genes.3063–3069(2003).doi:10.1038/sj.onc.1206446
4. Lin, J. Y. & Fisher, D. E. Melanocyte biology and skin pigmentation.445, 843–850(2007).
5. Yamaguchi,Y.&Hearing,V. J.Physiological factorsthatregulateskinpigmentation.BioFactors35,193–199(2009).
6. Yamaguchi,Y.,Brenner,M.&Hearing,V.J.Theregulationofskinpigmentation.JournalofBiologicalChemistry(2007).doi:10.1074/jbc.R700026200
7. Tomihari, M., Hwang, S.-H., Chung, J.-S., Cruz Jr., P. D. & Ariizumi, K. Gpnmb is amelanosome-associated glycoprotein that contributes to melanocyte/keratinocyteadhesioninaRGD-dependentfashion.Exp.Dermatol.18,586–595(2009).
8. Hoashi,T.etal.MART-1isrequiredforthefunctionofthemelanosomalmatrixproteinPMEL17/GP100and thematurationofmelanosomes. J.Biol. Chem.280, 14006–16(2005).
9. Yamaguchi,Y.&Hearing,V.J.MelanocytesandTheirDiseases.1–18(2014).
10. InamdarGS1,MadhunapantulaSV,R.G.TargetingtheMAPKpathwayinmelanoma:whysomeapproachessucceedandotherfail.BiochemPharmacol80,624–637(2011).
11. Halaban,R.TheRegulationofNormalMelanocyteProliferation.PigmentCellRes.13,4–14(2000).
12. D’Mello,S.A.N.,Finlay,G.J.,Baguley,B.C.&Askarian-Amiri,M.E.SignalingPathwaysinMelanogenesis.Int.J.Mol.Sci.17,(2016).
13. Lim,X.&Nusse,R.Wntsignalinginskindevelopment,homeostasis,anddisease.Cold

164
SpringHarb.Perspect.Biol.5,a008029(2013).
14. Bergmann,C.&Distler, J.H.W.CanonicalWnt signaling in systemic sclerosis.Lab.Investig.96,151–155(2016).
15. Vibert,L.etal.AnongoingroleforWntsignalingindifferentiatingmelanocytesinvivo.PigmentCellMelanomaRes.30,219–232(2017).
16. Fedon,Y.etal.inSkeletalMuscle-FromMyogenesistoClinicalRelations(InTech,2012).doi:10.5772/48330
17. Park,H.etal.MITFmediatescAMP-inducedproteinkinaseC-βexpressioninhumanmelanocytes.Biochem.J.578,571–578(2006).
18. Vredeveld, L. C. W. et al. Abrogation of BRAFV600E-induced senescence by PI3Kpathway activation contributes to melanomagenesis. Genes Dev. 26, 1055–1069(2012).
19. Blume-Jensen,P.,Janknecht,R.&Hunter,T.Thekitreceptorpromotescellsurvivalviaactivation of PI 3-kinase and subsequent Akt-mediated phosphorylation of Bad onSer136.Curr.Biol.8,779–782(1998).
20. Datta, S. R. et al. Akt phosphorylation of BAD couples survival signals to the cell-intrinsicdeathmachinery.Cell91,231–241(1997).
21. Cardinali,G.etal.Melanosometransferpromotedbykeratinocytegrowth factor inlightanddarkskin-derivedkeratinocytes.J.Invest.Dermatol.128,558–67(2008).
22. Seiberg, M. et al. The protease-activated receptor 2 regulates pigmentation viakeratinocyte-melanocyteinteractions.Exp.CellRes.254,25–32(2000).
23. Babiarz-Magee,L.,Chen,N.,Seiberg,M.&Lin,C.B.Theexpressionandactivationofprotease-activated receptor-2 correlate with skin color. Pigment Cell Res. (2004).doi:10.1111/j.1600-0749.2004.00133.x
24. Giehl,K.A.,Nägele,U.,Volkenandt,M.&Berking,C.Proteinexpressionofmelanocytegrowthfactors(bFGF,SCF)andtheirreceptors(FGFR-1,c-kit)inneviandmelanoma.J.Cutan.Pathol.34,7–14(2007).
