activation-induced cell death presented by exosomes delays t cell a membrane form of tnf{alpha}
TRANSCRIPT

of September 11, 2015.This information is current as
Cell DeathExosomes Delays T Cell Activation-Induced
Presented byαA Membrane Form of TNF-
William Grizzle and Robert P. KimberlyLiming Zhang, Shuangqin Zhang, Jianhua Wang, Xu Cao, Huang-Ge Zhang, Cunren Liu, Kaihun Su, Shaohua Yu,
http://www.jimmunol.org/content/176/12/7385doi: 10.4049/jimmunol.176.12.7385
2006; 176:7385-7393; ;J Immunol
Referenceshttp://www.jimmunol.org/content/176/12/7385.full#ref-list-1
, 19 of which you can access for free at: cites 64 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Errata
http://www.jimmunol.org/content/177/3/2025.2.full.pdfor:
next pageAn erratum has been published regarding this article. Please see
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2006 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on September 11, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on September 11, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on September 11, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on September 11, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

A Membrane Form of TNF-� Presented by Exosomes Delays TCell Activation-Induced Cell Death1
Huang-Ge Zhang,2*† Cunren Liu,* Kaihun Su,* Shaohua Yu,* Liming Zhang,*Shuangqin Zhang,* Jianhua Wang,* Xu Cao,‡ William Grizzle,‡ and Robert P. Kimberly2*
In common with many other cell types, synovial fibroblasts produce exosomes. In this study, we show that the exosomes producedby synovial fibroblasts obtained from individuals with rheumatoid arthritis (RASF), but not exosomes produced by synovialfibroblasts obtained from individuals with osteoarthritis, contain a membrane bound form of TNF-� as demonstrated by colloidalgold immunostaining of TNF-� and confirmed by both Western blot and mass spectrometry. The RASF-derived exosomes, but notexosomes derived from fibroblasts obtained from individuals with osteoarthritis, are cytotoxic for the L929 cell, a TNF-�-sensitivecell line, and stimulate activation of NF-�B and induction of collagenase-1 in RASF. These effects are blocked by addition ofsoluble TNFR1 (sTNFbp), suggesting that a TNF-�-signaling pathway mediates these biological activities. sTNFbp also reducedthe production of exosomes by RASF, suggesting the interruption of a positive amplification loop. Exosomes can transmit signalsbetween cells, and RASF exosomes, effectively taken up by anti-CD3-activated T cells, activated AKT and NF-�B and renderedthese activated T cells resistant to apoptosis. Neutralization of exosomal membrane TNF-� by sTNFbp partially reversed thisresistance, suggesting that not only TNF-� but also additional exosomal proteins may contribute to the development of apoptosisresistance. The Journal of Immunology, 2006, 176: 7385–7393.
E xosomes are formed by reverse budding of the membraneof late endosomes or multivesicular bodies (MVB)3 andare released into the extracellular space by fusion of MVB
with the plasma membrane. Although the function of exosomes isstill poorly understood, recent studies on the fate and functions ofexosomes, mostly in the immune system, have led to the hypoth-esis that exosomes represent a refined intercellular exchange de-vice allowing transfer of proteins and lipids between cells (1). Forexample, exosomes produced by IL-10-treated dendritic cells caneffectively prevent collagen II-induced arthritis in both rabbit andmouse models (2). Salivary gland epithelial cell exosomes packedwith a variety of autoantigens might have Ag-presenting functioncapable of stimulating either induction of T cell tolerance or anautoimmune response (3). Indeed, exosomes have been shown tobe a source of Ag for APCs and to participate in Ag presentationto T lymphocytes (4–6). The presence of biologically active mol-
ecules, such as TNF-�, within the exosomes might also affect theimmune response, and recently, a number of TNF family proteinshave been identified on exosomes derived from dendritic cells (7).
It is well recognized that TNF-� contributes to its pathogenesisof rheumatoid arthritis (RA), a chronic inflammatory disease char-acterized by destructive polyarthritis. Anti-TNF therapy for RAhas proven remarkably effective in decreasing inflammation, im-proving patient function, and attenuating cartilage and bone ero-sions (8–10). In virtually all animal models of arthritis (e.g., ad-juvant-induced, collagen-induced, and serum-induced arthritis),the symptoms are ameliorated by interference with the TNF-�-signaling pathways through the use of anti-TNF-� Abs, solubleTNFR fusion proteins or genetic manipulation (10, 11).
TNF occurs in two forms: a type II membrane protein (mTNF)and a soluble form (sTNF) that is derived from the membrane formby proteolytic cleavage. TNF is initially expressed as the 26-kDatransmembrane protein, and after cleavage at the membrane byTNF-�-converting enzyme, the 17-kDa soluble protein is released.The relative roles of mTNF and sTNF in terms of function havenot been determined, in large part due to the absence of physio-logically relevant models that enable the two forms of TNF to becompared. To date, the main evidence for an in vivo role formTNF has come from transgenic mice, in which noncleavableTNF is overexpressed in various organs (12). These studies havedemonstrated that mTNF plays a role in tissue destruction andautoimmune inflammation.
CD4� T cells are a major cell population in the inflammatoryinfiltrate in the joints of RA patients, and their topographic distri-bution in some of the histologically most typical cases is consistentwith an involvement in a germinal center reaction comparable tothe Ag-specific immune responses in secondary lymphoid organs(13, 14). However, the survival signals that protect the infiltratingCD4� T cells from activation-induced cell death (AICD) are stillnot identified.
In this study, we demonstrate that exosomes produced by syno-vial fibroblasts obtained from individuals with rheumatoid arthritis
*Division of Clinical Immunology and Rheumatology, Department of Medicine, Uni-versity of Alabama at Birmingham, Birmingham, AL 35294; †Birmingham VeteransAdministration Medical Center, Birmingham, AL 35233; and ‡Department of Pathol-ogy, University of Alabama at Birmingham, Birmingham, AL 35294
Received for publication August 30, 2005. Accepted for publication March 28, 2006.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported in part by grants from the National Institutes of Health(P30 AR48311, RO1 CA116092, RO1 CA107181), and by a Birmingham VeteransAffairs Medical Center Merit Review Grant (to H.-G.Z.). H.-G.Z. is a recipient of anArthritis Foundation Investigator Award.2 Address correspondence and reprint requests to Dr. Huang-Ge Zhang, University ofAlabama at Birmingham, 701 19th Street South, Lyons-Harrison Research Building,Room 473, Birmingham, AL 35294-0007. E-mail address: [email protected] or Dr. Robert P. Kimberly, University of Alabama at Birmingham, 1900University Avenue, Tinsley Harrison Tower, Room 429, Birmingham, AL 35294-0006. E-mail address: [email protected] Abbreviations used in this paper: MVB, multivesicular body; RA, rheumatoid ar-thritis; mTNF, membrane TNF; sTNF, soluble TNF; AICD, activation-induced celldeath; RASF, synovial fibroblasts obtained from individuals with RA; sTNFbp, sol-uble TNFR1; OA, osteoarthritis; OASF, synovial fibroblasts obtained from individ-uals with OA; MS, mass spectrometry; MMP-1, matrix metalloproteinase 1; PI, pro-pidium iodide.
The Journal of Immunology
Copyright © 2006 by The American Association of Immunologists, Inc. 0022-1767/06/$02.00
by guest on September 11, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

(RASF) contain a membrane-bound form of TNF-� that is cyto-toxic for L929 target cells, an effect eliminated by treatment witha soluble TNFR1 (sTNFbp). CD4 T cells cocultured with RASFexosomes, develop apoptosis resistance, and blockade of exosomalTNF-� led to partial restoration of CD4 T cell AICD. Taken to-gether, these observations suggest that RA synovial fibroblastsmay communicate with infiltrating CD4 T cells through RASFexosomes in the joints and such communication may lead to blunt-ing of AICD in these infiltrating T cells. The membrane-boundform of TNF-� on the RASF exosomes may mediate these effectson T cell AICD at least in part by sustaining the activities of bothAkt and NF-�B in activated T cells, thus blocking some effects ofcaspase activation.
Materials and MethodsCell culture
Primary synovial cell lines were established as described previously (15).In brief, small pieces of synovial tissue were digested with 1 mg/ml typeI collagenase (Sigma-Aldrich). After dissociation of the fibroblasts, thedigested cells were centrifuged at 1000 revolutions per minute for 5 minand plated in a 25-cm2 flask in 8 ml of DMEM supplemented with 10%FBS. The cells were cultured to 80% confluence before they were used inthe experiments. All experiments were performed using primary synovialcell lines. All of the synovial fibroblast cultures were prepared from freshsynovial tissue biopsy samples obtained through the Tissue ProcurementService of the University of Alabama at Birmingham (UAB) from patientswith RA or osteoarthritis (OA) obtained at the time of total joint replace-ment according to protocols approved by the Institutional Review Board.The diagnosis of RA satisfied the clinical criteria of the American Collegeof Rheumatology. All patients were female and between the ages of 45 and73 years; the 10 patients with RA were age matched with 10 patientswith OA.
The L929 cell line was maintained in DMEM supplemented with 5%FCS at 37°C in a 5% CO2 incubator. For most assays, the cells were platedat 4 � 104 cells/ml for 24 h. After washing with PBS, the cells wereresuspended in DMEM containing 2% FCS and returned to standard in-cubator conditions for 2 h before treatment.
