accumulation and translocation of cd metal and the cd-induced production of glutathione and...
TRANSCRIPT

ORIGINAL PAPER
Accumulation and translocation of Cd metal and the Cd-inducedproduction of glutathione and phytochelatins in Vicia faba L.
Radomır Cabala • L’udmila Slovakova •
Manal El Zohri • Hartmut Frank
Received: 3 February 2010 / Revised: 21 October 2010 / Accepted: 23 November 2010 / Published online: 8 December 2010
� Franciszek Gorski Institute of Plant Physiology, Polish Academy of Sciences, Krakow 2010
Abstract Translocation of cadmium (Cd) in the tissues of
Vicia faba, the water content in biomass, the biomass
production, and the glutathione and phytochelatin tissue
concentrations were studied and correlated with the plant
sensitivity and/or tolerance to Cd. The total concentrations
of Cd were determined by inductively coupled plasma/
mass spectrometry (ICP-MS), the concentrations of gluta-
thione (GSH) and phytochelatins 2 and 3 (PC2 and PC3)
were determined by on-line high performance liquid
chromatography/electrospray-ionization tandem mass
spectrometry (HPLC–ESI–MS–MS) in the roots and leaves
of the sensitive and the tolerant cultivars of V. faba grown
in Cd containing nutrient solutions (NS, 0–100 lmol l-1
Cd2?). Both the cultivars of V. faba accumulate a major
portion of Cd in the roots and only a minor part of ca. 4%
in the leaves. The differences between the cultivars con-
cerning Cd accumulation in leaves were apparent from
higher Cd concentrations in NS and the Cd amount in the
sensitive cultivar was approximately twice as high. In the
roots, the differences between the cultivars in the Cd
accumulation were only statistically significant with the
highest Cd concentrations in NS, with the tolerant cultivar
accumulating about 16% more of Cd compared to the
sensitive one. The biomass production of the sensitive
cultivar decreased approximately twice as fast with
increasing Cd concentration in NS. The biomass water
content decreased with increasing Cd concentration in NS
in both the cultivars. In general, the GSH concentration did
not linearly correlate with Cd accumulation, except for the
roots of the sensitive cultivar where it was independent,
and was higher in the sensitive cultivar than in the tolerant
one in both the leaves and roots. The GSH concentration in
leaves was approximately one order of magnitude higher
than that in the roots for both the cultivars. The relation-
ships between the PC and Cd concentrations in tissues were
found nonlinear. At lower Cd accumulation levels, the PC
concentrations followed an increase in the Cd accumula-
tion in both the roots and leaves, whereas at higher Cd
accumulations the relations differed between roots and
leaves. In the roots, the PC concentrations decreased with
increasing Cd accumulation, whereas the PC concentration
in the leaves followed the decrease in the Cd accumulation.
Keywords Vicia faba L. � Cd accumulation �Glutathione � Phytochelatins
Abbreviations
Cd Cadmium
CCd-leaves Cd concentration in the leaves
CCd-NS Cd concentration in the nutrient
solution
CCd-roots Cd concentration in the roots
Communicated by J. Ueda.
R. Cabala (&)
Department of Analytical Chemistry, Faculty of Science,
Charles University in Prague, Albertov 6,
128 43 Prague, Czech Republic
e-mail: [email protected]
L. Slovakova
Department of Plant Physiology, Faculty of Natural Sciences,
Comenius University in Bratislava, Bratislava, Slovak Republic
M. El Zohri
Botany Department, Faculty of Science,
Assuit University, Assuit, Egypt
H. Frank
Environmental Chemistry and Ecotoxicology,
University of Bayreuth, Bayreuth, Germany
123
Acta Physiol Plant (2011) 33:1239–1248
DOI 10.1007/s11738-010-0653-0

DW Dry weight
FW Fresh weight
GSH Glutathione, c-Glu-Cys-Gly
HPLC–ESI–MS–MS High performance liquid
chromatography electrospray-
ionization tandem mass
spectrometry
ICP-MS Inductively coupled plasma/mass
spectrometry
MTs Metallothioneins
NS Nutrient solution
PC, PCs Phytochelatin, phytochelatins
PC2 Phytochelatin 2 (c-Glu-Cys)2-Gly
PC3 Phytochelatin 3 (c-Glu-Cys)3-Gly
R Correlation coefficient
Introduction
Vicia faba beans are considered to be the main legume of
Middle Eastern countries, hence they have a potential
economic importance in many countries, especially in
Egypt. On the other hand, V. faba has proved to be very
sensitive biomonitor of water contaminants both in labo-
ratory and in situ treatments (Villalobos-Pietrini et al.
1994). Cadmium (Cd) is of special interest, as it is con-
sidered to be the fifth most toxic metal for vertebrates, and
the fourth most toxic metal for vascular plants (Jones
1939). It has been introduced into the environment mainly
through agricultural applications of Cd-contaminated
phosphate fertilizers, sewage sludge and pesticides and by
some industrial activities, such as metal mining, smelting
and founding, the use and disposal of batteries, and dis-
posal of metal-contaminated waste (Van Bruwaene et al.
1984; Cabrera et al. 1998). In addition to these anthropo-
genic emissions, Cd is also released by natural sources,
such as volcanoes and continental dusts or by weathering
of the underlying bedrock (Cabrera et al. 1998).
Plant species and even varieties differ significantly in
their resistance to Cd and in Cd accumulation capacity
(Hart et al. 1998; Zhao et al. 2002), but this is not neces-
sarily correlated with the tolerance to the metal (Greger
et al. 1991). For example, resistant and tolerant cultivars of
Silene cucubalus (Mathys 1975) and Agrostis (Mathys
1973) accumulate Cd to the same level. Cd accumulation
also differs among organs or tissues in the same plant.
Adriano (1986) reported that Cd concentrations in plant
tissues decrease in the order: old roots [ new roots [stems [ leaves [ reproductive organs.
Complexation, compartmentation and sequestration
belong to the main protective mechanisms of plants against
negative influence of heavy metals (e.g. Cd). Together with
metallothioneins (MTs), glutathione (GSH) and phyto-
chelatins (PCs), which plays an important role in detoxi-
fication of heavy metals, are members of a group of
biogenic ligands produced by plants (Meister 1995). GSH,
usually the most abundant intracellular nonprotein thiol
(tripeptide c-Glu-Cys-Gly), is enzymatically converted into
PCs, peptides of general structure (c-Glu-Cys)n-Gly,
n = 2–11, by glutamyl-cysteine dipeptidyl transpeptidase
(Beck et al. 2003). Cd2? catalyzed PC biosynthesis is
influenced, in dependence on their concentrations, by free
heavy metals and enzymes, and inhibited by Cd–PC and
Cd–GSH complexes. All the reactions form together a self-
regulating loop. The amount of PCs induced by Cd2?
seems to be roughly related to the Cd2? concentration and
the time. It is generally supposed that, within species-
specific and organ-specific limits of toxicity, the more
Cd2? is supplied, the greater induction of PCs is obtained
(Sanita di Toppi and Gabbrielli 1999). In addition,
Cd-stressed cells have to restore their physiological GSH
concentration by activating the enzyme catalyzing GSH
biosynthesis, which requires an energy supply (Ruegsegger
and Brunold 1992). Using purified recombinant PC-syn-
thase from Arabidopsis thaliana, it has been shown that the
Cd–(GSH)2 complex is a necessary substrate along with
free GSH, which explains the metal dependence of the PC
synthesis reaction mechanism (Vatamaniuk et al. 2000).
