regulation of nodule senescence in medicago truncatula
TRANSCRIPT

HAL Id: tel-03360702https://tel.archives-ouvertes.fr/tel-03360702
Submitted on 1 Oct 2021
HAL is a multi-disciplinary open accessarchive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come fromteaching and research institutions in France orabroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, estdestinée au dépôt et à la diffusion de documentsscientifiques de niveau recherche, publiés ou non,émanant des établissements d’enseignement et derecherche français ou étrangers, des laboratoirespublics ou privés.
Regulation of Nodule Senescence in Medicago truncatulaLi Yang
To cite this version:Li Yang. Regulation of Nodule Senescence in Medicago truncatula. Molecular biology. UniversitéCôte d’Azur, 2020. English. NNT : 2020COAZ6022. tel-03360702

Régulation de la sénescence nodositaire chez Medicago truncatula
Li YANG Institut Sophia-Agrobiotech (ISA), Equipe Symbiose
Présentée en vue de l’obtention du grade de docteur en: Sciences de la vie et de la santé, Interactions moléculaires et cellulaires d’Université Côte d’Azur Dirigée par :
Pr. Pierre FRENDO
Dr. Éric BONCOMPAGNI Soutenue le: 30 Septembre 2020
Devant le jury, composé de :
Paola FURLA, Président du jury Professeure, Université Côte d’Azur, Nice
Éric GIRAUD, Rapporteur Directeur de Recherche IRD, LSTM Montpellier
Andreas NIEBEL, Rapporteur Directeur de Recherche CNRS, LIPM Toulouse
Florian FRUGIER, Examinateur Directeur de Recherche CNRS, IPS2 Paris-Saclay
THÈSE DE DOCTORAT

Régulation de la sénescence nodositaire chez Medicago truncatula
Jury : Président du jury Pr. Paola FURLA, Professeure, Université Côte d’Azur, Nice Rapporteurs Dr. Andreas NIEBEL, Directeur de Recherche CNRS, LIPM Toulouse Dr. Éric GIRAUD, Directeur de Recherche IRD, LSTM Montpellier Examinateur Dr. Florian FRUGIER, Directeur de Recherche CNRS, IPS2 Paris-Saclay Codirecteurs de thèse Dr. Éric BONCOMPAGNI, Maître de conférences (HDR), Université Côte d’Azur Pr. Pierre FRENDO, Professeur, Université Côte d’Azur

RÉSUMÉ DE THÈSE Les légumineuses peuvent s'associer aux rhizobia pour développer de novo un organe
racinaire, la nodosité racinaire. La nodosité racinaire peut réduire l'azote atmosphérique en azote disponible pour l'hôte végétal. Ainsi, la symbiose fixatrice d'azote joue un rôle important dans l'agriculture avec des apports directs d'azote aux plantes cultivées.
Dans la première partie de la thèse, nous avons caractériser l'impact de la carence bactérienne en glutathion sur la différenciation bactérioïde et le fonctionnement des nodules lors de l'interaction symbiotique entre Medicago truncatula et Sinorhizobium meliloti. Des marqueurs physiologiques, biochimiques, cellulaires et génétiques ont été utilisés pour décrire le nodule fonctionnant dix et vingt jours après l'inoculation de la plante. Nos résultats ont montré que la carence bactérienne en glutathion n'affecte pas la différenciation bactérioïdienne. Cependant, elle induit un processus précoce de sénescence des nodosités chez M. truncatula.
Au cours de la sénescence des nodosités, les activités protéolytiques sont augmentées et se terminent par la dégradation finale des bactéroïdes et des cellules végétales. Par conséquent, les protéases se révèlent être les enzymes caractéristiques de la sénescence des nodosités. Au début de la sénescence nodositaire, une cystéine protéase de la famille des papaïnes, MtCP6 est un bon marqueur moléculaire pour l'initiation de la sénescence des nodules. En ce qui concerne la deuxième partie de la thèse, une analyse consistant en une série de délétion du promoteur de MtCP6 a été réalisée pour identifier les séquences régulatrices présentes sur le promoteur MtCP6 et impliquées dans la sénescence nodulaire. Afin de comprendre la régulation transcriptionnelle de la sénescence nodositaire chez M. truncatula. Ensuite, la région cis-régulatrice identifiée (67 pb, NS-box) a été validée pour fonctionner dans l'activation de la transcription dans la zone nodulaire III-IV. Nous avons pu montrer que les tétramères de NS-box peuvent induire la transcription dans la zone de sénescence de la nodosité. Afin de déterminer l'importance de NS-box par rapport au promoteur complet (–1720 pb), une analyse fonctionnelle a été réalisée avec suppression de la boîte NS sur le promoteur complet et sur le promoteur minimal (–242 pb). De plus, la validation les rôle potentiel de motif CW localisés dans la séquence NS-box en 5’ a été réalisée à l’aide de délétions sites spécifiques. Finalement, la technique de “yeast-one-hybrid” a été utilisée pour identifier des facteurs de transcription interagissant avec le fragment NS-box. Les résultats préliminaires sont présentés.
Dans leur ensemble, les résultats permettent une meilleure compréhension de la régulation et du fonctionnement de la sénescence nodulaire.
Mots clés : Sénescence nodulaire, Glutathion, Cystéine protéase, Régulation transcriptionnelle, Eléments cis-régulateur

ABSTRACT Leguminous plants are able to associate with rhizobium to develop de novo a root organ,
the root nodule. Root nodule can reduce atmospheric nitrogen into available nitrogen to plant host. Thus, the nitrogen-fixing symbiosis plays important roles in agriculture with direct nitrogen inputs to crops.
In the first part of the thesis, we intended to characterize the impact of the bacterial glutathione deficiency on the bacteroid differentiation and the nodule functioning during the symbiotic interaction between Medicago truncatula and Sinorhizobium meliloti. Physiological, biochemical, cellular and genetic markers were used to describe the nodule functioning at 10- and 20- days after inoculation. Our results showed that the bacterial glutathione deficiency does not affect bacteroid differentiation. However, it induces an early nodule senescence process in M. truncatula.
During nodule senescence, proteolytic activities are increased and terminated with the final degradation of bacteroids and plant cells. Hence, proteases are found to be the hallmarks of nodule senescence. At the onset of nodule senescence, a papain family cysteine protease, MtCP6, is a good molecular marker for the initiation of nodule senescence. In the second part of the thesis, serial promoter deletion analysis of a cysteine protease gene MtCP6 was conducted to identify cis regulatory element involved in the transcriptional regulation of nodule senescence of M. truncatula. Thereafter, the identified cis-regulatory region (67bp, NS-box) was validated to function in transcriptional activation in nodule zone III-IV. We have shown that the tetramer of the NS-box can induce the transcription at the onset of nodule senescence zone. In order to determine the significance of the NS-box, loss of function analyses were conducted with the deletion of the NS-box from the native MtCP6 promoter (–1720bp) and site-specific block deletions from the minimal MtCP6 promoter (–242bp). The results indicate a potential role of a CW motif localized at 5’ of the NS-box sequence. Finally, a yeast one hybrid experiment was performed to identify transcription factors interacting with NS-box with preliminary results presented.
Taken together, these data allow a better understanding of the regulation and the operation of the nodule senescence.
Key words: Nodule senescence, Glutathione, Cysteine protease, Transcriptional regulation, cis-regulatory element

Acknowledgements
Every step I have taken leads the way to the end of my thesis. I am indebted to so many people who have greatly supported me for the last four years. Language here is such a symbolic abbreviation which could not encapsulate all my grateful sensations.
First of all, I would like to thank the members of my thesis jury: Pr. Paola Furla, Dr. Éric Giraud, Dr. Andreas Niebel and Dr. Florian Frugier, for accepting the invitation of my PhD defence. All the jury members evaluated my thesis with scientific scrutiny, which also allowed a lively discussion in my defence.
I would show my foremost gratitude to my supervisor Prof. Pierre Frendo for accepting me to do PhD research in Symbiose team. He has been very responsible and committed in supervising me throughout my PhD. I am very grateful for the time he has invested in my PhD thesis, my research work and training me to be a scientist. He is also very considerate to include the GSH project in the course my PhD to guarantee enough workload for my graduation. Finally, it gives a great opportunity so that I could develop some knowledge of symbiosis based on observation rather than merely reading. He has also very clear thinking in science that guides me out of many subtleties.
My greatest acknowledgement is undoubtedly to my supervisor Dr. Éric Boncompagni. He has been the one always encouraging and supporting me with his positive thinking and inspiring instructions. He is the first one showing me all the manipulation basics in the lab and always providing me his greatest help. He has been harbouring his warming heart that healing me even during the downs of my PhD. Life is a long journey, and I am so grateful that I was once here meeting someone like him. He has given me countless opportunities to develop as a scientist as well as to follow my own willingness.
I am so obliged to Dr. Fernanda de Carvalho-Niebel for all the inspiring communications and detailed supervision in Toulouse, as well as the manuscript modifications. It was a great opportunity to collaborate in her lab in Auzeville where I conducted the yeast screening. She is holding the symbol of wisdom in Chinese Zodiac, providing wise ideas in research and efficient communications in science. She has put forward the project with considerable contributions.
Another one I would thank in Toulouse is Lisa Frances. She is so kind herself and has helped me with many practical considerations. She has facilitated the research with her skills and commitments, which has ensured the experiments (yeast and GUS fluorimetric assay) work out efficiently. I am grateful for all the other members in the ENOD team and have enjoyed the time with a collaborative and motivating environment.

Particular thanks to Dr. Andreas Niebel for all his help and the kind gift (cDNA library for screening). He has also offered me the help in spelling and grammar modifications as a reporter of the thesis jury. Many thanks for the dedicated language checking and scientific instructions.
I would show my sincere appreciation to Dr. Florian Frugier, who has enlightened the project with expertise suggestions as well as multiple practical considerations following my PhD project and in my PhD defence. I have been greatly inspired by his research ideas.
I am so grateful to Dr. Éric Galiana, who has also been very nice and helpful in the thesis committee.
Thanks to Pr. Paola Furla, who was genial in my defence and provided many solutions to improve my thesis manuscript.
Thanks to Dr. Éric Giraud. I am impressed by his efficiency of the report and the discussion of my defence, which conclude many questions and limitations of my thesis. I am also very thankful for his genial encouragement in my research career.
I am very grateful for all the members in Symbiose team. Thanks to Alexandre, Laurence, Mélanie, Aurélie, Claude, Isabelle, Karine and Nicolas, who have contributed to the dynamic ambience in the lab.
Thank Geneviève for being a very tranquil company around the lab bench and providing me many helps.
Thank Marc for being so peaceful himself. He has been very flexible to help me, especially in plant growth.
Thank Marie for spending so much energy in the lab affairs that guarantees our daily manipulations.
Thanks to Renaud who has organized the hiking club that was relaxing and memorable.
Thank Camille for filling my PhD life with her sunshine and colours. She has been very helpful to me in research and a very good friend in life.
Thank Martina for providing me many experiences in symbiosis study when I arrived in the lab.
Thank other doctoral students, Gaurav and Antoine, who have been very supportive and helpful.
Thank the interns Marie Francois and Pietro Giraud, who have performed pretty Haribo nodule staining.
Thank Olivier for his considerate help in microscopy and his profound work has anchored the CP6 study.
Thank Bruno for his kind help and generously providing me the cloning vectors. It has been the first turning point of my project after discussing with him.

Thank Joffrey for his generous help in technics and friendly considerations in daily life.
Thanks to Julie for providing me very detailed instructions in flow cytometry.
Last but not least, I have had very happy hours with YanYan(焱焱), YuSha(昱莎), Bin(滨), Chen(晨),
Jianlong(建龙), FengLuan(凤銮), YongPan(永攀), XinYue(馨月) and RuoHan(若涵). Thank you all for
being there for some time in the last four years and backing me up in all kinds of aspects.
Special thanks to Jianlong, who has provided me very generous and useful help in molecular biology techniques in the beginning of my PhD project.
And to all my families and beloved, no matter how far I am away, all my dreams are bound to you. It is definitely a life-long memory of one’s PhD thesis, and mine is very special with you all. I have had the fortune to see how life can be led; finally, my sketches are still on the way.
Antibes, autumn 2020


Table of Contents RÉSUMÉ DE THÈSE ............................................................................................................. 4
ABSTRACT .......................................................................................................................... 5
Acknowledgements ........................................................................................................... 6
Table of Contents ............................................................................................................. 10
Figure list ......................................................................................................................... 13
Table list .......................................................................................................................... 15
GENERAL INTRODUCTION ................................................................................................ 19 I. Nitrogen nutrition ............................................................................................................ 21
I.1 Anthropogenic changes to the global nitrogen cycle ........................................................................ 21 I.2 Plant nitrogen nutrition ..................................................................................................................... 23
II. Biological nitrogen fixation (BNF) ...................................................................................... 27 II.1 Biological nitrogen fixers .............................................................................................................. 27 II.2 Legume-Rhizobia symbiosis .......................................................................................................... 30
III. Medicago truncatula and Sinorhizobium meliloti symbiotic model system ....................... 33 III.1 Medicago truncatula .................................................................................................................... 33 III.2 Establishment of symbiosis........................................................................................................... 37
III.2.1 Recognition of legumes and rhizobia ....................................................................................... 37 III.2.2 Bacterial infection is coupled with nodule organogenesis ...................................................... 38 III.2.1 Bacteroid differentiation ......................................................................................................... 41 III.2.2 Nitrogen fixation in bacteroids ................................................................................................ 48
IV. Senescence ................................................................................................................... 50 IV.1 Senescence as a common process ................................................................................................ 50 IV.2 Regulation of senescence ............................................................................................................. 52
IV.2.1 Regulation of plant senescence ............................................................................................... 52
The coordinated death mechanism of plant-host and rhizobia (a review, Kazmierczak et al., 2020)…………...61
IV.2.2 Legume-rhizobium symbiotic nodule senescence ................................................................... 55
The involvement of proteases in the nodule senescence (a review, Yang, Li et al., 2020).………………...……98
OBJECTIVES .................................................................................................................... 113
CHAPTER I ...................................................................................................................... 115 I. Introduction ................................................................................................................... 117
I.1 Redox homeostasis in nodulation ................................................................................................... 117 I.2 Glutathione in prokaryotes ............................................................................................................. 119 I.3 Role of glutathione in nodule functioning. ...................................................................................... 121
II. Results ............................................................................................................................ 123 III. Discussion and perspectives ........................................................................................... 141
CHAPTER II ..................................................................................................................... 147 I. Introduction ................................................................................................................... 149
I.1 Transcriptional regulatory system in eukaryotes ............................................................................ 149

I.1.1 Transcriptional regulation ...................................................................................................... 149 I.1.2 Cis-regulatory elements ......................................................................................................... 152
I.2 Cysteine proteases (CPs) in nodule senescence .............................................................................. 155 I.2.1 Role of cysteine protease 6 (MtCP6) in nodule senescence .................................................. 157
II. Results (manuscript) ....................................................................................................... 161 III. Discussion ....................................................................................................................... 197 IV. Perspectives ............................................................................................................... 198
CHAPTER III .................................................................................................................... 203 I. Introduction ................................................................................................................... 205
I.1 Searching for NS-box-interacting factors ........................................................................................ 205 I.2 TFs associated with nodule senescence processes in legume plants .............................................. 206 I.3 Search for trans-regulatory elements interacting with NS-box ....................................................... 207
II. Material and Methods .................................................................................................... 208 III. Results ............................................................................................................................ 209 IV. Discussion .................................................................................................................. 215 V. Summary of potential transcriptional actors for MtCP6 expression ................................. 223 VI. Perspectives ............................................................................................................... 225
GENERAL DISCUSSION & PERSPECTIVES ......................................................................... 231 I. Disruption of symbiotic partnership during root nodule senescence ............................... 233 II. Deciphering nodule senescence mechanism at transcriptional level ............................... 236 III. The future of symbiotic nitrogen fixation ........................................................................ 240
ANNEX I ......................................................................................................................... 243 I. Plant and bacteria materials ........................................................................................... 245
I.1 Medicago truncatula ....................................................................................................................... 245 I.1.1 Seed sterilization and germination ........................................................................................ 245
I.2 Nicotiana benthamiana plant growth conditions ............................................................................ 246 I.3 Bacterial materials ........................................................................................................................... 246
II. Molecular biology methods ............................................................................................ 247 II.1 Cloning ........................................................................................................................................ 247 II.2 Transforming bacteria competent cells ...................................................................................... 249 II.3 Composite transgenic plants ...................................................................................................... 250 II.4 Yeast-One-Hybrid screening (Y1H) ............................................................................................. 253
II.4.1 Yeast strain, culture conditions and transformation. ............................................................ 253 II.4.2 Generation and selection of a 4x NS-box YM4271 yeast reporter strain .............................. 254 II.4.3 Yeast-one hybrid screen of a nodule cDNA library ................................................................ 255
III. Phenotyping ................................................................................................................... 257 III.1 Plant phenotyping ...................................................................................................................... 257 III.2 Histology and Microscopic observation ...................................................................................... 259
ANNEX II ........................................................................................................................ 263
REFERENCES .................................................................................................................. 277

13
Figure list (not including figures in publications/manuscript)
Figure 1 Nitrogen-cycle intermediates and major processes of the nitrogen cycle.
Figure 2 Historical changes rely on nitrogen fertilizer in intensive agriculture.
Figure 3 Schematic routes of nitrogen uptake from the rhizosphere, transportation and assimilation, and remobilization inside the plant.
Figure 4 Phylogenetic 16S tree in prokaryotes carrying nif genes.
Figure 5 Phylogeny of the Papilionoideae subfamily.
Figure 6 Unrooted phylogenetic tree of 16S ribosomal DNA sequences from selected a-, b- and g- proteobacteria.
Figure 7 Medicago mt4.0 and mt5.0.
Figure 8 Gene expression atlas of M. truncatula.
Figure 9 Schematic representation the coordinated bacterial infection and nodule organogenesis.
Figure 10 Plant symbiotic intracellular signalling pathways.
Figure 11 Overview of nodule zones and bacteroid differentiation stages in indeterminate nodules.
Figure 12 Size, shape, and DNA content of free-living, cultured S. meliloti bacteria and bacteroids isolated from nitrogen-fixing M. truncatula nodules.
Figure 13 Endoreduplication of the host plant cells is coupled with bacteroid differentiation in nodules.
Figure 14 Nitrogenase regulation, gene and structure.
Figure 15 Idealized growth and viability curves terminated at activating senescence.
Figure 16 Restoration of the atmyb2-1 plants to the wild type (WT), morphologically and in terms of levels of endogenous cytokinin, by intact AtMYB2 or by PAtMYB2-AtCKX1.
Figure 17 The nitrogen-fixing and senescent nodules of M. truncatula.
Figure 18 Microscopic analysis of nodule senescence in M. truncatula.
Figure 19 Microscopic analysis of the dark-induced nodule senescence in M. truncatula.
Figure 20 The cDNA-AFLP samples for transcriptome analysis nodule senescence in M. truncatula.
Figure 21 Antioxidant enzymes of legume nodules.
Figure 22 Expression of gshB in laser-dissected zones of 15-day-old nodules.
Figure 23 Model of the transcriptional machinery.
Figure 24 The feedforward loop, single-input module (SIM) and dense-overlapping regulons (DOR) found in transcription networks.
Figure 25 Classes of known regulatory elements.