Isolation of RASF and synovial fibroblasts obtained fromindividuals with OA (OASF) exosomes
RASF or OASF cells were grown to 80% confluence in DMEM supple-mented with 10% FCS, washed with a large volume PBS (20 ml/10-cmdish), and cultured with sera-free CD293 medium (Invitrogen Life Tech-nologies) for 48 h before the supernatants were harvested.
Exosomes were collected from the medium by differential centrifuga-tion. In brief, cells were removed by centrifugation for 10 min at 200 � g.The supernatants were collected and centrifuged sequentially according tothe following protocol: twice for 10 min at 500 � gmax, once for 15 min at2,000 � gmax, once for 30 min at 10,000 � gmax, and once for 60 min at70,000 � gmax using an SW28 rotor (Beckman Instruments). The exo-somes pelleted during the final centrifugation step were resuspended inPBS and repelleted at 70,000 � gmax. The repelleted exosomes were re-suspended in 5 ml of 2.6 M sucrose, 20 mM Tris-HCl (pH 7.2), and floatedonto a linear sucrose gradient (2.0–0.25 M sucrose, 20 mM Tris-HCl (pH7.2)) in an SW41 tube for 16 h at 270,000 � gmax to remove nonmem-branous protein (complexes). Gradient fractions (1 ml) were collected fromthe bottom of the tube and washed with PBS by centrifugation for 1 h at70,000 � gmax. The exosomes were then resuspended in PBS with a mix-ture of proteinase inhibitors (Roche), and the protein content was measuredusing a BCA protein assay kit (Bio-Rad). The aliquots were stored at�20°C for examination of the presence of exosomes by electron micros-copy. Usually, 300 �g of exosomal proteins was obtained from 1 � 108
RASF cells. The quality of each preparation was checked by electronmicroscopy.
Electron microscopy
Droplets of the sucrose-purified exosomes were placed directly on carbon-coated Formvar nickel-grids and left to adsorb onto the grids at 4°C for 1 h,then fixed with electromicromicroscopy-grade 2% paraformaldehyde(Fluka) and electromicromicroscopy-grade 0.1% glutaraldehyde (Fluka) at4°C for 1 h. After blocking with 2% cold fish gelatin (Sigma-Aldrich)/0.1%BSA/PBS for 20 min, the grids were immunolabeled using a mouse mono-clonal anti-TNF-� Ab (Santa Cruz Biotechnology) for 1 h at room tem-
perature. The unbound Ab was removed by four washes, and then revealedby incubation with 12-nm anti-mouse IgG-gold particles (The JacksonLaboratory). After extensive washes with PBS, the grids were stained with1% aqueous uranyl acetate before observation under a Hitachi H7000 elec-tron microscope (Electronic Instruments).
Mass spectrometry (MS) identification of proteins
The protein bands of interest from the colloidal blue-stained SDS-PAGEgel were excised, destained, dried, and then rehydrated and digested “in-gel” with trypsin overnight at 37°C for liquid chromatography-tandem MSidentification using a method as described previously (16). The MassLynx3.5 software (Micromass) was used for instrument operation, data acqui-sition, and analysis. The search for amino acid sequence similarity wasperformed using the basic local alignment search tool (BLAST) and/orScanps available from ExPASy (at �www.expasy.ch�).
ATPLite-M assay to evaluate sTNFRI blockade of the TNF-�-mediated cytotoxic effect of exosomes against L929 cells
L929 murine fibrosarcoma cells were grown in DMEM supplemented with10% FBS, penicillin (100 U/ml), streptomycin sulfate (100 �g/ml), andL-glutamine (2 mM) (DMEM/FCS). The cells were seeded in 96-well mi-crotiter plates at 4 � 104 cells/well and, the next day, serial dilutions ofRASF exosomes in DMEM with or without sTNFbp was added to the cellsin the presence of actinomycin D (1 �g/ml). After a 24-h incubation, thenumber of surviving cells was determined using the ATPlite assay as de-scribed by the manufacturer (Packard Instruments).
Preparation of cell nuclear extracts and colorimetric NF-�Bassay
The indicated numbers of RASF or OASF cells or CD4 T cells were stim-ulated with RASF or OASF exosomes alone or together with sTNFbp (1�g/ml) for the indicated time periods. Nuclear extracts were prepared fromthe harvested cells using a Nuclear Extract kit (Active Motif) and theNF-�B DNA-binding activity was detected using the TransAM NF-�BFamily Transcription Factor assay kit (Active Motif) according to the man-ufacturer’s protocol. Briefly, microwells coated with a double-stranded oli-gonucleotide containing the NF-�B consensus sequence were incubatedwith the nuclear extract for 1 h at room temperature and the washedthree times. The captured active transcription factor was incubated for1 h with Abs specific for the p65 or p50 NF-�B subunits, then for 1 hwith anti-rabbit IgG-coupled HRP. After incubation with developingsolution for 10 min, the OD was measured at 450 nm using a microtiterplate spectrophotometer.
Evaluation of the induction of collagenase production byexosomes
RASFs or OASFs were grown to 80% confluence in DMEM plus 10%FBS. The cells were then washed with PBS and cultured for 24 h in DMEMsupplemented with 2% FBS. The cells were then stimulated with RASF orOASF exosomes, with or without sTNFbp, as described above for 24 h.The supernatants were then collected for ELISA. Levels of collagenasewere determined by sandwich ELISA (Amersham Pharmaceutical). Theassay is a two-site ELISA sandwich format. Standards and samples wereincubated in microtiter wells precoated with rabbit anti-matrix metallopro-teinase 1 (MMP-1) Ab. The sample RASF supernatant was added at dif-ferent dilutions, incubated for 1 h, then a second polyclonal Ab to MMP-1was added. The binding of the second Ab bound to the wells was detectedwith donkey anti-rabbit conjugated with HRP using tetramethylbenzidine-hydrogen peroxide, in dimethylformamide as the developing agent. Thereaction was stopped by addition of an acid solution, and the absorbance ofthe reaction mixture read at 450 nm using a microtiter plate spectropho-tometer. The concentration of MMP-1 in a sample was determined byinterpolation from a standard curve. The sensitivity and linear range of thisstandard curve was 6.25–100 ng/ml.
Isolation of human CD4� T cells from peripheral blood
Peripheral blood obtained from healthy donors was diluted 1/1 with RPMI1640 medium (Invitrogen Life Technologies) and layered onto Ficoll-Paque (Pharmacia). Samples were centrifuged at 300 � g for 30 min andthe layer of mononuclear cells was collected and washed in RPMI 1640before removal of adherent cells by a 1-h incubation at 37°C in a petri dish.Nonadherent cells were collected and CD4� T cells were negatively iso-lated using the MACS CD4� T cell isolation kit (Miltenyi Biotec) accord-ing to the manufacturer’s instructions. The resulting CD4� T cell popula-tion (of �90% purity) was split for the in vitro culture and for analysis ofcell purity by flow cytometry.
7386 EXOSOMES MEDIATED T CELL AICD RESISTANCE
by guest on September 11, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

T cell activation
All T cells were cultured in RPMI 1640 supplemented with 10% heatinactivated normal human serum, 2 mM L-glutamine, 25 mM HEPESbuffer, 100 U/ml penicillin, and 100 �g/ml streptomycin at a cell densityof 1 � 106 T cells/ml. CD4� T cells were activated with 0.05 �g/mlanti-CD3 mAb (OKT3) (BD Pharmingen). Twenty-four hours after stim-ulation RASF exosomes were added, and this was repeated daily for 5days. Then, T cells were harvested for measurement of proliferation by[3H]thymidine incorporation and induction of IFN-� in the cultured super-natants using a standard ELISA method as described previously. In brief,18 h before harvesting the cells were pulsed with [3H]thymidine (1 �Ci/ml), harvested onto filter paper, and [3H]thymidine incorporation deter-mined by scintillation counting. The proliferation assays were performed intriplicate.
FACS analysis of apoptotic T cells
Induction of apoptosis in activated T cells as a result of anti-CD3 Abcross-linking was determined by flow cytometry analyses with PI and An-nexin VFITC staining (17). Briefly, anti-CD3-stimulated CD4 T cells wereharvested at the indicated time point after treatment with RASF exosomes.Early apoptotic cells were defined as single positive (annexin V�PI�) anddead cells as double positive (annexin V�PI�). The cells were also stainedwith CD4-allophycocyanin and anti-Fas-FITC or with CD4-allophycocya-nin and FasL-biotin plus streptavidin-FITC and analyzed by flow cytom-etry using a FACSCalibur flow cytometer and CellQuest software (BDBiosciences). Appropriate isotypic controls were used in every experiment.
Western blot analysis
CD4 T cells either treated or untreated with RASF exosomes were culturedand harvested at different time points. Cell pellets were washed once withPBS and lysed by SDS sample buffer containing 62.5 mM Tris-HCl (pH6.8), 2% (w/v) SDS, 10% glycerol, 50 mM DTT, and 0.1% (w/v) brom-phenol blue. The samples were then heated at 95°C and loaded on to a 10%polyacrylamide gel. After electrophoresis, the gels were blotted onto ni-trocellulose membranes, which were then incubated with blocking buffercontaining 1� TBS, 0.1% Tween 20, and 5% (w/v) nonfat dry milk for 1 hat room temperature. Proteins were detected using an anti-Akt, anti-phospho-Akt (Cell Signaling Technology), anti-I�B-�, Bcl-2, Bcl-xL, anti-�-Actin (BD Pharmingen), and FLIP (Alexis Biochemicals). The primaryAb was detected with an HRP-conjugated secondary Ab and visualized bychemiluminescent detection (Kirkegaard & Perry Laboratories).