25. Wolnicka-glubisz,A.,Pecio,A.,Podkowa,D.&Plonka,P.M.HGF/SFIncreasesNumberof Skin Melanocytes but Does Not Alter Quality or Quantity of FollicularMelanogenesis.8,1–11(2013).

165
26. Belleudi, F., Cardinali, G., Kovacs, D., Picardo, M. & Torrisi, M. R. KGF PromotesParacrineActivationoftheSCF/c-KITAxisfromHumanKeratinocytestoMelanomaCells.Transl.Oncol.3,80–90(2010).
27. Choi,W.etal.Thefibroblast-derivedparacrinefactorneuregulin-1hasanovelroleinregulating theconstitutivecolorandmelanocyte function inhumanskin. J.Cell Sci.123,3102–3111(2010).
28. Esper, R. M., Pankonin, M. S. & Loeb, J. A. Neuregulins: Versatile growth anddifferentiation factors in nervous system development and human disease. BrainResearchReviews51,161–175(2006).
29. Innominato,P.F.,Libbrecht,L.&vandenOord,J.J.Expressionofneurotrophinsandtheirreceptorsinpigmentcelllesionsoftheskin.J.Pathol.194,95–100(2001).
30. Scott, G. a,McClelland, L. a& Fricke,A. F. Semaphorin7apromotes spreading anddendricity inhumanmelanocytes throughbeta1-integrins. J. Invest.Dermatol.128,151–161(2008).
31. Yamaguchi,Y.etal.Dickkopf1(DKK1)regulatesskinpigmentationandthicknessbyaffectingWnt/beta-cateninsignalinginkeratinocytes.FASEBJ.22,1009–1020(2008).
32. Kaur,A.etal.sFRP2intheagedmicroenvironmentdrivesmelanomametastasisandtherapyresistance.Nature532,250–4(2016).
33. Kim,M.,Han,J.H.,Kim,J.-H.,Park,T.J.&Kang,H.Y.SecretedFrizzled-RelatedProtein2(sFRP2)FunctionsasaMelanogenicStimulator;theRoleofsFRP2inUV-InducedHyperpigmentaryDisorders.J.Invest.Dermatol.136,236–44(2016).
34. Aoki,H.,Moro,O.,Tagami,H.&Kishimoto,J.Geneexpressionprofilinganalysisofsolarlentigoinrelationtoimmunohistochemicalcharacteristics.Br.J.Dermatol.156,1214–1223(2007).
35. Kovacs, D. et al. Role of fibroblast-derived growth factors in regulatinghyperpigmentationofsolarlentigo.Br.J.Dermatol.163,1020–1027(2010).
36. Bastonini,E.,Kovacs,D.&Picardo,M.Skinpigmentationandpigmentarydisorders:Focusonepidermal/dermalcross-talk.AnnalsofDermatology28,279–289(2016).
37. Chen,N.et al. The roleof keratinocyte growth factor inmelanogenesis:Apossiblemechanismfortheinitiationofsolarlentigines.Exp.Dermatol.19,865–872(2010).

166
38. Niehrs, C. Function and biological roles of theDickkopf family ofWntmodulators.7469–7481(2006).doi:10.1038/sj.onc.1210054
39. Yamaguchi,Y.etal.Theeffectsofdickkopf1ongeneexpressionandWntsignalingbymelanocytes: mechanisms underlying its suppression of melanocyte function andproliferation.J.Invest.Dermatol.127,1217–25(2007).
40. Park,H.etal.DistinctrolesofDKK1andDKK2intumorangiogenesis.Angiogenesis17,221–234(2014).
41. Li,Y.etal.DKK3regulatescellproliferation,apoptosisandcollagensynthesisinkeloidfibroblastsviaTGF-b1/Smadsignalingpathway.Biomed.Pharmacother.91,174–180(2017).
42. Rachner, T. D. et al. P38 regulates theWnt inhibitor Dickkopf-1 in breast cancer.Biochem.Biophys.Res.Commun.466,728–732(2015).