14
Figure 26 Histochemical analysis of MtCP6 expression throughout nodule development or induced nodule senescence.
Figure 27 Transcriptional activation of CP genes at the onset of nodule senescence.
Figure 28 Schematic representation of the transcriptional machinery under control of activator (A) and repressor (B) TFs, respectively.
Figure 29 Schematic overview of the yeast one-hybrid screening procedure.
Figure 30 Yeast-one-hybrid (Y1H) screening strategy to identify NS-box interacting proteins.
Figure 31 Evaluating the specificity of the interaction of the erf candidates to the 4x NS reporter in yeast.
Figure 32 MtERF091 (MtERF1-1) homologues function in leguminous plants.
Figure 33 Analysis of the DNA-binding domain (ERF domain) from the ERF candidates and the homologs in Arabidopsis.
Figure 34 A GCCcore-like motif is present in NS-box.
Figure 35 Summary of transcriptional elements presumably involved in the expression of MtCP6 during nodule senescence.
Figure 36 Flow chart of perspectives to characterize TF candidates.
Figure S1 Example of transgenic roots of M. truncatula generated by A. rhizogenes Arqua1.
Figure S2 Heterogenous nodule size appears at 20- and 28dpi following declining nitrogen fixation capacity.
Figure S3 Kinetics of histochemical localization of β-glucuronidase (GUS) activity in nodules carrying promoter-GUS constructions for the MtCP6.
Figure S4 Putative cis-regulatory elements present in the minimal functional MtCP6 promoter (–242bp).
Figure S5 Re-growth and yeast colony PCR of Y1H candidates.
Figure S6 Alignment of ERF candidate sequences (translated protein sequences) to the identical putative ERFs (protein sequence from Mt4.0).
Figure S7 Expression profile of ERF-091, -092, -069 and -073 on MTGEA: M. truncatula gene expression atlas / Noble foundation (https://mtgea.noble.org/v3).

15
Table list (not including tables in publications/manuscript)
Table 1 Examples of organisms that can carry out nitrogen fixation.
Table 2 Associations between rhizobia and host plants.
Table 3 Cell types of developmental nodule senescence.
Table 4 Comparison of developmental and dark stress induced nodule senescence.
Table 5 Relative expressions of cysteine protease encoding genes and putative DNA-binding protein encoding genes during root nodule senescence.
Table 6 Phenotypes of GSH deficient rhizobia strains.
Table 7 Cysteine protease genes of M. truncatula involved in nodule senescence with different ID.
Table 8 List of AD in-frame His+ candidates.
Table S1 Strains and plasmids used during the course of this thesis.
Table S2 Examples for BP and three-entry multisite LR reaction mix.
Table S3 List of primers used in yeast-one-hybrid screen strategy
Table S4 Web-based resources for plant cis-regulatory sequence analysis used in the course of this thesis.

16

17
Smoothest ice
A paradise
To him, who is a dancer nice
Friedrich Nietzsche

18

19
GENERAL INTRODUCTION

20

GENERAL INTRODUCTION
21
I. Nitrogen nutrition
I.1 Anthropogenic changes to the global nitrogen cycle
Nitrogen (N) is one of the primary nutrition elements essential for life, as component of many organic molecules in cell, particularly: amino acids (proteins), nucleic acids (DNA and RNA), and macromolecules such as chlorophyll. In fact, there is an enormous N stock in the ecosphere, but mostly unavailable as in atmospheric dinitrogen (N2), accounting for 78% of Earth’s atmosphere (Stevens, 2019), or deeply in rocks and sediments. Thus available N for the plant is often limiting in natural conditions, albeit the global N status is changing with increasing rates of N production, N deposition and agricultural N application (Reay et al., 2008).
The global N production available for use by living organisms is either achieved by natural processes such as lightning and biological N fixation (BNF) or anthropogenic activities like the Haber Bosch process, in a reactive form (Nr) which includes inorganic reduced forms of N (e.g., NH3 and NH4+), inorganic oxidized forms (e.g., NOx, HNO3, N2O, and NO3–), and organic compounds (e.g., urea, amines, and proteins), by contrast to unreactive N2 gas (Galloway et al., 2008) (Figure 1). Human activities transform the global N cycle over the past century, bringing about a doubled global N cycling in comparation to natural N production (anthropogenic Nr production: 210 Tg N per year, natural Nr production: 203 Tg N per year) (Fowler et al., 2013). The changes in Nr production are reflecting an increasing combustion of fossil fuels, high N demand in agriculture and industry, and inefficient N utilization (Galloway
et al., 2008). Excessive N is largely lost into land, water and air, causing a cascade of environmental problems.
The rising Nr emissions is mostly thought to be deposited on terrestrial ecosystems, in addition to the open ocean and coastal zones (Reay et al., 2008). For example, from 1900 to 1980, deposition of oxidized N (such as NOX and N2O) in Europe increased by three to four times, whereas deposition of reduced N (such as ammonia, NH3) doubled (Engardt et al., 2017). However recently, the N concentrations in plant tissues, often considered an indicator of the amount of N taken up by plants or their N status, have been recently shown declined globally between 1980 and 2017 despite globally increasing availability of N (Craine et al., 2018), reflecting an inefficient N use.
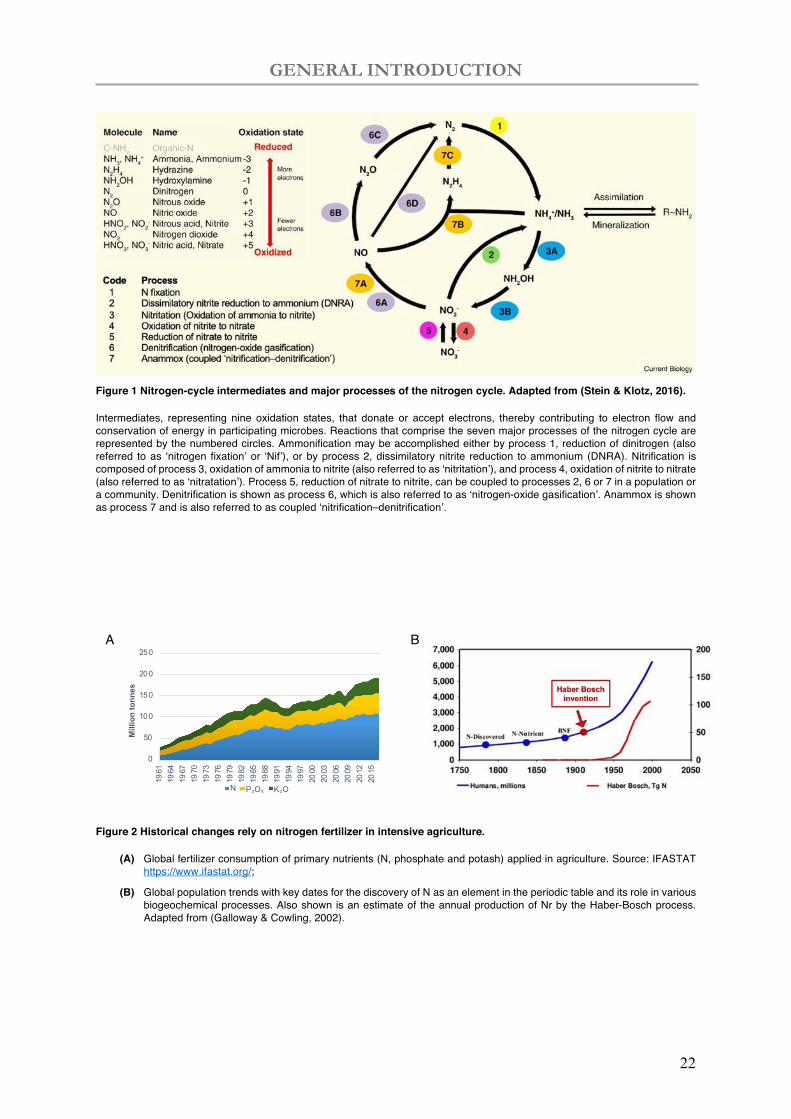
GENERAL INTRODUCTION
22
Figure 1 Nitrogen-cycle intermediates and major processes of the nitrogen cycle. Adapted from (Stein & Klotz, 2016).
Intermediates, representing nine oxidation states, that donate or accept electrons, thereby contributing to electron flow and conservation of energy in participating microbes. Reactions that comprise the seven major processes of the nitrogen cycle are represented by the numbered circles. Ammonification may be accomplished either by process 1, reduction of dinitrogen (also referred to as ‘nitrogen fixation’ or ‘Nif’), or by process 2, dissimilatory nitrite reduction to ammonium (DNRA). Nitrification is composed of process 3, oxidation of ammonia to nitrite (also referred to as ‘nitritation’), and process 4, oxidation of nitrite to nitrate (also referred to as ‘nitratation’). Process 5, reduction of nitrate to nitrite, can be coupled to processes 2, 6 or 7 in a population or a community. Denitrification is shown as process 6, which is also referred to as ‘nitrogen-oxide gasification’. Anammox is shown as process 7 and is also referred to as coupled ‘nitrification–denitrification’.
Figure 2 Historical changes rely on nitrogen fertilizer in intensive agriculture.
(A) Global fertilizer consumption of primary nutrients (N, phosphate and potash) applied in agriculture. Source: IFASTAT https://www.ifastat.org/;
(B) Global population trends with key dates for the discovery of N as an element in the periodic table and its role in various biogeochemical processes. Also shown is an estimate of the annual production of Nr by the Haber-Bosch process. Adapted from (Galloway & Cowling, 2002).
0
50
100
150
200
250
1961
1964
1967
1970
1973
1976
1979
1982
1985
1988
1991
1994
1997
2000
2003
2006
2009
2012
2015
Mill
ion
tonn
es
N P₂O₅ K₂O
A B

GENERAL INTRODUCTION
23
To feed a growing population which is already over 7 billion, high demand in food production requires intensive agriculture which is largely dependent on the application of inorganic fertilizers (Figure 2A). The agricultural production has been considerable promoted since the industrial revolution (~1840), especially after the invention of Haber Bosch process (Figure 2B) in 1913. As consequences, the change in Nr production is enormous from ~15 Tg N in 1860 to 156 Tg N in 1995 and further increased to 187 Tg N in 2005 (Galloway et al., 2008). The overuse of N fertilizers has led to series of negative impacts as in reduced biodiversity, green-house-gases emissions, polluted air and water quality, soil fertility degeneration and on human health risks.
Even though contemporary focus is on solving environmental problems induced by surplus fertilizers in developed countries, an increasing attention is also drawn on developing countries, wherein small-hold farmers are suffering from the lack of nutrient fertilizers (Foley
et al., 2011; Mueller et al., 2012). In these regions, scarce N availability intensifies to the existing food shortage problem mostly caused by failure of affording N fertilizers or due to ineffective N management. While N fertilizers may continue to dominate in intensive agriculture management for some time, a more sustainable agriculture system is ultimately required to keep the N cycle in reasonable balance for the sake of long-term utilization as well as world global conservation. Therein, we may need also in a short-term period of an integrated system using a precise management with high efficiency in N usage, as well as the combination of using biological N fixers such as legumes.
I.2 Plant nitrogen nutrition
As primary producers, plants rely on a large aboveground surface area to collect carbon dioxide and sunlight and a large underground surface area to collect the water and mineral nutrients needed to support their growth and development (Oldroyd & Leyser, 2020). Plants optimize their uptake and utilization strategies according to nutrient availability in soil. Typically, roots are well developed in harsh conditions with limited soil nutrients, which keep a limited growth of the upper part resulted in a high root: shoot ratio. Conversely, when soil resources are sufficient, a minimum root system is maintained, and vegetative growth is extended.
A widely accepted view is that the epidermal and cortical cells of the root are the main sites of uptake of ions. A proportion of these ions is subsequently diverted into vacuoles of the cells of these regions, a proportion is consumed on growth of the root, while the remainder is free to diffuse inwards along concentration gradients in the root cytoplasm, through the endodermis, to cells in the stele engaged in loss of ions to the xylem. Under situations where

GENERAL INTRODUCTION
24
plants are unable to access N and Pi from their immediate environment, they turn to these microorganisms to find new sources of these limiting nutrients. These processes regulate the plants’ receptiveness to their microbial communities, promoting symbiotic associations.
Plant nitrogen uptake relies mainly on inorganic forms
Plants mostly uptake N in inorganic forms like nitrate in aerobic soils and ammonium in flooded wetlands or acidic soils (Figure 3) (Marschner, 2011; Xu et al., 2012). In return, the resulted acidification or alkalization of the rhizosphere by N uptake changes the soil N availability for plants (Marschner, 2011). Alternatively, plants are proven to be able to utilize organic compounds, such as amides, amino acids. With nitrate and ammonium concentrations in soil solutions ranging from 100µM to 10mM, numerous membrane transporters was composed to cope with different N forms and concentrations (Figure 3) (Miller et al., 2007; Xu et al., 2012; O'Brien et al., 2016).
Plants take different routes for nitrogen transportation and storage
Upon absorption of different N sources, biosynthetic activities associated with N assimilation are also found varied in plant roots. Plants exhibit different distribution patterns of the transportation and storage in terms of N forms. In conditions where roots contain a large amount of N beyond its requirements, the remainder will be thereafter transported to shoots for utilization or storage. One extreme of nitrate response spectrum is in woody plants, in which nitrate reduction is being accomplished entirely by their roots, while it is impossible to detect free nitrate in the xylem but high levels of organic solutes of N are present (Pate, 1973) (Figure 3). The other extremes are in plants such as Borago spp. and Xanthium
pennsylvanjcum (Pate, 1973), such species have roots genetically deficient in the ability to synthesize enzymes of the nitrate reduction pathway (Figure 3). Most herbaceous species like pea, however, localize in the middle of the spectrum, are able to maintain active reductases in both their root and shoot systems (Pate, 1973).
Analyses of xylem and phloem contents suggest that the xylem supplies target organs with mostly nitrate and a limited range of amino acids, particularly the amide glutamine, whilst the phloem appears to deliver considerably less nitrate and amide but substantial amounts of sucrose and a wide range of amino acids at somewhat higher concentration than in the xylem
(Pate, 1973). As consequences, xylem nutrition brings in nutrients having on average a C:N ratio of approximately 2:1, whilst in the phloem solutes are dispensed having a C/N ratio of approximately 22:1 (Pate, 1973). The leaves are the N target organs during vegetative stage
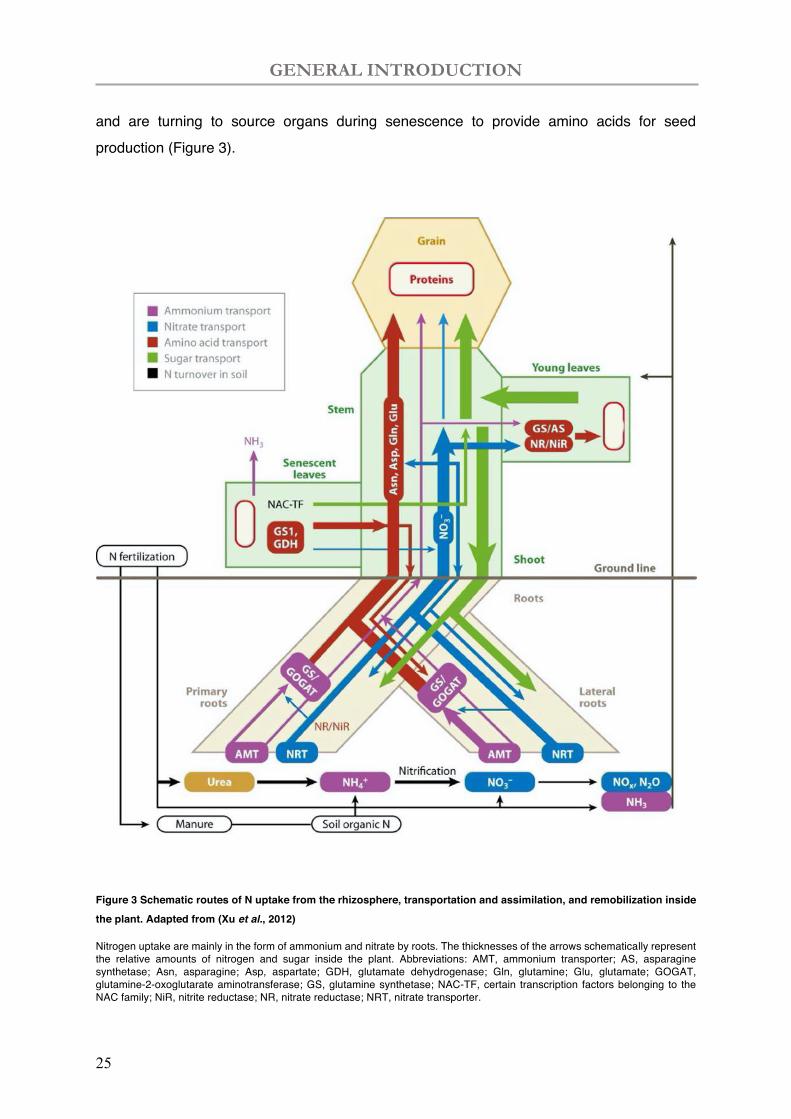
GENERAL INTRODUCTION
25
and are turning to source organs during senescence to provide amino acids for seed production (Figure 3).
Figure 3 Schematic routes of N uptake from the rhizosphere, transportation and assimilation, and remobilization inside the plant. Adapted from (Xu et al., 2012)
Nitrogen uptake are mainly in the form of ammonium and nitrate by roots. The thicknesses of the arrows schematically represent the relative amounts of nitrogen and sugar inside the plant. Abbreviations: AMT, ammonium transporter; AS, asparagine synthetase; Asn, asparagine; Asp, aspartate; GDH, glutamate dehydrogenase; Gln, glutamine; Glu, glutamate; GOGAT, glutamine-2-oxoglutarate aminotransferase; GS, glutamine synthetase; NAC-TF, certain transcription factors belonging to the NAC family; NiR, nitrite reductase; NR, nitrate reductase; NRT, nitrate transporter.