PCs belong to the group of sulfhydryl-rich peptides,
which exhibit strong metal-binding properties. Originally,
the basic physiological function of PCs has been suggested
to keep the homeostasis of heavy-metal nutrients, such as
Cu and Zn, in the cytoplasm (Steffens 1990). However,
there is currently no direct evidence that PCs have func-
tions other than in metal detoxification (Cobbett 2000). In
heavy-metal tolerance, PCs have shown various effects.
They seem to be involved in the main mechanism for high
tolerance to Cd in tomato and Arabidopsis (Howden et al.
1995). In other cases, such as S. vulgaris, the PCs seem to
have a negative influence on the metal tolerance, as the PC
production depletes the GSH in the cell, needed for pro-
tection against free radicals generated by the metal toxicity
(Ric De Voss et al. 1992). Studies on Cd-tolerant and
Cd-sensitive lines of the plant Datura innoxia, also known
as Jimsonweed, show that both lines accumulate similar
levels of PC–Cd complexes and synthesize similar
amounts of PCs at similar rates, but the onset of synthesis
of the PC–Cd complexes is more rapid in the metal-
tolerant plant line than in the metal-sensitive line (Yen
et al. 1999).
An important objective for scientific research is to select
crop species and cultivars, which are tolerant to Cd and
retain it in the roots, i.e. translocate less Cd from a con-
taminated medium to the biomass. This idea has
1240 Acta Physiol Plant (2011) 33:1239–1248
123
successfully been applied with durum wheat and sunflower
(Penner et al. 1995; Li et al. 1995). Accordingly, the aims
of this study were to investigate (a) the differences in the
Cd accumulation, the GSH and PC concentrations in bio-
mass between Cd-sensitive and Cd-tolerant cultivars of
V. faba; (b) the differences in Cd translocation patterns
within the plant between the cultivars; (c) the correlations
among the Cd, GSH and PC concentrations in the plant
tissues of both the cultivars.
Materials and methods
Reagents, chemicals and standards
Cadmium chloride (p.a.) was used as supplied (Sigma-
Aldrich, Steinheim, Germany). Bidistilled water (Buchi
F-285, Flawil, Switzerland) and all other reagents and sol-
vents were of analytical grade quality and used as received.
PC standards (lyophilized) were kindly provided by prof.
M. H. Zenk (University of Wittenberg-Halle, Germany).
Plant material and cultivation
Seeds of Cd-tolerant (Giza 40) and Cd-sensitive (Giza 2)
cultivars of broad bean (V. faba L.), obtained from the
Cairo Agricultural Research Center (Cairo, Egypt), were
germinated on moist filter paper in Petri dishes for 5 days.
The seedlings were further grown in 1/10 strength modified
Hoagland’s solution in a greenhouse with the 16/8 light
cycle (light intensity 35,000 lux) and the day/night tem-
perature of 23/18�C. When the plants were 19 days old,
they were transferred to a fresh nutrient solution containing
CdCl2 at concentrations of 0, 1, 10, 40 and 100 lmol l-1,
five replicates of each. After another 15 days, the samples
were collected: roots and leaves separated, washed with
deionized water, weighed, frozen in liquid nitrogen, and
stored at –80�C until the sample preparation.
Sample preparation, separation and quantification
A detailed description of the sample preparation, separation
and quantification method with the Cd detection limit,
0.5 lg l-1 (ICP-MS) and the limits of quantification
(LOQ), 0.2 lmol kg-1 (HPLC–ESI–MS–MS), for GSH,
PC2 and PC3 has been published in El Zohri et al. (2005).
Statistical testing
To judge rightly and exactly the statistical significance of
differences between the data compared t test at a confi-
dence level (1 - a) equal to 0.95 was applied.
Results
During the investigation, a huge multidimensional dataset
was obtained, composed of total Cd-, GSH- and PCs-con-
centrations in plant tissues and the biomass parameters
corresponding to respective Cd concentrations in the
nutrient solution. This large set of data was difficult to treat
and present in full and thus simple two-dimensional
parameter correlations are presented and discussed in the
text below.
Correlation between external and internal
Cd concentration
The general response of the broad been to the increasing
Cd concentration in the nutrient solution (CCd-NS) differs
for roots and leaves as well as for the two cultivars. The Cd
uptake into the roots and translocation into the leaves was
found time-dependent and influenced by the total Cd con-
centration in the medium.
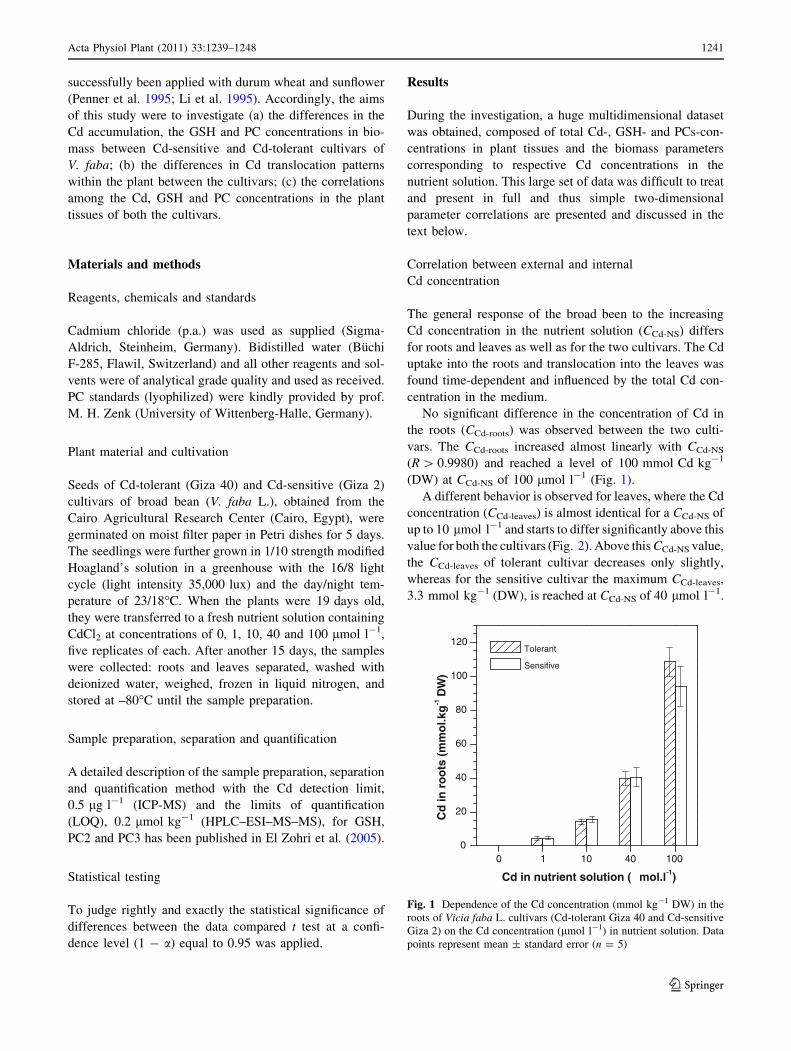
No significant difference in the concentration of Cd in
the roots (CCd-roots) was observed between the two culti-
vars. The CCd-roots increased almost linearly with CCd-NS
(R [ 0.9980) and reached a level of 100 mmol Cd kg-1
(DW) at CCd-NS of 100 lmol l-1 (Fig. 1).
