transcriptome analysis of a bacterially induced basal and hypersensitive response of medicago...
TRANSCRIPT

Transcriptome analysis of a bacterially induced basaland hypersensitive response of Medicago truncatula
Zoltan Bozso Æ Nicolas Maunoury Æ Agnes Szatmari ÆPeter Mergaert Æ Peter G. Ott Æ Laszlo R. Zsıros ÆErika Szabo Æ Eva Kondorosi Æ Zoltan Klement
Received: 26 August 2008 / Accepted: 19 April 2009 / Published online: 24 May 2009
� Springer Science+Business Media B.V. 2009
Abstract Research using the well-studied model legume
Medicago truncatula has largely focused on rhizobium
symbiosis, while little information is currently available for
this species on pathogen-induced transcriptome changes.
We have performed a transcriptome analysis of this species
with the objective of studying the basal (BR, no visible
symptoms) and hypersensitive response (HR, plant cell
death) in its leaves at 6 and at 24 h after infection by HR-
negative (hrcC mutant) and HR-inducing Pseudomonas
syringae pv. syringae strains, respectively. Although there
were no visible symptoms at the BR, the alterations in gene
expression were comparable to those found with the HR.
Both responses resulted in the transcriptional alteration of
hundreds of plant genes; however, the responses in the HR
were usually more intense. The reactions to HR-inducing
and HR-negative bacterial strains were significantly over-
lapping. Parallel up- or down-regulation of genes with the
same function occurred frequently. However, some plant
processes were regulated in one direction; for example,
most of the protein synthesis-related genes were activated
and all of the photosynthetic/chloroplast genes were sup-
pressed during BR. The possible roles of several functional
classes (e.g., cell rescue, signaling, defense, cell death, etc.)
of transcriptionally altered genes are discussed. The results
of the comparison with available mycorrhizal and nodule
expression data show that there is a significant overlap
between nodulation and the leaf defense response and
that during the early stage of the nodulation in roots,
Sinorhizobium meliloti induces a fluctuation in the tran-
scription of BR- and HR-responsive genes.
Keywords Basal resistance � Medicago truncatula �Microarray � Mycorrhiza � Nodulation �Pseudomonas syringae
Introduction
Plants are continuously exposed to infections caused by
different microbes, including bacteria, and have, in
response, developed various recognition mechanisms and
resistance responses. The most conspicuous, effective and,
therefore, the most studied plant defense response is the
plant cell death-associated hypersensitive response (HR),
which is a specific plant reaction to pathogens (Klement
1963). In the case of bacterial induced HR, the plant usu-
ally senses bacterial proteins, called effectors, which are
typically injected into the host cells by the bacterial type III
secretion system (TTSS). The important role of effector
proteins is supported by the fact that the integrity of the
TTSS is crucial for both HR induction in incompatible
interactions (no disease development because of resistance)
and for in planta multiplication of pathogen bacteria in the
compatible interactions (with disease development) (Alf-
ano and Collmer 2004). In the incompatible interactions,
the effectors are directly or indirectly recognized by plant
resistance (R) gene product(s), thereby inducing a signal
Electronic supplementary material The online version of thisarticle (doi:10.1007/s11103-009-9496-8) contains supplementarymaterial, which is available to authorized users.
Z. Bozso (&) � A. Szatmari � P. G. Ott �L. R. Zsıros � E. Szabo � Z. Klement
Plant Protection Institute of the Hungarian Academy of Sciences,
Herman O. 15, P.O. Box 102, Budapest, Hungary
e-mail: [email protected]
N. Maunoury � P. Mergaert � E. Kondorosi
Unite Propre de Recherche 2355, Institut des Sciences du
Vegetal, Centre National de la Recherche Scientifique, Avenue
de la Terrasse Batiment 23, 91198 Gif-sur-Yvette Cedex, France
123
Plant Mol Biol (2009) 70:627–646
DOI 10.1007/s11103-009-9496-8

cascade leading to the rapid death of plant cells within 5–
24 h. The HR is accompanied by the accumulation of
reactive oxygen species, including H2O2, and by a number
of other resistance-associated responses, such as the pro-
duction of pathogenesis-related (PR) proteins and phyto-
alexin (Hammond-Kosack and Jones 1996).
In addition to the HR, there is a more general plant
defense response that also recognizes saprophytic bacteria
and bacteria without functional TTSS and subsequently
inhibits their multiplication (Klement et al. 1999, 2003).
There are many synonyms in the literature for this general
recognition and defense system, such as basic, basal, gen-
eral resistance. or innate immunity (Jakobek and Lindgren
1993; de Torres et al. 2003; Gomez-Gomez and Boller
2002). In this paper, we call this general defense system
‘basal resistance’ (BR). The primary elicitors of the BR are
the common components of the microorganisms. These
molecules are also known as pathogen- or microbe-asso-
ciated molecular patterns (PAMPs/MAMPs) (Mackey and
McFall 2006). Flagellin, the protein subunits of flagella,
and the outer membrane-associated lipopolysaccharides
(LPS) of Gram-negative bacteria are the best known sur-
face-derived elicitors of the general response (Dow et al.
2000; Gomez-Gomez and Boller 2002). Disruption of the
cell integrity leading to the leakage of bacterial contents
may also trigger general defense responses. To date, a cold-
shock protein and a bacterial elongation factor have been
identified as endogenous bacterial elicitors of the BR
(Kunze et al. 2004; Felix and Boller 2003). Detection of
bacterial presence induces different defense-associated
physiological and structural changes in plant cells (Best-
wick et al. 1995; Ott et al. 1997; Bozso et al. 2005).
While the BR and HR can be distinguished, this does not
mean that these two plant defense responses are entirely
different. Although their recognition mechanisms and the
final outcome are obviously different, BR and HR use
partly similar or overlapping pathways. Hypersensitive
response-negative or saprophytic bacteria have been found
to be able to activate selected plant defense-related genes
that have been similarly activated by the HR-inducing
strains. The first report demonstrating gene activation
changes by HR-negative bacteria was published by
Jakobek and Lindgren (1993). Thereafter, it was shown that
not only whole bacterial cells but also bacterial compounds
can cause changes in a treated plant’s gene expression
pattern (Dow et al. 2000; Navarro et al. 2004). Genome-
wide transcriptional reprogramming has recently been
described in Arabidopsis thaliana during various plant
responses, including defense reactions and those that are
related to BR following bacterial infection (Thilmony et al.
2006; Truman et al. 2006). However, there is limited
information currently available on changes in gene acti-
vation during BR response in other plants. Our studies have
focused on Medicago truncatula, which has been selected
as a model legume for studying nitrogen-fixing nodule
development induced by the symbiont Sinorhizobium
meliloti. This plant also provides a suitable model for
investigating other plant–bacterial interactions and for
comparing plant reactions to pathogenic, symbiotic, or
mycorrhizal partners.
In this work we report on the transcriptional changes in
BR and HR occurring in M. truncatula leaves at 6 and 24 h
after bacterial inoculation. Our results show that the HR-
negative, BR-inducing Pseudomonas syringae pv. syringae
hrcC mutant elicits a dramatic transcriptional reprogram-
ming of the plant cells, although this reprogramming is less
pronounced than the transcriptional changes in HR induced
by wild-type P. syringae pv. syringae. We discuss the
possible function of particular classes of genes activated
during BR and compare defense and symbiosis related
transcriptional patterns.
Materials and methods
Plant materials and bacteria
Medicago truncatula cv. R108 plants were grown in mixed
soil at 23�C under a 16/8-h light/dark photoperiod in a
growth chamber. Leaves of 4- to 6-week-old plants were
used for infiltration.
Pseudomonas syringae pv. syringae 61 (P. syringae)
HR-positive wild-type strain and P. syringae pv. syringae
61 hrcC (P. syringae hrcC) HR-negative mutant strain
(61-143 1530B, Alan Collmer, Cornell University, Ithaca,
USA) were grown overnight at 27�C on King’s B medium
(King et al. 1954) which, in the case of the P. syringae
hrcC mutant was supplemented with 50 lg/ml kanamycin.
For infiltrations, bacteria were suspended in distilled water
at a density of 108 cells ml-1. The bacterial concentration
of the suspensions was estimated by optical density read-
ings at 600 nm. Infiltrations of M. truncatula leaves with
the bacterial suspension or distilled water were carried out
by injection with a syringe.
Sample preparation, Cy labeling, and microarray
hybridization
The experiments were performed according to the
requirements of the Minimum Information About a
Microarray Experiment (MIAME) (Brazma et al. 2001).
Experimental design
Medicago leaf samples injected with water or with the
bacterial suspension were harvested 6 and 24 h post
628 Plant Mol Biol (2009) 70:627–646
123

inoculation (hpi). For RNA preparation, approximately
100 mg leaf tissue was collected from four different plants,
frozen in liquid nitrogen, and stored at -70�C for future
analysis. Two independent experiments were performed on
leaves of different plant generations, with a 2-week interval
between leaf harvests.
RNA extraction and labeling
Total RNA purification was carried out using the plant total
RNA purification kit (Viogene Biotek, Taipei, Taiwan).
RNA quality was checked on agarose gels. cDNA synthesis
and labeling were performed using the Stratagene Fairplay
II Kit (Statagene, La Jolla, CA) according to the manu-
facturer’s instructions. The bacterial and water-treated
samples were labeled with Cy5 and Cy3, respectively. The
labeled probes were concentrated to 10 ll by means of a
vacuum concentrator. The efficiency of labeling was
checked by running 1 ll samples on a mini agarose gel,
which was photographed in a laser scanner (LS4; Genomic
Solutions, Huntingdon, UK).
Arrays
The M. truncatula 16 kOLI-1 array was used (produced
by the Department of Genetics, Institute of Genome
Research, Bielefeld University, Germany; www.genetic.
uni-bielefeld.de) and prepared for hybridization according
to the manufacturer’s instructions. This array represents
16,086 tentative consensus sequences of the TIGR M.
truncatula Gene index 5 and contains 70-mer oligonu-
cleotide probes in duplicate.
Hybridization conditions
The labeled cDNA samples (9 ll) were diluted with
hybridization buffer (Micromax; Perkin-Elmer, Foster
City, CA) to 120 ll, heated to 95�C for 1 min, loaded on
the prepared array slides, and then processed using an
automated hybridization station (GeneMachines HybSta-
tion; Genomic Solutions, Holliston, MA). The hybridiza-
tion was carried out at 65�C for 3 h, then at 55�C for 3 h,
and finally at 50�C for 12 h. This was followed by a
medium-stringency wash, a high-stringency wash, and a
wash with post-wash buffer at 42�C, each at room tem-
perature for 5 min. Wash buffers were from Genomic
Solutions.
Measurements
The hybridization signals were measured by scanning the
slides with a GenePix 4000B scanner piloted with GenePix
Pro software (Molecular Devices, Sunnyvale, CA).
Normalization
Data processing and normalization were performed with
Midas software (Saeed et al. 2003). Before data analysis,
the data files produced by GenePix Pro were converted
with an Express Converter (from the TIGR website) to a
MIDAS compatible tav file. After local background sub-
tractions, the signal intensities were normalized by the
Lowess (Lockfitt) method using default parameters of the
software (block mode, smoothing parameter 0.33, refer-
ence Cy3).
Data analysis
The spots that showed low intensity in both channels
(\5000; this level was about 50% more intense than the
values of the highest level of the empty control spots) were
omitted from further analyses because these spots usually
show high expression variability and, therefore, are a major
cause of inconsistencies in microarray results. The expres-
sion data of duplicated probes were averaged for both
channels and the expression ratios determined. Using the
results from duplicated experiments, we further analyzed
those genes showing at least a twofold up- or down-regu-
lation in both replicates (Supplementary Table 1) To con-
firm the result of differentially expressed genes, we
performed Rank Products analysis using the appropriate
software (from the Glasgow Microarray Analysis website).
The Rank Products Method gives reliable and consistent
results even in the case of a reduced number of repeats
(Breitling et al. 2004). Because at 6 hpi those genes that had
been activated or repressed in both P. syringae hrcC- and
P. syringae-injected leaves changed their activity exclu-
sively in the same direction, the two independent data sets
of these treatments (four data sets) could be used as a repeat
and analyzed in terms of significance. Ten random permu-
tations were used for estimating false discovery rates
(FDR). The genes that were above the selection limit (10%
FDR) by the rank analysis were compared to the list of the
genes that showed at least a twofold up- or down-regulation
in both wild-type P. syringae- and hrcC mutant-injected
leaves. The results of the analyses showed that at 10% FDR
almost all (96%) common genes that were scored as dif-
ferentially expressed on the basis of fold-method (C2,
B0.5) were also scored as being differentially regulated by
the Rank Product analyses (Supplementary Table 2).
Quantitative RT-PCR analysis of gene expression
Total RNA (2.5 lg) was used for the synthesis of 20 ll
cDNA (RevertAid H Minus First Strand cDNA Synthesis
kit; Fermentas, Vilnius, Lithuania). Specific primers were
designed for both up- and down-regulated genes expressed
Plant Mol Biol (2009) 70:627–646 629
123

at different levels. The primers and cycling parameters for
real-time PCR are given in Supplementary Table 3. For the
PCR analyses, 2.5 ll from a tenfold dilution of the cDNA
stock were used (prepared as described above) in 15-ll
reactions. The final concentrations of the primers was
3 lM. The PCR was performed using the iQ SYBR Green
2x Supermix (Bio-Rad, Hercules, CA) on the DNA Engine
Opticon 2 (MJ Research, Waltham, MA). Measured C(T)
values were always normalized to actin (accession no.
TC81607) internal control values. Each sample was mea-
sured at least twice, and standard deviations were calcu-
lated. Values of the water-inoculated controls were given
the arbitrary value of 1; other sample values were corre-
lated to the water controls.
Results and discussion
Experimental design and data analysis
To obtain an overall view of the transcriptional changes
that occur during BR and HR induced by bacteria in plant
cells, we performed microarray experiments with M.
truncatula 16 kOLI chips. Medicago truncatula leaves
were injected with the HR-negative TTSS mutant P.
syringae hrcC, which induces only BR, and the wild-type
P. syringae, which triggers both BR and HR. The response
of the leaves to the injected bacterial samples was com-
pared to the water-infiltrated control samples. To discrim-
inate plant responses related to mechanical stress caused by
the infiltration procedure, we also compared the response
of the water-infiltrated leaves to non-inoculated leaf sam-
ples. The leaf samples were collected at 6 and at 24 hpi,
except for leaves infiltrated with the wild-type P. syringae,
which could only be harvested at 6 hpi as thereafter (at
about 7–8 hpi) a tissue collapse occurred due to the HR.