For Western blot analysis of exosomal proteins, 10 �g of RASF orOASF exosomes or human rTNF-� (eBioscience) as an additional molec-ular marker were then prepared in lysis buffer (20 mM Tris-HCl (pH 7.4),150 mM NaCl, 10% glycerol, Nonidet NP40 0.2%, supplemented with aprotease inhibitor mixture; Roche Biochemicals). The proteins were re-solved by SDS-PAGE, transferred to nitrocellulose membranes by elec-troblotting, and the nonspecific binding sites were blocked by incubation inTBS containing 0.5% Tween 20 and 5% (w/v) dry milk. Immunoblot anal-yses were performed with the indicated Abs. Bound primary Abs werevisualized with HRP-conjugated goat anti-mouse-IgG (Jackson Immu-noResearch Laboratories) and chemiluminescent detection (Kirkegaard &Perry Laboratories).
Akt kinase assay
CD4 T cells were harvested at specific times after treatment with exosomesand lysed in lysis buffer. After equalizing the protein to 1.0 mg/200 �l ina 1.5-ml Eppendorf tube, a rabbit anti-mouse Akt Ab (Cell Signaling Tech-nology) was added, and the samples were rotated overnight at 4°C. Then,50 �l of protein A/G agarose bead slurry (Roche) was added, and thesamples were rotated at room temperature for 2 h. The beads were washedtwice with lysis buffer and kinase buffer, and the samples were suspendedin 50 �l kinase buffer supplemented with 10 mM ATP and 1 �g of GSK-3fusion protein (Cell Signaling Technology) and incubated at 30°C for 30min. The phosphorylation of GSK-3 fusion protein was analyzed by West-ern blot with phosphor-GSK-3 (Ser2119) Ab.
TaqMan RT-PCR
An ABI PRISM 7700 sequence detection system (Applied Biosystems)was used for amplification and detection of the gene transcripts of interest.In each TaqMan run, serial 5-fold dilutions of a single-stranded cDNAderived from a commercially available human positive control RNA (Ap-plied Biosystems) were amplified to create a standard curve, and values ofunknown samples were estimated relative to this standard curve. Standardcurves showed a linear relationship between the copy number (defined as1 ng of 1000-bp DNA � 9.1 � 1011 molecules) of the original internal
standard and the number of PCR cycles that were required to exceed apreset threshold, according to the method described previously (18). PCRsfor each sample were run in duplicate in three 5-fold serial dilutions. Fromthese standard curves, the relative amount of cDNA for the 18S ribosomalRNA, GAPDH, and each gene was determined and expressed as a ratio ofthe amount of this material in cells treated or untreated with RASF exo-somes after samples were standardized to the relative expression of the 18Sribosomal RNA and GAPDH.
Statistical analysis
Data are presented as arithmetic means of at least three independent ex-periments � SEM. Statistical analysis was performed using the Student-Newman-Keuls multiple range test and Duncan’s multiple range test. A pvalue of 0.05 was considered to be statistically significant.
ResultsThe membrane-bound form of TNF-� is detected on exosomesproduced by RASFs, but not OASFs
Many types of normal cells and tumor cells have now been shownto produce exosomes (19–26). Electron microscopic analysis ofthe supernatants of RASFs and OASFs that had been cultured for48 h indicated that these cells produce exosomes.
The function of exosomes remains largely unknown, but re-cently, a number of TNF family proteins have been identified onexosomes derived from dendritic cells. Western blotting of theexosomes produced by RASFs and OASFs indicated that TNF-� isassociated with the exosomes isolated from the supernatants ofRASFs but not OASFs (Fig. 1A). The presence of TNF-� on
FIGURE 1. Identification of a membrane form of TNF-� on the RASFexosomes. A, Exosomes were analyzed by Western blotting with a mono-clonal anti-TNF-� Ab. RASF or OASF exosomes were purified on sucrosedensity gradients from culture supernatants of primary synovial fibroblastsisolated from patients with either RA (n � 5) or OA (n � 5) as describedin the text. The exosomes were then lysed using a protein lysate buffer(Roche). A total of 10 �g of total protein lysate was either immunoblottedwith a mouse monoclonal anti-human TNF-� Ab (A, upper panel) or witha mouse monoclonal anti-14.3.3-� � Ab (A, bottom panel). To determinethe molecular mass of exosomal TNF-�, a recombinant soluble TNF-� (1�g) was mixed with lysates of RASF exosomes (B, lane 1), or OASFexosomes (lane 2) or lysate buffer (lane 3). The transferred membrane wasimmunoblotted with a mouse monoclonal anti-human TNF-� Ab. C, Im-munoelectron microscopy analysis of RASF vesicles. RASF or OASF exo-somes were purified from the supernatants of the primary synovial fibro-blasts isolated from either RA or OA patients by sucrose gradient andanalyzed by immunogold labeling using Ab specific for membrane TNF-�(bar, 200 nm). This micrograph is representative of two independent ex-periments with similar results of all five individual RASF and OASFexosomes.
7387The Journal of Immunology
by guest on September 11, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

exosomes is also evident with TNF-� colloidal gold immunostain-ing (Fig. 1C). Comparison of the molecular mass of TNF-� fromthe RASF exosomes with soluble TNF-� indicates that exosomesexpress the membrane-bound form and not the soluble form ofTNF-� (Fig. 1B). The 14-3.3-� protein is present in exosomesproduced by many different types of cells including RASF andOASF, and similar amounts of 14-3.3-� protein were detected inboth RASF and OASF exosomes (bottom panel, Fig. 1A). Thisfinding suggests that the absence of detectable membrane TNF-�on OASF exosomes is not due to lesser amounts of OASF exo-somes loaded on SDS-PAGE and that, therefore, the association ofTNF-� with RASF exosomes is a specific effect.
RASF exosomal TNF-� induces cytotoxicity of L929 cells
To determine whether the TNF-� associated with the RASF exo-somes is biologically active and to further confirm the association,we evaluated the ability of the exosomes to induce cytotoxicity ofL929 cells. Stimulation with TNF-� in L929 cells leads to celldeath and has been used for evaluation of the cytotoxic activity ofTNF-� (27, 28). Using the ATPLite assay, we found that RASFexosomes had a cytotoxic effect on L929 cells, but that OASFexosomes had only a minimal effect (Fig. 2A). The cytotoxic effectof the RASF exosomes correlated with their concentration (Fig.2A). To further determine whether the cytotoxic effect of the RASFexosomes on L929 cell was dependent on TNF-�, the RASF orOASF exosomes were preincubated with sTNFbp at different con-centrations for 1 h at 37°C before addition to the L929 cell cul-tures. Addition of sTNFbp blocked the cytotoxic effect of theRASF exosomes on the L929 cells in a concentration-dependentmanner (Fig. 2B), further suggesting that the RASF exosome-in-duced cytotoxicity of L929 is mediated by a TNF-�-signalingmechanism. The addition of sTNFbp has no effect on the low lev-els of cytotoxic activity of the OASF exosomes.
RASF exosomes induce activation of the NF-�B-signalingpathway in RASFs
TNF-�-mediated activation of the NF-�B pathway has been shownto play an important role in the resistance of RASF to apoptosisand may possibly drive the hyperplasia of rheumatoid synoviumthrough up-regulation of the NF-�B-associated proto-oncogenes.Therefore, we analyzed whether NF-�B is activated in RASFs bytreatment with RASF exosomes in the presence or absence ofsTNFbp (1 �g/ml). Nuclear proteins were extracted at differenttime points after treatment, and their ability to bind the NF-�Bconsensus sequence was quantitated by ELISA. OASF exosomes,or the exosome diluent (PBS), were used as controls. Stimulationof RASF cells with RASF exosomes (50 ng/ml) resulted in amarked increase in the binding activities of NF-�B subunits p50(Fig. 3A) and p65 (Fig. 3B), with the effect being more pronouncedin the case of p50 subunit (Fig. 3A). The activation of NF-�B wasdetected within 30 min after addition of RASF exosomes. Thisactivity was not observed on treatment with OASF exosomes, orPBS (Fig. 3A), and the activation was completely neutralized byaddition of sTNFbp, suggesting that activation of NF-�B is a spe-cific effect of RASF exosomes and is mediated by TNF-�. Simi-larly, stimulation of OASF cells with RASF exosomes also in-duced NF-�B activity similar to that in RASF (data not shown),
FIGURE 2. TNF-� associated with RASF exosomes are cytotoxic toL929 cell. A total of 4 � 104 cells/ml murine fibroblast cells L929 wereseeded in supplemented DMEM in 96-well microtiter plates. After 24 h,cells were treated with RASF or OASF exosomes at the concentrationsindicated (A) or cells were treated with RASF or OASF exosomes (50ng/ml) that had been preincubated with sTNFbp for 1 h at 37°C at theconcentrations indicated (B). The viability of the cells was assessed after24 h of incubation by the ATPLite assay as described in Materials andMethods. Data are presented as percentage of cytotoxicity for L929 cellline induced by the exosomes isolated from each RASF (n � 10) or OASF(n � 10). Sample size n � 10 with triplicates, mean � SEM. If the p valueis �0.05, indicating that these groups are significantly different, � � 0.05,Duncan’s multiple range test.