A different behavior is observed for leaves, where the Cd
concentration (CCd-leaves) is almost identical for a CCd-NS of
up to 10 lmol l-1 and starts to differ significantly above this
value for both the cultivars (Fig. 2). Above this CCd-NS value,
the CCd-leaves of tolerant cultivar decreases only slightly,
whereas for the sensitive cultivar the maximum CCd-leaves,
3.3 mmol kg-1 (DW), is reached at CCd-NS of 40 lmol l-1.
0 1 10 40 1000
20
40
60
80
100
120
Cd
in r
oo
ts (
mm
ol.k
g-1 D
W)
Cd in nutrient solution (µmol.l-1)
Tolerant
Sensitive
Fig. 1 Dependence of the Cd concentration (mmol kg-1 DW) in the
roots of Vicia faba L. cultivars (Cd-tolerant Giza 40 and Cd-sensitive
Giza 2) on the Cd concentration (lmol l-1) in nutrient solution. Data
points represent mean ± standard error (n = 5)
Acta Physiol Plant (2011) 33:1239–1248 1241
123
Correlation between the Cd concentration
and biomass production
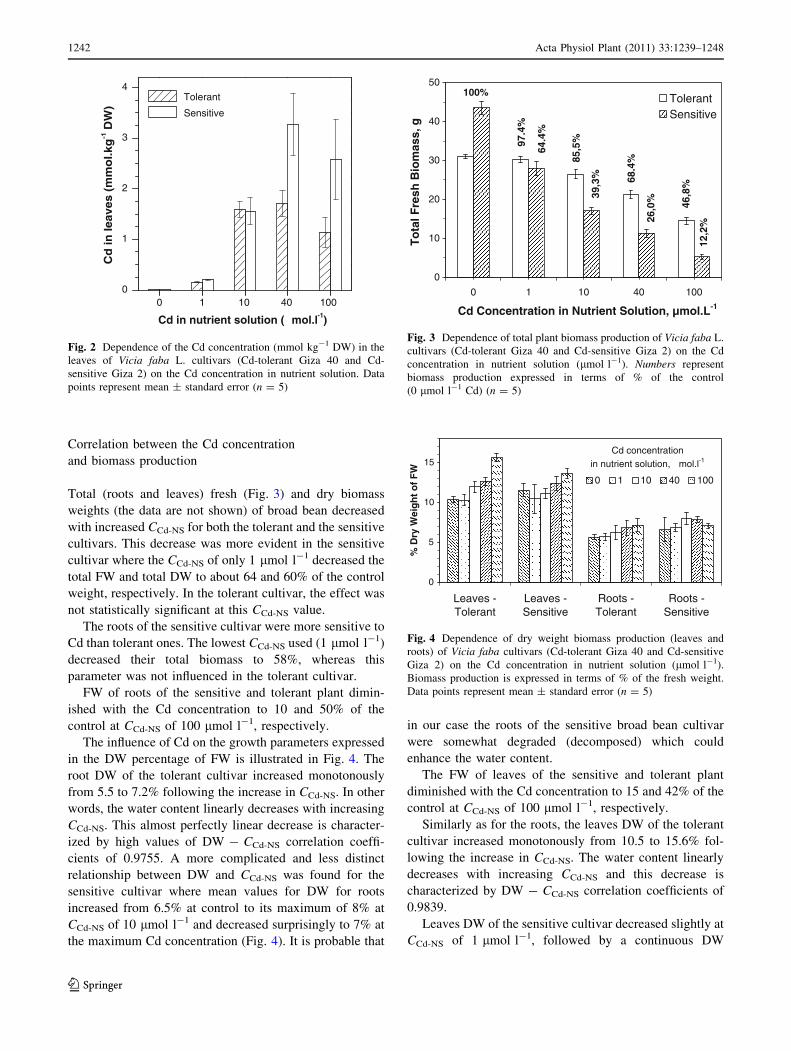
Total (roots and leaves) fresh (Fig. 3) and dry biomass
weights (the data are not shown) of broad bean decreased
with increased CCd-NS for both the tolerant and the sensitive
cultivars. This decrease was more evident in the sensitive
cultivar where the CCd-NS of only 1 lmol l-1 decreased the
total FW and total DW to about 64 and 60% of the control
weight, respectively. In the tolerant cultivar, the effect was
not statistically significant at this CCd-NS value.
The roots of the sensitive cultivar were more sensitive to
Cd than tolerant ones. The lowest CCd-NS used (1 lmol l-1)
decreased their total biomass to 58%, whereas this
parameter was not influenced in the tolerant cultivar.
FW of roots of the sensitive and tolerant plant dimin-
ished with the Cd concentration to 10 and 50% of the
control at CCd-NS of 100 lmol l-1, respectively.
The influence of Cd on the growth parameters expressed
in the DW percentage of FW is illustrated in Fig. 4. The
root DW of the tolerant cultivar increased monotonously
from 5.5 to 7.2% following the increase in CCd-NS. In other
words, the water content linearly decreases with increasing
CCd-NS. This almost perfectly linear decrease is character-
ized by high values of DW - CCd-NS correlation coeffi-
cients of 0.9755. A more complicated and less distinct
relationship between DW and CCd-NS was found for the
sensitive cultivar where mean values for DW for roots
increased from 6.5% at control to its maximum of 8% at
CCd-NS of 10 lmol l-1 and decreased surprisingly to 7% at
the maximum Cd concentration (Fig. 4). It is probable that
in our case the roots of the sensitive broad bean cultivar
were somewhat degraded (decomposed) which could
enhance the water content.
The FW of leaves of the sensitive and tolerant plant
diminished with the Cd concentration to 15 and 42% of the
control at CCd-NS of 100 lmol l-1, respectively.
Similarly as for the roots, the leaves DW of the tolerant
cultivar increased monotonously from 10.5 to 15.6% fol-
lowing the increase in CCd-NS. The water content linearly
decreases with increasing CCd-NS and this decrease is
characterized by DW - CCd-NS correlation coefficients of
0.9839.