Infiltration of P. syringae hrcC induced no visible symp-
toms. The leaf RNA samples were hybridized to 16k M.
truncatula microarrays. Following data normalization and
filtering (see Materials and methods), those genes showing
at least a twofold change in expression, either activation or
repression (C2, B0.5), as a consequence of the treatments
were selected as being differentially expressed (Supple-
mentary Table 1).
Bacterial infiltration resulted in major changes in gene
activities while water infiltration resulted in only minor
changes. The number of overlapping genes up- or down-
regulated by the bacterial and water-injected samples were
low (e.g., at 6 hpi, only 27 common genes were found
between the water- and P. s. hrcC-infiltrated samples),
which showed that most of the changes were caused by the
bacterial treatments (data not shown). The results obtained
by the microarray analysis were confirmed with real-time
PCR for 15 selected genes (Supplementary Table 4).
Although there were differences between microarray and
PCR data in terms of the intensity of the transcriptional
alterations, the direction of changes was consistent, which
confirmed the reliability of the microarray results.
Comparison of the different bacterial treatments
Despite the absence of symptoms, the injection of the HR-
minus P. syringae hrcC provoked a dramatic change in the
transcription profile by activating 781 genes and repressing
1116 ones at 6 hpi; this response was comparable to
the changes induced by the HR-positive bacteria, which
triggered the up-regulation of 705 genes and the down-
regulation of 845 genes (Table 1). At 6 hpi, the up- or
down-regulated genes of these treatments represented
about 10–12% of the genes present on the microarray; in
comparison, approximately 38–40% of the genes did not
show any significant changes in their activity and 50%
were undetectable in our experimental conditions. There
was a significant overlap (700 genes at 6 hpi) between BR
and HR. Moreover, the directions of these gene expression
modifications were the same during both response, with the
exception of three genes: TC 87010, TC 77756 and
TC86996 (Figs. 1a, 2). Among the genes that showed
modified activation only after a particular treatment could
be the genes that are specific for HR or BR. However, more
detailed time course experiments would be required to
confirm their specificity for HR or BR.
The decline of the numbers of significantly modified
genes (Table 1) and that of the fold-change of genes
(Fig. 1b) revealed that the transcriptional response
decreased from 6 to 24 hpi in leaves injected with P. sy-
ringae hrcC. Several genes showed significant changes in
activity at both 6 and 24 hpi (Figs. 1b, 2).
Classification of transcriptionally modified genes
induced during BR
Those genes that significantly changed their activity during
BR were classified into functional groups based on the
classification of the Institute for Genomic Research TC
annotation and the MENS (Medicago EST Navigation Sys-
tem) on-line databases. The largest group is composed of
genes without known functions or similarities to other genes
(e.g., 35% at 6 hpi in hrcC mutant-injected leaves). Most
groups contain both activated and repressed genes. However,
certain groups, such as photosynthesis/chloroplast and
ribosomal/protein synthesis genes, changed their transcrip-
tional activity specifically in one direction. At 6 hpi in leaves
injected with P. syringae hrcC, all photosynthesis/chlor-
oplast-related genes (188) were down-regulated (except for
one, ferrodoxin, TC77615; data not shown). In contrast, in
630 Plant Mol Biol (2009) 70:627–646
123
these samples, most of the ribosomal/protein synthesis-
related genes were activated (75 activated and seven
repressed) (Supplementary Table 5). The tendency of these
changes was in agreement with the results obtained after
comparable bacterial treatments in other plants, such as
Arabidopsis and tomato (Mysore et al. 2002; Truman et al.
2006). Certain of the responsive genes behaved as expected,
based on knowledge from other screening methods and other
pathosystems, such as PR genes, receptor-like kinases rec-
ognizing MAMPS, transcription factors, among others.
Cell signaling
The recognition of infecting bacteria by plant cells occurs
in at least two steps. The first step is the fast recognition of
bacterial MAMPs by cell-surface receptors. The second
step is the sensing of the injected bacterial effector proteins
inside the plant cells. The period required for building up
the TTSS system of bacteria is the primary determinant of
the length of this second step. In the case of P. syringae
strain 61, the length of this period (the so-called bacterial
induction time), is about 0.5–1 h (Bozso et al. 1999).
Although our samples were taken after these recognition
events, there were numerous changes in the expression of
signaling genes (Table 2). Transcriptional regulation of
signaling components could be a mechanism for enhancing
or weakening the intensity of defense reactions.
Kinase genes were the most abundant and the most
activated genes in this functional class at 6 hpi for both
types of bacterial treatments, suggesting that the kinases
8
10
8
10(A)
1
P.s. at 6 hpi(B)
1
P.s. hrcC at 24 hpi
4
6
8
4
6
8
-2
0
2
-10 -8 -6 -4 -2 0 2 4 6 8 10
-2
0
2
-10 -8 -6 -4 -2 0 2 4 6 8 10
P.s. hrcC at 6 hpi P.s. hrcC at 6 hpi
-8
-6
-4
-8
-6
-4
-10-103 4 3 4
2 2
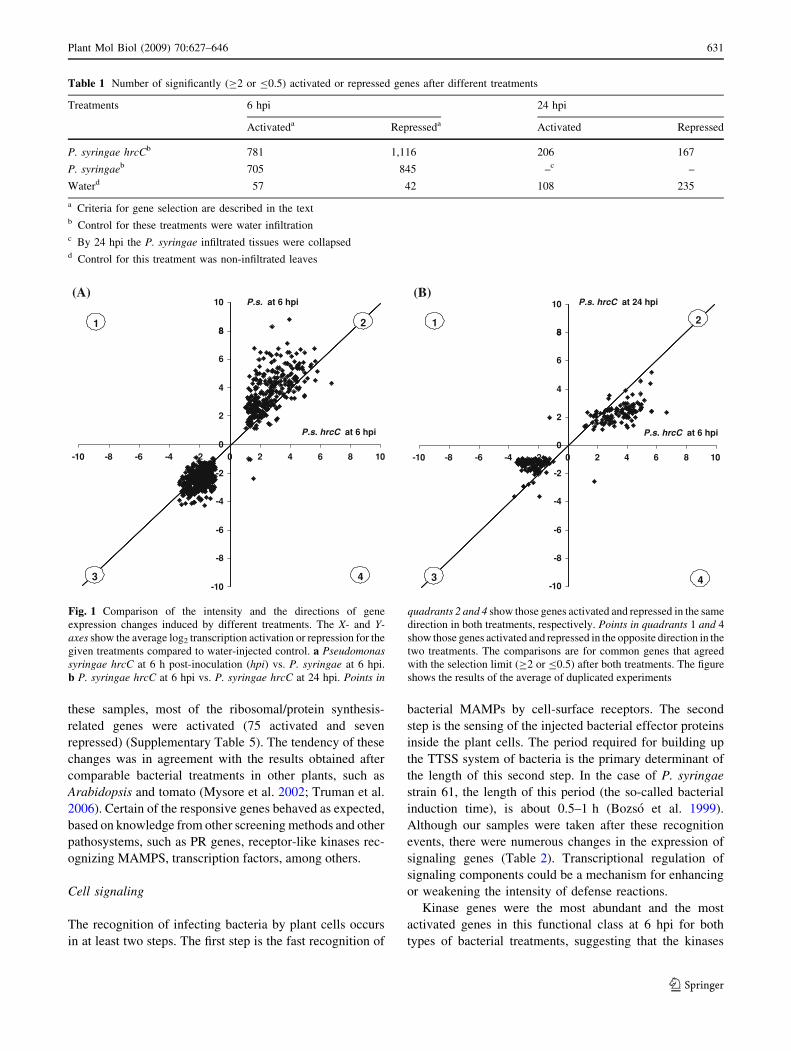
Fig. 1 Comparison of the intensity and the directions of gene
expression changes induced by different treatments. The X- and Y-axes show the average log2 transcription activation or repression for the
given treatments compared to water-injected control. a Pseudomonassyringae hrcC at 6 h post-inoculation (hpi) vs. P. syringae at 6 hpi.
b P. syringae hrcC at 6 hpi vs. P. syringae hrcC at 24 hpi. Points in
quadrants 2 and 4 show those genes activated and repressed in the same
direction in both treatments, respectively. Points in quadrants 1 and 4
show those genes activated and repressed in the opposite direction in the
two treatments. The comparisons are for common genes that agreed
with the selection limit (C2 or B0.5) after both treatments. The figure
shows the results of the average of duplicated experiments
Table 1 Number of significantly (C2 or B0.5) activated or repressed genes after different treatments
Treatments 6 hpi 24 hpi
Activateda Represseda Activated Repressed
P. syringae hrcCb 781 1,116 206 167
P. syringaeb 705 845 –c –
Waterd 57 42 108 235
a Criteria for gene selection are described in the textb Control for these treatments were water infiltrationc By 24 hpi the P. syringae infiltrated tissues were collapsedd Control for this treatment was non-infiltrated leaves
Plant Mol Biol (2009) 70:627–646 631
123

are one of the most important regulators of these resistance
responses. Several phosphatase genes were also up- or
down-regulated. While, at 6 hpi in P. syringae hrcC-
injected leaves, different phosphatase 2A subunits were
found to be activated, protein phosphatase 2C genes were
repressed. Phosphatase 2A (PP2Ac) in tomato and tobacco
is a negative regulator of plant defense responses by
influencing the PR-protein level and the development of
cell death (He et al. 2004). Protein phosphatase 2C proteins
can be negative regulators of stress-induced MAPK path-
ways, abscisic acid (ABA) signaling, and receptor signal-
ing (Schweighofer et al. 2004). Activated phosphatases
may have a role in feed-back regulation, whereas the
repression of other phosphatases may be required for the
activation of other plant defense responses.
Many receptor-like protein kinases were found among
the up- and down regulated genes. These receptors may
play a key role in sensing the presence of the bacterial
cells. In Arabidopsis, FLS2 (At5g64330) is responsible for
the recognition of a conserved region of bacterial flagellin
(Gomez-Gomez and Boller 2002). The probable homolog
of FLS2 in M. truncatula, TC89068 (67% of identity), was
activated at 6 hpi in P. syringae hrcC-treated leaves. Some
members of the S receptor-like kinase (self-incompatibil-
ity) gene family showed transcriptional activation in tissues
infected with HR-negative bacteria, suggesting that they
may be involved in the development of BR (Pastuglia et al.
2002). Although the activated M. truncatula S receptor-like
kinase TC93750 does not show close similarity to the
above-mentioned S gene receptors (data not shown), it
could have a role in pathogen perception.
Pseudomonas syringae hrcC treatments also activated
receptor-like genes of the yeast pheromone class. These
types of G protein-coupled receptors are peptide hormone-
sensing molecules in yeast (Bardwell 2005). The two
activated M. truncatula pheromone receptor-like genes
(TC7717, TC87246) are similar (E value = 1e-18 and
3e-38, respectively) to two Arabidopsis genes (At2g26530
and At2G15760), which complemented the yeast phero-
mone receptor mutants (Hirayama et al. 1997). The
microarray data of these Arabidopsis genes (data from
Genevestigator website) and the MtGI expressed sequence
tag (EST) data of the Medicago homologs (data from
M. truncatula gene index website) indicate the activation
of these genes during different defense-related processes
(data not shown).Thus, it can not be excluded that the
pheromone receptor-like genes may have signaling roles in
resistance responses that include the BR.
Transcription factors
Several members of WRKY, MYB, NAC/NAM, bZIP, and
AP2 transcription factors (TFs) were activated, and other
MYB, MYB-like, bZIP, and bHLH TFs were repressed in
P. syringae hrcC-infiltrated leaves at 6 hpi (Table 3).
These types of TFs are involved in various pathogen- or
elicitor-induced resistance reactions (Eulgem 2005).
To determine whether the transcriptional changes are
conserved among different plant species, we compared the
expression of M. truncatula and Arabidopsis TF homologs.
The Arabidopsis homologs of the Medicago TFs were
identified by BlastTX (available from the TAIR website)
(Table 3). The expression of these Arabidopsis genes in
response to various treatments was collected with the
Genevestigator website (Zimmermann et al. 2004). Table 3
shows that the Arabidopsis homologs were activated or
repressed after various microbial infections and other bio-
logical stresses and that they mostly changed in the same
direction as their M. truncatula homologs. Pseudomonas and
Agrobacterium triggered similar responses in Arabidopsis,
which suggests the general nature of the BR. The plant cell
death-inducing necrotroph fungi Alternaria brassicola and
Botrytis cinerea—but not the biotrophic Erysiphe species—
activated many of these TF homologs, implying that the
plant response after bacterial infection resembles, to a cer-
tain extent, the reaction to necrotroph pathogens.
Most of the Arabidopsis homologs of down-regulated
M. truncatula TFs were also repressed in response to
bacterial treatments, and no homologs were regulated in
Fig. 2 Quantitative comparison of gene expression showing the
number of transcriptionally modified genes in wild-type P. syringae(P.s. 6 hpi) and in P. syringae hrcC mutant (P.s. hrcC 6 and 24 hpi)
infiltrated tissues. Numbers indicate the number of genes influenced in
common (overlapped areas) or separately (not overlapping areas).
