a genomics approach to the early stages of triterpene saponin biosynthesis in medicago truncatula
TRANSCRIPT

A genomics approach to the early stages of triterpenesaponin biosynthesis in Medicago truncatula
Hideyuki Suzuki1,�, Lahoucine Achnine1,�, Ran Xu2, Seiichi P. T. Matsuda2 and Richard A. Dixon1,�
1Plant Biology Division, The Samuel Roberts Noble Foundation, 2510 Sam Noble Parkway, Ardmore, OK 73401, and2Departments of Chemistry and Biochemistry and Cell Biology, Rice University, Houston, TX 77005, USA
Received 11 July 2002; revised 20 September 2002; accepted 27 September 2002.�For correspondence (fax þ1 580 224 6692; e-mail [email protected]).�These authors contributed equally to this work.
Summary
The saponins of the model legume Medicago truncatula are glycosides of at least five different triterpene
aglycones: soyasapogenol B, soyasapogenol E, medicagenic acid, hederagenin and bayogenin. These agly-
cones are most likely derived from b-amyrin, a product of the cyclization of 2,3-oxidosqualene. Mining
M. truncatula EST data sets led to the identification of sequences putatively encoding three early enzymes
of triterpene aglycone formation: squalene synthase (SS), squalene epoxidase (SE), and b-amyrin synthase
(b-AS). SS was functionally characterized by expression in Escherichia coli, two forms of SE by comple-
mentation of the yeast erg1 mutant, and b-AS by expression in yeast. b-Amyrin was the sole product of the
cyclization of squalene epoxide by the recombinant M. truncatula b-AS, as judged by GC–MS and NMR.
Transcripts encoding b-AS, SS and one form of SE were strongly and co-ordinately induced, associated with
accumulation of triterpenes, upon exposure of M. truncatula cell suspension cultures to methyl jasmonate.
Sterol composition remained unaffected by jasmonate treatment. Molecular verification of induction of the
triterpene pathway in a cell culture system provides a new tool for saponin pathway gene discovery by DNA
array-based approaches.
Keywords: triterpene, sterol, saponin, methyl jasmonate, elicitation, functional expression.
Introduction
Triterpene glycoside saponins are attracting increasing
interest in view of their multiple biological activities. These
both positively and negatively impact plant traits. Thus,
whereas some saponins display allelopathic (Waller et al.,
1993), antimicrobial (Nagata et al., 1985; Osbourn, 1996;
Papadopoulou et al., 1999), and anti-insect (Pedersen et al.,
1976; Tava and Odoardi, 1997) activity, they can also be
toxic to monogastric animals, act as antipalatability factors,
or negatively impact forage digestibility in ruminants
(Cheeke, 1976; Oleszek, 1997; Oleszek et al., 1999; Small,
1996). Other saponins have potentially useful pharmacol-
ogical activities, including anticholesterolemic (Cheeke,
1976), anticancer (Haridas et al., 2001; Park et al., 2001),
adjuvant (Behboudi et al., 1999; Marciani et al., 2000), and
hemolytic (Jones and Elliott, 1969) activity. Despite the
obvious interest in facilitating or inhibiting production of
triterpene saponins for crop improvement or develop-
ment of pharmacological agents, most of the steps in their
biosynthesis remain uncharacterized at the molecular
level.
We have chosen the model legume Medicago truncatula
as a suitable species for a functional genomic approach to
triterpene saponin biosynthesis in view of the availability of
extensive EST resources (Bell et al., 2001) and the interest-
ing saponin profile of this species (Huhman and Sumner,
2002). Metabolic profiling of M. truncatula roots using
reverse-phase HPLC and electrospray ionization mass spec-
trometry showed the presence of a more complex mixture
of triterpenes than found in the closely related and pre-
viously well-studied species alfalfa (M. sativa) (Massiot
et al., 1988; Oleszek and Jurzysta, 1990; Oleszek et al.,
1992; Tava et al., 1993). Five different triterpene aglycones,
soyasapogenol B, soyasapogenol E, medicagenic acid,
hederagenin, and bayogenin were found to be the core
of the 37 M. truncatula saponins identified (Huhman and
Sumner, 2002). All these aglycones are most likely derived
from b-amyrin, the initial product of cyclization of 2,3-oxi-
dosqualene, as shown in Figure 1.
To utilize the genomic resources of M. truncatula for
gene discovery in the saponin pathway, it is necessary to
The Plant Journal (2002) 32, 1033–1048
� 2002 Blackwell Publishing Ltd 1033

Figure 1. The biosynthesis of b-amyrin and cycloartenol, and the involvement of cytochrome P450 and glycosyltransferase enzymes in the biosynthesis of the triterpene aglycones and selected conjugates found inMedicago truncatula.
1034
Hid
eyu
kiS
uzu
kiet
al.
�B
lackw
ell
Pu
blish
ing
Ltd
,T
he
Pla
nt
Jo
urn
al,
(2002),
32,
1033–1
048

functionally characterize one or more saponin biosynthetic
genes prior to DNA array-based approaches. The first com-
mitted step in triterpene biosynthesis in Medicago is cata-
lyzed by a specific oxidosqualene cyclase (OSC), b-amyrin
synthase (b-AS). In higher plants, oxidosqualene is a pre-
cursor common to the biosynthesis of both steroids and
triterpenoids (Abe et al., 1993). In sterol biosynthesis in
animals and fungi, the cyclization of 2,3-oxidosqualene
leads to the formation of lanosterol, whereas cycloartenol
is the first cyclized sterol precursor in plants. b-AS has been
functionally characterized from Panax ginseng (Kushiro
et al., 1998), pea (Morita et al., 2000) and Arabidopsis
thaliana (Husselstein-Muller et al., 2001), and is closely
related to plant cycloartenol synthase, which has also been
cloned and functionally characterized (Corey et al., 1993;
Hayashi et al., 2000). Surprisingly, a recently characterized
monocot b-AS from oat is phylogenetically distinct from
dicot b-AS enzymes (Haralampidis et al., 2001). b-AS may
produce one or more products from the cyclization of 2,3-
oxidosqualene, depending on the plant source (Abe et al.,
1993; Husselstein-Muller et al., 2001; Kushiro et al., 1998,
2000).
The two enzymes preceding OSC, namely squalene
synthase (SS) and squalene epoxidase (SE), have been well
characterized in mammals and yeast (Jandrositz et al.,
1991; Laden et al., 2000; Lee et al., 2000; Pandit et al.,
2000). SS has been functionally characterized from Arabi-
dopsis (Kribii et al., 1997; Nakashima et al., 1995). However,
although plant SE genes have been annotated based upon
sequence similarity to the mammalian and yeast enzymes
(Schafer et al., 1999), plant SE has not been functionally
characterized. SE is membrane associated, requires NADPH
cytochrome P450 reductase and, in mammals, additional
soluble protein factors for its activity (Laden et al., 2000;
Shibata et al., 2001). It is not known whether specific forms
of SS and SE might be differentially associated with sterol
and triterpene biosynthesis in plants.
We here describe an EST data mining approach for iso-
lation of candidate SS, SE and b-AS genes from M. trunca-
tula, and the functional identification of these genes by
heterologous expression in E. coli or yeast. The corre-
sponding cDNA sequences have been used as probes for
development of an inducible cell culture system for triter-
pene pathway gene discovery.
Results
Identification of early triterpene pathway genes in
M. truncatula by mining EST data sets
Candidate ESTs for the first steps of the saponin biosyn-
thetic pathway in M. truncatula were identified by mining
publicly available EST data sets representing cDNA
libraries from a variety of different organs and biotic/
abiotic treatments (Table 1). To obtain full-length mRNA
sequences for the putative SS, SE and b-AS genes, we
retrieved and analyzed EST clones found in cDNA libraries
from M. truncatula roots, nodulated roots, stems, cell sus-
pension cultures and leaves, and from some of the above
tissues following treatments such as insect damage, elici-
tation with yeast extract, drought, or phosphate starvation.
In the TIGR M. truncatula Gene Index (MtGI) database
(Quackenbush et al., 2000; http://www.tigr.org/tdb/mtgi/),
the putative SS ESTs were clustered into one tentative
consensus, whereas putative SE and b-AS ESTs were each
clustered into three tentative consensuses (Table 1). Two
SE genes, SE1 and SE2, are each present in a single copy
in the M. truncatula genome, as shown by DNA gel blot
analysis in Figure 2(b,c). Neither SE1 nor SE2 has restriction
Table 1 Medicago truncatula EST clones and tentative consensus (TC) sequences annotated as squalene synthase (SS), squaleneepoxidase (SE), and b-amyrin synthase (b-AS)
Gene nameTentative consensus (TC)sequence numbers
Number of EST clones per TC (number of full-length clones)[library distribution (number of clones per library)]
Squalene synthase (SS) �TC35874 12 (7) [a(3), b(2), e(1), f(2), �g(4)]�TC28416 7 (4) [�c(1), d(1), f(1), g(2), h(1), l(1)]
Squalene epoxidase (SE) TC29021 3 (0) [c(1), g(2)]�TC37711 2 (1) [�f(2)]TC28833 3 (0) [e(1), j(2)]
b-amyrin synthase (b-AS) TC28834 2 (0) [b(1), c(1)]�TC28878 8 (3) [b(1), c(1), e(1),�g(5)]
Data are from cDNA libraries from a number of different tissue sources sequenced at the Samuel Roberts Noble Foundation. Further detailscan be found in the TIGR M. truncatula Gene Index (MtGI) at (http://www.tigr.org/tdb/mtgi/). TC sequences are assembled from ESTs, andmay represent full-length transcripts. TC annotations contain information on the source library and abundance of ESTs. The tissue sourcesof the cDNA libraries are: a, root (6593); b, stem (10 314); c, developing leaf (7831); d, phosphate-starved leaf (9034); e, drought-inducedwhole plants (8416); f, yeast-elicited cell culture (8926); g, insect-damaged leaf (9921); h, developing flower (3404); i, nodulated root (29 721);and j, germinating seed (451). The numbers in brackets refer to the total number of ESTs sequenced in each library as of December 2001.The symbol (�) denotes the library from which the full-length clones used in the present work were isolated, and the corresponding TC.
� Blackwell Publishing Ltd, The Plant Journal, (2002), 32, 1033–1048
Triterpene saponin biosynthesis 1035