Leaves DW of the sensitive cultivar decreased slightly at
CCd-NS of 1 lmol l-1, followed by a continuous DW
0 1 10 40 1000
1
2
3
4
Cd
in le
aves
(m
mo
l.kg
-1 D
W)
Cd in nutrient solution (µmol.l-1)
Tolerant
Sensitive
Fig. 2 Dependence of the Cd concentration (mmol kg-1 DW) in the
leaves of Vicia faba L. cultivars (Cd-tolerant Giza 40 and Cd-
sensitive Giza 2) on the Cd concentration in nutrient solution. Data
points represent mean ± standard error (n = 5)
0
10
20
30
40
50
0 1 10 40 100
Cd Concentration in Nutrient Solution, µmol.L-1
To
tal F
resh
Bio
mas
s, g
TolerantSensitive
100%
97.4
%
64.4
%
85,5
%
39,3
% 68.4
%
26,0
% 46,8
%
12,2
%
Fig. 3 Dependence of total plant biomass production of Vicia faba L.
cultivars (Cd-tolerant Giza 40 and Cd-sensitive Giza 2) on the Cd
concentration in nutrient solution (lmol l-1). Numbers represent
biomass production expressed in terms of % of the control
(0 lmol l-1 Cd) (n = 5)
0
5
10
15
Leaves -Tolerant
Leaves -Sensitive
Roots -Tolerant
Roots -Sensitive
% D
ry W
eig
ht
of
FW
0 1 10 40 100
Cd concentration in nutrient solution, µmol.l-1
Fig. 4 Dependence of dry weight biomass production (leaves and
roots) of Vicia faba cultivars (Cd-tolerant Giza 40 and Cd-sensitive
Giza 2) on the Cd concentration in nutrient solution (lmol l-1).
Biomass production is expressed in terms of % of the fresh weight.
Data points represent mean ± standard error (n = 5)
1242 Acta Physiol Plant (2011) 33:1239–1248
123

increase up to 13.6% at the highest Cd concentration which
is the opposite tendency to the roots.
This irregular correlation between the water content in
FW and CCd-NS is reflected in the correlation coefficient
values below 0.42 for both roots and leaves of the sensitive
cultivar.
Correlation between tissue concentrations
of Cd, GSH and PCs
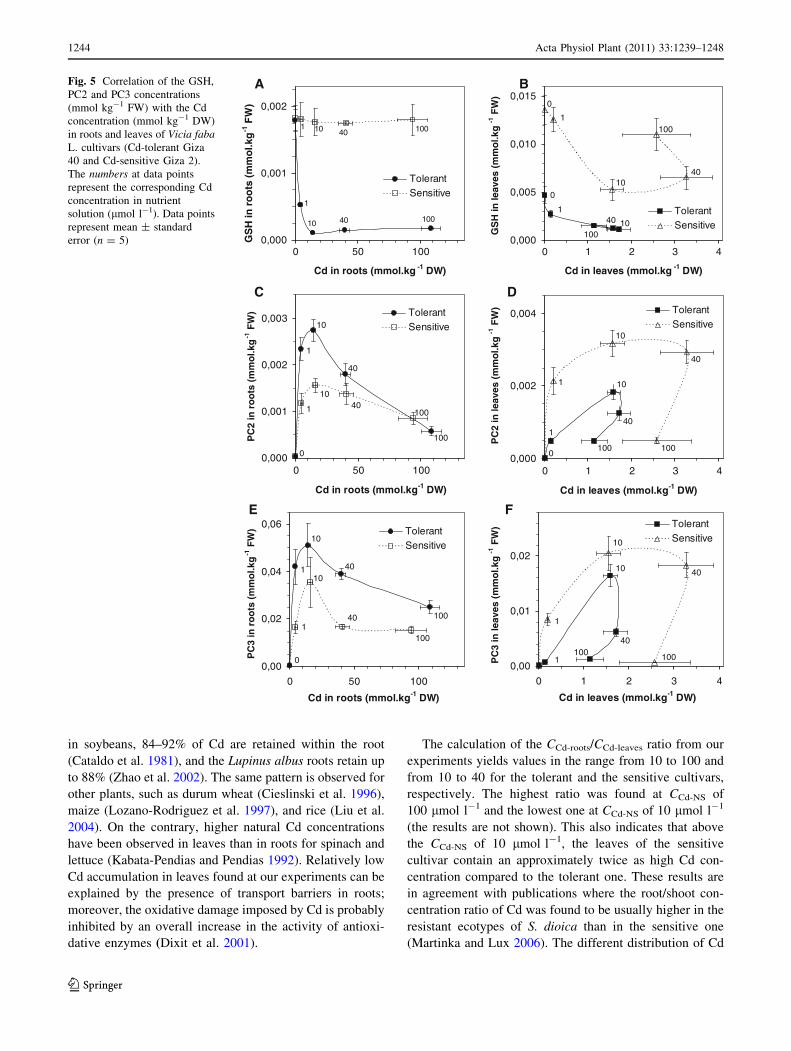
The response of the tissue GSH and PC concentrations
toward Cd accumulation was generally stronger in roots of
the tolerant cultivar and in leaves of the sensitive one.
The GSH concentration in roots of the sensitive cul-
tivar was independent on the CCd-NS (Fig. 5a) and
remained approximately at the same level as in control
broad bean plants. The GSH concentration was approxi-
mately ten times higher in roots of the sensitive cultivar
than in the tolerant one at higher Cd root concentrations.
On the contrary, a dramatic decrease in the GSH root
concentration occurred in the tolerant cultivar by appli-
cation of CCd-NS as low as 1 lmol l-1 and continued up
to CCd-NS of 10 lmol l-1. At higher Cd accumulation in
roots, the GSH level remained low and almost unchanged
(Fig. 5a).
Quite different results were observed for the leaves. The
GSH concentration in both the cultivars was generally
higher than in roots and differed between the two cultivars
even in the control plants (in all other cases, the concen-
trations of GSH and PCs were identical for the control
plants within the experimental precision). In the sensitive
cultivar, following an increase in the Cd accumulation (see
Fig. 2), the GSH concentration decreased to a minimum,
corresponding to the Cd accumulation at a CCd-NS of
10 lmol l-1 (Fig. 5b), whereas with a higher Cd accu-
mulation the GSH content was elevated but this increase
did not correspond to the amount of Cd determined in the
leaf tissues. In the tolerant cultivar, the amount of GSH in
leaves was three times lower than in the sensitive one. The
curve describing the correlation between the GSH con-
centration and the Cd tissue accumulation forms a loop
where the GSH concentration decreases continuously to its
minimum at CCd-NS of 10 lmol l-1 with increasing Cd
accumulation and remains almost unchanged at Cd accu-
mulation values corresponding to increased CCd-NS (com-
pare Figs. 2, 5b).
In both the cultivars, the production of PCs (PC2 and
PC3) was measured as the response to an increased CCd-NS
and correlated with Cd accumulation in broad bean tissues.
Application of Cd at very low concentrations induced
biosynthesis of PCs, as confirmed by the increase in the PC
concentrations from values below the limit of quantifica-
tion in control plants of both the cultivars to values above
them for the exposed plants. Careful inspection of Fig. 5
yields the following findings.
The PC concentrations in roots were higher for the tol-
erant cultivar than for the sensitive one (one exception for
CCd-NS of 100 lmol l-1 can be seen in Fig. 5c), whereas in
the leaves the relation was reciprocal. The highest PC2
concentration in leaves (Fig. 5d) is approximately one-
third lower compared to roots (Fig. 5c) for the tolerant
cultivar, whereas the highest PC2 in leaves is approxi-
mately twice as high as that in roots for the sensitive cul-
tivar. The highest PC3 concentration in leaves (Fig. 5f) is
approximately three times lower than in roots (Fig. 5e) of
the tolerant cultivar, whereas in the sensitive one it is 1.7
times lower than that in roots (opposite to PC2). This effect
corresponds to a higher Cd amount sequestered in the root
tissues as a consequence of an increased Cd concentration
in the cultivation medium.