The areas are proportional to the respective numbers. Numbers inparentheses represent activated and repressed genes, respectively,
followed by oppositely influenced genes, when applicable. The figure
represents the results of duplicated experiments. Criteria for gene
selection are described in the text
632 Plant Mol Biol (2009) 70:627–646
123
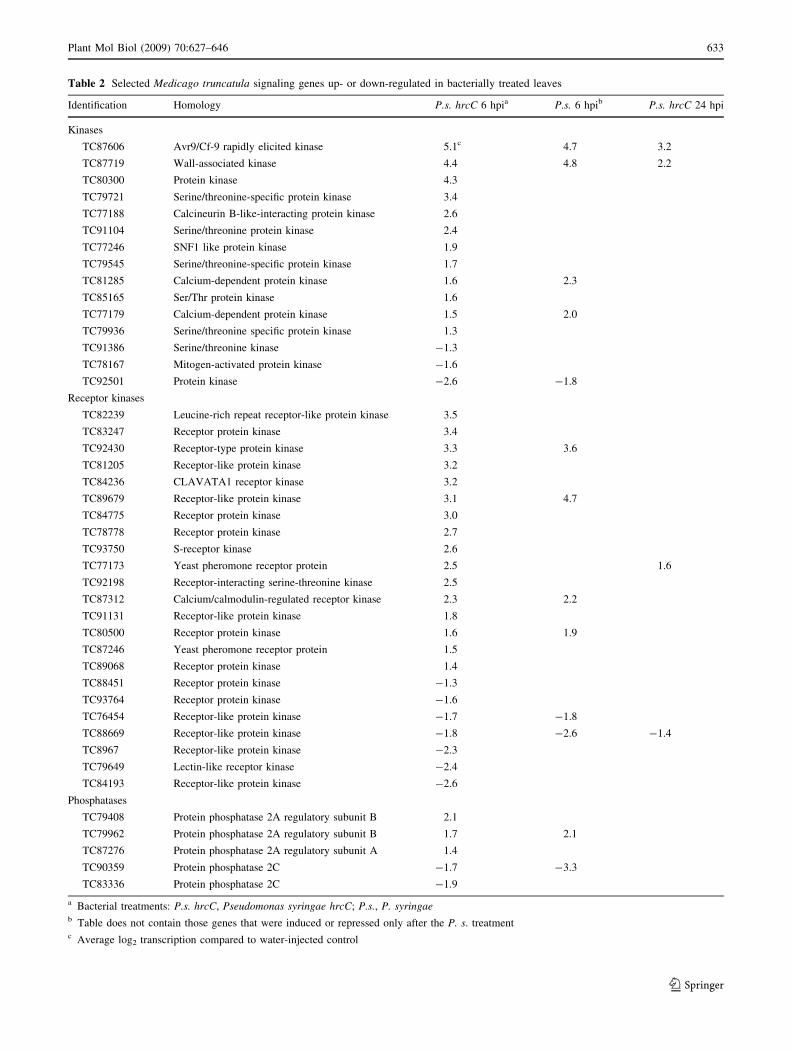
Table 2 Selected Medicago truncatula signaling genes up- or down-regulated in bacterially treated leaves
Identification Homology P.s. hrcC 6 hpia P.s. 6 hpib P.s. hrcC 24 hpi
Kinases
TC87606 Avr9/Cf-9 rapidly elicited kinase 5.1c 4.7 3.2
TC87719 Wall-associated kinase 4.4 4.8 2.2
TC80300 Protein kinase 4.3
TC79721 Serine/threonine-specific protein kinase 3.4
TC77188 Calcineurin B-like-interacting protein kinase 2.6
TC91104 Serine/threonine protein kinase 2.4
TC77246 SNF1 like protein kinase 1.9
TC79545 Serine/threonine-specific protein kinase 1.7
TC81285 Calcium-dependent protein kinase 1.6 2.3
TC85165 Ser/Thr protein kinase 1.6
TC77179 Calcium-dependent protein kinase 1.5 2.0
TC79936 Serine/threonine specific protein kinase 1.3
TC91386 Serine/threonine kinase -1.3
TC78167 Mitogen-activated protein kinase -1.6
TC92501 Protein kinase -2.6 -1.8
Receptor kinases
TC82239 Leucine-rich repeat receptor-like protein kinase 3.5
TC83247 Receptor protein kinase 3.4
TC92430 Receptor-type protein kinase 3.3 3.6
TC81205 Receptor-like protein kinase 3.2
TC84236 CLAVATA1 receptor kinase 3.2
TC89679 Receptor-like protein kinase 3.1 4.7
TC84775 Receptor protein kinase 3.0
TC78778 Receptor protein kinase 2.7
TC93750 S-receptor kinase 2.6
TC77173 Yeast pheromone receptor protein 2.5 1.6
TC92198 Receptor-interacting serine-threonine kinase 2.5
TC87312 Calcium/calmodulin-regulated receptor kinase 2.3 2.2
TC91131 Receptor-like protein kinase 1.8
TC80500 Receptor protein kinase 1.6 1.9
TC87246 Yeast pheromone receptor protein 1.5
TC89068 Receptor protein kinase 1.4
TC88451 Receptor protein kinase -1.3
TC93764 Receptor protein kinase -1.6
TC76454 Receptor-like protein kinase -1.7 -1.8
TC88669 Receptor-like protein kinase -1.8 -2.6 -1.4
TC8967 Receptor-like protein kinase -2.3
TC79649 Lectin-like receptor kinase -2.4
TC84193 Receptor-like protein kinase -2.6
Phosphatases
TC79408 Protein phosphatase 2A regulatory subunit B 2.1
TC79962 Protein phosphatase 2A regulatory subunit B 1.7 2.1
TC87276 Protein phosphatase 2A regulatory subunit A 1.4
TC90359 Protein phosphatase 2C -1.7 -3.3
TC83336 Protein phosphatase 2C -1.9
a Bacterial treatments: P.s. hrcC, Pseudomonas syringae hrcC; P.s., P. syringaeb Table does not contain those genes that were induced or repressed only after the P. s. treatmentc Average log2 transcription compared to water-injected control
Plant Mol Biol (2009) 70:627–646 633
123

the opposite direction. Thus, TFs involved in defense
mechanisms against different pathogens may be conserved.
Cell rescue
One of the most important challenges for a plant cell after
microbial infection is to eliminate the various induced and
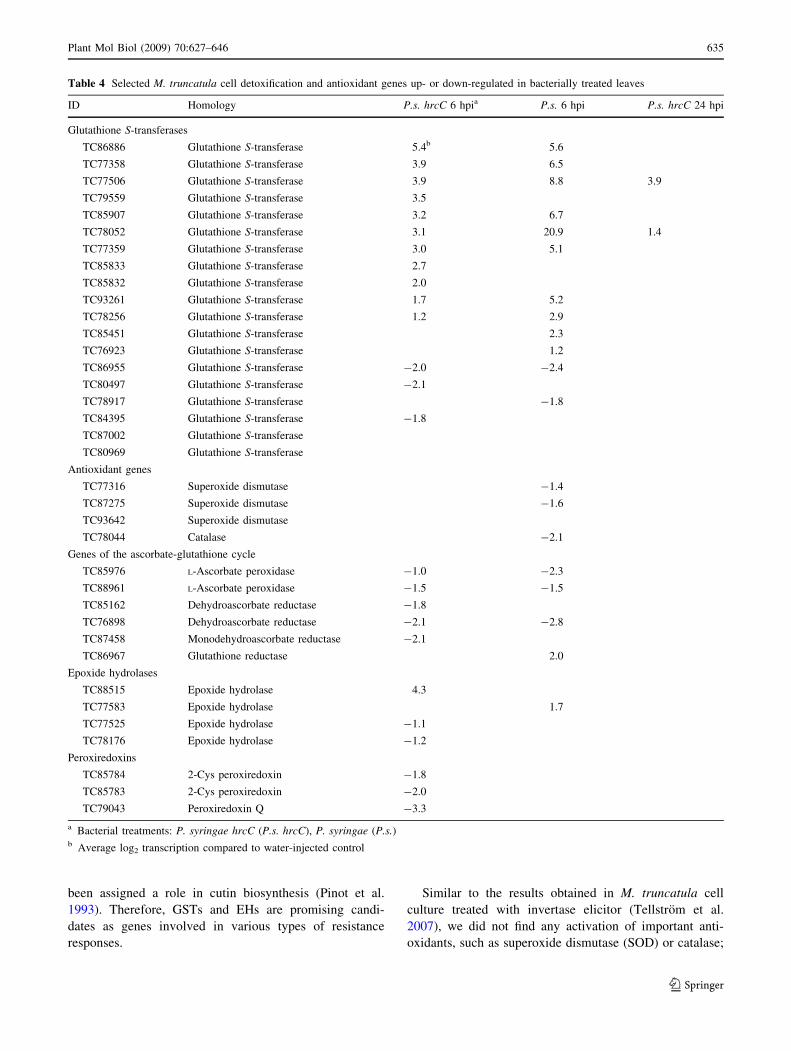
harmful processes that may damage cell integrity. Gluta-
thione S-transferases (GSTs) and epoxide hydrolases (EHs)
are key enzymes of detoxification (Sheehan et al. 2001;
Newman et al. 2005). Both activated and repressed GSTs
and EHs were found, and the GSTs were particularly
abundant (Table 4). It has been reported that the levels of
GST isoenzymes are regulated both transcriptionally and
translationally during bacterially induced BR and HR
(Jones et al. 2004). At 6 hpi, 10 and 11 GST isoenzyme
genes were activated by P. syringae and P. syringae hrcC
respectively, and the activation was stronger in the first
case. On the other hand, only 2 and 3 (P. syringae and P.
syringae hrcC respectively) GST isoenzyme genes were
repressed. Epoxide hydrolases play a critical role in
detoxifying reactive metabolites in both plants and animals
(Murray et al. 1993). The tobacco EH gene was found to be
activated during BR after injection of the P. syringae hrcC
mutant (Szatmari et al. 2006) and by tobacco mosaic virus
infection (Guo et al. 1998). Additionally, EHs have also
Table 3 Comparison of the expression of M. truncatula basal resistance-related transcription factors with their Arabidopsis homologs
IDType of transcription
factorArabidopsis
homolog genes E-value A. b
rass
icio
la a
A. t
umef
acie
ns
B. c
iner
ea
E. c
icho
race
arum
E. o
ront
ii
F. o
ccid
enta
lis
M. p
ersi
cae
nem
atod
e
P. i
nfes
tans
P. r
apae
P.
tom
ato
hrcC
b
P.
tom
atob
P.
tom
ato
avrR
pm1b
P.
phas
eoli
cola
P.s
.hrc
C
6 hp
i
P.s
. 6
hpi
Activation or repression of TF’s after different treatments TF’s activated in P.s. hrcC injected
leaves of M. truncatula in Arabidopsis in M.
truncatulaTC78376 WRKY AT1G80840 1,00E-51 Ac A A R A A A A A A ATC80208 WRKY AT4G18170 4,00E-39 A A A A A A A A A A A A A ATC86084 WRKY AT2G24570 5,00E-87 A A A A A A
TC89937 AP2 AT5G61890 2,00E-28 A A A A A A A A TC87696 AP2 AT4G23750 9,00E-28 A A R R R R R R R A TC88428 MYB AT1G66230 2,00E-73 R R R A R R A A TC86588 MYB AT1G06180 2,00E-63 A A A A A A A ATC78189 NAC/NAM AT1G01720 1,00E-97 A A R A A A A A TC88609 NAC/NAM AT1G69490 3,00E-85 R A A R R A R A A A A ATC81706 Zinc finger AT3G28210 5,00E-64 A A A A A A A ATC77581 bHLH AT1G32640 2,00E-64 R A A R A A A A TC80273 At-hook AT5G49700 2,00E-52 A A A R A R A A A A ATC85773 bZIP AT4G34590 1,00E-29 A A R A A
TC77604 SAR AT5G27120 0.0 A ATF’s repressed in P.s. hrcC
injected leaves of M. truncatula
TC80810 ccaat-box binding AT5G63470 1,00E-71 A R R RTC89017 Zinc finger AT1G68520 2,00E-33 R A R R R R R RTC87014 golden2-like AT2G20570 2,00E-28 R R R R R R RTC87249 bHlH AT4G00050 3,00E-22 R R R R RTC88329 squamosa AT2G47070 2,00E-85 R R RTC80869 bZIP AT1G49720 5,00E-35 A R
TC86792 pseudo-response
regulator AT5G02810 e-128 R R R R R TC77052 MYB AT1G74840 1,00E-44 R RTC78276 MYB AT3G46130 4,00E-52 R R R R R R R R R
BR, Basal resistance; TF, transcription factorsa A. brassiciola, Alternaria brassiciola; A. tumefaciens, Agrobacterium tumefaciens; B. cinereas, Botrytis cinerea; E. cichoracearum, Erysiphecichoracearum; E. orontii, Erysiphe orontii; F. occidentalis, Frankliniella occidentalis (thrips); M. persicae, Myzus persicae (aphids); nema-tode, Heterodera. schachti; P. infestans, Phytophthora. infestans; P. rapae, Pieris rapae (caterpillars); P. tomato hrcC, P. syringae pv. tomatoDC3000 hrcC; P. tomato, P. syringae pv. tomato DC3000; P. tomato avrRpm1, P. syringae pv. tomato DC3000 avrRpm1; P. phaseolicola,
P. savastanoi pv. phaeolicola; P.s. hrcC, P. syringae pv. syringae 61 hrcC; P.s., P. syringae pv. syringae 61b In the case of P. s. tomato, the given gene is considered as activated or repressed if the particular gene showed up- or down-regulation at least
at one presented time point (2, 6, 24 hpi)c A, Activated (C2); R, repressed (B0.5). Gray shading indicates if the direction of transcription of Arabidopsis genes was the same as that of the
M. truncatula TFs
634 Plant Mol Biol (2009) 70:627–646
123
been assigned a role in cutin biosynthesis (Pinot et al.
1993). Therefore, GSTs and EHs are promising candi-
dates as genes involved in various types of resistance
responses.
Similar to the results obtained in M. truncatula cell
culture treated with invertase elicitor (Tellstrom et al.
2007), we did not find any activation of important anti-
oxidants, such as superoxide dismutase (SOD) or catalase;
Table 4 Selected M. truncatula cell detoxification and antioxidant genes up- or down-regulated in bacterially treated leaves
ID Homology P.s. hrcC 6 hpia P.s. 6 hpi P.s. hrcC 24 hpi
Glutathione S-transferases
TC86886 Glutathione S-transferase 5.4b 5.6
TC77358 Glutathione S-transferase 3.9 6.5
TC77506 Glutathione S-transferase 3.9 8.8 3.9
TC79559 Glutathione S-transferase 3.5
TC85907 Glutathione S-transferase 3.2 6.7
TC78052 Glutathione S-transferase 3.1 20.9 1.4
TC77359 Glutathione S-transferase 3.0 5.1
TC85833 Glutathione S-transferase 2.7
TC85832 Glutathione S-transferase 2.0
TC93261 Glutathione S-transferase 1.7 5.2
TC78256 Glutathione S-transferase 1.2 2.9
TC85451 Glutathione S-transferase 2.3
TC76923 Glutathione S-transferase 1.2
TC86955 Glutathione S-transferase -2.0 -2.4
TC80497 Glutathione S-transferase -2.1
TC78917 Glutathione S-transferase -1.8
TC84395 Glutathione S-transferase -1.8
TC87002 Glutathione S-transferase
TC80969 Glutathione S-transferase
Antioxidant genes
TC77316 Superoxide dismutase -1.4
TC87275 Superoxide dismutase -1.6
TC93642 Superoxide dismutase
TC78044 Catalase -2.1
Genes of the ascorbate-glutathione cycle
TC85976 L-Ascorbate peroxidase -1.0 -2.3
TC88961 L-Ascorbate peroxidase -1.5 -1.5
TC85162 Dehydroascorbate reductase -1.8
TC76898 Dehydroascorbate reductase -2.1 -2.8
TC87458 Monodehydroascorbate reductase -2.1
TC86967 Glutathione reductase 2.0
Epoxide hydrolases
TC88515 Epoxide hydrolase 4.3
TC77583 Epoxide hydrolase 1.7
TC77525 Epoxide hydrolase -1.1
TC78176 Epoxide hydrolase -1.2
Peroxiredoxins
TC85784 2-Cys peroxiredoxin -1.8
TC85783 2-Cys peroxiredoxin -2.0
TC79043 Peroxiredoxin Q -3.3
a Bacterial treatments: P. syringae hrcC (P.s. hrcC), P. syringae (P.s.)b Average log2 transcription compared to water-injected control
Plant Mol Biol (2009) 70:627–646 635
123

to the contrary, these genes were repressed in the
P. syringae-infiltrated leaves at 6 hpi. Genes of the ascor-
bate–glutathione cycle, which is another important plant
antioxidant system, were also mostly down-regulated at
6 hpi in bacterially infiltrated leaves. At 6 hpi in the
P. syringae hrcC mutant-treated leaves, repression of three
peroxiredoxin genes was also found. Peroxiredoxins can
reduce H2O2 and alkyl peroxides to water and alcohols,
respectively (Dietz 2003). In Arabidopsis leaves, regulation
of peroxiredoxin at the protein level is complex and, sim-
ilarly to that of GSTs, it goes both up and down at the
transcriptional and protein level after bacterial treatments
(Jones et al. 2004). In our experiments, all peroxiredoxin
genes on the Medicago chip were repressed. It is possible
that the different experimental systems (plant and sampling
time) or the absence of the probes of the isoforms on the
16kOLI chip caused this difference with Arabidopsis
results.