GENERAL INTRODUCTION
26
The absorbed nitrate is partially reduced and assimilated in the roots, and the larger part is transported to the shoot, where it is first reduced to nitrite by nitrate reductase in the cytoplasm and then further to ammonium by nitrite reductase in the plastids and glutamine synthetase (GS) in the plastids and cytoplasm (Xu et al., 2012) (Figure 3). The reduction of nitrate to ammonium requires reducing power which is provided by photosynthesis which produces both energy (ATP) and reducing power (NADPH). Ammonium is further assimilated into amino acids catalysed by the GS/glutamine-2-oxoglutarate aminotransferase (GOGAT) cycle (Figure 3) (Xu et al., 2012). While GS and Asparagine synthetase (AS) are crucial in primary N metabolism, the mitochondrial NADH–glutamate dehydrogenase (GDH) can alternatively incorporate ammonium into Glu in response to high levels of ammonium under stress (Masclaux-Daubresse et al., 2010).
Nitrogen is reutilized via remobilization during plant senescence
The initiation of leaf senescence is considered as the transition from carbon to nitrogen source. During leaf senescence, internal redistribution of N occurs, and leaf proteins are degraded, particularly from chloroplast, and remobilized to other organs, e.g. seeds in the reproductive stage (Figure 3) (Masclaux-Daubresse et al., 2010). In vegetative tissues, Rubisco enzyme is not only a photosynthetic enzyme, but also the repository of recoverable N, in which protein breakdown only starts after the synthesis in young growing leaves (Thomas, 2013). The N remobilization is thus regulated sequentially by chloroplastic and vacuolar proteases (Desclos et al., 2009; Masclaux-Daubresse et al., 2010; Xu et al., 2012). During N remobilization of leaf senescence, a number of proteases encoding genes are strongly induced (Masclaux-Daubresse et al., 2010).
Plants response to nitrogen availability
Plant nutrient is systematically regulated by an integrative root and shoot signalling system with a variety of hormones moving between the root and the shoot to both signal nutrient availability and coordinate plant development (Oldroyd & Leyser, 2020). There are two scenarios in case of the nitrogen regulation: positive regulation by root N concentration and negative feedback by the shoots. The root response to N involves four signalling processes: local signalling in the root associated with perception of local N; root-shoot-root signalling indicating the presence of roots experiencing low N concentration; root-shoot-root signalling indicating the presence of roots experiencing locally high N concentration; and a

GENERAL INTRODUCTION
27
systemic inhibitory signal that suppresses root N-foraging activities when shoot N concentration is sufficient (Oldroyd & Leyser, 2020).
Nitrate response has been well studied in plants. There are two types of nitrate transporters in higher plants: NRT2 are high-affinity nitrate transporters, while most members of the NRT1 family are low-affinity nitrate transporters (Tsay et al., 2007). An exception is a plasma membrane transporter NRT1.1 (CHL1), which is a master gene in apical part of root (Dechorgnat et al., 2011), playing roles as nitrate transporter and nitrate sensor of activating the Arabidopsis Nitrate Response 1 (ANR1) to promote nitrate-stimulated lateral root proliferation (Zhang & Forde, 1998; Remans et al., 2006; Ho et al., 2009). Two low-affinity nitrate transporters in Arabidopsis (AtNRT1.5 and AtNRT1.8) loading and unloading into the root stele or from the shoot vasculature (Lin et al., 2008; Li et al., 2010), and AtNRT1.9 in root companion cells facilitates the loading of nitrate into the root phloem (Wang & Tsay, 2011).
The ammonium transporters (Amt/MEP/Rh family) is widely distributed in nature with high affinity to facilitate the growth on very low concentrations (<5µM) of ammonium salts (Khademi et al., 2004). In high external ammonium concentration, modification of the cytosolic C-terminus of AtAMT1.1 and AtAMT1.2 allows rapid inactivation of the trimer complex to prevent toxic ammonium accumulation (Loque et al., 2007) (Neuhäuser et al., 2007). Ammonium is also shown to trigger lateral root development mediated by AMTs (e.g., LjAMT1;3) (Lima et al., 2010). In Arabidopsis thaliana, the GDP mannose pyrophosphorylase (GMPase) confers a hypersensitivity to NH4+ (Qin et al., 2008).
II. Biological nitrogen fixation (BNF)
II.1 Biological nitrogen fixers
Another major source of N production on earth is from the BNF (Burris & Roberts, 1993). N fixation by BNF was estimated to contribute a total of approximately 50-70 Tg N in agricultural systems (Herridge et al., 2008), and accounting for 60% of the earth N fixation (Zahran, 1999; Canfield et al., 2010). BNF is less environmentally problematic as the fixed N are used in situ. The major limitation is that BNF has not found in Eukaryotes, but it is widely distributed in bacteria and Archaea, namely diazotrophic organisms (Dixon & Kahn, 2004). They are evolved to use nitrogenase (encoding by nif genes) in fixing dinitrogen to ammonia into biosphere, in phylogenetic groups including all subdivisions of the Proteobacteria, green sulphur bacteria (Chlorbi), Firmibacteria, cyanobacteria and Archaea (Figure 4) (Franche et
al., 2009).

GENERAL INTRODUCTION
28
Figure 4 Phylogenetic 16S tree with prokaryotes carrying nif genes. Adapted from (Franche et al., 2009).
A larger part of the N fixing Prokaryotes are free-living (nonsymbiotic) in the soil (Table
1), including cyanobacteria, Anabaena and genera such as Azotobacter, Beijerinckia, Clostridium (Franche et al., 2009; Taiz et al., 2015). The ability to form a N-fixing symbiosis appears to have evolved relatively recently in land plants, approximately 65 million years ago (Doyle, 2011). In the N-fixing organisms, diverse physiologies are found (Table 1) (Taiz et al., 2015):
i) Aerobic: which maintain reduced O2 conditions (microaerobic conditions) through their high levels of respiration (e.g., Azotobacter). Others, such as Gloeothece, evolve to produce O2 photosynthetically during the day and fix nitrogen during the night.
ii) Facultative: which are able to grow under both aerobic and anaerobic conditions, generally fix nitrogen only under anaerobic conditions (e.g., Klebsiella).
iii) Anaerobic: in which O2 does not pose a problem, because it is absent in their habitat. These anaerobic organisms can be either photosynthetic (e.g., Rhodospirillum), or nonphotosynthetic (e.g., Clostridium).
Alternatively, some other bacteria form host-specific mutualistic associations with higher plants, e.g. Rhizobia & leguminous plants, Frankia & dicotyledonous species (actinorhizal plants) (Table 1) (Franche et al., 2009; Taiz et al., 2015). In the case of Gunnera, these organs are existing stem glands that develop independently of the symbiont. In the case of legumes and actinorhizal plants, the N-fixing bacteria induce the plant to form root nodules. (Taiz et al., 2015) In addition, sugarcane and the tiny water fern Azolla can also form associations. with

GENERAL INTRODUCTION
29
the cyanobacteria Nostoc and Anabaena, respectively (Taiz et al., 2015). N fixing shrub such as gorse, broom, etc. are often pioneers stabilizing soils and increasing N content.
Table 1 Examples of organisms that can carry out nitrogen fixation. Adapted from (Taiz et al., 2015).
Symbiotic nitrogen fixation
Host plant N-fixing symbionts Legumes, Parasponia Rhizobia
Actinorhizal Frankia
Gunnera Nostoc
Azolla Anabaena
Sugarcane Acetobacter
Free-living nitrogen fixation
Type N-fixing genera Cyanobacteria Anabaena, Calothrix, Nostoc
Aerobic: Azospirillum, Azotobacter, Beijerinckia, Derxia
Facultative: Bacillus, Klebsiella
Anaerobic:
Nonphotosynthetic Clostridium, Methanococcus (archaebacterium)
Photosynthetic Chromatium, Rhodospirillum
Amongst the symbiotic interactions, the actinorhizal symbioses are mutualistic
relationships between the actinomycete genus Frankia and a number of dicotyledonous plant genera belonging to eight diverse plant families. The symbiosis occurs in a new root organ called the root nodules. Like in legume/rhizobia symbioses, bacteria can enter the plant root either intracellularly through an infection thread formed in a curled root hair, or intercellularly without root hair involvement, and the entry mechanism is determined by the host plant species. Nodule primordium formation is induced in the root pericycle as for lateral root primordia. Mature actinorhizal nodules are coralloid structures consisting of multiple lobes, each of which represents a modified lateral root without a root cap, a superficial periderm and with infected cells in the expanded cortex. The actinorhizal plants share a predilection for marginally fertile soils and the majority are pioneers on nitrogen-poor sites. In addition, many actinorhizal species are able to tolerate environmental stresses such as high salinity, drought, and cold. They inhabit a variety of ecosystems, including coastal dunes, alpine communities, arctic tundra, and forests. Actinorhizal plants are especially important in high latitude regions, such as Scandinavia, Canada, and Alaska, where actinorhizal plants are capable of vigorous

GENERAL INTRODUCTION
30
growth (Wall, 2000). Much of the new nitrogen entering these ecosystems comes from the actinorhizal symbioses that, on the whole, account for over 15% of the biologically fixed nitrogen worldwide.
II.2 Legume-Rhizobia symbiosis
The most studied nitrogen-fixing symbiosis is between Fabaceae and rhizobia (Azorhizobium, Bradyrhizobium, Photorhizobium, Rhizobium, and Sinorhizobium) (Table 2) (Poole et al., 2018). Such symbioses lead to the formation of de novo organ, the root nodule. Once the primary source of N for agricultural systems, Symbiotic Nitrogen Fixation (SNF) in legumes (approximately 50 million tons per year) contributes to less than half of what is provided by chemical fertilizer N (Canfield et al., 2010). Legumes have evolved about 60 Million years ago. However, even if several lineages are evolved with N-fixing symbiosis, not all legumes are undergoing N-fixing symbiosis and each legume species has its own cognate rhizobia partners.
Table 2 Associations between rhizobia and host plants. Adapted from (Poole et al., 2018).
Rhizobia Plant host species Azorhizobium caulinodans Sesbania rostrata
Azorhizobium doebereinerae Sesbania virgata
Bradyrhizobium japonicum Glycine max
Bradyrhizobium spp. Cajanus cajun, Glycine max, Aeschynomene americana, Aeschynomene indica, Aeschynomene rudis, Arachis duranensis and Arachis hypogaea
Burkholderia spp. Mimosa pudica
Cupriavidus sp. AMP6 Mimosa asperata
Cupriavidus taiwanensis Mimosa pudica
Mesorhizobium haukuii Sesbania sesban
Mesorhizobium loti Lotus japonicus and Lotus corniculatus
Rhizobium etli Phaseolus vulgaris
Rhizobium gallicum Phaseolus vulgaris
Rhizobium leguminosarum bv. phaseoli Phaseolus vulgaris
Rhizobium leguminosarum bv. trifolii Trifollium spp.
Rhizobium leguminosarum bv. viciae Pisum sativum, Lens culinaris, Vicia cracca, Vicia hirsuta and Vicia faba
Rhizobium sp. strain IRBG74 Sesbania cannabina, Sesbania bispinosa, Sesbania cannabina, Sesbania exasperata, Sesbania formosa, Sesbania grandiflora, Sesbania madagascariensis, Sesbania macrantha and Sesbania pachycarpa
Rhizobium tropici Phaseolus vulgaris
Sinorhizobium fredii NGR234 112 genera
Sinorhizobium fredii USDA257 79 genera
Sinorhizobium meliloti and Sinorhizobium medicae
Medicago sativa and Medicago truncatula

GENERAL INTRODUCTION
31
Nitrogen fixing symbiosis with leguminous plants (Leguminosae)
Leguminosae (Fabaceae) is the third largest angiosperm family with nearly 770 genera and over 19,500 species (Azani et al., 2017). As for economic importance, the legume family is second after Poaceae, providing nutrition sources to benefit health and livelihoods. Legumes are long-recognized with three subfamilies: Caesalpinioideae, Mimosoideae, and Papilionoideae. However recently, a new classification of six subfamilies: Cercidoideae, Detarioideae, Duparquetioideae, Dialioideae, Papilionoideae and the recircumscribed Caesalpinioideae, is published on agreement of the legume systematics community based on emerging updating phylogenetic analyses (Azani et al., 2017). Papilionoideae is the largest subfamily with 14 000 species (Figure 5) (Azani et al., 2017), wherein we found many nodulated legume species such as Glycine max (soybean), Phaseolus vulgaris (common bean), Cicer arietinum (chickpea), Vicia faba (broad bean) and Pisum sativum (pea), as well as two well-studied model legumes (Lotus japonicus and Medicago truncatula) (Figure 5).
Figure 5 Phylogeny of the Papilionoideae subfamily. Adapted from (Azani et al., 2017).
Depicted on a 95% majority-rule Bayesian consensus tree based on analysis of peptide sequences from 81 plastid encoded proteins, subsampling representative taxa from forthcoming phylogenomic analyses.
The rhizobia symbiotic partner
The bacterial partner of the symbiosis is from Alphaproteobacteria and Betaproteobacteria groups, collectively called rhizobia, currently consisting 98 species in 13
genera and referred to as a- or b-rhizobia (https://www.rhizobia.co.nz/taxonomy/rhizobia)

GENERAL INTRODUCTION
32
(Figure 6) (Masson-Boivin et al., 2009). Most of these bacterial species are in the
Rhizobiaceae family in the a-proteobacteria and are in either the Rhizobium, Mesorhizobium,
Ensifer (Sinorhizobium), or Bradyrhizobium genera. All a-rhizobial genera belong to the
Rhizobiales order, whereas b-rhizobial genera belong to the Burkholderiales order. It is important to note that there are other non-rhizobial species present in these genera. For example, in the Rhizobium genus, there is a well-known non-nitrogen-fixing Agrobacterium tumefaciens.
Figure 6 Unrooted phylogenetic tree of 16S ribosomal DNA sequences from selected a-, b- and g- proteobacteria. Adapted from (Masson-Boivin et al., 2009).
Genera in bold font contain rhizobia. Rhizobial species or strains whose genomes have been fully sequenced are indicated between brackets. Genome sequences have been published for C. taiwanensis, M. loti, S. meliloti, R. leguminosarum, R. etli, A. caulinodans, Sinorhizobium sp. strain NGR234, B. japonicum and Bradyrhizobium sp. BTAi1 and ORS278. Rhizobial genomes with sequencing projects in progress are indicated in square brackets.

GENERAL INTRODUCTION
33
III. Medicago truncatula and Sinorhizobium meliloti symbiotic model system
III.1 Medicago truncatula
M. truncatula belongs to the galegoid clade (also known as the inverted repeat-lacking clade, IRLC). M. truncatula is the preeminent model legumes in the study of symbiosis of indeterminate nodules (for determinate: L. japonicus). It is autogamous, with a short generation time and large seed production. It is a diploid specie (2n=16) with a fully sequenced genome of approximately 500 Mb (http://data.kew.org/cvalues/) (Arumuganathan & Earle, 1991), making the genomic and genetic manipulations amendable. A knowledge-based database of this model legume is available at the website LeGOO (Legume Graph-Oriented Organizer) (https://www.legoo.org) (Carri Re et al., 2020).
Medicago genome analysis
The Medicago genome sequencing was initiated in 2003 and published in 2011 (Mt3.5) based on BACs (bacterial artificial chromosomes) sequencing (Young et al., 2011). In total, there are 62,388 gene loci in Mt3.5, with 3,692 (5.9%) putative TF genes which are significantly nodule-enhanced (Young et al., 2011). A refined genome version (Mt4.0; Figure 7A) was released in 2014 using Illumina technology and ALLPATHS-LG assembly method (Tang et al., 2014). A 30% additional sequences are encompassed in Mt4.0 (~360 Mb) than Mt3.5 (246Mb). Mt4.0 includes 50,894 genes and is annotated with a hybrid approach of EST, RNA-seq, protein and gene prediction evidences (Tang et al., 2014). The latest version of M. truncatula genome sequences (Mt5.0; Figure 7B) was released in 2018 and developed using PacBio sequencing by Laboratory of Plant-Microbe Interactions (LIPM) (Pecrix et al., 2018). Based on that coregulated genes often colocalize, symbiosis-related islands (SRIs) are proposed for the genomic clustering of coregulating genes, provoking SRIs upregulated in nodules or expressed in the nodule differentiation zone (270 and 211, respectively) (Pecrix et al., 2018). The nodule enhanced 270 SRIs are consist of 1,960 genes in total, with a mean size of ~46 kb with ~7.3 coregulated genes on average, representing ~87% of the expressed genes in the islands (Pecrix et al., 2018). Many nodule-associated genes are present within SRIs, e.g., nodule-specific cysteine-rich peptides (NCRs), glycine-rich proteins (GRPs) or calmodulin-like proteins (CaML), MtENOD11, MtSYMREM1, MtSYMCRK, MtIRE, as well as genes encoding ERF transcription factors (Pecrix et al., 2018). The coregulation of