sites for BamHI, SalI, or XbaI. EcoRI cuts once in SE2, but
does not cut SE1; thus, the common major band in the
EcoRI lanes in Figure 2(b,c) is likely due to cross-hybridiza-
tion with the other SE gene. SE1 and SE2 share 76.6%
sequence identity at the nucleic acid level. b-AS and SS
are both present in two copies in M. truncatula (Figure 2a,d).
DNA sequence alignment using the Clustal program
indicated that M. truncatula (GenBank Accession #
AJ430610) and soybean (Glycine max) SS proteins are
closely related (88.6% at the amino acid level), as would
be expected, since both species belong to the Fabaceae.
M. truncatula SE1 (GenBank Accession # AJ430609) and
SE2 (GenBank Accession # AJ430608) proteins are more
closely related to P. ginseng putative SE, with 77.1 and
74.4% sequence identity, respectively, than to Arabidopsis
and Brassica SEs (Figure 3a). The two distinct types of plant
OSC, cycloartenol synthase and b-AS, exhibit a relatively
high level of amino acid sequence identity, even though
their reaction products are quite distinct (Hayashi et al.,
2000; Kushiro et al., 1998). Alignment of known plant
cycloartenol synthase and b-AS proteins indicates that
the putative M. truncatula b-AS (GenBank Accession #
AJ430607) falls into the b-amyrin synthase group (Fig-
ure 3c). M. truncatula b-AS protein was closely related to
pea (Pisum sativum) b-AS with 94.7% sequence identity.
Figure 3(b) shows amino acid sequence alignments of
M. truncatula SE1 and SE2 with the enzymes from human,
rat, and yeast. There is a high degree of sequence con-
servation in certain key regions, particularly around the
squalene and FAD binding domains. The M. truncatula
Figure 2. DNA gel blot analysis of triterpene pathway genes in Medicago truncatula.Genomic DNA was cut with the enzymes shown (B¼BamHI, S¼SalI, E¼EcoRI, X¼XbaI), fragments resolved by agarose gel electrophoresis, and blots probedwith cDNAs encoding squalene synthase (a), squalene epoxidase 1 (b), squalene epoxidase 2 (c), and b-amyrin synthase (d). Molecular size markers (in Kb) areshown on the left.
Figure 3. Sequence analysis of Medicago truncatula SE and b-AS genes.(a) Dendrogram comparison of the sequences of oxidosqualene cyclases. The dendrogram was created using the Clustal W and TreeViewPPC programs. Key andGenBank accession numbers of the oxidosqualene cyclase superfamily: bAS, b-amyrin synthase: M. truncatula bAS (this study), A. strigosa bAS (AsbAS,AJ311789), P. ginseng bAS (PgAbAS, B009030); G. glabra bAS (GgbAS, AB037203); P. sativum bAS (PsbAS, AB034802). LS, lanosterol synthases: Homo sapiensLS (U22526); Rattus norvegicus LS (U31352); Saccharomyces cerevisiae LS (U04841). SHC, squalene-hopene cyclase: Alicyclobacillus acidocaldarius SHC(AB007002). LuS, lupeol synthases: Arabidopsis thaliana LuS (AtLuS, AAD05032); O. europaea LuS (OeLuS, AB025343); Taraxacum officinale LuS (ToLuS,AB025345). CAS, cycloartenol synthases: M. truncatula CAS (this study), A. strigosa CAS (AsCAS, AJ311790); O. sativa CAS (OsCAS, AF169966); Luffa cylindricaCAS (LcCAS, AB033334); G. glabra CAS (GgCAS, AB025968); P. sativum CAS (PsCAS, D89619); P. ginseng CAS (PgCAS, AB009029); O. europaea CAS (OeCAS,AB025344).(b) Amino acid sequence alignments of M. truncatula putative squalene epoxidases 1 and 2 with published squalene epoxidases from other organisms. Thehighly conserved squalene and FAD binding domains are highlighted with boxes in the N- and C-terminal portions of the protein, respectively. The arrow marksthe position in the N-terminus at which the M. truncatula proteins were truncated for expression in yeast.(c) Dendrogram comparing the sequences of squalene epoxidases. The dendrogram was created using the Clustal W and TreeViewPPC programs. Key andGenBank accession numbers: Arabidopsis thaliana SE1 (AtSE1), A. thaliana SE2 (AtSE2), A. thaliana SE3 (AtSE3), A. thaliana SE4 (AtSE4, AJ005930); A. thalianaSE5 (AtSE5, AJ005927); A. thaliana SE6 (AtSE6, AJ005929); Brassica napus SE1 (BnSE1, AJ005931), B. napus SE2 (BnSE2, AJ005928), M. truncatula SE1 (thisstudy), M. truncatula SE2 (this study), rat SE (D37920), human SE (D78130), yeast erg1 (M64994), Candida SE (D88252).
� Blackwell Publishing Ltd, The Plant Journal, (2002), 32, 1033–1048
1036 Hideyuki Suzuki et al.

� Blackwell Publishing Ltd, The Plant Journal, (2002), 32, 1033–1048
Triterpene saponin biosynthesis 1037

enzymes are more similar to the mammalian enzymes than
to the enzyme from yeast.
Transcript abundance of early saponin pathway genes in
M. truncatula
Abundance of putative saponin biosynthetic enzyme tran-
scripts in different organs was first assessed in silico from
analysis of EST occurrence in the various cDNA libraries
using the data available in the TIGR M. truncatula Gene
Index database (Table 1). Putative triterpene pathway genes
appeared to be expressed at a higher level in insect-
damaged leaves than in control leaves based on relative
EST abundance (Table 1). Three SS full-length clones
(including the one functionally characterized below) and
one truncated clone were found among the 9921 clones
sequenced from the insect-damaged leaf library, but only
one truncated SE clone (NF026F08IN) was found. Two full-
length and two truncated b-AS clones were found, includ-
ing the one functionally characterized below. The highest
abundance was five ESTs for b-AS from the insect-
damaged leaf library.
The essential features of the in silico transcript abun-
dance analysis were confirmed by RNA gel blot analysis
in several cases, using RNA samples from the original
preparations used for cDNA library construction (Figure 4).
To distinguish between SE1 and SE2 transcripts, N-terminal
gene-specific probes were generated corresponding to the
50-untranslated regions and the open reading frames 50 of
the FAD binding domains. Putative SS transcripts were
remarkably abundant in roots, whereas flower, leaf, petiole,
cell culture, and stem showed a lower level of expression.
The analysis of M. truncatula SE1 transcripts shows that
this gene is expressed weakly in all tissues examined.
Exposure of M. truncatula cell suspension cultures to yeast
elicitor for 24 h resulted in an enhancement of SS, but not
SE1, transcripts. SE2 transcripts were also expressed in all
tissues examined, but at higher levels than SE1 transcripts
and with evidence of weak induction in cell cultures by
yeast elicitor. Putative b-AS transcripts were most highly
expressed in flower and root and induced from a very low
basal level in yeast-elicited cell cultures.
Functional characterization of M. truncatula squalene
synthase
SS catalyzes the reductive dimerization of two molecules
of farnesyl diphosphate (FPP) in a two-step reaction to
Figure 4. RNA gel blot analysis of tissue distribution of Medicago truncatula triterpene pathway transcripts.(a) Total RNA was isolated from the tissues shown, resolved by agarose gel electrophoresis, blotted, and probed with full-length M. truncatula squalene synthase(SS), N-terminal fragments from squalene epoxidase 1 (SE1) and squalene epoxidase 2 (SE2), and full-length b-amyrin synthase (b-AS) cDNAs. Cell suspensioncultures were of root origin and were induced with yeast elicitor (YE).(b) Quantitation of the data in (a). Relative transcript abundance was calculated by dividing the intensity of each transcript to that of the corresponding 18S RNAtranscript level. C¼ cell culture, C(E)¼ elicited cell culture, F¼ flower, L¼ leaf, P¼peptide, R¼ root, S¼ stem.
� Blackwell Publishing Ltd, The Plant Journal, (2002), 32, 1033–1048
1038 Hideyuki Suzuki et al.