Defense-associated genes
A large portion of genes showing altered activation in
response to bacterial treatments demonstrated a similarity
to genes known to be associated directly or indirectly with
plant defense responses (Table 5). In our samples, several
PR-10 and PR-10-like genes were abundantly and strongly
activated. The cytosol localized PR-10 proteins are induced
by phytopathogens and environmental stresses (Breda et al.
1996; Liu and Ekramoddoullah 2006). In addition to their
ribonuclease activity (Bantignies et al. 2000), the PR-10
and PR-10-related proteins can bind plant hormones, such
as brassinosteroids (Markovic-Housley et al. 2003) and
cytokinins (Koistinen et al. 2005). Both hormones can
induce or influence disease resistance (Nakashita et al.
2003; Carimi et al. 2003; Novacky 1972). Thus, the
accumulation of PR-10 may have multiple effects in plant
defense including a role in BR.
Other PR-proteins, such as chitinases, also were up- and
down-regulated in bacterially treated leaves (Table 5).
Some chitinases hydrolyze not only chitin but also bacterial
cell-wall peptidoglycans. The induction of such chitinase–
lysozyme activities in the tobacco cell wall was found to
correlate well with the BR (Ott et al. 2006) and overex-
pression of a particular chitinase gene in pepper, resulting
in enhanced bacterial disease resistance against P. syringae
(Hong and Hwang 2006).
Photoassimilate-responsive (PAR)-like genes (TC77234
and TC77235) are examples of the crosstalk between
sugar metabolism and defense responses against pathogens
that were up-regulated in bacterially infiltrated samples
(Herbers et al. 1995; Takemoto et al. 2003; Szatmari et al.
2006). Genes that are involved in ethylene and polyamine
synthesis (S-adenosyl-L-methionine synthetases) were also
activated both in P. syringae hrcC- and P. syringae-treated
leaves at 6 hpi. Interestingly, other key enzymes of ethyl-
ene synthesis, such as ACC synthase, and ACC oxidase,
were only up-regulated by the HR-inducing strain (data not
shown). Despite the general constitutive nature of R-pro-
teins (resistance genes), some nucleotide binding-type R
proteins were also activated at 6 hpi in Pseudomonas-
infiltrated leaves.
Examples of down-regulated defense-related genes are
genes of the nonspecific lipid-transfer protein (nsLTP)
class which have proven antibacterial and antifungal
activity (Garcia-Olmedo et al. 1995). Their repression
suggests, however, that the nsLTPs have no direct anti-
bacterial role in Medicago BR (data not shown).
Plant cell death-related genes
Plant cell death is the most characteristic response during
HR-type resistance. The cell death process following
elicitor- or pathogen-induced defense is not an uncon-
trolled cell collapse but a genetically determined and pro-
grammed cell death (PCD) (Greenberg and Yao 2004).
At 6 hpi, we found both up- and down-regulated cell
death-associated genes in leaves injected with Pseudomo-
nas strains. The genes of QM-like protein (TC88241) and
pirin (TC88605) were moderately induced in P. syringae
hrcC-treated samples (about three to fivefold), but they
were more induced in tissues undergoing the HR (approx.
30- to 44-fold). The ESTs of the QM-like protein gene in
the MtGI database (M. truncatula gene index website) were
most frequently isolated from elicitor-treated roots, con-
firming the activation of this gene in defense responses.
This protein is also known as 60S ribosomal protein L10.
The high induction of the QM-like gene (about 30-fold
activation during HR), compared to the low induction of
other ribosome structural genes (two- to three-fold activa-
tion; Supplementary Table 5) suggests that this gene may
have additional role(s) besides its ribosomal function. In
animal cells, the QM-like proteins are involved in cell
death via interaction with transcription factors (Oh et al.
2002). A tomato QM-like protein has been found to confer
protection to yeast against oxidative stress, and it can
rescue yeast from the lethality mediated by the mammalian
proapoptotic protein Bax (Chen et al. 2006).
Pirin is conserved from prokaryotic to eukaryotic
organisms, and their homologs play important roles in a
number of different biological processes, including apop-
tosis. In plants, the function of pirin(s) has not yet been
elucidated. A tomato pirin is induced during PCD, while
PCD inhibitors reduce the pirin transcription (Orzaez et al.
2001). Pirin may take part in signaling (Lapik and Kauf-
man 2003) and transcriptional regulation via interaction
with other proteins (Wendler et al. 1997), but it may also
636 Plant Mol Biol (2009) 70:627–646
123
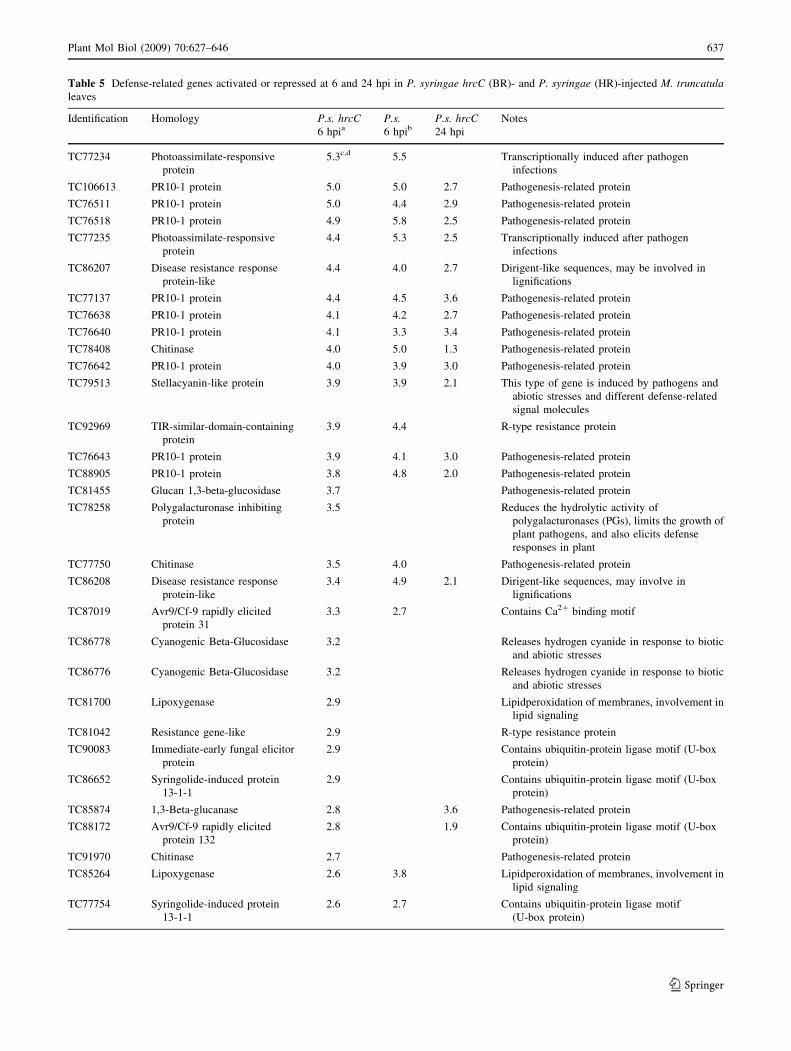
Table 5 Defense-related genes activated or repressed at 6 and 24 hpi in P. syringae hrcC (BR)- and P. syringae (HR)-injected M. truncatulaleaves
Identification Homology P.s. hrcC6 hpia
P.s.6 hpib
P.s. hrcC24 hpi
Notes
TC77234 Photoassimilate-responsive
protein
5.3c,d 5.5 Transcriptionally induced after pathogen
infections
TC106613 PR10-1 protein 5.0 5.0 2.7 Pathogenesis-related protein
TC76511 PR10-1 protein 5.0 4.4 2.9 Pathogenesis-related protein
TC76518 PR10-1 protein 4.9 5.8 2.5 Pathogenesis-related protein
TC77235 Photoassimilate-responsive
protein
4.4 5.3 2.5 Transcriptionally induced after pathogen
infections
TC86207 Disease resistance response
protein-like
4.4 4.0 2.7 Dirigent-like sequences, may be involved in
lignifications
TC77137 PR10-1 protein 4.4 4.5 3.6 Pathogenesis-related protein
TC76638 PR10-1 protein 4.1 4.2 2.7 Pathogenesis-related protein
TC76640 PR10-1 protein 4.1 3.3 3.4 Pathogenesis-related protein
TC78408 Chitinase 4.0 5.0 1.3 Pathogenesis-related protein
TC76642 PR10-1 protein 4.0 3.9 3.0 Pathogenesis-related protein
TC79513 Stellacyanin-like protein 3.9 3.9 2.1 This type of gene is induced by pathogens and
abiotic stresses and different defense-related
signal molecules
TC92969 TIR-similar-domain-containing
protein
3.9 4.4 R-type resistance protein
TC76643 PR10-1 protein 3.9 4.1 3.0 Pathogenesis-related protein
TC88905 PR10-1 protein 3.8 4.8 2.0 Pathogenesis-related protein
TC81455 Glucan 1,3-beta-glucosidase 3.7 Pathogenesis-related protein
TC78258 Polygalacturonase inhibiting
protein
3.5 Reduces the hydrolytic activity of
polygalacturonases (PGs), limits the growth of
plant pathogens, and also elicits defense
responses in plant
TC77750 Chitinase 3.5 4.0 Pathogenesis-related protein
TC86208 Disease resistance response
protein-like
3.4 4.9 2.1 Dirigent-like sequences, may involve in
lignifications
TC87019 Avr9/Cf-9 rapidly elicited
protein 31
3.3 2.7 Contains Ca2? binding motif
TC86778 Cyanogenic Beta-Glucosidase 3.2 Releases hydrogen cyanide in response to biotic
and abiotic stresses
TC86776 Cyanogenic Beta-Glucosidase 3.2 Releases hydrogen cyanide in response to biotic
and abiotic stresses
TC81700 Lipoxygenase 2.9 Lipidperoxidation of membranes, involvement in
lipid signaling
TC81042 Resistance gene-like 2.9 R-type resistance protein
TC90083 Immediate-early fungal elicitor
protein
2.9 Contains ubiquitin-protein ligase motif (U-box
protein)
TC86652 Syringolide-induced protein
13-1-1
2.9 Contains ubiquitin-protein ligase motif (U-box
protein)
TC85874 1,3-Beta-glucanase 2.8 3.6 Pathogenesis-related protein
TC88172 Avr9/Cf-9 rapidly elicited
protein 132
2.8 1.9 Contains ubiquitin-protein ligase motif (U-box
protein)
TC91970 Chitinase 2.7 Pathogenesis-related protein
TC85264 Lipoxygenase 2.6 3.8 Lipidperoxidation of membranes, involvement in
lipid signaling
TC77754 Syringolide-induced protein
13-1-1
2.6 2.7 Contains ubiquitin-protein ligase motif
(U-box protein)
Plant Mol Biol (2009) 70:627–646 637
123
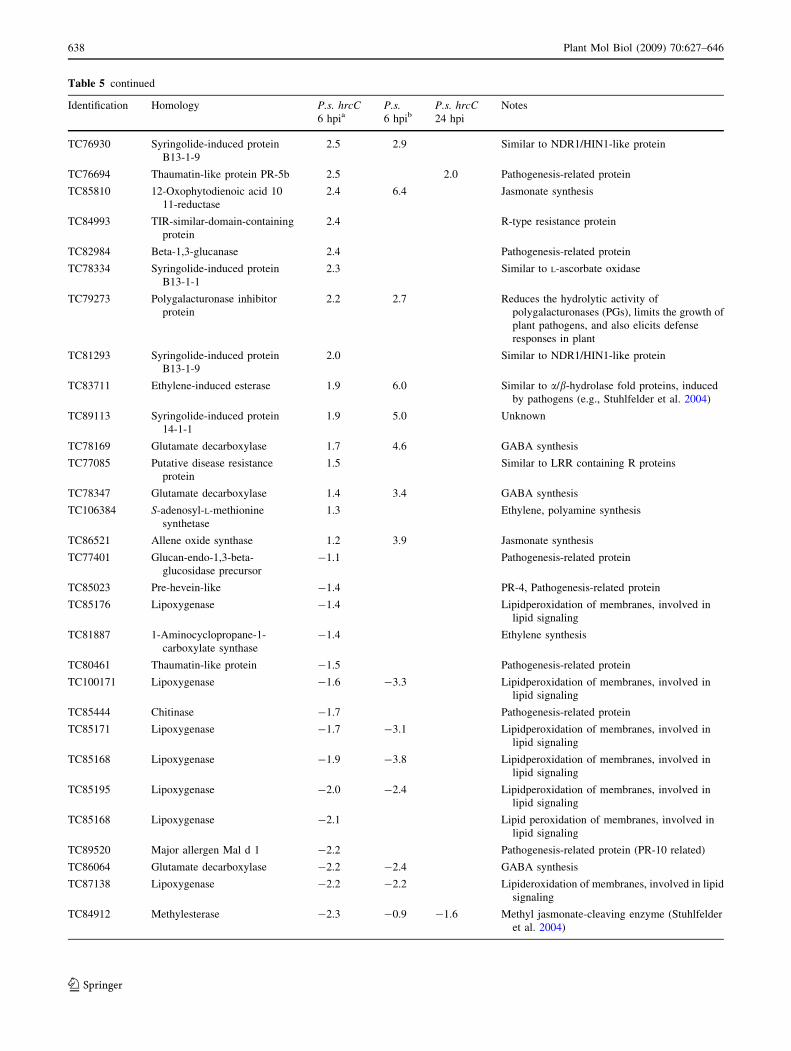
Table 5 continued
Identification Homology P.s. hrcC6 hpia
P.s.6 hpib
P.s. hrcC24 hpi
Notes
TC76930 Syringolide-induced protein
B13-1-9
2.5 2.9 Similar to NDR1/HIN1-like protein
TC76694 Thaumatin-like protein PR-5b 2.5 2.0 Pathogenesis-related protein
TC85810 12-Oxophytodienoic acid 10
11-reductase
2.4 6.4 Jasmonate synthesis
TC84993 TIR-similar-domain-containing
protein
2.4 R-type resistance protein
TC82984 Beta-1,3-glucanase 2.4 Pathogenesis-related protein
TC78334 Syringolide-induced protein
B13-1-1
2.3 Similar to L-ascorbate oxidase
TC79273 Polygalacturonase inhibitor
protein
2.2 2.7 Reduces the hydrolytic activity of
polygalacturonases (PGs), limits the growth of
plant pathogens, and also elicits defense
responses in plant
TC81293 Syringolide-induced protein
B13-1-9
2.0 Similar to NDR1/HIN1-like protein
TC83711 Ethylene-induced esterase 1.9 6.0 Similar to a/b-hydrolase fold proteins, induced
by pathogens (e.g., Stuhlfelder et al. 2004)
TC89113 Syringolide-induced protein
14-1-1
1.9 5.0 Unknown
TC78169 Glutamate decarboxylase 1.7 4.6 GABA synthesis
TC77085 Putative disease resistance
protein
1.5 Similar to LRR containing R proteins
TC78347 Glutamate decarboxylase 1.4 3.4 GABA synthesis
TC106384 S-adenosyl-L-methionine
synthetase
1.3 Ethylene, polyamine synthesis
TC86521 Allene oxide synthase 1.2 3.9 Jasmonate synthesis
TC77401 Glucan-endo-1,3-beta-
glucosidase precursor
-1.1 Pathogenesis-related protein
TC85023 Pre-hevein-like -1.4 PR-4, Pathogenesis-related protein
TC85176 Lipoxygenase -1.4 Lipidperoxidation of membranes, involved in
lipid signaling
TC81887 1-Aminocyclopropane-1-
carboxylate synthase
-1.4 Ethylene synthesis
TC80461 Thaumatin-like protein -1.5 Pathogenesis-related protein
TC100171 Lipoxygenase -1.6 -3.3 Lipidperoxidation of membranes, involved in
lipid signaling
TC85444 Chitinase -1.7 Pathogenesis-related protein
TC85171 Lipoxygenase -1.7 -3.1 Lipidperoxidation of membranes, involved in
lipid signaling
TC85168 Lipoxygenase -1.9 -3.8 Lipidperoxidation of membranes, involved in
lipid signaling
TC85195 Lipoxygenase -2.0 -2.4 Lipidperoxidation of membranes, involved in
lipid signaling
TC85168 Lipoxygenase -2.1 Lipid peroxidation of membranes, involved in
lipid signaling
TC89520 Major allergen Mal d 1 -2.2 Pathogenesis-related protein (PR-10 related)
TC86064 Glutamate decarboxylase -2.2 -2.4 GABA synthesis
TC87138 Lipoxygenase -2.2 -2.2 Lipideroxidation of membranes, involved in lipid
signaling
TC84912 Methylesterase -2.3 -0.9 -1.6 Methyl jasmonate-cleaving enzyme (Stuhlfelder
et al. 2004)
638 Plant Mol Biol (2009) 70:627–646
123
have an enzymatic role in biological redox reactions (Pang
et al. 2004). Constitutively expressed pirin in epithelial
cells showed that pirin up-regulation is associated with
apoptosis (Gelbman et al. 2007).