GENERAL INTRODUCTION
34
neighbouring gene sets are impacted by local chromatin structure determined by epigenetic regulators like DNA methylation status, small non-coding RNAs (ncRNAs) and histone marks (Pecrix et al., 2018). CHH (three-nucleotide cytosine context where H can be A, C or T) DMRs (differentially methylated regions) of the differentiation islands were found to be maximal at the gene promoter regions (Pecrix et al., 2018). The Mt5.0 genome browser integrates various data sources including omics, interactome and mutant collections.
Figure 7 Medicago Mt4.0 and Mt5.0.
(A) Increased number of chromosome-anchored sequences in Medicago Mt4.0 compared to Mt3.5. Red-coloured portion of the chromosomes represent BAC sequences used in Mt3.5, while the white regions on the chromosomes represent newly anchored sequences in Mt4.0. (Tang et al., 2014).
(B) Medicago Mt5.0 indicated with symbiosis-island. Symbiosis-related genomic islands represent physical clusters of genes strongly up- or downregulated when comparing whole nodules versus root systems (blue and red lines, respectively) or differentially regulated between laser-dissected nodule zones (specifically expressed in the nodule apex or differentiation zone: yellow and light blue, respectively). (Pecrix et al., 2018).
Gene expression analysis in the nodulation process
Transcriptome analyses have identified hundreds of plant and bacterial genes that are differentially expressed during nodule development and differentiation. Small-scale transcriptome studies have been carried out on different organs in multiple legume species under a variety of experimental conditions (Becker et al., 2004; Colebatch et al., 2004; El
A B

GENERAL INTRODUCTION
35
Yahyaoui et al., 2004; Mitra et al., 2004; Uchiumi et al., 2004). A more comprehensive Gene Expression Atlas (MtGEA; https://mtgea.noble.org/v3/) is used in transcriptomic analysis based on Affymetrix data of a wide range of biological conditions (Benedito et al., 2008). Organ-specific genes were largely uncovered in the nodules (1,354 genes) and seeds (3,228 genes) (Figure 8A), implying a peculiar function in the specialized organs (Benedito et al., 2008). RNAseq data was coupled with laser-capture-microdissected (LCM) approach (Figure 8B) to shed light on spatial analysis of plant and bacterial gene expression in different nodule zones (SYMbiMICS: https://iant.toulouse.inra.fr/symbimics/) (Roux et al., 2014). Differential and co-expressive analysis of the LCM RNAseq data highlighted 16 expression patterns illustrating successive waves of gene expressions involved from the nodule apical meristem to the N-fixation zone (Figure 8B) (Pecrix et al., 2018).
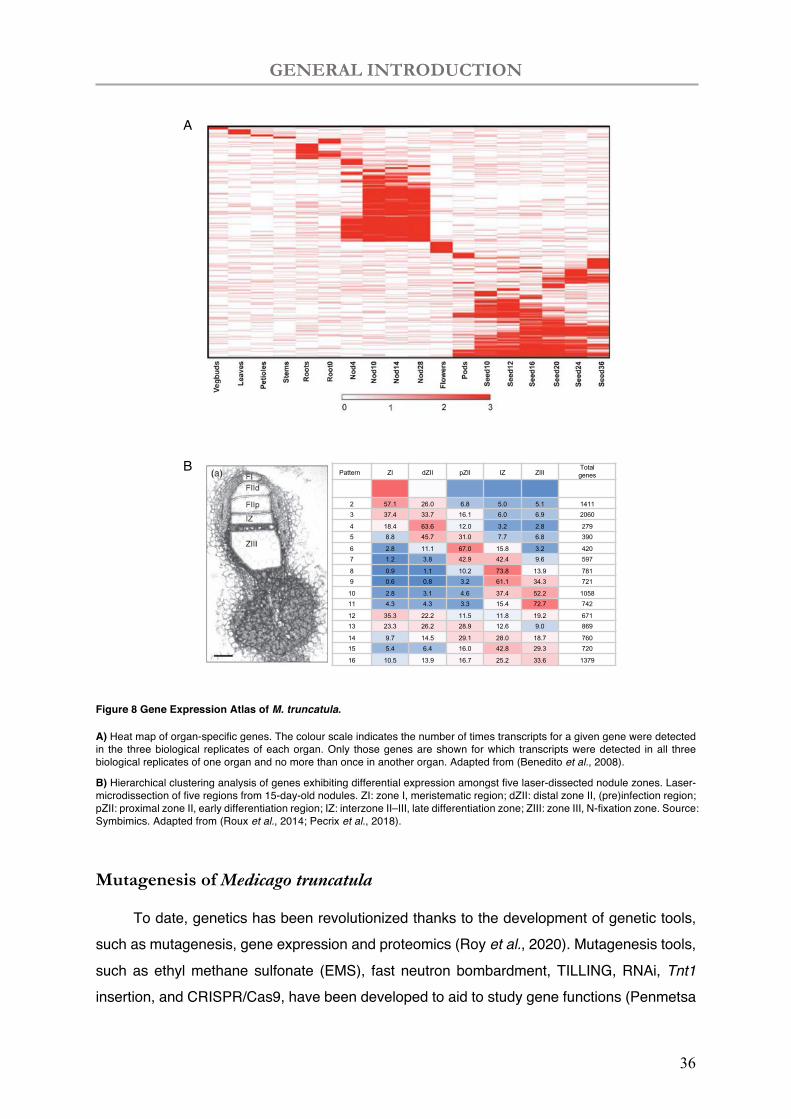
GENERAL INTRODUCTION
36
Figure 8 Gene Expression Atlas of M. truncatula.
A) Heat map of organ-specific genes. The colour scale indicates the number of times transcripts for a given gene were detected in the three biological replicates of each organ. Only those genes are shown for which transcripts were detected in all three biological replicates of one organ and no more than once in another organ. Adapted from (Benedito et al., 2008).
B) Hierarchical clustering analysis of genes exhibiting differential expression amongst five laser-dissected nodule zones. Laser-microdissection of five regions from 15-day-old nodules. ZI: zone I, meristematic region; dZII: distal zone II, (pre)infection region; pZII: proximal zone II, early differentiation region; IZ: interzone II–III, late differentiation zone; ZIII: zone III, N-fixation zone. Source: Symbimics. Adapted from (Roux et al., 2014; Pecrix et al., 2018).
Mutagenesis of Medicago truncatula
To date, genetics has been revolutionized thanks to the development of genetic tools, such as mutagenesis, gene expression and proteomics (Roy et al., 2020). Mutagenesis tools, such as ethyl methane sulfonate (EMS), fast neutron bombardment, TILLING, RNAi, Tnt1 insertion, and CRISPR/Cas9, have been developed to aid to study gene functions (Penmetsa
A
B Pattern ZI dZII pZII IZ ZIIITotalgenes
2 57.1 26.0 6.8 5.0 5.1 14113 37.4 33.7 16.1 6.0 6.9 2060
4 18.4 63.6 12.0 3.2 2.8 2795 8.8 45.7 31.0 7.7 6.8 390
6 2.8 11.1 67.0 15.8 3.2 4207 1.2 3.8 42.9 42.4 9.6 597
8 0.9 1.1 10.2 73.8 13.9 7819 0.6 0.8 3.2 61.1 34.3 721
10 2.8 3.1 4.6 37.4 52.2 105811 4.3 4.3 3.3 15.4 72.7 742
12 35.3 22.2 11.5 11.8 19.2 67113 23.3 26.2 28.9 12.6 9.0 869
14 9.7 14.5 29.1 28.0 18.7 76015 5.4 6.4 16.0 42.8 29.3 720
16 10.5 13.9 16.7 25.2 33.6 1379

GENERAL INTRODUCTION
37
& Cook, 2000; Tadege et al., 2008; Pislariu et al., 2012; Roy et al., 2020). In model species like M. truncatula, forward and reverse genetics have accelerated the understanding of the molecular and cellular processes of the symbiotic relationship. However, reverse genetic approaches are limited to reveal gene functions when gene family is complex or abundant, e.g., the NCRs. T-DNA insertion is largely used mostly in laboratory small scale. M. truncatula mutant population at the Samuel Roberts Noble Foundation is the largest legume mutant collection and were generated using the tobacco retrotransposon Tnt1 (Tadege et al., 2008; Pislariu et al., 2012).
III.2 Establishment of symbiosis
Under N limited conditions, leguminous plants and rhizobia elaborate to develop de novo N fixing root organ, called the root nodule. In this mutualistic relationship, the plant provides the bacteria partner with photosynthates and protective niche, and in return, the symbiotic bacteria fix atmosphere N to supply the plant with ammonium. The lifespan of nodules is a cascade of the initial signal exchange, infection process, development of the functional N fixing nodules and senescence before the final organ death, regulated by specific genes from both the plant and bacteria side with precise timing.
III.2.1 Recognition of legumes and rhizobia
Symbiotic recognition initiates the nodulation process
Establishment of symbiosis requires recognition of both partners with specific signalling. Under N starvation, plant emit metabolites (flavonoid) into rhizosphere that trigger the expression of bacterial nod genes into lipochitooligosaccharidic nodulation factors (Nod factors). The nod, nif and fix genes are located on large plasmids (Kondorosi et al., 1984). The function of nodulation (nod) genes has been extensively investigated by rhizobia mutants. The mutants impaired in the symbiotic interaction are found mostly Nod− (nodulation deficient) or Fix− (impaired nitrogen fixation) (Kondorosi et al., 1984). Four common nod genes (nodABC and D) are conserved in most Rhizobium species and played a role determining root hair curling or nodule initiation, and some other genes for host specificity of nodulation (hsnABC and D; also called nodFEG and H or nodL, nodP and nodQ) differs in among rhizobia strains to determine the host range (Kondorosi & Kondorosi, 1986). The nod D, a LysR-type protein induced by the plant root secreted (iso)flavonoids and betaines (Kobayashi et al., 2004), is constitutively expressed to regulate the transcription of all the nod gene units. Originally, dual

GENERAL INTRODUCTION
38
transcription control is found with the activator nodD (Rostas et al., 1986) binding to a 47bp cis-regulatory element (nod-box), and an occasional expressing repressor protecting a 21bp stretch downstream of the n6 nod-box (Rostas et al., 1986; Kondorosi et al., 1989).
Nod factors accumulated in cell walls of root epidermis, perceived by plant receptors (Nod factor perception (NFP), or NFR1/NFR5 in L. japonicus) containing the chitin binding and LysM domain receptor kinases (e.g., LYK3 encoding by HCL gene) (Madsen et al., 2003; Radutoiu et al., 2003; Arrighi et al., 2006; Broghammer et al., 2012; Moling et al., 2014). Upon infection of rhizobia, specific plant proteins were found exclusively expressed in nodules (Legocki & Verma, 1979; Legocki & Verma, 1980), thereafter called nodulins, including early nodulins and late nodulins. Plant receptors are crucial for selection/host range determination (Radutoiu et al., 2007), collaborating with bacterial Nod factors. These receptor genes are also identified with plant mutants with the Nod- and Fix- phenotype, as well as defects in Nod factor responses (Roy et al., 2020).
III.2.2 Bacterial infection is coupled with nodule organogenesis
To allow rhizobial infection, a tubular-like infection thread (IT) is developed by the plant to allow the entry of the entrapped bacterial microcolony present at the surface of the root hair. The IT grows into root cortex wherein bacteria are released in the plant cell via endocytosis (Figure 9) (Brewin, 2004; Oldroyd et al., 2011). During the infection, cell divisions begin in the inner and subsequently mid-cortex and nodule primordium is thereby formed (Schultze & Kondorosi, 1998). Bacterial infection and nodule organogenesis are coordinated steps so that multiple genes are required for IT formation and nodulation.
Symbiotic signalling pathway
The plant receptor kinase complexes thus initiate nodule organogenesis in a successive signalling pathway controlling nodulation (Figure 10) (Oldroyd et al., 2011; Oldroyd, 2013). An activation of calcium oscillation in form of periodic spiking is believed to be as the centre of the symbiotic pathway (Oldroyd & Downie, 2004; Oldroyd & Downie, 2006; Charpentier et al., 2016), with which ion flux changes lead to root hair curling entraps a bacterial microcolony and a subsequent expression of early nodulation genes (Charpentier & Oldroyd, 2013). Nod factors signals transmit to a receptor-like kinase (LjSYMRK), DMI2/MtSYM2 in Medicago, which associates with the 3-hydroxy-3-methylglutaryl-CoA reductase (HMGR) to the production of mevalonate, which may function as a secondary messenger to the nucleus (Kevei et al., 2007). The ion channels, a complex of DMI1 (Lj POLLUX) and CNGC15,
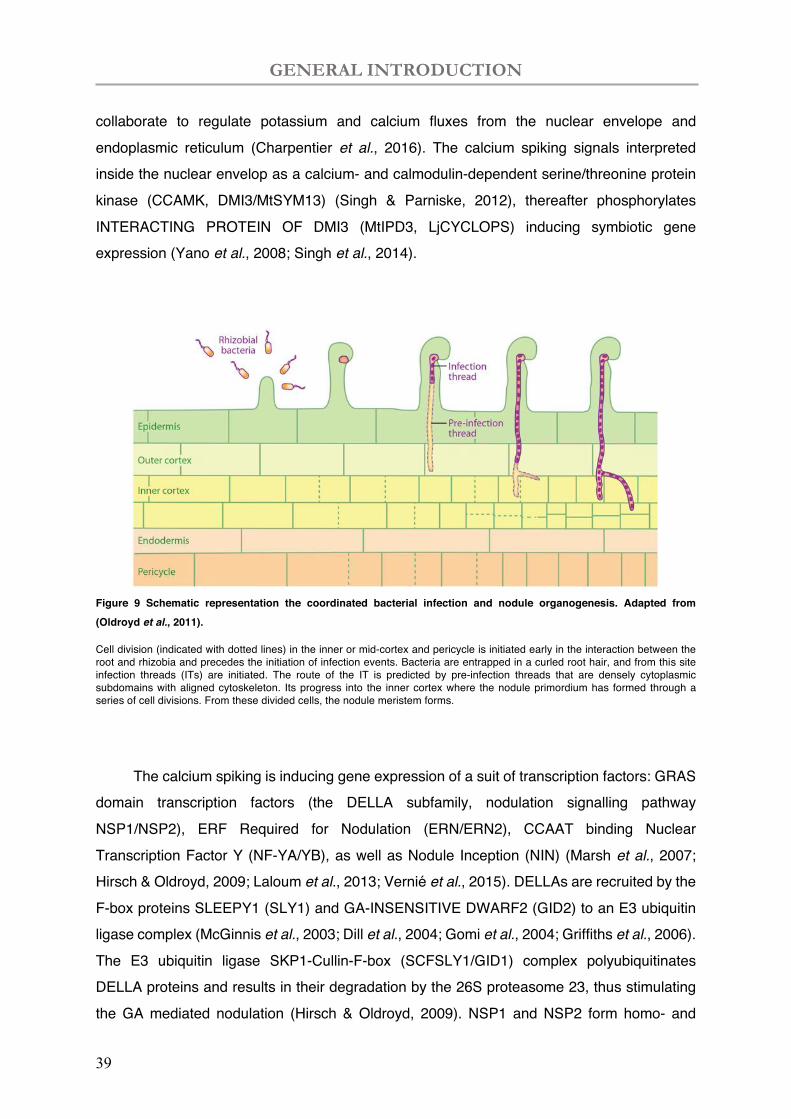
GENERAL INTRODUCTION
39
collaborate to regulate potassium and calcium fluxes from the nuclear envelope and endoplasmic reticulum (Charpentier et al., 2016). The calcium spiking signals interpreted inside the nuclear envelop as a calcium- and calmodulin-dependent serine/threonine protein kinase (CCAMK, DMI3/MtSYM13) (Singh & Parniske, 2012), thereafter phosphorylates INTERACTING PROTEIN OF DMI3 (MtIPD3, LjCYCLOPS) inducing symbiotic gene expression (Yano et al., 2008; Singh et al., 2014).
Figure 9 Schematic representation the coordinated bacterial infection and nodule organogenesis. Adapted from (Oldroyd et al., 2011).
Cell division (indicated with dotted lines) in the inner or mid-cortex and pericycle is initiated early in the interaction between the root and rhizobia and precedes the initiation of infection events. Bacteria are entrapped in a curled root hair, and from this site infection threads (ITs) are initiated. The route of the IT is predicted by pre-infection threads that are densely cytoplasmic subdomains with aligned cytoskeleton. Its progress into the inner cortex where the nodule primordium has formed through a series of cell divisions. From these divided cells, the nodule meristem forms.
The calcium spiking is inducing gene expression of a suit of transcription factors: GRAS domain transcription factors (the DELLA subfamily, nodulation signalling pathway NSP1/NSP2), ERF Required for Nodulation (ERN/ERN2), CCAAT binding Nuclear Transcription Factor Y (NF-YA/YB), as well as Nodule Inception (NIN) (Marsh et al., 2007; Hirsch & Oldroyd, 2009; Laloum et al., 2013; Vernié et al., 2015). DELLAs are recruited by the F-box proteins SLEEPY1 (SLY1) and GA-INSENSITIVE DWARF2 (GID2) to an E3 ubiquitin ligase complex (McGinnis et al., 2003; Dill et al., 2004; Gomi et al., 2004; Griffiths et al., 2006). The E3 ubiquitin ligase SKP1-Cullin-F-box (SCFSLY1/GID1) complex polyubiquitinates DELLA proteins and results in their degradation by the 26S proteasome 23, thus stimulating the GA mediated nodulation (Hirsch & Oldroyd, 2009). NSP1 and NSP2 form homo- and

GENERAL INTRODUCTION
40
heteropolymers interacting with promoters of early nodulins like ENOD11 (Journet et al., 2001; Hirsch et al., 2009; Cerri et al., 2012). NSP1/NSP2 activates the transcription of ENOD11 as well as the transcription factor ERN1 (Ethylene Response Factor Required for Nodulation1). ERN1 directly regulates ENOD11 (Cerri et al., 2012) via specific binding to a Nod factor responsive cis element, NF-box (Andriankaja et al., 2007), enabling the subsequent bacterial infection process.
Figure 10 Plant symbiotic intracellular signalling pathways. Adapted from (Zipfel & Oldroyd, 2017).
The Nod-LCOs are recognized by a receptor complex of the LysM receptor kinases NFR1 and NFR5 and the LRR-containing receptor kinase SYMRK (known as DMI2 in Medicago). Pathway activation leads to a burst of reactive oxygen species (ROS) and an influx of calcium across the plasma membrane. The 3-hydroxy-3-methylglutaryl-CoA reductase (HMGR) that associates with SYMRK is involved in the production of mevalonate, which may function as a secondary messenger to activate calcium oscillations in the nucleus. Several ion channels (DMI1, CNGC15 and MCA8) are located at the nuclear membrane, regulating counterflows of potassium and calcium. Nuclear calcium oscillations activate CCAMK, which phosphorylates CYCLOPS to promote the induction of symbiosis gene expression. CYCLOPS may form part of a large complex that contains a number of GRAS-domain-containing transcription factors (NSP1, NSP2 and DELLA).