produce squalene. This reaction is believed to proceed via
head-to-head coupling of two FPP molecules to form squa-
lene via a stable cyclopropylcarbinyl diphosphate inter-
mediate (Pandit et al., 2000). Functional expression of the
M. truncatula putative SS was accomplished in E. coli BL21
cells. SDS-PAGE analysis of total proteins showed that a 43-
kDa band, corresponding to the predicted size of the recom-
binant protein, appeared in extracts from IPTG induced
E. coli, but not in cultures harboring an empty pET-15b
vector (data not shown). To demonstrate squalene
synthase activity, harvested cells were disrupted, centri-
fuged, and the supernatant used for enzyme assay in the
presence of 14C-FPP, NADPH and Mg2þ. Reverse-phase
TLC was used to separate the product(s). No 14C-squalene
product is formed in extracts of E. coli transformed with
the empty vector (Figure 5a). In contrast, extracts from
E. coli transformed with pET15b containing recombinant
SS (pET-SS), in spite of the very small proportion of
soluble recombinant enzyme, catalyzed formation of a
labeled product that co-migrated with authentic squalene.
When NADPH was omitted, no squalene product was
observed. A strong reduction in squalene formation was
also observed on omitting Mg2þ from the incubation mix-
ture, the residual activity perhaps being supported by
endogenous Mg2þ.
We further characterized the M. truncatula SS reaction by
substituting Mg2þ in the incubation mixture with other
divalent cations. Mn2þ, Co2þ, and Fe2þ could substitute
for Mg2þ as cofactors in this reaction, whereas Ca2þ,
Cu2þ, or Zn2þcould not (Figure 5a).
Functional characterization of M. truncatula squalene
epoxidase
SE catalyzes the insertion of an oxygen atom across a
carbon–carbon double bond to form an epoxide in a
reaction more typical of P450-type reactions. Squalene
monoxygenases have been cloned and functionally
characterized from yeast, rat, and human (Jandrositz
et al., 1991; Laden et al., 2000; Sakakibara et al., 1995),
but not from plants. SE, encoded by the ERG1 gene in
yeast, is a key enzyme in the sterol biosynthetic pathway.
The KLN1 strain of yeast (MATa, erg1::URA3, leu2, ura3,
trp1) used here for the functional characterization of puta-
tive M. truncatula SE is an obligate ergosterol auxotroph;
disruption of ERG1 is lethal unless ergosterol is supplied to
cells growing under anaerobic conditions (Landl et al.,
1996).
To functionally characterize the two putative M. trunca-
tula squalene epoxidases, the SE1 and SE2 coding
sequences, with 47 amino acids truncated from the N-
teminus of SE1, and 52 amino acids truncated from the
N-teminus of SE2, and the ERG1 ORF as a positive control
were cloned into the pWV3 vector, containing the LEU2
selectable marker, under the control of the constitutive
pADH1 promoter. The N-terminal truncation sites were
chosen by comparison with the yeast protein, which has
a short N-terminus compared to plant or mammalian SE
(Figure 3b); we thought that this truncation might therefore
facilitate expression of the plant enzyme in yeast. Selection
of transformants for the Leuþ phenotype was made in SD
medium supplied with ergosterol and tryptophan under
anaerobic conditions, as shown in Figure 5(b), panel 1.
As expected, KLN1 did not grow because the medium
was deprived of Leu (Landl et al., 1996). When plated in
YPD (or SDþ trp) medium without ergosterol under anae-
robic conditions, the transformants were not viable (Fig-
ure 5b, panel 2), whereas under aerobic conditions, they
exhibited strong growth (Figure 5b, panel 3). pWV3 trans-
formants were not able to grow under either condition,
showing that the SE or ERG1 (positive control) inserts
contributed to this growth. Thus, the growth of the trans-
formants is oxygen dependent, as is the SE reaction. These
data show that the ergosterol biosynthetic pathway in the
yeast erg1 knockout could be re-constituted by heterolo-
gous complementation with M. truncatula SE with or with-
out truncation of the N-terminus.
Functional characterization of M. truncatula b-amyrin
synthase
The enzyme encoded by EST NF051E06IN, when expressed
in yeast, cyclized oxidosqualene to form product(s) that co-
migrated with b-amyrin on TLC (data not shown), whereas
the yeast strain SMY8 harboring the empty vector did not
form this compound(s). GC analysis revealed that the
recombinant M. truncatula b-AS only produced a single
cyclization product from oxidosqualene, as shown in Fig-
ure 5(c). The GC relative retention time (Rt) of the cyclization
product, TMS ether, was identical to that of authentic b-
amyrin TMS ether (Rt¼ 1.23, relative to epicoprostanol
TMS ether). The mass spectra (MS) of the enzymatic pro-
duct, b-amyrin standard and their co-injection agreed with
each other. (EI-MS): (TMS ether) m/z¼ 498 [M]þ (6%), 483
[M-CH3]þ (3%), 408 [M-Me3SiOH]þ (2%), 393 [M-Me3SiOH-
CH3]þ (3%), 218 (C-ring fragment, 100%), 203 [m/z 218 –
CH3]þ (39%). NMR data further confirmed the identification
of b-amyrin. Key 1H-NMR signals of the authentic sample
matched those of the NF051E06IN product to �0.001 p.p.m.
(500 MHz, CDCl3, tetramethylsilane as internal standard): d5.184 (t, 3.5 Hz, 1H, H-12), 3.223 (ddd, 11.2, 6.0, 4.7 Hz, 1H, H-
3), 1.135 (d, 1.0Hz, 3H, H-27), 0.998 (s, 3H, H-23), 0.968 (s, 3H,
H-26), 0.938 (s, 3H, H-25), 0.872 (s, 6H, H-29, H-30), 0.832 (s,
3H, H-28), 0.792 (s, 3H, H-24), 0.742 (d, 11.7, 1.9 Hz, 1H, H-5).
The observed MS and NMR data agreed with literature
values for b-amyrin (Segura et al., 2000). A 290-ml yeast
culture produced 1.7 mg of b-amyrin (>98% pure) from
14 mg of oxidosqualene substrate.
� Blackwell Publishing Ltd, The Plant Journal, (2002), 32, 1033–1048
Triterpene saponin biosynthesis 1039

� Blackwell Publishing Ltd, The Plant Journal, (2002), 32, 1033–1048
1040 Hideyuki Suzuki et al.