The PC3 concentration is approximately one order of
magnitude higher than that of PC2 for leaves and roots of
both the cultivars. The courses of the dependence of PC3
concentration on the Cd accumulation follow the shape of
the courses for PC2 for roots and leaves of both the cul-
tivars, but the shapes are fundamentally different between
roots and leaves.
Discussion
Correlation between external and internal
Cd concentration
In accordance with the published results, the Cd uptake
into the roots and translocation into the leaves was found
time-dependent (Dixit et al. 2001) and influenced by the
total Cd concentration in the medium (Szalai et al. 2002).
In agreement with our results, a continuous increase in the
Cd amount in roots was also observed in maize (Szalai
et al. 2002) and wheat (Sun et al. 2005) where its con-
centration was much higher in the roots than in the leaves.
An approximately linear correlation between CCd-roots and
CCd-NS is described in this paper. Results for V. faba could
be an evidence for simple (passive) partitioning of Cd
between NS and the root tissue in V. faba. Similarly, in
Silene dioica, the young parts of roots significantly par-
ticipate in substantial uptake of Cd, as most Cd is present in
the rhizodermal cell walls (Martinka and Lux 2006).
Unfortunately, our experimental technique applied did not
allow to differentiate the Cd binding locations within the
tissues and cells. We have found that the amount of Cd in
leaves was approximately 30 times lower than in roots for
both the cultivars. This translocation pattern is observed in
some bush bean (Phaseolus vulgaris) which accumulates
Cd in the root more than in the shoot (Rodecap et al. 1981),
Acta Physiol Plant (2011) 33:1239–1248 1243
123
in soybeans, 84–92% of Cd are retained within the root
(Cataldo et al. 1981), and the Lupinus albus roots retain up
to 88% (Zhao et al. 2002). The same pattern is observed for
other plants, such as durum wheat (Cieslinski et al. 1996),
maize (Lozano-Rodriguez et al. 1997), and rice (Liu et al.
2004). On the contrary, higher natural Cd concentrations
have been observed in leaves than in roots for spinach and
lettuce (Kabata-Pendias and Pendias 1992). Relatively low
Cd accumulation in leaves found at our experiments can be
explained by the presence of transport barriers in roots;
moreover, the oxidative damage imposed by Cd is probably
inhibited by an overall increase in the activity of antioxi-
dative enzymes (Dixit et al. 2001).
The calculation of the CCd-roots/CCd-leaves ratio from our
experiments yields values in the range from 10 to 100 and
from 10 to 40 for the tolerant and the sensitive cultivars,
respectively. The highest ratio was found at CCd-NS of
100 lmol l-1 and the lowest one at CCd-NS of 10 lmol l-1
(the results are not shown). This also indicates that above
the CCd-NS of 10 lmol l-1, the leaves of the sensitive
cultivar contain an approximately twice as high Cd con-
centration compared to the tolerant one. These results are
in agreement with publications where the root/shoot con-
centration ratio of Cd was found to be usually higher in the
resistant ecotypes of S. dioica than in the sensitive one
(Martinka and Lux 2006). The different distribution of Cd
0,000
0,001
0,002
0 50 100
Cd in roots (mmol.kg -1 DW)
GS
H in
ro
ots
(m
mo
l.kg
-1 F
W)
TolerantSensitive
100
100
40
40
10
10
1
1
0,000
0,001
0,002
0,003
0 50 100
Cd in roots (mmol.kg-1 DW)
PC
2 in
ro
ots
(m
mo
l.kg
-1 F
W) Tolerant
Sensitive
0
100
10040
40
10
10
1
1
0,00
0,02
0,04
0,06
0 50 100
Cd in roots (mmol.kg-1 DW)
PC
3 in
ro
ots
(m
mo
l.kg
-1 F
W) Tolerant
Sensitive
100
100
40
40
10
101
1
0
0,000
0,005
0,010
0,015
0 1 2 3 4
Cd in leaves (mmol.kg -1 DW)
GS
H in
leav
es (
mm
ol.k
g-1
FW
)
TolerantSensitive
100
100
40
40
10
10
1
1
0
0
0,000
0,002
0,004
0 1 2 3 4
Cd in leaves (mmol.kg-1 DW)P
C2
in le
aves
(m
mo
l.kg
-1 F
W) Tolerant
Sensitive
100100
40
40
10
101
1
0
0,00
0,01
0,02
0 1 2 3 4
Cd in leaves (mmol.kg-1 DW)
PC
3 in
leav
es (
mm
ol.k
g-1
FW
) TolerantSensitive
100100
40
40
10
10
1
1
A B
C D
F E
Fig. 5 Correlation of the GSH,
PC2 and PC3 concentrations
(mmol kg-1 FW) with the Cd
concentration (mmol kg-1 DW)
in roots and leaves of Vicia fabaL. cultivars (Cd-tolerant Giza
40 and Cd-sensitive Giza 2).
The numbers at data points
represent the corresponding Cd
concentration in nutrient
solution (lmol l-1). Data points
represent mean ± standard
error (n = 5)
1244 Acta Physiol Plant (2011) 33:1239–1248
123

among the tissues could explain the differences in the
sensitivity to this metal (Lozano-Rodriguez et al. 1997).
Correlation between the Cd concentration
and biomass production
It was published that the Cd-treated plants showed lower
solute potential values, which resulted in higher turgor
potential values. This observation may suggest that Cd
decreases cell wall extensibility, which may cause a
reduced cell expansion (Poschenrieder et al. 1989). The
irregular correlation between the water content in FW and
CCd-NS found at our plants is inconsistent with most pub-
lished results. Indeed, in agreement with our results,
a longer cultivation of roots of sensitive Mung bean in
Cd-medium caused degradation processes and desiccation
(Slovakova et al. 2007). Similarly, it was published that
higher Cd concentrations caused growth retardation and the
roots appeared to be darker than the control lettuce plants,
perhaps due to root death (Maier et al. 2003). Roots of Cd-
tolerant pea mutant developed and growth by Cd concen-
trations by which root growth of wild-type plants was
completely inhibited (Tsyganov et al. 2007). To increased
Cd concentration in S. vulgaris, the twice as high Cd
concentration was necessary to inhibit the growth of tol-
erant plant roots compared to the roots of the sensitive one
(Knecht et al. 1994). Exposure to Cd reduced transpiration
in Brassica juncea and this correlated with reduced leaf
expansion growth whereas the photosynthesis was not
affected (Haag-Kerwer et al. 1999). Disturbances in
physiological functions may also be caused by malfunc-
tioning of plasma membrane. A longer exposure of a pea
plant to Cd was associated with increased degree of un-
saturation of the fatty acids, causing changes in the root
plasma membrane phospholipid composition and fluidity
(Hernandez and Cooke 1997).
Correlation between tissue concentrations
of Cd, GSH and PCs
In many works about molecular response of plant to Cd
exposure is presented that legumes have the unique
capacity to synthesize in higher ratio homoglutathione
(hGSH) and homophytochelatins (hPCs) than GSH and
PCs. This was reported for Vigna angularis (Oven et al.
2001) and for soybean (Vazqez et al. 2009). But in other
kind of legumes such as pea and white lupin (Vazqez et al.