FIGURE 3. TNF-� associated with RASF exosomes induce activationof NF-�B in RASF cells. A total of 1 � 107 cells of RASFs or OASFs werestimulated with RASF or OASF exosomes (50 ng/ml), respectively, in thepresence of sTNFbp (1 �g/ml) or PBS as a control. The nuclei were thenextracted at the time points indicated. NF-�B DNA-binding activity wasmeasured using a colorimetric NF-�B assay (A and B) and the induction ofMMP-1 in the culture supernatants was measured by ELISA at hours 6, 12,and 24 after exosome stimulation (C). Values represent the means of trip-licate determinations (n � 5). Data are presented as means � SEM fromthree independent experiments. �, p 0.05; ��, p 0.01, calculated asuntreated cells vs stimulated cells. The data at each time point were derivedusing exosomes isolated from RASF or OASFs from five patients analyzedindividually.
7388 EXOSOMES MEDIATED T CELL AICD RESISTANCE
by guest on September 11, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

suggesting that activation of NF-�B activity mediated by RASFexosomes is not cell-type specific, but is specific to RASFexosomes.
As NF-�B is a well-characterized activator of MMP-1 (29–31),we next examined the effect of RASF exosomal TNF-� on induc-tion of MMP-1. The levels of MMP in the supernatants of cellstreated as described above were assayed by ELISA. In agreementwith the RASF exosome-mediated activation of NF-�B, we foundthat RASF exosomes induce the expression of MMP-1 in RASF ina time-dependent manner (Fig. 3C). The production of MMP-1 inthe cells treated with RASF exosomes plus sTNFbp was signifi-cantly lower ( p 0.0018) (Fig. 3C) and treatment with OASFexosomes or exosome diluent had little effect on induction ofMMP-1.
Production of RASF exosomes is enhanced by TNF-�stimulation
As the production of exosomes is regulated by the proteasomedegradation pathway, and the TNF-�-signaling pathway involvesthe ubiquitination pathway, we next examined whether the pro-duction of RASF exosomes is enhanced by TNF-� stimulation.RASFs were starved by culturing in DMEM medium with 2% FBSfor 16 h, and then stimulated with TNF-� at the different concen-trations for an additional 48 h. The exosomes in the supernatantswere purified by differential centrifugation, and the production ofexosomes was quantified using the Bradford protein assay. Theeffect of TNF-� on the production of exosomes by OASF wasanalyzed similarly. We found that the production of RASF exo-somes was enhanced significantly by addition of TNF-�-stimu-lated RASFs (Fig. 4A, p 0.0011) although TNF-� only enhancedthe yield of OASF exosomes slightly. As the membrane-boundform of TNF-� is detected on the RASF exosomes, the effect ofsTNFbp on the production of RASF exosomes was also examined.The yield of RASF exosomes was reduced in the RASFs culturedin the presence of sTNFbp in a concentration-dependent manner(Fig. 4B). In contrast, there was no effect on OASF exosome pro-duction. Therefore, the production of RASF exosomes is regulated
by a TNF-� pathway through both paracrine and autocrinemechanisms.
Coculture of RASF exosomes with CD4 T cells results insustained T cell proliferation
The infiltration of autoreactive CD4� T cells into synovium hasbeen considered the major instigator of joint inflammation, andboth hyperproliferation and decreased AICD of infiltrating T cellshave been proposed as mechanisms underlying the pathogenesis ofRA (13–15, 32). Therefore, we investigated whether incubation ofRASF exosomes with anti-CD3 Ab-activated CD4 T cells mighthave effects on CD4 T cell proliferation and subsequent AICD.RASF exosomes or OASF exosomes were added daily, starting withthe second day of culture of CD4 T cells that had been stimulated withanti-CD3 Ab. The effects of RASF exosomes treatment on T cellactivation were determined by [3H]thymidine incorporation assay,IL-2 and IFN-� induction in the cell culture supernatants. RASF exo-somes exhibited no significant effects on T cell proliferation or oninduction of IL-2 or IFN-� by CD4 T cells during the initial phases ofculture in comparison with the T cells stimulated with anti-CD3 alone.However, by day 6, the T cells cocultured with RASF exosomesmaintained proliferation (Fig. 5A), induction of IFN-� (Fig. 5B), andIL-2 (Fig. 5C) in a RASF-exosome dose-dependent manner whereasequal concentrations of the OASF exosomes did not have a compa-rable effect (data not shown). Preincubation of RASF exosomes with
FIGURE 4. TNF-� associated with RASF exosomes and exogenousTNF-� enhance the production of exosomes. RASF or OASF cells grownat 80% confluency were treated with a rTNF-� at different concentrations(A) or with a elevated concentrations of sTNFbp (B) for 48 h, then exo-somes were isolated and purified as described in Materials and Methods.The production of exosomes in the cell culture supernatants was quantifiedby the Bradford assay. Data are representative of at least three independentexperiments performed in triplicate. The data were derived using exosomesisolated from RASF or OASFs from 10 patients analyzed individually.There was a statistically significant enhancement of production of RASFexosomes in the TNF-�-stimulated RASF compared with the OASF. ��,p 0.01.
FIGURE 5. RASF exosomes sustain the proliferation of CD4 T cells atthe late phase of T cell activation. A total of 105 cells sorted of MACS-purified CD4 T cells were incubated in complete medium with 1 �g/mlplate-bound anti-CD3 in 96-well U-bottom plates (Nunc) for 1 day andthen different amounts of RASF exosomes were daily added to the anti-CD3-activated CD4 T cell culture. The proliferative response was deter-mined at days 2, 4, 6 of coculture by [3H]TdR incorporation. [3H]TdR wasadded for the remaining 18 h of culture. Proliferation was measured usinga liquid scintillation counter. Data are presented as mean cpm � SD oftriplicate cultures (A). Supernatants were harvested at day 6 after cultureand secreted IFN-� () and IL-2 (C) were measured by ELISA. The pro-portion of early apoptotic (annexin V�PI�) plus dead cells (annexinV�PI�) (percentage) was determined by flow cytometric analysis follow-ing annexin V and PI staining 6 days after the culture. One representativeof five independent experiments is shown (D). Results obtained from CD4T cells isolated five individuals of health subjects, in three independentexperiments, were pooled and expressed as mean � SD (E). The asteriskabove the bar indicates exosomes treatment groups that were significantlydifferent from the control group. �, p 0.05; ��, p 0.01. The percentagesof CD4 T cell death at day 2, 4, and 6 after anti-CD3 stimulation weredetermined (F). Data are presented as percentage of death of the CD4 T cellisolated from each health individuals (n � 5) with triplicates, mean �SEM, �, p 0.05; ��, p 0.01.
7389The Journal of Immunology
by guest on September 11, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
sTNFbp significantly reversed the effect of RASF exosomes in sus-taining T cell proliferation, suggesting that RASF exosome-associatedTNF-� plays a role in sustaining T cell proliferation. To further definethe nature of sustained T cell proliferation after treatment with RASFexosomes, we used FACS analysis to assess the effect of RASF exo-somes on T cell AICD at day 6 after anti-CD3 stimulation (Fig. 5D).Treatment with RASF exosomes at concentrations of 100 ng/ml sig-nificantly reduced the cell death of activated T cells up to 42% (Fig.5E), while the equivalent concentration of OASF exosomes did not.Further, the reduction in cell death varied in a dose-dependent fashion(Fig. 5F) and as the reciprocal of the effect on cell proliferation asshown in Fig. 5A. These results suggest that RASF exosomes mayplay a role in the induction of resistance to AICD in T cells and thatthe cross-talk between T cells and RASF via exosomes might reduceapoptosis of anti-CD3-activated T cells, leading to CD4 T cell con-tinuation of proliferation.