Development of an inducible cell culture system for
functional genomics approaches to further understand
triterpene saponin biosynthesis in M. truncatula
The reactions of triterpene biosynthesis beyond the initial
cyclization step catalyzed by b-AS are complex, and none of
the enzymes involved in Medicago has been characterized
at the molecular level. However, it is possible to present a
putative pathway based on the nature of the saponins
identified in M. truncatula and analogies with parallel oxi-
dative reactions involved in brassinosteroid biosynthesis
(Bishop et al., 1999; Fujioka et al., 2000; Huhman and
Sumner, 2002; Noguchi et al., 2000). It is likely that the
oxidative reactions are catalyzed by a series of cytochrome
P450 monooxygenases, with the final reactions catalyzed
by glycosyltransferases (GTs), as outlined in Figure 1. To
utilize existing information on P450s and GTs from the
M. truncatula genomic databases, it is necessary to develop
a system in which the saponin pathway can be rapidly and
reproducibly induced as a basis for micro- and macro-array
experiments.
Extractation and quantitation of the multiple M. trunca-
tula triterpene saponins is not trivial (Huhman and Sumner,
2002), and is therefore not the best assay method for
determining expression of the triterpene pathway. We
therefore decided to measure changes in transcript levels
by RNA gel blot analysis, using the functionally confirmed
Medicago b-AS, SE, and SS as probes, in a series of experi-
ments designed to investigate conditions for inducing tri-
terpene synthesis in M. truncatula root cell suspension
cultures. We tested a variety of potential elicitors, including
methyl jasmonate (MeJA), yeast elicitor (YE), salicylic acid
(SA), and abscisic acid (ABA), for their ability to induce
saponin pathway transcripts in the cultures. YE weakly
induced SS, SE2, and b-AS, as previously shown in Figure 4.
Enhancement of b-AS transcript levels was two- and sixfold
at 12 h post-elicitation with YE and SA, respectively (data
not shown). b-AS transcripts were induced to a maximum
level of 2.5-fold 1 h after exposure to ABA (data not shown).
Strongest elicitation of b-AS transcripts was found with
MeJA, which induced an increase of up to 30-fold by 8–
24 h post-elicitation, as shown in Figure 6(a,b). SS tran-
scripts were coordinately induced with b-AS transcripts
in response to MeJA. SE1 transcripts were not significantly
induced by MeJA, whereas SE2 transcript induction closely
followed that of b-AS (Figure 6a,b), indicating that SE2, but
not SE1, may function specifically in the formation of
triterpenoids. In contrast, elicitation with MeJA caused a
significant reduction in cycloartenol synthase transcript
levels.
Treatment of cell cultures with MeJA was confirmed to
induce accumulation of triterpene saponins, as assessed by
chemical extraction and analysis by LC/MS (Figure 7a).
Small amounts of hederagenin glycoside were present in
unelicited cultures. These increased approximately 10-fold
by 24 h post-elicitation, and glycosides of soyasapogenols
B and E appeared. The latter compounds were not detected
in the unelicited cultures.
The reduction in cycloartenol synthase transcripts in
response to MeJA could indicate preferential channeling
of oxidosqualene from biosynthesis of housekeeping ster-
ols into the triterpene pathway. However, GC/MS analysis
of control and elicited cell cultures revealed no significant
change in sterol profiles at 24 h post-elicitation, as shown in
Figure 7(b). The major difference between elicited and con-
trol cultures was the appearance, following elicitation, of a
small peak shown to correspond to the trimethylsilyl ether
of b-amyrin.
MeJA has been suggested to be a signal molecule for the
biosynthesis of phytoalexins (Gundlach et al., 1992). In
Medicago species, the phytoalexins are isoflavonoids
derived from the phenylpropanoid/flavonoid pathway
(Dixon, 1999). To determine whether the phenylpropanoid
pathway is co-induced with the triterpene biosynthetic
pathway following exposure of cells to MeJA, membranes
were probed with labeled M. truncatula phenylalanine
ammonia lyase (PAL) and chalcone synthase (CHS) cDNAs.
PAL transcripts were only weakly induced by MeJA, with a
Figure 5. Functional characterization of Medicago truncatula squalene synthase, squalene epoxidase, and b-amyrin synthase.(a) M. truncatula squalene synthase (SS) was expressed in Escherichia coli BL21(DE3, pLyS) using the pET-15b expression vector. The enzyme was assayed byradio-TLC as described in Experimental procedures. Lane 1: extract from E. coli harboring pET-15b empty vector assayed in the presence ofNADPHþMgCl2þDTTþKFþ 14C-FPPþ50 mM Tris–HCl (pH 7.6) (negative control). Lane 2: extract from E. coli harboring pET-SS assayed as in lane 1 (positivecontrol). Lanes 3–11: extracts from E. coli harboring pET-SS assayed with different components in the reaction mixture. Lane 3: without NADPH; lane 4, withoutDTT; lane 5, without MgCl2; lanes 6–11, MnCl2, CaCl2, CoCl2, CuCl2, FeCl2, ZnCl2 in place of MgCl2; lane 12, authentic 14C-squalene. SQ, squalene; FOH, farnesol.(b) Complementation of the yeast erg1 mutant by M. truncatula squalene epoxidase. (1) Selection of transformants for the Leuþ phenotype in SD mediumsupplied with ergosterol and tryptophan under anaerobic conditions. (2) Plating of yeast cells in YPD (or SDþ trp) medium without ergosterol under anaerobicconditions. The transformants were not viable. The same result was obtained with SD medium plus tryptophan. (3) Growth of yeast cells in YPD medium withoutergosterol under aerobic conditions. KLN1¼non-transformed KLN1 yeast strain; pWV3¼KLN1 yeast transformed with the pWV3 yeast expression vector only;pWV3-SE1 and pWV3-SE2¼KLN1 yeast transformed with the pWV3 yeast expression vector containing SE1 and SE2 ORFs, respectively; pWV3-SE1D47 andpWV3-SE2D52¼KLN1 yeast transformed with the pWV3 yeast expression vector containing SE1 and SE2, with 47 and 52 amino acids truncated from the N-termini, respectively.(c) Functional expression of M. truncatula b-amyrin synthase in yeast. Gas chromatogram showing the product from the incubation of recombinant b-amyrinsynthase with oxidosqualene. A single product, representing the TMS ether of b-amyrin, was produced. The internal standard peak is the TMS ether ofepicoprostanol.
� Blackwell Publishing Ltd, The Plant Journal, (2002), 32, 1033–1048
Triterpene saponin biosynthesis 1041
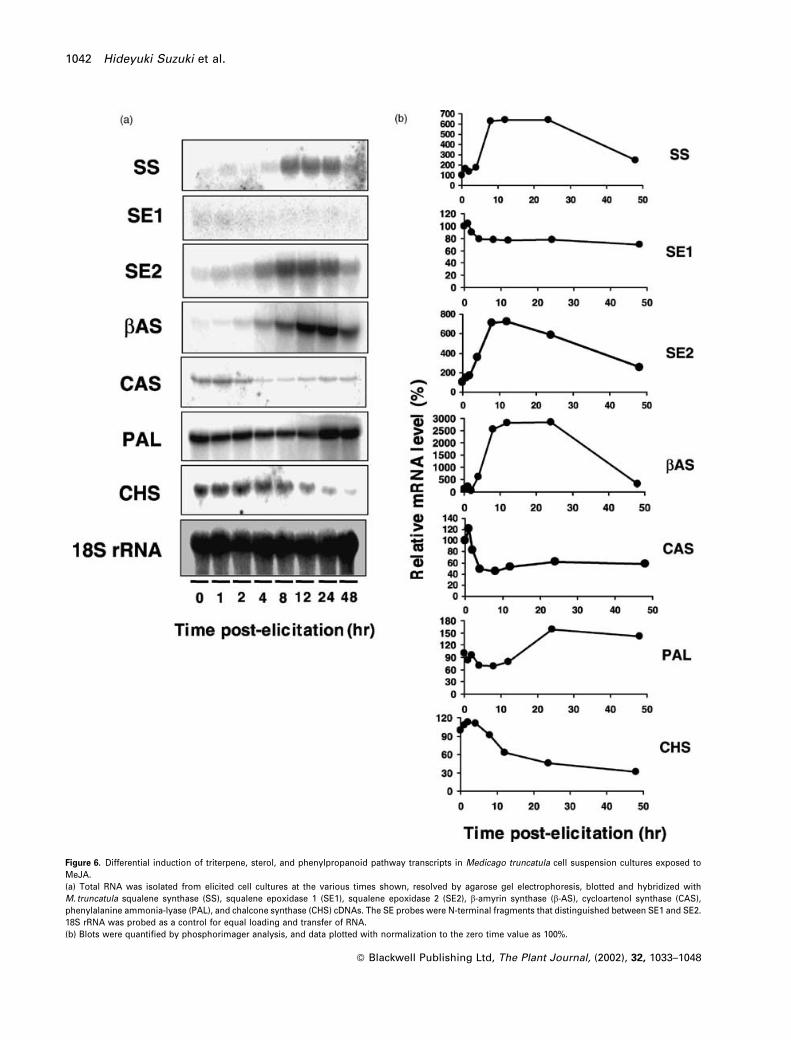
Figure 6. Differential induction of triterpene, sterol, and phenylpropanoid pathway transcripts in Medicago truncatula cell suspension cultures exposed toMeJA.(a) Total RNA was isolated from elicited cell cultures at the various times shown, resolved by agarose gel electrophoresis, blotted and hybridized withM. truncatula squalene synthase (SS), squalene epoxidase 1 (SE1), squalene epoxidase 2 (SE2), b-amyrin synthase (b-AS), cycloartenol synthase (CAS),phenylalanine ammonia-lyase (PAL), and chalcone synthase (CHS) cDNAs. The SE probes were N-terminal fragments that distinguished between SE1 and SE2.18S rRNA was probed as a control for equal loading and transfer of RNA.(b) Blots were quantified by phosphorimager analysis, and data plotted with normalization to the zero time value as 100%.
� Blackwell Publishing Ltd, The Plant Journal, (2002), 32, 1033–1048
1042 Hideyuki Suzuki et al.

maximum increase of only 1.5-fold at 24 h post-elicitation.
More strikingly, CHS transcript levels decreased in parallel
to the increase in b-AS mRNA (Figure 6a,b).
Discussion
Mining M. truncatula EST data sets yielded candidate
clones for the enzymes that catalyze the early steps of
triterpene biosynthesis, namely SS, SE, and b-AS. Several
of the ESTs proved to originate from full-length clones,
which were functionally characterized by heterologous
expression. Divalent cation-dependent production of squa-
lene from farnesyl diphosphate in E. coli extracts provided
functional confirmation for heterologous expression of
M. truncatula SS. A. thaliana SS has been functionally
expressed, and shown to produce squalene in the presence
of Mg2þ and NADPH, and dehydrosqualene in the presence
of Mn2þ but absence of NADPH (Nakashima et al., 1995).
The M. truncatula squalene synthase could use Mn2þ or
Mg2þ equally well as cofactors for squalene formation in
the presence of NADPH. Interestingly, the intact full-length
Arabidopsis SS1 cannot complement a yeast SS mutant,
even though the yeast cells expressing the Arabidopsis
enzyme contain detectable SS activity when assayed in vi-
tro. This has been shown to be due to a requirement for a
specific C-terminal portion of the yeast SS for metabolic
channeling of squalene through the yeast sterol pathway
(Kribii et al., 1997). This is an interesting feature from the
point of view of the organization of potential metabolic
complexes necessary for channeling of squalene into either
the triterpene or the sterol pathway in plants. The fact that
M. truncatula possesses two isoforms of the subsequent
enzyme in the pathway, SE1 and SE2, raises the question of
whether these may have different biochemical functions in
relation to triterpene and sterol biosynthesis. This idea is
supported by the co-induction of SE2, but not SE1, with b-
AS in MeJA-treated cell cultures.
Mammalian SE plays a pivotal role in cholesterol bio-
synthesis, and the enzyme is expressed at low levels in
most tissues (Ono and Bloch, 1975; Yamamoto and Bloch,
1970). Detailed enzymological characterization of human
SE has been reported (Laden et al., 2000). In yeast, the
squalene epoxidase Erg1p exhibits dual localization in the
endoplasmic reticulum and in lipid particles (Leber et al.,
1998). Although plant genes with sequence similarity to
mammalian SE have been described in the literature, the
present report is, to the best of our knowledge, the first to
document the functional characterization of a plant SE. The
two isoforms of M. truncatula squalene epoxidase, SE1 and
SE2, share 82.1% amino acid identity. Both M. truncatula
SEs could complement the ergosterol biosynthetic path-
way in the Erg1 knockout yeast strain KLN1. This is inter-
esting in view of the failure of Arabidopsis SS to correctly
couple with the sterol biosynthetic machinery in yeast
(Kribii et al., 1997), and the complexity of the mammalian
squalene epoxidase reaction which requires, in addition to
NADPH cytochrome P450 reductase, a specific lipid transfer
protein for transfer of squalene to the enzyme (Shibata et al.,
2001). This also suggests that in spite of the differen-
tial induction of the two Medicago SEs in planta, with its
Figure 7. Triterpene and sterol profiles in M. truncatula cell suspension cultures.(a,b) Accumulation of triterpene saponins in response to MeJA. The traces show portions of selective ion chromatograms of extracts from unelicited (a) and 24-hMeJA-elicited (b) M. truncatula cell suspension cultures. 1, rhamnose-hexose-hexose-hederagenin; 2, 3-rhamnose-galactose-glucose-soyasapogenol B; 3,rhamnose-hexose-hexose-soyasapogenol E.(c,d) Effects of MeJA on sterol levels. The traces show portions of selective ion chromatograms of extracts from unelicited (c) and 24-h MeJA-elicited (d)M. truncatula cell suspension cultures. All compounds are detected as the TMS ethers. 1, stigmasterol; 5, [22E]-ergosta-7,22-dien-3a-ol; 6, b-amyrin; 8, stigmata-7,24[28]-dien-33a-ol. Compounds 2, 3, 4 and 7 are unidentified sterols.
� Blackwell Publishing Ltd, The Plant Journal, (2002), 32, 1033–1048
Triterpene saponin biosynthesis 1043