43. Hartman,M.L.etal.Geneexpressionprofilingidentifiesmicrophthalmia-associatedtranscription factor (MITF) and dickkopf-1 (DKK1) as regulators ofmicroenvironment-driven alterations in melanoma phenotype. PLoS One (2014).doi:10.1371/journal.pone.0095157
44. Yamaguchi,Y.,Morita,A.,Maeda,A.&Hearing,V.J.RegulationofSkinPigmentationandThicknessbyDickkopf1(DKK1).J.Investig.DermatologySymp.Proc.14,73–75(2009).
45. Yamaguchi,Y.etal.TheEffectsofDickkopf1onGeneExpressionandWntSignalingbyMelanocytes :MechanismsUnderlyingItsSuppressionofMelanocyteFunctionandProliferation.J.Invest.Dermatol.127,1217–1225(2007).
46. Hirobe,T.,Hasegawa,K.,Furuya,R.,Fujiwara,R.&Sato,K.Effectsoffibroblast-derivedfactors on theproliferation anddifferentiation of humanmelanocytes in culture. J.Dermatol.Sci.71,45–57(2013).
47. Chen, J. et al. LRP6 dimerization through its LDLR domain is required for robustcanonicalWntpathwayactivation.26,1068–1074(2014).
48. Embryology, M., Krebsforschungszentrum, D. & Niehrs, C. LDL-receptor-relatedprotein6isareceptorforDickkopfproteins.1,321–325(2001).
49. Sarkar,D.etal.mda-7(IL-24)mediatesselectiveapoptosisinhumanmelanomacells

167
byinducingthecoordinatedoverexpressionoftheGADDfamilyofgenesbymeansofp38MAPK.99,1–6(2002).
50. Gilmour,J.etal.AcrucialrolefortheubiquitouslyexpressedtranscriptionfactorSp1atearlystagesofhematopoieticspecification.Development141,2391–2401(2014).
51. Moretti,S.etal.NewInsightsintothePathogenesisofVitiligo :ImbalanceofEpidermalCytokinesatSitesofLesions.87–92(2002).
52. Grimes,P.E.Newinsightsandnewtherapiesinvitiligo.Jama293,730(2005).
53. Regazzetti,C.etal.TranscriptionalAnalysisofVitiligoSkinRevealstheAlterationofWNT Pathway: A Promising Target for Repigmenting Vitiligo Patients. J. Invest.Dermatol.(2015).doi:10.1038/jid.2015.335
54. Oh, S. H. et al. DKK1 is highly expressed in the dermis of vitiligo lesion: Is thereassociationbetweenDKK1andvitiligo?JournalofDermatologicalScience66,163–165(2012).
55. Cario-André,M.,Pain,C.,Gauthier,Y.,Casoli,V.&Taieb,A.Invivoandinvitroevidenceofdermalfibroblastsinfluenceonhumanepidermalpigmentation.PigmentCellRes.(2006).doi:10.1111/j.1600-0749.2006.00326.x
56. Kitisin,K.etal.Tgf-Betasignalingindevelopment.Sci.STKE2007,cm1(2007).
57. Kuphal, S. et al. Expression of Dickkopf genes is strongly reduced in malignantmelanoma.Oncogene25,5027–5036(2006).
58. Kitisin,K.etal.TGF-betaSignalinginDevelopment.Sci.STKE2007,cm1-cm1(2007).
59. Wang, X. J., Han, G., Owens, P., Siddiqui, Y. & Li, A. G. Role of TGF beta-mediatedinflammation in cutaneous wound healing. J.Investig.Dermatol.Symp.Proc. 11, 112–117(2006).
60. Verrecchia, F.&Mauviel,A.Transforming growth factor-beta and fibrosis.World J.Gastroenterol.13,3056–62(2007).
61. Soma,Y.&Grotendorst,G.R.TGF?????stimulatesprimaryhumanskinfibroblastDNAsynthesisviaanautocrineproductionofPDGF???relatedpeptides.J.Cell.Physiol.140,246–253(1989).
62. Samuel,G.H.,Bujor,A.M.,Nakerakanti,S.S.,Hant,F.N.&Trojanowska,M.Autocrine

168
transforminggrowthfactorβsignalingregulatesextracellularsignal-regulatedkinase1/2 phosphorylation via modulation of protein phosphatase 2A expression insclerodermafibroblasts.FibrogenesisTissueRepair3,25(2010).