Up-regulated NF-�B and Akt activities are associated with anRASF exosome-mediated reduction in T cell AICD
Previous investigations showed that TNF-� stimulation of T cellsleads to the activation of both NF-�B and Akt. The activation ofNF-�B and Akt plays a critical role in cell survival signaling path-ways and development of autoimmune diseases in both humanautoimmune diseases and mouse models (33, 34). We examinedactivities of NF-�B and Akt at day 6 after T cells were exposed toRASF exosomes. The results of Western blot analyses of Akt in-dicate that levels of phosphorylated Akt were increased by stim-ulation of primary CD4 T cells with RASF exosomes in a dose-dependent manner (Fig. 6A, top panel). This induction effect wasspecific for phosphorylated Akt, because there were no changesdetected in total amount of Akt between treatments (Fig. 6A, sec-ond panel from the top). We also evaluated the phosphorylationstatus of GSK-3�, a known substrate of Akt. The phosphorylation
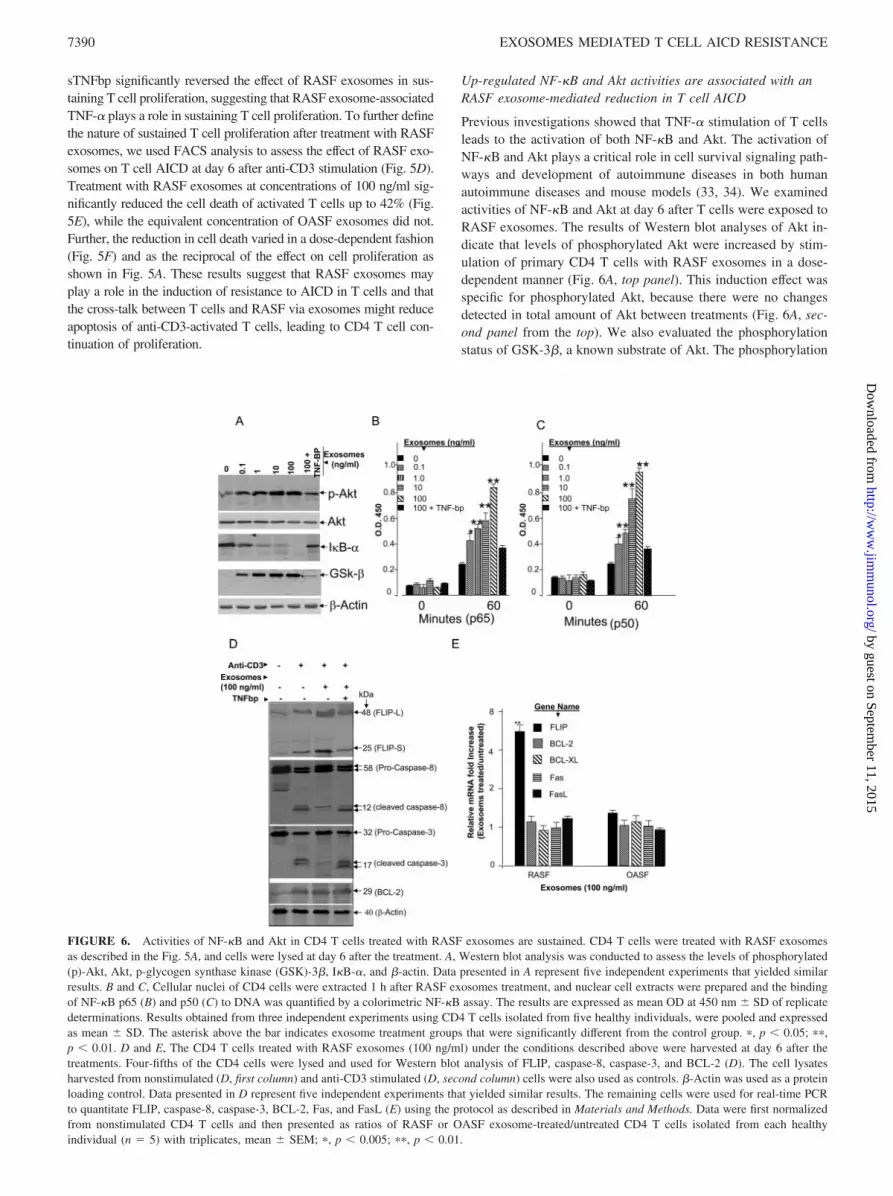
FIGURE 6. Activities of NF-�B and Akt in CD4 T cells treated with RASF exosomes are sustained. CD4 T cells were treated with RASF exosomesas described in the Fig. 5A, and cells were lysed at day 6 after the treatment. A, Western blot analysis was conducted to assess the levels of phosphorylated(p)-Akt, Akt, p-glycogen synthase kinase (GSK)-3�, I�B-�, and �-actin. Data presented in A represent five independent experiments that yielded similarresults. B and C, Cellular nuclei of CD4 cells were extracted 1 h after RASF exosomes treatment, and nuclear cell extracts were prepared and the bindingof NF-�B p65 (B) and p50 (C) to DNA was quantified by a colorimetric NF-�B assay. The results are expressed as mean OD at 450 nm � SD of replicatedeterminations. Results obtained from three independent experiments using CD4 T cells isolated from five healthy individuals, were pooled and expressedas mean � SD. The asterisk above the bar indicates exosome treatment groups that were significantly different from the control group. �, p 0.05; ��,p 0.01. D and E, The CD4 T cells treated with RASF exosomes (100 ng/ml) under the conditions described above were harvested at day 6 after thetreatments. Four-fifths of the CD4 cells were lysed and used for Western blot analysis of FLIP, caspase-8, caspase-3, and BCL-2 (D). The cell lysatesharvested from nonstimulated (D, first column) and anti-CD3 stimulated (D, second column) cells were also used as controls. �-Actin was used as a proteinloading control. Data presented in D represent five independent experiments that yielded similar results. The remaining cells were used for real-time PCRto quantitate FLIP, caspase-8, caspase-3, BCL-2, Fas, and FasL (E) using the protocol as described in Materials and Methods. Data were first normalizedfrom nonstimulated CD4 T cells and then presented as ratios of RASF or OASF exosome-treated/untreated CD4 T cells isolated from each healthyindividual (n � 5) with triplicates, mean � SEM; �, p 0.005; ��, p 0.01.
7390 EXOSOMES MEDIATED T CELL AICD RESISTANCE
by guest on September 11, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

of GSK-3 � correlated directly with the Akt phosphorylation sta-tus, supporting the view that the Akt pathway is functionally active(Fig. 6A, fourth panel from the top). Similarly, NF-�B activity of6-day cultured CD4 T cells was increased at 60 min after CD4 Tcells were stimulated with RASF exosomes (Fig. 6, B and C). Theactivity of NF-�B increased in a manner dependent on the dose ofRASF exosomes. Amounts of I�B-�, which is an NF-�B-specificinhibitor, was also reduced as the concentration of RASF exo-somes increased (Fig. 6A, third panel from the top), which is cor-related with increased activation of NF-�B. These results suggestthat a significant activation of both NF-�B and Akt were observedin CD4 T cells after 6 days of anti-CD3 stimulation in the pres-ence, but not absence, of RASF exosomes. It is unlikely that dif-ferences in activity of either Akt or NF-�B resulted from thechanges of global protein expression because the levels of �-actinloaded on each lane were not affected by RASF exosome treat-ments (Fig. 6A, bottom panel).
Because activation of Akt or NF-�B has been shown to up-regulate a number of apoptosis-associated proteins (35–37), weexamined the effects of RASF exosomes on the expression ofFLIP, Bcl-2, and on the cleavage of both caspase-8 and caspase-3.CD4-T cells stimulated with anti-CD3 and treated with RASF exo-somes have more FLIP (Fig. 6D, third column) but similar Bcl-2detected in the cell lysates compared with lysates from CD3-stim-ulated cells not treated with exosomes (second column). This resultparalleled partially blocked cleavage of both capase-8 and -3, Ad-dition of sTNFbp restored this cleavage process (Fig. 6D, fourthcolumn), suggesting that RASF exosome-associated membraneTNF-� may play a role in blocking caspase activation throughup-regulation of the FLIP pathway. RASF exosome-mediatedblockade of caspase-8 and -3 activation was seen in the context ofTCR signaling as non-CD3-stimulated cells did not have eithercaspase-8 or caspase-3 activated (Fig. 6D, first column). To deter-mine whether the RASF-exosome-mediated induction of FLIPprotein had a corresponding increase in mRNA, the expression ofFLIP mRNA was quantified by TaqMan PCR analysis. FLIPmRNA was significantly up-regulated after CD4 T cells weretreated with RASF exosomes (Fig. 6E). This result was not due tothe amount of mRNA used for the TaqMan PCR analysis as therewere no differences in the amount of other apoptosis-related genesamplified (Fig. 6E).
DiscussionRASFs and OASFs produce exosomes with a size distribution sim-ilar to that seen for exosomes from other sources and consistentwith the size criterion for exosomes proposed by Thery et al. (26).The array of associated proteins identified by using the liquid chro-matography-tandem MS analysis (Table I) is consistent with thoseidentified in exosomes from other cell types, such as B lympho-cytes and dendritic cells (5, 38, 39) and indicates that the RASFexosomes are derived from the MVB. Notably, however, RASFexosomal proteins include TNF-� which is absent from OASFexosomes. The presence of TNF-� was of particular interest asTNF-� plays a pathogenic role in RA, and the membrane form ofTNF-� has been recognized as playing an important role in theinflammatory response and induction of arthritis. Here, we showthat the membrane form of TNF-� is associated with exosomesproduced by RASF. Three lines of evidences support this conclu-sion. First, TNF-� was detected by direct immunocolloidal goldstaining and Western blot analysis of RASF but was absent inOASF exosomes. Second, RASF exosomes induced the cytotox-icity of TNF-�-sensitive L929 cells, with soluble TNFR blockingthis cytotoxic effect. Finally, RASF exosomes were potent induc-ers of NF-�B activation in the RASF that led to the degradation ofI�B-� and induction of MMP-1. Importantly, RASF exosomes notonly stimulated TNF-� signaling, leading to induction of MMP-1and activation of NF-�B, but they also induced resistance to apo-ptosis of activated T cells in a paracrine manner.
It is well-recognized that activation of both Akt and NF-�B areregulated by TNF-�-signaling pathway and that activated Akt andNF-�B have pathogenic effects in the development of rheumatoidarthritis (15). Our data indicate that the effects of RASF exosomeson induction of MMP-1 by RASFs and on AICD defects of CD4T cells are mediated by TNF-� with activation of Akt and NF-�Bpathways. Furthermore, blockade of the activation of caspase-8and caspase-3 and the up-regulation of anti-apoptosis gene FLIPmay play a role in AICD defect.