implications for differential function, both forms might be
able to participate in sterol biosynthesis in plant cells, as in
the heterologous yeast system.
The formation of b-amyrin by cyclization of 2,3-oxidos-
qualene is a complex reaction believed to occur via the
‘chair-chair-chair’ conformation of the substrate. The OSCs
lanosterol and cycloartenol synthase have been extensively
studied in mammals and yeast (Abe and Prestwich, 1995;
Corey et al., 1993, 1994, 1996; Morita et al., 1997). P. ginseng
b-AS has been previously cloned and functionally
expressed in yeast (Kushiro et al., 1998). Recently, cDNAs
encoding three proteins from A. thaliana with 49–59% iden-
tity to cycloartenol synthase were functionally expressed.
The products of one of these enzymes consisted of a
mixture of lupeol, b-amyrin, and a-amyrin (15 : 55 : 30) (Hus-
selstein-Muller et al., 2001). This enzyme is distinct from a
lupeol synthase characterized from the same source (Her-
rera et al., 1998). In the present study, M. truncatula b-AS
catalyzed the formation of b-amyrin alone, with no minor
products, as also observed for the Panax enzyme (Kushiro
et al., 1998).
Previous studies have shown effects of sucrose and
mineral nutrients on saponin production in plant cell sus-
pension cultures, but these effects were neither large nor
rapid (Akalezi et al., 1999; Fulcheri et al., 1998). M. truncatula
root cell suspension cultures produce low levels of triter-
pene saponins and have correspondingly low-steady state
levels of SS, SE and, particularly b-AS transcripts. We have
now determined conditions for rapid induction of triterpene
biosynthesis in the cultures following exposure to MeJA.
Jasmonates are important stress signaling molecules
known to be good elicitors for a wide range of secondary
metabolites such as polyamines, coumaryl conjugates,
anthraquinones, naphthoquinones, polysaccharides, terpe-
noids, alkaloids, and phenylpropanoids from different plant
origins (Memelink et al., 2001). In Medicago cell suspension
cultures, exposure to MeJA downregulates the flavonoid
branch of phenylpropanoid biosynthesis, as assessed by
CHS steady state transcript levels, but induces the appear-
ance of glycosides of the triterpenes hederagenin and
soyasapogenols B and E. In contrast, exposure of the cells
to yeast elicitor results in a strong induction of the phenyl-
propanoid pathway associated with accumulation of iso-
flavonoid phytoalexins (H. Suzuki, J.W. Blount, and R.A.
Dixon, unpublished results), but with little effect on triter-
pene biosynthesis. The downregulation of cycloartenol
synthase transcripts following exposure to MeJA suggests
preferential channeling of oxidosqualene from sterol synth-
esis to triterpene synthesis following elicitation, in a similar
manner to the regulation at the sesquiterpene/sterol branch
in elicited tobacco cell cultures (Vogeli and Chappell, 1988).
However, metabolic profile analysis failed to demonstrate
a reduction in sterol levels in MeJA-elicited M. truncatula
cell cultures, at least at the 24-h time point tested. Thus,
although cycloartenol synthase transcript levels are down-
regulated by MeJA treatment, they do not completely dis-
appear but rather reach a new steady state, which may still
be sufficient to support sterol biosynthesis.
MeJA is a wound signal for activation of plant responses
to insect attack (McConn et al., 1997). It is interesting that
high expression of b-AS appears to be associated with
insect herbivory in M. truncatula. There is a correlation
between growth of the yellow mealworm (Tenebrio moli-
tor) and saponin content in M. sativa, with glycosides of me-
dicagenic acid exhibiting the highest degree of antifeedant
activity (Pracros, 1988). Preliminary results indicate that
regurgitant from tobacco hornworm (Korth and Dixon,
1997) can induce b-AS transcripts both in intact plants
(K. Korth, personal communication) and cell cultures
(L. Achnine and H. Suzuki, unpublished results) of M.
truncatula. The cell culture system may therefore prove a
use ful model for the study of anti-insect responses in
legumes.
The later enzymes of triterpene biosynthesis are primarily
cytochrome P450s and glycosyl transferases. These
enzymes exist as large supergene families in plants (Chap-
ple, 1998; Vogt and Jones, 2000), with approximately 250
members of the P450 family estimated from current
M. truncatula EST information. Glycosyltransferase activity
has been shown to correlate with saponin production in
root cultures of Gypsophila paniculata (Herold & Henry,
2001), but specific triterpene glycosyltransfereases remain
to be characterized at the molecular level. The ability to
differentially upregulate two major pathways of natural
product metabolism (i.e. triterpenes and phenylpropa-
noids/flavonoids) in a cell culture system facilitates the
design of DNA macro- and microarray experiments for
selection of candidate P450 and glycosyltransferase genes
from our extensive EST collections (Bell et al., 2001). These
can then be functionally characterized in yeast (P450s)
or E. coli (GTs). It is possible that the cytochrome P450s
might represent important regulatory steps in triterpene
biosynthesis.
Experimental procedures
Plant material
M. truncatula Gaerth ‘Jemalong’ (line A17) plants were grownunder greenhouse conditions in 11 cm diameter pots in Metro-mix 250 or 350 (Scott, Marysville, OH, USA), nine plants per potwith an 18-h light/258C and 6-h dark/228C photoperiod. Cell sus-pension cultures were initiated from roots of line A17, maintainedin a modified Schenk and Hildebrandt medium, as describedpreviously for alfalfa cultures (Dixon et al., 1981), and subculturedevery 10–14 days.
Six days after subculture, dark grown M. truncatula A-17 rootsuspension cultures (75-ml batches) were treated with MeJA(500 mM), YE (50 mg glucose equivalents ml�1), SA (500 mM), orABA (500 mM), harvested at various times after elicitation and
� Blackwell Publishing Ltd, The Plant Journal, (2002), 32, 1033–1048
1044 Hideyuki Suzuki et al.