Plant cysteine proteases are implicated in regulating the
cell death process. Cystatin is a cysteine protease inhibitor
induced during defense responses to biotic and abiotic
stresses (Sanmartin et al. 2005). Pharmacological cysteine
protease (caspase) inhibitors as well as an overexpression
of cystatin can reduce the degree of plant cell death
(Belenghi et al. 2003). In our experiments, cystatin
(TC77459) was found to be activated during P. syringae
mutant-induced BR (approx. 219); comparatively, it was
activated at even higher levels in tissues undergoing HR
(approx. 459).
The transcription of a Medicago ACD2 homolog
(TC78693) was repressed in P. syringae hrcC-infiltrated
tissues at 6 hpi (about 0.3-fold). In Arabidopsis, an ACD2
mutation has an accelerated cell death phenotype. ACD2 is
involved in chlorophyll breakdown, and the cell death
phenotype of this mutant is the consequence of an accu-
mulation of a toxic chlorophyll intermediate (Mach et al.
2001). The ACD2 mutation in plants cause enhanced dis-
ease resistance and salicylic acid accumulation.
The bax inhibitor-like protein, (BI-1), was activated at 6
hpi during HR. BI-1, which is conserved in animals and
plants and involved in the development of and responses to
biotic and abiotic stresses, can suppress cell death induced
by mitochondrial dysfunction, reactive oxygen species, or
elevated cytosolic Ca2? levels (Huckelhoven 2004).
The transcriptional changes of the above presented cell-
death related genes do not completely explain the result of
the reactions (i.e. cell death or symptomless reaction). The
outcome of such interactions is likely to depend on the
balance of these processes. In HR, the balance may be in
favor of PCD, while in BR, the repressing PCD pathway
may prevail. Since the regulation of PCD is very complex,
other factors not detected by our experiments may also
influence the process.
Protein synthesis-related genes
The need for increased protein synthesis during the BR
response at 6 hpi was indicated by the high number of
activated of ribosomal and protein synthesis genes (75
activated and seven repressed; Supplementary Table 5).
This activation did not last until 24 hpi. Only a few of the
same ribosomal genes were up-regulated by HR-inducing
P. syringae, while another set of these genes was repressed
(one activated and 30 repressed genes; Supplementary
Table 5). The reason for this general repression during HR
could be the dismantling of the protein synthesis apparatus
or, alternatively, a specific regulation during cell collapse.
None of the genes that were induced during BR were
repressed in HR, and the repressed genes in wild-type
infiltrated tissue were not activated in BR. Thus, the
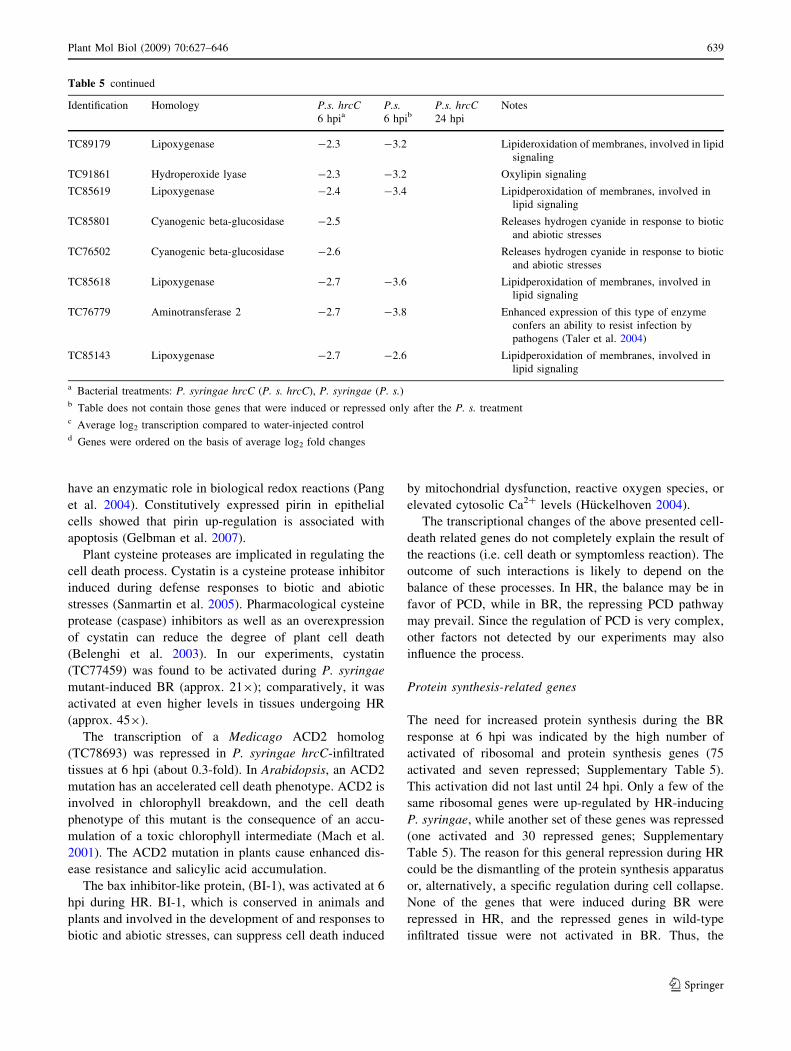
Table 5 continued
Identification Homology P.s. hrcC6 hpia
P.s.6 hpib
P.s. hrcC24 hpi
Notes
TC89179 Lipoxygenase -2.3 -3.2 Lipideroxidation of membranes, involved in lipid
signaling
TC91861 Hydroperoxide lyase -2.3 -3.2 Oxylipin signaling
TC85619 Lipoxygenase -2.4 -3.4 Lipidperoxidation of membranes, involved in
lipid signaling
TC85801 Cyanogenic beta-glucosidase -2.5 Releases hydrogen cyanide in response to biotic
and abiotic stresses
TC76502 Cyanogenic beta-glucosidase -2.6 Releases hydrogen cyanide in response to biotic
and abiotic stresses
TC85618 Lipoxygenase -2.7 -3.6 Lipidperoxidation of membranes, involved in
lipid signaling
TC76779 Aminotransferase 2 -2.7 -3.8 Enhanced expression of this type of enzyme
confers an ability to resist infection by
pathogens (Taler et al. 2004)
TC85143 Lipoxygenase -2.7 -2.6 Lipidperoxidation of membranes, involved in
lipid signaling
a Bacterial treatments: P. syringae hrcC (P. s. hrcC), P. syringae (P. s.)b Table does not contain those genes that were induced or repressed only after the P. s. treatmentc Average log2 transcription compared to water-injected controld Genes were ordered on the basis of average log2 fold changes
Plant Mol Biol (2009) 70:627–646 639
123

repression may not be the simple result of developing HR
and cell destruction, but rather a regulated process that is
different during BR and HR. Our previous observations
showed that temporal inhibition of protein synthesis is able
to block or delay the development of BR and allow non-
pathogenic bacteria to grow or survive (Bozso et al. 1999;
Klement et al. 2003). The above-described results are
corroborating these earlier observations and show that
intense translation is crucial for developing BR.
Comparison of early (6 hpi) versus late (24 hpi)
transcriptional responses in BR
Some of the plant responses were maintained from 6 to 24
hpi, but there were a number of significant alterations in the
transcriptome as well that are summarized here. (1) The
repression of photosynthesis/cloroplast-related genes was
decreased (188 vs. 29 at 6 and 24 hpi, respectively), sug-
gesting that the recovery of plant photosynthetic capacity
and chloroplast-related metabolic processes are important
for effective long-lasting defense (data not shown). (2) No
activated ribosome/protein synthesis-related genes were
found at 24 hpi, implying the decreased requirement of
protein synthesis during this phase of defense. (3) Only few
early transcription factors and genes involved in signal
transduction retained their altered activation, while new
ones were found to be up- or down-regulated at 24 hpi
(data not shown). These factors may regulate the BR
response at a later phase. (4) The defense-related and
phenylpropanoid/flavonoid synthesis genes that were
highly up-regulated at 6 hpi were also activated at 24 hpi,
and only a few newly up- or down-regulated genes from
these classes were found. It seems that these pathways are
continuously activated and may be regulated by those TFs
and/or other signal components that also show permanent
activity during different BR stages. (5) At 24 hpi, cell-wall
fortification genes were still activated (data not shown); in
particular, the repetitive proline-rich protein-encoding
genes and the lignification genes (e.g., genes of dirigent
and laccase-like proteins).
Comparison of resistance responses in leaves
with responses to root symbionts
A comparison of the different types of plant–microbe
interactions may help identify common responses and
specific ones for a particular interaction. In this respect,
M. truncatula is a model for both pathogenic and symbiotic
interactions. We therefore compared our leaf defense
expression data with some published symbiotic transcrip-
tion results. Several papers describe transcriptional changes
during rhizobial and arbuscular mycorrhizal interactions of
M. truncatula. These studies are based either on in silico
analysis of different EST databases or on microarray
experiments in which transcriptional responses to Glomus
fungi inducing arbuscular mycorrhiza and to S. meliloti
bacteria forming nitrogen-fixing nodules are described
(e.g., Fedorova et al. 2002; Liu et al. 2003, 2007; Mergaert
et al. 2003; Brechenmacher et al. 2004; El Yahyaoui et al.
2004; Hohnjec et al. 2005).
Comparison of BR and arbuscular mycorrhizal (AM)
fungi-induced transcriptional changes
Our dataset of the Pseudomonas-infiltrated leaf samples (6
and 24 dpi) was compared with the ones of roots infected
with Glomus mosseae or G. intraradices at 28 days after
infection (Hohnjec et al. 2005). This latter dataset was
produced with the same type of microarrays as the ones
presented here, thus facilitating a comparison between the
sets of data. The comparison of our leaf BR and HR results
with at least twofold Glomus-activated genes in root results
in 36 common genes (Table 6). Most of the Glomus-
activated genes (24 of 36 genes) were repressed at 6 hpi in
either or both BR and HR responses to Pseudomonas.
Examples of these counter-regulated genes were cell wall-
related genes and some TFs. The opposite regulation of
these genes suggests that cell-wall modifications may differ
at this phase of the symbiotic and defense interactions. Of
the 50 most strongly mycorrhizal repressed genes pub-
lished by Hohnjec et al. (2005), 12 showed significant
transcriptional alteration—eight in the same and four in the
opposite direction—at 6 hpi in P. syringae-infiltrated
leaves (Table 6). The four counter-regulated genes can be
part of plant defense reactions suppressed by the fungus to
avoid the plant resistance (e.g., the thaumatin-like genes
that may have antimicrobial activity).
It is assumed that defense- and other stress-related
genes are up-regulated at the initial phase of the AM
fungi–plant interaction (Brechenmacher et al. 2004;
Deguchi et al. 2007). Therefore, our leaf expression data
were also compared with early (6 hpi and 8 dpi) AM
fungi-induced transcriptional changes published by Si-
ciliano et al. (2007) and Liu et al. (2003). These com-
parisons resulted in a limited number of commonly up-
regulated genes—seven of 104 and four of 14, respec-
tively (data not shown). The restricted similarity between
the transcriptional alteration of a bacterially induced leaf
resistance response and a fungal symbiotic interaction
implies that—at least in these phases of interactions—
basically other sets of genes are required. However, more
detailed time course analyses of both interactions could
reveal additional common genes, especially at early
phases of infections.