63. Kopp,J.etal.AbrogationofTransformingGrowthFactor-SignalingbySMAD7InhibitsCollagenGel Contraction ofHumanDermal Fibroblasts. J. Biol. Chem.280, 21570–21576(2005).
64. Wang,R.etal.Hypertrophicscartissuesandfibroblastsproducemoretransforminggrowth factor-beta1mRNA and protein than normal skin and cells.Wound RepairRegen.8,128–37
65. Rolfe,K.J., Irvine,L.M.,Grobbelaar,A.O.&Linge,C.Differentialgeneexpressioninresponsetotransforminggrowthfactor-?1byfetalandpostnataldermalfibroblasts.WoundRepairRegen.15,897–906(2007).
66. Ravanti,L.etal.Transforminggrowthfactor-betainducescollagenase-3expressionbyhuman gingival fibroblasts via p38mitogen-activated protein kinase. J. Biol. Chem.274,37292–300(1999).
67. Penn,J.W.,Grobbelaar,A.O.&Rolfe,K.J.TheroleoftheTGF-βfamilyinwoundhealing,burnsandscarring:areview.Int.J.BurnsTrauma2,18–28(2012).
68. Akhmetshina,A.etal.Activationof canonicalWntsignalling is required forTGF-β-mediatedfibrosis.Nat.Commun.3,735(2012).
69. Tang, J. & Li, Q. Primary culture of human face skinmelanocytes for the study ofhyperpigmentation.891–898(2014).doi:10.1007/s10616-013-9643-6
70. Dawson-baglien,E.M.,Winkler,P.A.,Bruewer,A.R.,Petersen-jones,S.M.&Bartoe,J.T. Isolation and cultivation of canine uveal melanocytes. 1–6 (2014).doi:10.1111/vop.12208
71. Hsu,M.,Li,L.&Herlyn,M.CultivationofNormalHumanEpidermalMelanocytesintheAbsenceofPhorbolEsters.107,(1982).
72. Granger, C. et al. Biological processes in solar lentigo : insights brought byexperimentalmodels.174–177(2016).doi:10.1111/exd.12937
73. Cornali.A.G.,Bardawill,C.J.,andDavid,M.Determinationofserumproteinbymeansofthebiuretreaction.J.Bid.Chem.751–766(1946).

169
74. Placet,V.etal.Developmentandcharacterizationofahumandermalequivalentwithphysiological mechanical properties ´. 1–8 (2011). doi:10.1111/j.1600-0846.2011.00564.x
75. Bride,J.&Viennet,C.Indicationoffibroblastapoptosisduringthematurationofdisc-shaped mechanically stressed collagen lattices. 312–317 (2004).doi:10.1007/s00403-003-0438-z
76. Goorochurn,R.etal.DifferentialmorphologicalandfunctionalfeaturesoffibroblastsexplantedfromSolarLentigo.Br.J.Dermatol.(2017).doi:10.1111/bjd.15386
77. Yamaguchi,Y.,Morita,A.,Maeda,A.&Hearing,V.J.Regulationofskinpigmentationandthicknessbydickkopf1(DKK1).doi:10.1038/jidsymp.2009.4
78. Yamaguchi,Y.etal.Mesenchymal–epithelialinteractionsintheskin.J.CellBiol.165,275–285(2004).
79. Misery,L.etal.Sensitivescalp:doesthisconditionexist?Anepidemiologicalstudy.ContactDermatitis58,234–238(2008).
80. Misery,L.etal.Sensitivescalp:Doesthisconditionexist?Anepidemiologicalstudy.ContactDermatitis58,234–238(2008).
81. Misery,L.,Loser,K.&Ständer,S.Sensitiveskin.JournaloftheEuropeanAcademyofDermatologyandVenereology30,2–8(2016).
82. Isoda,K.etal.Efficacyofthecombineduseofafacialcleanserandmoisturizersforthecareofmildacnepatientswithsensitiveskin.J.Dermatol.42,181–188(2015).
83. Richters,R.etal.Whatissensitiveskin?Asystematic literaturereviewofobjectivemeasurements.SkinPharmacologyandPhysiology28,75–83(2015).