frozen at �808C. Control cells were treated with the same volumeof distilled water.
Extraction and quantitation of triterpene saponins and
sterols
Saponin extracts were obtained using a solid phase extractionprocedure as previously described (Oleszek, 1988; Oleszek et al.,1990). Six grams fresh weight of cells was extracted in 80%methanol for 24 h. The extracts were concentrated under a nitro-gen stream to yield an aqueous solution that was diluted to a finalconcentration of 35% methanol (v/v) and loaded onto a 35-ml, 10-g,C18 SPE extraction cartridge (Waters, Milford, MS). The SPEcartridge was washed with two column volumes each of HPLCgrade water and 35% methanol. The saponins were eluted withtwo column volumes of 100% methanol. The methanol fractionwas dried under vacuum, re-suspended in methanol to a finalconcentration of c. 400 ng ml�1 and analyzed by gradient elution,reverse-phase HPLC with simultaneous online UV and mass selec-tive detection (Huhman and Sumner, 2002).
For determination of sterol levels, 6.0 mg of lyophilized tissuewas extracted with 1.5 ml water and 1.5 ml chloroform at 508C for2 h. The sample was centrifuged at 2900 g for 30 min and 1.0 ml ofthe organic layer collected and dried under a stream of nitrogen.Lipids were hydrolyzed in 0.8 ml chloroform and 0.5 ml of metha-nol:HCl (97 : 3) for 2 h at 508C. The samples were washed with three0.5-ml aliquots of HPLC grade water to remove acid, dried, re-suspended in 50 ml pyridine, and derivatized with 50 ml MSTFA (N-methyl-N-trimethylsilyltriflouroacetamide) plus 1% TMCS (tri-methylchlorosilane). One microliter of sample was injected usingan HP 6890 series autosampler onto an HP 6890 GC with a DB-5MScolumn (60 m, 0.25-mm film thickness, 0.25 mm i.d., J & W Scien-tific). Spectroscopic data were acquired with an HP 5973 massselective detector scanning from 50 to 650 at a scan rate of 2.48scans sec�1. Best matches were assigned by searching the NISTlibrary following spectral de-convolution by AMDIS software.
Functional expression of M. truncatula SS, SE, and b-AS
Expression of M. truncatula EST clone NF066G09IN, a putativesqualene synthase, was performed by amplification of the openreading frame from pBluescript II SKþ (Stratagene, La Jolla, CA)with introduction of NcoI and BamHI sites (50-CCATGCCATGG-GAAGTATAAAAGCGATTTTGAAGAATC-30 for the upstream pri-mer and 50-CGGGATCCTTAGTTATTGTGACGATTGGCAGAGAG-30
for the downstream primer). The PCR product was purified, ligatedinto pGEMTeasy vector (Promega, Madison, WI, USA), sequenced,excised and re-cloned between the NcoI and BamHI sites of thepET15b expression vector (Novagen, Madison, WI, USA). E. coliBL21 (DE3, pLyS) cells harboring the expression construct weregrown to an OD600 of 0.6, and expression was induced by additionof isopropyl 1-thio-b-D-galactopyranoside (IPTG) to a final con-centration of 0.5 mM, with further incubation for 3 h. Cell lysateswere prepared and the crude extract used for the enzyme assay.The reaction mixture contained, in a total volume of 100 ml, 11.4 mM
[1,2–14C]-FPP (125 nCi; American Radiolabeled Chemicals, St.Louis, MO), 3 mM NADPH, 5 mM MgCl2, 0.1 mM dithiothreitol,100 mM KF, 50 mM Tris–HCl (pH 7.6), and enzyme (70mg of pro-tein). The reaction mixture was incubated at 308C for 1 h and thereaction stopped by addition of 100 ml ethyl acetate. Lipids wereextracted with ethyl acetate and 10 ml subjected to TLC on silica gelreverse phase plates (RP-18) (J.T. Baker, Phillipsburg, NJ). Theplates were developed with acetone: water (19 : 1, v/v). After devel-
opment, plates were exposed and analyzed with a bio-imageanalyzer (Molecular Dynamics, Sunnyvale, CA).
Functional identification of putative squalene epoxidasesencoded by M. truncatula EST clones NF065G06EC (SE1) andNF102D09LF (SE2) was achieved by heterologous expression inthe Erg1 knockout yeast mutant KLN (MATa, erg1::URA3, leu2,ura3, trp) (Landl et al., 1996). The PCR fragments with introducedBamHI and XhoI sites were amplified with the following primers:for the pWV3-SE1 construct, 50-CGCGGATCCATGATAGACCCC-TACGGTTTCGGGTGG-30 for upstream and 50- CCGCTCGAGT-TATGCATCTGGAGGAGCTCTATAAT-30 for downstream; forthe pWV3-D47SE1 construct, 50-CGCGGATCCATGTCTTTTAATCC-CAACGGCGATGTTG-30 for upstream; for the pWV3-SE2 construct,50-CGCGGATCCATGGATCTATACAATATCGGTTGGAATTTA-30 forupstream and 50-CCGCTCGAGTCAAAATGCATTTACCGGGGGAG-CTC-30 for downstream; for the pWV3-D52SE2 construct, 50-CGC-GGATCCATGTCGGACAAACTTAACGGTGATGCTG-30 forupstream.For amplification of the yeast Erg1 sequence, 50-CGGGATC-CATGTCTGCTGTTAACGTTGCACCTGAATTG-30 was used for theupstream primer and 50-CCGCTCGAGTTAACCAATCAACTCAC-CAAACAAAAATGGG-30 for downstream. The PCR products werepurified, subcloned into pGEMTeasy vector, sequenced, excised,and re-cloned between the BamHI and XhoI sites of the pWV3 yeastexpression vector. The SE1 and SE2 ORFs, SE1 with 47 amino acidstruncated from the N-teminus, SE2 with 52 amino acids truncatedfrom the N-teminus, and the Erg1 ORF as a positive control, wereunder control of the constitutive ADH1 promoter, and the pWV3vector contained the Leu2 selectable marker for yeast expression.Anaerobic conditions were achieved by culturing the yeast strainsin an Anaerocult A chamber (VWR Scientific Products, Atlanta,GA). Ergosterol (final concentration 20 mg ml�1) was dissolved inTween 80:ethanol (1 : 1, v/v), with a final Tween 80 concentration of0.5% (v/v) in the medium. EST clone NF051E06IN contained anapparent full-length oxidosqualene cyclase (OSC) gene in pBlue-script SK– vector. The plasmid was digested with NotI, XhoI, andScaI, to release the 2.8-kb insert with NotI and XhoI termini (ScaIwas included to cut the 2.9-kb vector into 1.1- and 1.8-kb fragments,facilitating purification). The insert was subcloned into the yeastexpression vector pRS426GalR that contains the URA3 selectablemarker, the 2m origin of replication, and Gal promoter. This high-copy expression construct was named pRX10.2, and was trans-formed into yeast lanosterol synthase mutant SMY8 (MATaerg7::HIS3 hem1::TRP1 ura3–52 trp1-D63 leu2–3112 his3-D200ade2 Galþ). The transformants were selected on synthetic com-plete medium (containing 2% dextrose) lacking uracil and supple-mented with heme (13 mg ml�1), ergosterol (20 mg ml�1), andTween-80 (5 ml ml�1). SMY8 harboring empty vector pRS426Galwas used as negative control in the following assay.
A 5-ml yeast culture was induced with 2% (w/v) galactose andgrown to saturation. The harvested yeast cells were re-suspendedin 200 mM sodium phosphate buffer (pH 6.4), lysed by vortexingwith glass beads, and incubated with 1 mg ml�1 oxidosqualeneand 0.1% Tween-80. The reaction was incubated at room tempera-ture for 24 h and quenched with two volumes of ethanol. Aftercentrifugation, the supernatant was transferred into a glass tube,and the cell debris was extracted with two further volumes ofethanol. The combined ethanol extract was dried under a nitrogenstream, re-dissolved in ethyl acetate and filtered through a smallsilica gel plug to remove cell debris and some polar components.The crude extract was derivatized to form trimethylsilyl (TMS)ethers by treatment with 50 ml of bis(trimethylsilyl)trifluoroaceta-mide-pyridine (1 : 1, v/v) at 408C for 2 h and was analyzed by GC-FIDand GC–MS, with epicoprostanol (an unnatural C-30 sterol) TMSether as internal standard and authentic b-amyrin TMS ether as
� Blackwell Publishing Ltd, The Plant Journal, (2002), 32, 1033–1048
Triterpene saponin biosynthesis 1045