GENERAL INTRODUCTION
41
The transcription factor NIN is a master gene of nodule organogenesis
During the infection, cells in the inner and subsequently mid-cortex begin dividing and nodule primordium is thereby formed (Schultze & Kondorosi, 1998). The TF NIN was firstly identified in L. japonicus (Schauser et al., 1999), and subsequently evidenced by multiple groups as a master regulatory gene of nodule organogenesis through cytokinin promotion (Marsh et al., 2007; Vernié et al., 2015). NIN has been shown to regulate many nodulation related genes such as Rhizobium-induced peroxidase 1 (RIP1) (Cook et al., 1995), ENOD11 (Cerri et al., 2012), Nodulation Pectate Lyase (NPL) (Xie et al., 2012), NF-YA1/2 (Soyano et al., 2013) and a number of CLE encoded peptides involved in autoregulation of nodulation (Nakajima et al.) (Soyano et al., 2014). NIN is able to inhibit nodulation in response to nitrate, together with a family of NIN-like proteins (NLPs) through interaction of their carboxy-terminal PB1 domains (Lin et al., 2018). NLPs are found to be nitrate responsive (Lin et al., 2018; Nishida et al., 2018). Furthermore, different cis-regulatory sequences in the NIN promoter are revealed to regulate bacterial infection and nodule organogenesis (Yoro et al., 2014). In addition, a remote cis-regulatory region 5kb upstream of the start codon was identified for NIN expression initiate nodule primordium formation, containing putative cytokinin response elements (Liu et al., 2019). Ectopic expression of NIN and the NF-YA1 is able to trigger cortical cell divisions (Soyano et al., 2013).
III.2.1 Bacteroid differentiation
Nodule are developed into determinate and indeterminate nodules in
different leguminous plants
When infection thread reaches the primordia, bacteria are released. Bacteria reside in plant cells as an organelle-like vesicles, with a half plant and half bacteria derived double membrane (peribacteroid membrane, PBM), collectively termed symbiosome. In order to perform nitrogen fixation, both plant and bacterial partners undergo differentiation, therein bacteroid differentiation are classified into two types as non-terminal differentiation and terminal differentiation.
1) Non-terminal bacteroid differentiation: In most legume clades like the ones comprising Lotus japonicus or common bean (P. vulgaris), bacteroids remain similar in size and shape to the free-living rhizobia. They also retain full capacity to resume growth when they are extracted from nodules (Mergaert et al., 2006).

GENERAL INTRODUCTION
42
2) Terminal bacteroid differentiation: in galegoid (temperate) legumes like M. truncatula, the endosymbiotic bacteria and the host cells undergo the terminal differentiation.
In consequences, nodules are accordingly developmentally classified in two types: 1) Determinate nodules are round in shape without a persistent meristem, in the model
plant L. japonicus and some (Ruvkun et al.)tropical legumes such as Phaseolus, Glycine, and Vigna species.
2) Indeterminate nodules are in cylindric shape caused by continuously generated cells from an apical meristem which was established from nodule primordia, mostly present in temperate legumes, e.g., the model plant M. truncatula. The meristem cells continue to grow into differentiation processes, thus lead to a typical zonation in mature indeterminate nodules.
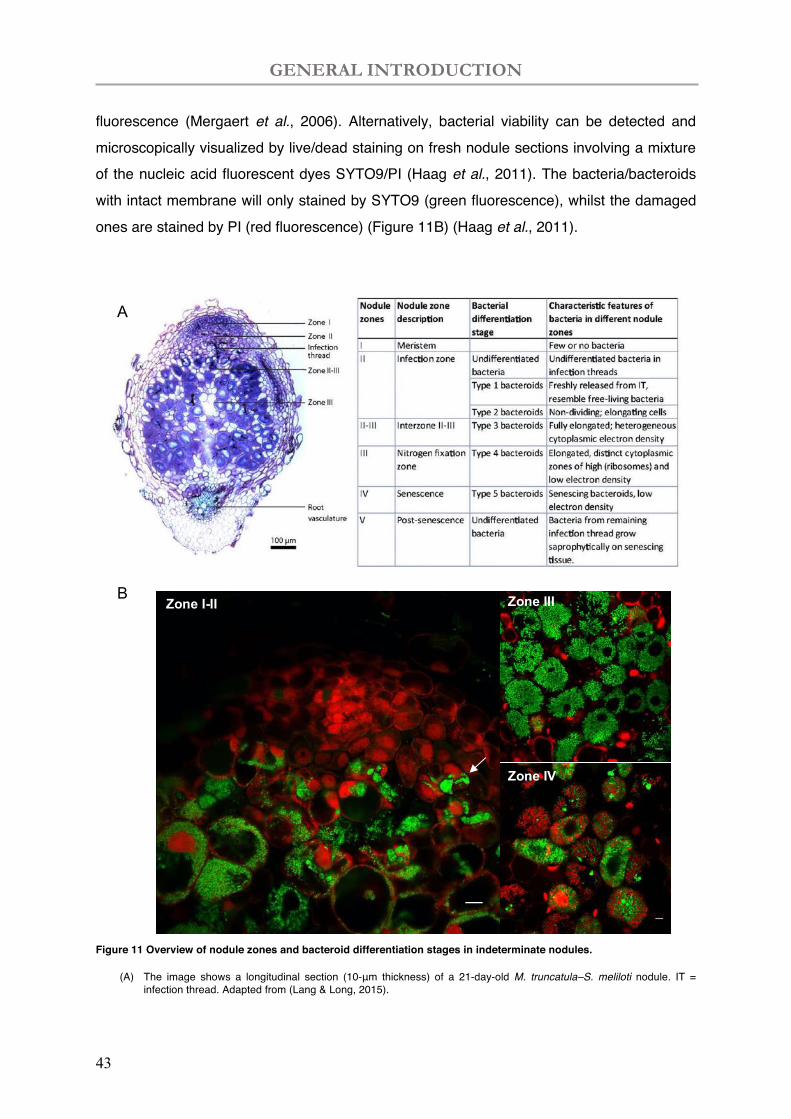
Zonation in indeterminate nodules are as shown in Figure 11A and Figure 11B: meristem (Zone I); infection zone (zone II) where bacteria are released and infect plant cells; a large nitrogen fixation zone (zone III) where the main nitrogen fixation function of nodules happen; a conically enlargement of senescence zone (zone IV) along with time or induced by stresses. At last, a Zone V is also mentioned when cytoplasm of nodule cells is fully degraded, and saprophytic bacteria colonized these senescent nodules. While the development of zonation is gradual without a sharp border, an interzone (zone IZ) is defined between zone II-III corresponding to the bacteria terminal differentiation into bacteroid and characterized by amyloplast accumulation (Vasse et al., 1990). We can also observe the interzone between zone III-IV where the nodule senescence related genes are induced (Perez Guerra et al., 2010; Pierre et al., 2014).
Bacteria grow and divide in the symbiosome, presenting different characteristics in the indeterminate nodule zones (Lang & Long, 2015). Seldom bacteria can be found in nodule meristem in which infection thread established (zone I; Figure 11B) (Lang & Long, 2015). In nodule zone II, three types of bacteria can be found: small and rod-shaped free-living bacteria, freshly released free bacteria and elongating bacteria (zone II; Figure 11B) (Lang & Long, 2015). The bacterial and plant cell elongation mainly occur in the interzone II-III, afterwards bacteroids are fully elongated or in Y-shape occupying the cytosol of host cells in zone III with an organized manner (zone III; Figure 11B) (Kondorosi et al., 2013). The infected cells of nitrogen-fixing nodules are interspersed with uninfected cells that are involved in transport of sugars and nitrogenous compounds in connection with the vascular bundles (zone III; Figure 11B) (Peiter & Schubert, 2003; Godiard et al., 2011). The respiratory activity of living bacteria/bacteroids can be detected by 5-cyano-2,3-di-4-tolyl tetrazolium chloride (CTC)
GENERAL INTRODUCTION
43
fluorescence (Mergaert et al., 2006). Alternatively, bacterial viability can be detected and microscopically visualized by live/dead staining on fresh nodule sections involving a mixture of the nucleic acid fluorescent dyes SYTO9/PI (Haag et al., 2011). The bacteria/bacteroids with intact membrane will only stained by SYTO9 (green fluorescence), whilst the damaged ones are stained by PI (red fluorescence) (Figure 11B) (Haag et al., 2011).
Figure 11 Overview of nodule zones and bacteroid differentiation stages in indeterminate nodules.
(A) The image shows a longitudinal section (10-μm thickness) of a 21-day-old M. truncatula–S. meliloti nodule. IT = infection thread. Adapted from (Lang & Long, 2015).
B Zone I-II Zone III
Zone IV
A
GENERAL INTRODUCTION
44
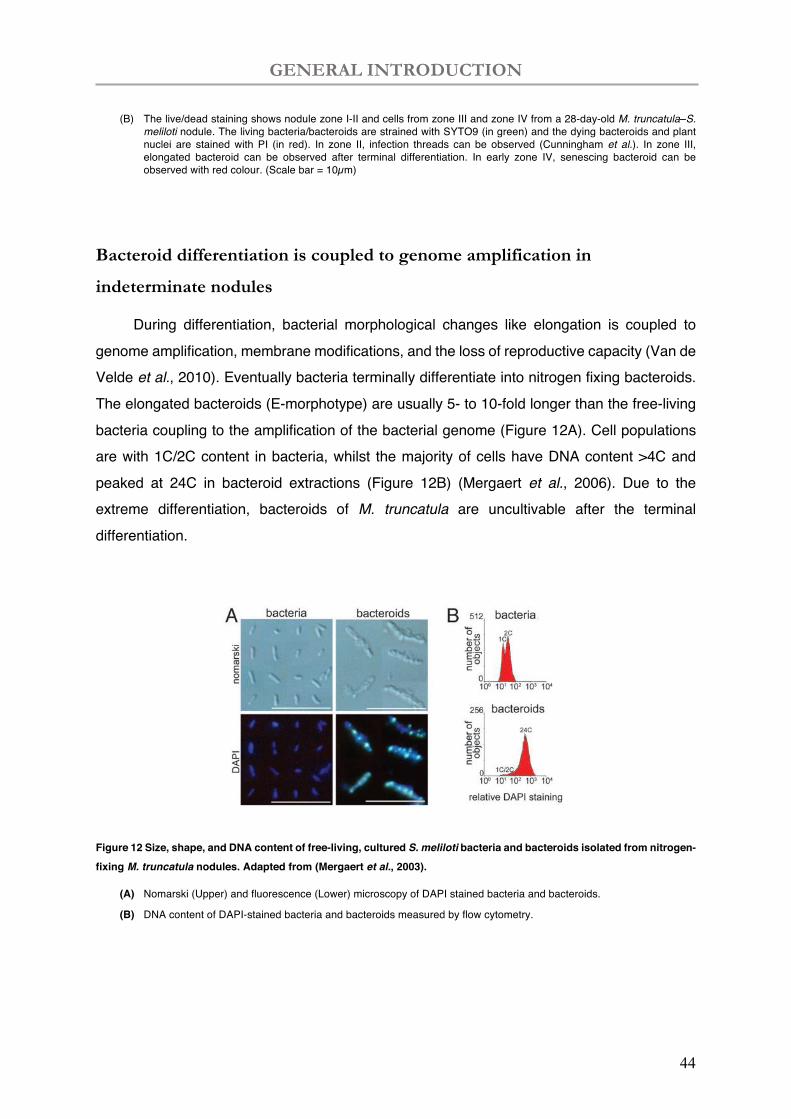
(B) The live/dead staining shows nodule zone I-II and cells from zone III and zone IV from a 28-day-old M. truncatula–S. meliloti nodule. The living bacteria/bacteroids are strained with SYTO9 (in green) and the dying bacteroids and plant nuclei are stained with PI (in red). In zone II, infection threads can be observed (Cunningham et al.). In zone III, elongated bacteroid can be observed after terminal differentiation. In early zone IV, senescing bacteroid can be observed with red colour. (Scale bar = 10µm)
Bacteroid differentiation is coupled to genome amplification in
indeterminate nodules
During differentiation, bacterial morphological changes like elongation is coupled to genome amplification, membrane modifications, and the loss of reproductive capacity (Van de Velde et al., 2010). Eventually bacteria terminally differentiate into nitrogen fixing bacteroids. The elongated bacteroids (E-morphotype) are usually 5- to 10-fold longer than the free-living bacteria coupling to the amplification of the bacterial genome (Figure 12A). Cell populations are with 1C/2C content in bacteria, whilst the majority of cells have DNA content >4C and peaked at 24C in bacteroid extractions (Figure 12B) (Mergaert et al., 2006). Due to the extreme differentiation, bacteroids of M. truncatula are uncultivable after the terminal differentiation.
Figure 12 Size, shape, and DNA content of free-living, cultured S. meliloti bacteria and bacteroids isolated from nitrogen-fixing M. truncatula nodules. Adapted from (Mergaert et al., 2003).
(A) Nomarski (Upper) and fluorescence (Lower) microscopy of DAPI stained bacteria and bacteroids.
(B) DNA content of DAPI-stained bacteria and bacteroids measured by flow cytometry.

GENERAL INTRODUCTION
45
Symbiotic plant cells undergo endoreduplication
Coordinately, the symbiotic plant cells are elongated and coupled with endoreduplication, eventually differentiate into polyploid cells. In Figure 13A, bacteria are released in plant cells in postmitotic stage with 4C DNA content (C being the haploid DNA content) (Kondorosi et al., 2013). Upon activation of NCRs and GRPs, the symbiotic cells further differentiate into 8C, 16C and 32C couple with the endoreduplication (genome replication without mitosis and cytokinesis) and cell enlargement (Figure 13A) (Mergaert et al., 2006; Kondorosi et al., 2013). Furthermore, in coordination with the host cells, bacterial infection (zone II) also leads to the enlargement of symbiotic cells and endoreduplication cycles resulted in high ploidy levels of 32C and 64C (Figure 13) (Cebolla et al., 1999; Vinardell et al., 2003; Kondorosi et al., 2013), eventually forming the fully elongated and huge polyploid cells. The symbiotic cells with high ploidy levels are therefore able to undergo extreme cell growth, host thousands of bacteroids and sustain the energy-demanding N fixation (Mergaert et al., 2006). In Figure 13B, a flow cytometry analysis shows proportions of cell ploidy levels at different stages of nodule development (Vinardell et al., 2003).
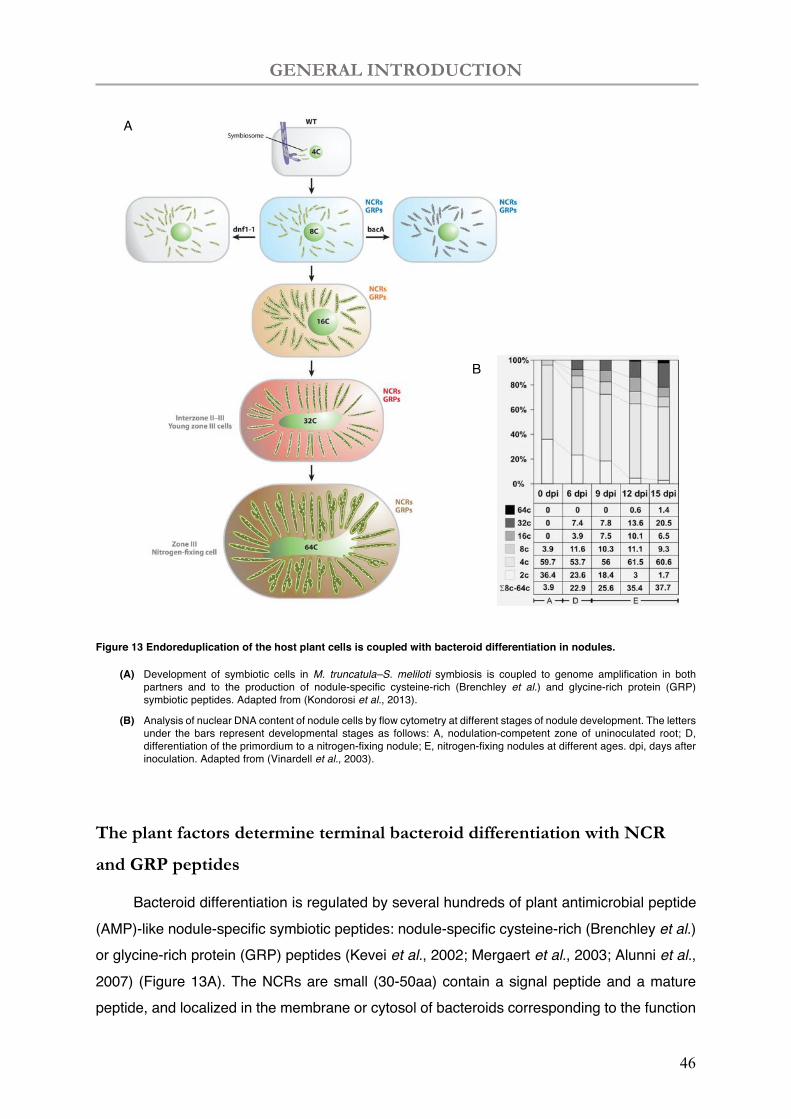
GENERAL INTRODUCTION
46
Figure 13 Endoreduplication of the host plant cells is coupled with bacteroid differentiation in nodules.
(A) Development of symbiotic cells in M. truncatula–S. meliloti symbiosis is coupled to genome amplification in both partners and to the production of nodule-specific cysteine-rich (Brenchley et al.) and glycine-rich protein (GRP) symbiotic peptides. Adapted from (Kondorosi et al., 2013).
(B) Analysis of nuclear DNA content of nodule cells by flow cytometry at different stages of nodule development. The letters under the bars represent developmental stages as follows: A, nodulation-competent zone of uninoculated root; D, differentiation of the primordium to a nitrogen-fixing nodule; E, nitrogen-fixing nodules at different ages. dpi, days after inoculation. Adapted from (Vinardell et al., 2003).
The plant factors determine terminal bacteroid differentiation with NCR
and GRP peptides
Bacteroid differentiation is regulated by several hundreds of plant antimicrobial peptide (AMP)-like nodule-specific symbiotic peptides: nodule-specific cysteine-rich (Brenchley et al.) or glycine-rich protein (GRP) peptides (Kevei et al., 2002; Mergaert et al., 2003; Alunni et al., 2007) (Figure 13A). The NCRs are small (30-50aa) contain a signal peptide and a mature peptide, and localized in the membrane or cytosol of bacteroids corresponding to the function
A
B