640 Plant Mol Biol (2009) 70:627–646
123
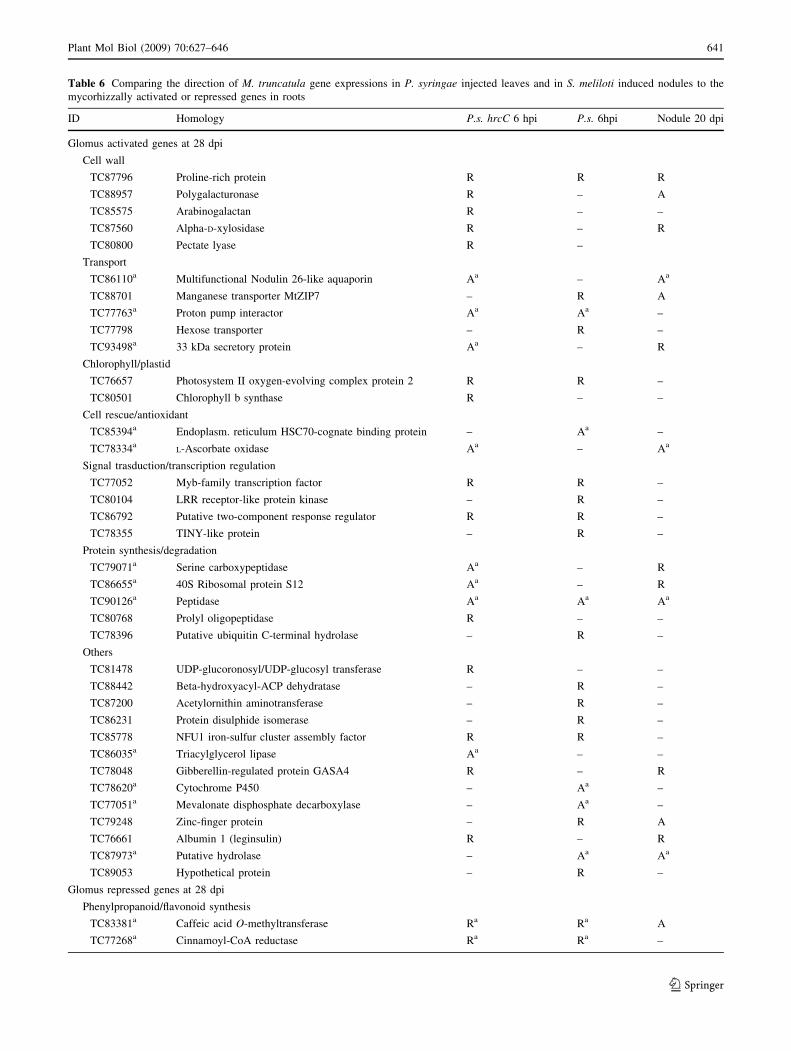
Table 6 Comparing the direction of M. truncatula gene expressions in P. syringae injected leaves and in S. meliloti induced nodules to the
mycorhizzally activated or repressed genes in roots
ID Homology P.s. hrcC 6 hpi P.s. 6hpi Nodule 20 dpi
Glomus activated genes at 28 dpi
Cell wall
TC87796 Proline-rich protein R R R
TC88957 Polygalacturonase R – A
TC85575 Arabinogalactan R – –
TC87560 Alpha-D-xylosidase R – R
TC80800 Pectate lyase R –
Transport
TC86110a Multifunctional Nodulin 26-like aquaporin Aa – Aa
TC88701 Manganese transporter MtZIP7 – R A
TC77763a Proton pump interactor Aa Aa –
TC77798 Hexose transporter – R –
TC93498a 33 kDa secretory protein Aa – R
Chlorophyll/plastid
TC76657 Photosystem II oxygen-evolving complex protein 2 R R –
TC80501 Chlorophyll b synthase R – –
Cell rescue/antioxidant
TC85394a Endoplasm. reticulum HSC70-cognate binding protein – Aa –
TC78334aL-Ascorbate oxidase Aa – Aa
Signal trasduction/transcription regulation
TC77052 Myb-family transcription factor R R –
TC80104 LRR receptor-like protein kinase – R –
TC86792 Putative two-component response regulator R R –
TC78355 TINY-like protein – R –
Protein synthesis/degradation
TC79071a Serine carboxypeptidase Aa – R
TC86655a 40S Ribosomal protein S12 Aa – R
TC90126a Peptidase Aa Aa Aa
TC80768 Prolyl oligopeptidase R – –
TC78396 Putative ubiquitin C-terminal hydrolase – R –
Others
TC81478 UDP-glucoronosyl/UDP-glucosyl transferase R – –
TC88442 Beta-hydroxyacyl-ACP dehydratase – R –
TC87200 Acetylornithin aminotransferase – R –
TC86231 Protein disulphide isomerase – R –
TC85778 NFU1 iron-sulfur cluster assembly factor R R –
TC86035a Triacylglycerol lipase Aa – –
TC78048 Gibberellin-regulated protein GASA4 R – R
TC78620a Cytochrome P450 – Aa –
TC77051a Mevalonate disphosphate decarboxylase – Aa –
TC79248 Zinc-finger protein – R A
TC76661 Albumin 1 (leginsulin) R – R
TC87973a Putative hydrolase – Aa Aa
TC89053 Hypothetical protein – R –
Glomus repressed genes at 28 dpi
Phenylpropanoid/flavonoid synthesis
TC83381a Caffeic acid O-methyltransferase Ra Ra A
TC77268a Cinnamoyl-CoA reductase Ra Ra –
Plant Mol Biol (2009) 70:627–646 641
123
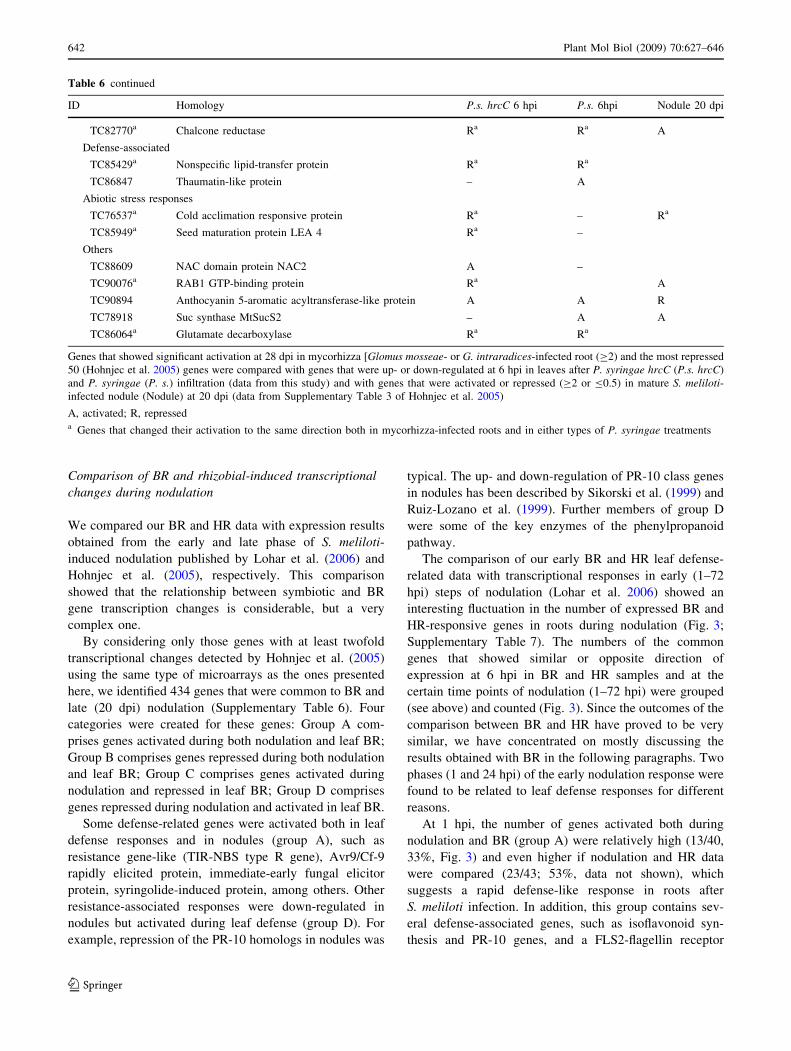
Comparison of BR and rhizobial-induced transcriptional
changes during nodulation
We compared our BR and HR data with expression results
obtained from the early and late phase of S. meliloti-
induced nodulation published by Lohar et al. (2006) and
Hohnjec et al. (2005), respectively. This comparison
showed that the relationship between symbiotic and BR
gene transcription changes is considerable, but a very
complex one.
By considering only those genes with at least twofold
transcriptional changes detected by Hohnjec et al. (2005)
using the same type of microarrays as the ones presented
here, we identified 434 genes that were common to BR and
late (20 dpi) nodulation (Supplementary Table 6). Four
categories were created for these genes: Group A com-
prises genes activated during both nodulation and leaf BR;
Group B comprises genes repressed during both nodulation
and leaf BR; Group C comprises genes activated during
nodulation and repressed in leaf BR; Group D comprises
genes repressed during nodulation and activated in leaf BR.
Some defense-related genes were activated both in leaf
defense responses and in nodules (group A), such as
resistance gene-like (TIR-NBS type R gene), Avr9/Cf-9
rapidly elicited protein, immediate-early fungal elicitor
protein, syringolide-induced protein, among others. Other
resistance-associated responses were down-regulated in
nodules but activated during leaf defense (group D). For
example, repression of the PR-10 homologs in nodules was
typical. The up- and down-regulation of PR-10 class genes
in nodules has been described by Sikorski et al. (1999) and
Ruiz-Lozano et al. (1999). Further members of group D
were some of the key enzymes of the phenylpropanoid
pathway.
The comparison of our early BR and HR leaf defense-
related data with transcriptional responses in early (1–72
hpi) steps of nodulation (Lohar et al. 2006) showed an
interesting fluctuation in the number of expressed BR and
HR-responsive genes in roots during nodulation (Fig. 3;
Supplementary Table 7). The numbers of the common
genes that showed similar or opposite direction of
expression at 6 hpi in BR and HR samples and at the
certain time points of nodulation (1–72 hpi) were grouped
(see above) and counted (Fig. 3). Since the outcomes of the
comparison between BR and HR have proved to be very
similar, we have concentrated on mostly discussing the
results obtained with BR in the following paragraphs. Two
phases (1 and 24 hpi) of the early nodulation response were
found to be related to leaf defense responses for different
reasons.
At 1 hpi, the number of genes activated both during
nodulation and BR (group A) were relatively high (13/40,
33%, Fig. 3) and even higher if nodulation and HR data
were compared (23/43; 53%, data not shown), which
suggests a rapid defense-like response in roots after
S. meliloti infection. In addition, this group contains sev-
eral defense-associated genes, such as isoflavonoid syn-
thesis and PR-10 genes, and a FLS2-flagellin receptor
Table 6 continued
ID Homology P.s. hrcC 6 hpi P.s. 6hpi Nodule 20 dpi
TC82770a Chalcone reductase Ra Ra A
Defense-associated
TC85429a Nonspecific lipid-transfer protein Ra Ra
TC86847 Thaumatin-like protein – A
Abiotic stress responses
TC76537a Cold acclimation responsive protein Ra – Ra
TC85949a Seed maturation protein LEA 4 Ra –
Others
TC88609 NAC domain protein NAC2 A –
TC90076a RAB1 GTP-binding protein Ra A
TC90894 Anthocyanin 5-aromatic acyltransferase-like protein A A R
TC78918 Suc synthase MtSucS2 – A A
TC86064a Glutamate decarboxylase Ra Ra
Genes that showed significant activation at 28 dpi in mycorhizza [Glomus mosseae- or G. intraradices-infected root (C2) and the most repressed
50 (Hohnjec et al. 2005) genes were compared with genes that were up- or down-regulated at 6 hpi in leaves after P. syringae hrcC (P.s. hrcC)
and P. syringae (P. s.) infiltration (data from this study) and with genes that were activated or repressed (C2 or B0.5) in mature S. meliloti-infected nodule (Nodule) at 20 dpi (data from Supplementary Table 3 of Hohnjec et al. 2005)
A, activated; R, represseda Genes that changed their activation to the same direction both in mycorhizza-infected roots and in either types of P. syringae treatments
642 Plant Mol Biol (2009) 70:627–646
123
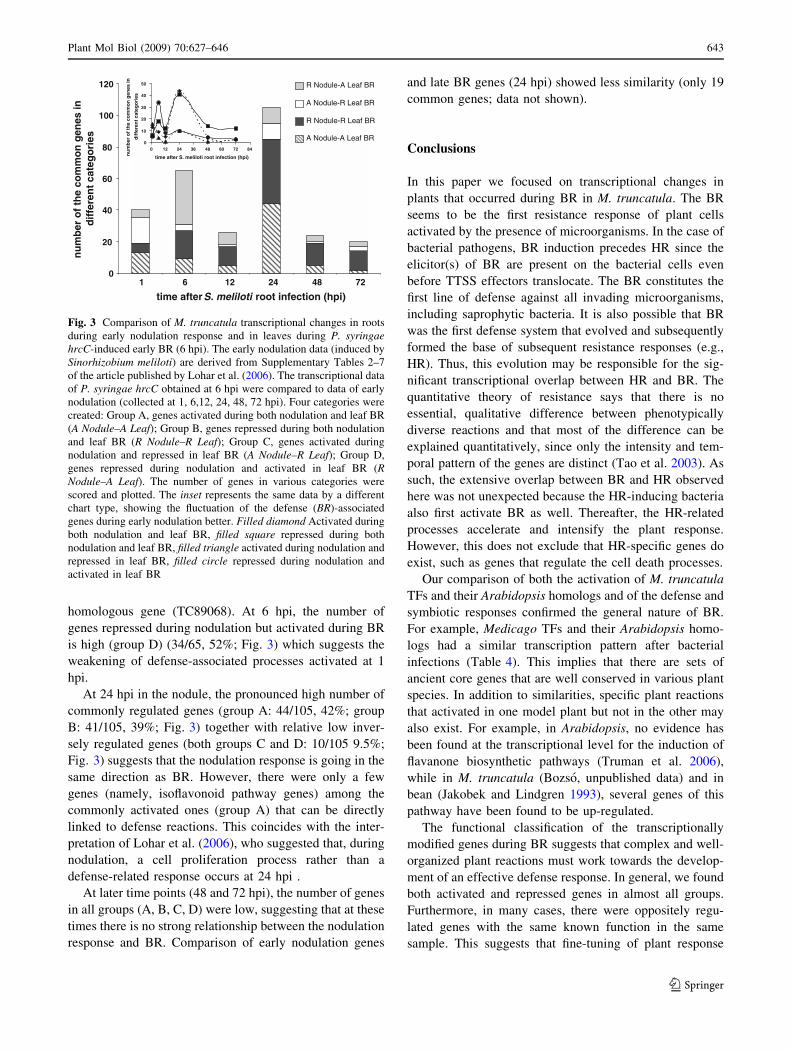
homologous gene (TC89068). At 6 hpi, the number of
genes repressed during nodulation but activated during BR
is high (group D) (34/65, 52%; Fig. 3) which suggests the
weakening of defense-associated processes activated at 1
hpi.