external standard. Co-injection of crude product(s) and b-amyrinstandard was performed on GC–MS.
GC analysis employed a Hewlett-Packard 6890 system equippedwith a Rtx-5 capillary column (Restek, 30 m� 0.25 mm i.d., 0.10 mmdf). A 5-ml aliquot was injected at 2808C with a split ratio of 40 : 1,helium flow was at 20 cm sec�1, and the following temperatureprogram was applied: 1008C for 2 min, rising to 2808C at208C min�1, holding at 2808C for 30 min. The flame ionizationdetector was at 2808C. GC–MS was performed on a Hewlett-Pack-ard 5890 A instrument equipped with a DB-5ms column (J & W, 60m� 0.25 mm i.d., 0.10 mm df). Separation was achieved with split-less injection (1-min delay) at 2008C, helium flow at 30 cm sec�1
(1 ml min�1) and the identical temperature program as above.Mass spectra (m/z 35–500) were obtained on a ZAB-HF reverse-geometry double-focusing instrument at 70 eV with an electron-impact ion source (2008C). The accelerating voltage was 8 kV andthe resolution was 1000 (10% valley).
A 300-ml SMY8[RX10.2] yeast culture was processed similarly toobtain enough enzymatic product(s) for NMR analysis. The etha-nolic supernatant of the in vitro catalytic reaction was evaporatedto dryness and re-dissolved in ethyl acetate. The crude mixturewas filtered through a silica plug and then separated by silica gelcolumn chromatography to remove excess oxidosqualene sub-strate, exogenous ergosterol, and fatty acids. Polycyclic triterpenealcohols co-migrate with b-amyrin on TLC, and fractions withmaterial in this region were pooled and analyzed by 1H-NMRand GC–MS, which showed b-amyrin uncontaminated by othertriterpene alcohol isomers (detection limit 2%). NMR spectra offree sterols were obtained on a Bruker AMX500 spectrometer(500 MHz for 1H) at 258C in CDCl3 solution and referenced tointernal tetramethylsilane.
DNA and RNA gel blot analysis
DNA was extracted from M. truncatula plants by standard methods(Sambrook et al., 1989). Southern blotting and hybridization werecarried out as described previously (Church and Gilbert, 1984). TheSS, SE1, SE2, and b-AS probes were amplified as complete ORFsfrom the EST clones NF066G09IN, NF065G06EC (SE1),NF102D09LF (SE2), and NF051E06IN, respectively. For generationof gene-specific probes to distinguish between the SE1 and SE2transcripts, pBluescriptSE1 was double digested with EcoRI andNaeI, and pBluescriptSE2 was digested with EcoRI and AatII,resulting, respectively, in 195- and 247-bp fragments specific forthe N-termini of the two open reading frames.
Thirty micrograms of M. truncatula RNA from root cell suspen-sion culture or other tissues was separated by electrophoresis in a1% agarose gel containing 0.66 M formaldehyde and then blottedonto a Hybond-Nþ membrane (Amersham). The gene-specific SE1and SE2 probes and the entire cDNA fragments of SS, b-AS, andthe M. truncatula cycloartenol synthase (NF015H10LF), phenylala-nine ammonia lyase (NF011C12ST), and chalcone synthase(NF044D07EC) EST clones were radiolabeled with [32P] dCTP usinga Ready-to-go DNA Labeling Beads (-dCTP) kit (Amersham) andused as probes.
Acknowledgements
The authors thank Drs R. Leber and F. Turnowsky (Institute ofMolecular Biology, Graz University, Austria) for providing theKLN1 yeast strain, Dr Wayne Versaw (Noble Foundation) for thepWV3 yeast expression vector, Angela Scott for EST sequencing,David Huhman for LC/MS analysis of saponins, Corey Broeckling
and Jihai Pang for GC/MS analysis of sterols and b-amyrin, and DrsLiangjiang Wang and Robert Gonzales for critical reading of themanuscript. This work was supported by the Samuel RobertsNoble Foundation and in part by grants to R.A.D. from the NationalScience Foundation (0109732) and to S.P.T.M. from the USDA(2001-35315-10157) and the Robert A. Welch Foundation (C-1323).
References
Abe, I. and Prestwich, G.D. (1995) Molecular cloning, characteriza-tion, and functional expression of rat oxidosqualene cyclasecDNA. Proc. Natl Acad. Sci. USA, 92, 9274–9278.
Abe, I., Rohmer, M. and Prestwich, G.D. (1993) Enzymatic cycliza-tion of squalene and oxidosqualene to sterols and triterpenes.Chem. Rev. 93, 2189–2206.
Akalezi, C.O., Liu, S., Li, Q.S., Yu, J.T. and Zhong, J.J. (1999)Combined effects of initial sucrose concentration and inoculumsize on cell growth and ginseng saponin production by sus-pension cultures of Panax ginseng. Process Biochem. 34,639–664.
Behboudi, S., Morein, B. and VillacresEriksson, M.C. (1999) Quil-laja saponin formulations that stimulate proinflammatory cyto-kines elicit a potent acquired cell-mediated immunity. Scand.J. Immunol. 50, 371–377.
Bell, C., Dixon, R.A., Farmer, A.D. et al. (2001) The Medicagogenome initiative: a model legume database. Nucl Acids Res.29, 114–117.
Bishop, G.J., Nomura, T., Yokota, T., Harrison, K., Noguchi, T.,Fujioka, S., Takatsuto, S., Jones, J.D.G. and Kamiya, Y. (1999)The tomato DWARF enzyme catalyses C-6 oxidation in brassi-nosteroid biosynthesis. Proc. Natl Acad. Sci. USA, 96, 1761–1766.
Chapple, C. (1998) Molecular genetic analysis of plant cytochromeP450-dependent monooxygenases. Annu. Rev. Plant Physiol.Plant Mol. Biol. 49, 311–343.
Cheeke, P.R. (1976) Nutritional and physiological properties ofsaponins. Nutr. Report Int. 13, 315–324.
Church, G.M. and Gilbert, W. (1984) Genomic sequencing. Proc.Natl Acad. Sci. USA, 81, 1991–1995.
Corey, E.J., Matsuda, S.P.T., Baker, C.H., Ting, A.Y. and Cheng, H.(1996) Molecular cloning of a Schizosaccharomyces pombecDNA encoding lanosterol synthase and investigation of con-served tryptophan residues. Biochem. Biophys. Res. Comm.219, 327–331.
Corey, E.J., Matsuda, S.P.T. and Bartel, B. (1993) Isolation of anArabidopsis thaliana gene encoding cycloartenol synthase byfunctional expression in a yeast mutant lacking lanosterolsynthase by the use of a chromatographic screen. Proc. NatlAcad. Sci. USA, 90, 11628–11632.
Corey, E.J., Matsuda, S.P.T. and Bartel, B. (1994) Molecular clon-ing, characterization, and overexpression of ERG7, the Sacchar-omyces cerevisiae gene encoding lanosterol synthase. Proc.Natl Acad. Sci. USA, 91, 2211–2215.
Dixon, R.A. (1999) Isoflavonoids: biochemistry, molecular biologyand biological functions. In Comprehensive Natural ProductsChemistry, Vol. 1 (Sankawa, U., ed.). Oxford: Elsevier, pp.773–823.
Dixon, R.A., Dey, P.M., Murphy, D.L. and Whitehead, I.M. (1981)Dose–responses for Colletotrichum lindemuthianum elicitor-mediated enzyme induction in French bean cell suspensioncultures. Planta, 151, 272–280.
Fujioka, S., Noguchi, T., Watanabe, T., Takatsuto, S. and Yoshida,S. (2000) Biosynthesis of brassinosteroids in cultured cells ofCatharanthus roseus. Phytochemistry, 53, 549–553.
� Blackwell Publishing Ltd, The Plant Journal, (2002), 32, 1033–1048
1046 Hideyuki Suzuki et al.