GENERAL INTRODUCTION
47
in bacteroid differentiation (Mergaert et al., 2003; Van de Velde et al., 2010). The corresponding gene expressions are only in infected cells and localized in zones with their presumed function produced in zone II, interzone II–III, and zone III (Mergaert et al., 2003; Van de Velde et al., 2010; Kondorosi et al., 2013).
In a dnf1 mutant of M. truncatula, bacteria can be release in plant cell, but the subsequent differentiation is blocked (Wang et al., 2010). The DNF1 protein is a nodule-specific subunit of the signal peptidase complex (SPC) (Wang et al., 2010). SPC is an early component in the protein secretory pathway in the endoplasmic reticulum (ER) and removes the signal peptide of secretory proteins during their translocation into the ER lumen (Van de Velde et al., 2010; Wang et al., 2010). The bacterial terminal differentiation requires NCR trafficking into bacteroid, but the bacteroid differentiation is obstructed in dnf1 mutant when the processing of NCRs blocked at ER (Van de Velde et al., 2006). One of the cationic peptides, NCR247, is expressed in the older zone II and in the interzone II-III (Farkas et al., 2014). NCR247 can penetrate bacteria membranes and form complex with bacterial proteins to affect bacterial cell division, translation, and protein folding (Farkas et al., 2014). In contrast, ectopic expression of NCRs in L. japonicus, for which bacteroids are normally U-morphotype (similar morphology than free-living bacteria), induce bacteroid morphological changes reminiscent of terminally differentiation with elongated bacteroids (Van de Velde et al., 2010).
GRP carry a secretory signal peptide and a glycine rich mature peptide of approximately 100 amino acids (Sachetto-Martins et al., 2000; Kevei et al., 2002; Mangeon et al., 2010; Kondorosi et al., 2013). Most other GRPs are developmentally regulated by biotic and abiotic factors, e.g., pathogens (Molina et al., 1997), osmotic stress (Kim et al., 2007), cold shock (Carpenter et al., 1994; Kim et al., 2007) and wounding (Showalter et al., 1991). On the contrary, the nodule specific GRPs requires rhizobia infection but are not responding to abiotic factors (Kevei et al., 2002). MsnodGRP1 and MsnodGRP2 transcripts were found abundant in interzone II-III, suggesting a role in nodule differentiation (Kevei et al., 2002).
Bacterial partnership in bacteroid differentiation: BacA and CtrlA
On the bacterial side, BacA, a membrane protein from the ATP-binding cassette (ABC) transporter superfamily, plays an important role in bacteroids differentiation (Maunoury et al., 2010; Haag et al., 2011; Kondorosi et al., 2013). In the S. meliloti mutant bacA, bacteria succeed to release in host cells but is deficient in bacteroid differentiation, resulting in immediate nodule senescence (Maunoury et al., 2010). BacA is suggested to protect bacteria against the antimicrobial actions of NCRs which induce membrane permeabilization and

GENERAL INTRODUCTION
48
bacterial killing in vitro. In contrast, BacA is dispensable for bacterial survival in an M. truncatula dnf1 mutant (Haag et al., 2011). Therefore, BacA is critical to long-term host infection via allowing the bacterial persistence in symbiotic cells rather than rapid bacterial death (Haag et al., 2011).
As a transcriptional regulator, CtrA is considered as the master regulator of cell cycles in some α-proteobacteria such as Caulobacter crescentus as a model system (Laub et al., 2000; Laub et al., 2002; Brilli et al., 2010). In S. meliloti, CtrA depletion leads to bacterial cell elongation, increase of DNA content and abolished cell division, which are similar to bacteroid differentiation, but the membrane integrity was maintained (Pini et al., 2015). Through microarray-based gene expression analysis, CtrA were found to regulate the transcription of at least 126 genes in S. meliloti, elevating genes such as cell cycle regulators (i.e. ctrA, divJ, divK, sciP), motility genes (i.e. mcp, che, flg, flaAB, fli), cell envelope components and several hypothetical proteins (Pini et al., 2015). CtrA was also shown as a repressor for cell division regulators minC and minD. Therefore, CtrA down-regulation during bacteroid differentiation increases minCD expression, subsequently blocking cell division by inhibiting the FtsZ polymerization and Z-sring formation (Cheng et al., 2007; Pini et al., 2015). CtrA can bind to two consensus sites (TTAA-N7-TTAAC) and (TTAACCAT) to regulate cell cycle genes (Laub et al., 2002). In S. meliloti, these binding sites are detected in the promoter regions of mcpZ, pilA1, flaA and divJ via a large-scale Chromatin Immunoprecipitation assay (Johnson et al., 2007; Pini et al., 2015).
III.2.2 Nitrogen fixation in bacteroids
The bacteroids are able to fix atmosphere nitrogen into ammonia in microaerobic conditions. This BNF process allows for the production of NH3/NH4+ that can be utilized by plants (Stein and Klotz, 2016). Similar as in the Harbor-Bosch process, BNF requires high energy input to break the triple band in N2. Instead of using fossil fuels, the redox reaction of BNF is associated with electron transfer processes with ATP hydrolysis-16 moles of ATP to produce 2 moles of ammonia. The equation of BNF process is as in below:
N2 + 8H++ 8e−+ 16Mg-ATP → 2NH3 + H2 + 16Mg-ADP + 16Pi

GENERAL INTRODUCTION
49
Nitrogen fixation is catalysed by the nitrogenase enzyme complexes
This reaction is catalysed by the nitrogenase enzyme complexes (Oldroyd & Dixon, 2014), encoding by numerous nif genes, including regulators. The nitrogenase is stringently regulated at transcriptional level (Figure 14A), responding to N resources and oxygen level. NifA is the master regulator of nif genes, co-dependent with the RNA polymerase sigma factor, σ54 (Dixon & Kahn, 2004). The fixLJ genes are positive regulators of the symbiotic expression of the N fixation gens in a cascade: the fixLJ genes activate nifA, which in turn activates nifHDK and fixABCX (David et al., 1988). The nitrogenase complex consists of a reductase component (Fe protein) and a catalyst component (MoFe protein) (Figure 14B) (Burgess & Lowe, 1996; Seefeldt et al., 2009; Oldroyd & Dixon, 2014). The Fe protein act as electron donors reduced by ferredoxin and flavodoxin, which is dependent on MgATP hydrolysis (Dixon & Kahn, 2004). Three metalloclusters, [4Fe-4S], P cluster and FeMo-co [Figure 14B(c)], provide an electron transfer conduit and the catalytic site for N reduction, respectively (Oldroyd & Dixon, 2014).
However, O2 can irreversibly damage the Fe protein and the MoFe protein, therefore most of the BNF process must be protected from O2 in various physiological strategies, including anaerobic growth conditions, consumption of excess O2 by respiration, O2 diffusion barriers, or compartmentation of the enzyme spatially or temporally (Dixon & Kahn, 2004). In case of legumes nodules, gas permeability is restricted by the nodule cortex as the barrier for O2 diffusion. In addition, an O2-binding protein, leghaemoglobin, which also gives the nodules a pink colour, is present in the cytoplasm of infected nodule cells to facilitate the bacteroid respiration under microoxic conditions. These conditions within nodules are achieved by restricted gaseous diffusion into nodules and high rates of respiration facilitated by oxygen-binding and oxygen-transporting leghaemoglobin in the cytoplasm of plant cells (Ott et al., 2005). Free-oxygen concentrations in nodule cells are as low as 10–8 M in the fixation zone, permitting nitrogenase to function (Tjepkema & Yocum, 1974). Thus, oxygen is maintained in nodules at a level that support respiration but sufficiently low to avoid nitrogenase inactivation (Kuzma et al. 1993). Finally, the released ammonia must be assimilated immediately in root nodules to avoid toxicity.

GENERAL INTRODUCTION
50
Figure 14 Nitrogenase regulation, gene and structure.
(A) Regulatory cascades controlling nif transcription in S. meliloti. Adapted from (Dixon & Kahn, 2004).
(B) Nitrogenase structure and genes required for its biosynthesis. Adapted from (Oldroyd & Dixon, 2014).
(a) The nitrogen fixation (nif) gene cluster from Klebsiella oxytoca. The nitrogenase structural genes (nifH, nifD and nifK) are coloured according to the crystal structure shown in (b). Remaining genes are colour-coded according to their functions: dark red, Fe protein maturation (nifM); light blue, FeMoco biosynthesis (nifY, nifE, nifN, nifX, nifV, nifB, nifQ); yellow, Fe–S cluster biosynthesis (nifU, nifS, nifZ); green, electron transport (nifJ, nifF); orange, transcriptional regulation (nifL, nifA); grey, unknown function (nifT, nifW).
(b) Structure of the nitrogenase enzyme complex showing the MoFe and Fe protein components, with the three metalloclusters revealed on the left-half of the complex (abbreviated as F, [4Fe4S]; P, P cluster and M, FeMo-co, respectively). ATP hydrolysis by the Fe protein, the route of electron transfer to the catalytic site and the enzyme reaction are also illustrated on the left half of the structure.
(c) Structures of the three metalloclusters in nitrogenase. Genes required for the biosynthesis of each cluster are illustrated below.
IV. Senescence
IV.1 Senescence as a common process
Terminology of senescence
The word “senescence” is originated from the Latin senescere, i.e., to grow old, reflecting its association with senility (Thomas, 2013). For semelparous species, the ending timepoint culminates with mating under monocarpic senescence, whereas in iteroparous species, senescence splits in the entities (e.g. whole plant) and the parts (e.g. organs), so that reproduction of parts takes place repeatedly (Thomas, 2013). There is a long-term semantic
A B
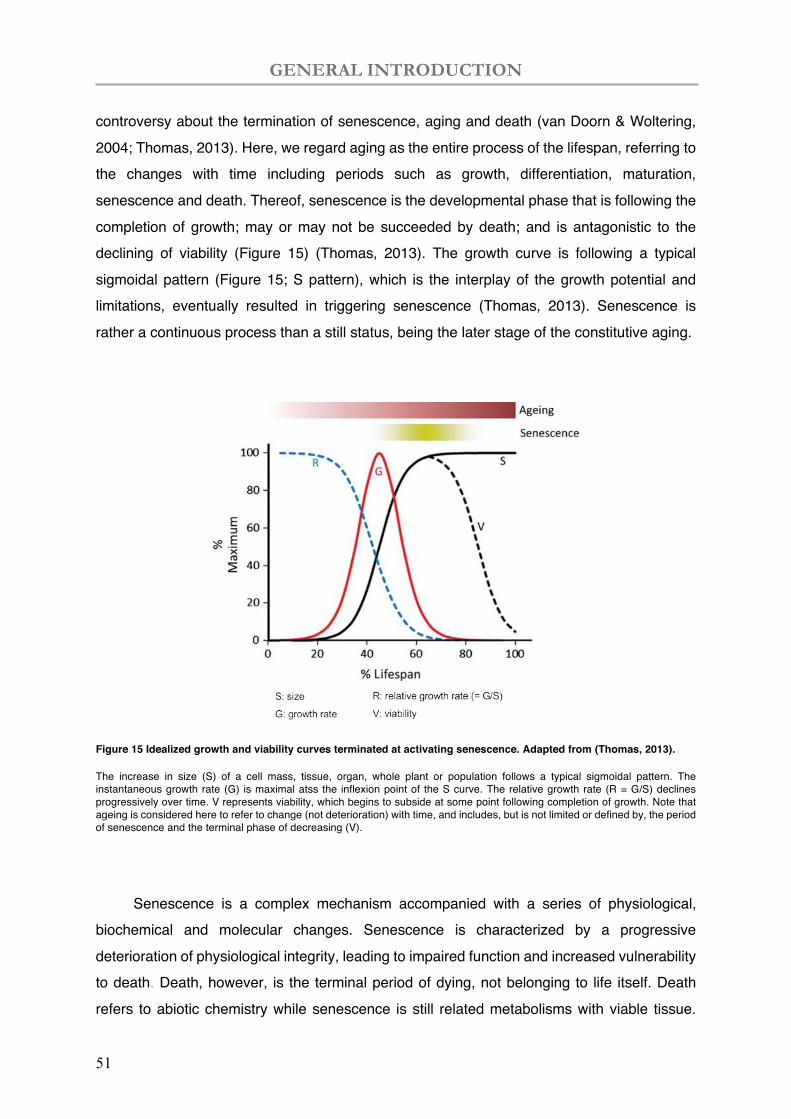
GENERAL INTRODUCTION
51
controversy about the termination of senescence, aging and death (van Doorn & Woltering, 2004; Thomas, 2013). Here, we regard aging as the entire process of the lifespan, referring to the changes with time including periods such as growth, differentiation, maturation, senescence and death. Thereof, senescence is the developmental phase that is following the completion of growth; may or may not be succeeded by death; and is antagonistic to the declining of viability (Figure 15) (Thomas, 2013). The growth curve is following a typical sigmoidal pattern (Figure 15; S pattern), which is the interplay of the growth potential and limitations, eventually resulted in triggering senescence (Thomas, 2013). Senescence is rather a continuous process than a still status, being the later stage of the constitutive aging.
Figure 15 Idealized growth and viability curves terminated at activating senescence. Adapted from (Thomas, 2013).
The increase in size (S) of a cell mass, tissue, organ, whole plant or population follows a typical sigmoidal pattern. The instantaneous growth rate (G) is maximal atss the inflexion point of the S curve. The relative growth rate (R = G/S) declines progressively over time. V represents viability, which begins to subside at some point following completion of growth. Note that ageing is considered here to refer to change (not deterioration) with time, and includes, but is not limited or defined by, the period of senescence and the terminal phase of decreasing (V).
Senescence is a complex mechanism accompanied with a series of physiological, biochemical and molecular changes. Senescence is characterized by a progressive deterioration of physiological integrity, leading to impaired function and increased vulnerability to death. Death, however, is the terminal period of dying, not belonging to life itself. Death refers to abiotic chemistry while senescence is still related metabolisms with viable tissue.

GENERAL INTRODUCTION
52
Recently, senescence is under scientific scrutiny thanks to modern advances in biotechnical approaches. However, the hallmarks of senescence in animals (e.g., somatic mutation, telomere attrition and the costs of repair and maintenance) seem not to be significant in plants, but other factors (e.g. epigenetic alterations, loss of proteostasis, deregulated nutrient sensing, mitochondrial dysfunction, cellular senescence, and altered intercellular communication) still matter (Pladys & Vance, 1993; Lopez-Otin et al., 2013; Thomas, 2013).
Developmental and induced senescence
Senescence and disease may seem to be different process in a distant view. Senescence can be classified as developmental senescence and induced senescence based on causality. Generally, developmental senescence is believed to be cell programmed. Comparatively, induced senescence refers to environmental changes. However, they would share a lot of similarities, if we regard time as an abiotic factor in developmental senescence, thereby an accelerated process of senescence would be seen in induced senescence. Multiple physiological processes involved senescence-like and defence-like mechanisms. In this context, defence reactions such as programmed cell death occurring during incompatible interactions share common symptoms with senescence (Lim et al., 2007). The common elements and the specificity of each reactions still need to be elucidated as there are multiple processes depending on the plant species and the environmental constraints. Nevertheless, both senescence and defence reactions involve genetic reprograming which allow the realization of the different physiological responses. Programmed senescence precedes according to a timetable affected by developmental and environmental factors and mediated by genetic programs (Jansson & Thomas, 2008).
IV.2 Regulation of senescence
IV.2.1 Regulation of plant senescence
Plant senescence is under molecular control such as phytohormone signalling, redox transduction, kinases and transcriptional factors (Lim et al., 2007; Thomas et al., 2009; Kirkwood & Melov, 2011; Thomas, 2013). Plant hormones including salicylic acid, abscisic acid, jasmonic acid and ethylene advance leaf senescence, whereas others like cytokinin, gibberellins, and auxins delay this process (Sarwat et al., 2013). Depending on the different plant species and the multiple senescence processes linked to induced senescence or organ specificity, the implication of the different hormones could be different. A striking example is
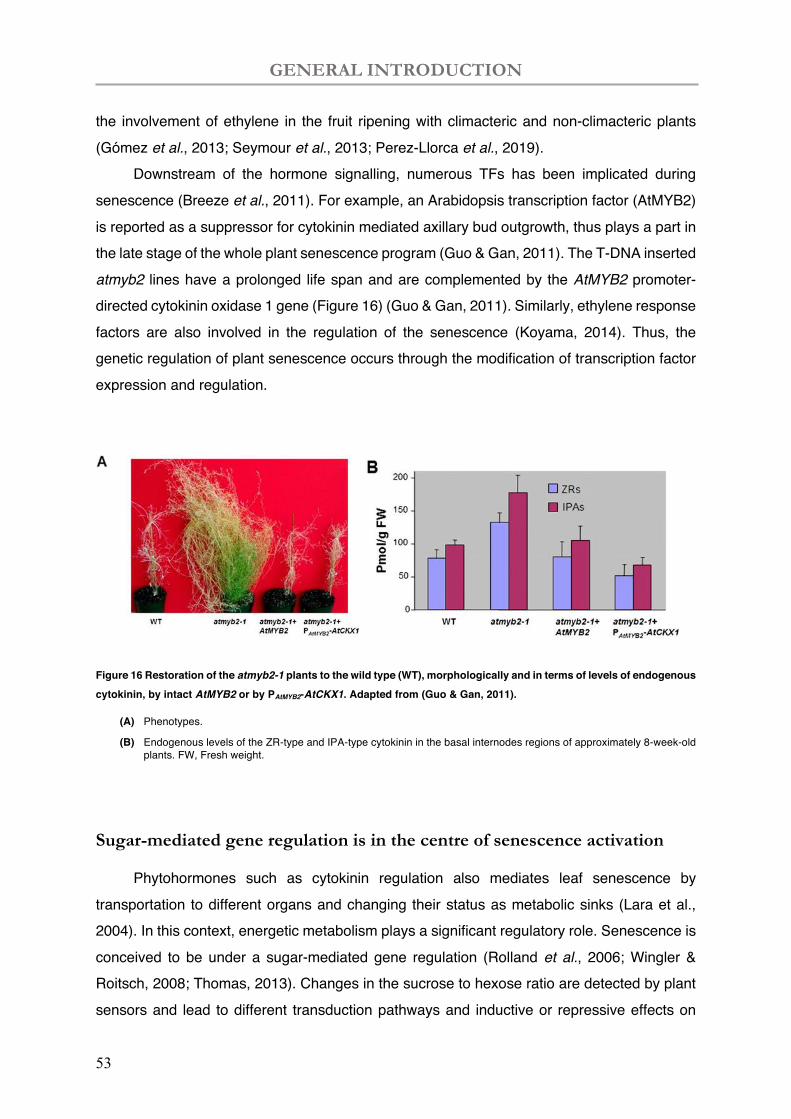
GENERAL INTRODUCTION
53
the involvement of ethylene in the fruit ripening with climacteric and non-climacteric plants (Gómez et al., 2013; Seymour et al., 2013; Perez-Llorca et al., 2019).
Downstream of the hormone signalling, numerous TFs has been implicated during senescence (Breeze et al., 2011). For example, an Arabidopsis transcription factor (AtMYB2) is reported as a suppressor for cytokinin mediated axillary bud outgrowth, thus plays a part in the late stage of the whole plant senescence program (Guo & Gan, 2011). The T-DNA inserted atmyb2 lines have a prolonged life span and are complemented by the AtMYB2 promoter-directed cytokinin oxidase 1 gene (Figure 16) (Guo & Gan, 2011). Similarly, ethylene response factors are also involved in the regulation of the senescence (Koyama, 2014). Thus, the genetic regulation of plant senescence occurs through the modification of transcription factor expression and regulation.
Figure 16 Restoration of the atmyb2-1 plants to the wild type (WT), morphologically and in terms of levels of endogenous cytokinin, by intact AtMYB2 or by PAtMYB2-AtCKX1. Adapted from (Guo & Gan, 2011).
(A) Phenotypes.
(B) Endogenous levels of the ZR-type and IPA-type cytokinin in the basal internodes regions of approximately 8-week-old plants. FW, Fresh weight.
Sugar-mediated gene regulation is in the centre of senescence activation
Phytohormones such as cytokinin regulation also mediates leaf senescence by transportation to different organs and changing their status as metabolic sinks (Lara et al., 2004). In this context, energetic metabolism plays a significant regulatory role. Senescence is conceived to be under a sugar-mediated gene regulation (Rolland et al., 2006; Wingler & Roitsch, 2008; Thomas, 2013). Changes in the sucrose to hexose ratio are detected by plant sensors and lead to different transduction pathways and inductive or repressive effects on