2009) the prevalent type were PCs synthesized form GSH.
Works with Lotus japonicus (Loscos et al. 2006; Ramos
et al. 2007) indicate that synthesis of both kind of hGSH
and hPCs as well as GSH and PCs is also tissue specific
and Lotus is able to synthesize both of them. The HPLC–
ESI–MS–MS analytical method used for determination of
GSH and PCs (El Zohri et al. 2005) proved absence of
hGSH and hPCs in V. faba. The possible oxidation of GSH
and PCs in the sample was studied and discussed in the
same article. Briefly, in plant extracts prepared with addi-
tion of DTT at the very beginning of the extraction, GSH
and PCs were completely reduced, whereas in its absence
they were present predominantly in the oxidized form. This
is especially important for V. faba homogenates which are
known to contain high concentrations of redox-active
polyphenols, oxidative enzymes, and transition metals, all
of which promote oxidation during sample preparation.
Our results show that the response of GSH concentration
in roots to Cd is in agreement with the generally accepted
opinion that the formation of a Cd–PC complex reduces the
free Cd concentration in cytosol and could lead to a
decrease in the GSH content (Lozano-Rodriguez et al.
1997). Similarly, it has been published that the GSH con-
tent of CdCl2-treated cells of Mesorhisobium huakui,
which express gene for PC-synthase of A. thaliana,
decreased, whereas the GSH content in untreated cells was
higher (Sriprang et al. 2003). On the other hand, in the
sensitive cultivar the increased amount of GSH at higher
Cd content could result either from diminished PC syn-
thesis or from the enhanced synthesis of GSH as published
for poplar transformants with an augmented PC synthesis
(Noctor et al. 1996, 1998). Nonlinear relationships among
CCd-NS, GSH and Cd tissue concentrations are described in
this paper. Results point to relatively complicated feed-
back-controlled processes of production, regulation and
accumulation of the species of interest.
The differences in the PC content, found in our work,
clearly demonstrate that the tolerant cultivar could block
the Cd influx at the first barrier more effectively, in the
roots, whereas PCs probably bind Cd ions predominantly in
the leaves of the sensitive cultivar. As reported by Liu et al.
(2007), Cd resistance in both Brassica chinensis L. and
Brassica pekinensis (Lour.) Ropr. is based on an exclusion
mechanism in which the roots accumulate the metal ions
and then prevent the Cd translocation to the shoot. A
similar metal accumulation pattern has been published for
wheat. The PC concentrations in the stem and roots of
wheat were increased whereas no distinct difference was
found in the leaves and the PC synthesis was closely linked
to metal uptake and accumulation (Sun et al. 2005). Also,
tobacco plants overexpressing of Arabidopsis PC-synthase
gene showed higher content of PCs than controls and it was
higher in roots than in shoots (Pomponi et al. 2006). We
have found that in the roots the PC concentrations increase
up to their maximum at the Cd accumulation level corre-
sponding to CCd-NS of 10 lmol l-1 and decrease even at
increasing Cd accumulation. This behavior could be
explained by the limited PCs production capacity and/or
GSH depletion (compare Fig. 5a, c, e) which is more
Acta Physiol Plant (2011) 33:1239–1248 1245
123

apparent for the sensitive cultivar. The concentrations of
PCs in the leaves do not increase so steeply with increasing
Cd accumulation and beyond its maximum, at the Cd
accumulation level corresponding to CCd-NS equal to
10 lmol l-1, decreases sharply following only moderate
reduction in the Cd accumulation. This effect could be an
evidence of a Cd removal mechanism or a Cd input barrier
which is more effective in the tolerant cultivar than in the
sensitive one. Another evidence for a more effective Cd
removal mechanism in leaves follows from the fact that the
Cd accumulation level is substantially lower in the tolerant
cultivar above CCd-NS of 10 lmol l-1.
The fact that the highest PC concentrations are found at
the same levels of the Cd accumulation in respective tis-
sues corresponding to CCd-NS of 10 lmol l-1 could result
from the activity of a similar Cd-controlled feedback
mechanism of PC production and GSH level regulation in
both the cultivars.
Our measurements have shown that the PC3 concen-
tration was approximately one order of magnitude higher
than that of PC2 for leaves and roots of both the cultivars.
Synthesis of higher concentration of higher molecular
weight PC3 than PC2 was recorded also by Beraud et al.
(2007) as reaction of V. faba plants to the increased Cd
concentration exposure. Oven et al. (2001) has reported
about Cd-induced synthesis of the PCs in Azuki bean
where the PC2 concentration increased 15-fold upon
exposure to the Cd ions. PC(n ? 1)/PC(n) ratio is species
specific and depends on the Cd accumulation as can be
found in the literature (for Phragmites australis see Ederli
et al. 2004, for Salix viminalis Landberg and Greger 2004,
for Cuscuta reflexa Srivastava et al. 2004, for S. vulgaris
Knecht et al. 1994, for barley genotypes with different
tolerance to Cd Persson et al. 2006, for B. juncea Haag-
Kerwer et al. 1999).
In our experiments a comparatively good correlation
was found between PC2 and PC3 concentrations in leaves
of the sensitive cultivar (R2 = 0.9455, compare Fig. 5d
and f) and in roots of the tolerant cultivar (R2 = 0.9082,
compare Fig. 5c and e), whereas correlations were mod-
erate in leaves of the tolerant cultivar (R2 = 0.8907) and
roots of the sensitive cultivar (R2 = 0.7863). It is inter-
esting that the correlation is better in roots of the tolerant
cultivar where the PC concentrations are higher compared
to those in the sensitive cultivar and these relations are
opposite in the leaves. The correlations between GSH and
PC concentrations were only moderate and the corre-
sponding R2 are in the range from 0.4285 (GSH-PC2 in the
roots of the tolerant plants) to 0.8182 (GSH-PC3 in the
leaves of sensitive cultivar). The absence of a straightfor-
ward correlation between the GSH and PC concentrations
is a sign of a more complicated reaction mechanism of the
PC production from the GSH precursor.
As can be concluded from these findings, the tolerant
broad bean cultivar Giza 40 has a better ability to sequester
higher Cd amounts in roots which is confirmed by a higher
production of both PC2 and PC3 compared to the sensitive
cultivar Giza 2. The higher PCs production in leaves of the
Giza 2 cultivar indicate that much more of free Cd can
penetrate into the leaves and can negatively influence the
metabolic processes, e.g., photosynthesis (Gorinova et al.
2007), which consequently affects the whole plant metab-
olism and growth. As the next step of the investigation, it
would be necessary to study Cd accumulation and trans-
location, together with the GSH and PC tissue concentra-
tions, during the whole life cycle of V. faba, from the
germination period until the bean-fruit production.