At 24 hpi in the nodule, the pronounced high number of
commonly regulated genes (group A: 44/105, 42%; group
B: 41/105, 39%; Fig. 3) together with relative low inver-
sely regulated genes (both groups C and D: 10/105 9.5%;
Fig. 3) suggests that the nodulation response is going in the
same direction as BR. However, there were only a few
genes (namely, isoflavonoid pathway genes) among the
commonly activated ones (group A) that can be directly
linked to defense reactions. This coincides with the inter-
pretation of Lohar et al. (2006), who suggested that, during
nodulation, a cell proliferation process rather than a
defense-related response occurs at 24 hpi .
At later time points (48 and 72 hpi), the number of genes
in all groups (A, B, C, D) were low, suggesting that at these
times there is no strong relationship between the nodulation
response and BR. Comparison of early nodulation genes
and late BR genes (24 hpi) showed less similarity (only 19
common genes; data not shown).
Conclusions
In this paper we focused on transcriptional changes in
plants that occurred during BR in M. truncatula. The BR
seems to be the first resistance response of plant cells
activated by the presence of microorganisms. In the case of
bacterial pathogens, BR induction precedes HR since the
elicitor(s) of BR are present on the bacterial cells even
before TTSS effectors translocate. The BR constitutes the
first line of defense against all invading microorganisms,
including saprophytic bacteria. It is also possible that BR
was the first defense system that evolved and subsequently
formed the base of subsequent resistance responses (e.g.,
HR). Thus, this evolution may be responsible for the sig-
nificant transcriptional overlap between HR and BR. The
quantitative theory of resistance says that there is no
essential, qualitative difference between phenotypically
diverse reactions and that most of the difference can be
explained quantitatively, since only the intensity and tem-
poral pattern of the genes are distinct (Tao et al. 2003). As
such, the extensive overlap between BR and HR observed
here was not unexpected because the HR-inducing bacteria
also first activate BR as well. Thereafter, the HR-related
processes accelerate and intensify the plant response.
However, this does not exclude that HR-specific genes do
exist, such as genes that regulate the cell death processes.
Our comparison of both the activation of M. truncatula
TFs and their Arabidopsis homologs and of the defense and
symbiotic responses confirmed the general nature of BR.
For example, Medicago TFs and their Arabidopsis homo-
logs had a similar transcription pattern after bacterial
infections (Table 4). This implies that there are sets of
ancient core genes that are well conserved in various plant
species. In addition to similarities, specific plant reactions
that activated in one model plant but not in the other may
also exist. For example, in Arabidopsis, no evidence has
been found at the transcriptional level for the induction of
flavanone biosynthetic pathways (Truman et al. 2006),
while in M. truncatula (Bozso, unpublished data) and in
bean (Jakobek and Lindgren 1993), several genes of this
pathway have been found to be up-regulated.
The functional classification of the transcriptionally
modified genes during BR suggests that complex and well-
organized plant reactions must work towards the develop-
ment of an effective defense response. In general, we found
both activated and repressed genes in almost all groups.
Furthermore, in many cases, there were oppositely regu-
lated genes with the same known function in the same
sample. This suggests that fine-tuning of plant response
0
20
40
60
80
100
120
1 6 12 24 48 72
time after S. meliloti root infection (hpi)
R Nodule-A Leaf BR
A Nodule-R Leaf BR
R Nodule-R Leaf BR
A Nodule-A Leaf BR0
10
20
30
40
50
0 12 24 36 48 60 72 84
time after S. meliloti root infection (hpi)nu
mb
er o
f th
e co
mm
on
gen
es in
dif
fere
nt
cate
go
ries
nu
mb
er o
f th
e co
mm
on
gen
es in
d
iffe
ren
t ca
teg
ori
es
Fig. 3 Comparison of M. truncatula transcriptional changes in roots
during early nodulation response and in leaves during P. syringaehrcC-induced early BR (6 hpi). The early nodulation data (induced by
Sinorhizobium meliloti) are derived from Supplementary Tables 2–7
of the article published by Lohar et al. (2006). The transcriptional data
of P. syringae hrcC obtained at 6 hpi were compared to data of early
nodulation (collected at 1, 6,12, 24, 48, 72 hpi). Four categories were
created: Group A, genes activated during both nodulation and leaf BR
(A Nodule–A Leaf); Group B, genes repressed during both nodulation
and leaf BR (R Nodule–R Leaf); Group C, genes activated during
nodulation and repressed in leaf BR (A Nodule–R Leaf); Group D,
genes repressed during nodulation and activated in leaf BR (RNodule–A Leaf). The number of genes in various categories were
scored and plotted. The inset represents the same data by a different
chart type, showing the fluctuation of the defense (BR)-associated
genes during early nodulation better. Filled diamond Activated during
both nodulation and leaf BR, filled square repressed during both
nodulation and leaf BR, filled triangle activated during nodulation and
repressed in leaf BR, filled circle repressed during nodulation and
activated in leaf BR
Plant Mol Biol (2009) 70:627–646 643
123

involving activation or repression of the proper isoenzymes
should be important for the efficient establishment of
defense responses.
Our comparison of the transcriptome changes induced
by BR or HR and mycorrhizal/rhizobial gene alterations
showed that there is a significant overlap of genes involved
in defense and symbiotic plant responses. High similarities
were found with the early phase of S. meliloti infection,
which confirms that this period of symbiotic interaction is
similar to defense reactions and that the suppression of
these responses must be essential for successful coloniza-
tion. However, transcriptional alteration of a gene during
both the resistance response and symbiotic interaction does
not automatically indicate a direct defense function during
symbiosis. It may also imply other more general functions
of the gene that are activated after various stress responses.
The greatest challenge facing researchers in the field is
to isolate and identify the key components for the BR
response in the vast number of transcriptionally modified
genes. There are several approaches that can be used for
this purpose. A candidate gene approach or the identifica-
tion of the genes affected (either induced or repressed) by
bacterial BR suppressors is a promising method by which
to find important BR-related genes. In addition, the tran-
scriptome analysis of legume plants (including Medicago)
during different defense reactions against various patho-
gens and elicitors and a comparison of these data with the
results of other types of plants and with symbiotic
responses will facilitate in the determination of the function
of the genes.
Acknowledgments This research was supported by grants of the
Hungarian National Science Foundation, AT-049318 and K68386.
We thank the GODMAP platform (Gif-sur-Yvette, France) for the use
of their microarray facilities.
References
Alfano JR, Collmer A (2004) Type III secretion system effector
proteins: double agents in bacterial disease and plant defense.
Annu Rev Phytopathol 42:385–414. doi:10.1146/annurev.phyto.
42.040103.110731
Bantignies B, Seguin J, Muzac I, Dedaldechamp F, Gulick P, Ibrahim
R (2000) Direct evidence for ribonucleolytic activity of a PR-10-
like protein from white lupin roots. Plant Mol Biol 42:871–881.
doi:10.1023/A:1006475303115
Bardwell L (2005) A walk-through of the yeast mating pheromone
response pathway. Peptides 26:339–350. doi:10.1016/j.peptides.
2004.10.002
Belenghi B, Acconcia F, Trovato M, Perazzolli M, Bocedi A, Polticelli F,
Ascenzi P, Delledonne M (2003) AtCYS1, a cystatin from
Arabidopsis thaliana, suppresses hypersensitive cell death. Eur J
Biochem 270:2593–2604. doi:10.1046/j.1432-1033.2003.03630.x
Bestwick CS, Bennett MH, Mansfield JW (1995) Hrp mutant of
Pseudomonas syringae pv. phaseolicola induces cell wall
alterations but not membrane damage leading to the HR in
lettuce (Lactuca sativa). Plant Physiol 108:503–516
Bozso Z, Ott PG, Kecskes ML, Klement Z (1999) Effect of heat and
cycloheximide treatment of tobacco on the ability of Pseudo-monas syringae pv. syringae 61 hrp/hrmA mutants to cause HR.
Physiol Mol Plant Pathol 55:215–223. doi:10.1006/pmpp.
1999.0225
Bozso Z, Ott PG, Szatmari A, Czelleng A, Varga G, Besenyei E,
Sardi E, Banyai E, Klement Z (2005) Early detection of
bacterium-induced basal resistance in tobacco leaves with
diaminobenzidine and dichlorofluorescein diacetate. J Phytopa-
thol 153:596–607. doi:10.1111/j.1439-0434.2005.01026.x
Brazma A, Hingamp P, Quackenbush J, Sherlock G, Spellman P,
Stoeckert C, Aach J, Ansorge W, Ball CA, Causton HC,
Gaasterland T, Glenisson P, Holstege FC, Kim IF, Markowitz V,
Matese JC, Parkinson H, Robinson A, Sarkans U, Schulze-
Kremer S, Stewart J, Taylor R, Vilo J, Vingron M (2001)
Minimum information about a microarray experiment (MI-
AME)-toward standards for microarray data. Nat Genet 29:365–
371. doi:10.1038/ng1201-365
Brechenmacher L, Weidmann S, van Tuinen D, Chatagnier O,
Gianinazzi S, Franken P, Gianinazzi-Pearson V (2004) Expres-
sion profiling of up-regulated plant and fungal genes in early and
late stages of Medicago truncatula-Glomus mosseae interactions.
Mycorrhiza 14:253–262. doi:10.1007/s00572-003-0263-4
Breda C, Sallaud C, el-Turk J, Buffard D, de Kozak I, Esnault R,
Kondorosi A (1996) Defense reaction in Medicago sativa: a gene
encoding a class 10 PR protein is expressed in vascular bundles.
Mol Plant Microbe Interact 9:713–719
Breitling R, Armengaud P, Amtmann A, Herzyk P (2004) Rank
products: a simple, yet powerful, new method to detect
differentially regulated genes in replicated microarray experi-
ments. FEBS Lett 573:83–92. doi:10.1016/j.febslet.2004.07.055
Carimi F, Zottini M, Formentin E, Terzi M, Lo Schiavo F (2003)
Cytokinins: new apoptotic inducers in plants. Planta 216:413–
421
Chen C, Wanduragala S, Becker DF, Dickman MB (2006) Tomato
QM-like protein protects Saccharomyces cerevisiae cells against
oxidative stress by regulating intracellular proline levels. Appl
Environ Microbiol 72:4001–4006. doi:10.1128/AEM.02428-05
de Torres M, Sanchez P, Fernandez-Delmond I, Grant M (2003)
Expression profiling of the host response to bacterial infection:
the transition from basal to induced defence responses in RPM1-
mediated resistance. Plant J 33:665–676. doi:10.1046/j.1365-
313X.2003.01653.x
Deguchi Y, Banba M, Shimoda Y, Chechetka SA, Suzuri R, Okusako
Y, Ooki Y, Toyokura K, Suzuki A, Uchiumi T, Higashi S, Abe
M, Kouchi H, Izui K, Hata S (2007) Transcriptome profiling of
Lotus japonicus roots during arbuscular mycorrhiza development
and comparison with that of nodulation. DNA Res 14:117–133.
doi:10.1093/dnares/dsm014
Dietz KJ (2003) Plant peroxiredoxins. Annu Rev Plant Biol 54:93–
107. doi:10.1146/annurev.arplant.54.031902.134934
Dow M, Newman MA, von Roepenack E (2000) The induction and
modulation of plant defense responses by bacterial lipopolysac-
charides. Annu Rev Phytopathol 38:241–261. doi:10.1146/annurev.phyto.38.1.241
El Yahyaoui F, Kuster H, Ben Amor B, Hohnjec N, Puhler A, Becker
A, Gouzy J, Vernie T, Gough C, Niebel A, Godiard L, Gamas P
(2004) Expression profiling in Medicago truncatula identifies
more than 750 genes differentially expressed during nodulation,
including many potential regulators of the symbiotic program.
Plant Physiol 136:3159–3176. doi:10.1104/pp.104.043612
Eulgem T (2005) Regulation of the Arabidopsis defense transcrip-
tome. Trends Plant Sci 10:71–78. doi:10.1016/j.tplants.2004.
12.006
644 Plant Mol Biol (2009) 70:627–646
123

Fedorova M, van de Mortel J, Matsumoto PA, Cho J, Town CD,
VandenBosch KA, Gantt JS, Vance CP (2002) Genome-wide
identification of nodule-specific transcripts in the model legume
Medicago truncatula. Plant Physiol 130:519–537. doi:10.1104/
pp.006833
Felix G, Boller T (2003) Molecular sensing of bacteria in plants. The
highly conserved RNA-binding motif RNP-1 of bacterial cold
shock proteins is recognized as an elicitor signal in tobacco. J
Biol Chem 278:6201–6208. doi:10.1074/jbc.M209880200
Garcia-Olmedo F, Molina A, Segura A, Moreno M (1995) The
defensive role of nonspecific lipid-transfer proteins in plants.