GENERAL INTRODUCTION
54
gene transcription (Smeekens et al., 2010). A protein kinase SnRK1 is activated by darkness, nutrient starvation and high cellular concentrations of sucrose or low glucose, or both, conditions associated with the induction of senescence (Jongebloed et al., 2004; Baena-González & Hanson, 2017; Crepin & Rolland, 2019). The enzyme hexokinases (HXKs) function as glucose sensors and its mutants present senescence-delaying phenotypes, e.g., Arabidopsis HXK1 and rice HXK5 and HXK6 (Moore et al., 2003; Cho et al., 2006). Senescence is therefore part of the cell fate in coordination with source-sink communications through the vascular system, during which resources are translocated form the obsolete body parts to new developing structures (Guiboileau et al., 2010; Avila-Ospina et al., 2014).
Proteolytic activities are downstream of the senescence process
Proteases are involved in numerous processes in normal cell functioning including germination, development, senescence, immunity, and stress responses (Zhou et al., 2017; Liu et al., 2018). Proteinases are the central component of executioner in the death machinery which occurs during senescence or cell death (Diaz-Mendoza et al., 2016; Martínez et al., 2018; Buono et al., 2019; Park et al., 2019). They are mostly directly produced at the change of cell structure degradation and most likely localized to the cytosol or in vacuolar compartment. Some cysteine proteases such as δVPE are expressed in both senescence and non-senescence Programmed cell death (PCD) contexts (Nakaune et al., 2005), and metacaspases are expressed in both organ senescence and PCD (Breeze et al., 2011; Coll et al., 2014; Olvera-Carrillo et al., 2015). One well known plant PCD mechanism, is found during plant-pathogen interactions, the host cell death is featured with rapid tissue collapse, i.e., the hypersensitive response (HR), regulated transcriptionally and translationally (Dangl et al., 1996). In adjacent cells of the HR sites, plant defence genes are transcriptionally elicited to constrain pathogens (Somssich et al., 1988; Heath, 2000; Balint-Kurti, 2019).
In leave senescence, autophagy has been also implicated in the incidence of diverse age-related physiological and pathological changes, by degrading and recycling damaged macromolecules and organelles (Hanaoka et al., 2002; Vauclare et al., 2010; Minina et al., 2013; Avila-Ospina et al., 2014). Macroautophagy involves formation of autophagosomes, which are double membrane-bound structures enclosing macromolecules and organelle residues (Kundu & Thompson, 2005). During leaf senescence, several possible routes for internalization of chloroplast components by the central vacuole are proposed, such as autophagosome and senescence-associated vesicle (SAV) trafficking (Avila-Ospina et al., 2014). In SAVs, the senescence-associated cysteine-protease encoded by the SAG12
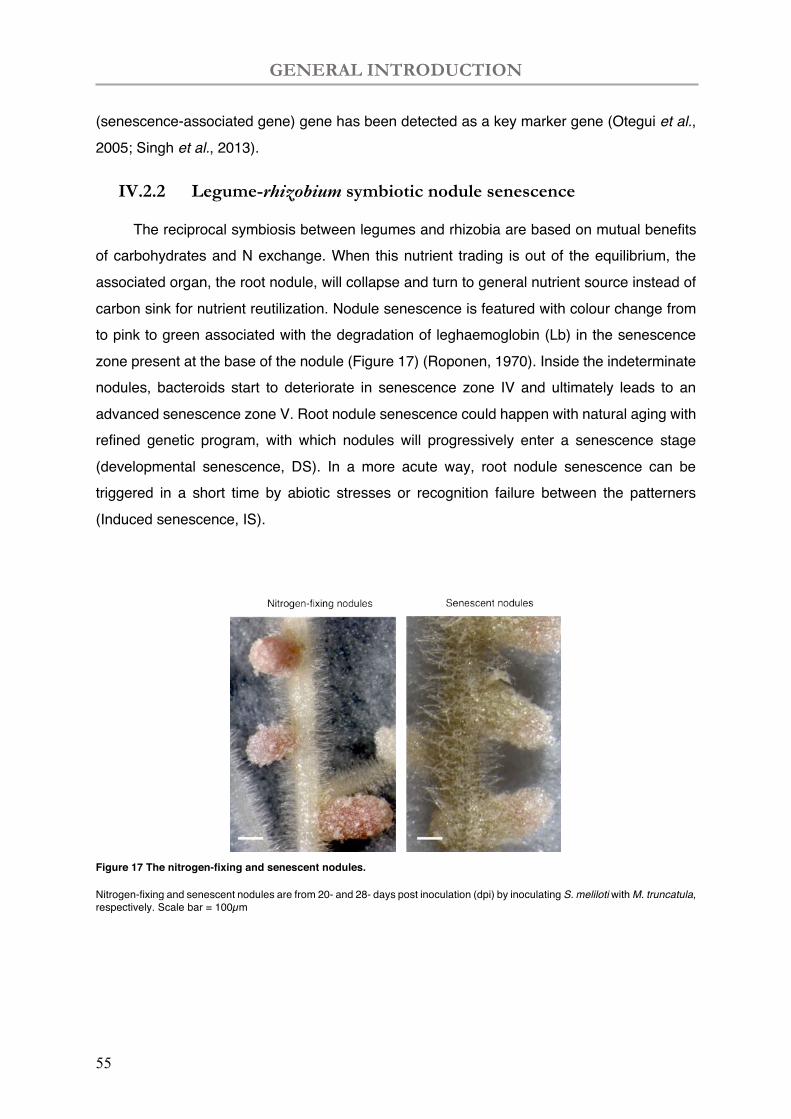
GENERAL INTRODUCTION
55
(senescence-associated gene) gene has been detected as a key marker gene (Otegui et al., 2005; Singh et al., 2013).
IV.2.2 Legume-rhizobium symbiotic nodule senescence
The reciprocal symbiosis between legumes and rhizobia are based on mutual benefits of carbohydrates and N exchange. When this nutrient trading is out of the equilibrium, the associated organ, the root nodule, will collapse and turn to general nutrient source instead of carbon sink for nutrient reutilization. Nodule senescence is featured with colour change from to pink to green associated with the degradation of leghaemoglobin (Lb) in the senescence zone present at the base of the nodule (Figure 17) (Roponen, 1970). Inside the indeterminate nodules, bacteroids start to deteriorate in senescence zone IV and ultimately leads to an advanced senescence zone V. Root nodule senescence could happen with natural aging with refined genetic program, with which nodules will progressively enter a senescence stage (developmental senescence, DS). In a more acute way, root nodule senescence can be triggered in a short time by abiotic stresses or recognition failure between the patterners (Induced senescence, IS).
Figure 17 The nitrogen-fixing and senescent nodules.
Nitrogen-fixing and senescent nodules are from 20- and 28- days post inoculation (dpi) by inoculating S. meliloti with M. truncatula, respectively. Scale bar = 100µm
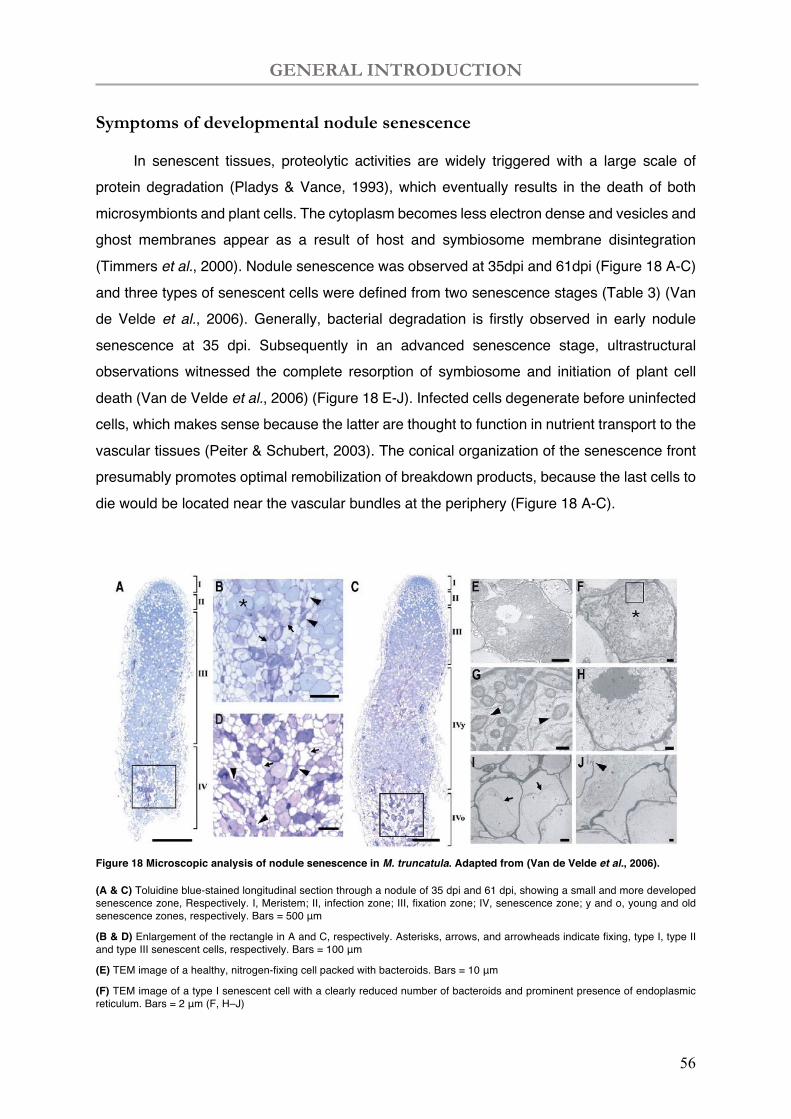
GENERAL INTRODUCTION
56
Symptoms of developmental nodule senescence
In senescent tissues, proteolytic activities are widely triggered with a large scale of protein degradation (Pladys & Vance, 1993), which eventually results in the death of both microsymbionts and plant cells. The cytoplasm becomes less electron dense and vesicles and ghost membranes appear as a result of host and symbiosome membrane disintegration (Timmers et al., 2000). Nodule senescence was observed at 35dpi and 61dpi (Figure 18 A-C) and three types of senescent cells were defined from two senescence stages (Table 3) (Van de Velde et al., 2006). Generally, bacterial degradation is firstly observed in early nodule senescence at 35 dpi. Subsequently in an advanced senescence stage, ultrastructural observations witnessed the complete resorption of symbiosome and initiation of plant cell death (Van de Velde et al., 2006) (Figure 18 E-J). Infected cells degenerate before uninfected cells, which makes sense because the latter are thought to function in nutrient transport to the vascular tissues (Peiter & Schubert, 2003). The conical organization of the senescence front presumably promotes optimal remobilization of breakdown products, because the last cells to die would be located near the vascular bundles at the periphery (Figure 18 A-C).
Figure 18 Microscopic analysis of nodule senescence in M. truncatula. Adapted from (Van de Velde et al., 2006).
(A & C) Toluidine blue-stained longitudinal section through a nodule of 35 dpi and 61 dpi, showing a small and more developed senescence zone, Respectively. I, Meristem; II, infection zone; III, fixation zone; IV, senescence zone; y and o, young and old senescence zones, respectively. Bars = 500 μm
(B & D) Enlargement of the rectangle in A and C, respectively. Asterisks, arrows, and arrowheads indicate fixing, type I, type II and type III senescent cells, respectively. Bars = 100 μm
(E) TEM image of a healthy, nitrogen-fixing cell packed with bacteroids. Bars = 10 μm
(F) TEM image of a type I senescent cell with a clearly reduced number of bacteroids and prominent presence of endoplasmic reticulum. Bars = 2 μm (F, H–J)
GENERAL INTRODUCTION
57
(G) Enlargement of the rectangle in F showing peribacteroid membrane degradation (arrowheads). Bars = 500 nm
(H) TEM image of a type I senescent cell undergoing complete resorption.
(I) TEM picture of a type III senescent cell showing signs of plant cell death, such as plasma membrane loosening (arrows).
(J) TEM image of a type II senescent cell showing loss of cellular integrity and plant cell wall breakage.
Table 3 Cell types of developmental nodule senescence. Adapted from (Van de Velde et al., 2006).
Cell types Symptoms Ultrastructure symptoms under TEM Degradation stages
Type I (35-61dpi)
Bacterial degradation:
loss of cell content;
cell rigidity maintained
(Figure 18B, arrows)
Bacteroid lysis: peribacteroid membrane degradation,
(Figure 18E, arrows);
Vesicular activity, prominent endoplamic reticulum
(Figure 18F, asterisk);
Symbiosome resorbed (Figure 18H)
First stage: bacteroid degradation
with a few dying plant
cells
Type II (35-61dpi)
Cell death
(Figure 18B,
arrowhead)
Cell wall breakage
Type III (61dpi)
Vacuolated cells
(Figure 18D, arrows)
Symbiosome resorption;
Plant cell death (departure of cell wall and plasma
membrane Figure 18I);
Advanced stage: resorbed symbiosome,
initiation of plant cell
decay and collapse
dpi: day post inoculation, TEM: transmission electron microscopy.
Symptoms of induced nodule senescence
Abiotic factors were evidenced to rapidly trigger nodule senescence, such as continuous dark treatment and extra N provision (Matamoros et al., 1999; Swaraj et al., 2001; Hernández-Jiménez et al., 2002; Perez Guerra et al., 2010). Induced senescence leads to slightly different symptoms of decaying nodules compared with developmental senescence, with a faster degradation of bacteroid content but remain of the symbiosomal structure (Table 4) (Perez Guerra et al., 2010). The start of the developmental nodule senescence was characterized by numerous vesicles, prominent endoplasmic reticulum, abundant mitochondria and Golgi apparatus, and symbiosome degradation (Figure 19A). During dark induced senescence, the bacteroid content condensed, the peribacteroid space increased (Figure 19B, arrows), and often different symbiosomes fused (Figure 19B, arrowheads). Strikingly, the PBMs remained intact, even when most of the bacteroid content had disappeared, in strong contrast to the developmental senescence, during which eventually the complete symbiosome had dissolved (Figure 19, compare C and D). After vacuolar collapse, the cytoplasm was completely degraded, the mitochondria disappeared, and the only cellular remains were apparently intact
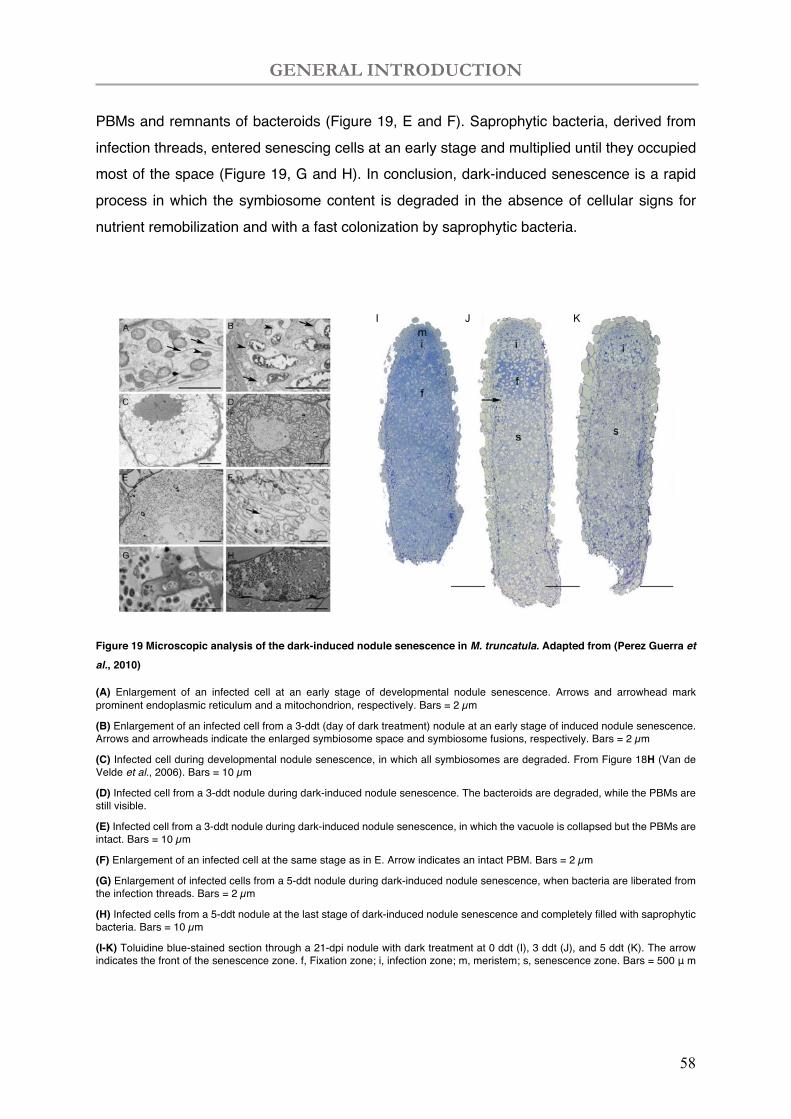
GENERAL INTRODUCTION
58
PBMs and remnants of bacteroids (Figure 19, E and F). Saprophytic bacteria, derived from infection threads, entered senescing cells at an early stage and multiplied until they occupied most of the space (Figure 19, G and H). In conclusion, dark-induced senescence is a rapid process in which the symbiosome content is degraded in the absence of cellular signs for nutrient remobilization and with a fast colonization by saprophytic bacteria.
Figure 19 Microscopic analysis of the dark-induced nodule senescence in M. truncatula. Adapted from (Perez Guerra et
al., 2010)
(A) Enlargement of an infected cell at an early stage of developmental nodule senescence. Arrows and arrowhead mark prominent endoplasmic reticulum and a mitochondrion, respectively. Bars = 2 µm
(B) Enlargement of an infected cell from a 3-ddt (day of dark treatment) nodule at an early stage of induced nodule senescence. Arrows and arrowheads indicate the enlarged symbiosome space and symbiosome fusions, respectively. Bars = 2 µm
(C) Infected cell during developmental nodule senescence, in which all symbiosomes are degraded. From Figure 18H (Van de Velde et al., 2006). Bars = 10 µm
(D) Infected cell from a 3-ddt nodule during dark-induced nodule senescence. The bacteroids are degraded, while the PBMs are still visible.
(E) Infected cell from a 3-ddt nodule during dark-induced nodule senescence, in which the vacuole is collapsed but the PBMs are intact. Bars = 10 µm
(F) Enlargement of an infected cell at the same stage as in E. Arrow indicates an intact PBM. Bars = 2 µm
(G) Enlargement of infected cells from a 5-ddt nodule during dark-induced nodule senescence, when bacteria are liberated from the infection threads. Bars = 2 µm
(H) Infected cells from a 5-ddt nodule at the last stage of dark-induced nodule senescence and completely filled with saprophytic bacteria. Bars = 10 µm
(I-K) Toluidine blue-stained section through a 21-dpi nodule with dark treatment at 0 ddt (I), 3 ddt (J), and 5 ddt (K). The arrow indicates the front of the senescence zone. f, Fixation zone; i, infection zone; m, meristem; s, senescence zone. Bars = 500 μ m
I J K