The important role of TNF-� signaling in T cell survivalthrough activation Akt was shown using TNFR knockout mice(40–43). Akt phosphorylation is defective in TNFR-deficient Tcells and the activation of Akt during T cell activation is an im-portant regulatory point for initiation and progression of the T cell
Table I. Identities of proteins found in synovial fibroblast-derived exosomes
Protein Name Acc. No. RASF OASF Unique Peptides Coverage (%)
TNF-� NP_038721 � � 6 25Myosin CAB05105 � � 4 8Heat sock protein-90 AAA36026 � � 4 25Ezrin Q96CU8 � � 2 12Moesin Moes_human � � 3 15Annexin VI Anx6_human � � 6 15Heat shock protein 70 A27077 � � 4 18Actin AAH08633 � � 7 25MHC class I Ag Q9TP25 � � 6 12Enolase 1 PO6733 � � 5 16Aldose reductase P151221 � � 2 12Heat shock 70 kDa P08107 � � 5 12Keratin 10 P13645 � � 2 1114-3-3-� 2BQ0_B � � 4 14Syntenin 000560 � � 4 12Complement C1q chain C C1HuQC � � 6 5Annexin II ANx2_human � � 5 18Protein phosphatae 1 PO8129 � � 6 15Kinesin-like protein Q9NS87 � � 2 12Fibronectin precursor CAA26536 � � 5 15
7391The Journal of Immunology
by guest on September 11, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

response. Overexpression of Akt resulted in the development ofautoimmune diseases (44, 45). Therefore, RASF exosome-medi-ated activation of Akt in anti-CD3 activation of CD4 T cells pointsto one mechanism that may play a contributing role in develop-ment of RA.
Our data suggest that RASF exosomes not only activate Akt butalso sustain NF-�B activity. Numerous lines of evidence havestrongly supported TNF-�-mediated activation of NF-�B as one ofkey regulators of T cell survival and of MMP induction of RASFcells. Activation of NF-�B has also been proposed as a pathogenicfactor contributing to hyperplasia of rheumatoid synovium. Ourdata support significant activation of NF-�B induced in T cells andRASF treated with RASF-derived exosomes, whereas OASF exo-somes caused only slight time-dependent NF-�B activation in CD4T cells. Our data also show that the rapid reduction of I�B-� cor-related directly with the NF-�B activation status, supporting theview that the NF-�B pathway is functionally active. The observa-tion that MMP induction in RASFs was blocked by addition ofsTNFbp further supports the notion that NF-�B is also activated inRASFs. Other data in the literature suggest that the activation ofNF-�B in T cells leads to up-regulation of both FLIP and Bcl-2(46–49). However, our data show that more FLIP but not Bcl-2was detected after RASF exosome treatment, suggesting that thiseffect may be due to the net result of multiple proteins packed inthe exosomes interacting with the NF-�B pathway. Further inves-tigation of which exosomal protein(s) play a role in the preferentialactivation of FLIP is underway.
Data generated by other investigators suggest that membrane-associated TNF-� is biologically active, and it has been presumedthat the cytotoxic and inflammatory effects of TNF-� are throughcell-to-cell contact (12). In contrast, our findings suggest thatmembrane-associated TNF-� may exercise its role through exo-somes, instead of, or at least in addition to, cell-cell contact. Exo-somal membrane proteins including TNF-� may play distinct rolein comparison with soluble TNF-�. TNF-� is first produced as a26-kDa transmembrane molecule (mTNF) which is cleaved by themetalloproteinase-disintegrin TNF-�-converting enzyme (50) togenerate a soluble 17-kDa molecule (sTNF). Synovial inflamma-tion and joint erosion appear to be dependent, at least in part, oncell-associated TNF-� signaling (12, 32, 51). Using a novel trans-genic mouse that expresses only a membrane-associated form ofTNF-�, Kollias and coworkers (12) demonstrated that expressionof the 26-kDa form of TNF-� was sufficient in itself to inducearthritis. These animals spontaneously develop a pattern of arthritissimilar to those of human RA at �6–8 wk of age (12). AlthoughsTNF is regarded as the main ligand for TNFR1, mTNF is con-sidered to be more effective than sTNF in activating TNFR2 (52).TNFR2 engagement leads to binding of TNFR-associated factors 1and 2, leading to NF-�B activation and cell survival (53). Synovialhyperplasia and joint erosion appear to be dependent, at least inpart, on cell-associated TNF-� signaling, with involvement ofTNF-R2 (54–56). We have demonstrated that RASF exosomalTNF-� strongly stimulates the activation of NF-�B in RASFs.Whether the preferential binding affinity of TNFR2 for the 26-kDaform of TNF-� on cell membranes is also true for TNF-� on RASFexosomes is currently being addressed through studies of TNFR1-or TNFR2-deficient cell lines.
In addition to TNF-�, other proteins packed in the RASF exo-somes may play a role modulating infiltrating immune cells. RASFexosomes can be endocytosed by autoreactive T cells and B cells,and by serving as a shuttle between cells (1, 2, 7, 19), they maydeliver proteins and/or lipids to the targeted cells. Thus, RASFexosomes are poised to alter the biological functions of multiplecell types, and one might consider therapeutic targeting of exo-
somes with additional agents beyond those binding and blockingTNF-�.
Evidence indicates that the ubiquitin pathway regulates the for-mation and rate of exosome budding through the cellular mem-brane, and TNF-� signaling plays a role in regulation of ubiquitin-mediated protein degradation (57–63). The observation thatTNF-� can regulate the production of exosomes has not been pre-viously recognized, and one might speculate that TNF-�-mediatedenhancement of the production of RASF exosomes may also en-hance the ubiquitination and immunogenicity of self proteins, ashas been proposed by other investigators (64). The production ofRASF exosomes may be also accelerated by the activation of path-ways that regulate assembly of ubiquinated proteins into exo-somes. Soluble N-ethylmaleimide-sensitive factor attachment re-ceptor-mediated fusion of a TNF-�-contained vesicular-likestructure is a requirement of this trafficking pathway (5). Our dataindicate that TNF-� stimulation leads to enhanced production ofRASF exosomes, and identification of specific pathways that reg-ulate induction of RASF exosomes may lead to new therapeutictargets for RA treatment.
AcknowledgmentsWe thank Dr. Fiona Hunter for editorial assistance, Landon Wilson at UABMass Spectrometry Shared Facility for the MALDI-TOF-MS mass finger-printing analysis.
DisclosuresThe authors have no financial conflict of interest.
References1. Couzin, J. 2005. Cell biology: the ins and outs of exosomes. Science 308:
1862–1863.2. Kim, S. H., E. R. Lechman, N. Bianco, R. Menon, A. Keravala, J. Nash, Z. Mi,
S. C. Watkins, A. Gambotto, and P. D. Robbins. 2005. Exosomes derived fromIL-10-treated dendritic cells can suppress inflammation and collagen-induced ar-thritis. J. Immunol. 174: 6440–6448.
3. Kapsogeorgou, E. K., R. F. Abu-Helu, H. M. Moutsopoulos, andM. N. Manoussakis. 2005. Salivary gland epithelial cell exosomes: a source ofautoantigenic ribonucleoproteins. Arthritis Rheum. 52: 1517–1521.
4. Hsu, D. H., P. Paz, G. Villaflor, A. Rivas, A. Mehta-Damani, E. Angevin,L. Zitvogel, and J. B. Le Pecq. 2003. Exosomes as a tumor vaccine: enhancingpotency through direct loading of antigenic peptides. J. Immunother. 26:440–450.
5. Vincent-Schneider, H., P. Stumptner-Cuvelette, D. Lankar, S. Pain, G. Raposo,P. Benaroch, and C. Bonnerot. 2002. Exosomes bearing HLA-DR1 moleculesneed dendritic cells to efficiently stimulate specific T cells. Int. Immunol. 14:713–722.
6. Clayton, A., J. Court, H. Navabi, M. Adams, M. D. Mason, J. A. Hobot,G. R. Newman, and B. Jasani. 2001. Analysis of antigen presenting cell derivedexosomes, based on immuno-magnetic isolation and flow cytometry. J. Immunol.Methods 247: 163–174.
7. Segura, E., C. Nicco, B. Lombard, P. Veron, G. Raposo, F. Batteux,S. Amigorena, and C. Thery. 2005. ICAM-1 on exosomes from mature dendriticcells is critical for efficient naive T-cell priming. Blood 106: 216–223.
8. Hyrich, K. L., A. J. Silman, K. D. Watson, and D. P. Symmons. 2004. Anti-TNF�treatment in rheumatoid arthritis: an update on safety. Ann. Rheum. Dis. 63:1538–1543.
9. Feldmann, M., F. M. Brennan, E. Paleolog, A. Cope, P. Taylor, R. Williams,J. Woody, and R. N. Maini. 2004. Anti-TNF� therapy of rheumatoid arthritis:what can we learn about chronic disease? Novartis Found Symp. 256: 53–69,69–73, 106–111, 266–109.
10. Toussirot, E., and D. Wendling. 2004. The use of TNF-� blocking agents inrheumatoid arthritis: an overview. Expert Opin. Pharmacother. 5: 581–594.
11. Williams, R. O., J. Ghrayeb, M. Feldmann, and R. N. Maini. 1995. Successfultherapy of collagen-induced arthritis with TNF receptor-IgG fusion protein andcombination with anti-CD4. Immunology 84: 433–439.
12. Georgopoulos, S., D. Plows, and G. Kollias. 1996. Transmembrane TNF is suf-ficient to induce localized tissue toxicity and chronic inflammatory arthritis intransgenic mice. J. Inflamm. 46: 86–97.
13. Weyand, C. M., and J. J. Goronzy. 2003. Ectopic germinal center formation inrheumatoid synovitis. Ann. NY Acad. Sci. 987: 140–149.
14. Weyand, C. M., A. Braun, S. Takemura, and J. J. Goronzy. 2001. Lymphoidmicrostructures in rheumatoid synovitis. Curr. Dir. Autoimmun. 3: 168–187.