In conclusion, V. faba accumulates more than 95% of Cd
in the roots and only less than 4% in the leaves. The differ-
ences between the cultivars were statistically apparent only
at higher levels of Cd accumulation. The CCd-leaves was found
to be nonlinearly dependent on CCd-NS, ranging from 0.01 up
to 1.71 and from 0.01 to 3.27 mmol kg-1 DW for tolerant
and sensitive cultivars, respectively. The differences
between the cultivars in Cd accumulation were apparent at
higher CCd-NS. On the contrary, in the roots, the CCd-roots
depended linearly on CCd-NS and was found in the range from
0 to 93.9 and 0 to 108.5 mmol kg-1 DW for tolerant and
sensitive cultivar, respectively. The difference between the
cultivars in the Cd accumulation in roots was only statisti-
cally significant with the highest CCd-NS. The production of
biomass and its water content decreased nonlinearly with
increasing Cd accumulation and this decrease was several
times steeper for the sensitive cultivar. The biomass weight
reached only 12.2 and 46.8% of the control for sensitive
and tolerant cultivar, respectively, at CCd-NS equal to
100 lmol l-1 Cd2?. The GSH concentration did not linearly
correlate with Cd accumulation, except for the roots of the
sensitive cultivar where the GSH concentration was almost
constant and equal to 0.0018 mmol kg-1 FW. The GSH
concentration was higher in the sensitive cultivar than that in
the tolerant one in both the leaves and roots. The GSH con-
centrations were in the range of 0.0002–0.0018 mmol kg-1
FW for roots, of 0.0001–0.005 mmol kg-1 FW and of
0.0001–0.0135 mmol kg-1 FW for leaves of tolerant and
sensitive cultivars, respectively. The relationships between
the PC and Cd concentrations in tissues were found nonlin-
ear. The concentrations of PC2 and PC3 were found in ranges
of 0–0.003 mmol kg-1 FW and 0–0.035 mmol kg-1 FW,
respectively. For roots, the PC concentrations were higher in
tolerant cultivars compared to the sensitive one, whereas the
relation was opposite in leaves (except at the highest Cd
accumulation where the difference in the PC concentrations
between the cultivars was negligible). The highest PC con-
centrations were found at CCd-NS of 10 lmol l-1. The rela-
tions among the Cd accumulation and GSH and PC tissue
1246 Acta Physiol Plant (2011) 33:1239–1248
123

contents were found nonlinear and relatively complex,
indicating the presence of some feedback control system in
the two cultivars.
Acknowledgments R. Cabala would like to acknowledge the
research project VZ MSM 0021620857 from the Ministry of Educa-
tion, Youth and Sports of the Czech Republic. L’. Slovakova grate-
fully acknowledges a partial financial support from the grants of the
Slovak grant agencies VEGA No. 1/4354/07, COST 0004-06 APVV.
References
Adriano DC (1986) Trace elements in the terrestrial environment.
Springer, New York
Beck A, Lendzian K, Oven M, Christmann A, Grill E (2003)
Phytochelatin synthase catalyzes key step in turnover of
glutathione conjugates. Phytochemistry 62:423–431
Beraud E, Cotelle S, Leroy P, Ferard JF (2007) Genotoxic effects and
induction of phytochelatins in the presence of Cd in Vicia fabaroots. Mutat Res 633:112–116
Cabrera C, Ortega E, Lorenzo ML, Lopez MC (1998) Cd contam-
ination of vegetable crops, farmlands, and irrigation waters. Rev
Environ Contam Toxicol 154:55–81
Cataldo DA, Garland TR, Wildung RE (1981) Cd uptake kinetics in
intact soybean plants. Plant Physiol 68:835–839
Cieslinski G, Nielsen GH, Hogue EJ (1996) Low-molecular-weight
organic acids in rhizosphere soils of durum wheat and their
effect on Cd bioaccumulation. Plant Soil 180:267–276
Cobbett CS (2000) Phytochelatins and their roles in heavy metal
detoxification. Plant Physiol 123:825–832
Dixit V, Pandey V, Shyam R (2001) Differential antioxidative
responses to Cd in roots and leaves of pea (Pisum sativum L. cv.
Azad). J Exp Bot 52:1101–1109
Ederli L, Reale L, Ferranti F, Pasqualini S (2004) Responses induced
by high concentration of Cd in Phragmites australis roots.
Physiol Plant 121:66–74
El Zohri M, Cabala R, Frank H (2005) Quantification of phytochel-
atins in plants by reversed-phase HPLC–ESI–MS–MS. Anal
Bioanal Chem 382:1871–1876
Gorinova N, Nedkovska M, Todorovska E, Simova-Stoilova L,
Stoyanova Z, Georgieva K, Demirevska-Kepova K, Atanasov A,
Herzig R (2007) Improved phytoaccumulation of Cd by
genetically modified tobacco plants (Nicotiana tabacum L.).
Physiological and biochemical response of the transformants to
Cd toxicity. Environ Pollut 145:161–170
Greger M, Brammer E, Lindberg S, Larsson G, Idestam-Almquist J
(1991) Uptake and physiological effects of Cd in sugar beet
(Beta vulgaris) related to mineral provision. J Exp Bot
42:729–737
Haag-Kerwer A, Schafe HJ, Heiss S, Walter C, Rausch T (1999) Cd
exposure in Brassica juncea caused a decline in transpiration
rate and leaf expansion without effect on photosynthesis. J Exp
Bot 50:1827–1835
Hart JJ, Welch RM, Norvell WA, Sullivan LA, Kochian LV (1998)
Characterization of Cd binding, uptake, and translocation in
intact seedlings of bread and durum wheat cultivars. Plant
Physiol 116:1413–1420
Hernandez LE, Cooke DT (1997) Modification of the root plasma
membrane lipid composition of Cd treated Pisum sativum. J Exp
Bot 48:1375–1381
Howden R, Goldsborough PB, Andersen CD, Cobett CS (1995) Cd-
sensitive, cad1 mutants of Arabidopsis thaliana are phytochel-
atin deficient. Plant Physiol 107:1059–1066
Jones JRE (1939) The relation between the electrolytic solution
pressures of the metals and their toxicity to the stickleback
(Gasterosteus aculeatus L.). J Exp Biol 16:425–437
Kabata-Pendias A, Pendias H (1992) Trace elements in soils and
plants, 2nd edn. CRC Press, Boca Raton
Knecht JA, van Dillen M, Koevoets PLM, Schat H, Verkeij JAC,
Ernst WHO (1994) Phytochelatins in Cd-sensitive and Cd-
tolerant Silene vulgaris. Plant Physiol 104:255–261
Landberg T, Greger M (2004) No phytochelatin (PC2 and PC3)
detected in Salix viminalis. Physiol Plant 121:481–487
Li YM, Chaney RL, Schneiter AA, Miller JF (1995) Screening for
low grain Cd phenotypes in sunflower, durum wheat and flax.