84. Greive,K.,Heinicke,I.,Barnes,T.&Adams,D.Evaluationofatopicaltreatmentforthereliefofsensitiveskin.Clin.Cosmet.Investig.Dermatol.8,405(2015).
85. Hu, L., Man, H., Elias, P. M. &Man, M.-Q. Herbalmedicines that benefit epidermalpermeabilitybarrierfunction.DermatologicaSin.33,90–95(2015).
86. TuY,GuH,LiN,PangQ,H.L.EffectsofPrinsepiautilisRoyleoilonthesynthesisofceramideandexpressionofceramidase.ChinJDermatol718–722(2012).
87. Majtán,J.,Kumar,P.,Koller,J.,Dragúnová,J.&Gabriz,J.Inductionofmetalloproteinase

170
9 secretion from human keratinocytes by pleuran (beta-glucan from Pleurotusostreatus).Z.Naturforsch.C.64,597–600(2009).
88. Bobovcak,M.,Kuniakova,R.,Gabriz,J.&Majtan,J.EffectofPleuran(beta-glucanfromPleurotus ostreatus) supplementation on cellular immune response after intensiveexercise ineliteathletes.Appl.Physiol.Nutr.Metab.Appl.Nutr.Metab.35,755–762(2010).
89. Jesenak,M.,Urbancek,S.,Majtan,J.,Banovcin,P.&Hercogova,J.β-Glucan-basedcream(containing pleuran isolated from pleurotus ostreatus ) in supportive treatment ofmild-to-moderateatopicdermatitis.J.Dermatolog.Treat.27,351–354(2016).
90. Hau,C.S.etal.HighCalcium,ATP,andPoly(I:C)AugmenttheImmuneResponsetoβ-Glucan inNormalHumanEpidermalKeratinocytes. J. Invest. Dermatol.131, 2255–2262(2011).
91. Lee,A.S.,Kim,J.S.,Lee,Y.J.,Kang,D.G.&Lee,H.S.Anti-TNF-αactivityofPortulacaoleraceainvascularendothelialcells.Int.J.Mol.Sci.13,5628–5644(2012).
92. Yang,X.etal.ProtectiveeffectsofethanolextractfromPortulacaoleraceaLondextransulphate sodium-induced mice ulcerative colitis involving anti-inflammatory andantioxidant.Am.J.Transl.Res.8,2138–2148(2016).
93. Karimi,G.,Hosseinzadeh,H.&Ettehad,N.Evaluationof thegastric antiulcerogeniceffectsofPortulacaoleraceaL.extractsinmice.Phyther.Res.18,484–487(2004).
94. Lei, X., Li, J., Liu, B., Zhang, N. & Liu, H. Separation and Identification of FourNewCompounds with Antibacterial Activity from Portulaca oleracea L. Molecules 20,16375–87(2015).
95. Rashed, A. Simple evaluation of the wound healing activity of a crude extract ofPortulacaoleraceaL.(growinginJordan)inMusmusculusJVI-1.J.Ethnopharmacol.88,131–136(2003).
96. Zhou, Y. X. et al. Portulaca oleracea L.: A review of phytochemistry andpharmacologicaleffects.BiomedRes.Int.2015,(2015).
97. Lee,S.,Kim,K.H.,Park,C.,Lee,J.-S.&Kim,Y.H.PortulacaoleraceaextractsprotecthumankeratinocytesandfibroblastsfromUV-inducedapoptosis.Exp.Dermatol.23,13–17(2014).

171
98. Zhao, H. et al. Portulaca oleracea L. aids calcipotriol in reversing keratinocytedifferentiation and skin barrier dysfunction in psoriasis through inhibition of thenuclearfactorkappaBsignalingpathway.Exp.Ther.Med.9,303–310(2015).
99. Raposo,G.&Marks,M.S.Thedarksideoflysosome-relatedorganelles:specializationoftheendocyticpathwayformelanosomebiogenesis.Traffic3,237–248(2002).
100. Gillbro, J.M.&Olsson,M. J. Themelanogenesis andmechanismsof skin-lighteningagents – existing and new approaches. 210–221 (2011). doi:10.1111/j.1468-2494.2010.00616.x