15. Zhang, H. G., Y. Wang, J. F. Xie, X. Liang, D. Liu, P. Yang, H. C. Hsu,R. B. Ray, and J. D. Mountz. 2001. Regulation of tumor necrosis factor �-me-diated apoptosis of rheumatoid arthritis synovial fibroblasts by the protein kinaseAkt. Arthritis Rheum. 44: 1555–1567.
7392 EXOSOMES MEDIATED T CELL AICD RESISTANCE
by guest on September 11, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

16. Yang, X., J. Wang, C. Liu, W. E. Grizzle, S. Yu, S. Zhang, S. Barnes,W. J. Koopman, J. D. Mountz, R. P. Kimberly, and H. G. Zhang. 2005. Cleavageof p53-vimentin complex enhances tumor necrosis factor-related apoptosis-in-ducing ligand-mediated apoptosis of rheumatoid arthritis synovial fibroblasts.Am. J. Pathol. 167: 705–719.
17. Hsu, H. C., J. Shi, P. Yang, X. Xu, C. Dodd, Y. Matsuki, H. G. Zhang, andJ. D. Mountz. 2001. Activated CD8� T cells from aged mice exhibit decreasedactivation-induced cell death. Mech. Ageing Dev. 122: 1663–1684.
18. Zhang, H. G., K. Hyde, G. P. Page, J. P. Brand, J. Zhou, S. Yu, D. B. Allison,H. C. Hsu, and J. D. Mountz. 2004. Novel tumor necrosis factor �-regulatedgenes in rheumatoid arthritis. Arthritis Rheum. 50: 420–431.
19. Fevrier, B., and G. Raposo. 2004. Exosomes: endosomal-derived vesicles ship-ping extracellular messages. Curr. Opin. Cell Biol. 16: 415–421.
20. Laulagnier, K., D. Grand, A. Dujardin, S. Hamdi, H. Vincent-Schneider,D. Lankar, J. P. Salles, C. Bonnerot, B. Perret, and M. Record. 2004. PLD2 isenriched on exosomes and its activity is correlated to the release of exosomes.FEBS Lett. 572: 11–14.
21. de Gassart, A., C. Geminard, D. Hoekstra, and M. Vidal. 2004. Exosome secre-tion: the art of reutilizing nonrecycled proteins? Traffic 5: 896–903.
22. Mears, R., R. A. Craven, S. Hanrahan, N. Totty, C. Upton, S. L. Young, P. Patel,P. J. Selby, and R. E. Banks. 2004. Proteomic analysis of melanoma-derivedexosomes by two-dimensional polyacrylamide gel electrophoresis and mass spec-trometry. Proteomics 4: 4019–4031.
23. Hegmans, J. P., M. P. Bard, A. Hemmes, T. M. Luider, M. J. Kleijmeer,J. B. Prins, L. Zitvogel, S. A. Burgers, H. C. Hoogsteden, and B. N. Lambrecht.2004. Proteomic analysis of exosomes secreted by human mesothelioma cells.Am. J. Pathol. 164: 1807–1815.
24. Janiszewski, M., A. O. Do Carmo, M. A. Pedro, E. Silva, E. Knobel, andF. R. Laurindo. 2004. Platelet-derived exosomes of septic individuals possessproapoptotic NAD(P)H oxidase activity: a novel vascular redox pathway. Crit.Care Med. 32: 818–825.
25. de Gassart, A., C. Geminard, B. Fevrier, G. Raposo, and M. Vidal. 2003. Lipidraft-associated protein sorting in exosomes. Blood 102: 4336–4344.
26. Thery, C., L. Zitvogel, and S. Amigorena. 2002. Exosomes: composition, bio-genesis and function. Nat. Rev. Immunol. 2: 569–579.
27. Eigler, A., J. Moeller, and S. Endres. 1995. Exogenous and endogenous nitricoxide attenuates tumor necrosis factor synthesis in the murine macrophage cellline RAW 264.7. J. Immunol. 154: 4048–4054.
28. Volpe, E. A. 1995. Extremely high production of thymidine by TNF-susceptibleL929 cells. Cancer Lett. 89: 169–176.
29. Vincenti, M. P., C. I. Coon, and C. E. Brinckerhoff. 1998. Nuclear factor �B/p50activates an element in the distal matrix metalloproteinase 1 promoter in inter-leukin-1�-stimulated synovial fibroblasts. Arthritis Rheum. 41: 1987–1994.
30. Barchowsky, A., D. Frleta, and M. P. Vincenti. 2000. Integration of the NF-�Band mitogen-activated protein kinase/AP-1 pathways at the collagenase-1 pro-moter: divergence of IL-1 and TNF-dependent signal transduction in rabbit pri-mary synovial fibroblasts. Cytokine 12: 1469–1479.
31. Tasaki, K., Y. Shintani, T. Saotome, A. Andoh, Y. Fujiyama, S. Hozawa, andT. Bamba. 2003. Pro-inflammatory cytokine-induced matrix metalloproteinase-1(MMP-1) secretion in human pancreatic periacinar myofibroblasts. Pancreatol-ogy 3: 414–421.
32. Youn, J., H. Y. Kim, J. H. Park, S. H. Hwang, S. Y. Lee, C. S. Cho, and S. K. Lee.2002. Regulation of TNF-�-mediated hyperplasia through TNF receptors,TRAFs, and NF-�B in synoviocytes obtained from patients with rheumatoid ar-thritis. Immunol. Lett. 83: 85–93.
33. Ohashi, P. S. 2002. T-cell signalling and autoimmunity: molecular mechanismsof disease. Nat. Rev. Immunol. 2: 427–438.
34. Patel, R. K., and C. Mohan. 2005. PI3K/AKT signaling and systemic autoim-munity. Immunol. Res. 31: 47–55.
35. Dohrman, A., T. Kataoka, S. Cuenin, J. Q. Russell, J. Tschopp, and R. C. Budd.2005. Cellular FLIP (long form) regulates CD8� T cell activation throughcaspase-8-dependent NF-�B activation. J. Immunol. 174: 5270–5278.
36. Bai, S., H. Liu, K. H. Chen, P. Eksarko, H. Perlman, T. L. Moore, andR. M. Pope. 2004. NF-�B-regulated expression of cellular FLIP protects rheu-matoid arthritis synovial fibroblasts from tumor necrosis factor �-mediated apo-ptosis. Arthritis Rheum. 50: 3844–3855.
37. Micheau, O., S. Lens, O. Gaide, K. Alevizopoulos, and J. Tschopp. 2001. NF-�Bsignals induce the expression of c-FLIP. Mol. Cell. Biol. 21: 5299–5305.
38. Wubbolts, R., R. S. Leckie, P. T. Veenhuizen, G. Schwarzmann, W. Mobius,J. Hoernschemeyer, J. W. Slot, H. J. Geuze, and W. Stoorvogel. 2003. Proteomicand biochemical analyses of human B cell-derived exosomes: potential implica-tions for their function and multivesicular body formation. J. Biol. Chem. 278:10963–10972.
39. Skokos, D., H. Goubran-Botros, M. Roa, and S. Mecheri. 2002. Immunoregula-tory properties of mast cell-derived exosomes. Mol. Immunol. 38: 1359–1362.
40. Horai, R., A. Nakajima, K. Habiro, M. Kotani, S. Nakae, T. Matsuki, A. Nambu,S. Saijo, H. Kotaki, K. Sudo, et al. 2004. TNF-� is crucial for the developmentof autoimmune arthritis in IL-1 receptor antagonist-deficient mice. J. Clin. Invest.114: 1603–1611.
41. Kim, E. Y., and H. S. Teh. 2004. Critical role of TNF receptor type-2 (p75) as acostimulator for IL-2 induction and T cell survival: a functional link to CD28.J. Immunol. 173: 4500–4509.
42. Kim, E. Y., and H. S. Teh. 2001. TNF type 2 receptor (p75) lowers the thresholdof T cell activation. J. Immunol. 167: 6812–6820.
43. Song, J., T. So, M. Cheng, X. Tang, and M. Croft. 2005. Sustained survivinexpression from OX40 costimulatory signals drives T cell clonal expansion. Im-munity 22: 621–631.
44. Parsons, M. J., R. G. Jones, M. S. Tsao, B. Odermatt, P. S. Ohashi, andJ. R. Woodgett. 2001. Expression of active protein kinase B in T cells perturbsboth T and B cell homeostasis and promotes inflammation. J. Immunol. 167:42–48.
45. Kelly, E., A. Won, Y. Refaeli, and L. Van Parijs. 2002. IL-2 and related cytokinescan promote T cell survival by activating AKT. J. Immunol. 168: 597–603.
46. Zhong, F., M. C. Davis, K. S. McColl, and C. W. Distelhorst. 2006. Bcl-2 dif-ferentially regulates Ca2� signals according to the strength of T cell receptoractivation. J. Cell Biol. 172: 127–137.
47. Grayson, J. M., A. J. Zajac, J. D. Altman, and R. Ahmed. 2000. Cutting edge:increased expression of Bcl-2 in antigen-specific memory CD8� T cells. J. Im-munol. 164: 3950–3954.
48. Ferreira, C., T. Barthlott, S. Garcia, R. Zamoyska, and B. Stockinger. 2000.Differential survival of naive CD4 and CD8 T cells. J. Immunol. 165: 3689–3694.
49. Linette, G. P., M. J. Grusby, S. M. Hedrick, T. H. Hansen, L. H. Glimcher, andS. J. Korsmeyer. 1994. Bcl-2 is upregulated at the CD4�CD8� stage duringpositive selection and promotes thymocyte differentiation at several controlpoints. Immunity 1: 197–205.