Trends Microbiol 3:72–74. doi:10.1016/S0966-842X(00)88879-4
Gelbman BD, Heguy A, O’Connor TP, Zabner J, Crystal RG (2007)
Upregulation of pirin expression by chronic cigarette smoking is
associated with bronchial epithelial cell apoptosis. Respir Res
8:10. doi:10.1186/1465-9921-8-10
Gomez-Gomez L, Boller T (2002) Flagellin perception: a paradigm
for innate immunity. Trends Plant Sci 7:251–256. doi:10.1016/
S1360-1385(02)02261-6
Greenberg JT, Yao N (2004) The role and regulation of programmed
cell death in plant-pathogen interactions. Cell Microbiol 6:201–
211. doi:10.1111/j.1462-5822.2004.00361.x
Guo A, Durner J, Klessig DF (1998) Characterization of a tobacco
epoxide hydrolase gene induced during the resistance response to
TMV. Plant J 15:647–656. doi:10.1046/j.1365-313x.1998.00241.x
Hammond-Kosack KE, Jones JDG (1996) Resistance gene-dependent
plant defense responses. Plant Cell 8:1773–1791
He X, Anderson JC, del Pozo O, Gu YQ, Tang X, Martin GB (2004)
Silencing of subfamily I of protein phosphatase 2A catalytic
subunits results in activation of plant defense responses and
localized cell death. Plant J 38:563–577. doi:10.1111/j.1365-
313X.2004.02073.x
Herbers K, Monke G, Badur R, Sonnewald U (1995) A simplified
procedure for the subtractive cDNA cloning of photoassimilate-
responding genes: isolation of cDNAs encoding a new class of
pathogenesis-related proteins. Plant Mol Biol 29:1027–1038.
doi:10.1007/BF00014975
Hirayama T, Ishida C, Kuromori T, Obata S, Shimoda C, Yamamoto
M, Shinozaki K, Ohto C (1997) Functional cloning of a cDNA
encoding Mei2-like protein from Arabidopsis thaliana using a
fission yeast pheromone receptor deficient mutant. FEBS Lett
413:16–20. doi:10.1016/S0014-5793(97)00871-5
Hohnjec N, Vieweg MF, Puhler A, Becker A, Kuster H (2005)
Overlaps in the transcriptional profiles of Medicago truncatularoots inoculated with two different Glomus fungi provide
insights into the genetic program activated during arbuscular
mycorrhiza. Plant Physiol 137:1283–1301. doi:10.1104/
pp.104.056572
Hong JK, Hwang BK (2006) Promoter activation of pepper class II
basic chitinase gene, CAChi2, and enhanced bacterial disease
resistance and osmotic stress tolerance in the CAChi2-over-
expressing Arabidopsis. Planta 223:433–448. doi:10.1007/
s00425-005-0099-6
Huckelhoven R (2004) BAX Inhibitor-1, an ancient cell death suppressor
in animals and plants with prokaryotic relatives. Apoptosis 9:299–
307. doi:10.1023/B:APPT.0000025806.71000.1c
Jakobek JL, Lindgren PB (1993) Generalized induction of defense
responses in bean is not correlated with the induction of the
hypersensitive reaction. Plant Cell 5:49–56
Jones AM, Thomas V, Truman B, Lilley K, Mansfield JW, Grant M
(2004) Specific changes in the Arabidopsis proteome in response to
bacterial challenge: differentiating basal and R-gene mediated
resistance. Phytochemistry 65:1805–1816. doi:10.1016/j.phyto
chem.2004.04.005
King EO, Ward MK, Raney DE (1954) Two simple media for the
demonstration of pyocyanine and fluorescein. J Lab Clin Med
22:301–307
Klement Z (1963) Rapid detection of the pathogenicity of phytopha-
togenic pseudomonads. Nature 199:299–300. doi:10.1038/
199299b0
Klement Z, Bozso Z, Ott PG, Kecskes ML, Rudolp K (1999)
Symptomless resistant response instead of the hypersensitive
reaction in tobacco leaves after infiltration of heterologous
pathovars of Pseudomonas syringae. J Phytopathol 147:467–
475. doi:10.1111/j.1439-0434.1999.tb03852.x
Klement Z, Bozso Z, Kecskes ML, Besenyei E, Czelleng A, Ott PG
(2003) Local early induced resistance of plants as the first line of
defence against bacteria. Pest Manag Sci 59:465–474.
doi:10.1002/ps.694
Koistinen KM, Soininen P, Venalainen TA, Hayrinen J, Laatikainen R,
Perakyla M, Tervahauta AI, Karenlampi SO (2005) Birch PR-10c
interacts with several biologically important ligands. Phytochem-
istry 66:2524–2533. doi:10.1016/j.phytochem.2005.09.007
Kunze G, Zipfel C, Robatzek S, Niehaus K, Boller T, Felix G (2004)
The N terminus of bacterial elongation factor Tu elicits innate
immunity in Arabidopsis plants. Plant Cell 16:3496–3507.
doi:10.1105/tpc.104.026765
Lapik YR, Kaufman LS (2003) The Arabidopsis cupin domain
protein AtPirin1 interacts with the G protein alpha-subunit GPA1
and regulates seed germination and early seedling development.
Plant Cell 15:1578–1590. doi:10.1105/tpc.011890
Liu JJ, Ekramoddoullah AKM (2006) The family 10 of plant
pathogenesis-related proteins: Their structure, regulation, and
function in response to biotic and abiotic stresses. Physiol Mol
Plant Pathol 68:3–13. doi:10.1016/j.pmpp.2006.06.004
Liu JY, Blaylock LA, Endre G, Cho J, Town CD, VandenBosch KA,
Harrison MJ (2003) Transcript profiling coupled with spatial
expression analyses reveals genes involved in distinct develop-
mental stages of an arbuscular mycorrhizal symbiosis. Plant Cell
15:2106–2123. doi:10.1105/tpc.014183
Liu J, Maldonado-Mendoza I, Lopez-Meyer M, Cheung F, Town CD,
Harrison MJ (2007) Arbuscular mycorrhizal symbiosis is
accompanied by local and systemic alterations in gene expres-
sion and an increase in disease resistance in the shoots. Plant J
50:529–544. doi:10.1111/j.1365-313X.2007.03069.x
Lohar DP, Sharopova N, Endre G, Penuela S, Samac D, Town C,
Silverstein KA, VandenBosch KA (2006) Transcript analysis of
early nodulation events in Medicago truncatula. Plant Physiol
140:221–234. doi:10.1104/pp.105.070326
Mach JM, Castillo AR, Hoogstraten R, Greenberg JT (2001) The
Arabidopsis-accelerated cell death gene ACD2 encodes red
chlorophyll catabolite reductase and suppresses the spread of
disease symptoms. Proc Natl Acad Sci USA 98:771–776.
doi:10.1073/pnas.021465298
Mackey D, McFall AJ (2006) MAMPs and MIMPs: proposed
classifications for inducers of innate immunity. Mol Microbiol
61:1365–1371. doi:10.1111/j.1365-2958.2006.05311.x
Markovic-Housley Z, Degano M, Lamba D, von Roepenack-Lahaye
E, Clemens S, Susani M, Ferreira F, Scheiner O, Breiteneder H
(2003) Crystal structure of a hypoallergenic isoform of the major
birch pollen allergen Bet v 1 and its likely biological function as
a plant steroid carrier. J Mol Biol 325:123–133. doi:10.1016/
S0022-2836(02)01197-X
Mergaert P, Nikovics K, Kelemen Z, Maunoury N, Vaubert D,
Kondorosi A, Kondorosi E (2003) A novel family in Medicagotruncatula consisting of more than 300 nodule-specific genes
coding for small, secreted polypeptides with conserved cysteine
motifs. Plant Physiol 132:161–173. doi:10.1104/pp.102.018192
Plant Mol Biol (2009) 70:627–646 645
123

Murray GI, Paterson PJ, Weaver RJ, Ewen SW, Melvin WT, Burke
MD (1993) The expression of cytochrome P-450, epoxide
hydrolase, and glutathione S-transferase in hepatocellular carci-
noma. Cancer 71:36–43. doi:10.1002/1097-0142(19930101)71:
1\36::AID-CNCR2820710107[3.0.CO;2-J
Mysore KS, Crasta OR, Tuori RP, Folkerts O, Swirsky PB, Martin
GB (2002) Comprehensive transcript profiling of Pto- and Prf-
mediated host defense responses to infection by Pseudomonassyringae pv. tomato. Plant J 32:299–315. doi:10.1046/j.1365-
313X.2002.01424.x
Nakashita H, Yasuda M, Nitta T, Asami T, Fujioka S, Arai Y,
Sekimata K, Takatsuto S, Yamaguchi I, Yoshida S (2003)
Brassinosteroid functions in a broad range of disease resistance
in tobacco and rice. Plant J 33:887–898. doi:10.1046/j.1365-
313X.2003.01675.x
Navarro L, Zipfel C, Rowland O, Keller I, Robatzek S, Boller T,
Jones JD (2004) The transcriptional innate immune response to
flg22. Interplay and overlap with AVR gene-dependent defense
responses and bacterial pathogenesis. Plant Physiol 135:1113–
1128. doi:10.1104/pp.103.036749
Newman JW, Morisseau C, Hammock BD (2005) Epoxide hydro-
lases: their roles and interactions with lipid metabolism. Prog
Lipid Res 44:1–51. doi:10.1016/j.plipres.2004.10.001
Novacky A (1972) Suppression of the bacterially induced hypersen-
sitive reaction by cytokinins. Physiol Plant Pathol 2:101–104.
doi:10.1016/S0048-4059(72)80004-3
Oh HS, Kwon H, Sun SK, Yang CH (2002) QM, a putative tumor
suppressor, regulates proto-oncogene c-yes. J Biol Chem
277:36489–36498. doi:10.1074/jbc.M201859200
Orzaez D, de Jong AJ, Woltering EJ (2001) A tomato homologue of
the human protein PIRIN is induced during programmed cell
death. Plant Mol Biol 46:459–468. doi:10.1023/A:10106185
15051
Ott PG, Szabo L, Balazs E, Klement Z (1997) Submicroscopic
evidence of bacterially induced resistance in tobacco leaves.
Acta Phytopathol Entomol Hung 32:265–280
Ott PG, Varga GJ, Szatmari A, Bozso Z, Klement E, Medzihradszky
KF, Besenyei E, Czelleng A, Klement Z (2006) Novel extracel-
lular chitinases rapidly and specifically induced by general
bacterial elicitors and suppressed by virulent bacteria as a marker
of early basal resistance in tobacco. Mol Plant Microbe Interact
19:161–172. doi:10.1094/MPMI-19-0161
Pang H, Bartlam M, Zeng Q, Miyatake H, Hisano T, Miki K, Wong
LL, Gao GF, Rao Z (2004) Crystal structure of human pirin: an
iron-binding nuclear protein and transcription cofactor. J Biol
Chem 279:1491–1498. doi:10.1074/jbc.M310022200
Pastuglia M, Swarup R, Rocher A, Saindrenan P, Roby D, Dumas C,
Cock JM (2002) Comparison of the expression patterns of two
small gene families of S genefamily receptor kinase genes during
the defence response in Brassica oleracea and Arabidopsisthaliana. Gene 282:215–225. doi:10.1016/S0378-1119(01)00
821-6
Pinot F, Bosch H, Alayrac C, Mioskowski C, Vendais A, Durst F,
Salaun JP (1993) [omega]-hydroxylation of oleic acid in Viciasativa microsomes (inhibition by substrate analogs and inacti-
vation by terminal acetylenes). Plant Physiol 102:1313–1318
Ruiz-Lozano JM, Roussel H, Gianinazzi S, Gianinazzi-Pearson V
(1999) Defense genes are differentially induced by a mycorrhizal
fungus and Rhizobium sp. in wild-type and symbiosis-defective
pea genotypes. Mol Plant Microbe Interact 12:976–984.
doi:10.1094/MPMI.1999.12.11.976
Saeed AI, Sharov V, White J, Li J, Liang W, Bhagabati N, Braisted J,
Klapa M, Currier T, Thiagarajan M, Sturn A, Snuffin M,
Rezantsev A, Popov D, Ryltsov A, Kostukovich E, Borisovsky I,
Liu Z, Vinsavich A, Trush V, Quackenbush J (2003) TM4: a
free, open-source system for microarray data management and
analysis. Biotechniques 34:374–378
Sanmartin M, Jaroszewski L, Raikhel NV, Rojo E (2005) Caspases.
Regulating death since the origin of life. Plant Physiol 137:841–
847. doi:10.1104/pp.104.058552
Schweighofer A, Hirt H, Meskiene I (2004) Plant PP2C phosphatases:
emerging functions in stress signaling. Trends Plant Sci 9:236–
243. doi:10.1016/j.tplants.2004.03.007
Sheehan D, Meade G, Foley VM, Dowd CA (2001) Structure,
function and evolution of glutathione transferases: implications
for classification of non-mammalian members of an ancient
enzyme superfamily. Biochem J 360:1–16. doi:10.1042/0264-
6021:3600001
Siciliano V, Genre A, Balestrini R, Cappellazzo G, deWit PJ,
Bonfante P (2007) Transcriptome analysis of arbuscular mycor-
rhizal roots during development of the prepenetration apparatus.
Plant Physiol 144:1455–1466. doi:10.1104/pp.107.097980
Sikorski MM, Biesiadka J, Kasperska AE, Kopciska J, Lotocka B,
Golinowski W, Legocki AB (1999) Expression of genes
encoding PR10 class pathogenesis-related proteins is inhibited
in yellow lupine root nodules. Plant Sci 149:125–137.
doi:10.1016/S0168-9452(99)00148-X
Stuhlfelder C, Mueller MJ, Warzecha H (2004) Cloning and
expression of a tomato cDNA encoding a methyl jasmonate
cleaving esterase. Eur J Biochem 271:2976–2983. doi:10.1111/
j.1432-1033.2004.04227.x
Szatmari A, Ott PG, Varga GJ, Besenyei E, Czelleng A, Klement Z,
Bozso Z (2006) Characterisation of basal resistance (BR) by
expression patterns of newly isolated representative genes in
tobacco. Plant Cell Rep 25:728–740. doi:10.1007/s00299-005-
0110-5
Takemoto D, Yoshioka H, Doke N, Kawakita K (2003) Disease
stress-inducible genes of tobacco: expression profile of elicitor-
responsive genes isolated by subtractive hybridization. Physiol
Plant 118:545–553. doi:10.1034/j.1399-3054.2003.00145.x
Taler D, Galperin M, Benjamin I, Cohen Y, Kenigsbuch D (2004) Plant R
genes that encode photorespiratory enzymes confer resistance
against disease. Plant Cell 16:172–184. doi:10.1105/tpc.016352
Tao Y, Xie Z, Chen W, Glazebrook J, Chang HS, Han B, Zhu T, Zou
G, Katagiri F (2003) Quantitative nature of Arabidopsisresponses during compatible and incompatible interactions with
the bacterial pathogen Pseudomonas syringae. Plant Cell
15:317–330. doi:10.1105/tpc.007591
Tellstrom V, Usadel B, Thimm O, Stitt M, Kuster H, Niehaus K
(2007) The lipopolysaccharide of Sinorhizobium meliloti sup-
presses defense-associated gene expression in cell cultures of the
host plant Medicago truncatula. Plant Physiol 143:825–837.
doi:10.1104/pp.106.090985
Thilmony R, Underwood W, He SY (2006) Genome-wide transcrip-
tional analysis of the Arabidopsis thaliana interaction with the
plant pathogen Pseudomonas syringae pv. tomato DC3000 and
the human pathogen Escherichia coli O157:H7. Plant J 46:34–
53. doi:10.1111/j.1365-313X.2006.02725.x
Truman W, de Zabala MT, Grant M (2006) Type III effectors
orchestrate a complex interplay between transcriptional networks
to modify basal defence responses during pathogenesis and
resistance. Plant J 46:14–33. doi:10.1111/j.1365-313X.2006.
02672.x
Wendler WM, Kremmer E, Forster R, Winnacker EL (1997)
Identification of pirin, a novel highly conserved nuclear protein.
J Biol Chem 272:8482–8489. doi:10.1074/jbc.272.43.27091
Zimmermann P, Hirsch-Hoffmann M, Hennig L, Gruissem W (2004)
Genevestigator. Arabidopsis microarray database and analysis
toolbox. Plant Physiol 136:2621–2632. doi:10.1104/pp.104.
046367
646 Plant Mol Biol (2009) 70:627–646
123