Fulcheri, C., Morard, P. and Henry, M. (1998) Stimulation of thegrowth and the triterpenoid saponin accumulation of Saponariaofficinalis cell and Gypsophila paniculata root suspension cul-tures by improvement of the mineral composition of the media.J. Agric. Food Chem. 46, 2055–2061.
Gundlach, H., Muller, M.J., Kutchan, T.M. and Zenk, M.H. (1992)Jasmonic acid is a signal transducer in elicitor-induced plant cellcultures. Proc. Natl Acad. Sci. USA, 89, 2389–2393.
Haralampidis, K., Bryan, G., Qi, X., Papadopoulou, K., Bakht, S.,Melton, R. and Osbourn, A. (2001) A new class of oxidosqualenecyclase directs synthesis of antimicrobial phytoprotectants inmonocots. Proc. Natl Acad. Sci. USA, 98, 13431–13436.
Haridas, V., Higuchi, M., Jayatilake, G.S., Bailey, D., Mujoo, K.,Blake, M.E., Arntzen, C.J. and Gutterman, J.U. (2001) Avicins:triterpenoid saponins from Acacia victoriae (Bentham) induceapoptosis by mitochondrial perturbation. Proc. Natl Acad. Sci.USA, 98, 5821–5826.
Hayashi, H., Hiraoka, N., Ikeshiro, Y., Kushiro, T., Morita, M.,Shibuya, M. and Ebizuka, Y. (2000) Molecular cloning and char-acterization of a cDNA for Glycyrrhiza glabra cycloartenolsynthase. Biol. Pharm. Bull. 23, 231–234.
Herold, M.C. and Henry, M. (2001) UDP-glucuronosyltransferaseactivity is correlated to saponin production in Gypsophila pani-culata root in vitro cultures. Biotechnol. Lett. 23, 335–337.
Herrera, J.B., Bartel, B., Wilson, W.K. and Matsuda, S.P. (1998)Cloning and characterization of the Arabidopsis thaliana lupeolsynthase gene. Phytochemistry, 49, 1905–1911.
Huhman, D.V. and Sumner, L.W. (2002) Metabolic profiling ofsaponin glycosides in Medicago sativa and Medicago truncatulausing HPLC coupled to an electrospray ion-trap mass spectro-meter. Phytochemistry, 59, 347–360.
Husselstein-Muller, T., Schaller, H. and Benveniste, P. (2001) Mole-cular cloning and expression in yeast of 2,3-oxidosqualene-triterpenoid cyclases from Arabidopsis thaliana. Plant Mol Biol.45, 75–92.
Jandrositz, A., Turnowsky, F. and Hogenauer, G. (1991) The geneencoding squalene epoxidase from Saccharomyces cerevisiae:cloning and characterization. Gene, 107, 155–160.
Jones, M. and Elliott, F.C. (1969) Two rapid assays for saponin inindividual alfalfa plants. Crop Sci. 9, 688–691.
Korth, K.L. and Dixon, R.A. (1997) Evidence for chewing insect-specific molecular events distinct from a general woundresponse in leaves. Plant Physiol. 115, 1299–1305.
Kribii, R., Arro, M., Del Arco, A., Gonzalez, V., Balcells, L.,Delourme, D., Ferrer, A., Karst, F. and Boronat, A. (1997) Cloningand characterization of the Arabidopsis thaliana SQS1 geneencoding squalene synthase – involvement of the C-terminalregion of the enzyme in the channeling of squalene through thesterol pathway. Eur. J. Biochem. 249, 61–69.
Kushiro, T., Shibuya, M. and Ebizuka, Y. (1998) b-Amyrin synthase:cloning of oxidosqualene cyclase that catalyzes the formation ofthe most popular triterpene among higher plants. Eur. J. Bio-chem. 256, 238–244.
Kushiro, T., Shibuya, M., Masuda, K. and Ebizuka, Y. (2000) A novelmultifunctional triterpene synthase from Arabidopsis thaliana.Tetrahedron Lett. 41, 7705–7710.
Laden, B.P., Tang, Y. and Porter, T.D. (2000) Cloning, heterologousexpression, and enzymological characterization of human squa-lene monooxygenase. Arch. Biochem. Biophys. 374, 381–388.
Landl, K.M., Klosch, B. and Turnowsky, F. (1996) ERG1, encodingsqualene epoxidase, is located on the right arm of chromosomeVII of Saccharomyces cerevisiae. Yeast, 12, 609–613.
Leber, R., Landl, K., Zinser, E., Ahorn, H., Spok, A., Kohlwein, S.D.,Turnowsky, F. and Daum, G. (1998) Dual localization of squalene
epoxidase, Erg1p, in yeast reflects a relationship betweenthe endoplasmic reticulum and lipid particles. Mol. Biol. Cell.9, 375–386.
Lee, H.K., Denner-Ancona, P., Sakakibara, J., Ono, T. and Pre-stwich, G.D. (2000) Photoaffinity labeling and site-directedmutagenesis of rat squalene epoxidase. Arch. Biochem. Bio-phys. 381, 43–52.
Marciani, D.J., Press, J.B., Reynolds, R.C., Pathak, A.K., Pathak, V.,Gundy, L.E., Farmer, J.T., Koratich, M.S. and May, R.D. (2000)Development of semisynthetic triterpenoid saponin deri-vatives with immune stimulating activity. Vaccine, 18, 3141–3151.
Massiot, G., Lavaud, C., Le Men-Olivier, L., Van Binst, G., Miller,S.P.F. and Fales, H.M. (1988) Structural elucidation of alfalfaroot saponins by mass spectrometry and nuclear magneticresonance analysis. J. Chem. Soc. Perkin Trans. I, 3071–3079.
McConn, M., Creelman, R.A., Bell, E., Mullet, J.E. and Browse, J.(1997) Jasmonate is essential for insect defense in Arabidopsis.Proc. Natl Acad. Sci. USA, 94, 5473–5477.
Memelink, J., Verpoorte, R. and Kijne, J.W. (2001) ORCAnization ofjasmonate-responsive gene expression in alkaloid metabolism.Trends Plant Sci. 6, 212–219.
Morita, M., Shibuya, M., Kushiro, T., Masuda, K. and Ebizuka, Y.(2000) Molecular cloning and functional expression of triterpenesynthases from pea (Pisum sativum). New alpha-amyrin-produ-cing enzyme is a multifunctional triterpene synthase. Eur. J.Biochem. 267, 3453–3460.
Morita, M., Shibuya, M., Lee, M.S., Sankawa, U. and Ebizuka, Y.(1997) Molecular cloning of pea cDNA encoding cycloartenolsynthase and its functional expression in yeast. Biol. Pharm.Bull. 20, 770–775.
Nagata, T., Tsushida, T., Hamaya, E., Enoki, N., Manabe, S. andNishino, C. (1985) Camellidins, antifugal saponins isolated fromCamellia japonica. Agric. Biol. Chem. 49, 1181–1186.
Nakashima, T., Inoue, T., Oka, A., Nishino, T., Osumi, T. and Hata,S. (1995) Cloning, expression, and characterization of cDNAsencoding Arabidopsis thaliana squalene synthase. Proc. NatlAcad. Sci. USA, 92, 2328–2332.
Noguchi, T., Fujioka, S., Choe, S., Takatsuto, S., Tax, F.E., Yoshida,S. and Feldmann, K.A. (2000) Biosynthetic pathways of brassi-nolide in Arabidopsis. Plant Physiol. 124, 201–209.
Oleszek, W. (1988) Solid phase extraction-fractionation of alfalfasaponins. J. Sci. Food Agric. 44, 43–49.
Oleszek, W. (1997) Alfalfa saponins: structure, biological activity,and chemotaxonomy. In Saponins Used in Food and Agriculture(Waller, G. and Yamasaki, K., eds). New York: Plenum Press, pp.155–170.
Oleszek, W., Junkuszew, M. and Stochmal, A. (1999) Determina-tion and toxicity of saponins from Amaranthus cruentus seeds.J. Agric. Food Chem. 47, 3685–3687.
Oleszek, W. and Jurzysta, M. (1990) High-performance liquidchromatography of alfalfa root saponins. J. Chromatog. 519,109–116.
Oleszek, W., Jurzysta, M., Ploszynski, M., Colquhoun, I.J., Price,K.R. and Fenwick, G.R. (1992) Zahnic acid tridesmoside andother dominant saponins from alfalfa (Medicago sativa L.) aerialparts. J. Agric. Food Chem. 40, 191–196.
Oleszek, W., Price, K.R., Colquhoun, I.J., Jurzysta, M., Ploszynski,M. and Fenwick, G.R. (1990) Isolation and identification of alfalfa(Medicago sativa L.) root saponins: their activity in relation to afungal bioassay. J. Agric. Food. Chem. 38, 1810–1817.
Ono, T. and Bloch, K. (1975) Solubilization and partial charac-terization of rat liver squalene epoxidase. J. Biol. Chem. 250,1571–1579.
� Blackwell Publishing Ltd, The Plant Journal, (2002), 32, 1033–1048
Triterpene saponin biosynthesis 1047

Osbourn, A. (1996) Saponins and plant defence – a soap story.Trends Plant Sci. 1, 4–9.
Pandit, J., Danley, D.E., Schulte, G.K., Mazzalupo, S., Pauly, T.A.,Hayward, C.M., Hamanaka, E.S., Thompson, J.F. and Harwood,H.J., Jr (2000) Crystal structure of human squalene synthase.J. Biol. Chem. 275, 30610–30617.
Papadopoulou, K., Melton, R.E., Leggett, M., Daniels, M.J. andOsbourn, A.E. (1999) Compromised disease resistance in sapo-nin-deficient plants. Proc. Natl Acad. Sci. USA, 96, 12923–12928.
Park, H.J., Kwon, S.H., Lee, J.H., Lee, K.H., Miyamoto, K. and Lee,K.T. (2001) Kalopanaxsaponin A is a basic saponin structure forthe anti-tumor activity of hederagenin monodesmosides. PlantaMed. 67, 118–121.
Pedersen, M.W., Barnes, D.K., Sorensen, E.L. et al. (1976) Effects oflow and high saponin selection in alfalfa on agronomic and pestresistance traits and the interrelationship of these traits. CropSci. 16, 193–199.
Pracros, P. (1988) Mesure de l’activite des saponins de la luzernepas les larves du ver de farine: Tenebrio molitor L. (Coleoptere,Tenebrionidae). II. Recherche des fractions de saponins respon-sables des effects antinutritionnels observes. Agronomie, 8,793–799.
Quackenbush, J., Liang, F., Holt, I., Pertea, G. and Upton, J. (2000)The TIGR gene indices: reconstruction and representation ofexpressed gene sequences. Nucl. Acids Res. 28, 141–145.
Sakakibara, J., Watanabe, R., Kanai, Y. and Ono, T. (1995) Mole-cular cloning and expression of rat squalene epoxidase. J. Biol.Chem. 270, 17–20.
Sambrook, J., Fritsch, E.F. and Maniatis, T. (1989) MolecularCloning: a Laboratory Manual. Cold Spring Harbor, NY: ColdSpring Harbor Laboratory Press.
Schafer, U.A., Reed, D.W., Hunter, D.G., Yao, K., Weninger, A.M.,Tsang, E.W.T., Reaney, M.J.T., MacKenzie, S.L. and Covello, P.S.
(1999) An example of intron junctional sliding in the genefamilies encoding squalene monooxygenase homologues inArabidopsis thaliana and Brassica napus. Plant Mol. Biol. 39,721–728.
Segura, M.J.R., Meyer, M.M. and Matsuda, S.P.T. (2000) Arabi-dopsis thaliana LUP1 converts oxidosqualene to multiple triter-pene alcohols and a triterpene diol. Org. Lett. 2, 2257–2259.
Shibata, N., Arita, M., Misaki, Y., Dohmae, N., Takio, K., Ono, T.and Inoue, K. (2001) Supernatant protein factor, which stimu-lates the conversion of squalene to lanosterol, is a cytosolicsqualene transfer protein and enhances cholesterol biosynth-esis. Proc. Natl Acad. Sci. USA, 98, 2244–2249.
Small, E. (1996) Adaptations to herbivory in alfalfa (Medicagosativa). Can. J. Bot. 74, 807–822.
Tava, A. and Odoardi, M. (1997) Saponins from Medicago spp:chemical characterization and biological activity against insects.In Saponins Used in Food and Agriculture (Waller, G. andYamasaki, K., eds). New York: Plenum Press, pp. 97–109.
Tava, A., Oleszek, W., Jurzysta, M., Berardo, N. and Odoardi, M.(1993) Alfalfa saponins and sapogenins: isolation and quantifi-cation in two different cultivars. Phytochem. Anal. 4, 269–274.
Vogeli, U. and Chappell, J. (1988) Induction of sesquiterpenecyclase and suppression of squalene synthetase activities inplant cell cultures treated with fungal elicitor. Plant Physiol.88, 1291–1296.
Vogt, T. and Jones, P. (2000) Glycosyltransferases in plant naturalproduct synthesis: characterization of a supergene family.Trends Plant Sci. 5, 380–386.
Waller, G.R., Jurzysta, M. and Thorne, R.L.Z. (1993) Allelopathicactivity of root saponins from alfalfa (Medicago sativa L.) onweeds and wheat. Bot. Bull. Acad. Sin. 34, 1–11.
Yamamoto, S. and Bloch, K. (1970) Studies on squalene epoxidaseof rat liver. J. Biol. Chem. 245, 7–16.
� Blackwell Publishing Ltd, The Plant Journal, (2002), 32, 1033–1048
1048 Hideyuki Suzuki et al.