50. Moss, M. L., S. L. Jin, J. D. Becherer, D. M. Bickett, W. Burkhart, W. J. Chen,D. Hassler, M. T. Leesnitzer, G. McGeehan, M. Milla, et al. 1997. Structuralfeatures and biochemical properties of TNF-� converting enzyme (TACE).J. Neuroimmunol. 72: 127–129.
51. Mageed, R. A., G. Adams, D. Woodrow, O. L. Podhajcer, and Y. Chernajovsky.1998. Prevention of collagen-induced arthritis by gene delivery of soluble p75tumour necrosis factor receptor. Gene Ther. 5: 1584–1592.
52. Grell, M., E. Douni, H. Wajant, M. Lohden, M. Clauss, B. Maxeiner,S. Georgopoulos, W. Lesslauer, G. Kollias, and K. Pfizenmaier. 1995. The trans-membrane form of tumor necrosis factor is the prime activating ligand of the 80kDa tumor necrosis factor receptor. Cell 83: 793–802.
53. Yeh, W. C., R. Hakem, M. Woo, and T. W. Mak. 1999. Gene targeting in theanalysis of mammalian apoptosis and TNF receptor superfamily signaling. Im-munol. Rev. 169: 283–302.
54. Dieude, P., J. Osorio, E. Petit-Teixeira, S. Moreno, S. Garnier, S. Cailleau-Moindrault, C. Stalens, S. Lasbleiz, T. Bardin, B. Prum, and F. Cornelis. 2004.A TNFR1 genotype with a protective role in familial rheumatoid arthritis. Ar-thritis Rheum. 50: 413–419.
55. Constantin, A., P. Dieude, V. Lauwers-Cances, B. Jamard, B. Mazieres,A. Cambon-Thomsen, F. Cornelis, and A. Cantagrel. 2004. Tumor necrosis factorreceptor II gene polymorphism and severity of rheumatoid arthritis. ArthritisRheum. 50: 742–747.
56. Barton, A., S. John, W. E. Ollier, A. Silman, and J. Worthington. 2001. Asso-ciation between rheumatoid arthritis and polymorphism of tumor necrosis factorreceptor II, but not tumor necrosis factor receptor I, in Caucasians. ArthritisRheum. 44: 61–65.
57. Hu, X. 2003. Proteolytic signaling by TNF�: caspase activation and I�B degra-dation. Cytokine 21: 286–294.
58. Zhou, J., T. Schmid, and B. Brune. 2003. Tumor necrosis factor-� causes accu-mulation of a ubiquitinated form of hypoxia inducible factor-1� through a nuclearfactor-�B-dependent pathway. Mol. Biol. Cell 14: 2216–2225.
59. Li, Y. P., S. H. Lecker, Y. Chen, I. D. Waddell, A. L. Goldberg, and M. B. Reid.2003. TNF-� increases ubiquitin-conjugating activity in skeletal muscle by up-regulating UbcH2/E220k. FASEB J. 17: 1048–1057.
60. Legler, D. F., O. Micheau, M. A. Doucey, J. Tschopp, and C. Bron. 2003. Re-cruitment of TNF receptor 1 to lipid rafts is essential for TNF�-mediated NF-�Bactivation. Immunity 18: 655–664.
61. Naito, Y., O. Handa, T. Takagi, T. Ishikawa, E. Imamoto, S. Nakagawa,T. Yamaguchi, N. Yoshida, H. Matsui, and T. Yoshikawa. 2002. Ubiquitin-pro-teasome inhibitor enhances tumour necrosis factor-�-induced apoptosis in ratgastric epithelial cells. Aliment Pharmacol. Ther. 16(Suppl. 2): 59–66.
62. Reid, M. B., and Y. P. Li. 2001. Tumor necrosis factor-� and muscle wasting: acellular perspective. Respir. Res. 2: 269–272.
63. Llovera, M., C. Garcia-Martinez, N. Agell, F. J. Lopez-Soriano, andJ. M. Argiles. 1997. TNF can directly induce the expression of ubiquitin-depen-dent proteolytic system in rat soleus muscles. Biochem. Biophys. Res. Commun.230: 238–241.
64. Rouard, H., B. Klonjkowski, J. Marquet, C. Lahet, S. Mercier, M. Andrieu,P. Maison, V. Molinier-Frenkel, M. Eloit, J. P. Farcet, et al. 2003. Adenoviraltransgene ubiquitination enhances mouse immunization and class I presentationby human dendritic cells. Hum. Gene Ther. 14: 1319–1332. Vol. 176, No. 12
7393The Journal of Immunology
by guest on September 11, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
CORRECTIONSCaprio-Young, J. C., J. J. Bell, H.-H. Lee, J. Ellis, D. Nast, G. Sayler, B. Min, and H. Zaghouani. 2006. Neonatally primedlymph node, but not splenic T cells, display a Gly-Gly motif within the TCR �-chain complementarity-determining region3 that controls affinity and may affect lymphoid organ retention. J. Immunol. 176: 357–364.
In Results, in the penultimate sentence of the second paragraph under the heading A Gly-Gly motif is conserved within theCDR3 of lymph node T cell hybridomas, reference to Figure 7a and 7b are reversed. The corrected sentence is shown below.
The lymph node TCR contains a rigid loop with an extended planar surface (Fig. 7b), whereas the splenic Th1 TCRpresents a round shape with a less extended surface (Fig. 7a).
Taieb, J., N. Chaput, N. Schartz, S. Roux, S. Novault, C. Menard, F. Ghiringhelli, M. Terme, A. F. Carpentier, G.Darrasse-Jese, F. Lemonnier, and L. Zitvogel. 2006. Chemoimmunotherapy of tumors: cyclophosphamide synergizes withexosome based vaccines. J. Immunol. 176: 2722–2729.
The tenth author’s last name is incorrect. The correct name is Guillaume Darrasse-Jeze.
Swiecki, M. K., M. W. Lisanby, F. Shu, C. L. Turnbough, Jr., and J. F. Kearney. 2006. Monoclonal antibodies for Bacillusanthracis spore detection and functional analyses of spore germination and outgrowth. J. Immunol. 176: 6076–6084.
In Table II, the data reported for GA2–3b in column six (�rmlD spores) should be negative (�) not 2-log shift (��).The corrected table is shown below.
Table II. mABs raised against irradiated B. anthracis spores orpurified B. anthracis exosporium
mAbs
Western FACS
Anti-E. coliBc1A
Anti-Deglyco,Bc1A
WT Sternespores
�bc1Aspores
�rmlDspores
AB2 NDa ND � � ��AF10b � � � � ��AH88 � � � � ��BD8b � � � ND NDBE12
b� � � � ��
BF1-4b
� � � � ��BF12 ND ND � � ��BG11b � � � ND NDCA3* � � � � ��DE3-1b � � � � ��DE12 � � � � ��FD3-4b � � � ND NDEF12b � � � ND NDAA2-1 ND ND � � ��BA10-1b � � � � ��DH4-1b � � � �� ��EA2-1 ND ND � � ��EA4-10b � � � � ��EA4-10-4 � � � � ��EG4-4b � � � � ��FH6-1b � � � �� ��GA2-3b � � � � �GB4-4b � � � ND NDGB4-6-2 ND ND � � ��HB2-2 ND ND � � ��IC801b � � � � ��JB5-1b � � � � ��JC8-5sb � � � � ��
a ND, Not determined, �, negative by Western or FACS; �, positive by Westernor l-log shift by FACS; ��, 2-log shift by FACS ABS-EF12, mAbs raised againstspores; AA2-I-JC8-5, mAbs and raised against exosporium.
b Included in Ref. 9.
The Journal of Immunology
Copyright © 2006 by The American Association of Immunologists, Inc. 0022-1767/06/$02.00

de Nadaı, P., A.-S. Charbonnier, C. Chenivesse, S. Senechal, C. Fournier, J. Gilet, H. Vorng, Y. Chang, P. Gosset, B.Wallaert, A.-B. Tonnel, P. Lassalle, and A. Tsicopoulos. 2006. Involvement of CCL18 in allergic asthma. J. Immunol.176: 6286–6293.
In Results, in the last sentence of the paragraph under the heading CCL18 is up-regulated in BAL and sera from AApatients, and in Figure 4C, the concentration of serum CCL18 is expressed incorrectly as “pg/ml” instead of “ng/ml.” Thecorrected sentence and figure are shown below.
CCL18 was significantly elevated in AA (73.9 � 11.2 ng/ml) compared with NA (31.7 � 5 ng/ml) subjects (Fig. 4C).
Zhang, H.-G., C. Liu, K. Su, S. Yu, L. Zhang, S. Zhang, J. Wang, X. Cao, W. Grizzle, and R. P. Kimberly. 2006. A membraneform of TNF-� presented by exosomes delays T cell activation-induced cell death. J. Immunol. 176: 7385–7393.
The third author’s first name is incorrect. The correct name is Kaihong Su.
Vaknin-Dembinsky, A., K. Balashov, and H. L. Weiner. 2006. IL-23 is increased in dendritic cells in multiple sclerosisand down-regulation of IL-23 by antisense oligos increases dendritic cell IL-10 production. J. Immunol. 176: 7768–7774.
During production, the figure from an unrelated article was inadvertently inserted as the image for Figure 8. The correctfigure is shown below. The error has been corrected in the online version, which now differs from the print version asoriginally published.
2025The Journal of Immunology