Crop Sci 35:137–141
Liu WJ, Zhu YG, Smith FA, Smith SE (2004) Do iron plaque and
genotypes affect arsenate uptake and translocation by rice
seedlings (Oryza sativa L.) grown in solution culture? J Exp Bot
55:1707–1713
Liu CP, Shen ZG, Li XD (2007) Accumulation and detoxification of
Cd in Brassica pekinensis and B. chinensis. Biol Plant
51:116–120
Loscos J, Naya L, Ramos J, Clemente MR, Matamoros MA, Becana
M (2006) A reassessment of substrate specificity and activation
of phytochelatin synthases from model plants by physiologically
relevant metals. Plant Physiol 140:1213–1221
Lozano-Rodriguez E, Hernandez LE, Bonay P, Carpena-Riuz RO
(1997) Distribution of Cd in shoot and root tissues of maize and
pea plants: physiological disturbances. J Exp Bot 48:123–128
Maier EA, Matthews RD, McDowell JA, Walden RR, Ahner BA
(2003) Environmental Cd levels increase phytochelatin and
glutathione in lettuce grown in a chelator-buffered nutrient
solution. J Environ Qual 32:1356–1364
Martinka M, Lux A (2006) Intraspecific variation of Silene dioica L.
in uptake and translocation of Cd related to endodermal
development. In: Teixeira da Silva JA (ed) Floriculture,
ornamental and plant biotechnology. Advances and topical
issues, vol III, 1st ed. edn. GSB, UK, pp 312–316
Mathys W (1973) Vergleichende Untersuchungen der Zinkaufnahme
von resistent und sensitiven Populationen von Agrostis tenuisSibth. Flora Jena 162:492–499
Mathys W (1975) Enzymes of heavy metal-resistant and nonresistant
populations of Silene cucubalus and their interactions with some
heavy metals in vitro and in vivo. Physiol Plant 33:161–165
Meister A (1995) Glutathione biosynthesis and its inhibition. Method
Enzymol 251:3–7
Noctor G, Strohm M, Jouanin L, Kunert KJ, Foyer CH, Rennenberg H
(1996) Synthesis of glutathione in leaves of transgenic poplar
overexpressing c-glutamylcysteine synthetase. Plant Physiol
11:1071–1078
Noctor G, Arisi ACM, Jouanin L, Kunert KJ, Rennenberg H, Foyer
CH (1998) Glutathione: biosynthesis, metabolism and relation-
ship to stress tolerance explored in transformed plants. J Exp Bot
49:623–647
Oven M, Raith K, Neubert RHH, Kutchan TM, Zenk MH (2001)
Homo-phytochelatins are synthesized in response to Cd in azuky
beans. Plant Physiol 126:1275–1280
Penner GA, Clark J, Bezte LZ, Lisle D (1995) Identification of RAPD
markers linked to a gene governing Cd uptake in durum wheat.
Genome 38:543–547
Persson DP, Hansen TH, Holm PE, Schjoerring JK, Hansen HCB,
Nielsen J, Cakmak I, Husted S (2006) Multi-elemental speciation
analysis of barley genotypes differing in tolerance to Cd toxicity
using SEC-ICP-MS and ESI-TOF-MS. J Anal Atom Spectrom
21:996–1005
Pomponi M, Censi V, Di Girolamo V, De Paolis A, Sanita di Toppi L,
Aromolo R, Costantino P, Cardarelli M (2006) Overexpression
of Arabidopsis phytochelatin synthase in tobaco plants enhances
Acta Physiol Plant (2011) 33:1239–1248 1247
123

Cd2? tolerance and accumulation but not translocation to the
shoot. Planta 223:180–190
Poschenrieder C, Gunse B, Barcelo J (1989) Influence of Cd on water
relations, stomatal resistance, and abscisic acid content in
expanding bean leaves. J Plant Physiol 90:1365–1371
Ramos J, Clemente MR, Naya L, Loscos J, Perez-Rentome C, Sato S,
Tabata S, Becana M (2007) Phytochelatin synthases of the model
legume Lotus japonicus. A small multigene family with
differential response to Cd and alternatively spliced variants.
Plant Physiol 143:1110–1118
Ric De Voss CHR, Vonk MJ, Vooijs R, Schat H (1992) Glutathione
depletion due to copper-induced phytochelatin synthesis causes
oxidative stress in Silene cucubalus. Plant Physiol 98:853–858
Rodecap KD, Tingey DT, Tibbs JH (1981) Cd-induced ethylene
production in bean plant. Z Pflanzenphysiol 105:65–74
Ruegsegger A, Brunold C (1992) Effect of Cd on ‘y-glutamylcysteine
synthesis in maize seedlings. Plant Physiol 99:428–433
Sanita di Toppi L, Gabbrielli R (1999) Response to Cd in higher
plants. Environ Exp Bot 41:105–130
Slovakova L, Klecova-Simonova E, Henselova M, Hudak J (2007)
Antioxidant enzymes of tolerant (Brassica juncea L.) and
susceptible [Vigna radiata (L.) Wilczek] plants to Cd. In: Blaha
L (ed) Influence of abiotic and biotic stressors to property of
plants 2007. Proceedings VURV v.v.i., Prague-Ruzyne,
pp 355–362
Sriprang R, Hayashi M, Ono H, Takagi M, Hirata K, Murooka Y
(2003) Enhanced accumulation of Cd2? by a Mesorhizobium sp.
transformed with a gene from Arabidopsis thaliana coding for
phytochelatin synthase. Appl Environ Microbiol 69:1791–1796
Srivastava S, Tripathi RD, Dwivedi UN (2004) Synthesis of
phytochelatins and modulation of antioxidants in response to
Cd stress in Cuscuta reflexa—an angiospermic parasite. J Plant
Physiol 161:665–674
Steffens JC (1990) The heavy metal-binding peptides of plants. Annu
Rev Plant Mol Biol 41:553–575
Sun Q, Wang XR, Ding SM, Yuan XF (2005) Effects of interaction
between Cd and plumbum on phytochelatins and glutathione
production in wheat (Triticum aestivum L.). J Integr Plant Biol
(Acta Bot Sin) 47:435–442
Szalai G, Janda T, Galan-Goldhirsh A, Paldi E (2002) Effect of Cd
treatment on phytochelatin synthesis in maize. Acta Biol Szeged
46:121–122
Tsyganov VE, Belimov AA, Borisov AY, Safronova VI, Georgi M,
Dietz KJ, Tikhonovich IA (2007) A chemically induced new pea
(Pisum sativum) mutant SGECdt with increased tolerance to, and
accumulation of, Cd. Ann Bot 99:227–237
Van Bruwaene R, Kirchmann R, Impens R (1984) Cd contamination
in agriculture and zootechnology. Experientia 40:43–52
Vatamaniuk OK, Mari S, Lu YP, Rea PA (2000) Mechanism of heavy
metal ion activation of phytochelatin (PC) synthase. J Biol Chem
275:31451–31459
Vazqez S, Goldsbrough P, Carpena RO (2009) Comparative analysis
of the contribution of phytochelatins to Cd and arsenic tolerance
in soybean and white lupin. Plant Physiol Biochem 47:63–67
Villalobos-Pietrini R, Flores-Marquez AR, Gomez-Arroyo S (1994)
Cytogenetic effects in Vicia faba of the polluted water from
rivers of the Tlaxcala hydrological system, Mexico. Rev Int
Contam Ambient 10:83–88
Yen TY, Villa JA, DeWitt JG (1999) Analysis of phytochelatin–Cd
complexes from plant tissue culture using nano-electrospray
ionization tandem mass spectrometry and capillary liquid
chromatography/electrospray ionization tandem mass spectrom-
etry. J Mass Spectrom 34:930–941
Zhao FJ, Hamon RE, Lombi E, McLaughlin MJ, McGrath SP (2002)
Characteristics of Cd uptake in two contrasting ecotypes of the
hyperaccumulator Thlaspi caerulescens. J Exp Bot 53:535–543
1248 Acta Physiol Plant (2011) 33:1239–1248
123