GENERAL INTRODUCTION
59
Table 4 Comparison of developmental and dark stress induced nodule senescence. Adapted from (Perez Guerra et al., 2010).
Developmental senescence (DS) Dark stress induced senescence (IS)
Factors progressive with time fast progressing upon dark stress
Vesicles numerous vesicles not pronounced vesicle mobilization
Plant cells prominent endoplasmic reticulum,
mitochondria and Golgi apparatus
absent symptoms
Symbiosome symbiosome degradation,
mixed with the cytoplasm
bacteroid content condensed,
peribacteroid space increased
PBMs degraded remained intact, fused
Saprophytic bacteria colonize in senescent cells
Transcriptomics of nodule senescence in M. truncatula
Transcriptomic approaches have been performed to get in sight to the transcriptional regulation of nodule senescence (Van de Velde et al., 2006; Seabra et al., 2012; Cabeza et al., 2014; Serova et al., 2017). Differential gene expression analysis has allowed the identification of the down-regulated genes associated with nodule function such as Lb and NCR peptides. In order to decipher transcriptional regulation mechanism related to nodule senescence, transcriptomic studies (Van de Velde et al., 2006; Seabra et al., 2012; Cabeza et al., 2014) have identified up-regulated genes associated with molecular degradation and nutrient remobilization, such as CPs and PAP (purple acid phosphatase). Moreover, the expression of MtCP1 to MtCP6, except for MtCP4, was found transiently upregulated upon dark treatment, whist the elevation of CP expressions is progressive upon time during developmental senescence (Perez Guerra et al., 2010).
Genetic approaches have revealed consecutive cascades in nodule senescence pathway, corresponding to the development of symptoms (Van de Velde et al., 2006). A transcriptome analysis revealed that developmental nodule senescence in M. truncatula is associated with a carbon sink-to-source transition and with the activation of various genes related to defence and stress. Signal transduction related genes were firstly elevated before the symptoms, which were found in healthy nodules. In addition to the refined transcriptional

GENERAL INTRODUCTION
60
program, nodule decaying is coupled with a subsequent induction of protease activities associated to ubiquitinoylation reaction. Finally, catabolic events were triggered to transport and reutilize nutrients from macromolecules, such as ATP-binding cassette proteins and phosphate transporters, amino acids, and metal ions. Interestingly, mutations in the STAY-GREEN gene, MtSGR 30 led to aberrant shoot senescence and delayed nodule senescence (Zhou et al., 2011). Silencing of two ferritin genes, MtFer2 and MtFer3, increased free iron levels in the nodule and induced early nodule senescence (Dhanushkodi et al., 2018). Nodules of the MtCas31 (cold acclimation specific 31) mutant, which is defective in a dehydrin protein that interacts with the leghaemoglobin MtLB120-1, had elevated transcript levels of senescence genes and reduced nitrogenase activity under drought stress (Li et al., 2018).
In most monocarpic legumes, pod filling often coincides with nodule senescence accompanied with a declined N fixation capacity in soybeans (Lawn & Brun, 1974), peas and common beans (Bethlenfalvay & Phillips, 1977). Exogenous application of N during pod filling increases both yield and seed protein content (Merbach & Schilling, 1980). Hence, extending the period of active N fixation by delaying the nodule senescence process might have a beneficial effect on crop yield and seed quality. The existence of soybean (Glycine max) varieties with a delayed nodule senescence and a longer active N fixation period (Espinosa‐Victoria et al., 2000) points to a genetic control of the onset of senescence, suggesting a molecular basis to modify timing and/or progression of developmental nodule senescence.

GENERAL INTRODUCTION
61
The coordinated death mechanism of plant-host and rhizobia is discussed
in the following review (Kazmierczak et al., 2020).
Kazmierczak T, Yang L, Boncompagni E, Meilhoc E, Frugier F, Frendo P, Bruand C, Gruber V, Brouquisse R 2020. Legume nodule senescence: a coordinated death mechanism between bacteria and plant cells. In: Frendo P, Frugier F, Masson-Boivin C eds. Regulation of Nitrogen-Fixing Symbioses in Legumes: Academic Press, 181-212.

GENERAL INTRODUCTION
62

CHAPTER SEVEN
Legume nodule senescence: acoordinated death mechanismbetween bacteria and plant cellsThéophile Kazmierczaka,d, Li Yangb, Eric Boncompagnib,Eliane Meilhocc, Florian Frugiera, Pierre Frendob, Claude Bruandc,Véronique Grubera and Renaud Brouquisseb,*aInstitute of Plant Sciences Paris Saclay (IPS2), CNRS, INRA, Univ Paris Diderot, Univ Paris-Sud,Univ d’Evry, Université Paris-Saclay, Gif-sur-Yvette, FrancebINRA, CNRS, Université Cote d’Azur, ISA, FrancecLaboratoire des Interactions Plantes-Microorganismes (LIPM), Université de Toulouse, INRA, CNRS,INSA, Castanet-Tolosan, FrancedBIOtransfer, Montreuil, France*Corresponding author: E-mail: [email protected]
Contents
1. Introduction
182 2. The regulation of nodule senescence 1852.1 Physiological and structural modifications
185 2.2 Redox state, ROS and RNS 188 2.3 Hormones 194 2.4 Proteases associated with bacteroid and plant cell degradation 1953. Transcriptomic characterization of nodule senescence
198 4. Concluding remarks - future prospects 201 Acknowledgments 203 References 203Abstract
AdvaISSNhttps
In the legume-rhizobia symbiosis, atmospheric dinitrogen (N2) fixation by rhizobia takesplace in a specific root organ, called the nodule. During biological N2 fixation, the plantprovides photosynthetic carbohydrates to bacteria and, in exchange, bacteria feed theplant with reduced nitrogen (N) under ammonia (NH3) form. Like most other organs,the root nodule has a limited lifespan, and eventually enters a senescence process char-acterized by a decline of N2 fixation and the coordinated death of both bacteria andplant cells. This senescence process characterized by a metabolic switch from a carbonsink to a nutrient source either results from nodule aging (developmental nodulesenescence) or can be triggered prematurely by adverse environmental conditions.Accumulating evidence suggests that redox and hormone signaling contribute to
nces in Botanical Research, Volume 940065-2296
://doi.org/10.1016/bs.abr.2019.09.013© 2020 Elsevier Ltd.All rights reserved. 181 j

182 Théophile Kazmierczak et al.
the triggering of senescence. Indeed, nodule senescence is a developmentally pro-grammed process in which antioxidants, reactive oxygen and N species (ROS/RNS), hor-mones and proteinases play key roles. This chapter reviews recent knowledge onnodule senescence and proposes models by which ROS, RNS, and likely hormonesintervene in the orchestration of this process.
1. Introduction
Plant-microbe interactions represent a major aspect of the formationand of the development of ecosystems. Plants often establish symbioticrelationships with specific fungi and bacteria to acquire the nutrients theyneed to support growth and development (Smith & Smith, 2011; VanDer Heijden, De Bruin, Luckerhoff, Van Logtestijn, & Schlaeppi, 2016;Venkateshwaran, Volkening, Sussman, & Ané, 2013; Wang et al., 2017).Amongst the different nutrients, nitrogen (N) has a major importance as itrepresents a crucial constituent of proteins, nucleic acids and chlorophylland is often in insufficient amounts in the soil under nitrate and/or ammoniaforms. As a result, plants belonging to the legume family have developed asymbiotic association with soil bacteria collectively called rhizobia to in-crease their N nutrition via the biological fixation of atmospheric dinitrogen(N2) (BNF). In BNF, the plant provides photosynthetic carbohydrates to thebacteria and, in exchange, bacteria feed the plant with reduced N underammonia (NH3) form. Legumes are a family of dicotyledonous plants ofthe Fabales order. It is the third most important family of flowering plantsafter the Orchidaceae and Asteraceae with more than 19,500 species(Christenhunsz & Byng, 2016). Economically, Fabaceae is the second largestfamily after Poaceae and is a valuable source of plant proteins for human foodand animal feeding.
The success of the symbiotic interaction relies on the molecular dialoguebetween the plant and the bacteria (Oldroyd, 2013). The plant root exudateswhich contain flavonoids and isoflavonoids induce the production byrhizobia of lipochitooligosaccharides, called Nodulation Factors (NFs;Ferguson et al., 2010). NFs are usually essential for the bacterial infectionprocess and the organogenesis of the root nodule, the organ which hoststhe N2-fixing bacteria. During the infection process, rhizobia attach toroot hairs in the susceptible zone of the host roots. The recognition of theNFs by the plant induces the root hair curling and the formation of an infec-tion pocket (Fig. 1). Then, bacteria enter into the root hairs through a

IV-Senescence
III-N2 Fixation
II-InfectionI-Division
1 dpi 4-5 dpi 3-5 wpi 3-5 wpi
Indeterminate nodule Determinate nodule
Rhizobium Infection thread Nodule primordium
Direction of increase of senescent tissueSenescent cellNitrogen fixing cell
EpCoEnVb
Fig. 1 Schematic representation of the establishment and of the development of thelegume-rhizobium symbiosis, and of the structure of mature indeterminate and deter-minate nodules. En, endodermis; Co, cortex; dpi, day post-inoculation; Ep, epidermis; Vb,vascular bundle; wpi, week post-inoculation.
Legume nodule senescence 183
membrane invagination and the formation of tubular structures, the infec-tion threads (Fig. 1), which allow the transit of the bacteria from the rootsurface to the cells of the neo-formed nodule (Timmers, Auriac, & Truchet,1999). Later, the bacteria are endocytosed into the plant cell in a new organ-elle-like structure, called symbiosome. Bacteria inside the symbiosome,called bacteroids, become able to reduce N2 to NH3 thanks to the activityof a specific enzymatic complex, the nitrogenase (Peters, Fisher, & Dean,1995).
Two predominant nodule types exist in legumes (Fig. 1). The noduletype depends on the host plant and is determined by the nature of the nodulemeristem which could be either transient or persistent (Oldroyd, 2013).Determinate nodules, with a transient meristem, are spherical since celldivisions occur only during a short period of time. The final growth ofthe nodule is achieved by cell enlargement. In contrast, indeterminate nod-ules, with a persistent meristem, are cylindrical as they grow during thewhole nodule life by increasing the number and the size of cells. Determi-nate nodules are generally formed in tropical legumes such as Phaseolus,Vigna and Glycine, whereas indeterminate nodules are found in temperatelegumes such as Medicago, Trifolium or Pisum.
Beside the nodule types, the bacteroids possess also three differentmorphotypes named U, S and E (Kondorosi, Mergaert, & Kereszt, 2013;

184 Théophile Kazmierczak et al.
Oono, Schmitt, Sprent, & Denison, 2010). The U-morphotype (for Undif-ferentiated) corresponds to bacteroids with minor morphological modifica-tions compared to the free-living bacteria and the ability to reverse from thesymbiotic lifestyle to the free-living state. The Se and E-morphotypescorrespond to large Spherical and large Elongated bacteroids, respectively.These bacteroids cannot return to free-living life and are termed “terminallydifferentiated”. This terminal differentiation is associated with the increase ofpermeability of the bacteroid membrane and of DNA content compared tofree-living bacteria (Mergaert et al., 2006). The bacteroid differentiation isdriven by the plant host via the production of a class of peptides callednodule-specific cysteine-rich peptides (NCRs; Van de Velde et al., 2010).To date, NCR peptides have been found to control the terminal differen-tiation of endosymbionts in inverted repeat-lacking clade (IRLC) legumesand in Aeschynomene spp. (Alunni et al., 2007; Czernic et al., 2015; Mergaertet al., 2003). At the functional level, NCRs induce membrane permeability(Lamouche, Bonadé-Bottino, Peter Mergaert, & Alunni, 2019) and haveintracellular bacteroid targets such as the cell division protein FtsZ and thechaperone protein GroEL, suggesting that NCRs interact with the bacterialmetabolism to induce the terminal differentiation (Farkas et al., 2014).
The cellular differentiation of both symbiotic partners is required toallow the symbiotic N2 fixation. The production of the nitrogenase enzy-matic complex in the bacteroid is a prerequisite for the reduction of N2
to NH3. This complex needs high amounts of energy, using sixteen ATPfor each N2 reduced to NH3, and as a result demands a high respirationrate in the nodule. However, the nitrogenase activity is extremely sensitiveto dioxygen (O2) implying that free O2 should be present at a very low levelin bacteroids (Peters et al., 1995). Multiple strategies allow to deal with thisparadox. To decrease the O2 level in the infected cells, the nodule inner cor-tex cell layer acts as an oxygen barrier (Tjepkema & Yocum, 1974). In par-allel, leghemoglobin (Lb), an efficient O2-binding protein, present atmillimolar concentration in the cytoplasm of infected cells (Appleby,1992), delivers O2 to the mitochondria and the bacteroids for respirationand decreases free O2 to the nanomolar range in the N2-fixing zone (Ottet al., 2005; Soupene, Foussard, Boistard, Truchet, & Batut, 1995). Finally,the bacteroid is able to function at a very low oxygen concentration thanksto a specific high affinity cbb3 cytochrome oxidase (Preisig, Zufferey,Thony-Meyer, Appleby, & Hennecke, 1996). The reduction of N2 toNH3 is accompanied by the interuption of NH3 assimilation into bacteroidamino acids. NH3 is secreted from the bacteroid into the plant cytosol, for

Legume nodule senescence 185
assimilation into the amides glutamine and asparagine, or into ureides (Day,Poole, Tyerman, & Rosendahl, 2001). In return, the plant provides carbonand energy sources to bacteroids in the form of dicarboxylic acids, particu-larly malate and succinate, which are produced from sucrose via sucrose syn-thase and glycolytic enzymes. Thus, the nodule presents multiplepeculiarities, involving a plant-microbe crosstalk associated with a meta-bolism switch which needs a high energy level under microoxic conditions.These working conditions involve a specific cellular redox control in bothsymbiotic partners. The contribution of reactive oxygen and N species(ROS and RNS) and the importance of the antioxidant defense in the for-mation and the functioning of root nodules have been largely demonstrated(Alloing, Mandon, Boncompagni, Montrichard, & Frendo, 2018; Damiani,Pauly, Puppo, Brouquisse, & Boscari, 2016; Frendo, Matamoros, Alloing, &Becana, 2013; Puppo, Pauly, Boscari, Mandon, & Brouquisse, 2013).
Whereas formation and functioning of the root nodule has beenextensively analyzed the rupture of the interaction leading to the nodulesenescence has not been much characterized and only few reviews havebeen dedicated to nodule senescence (Bruand & Meilhoc, 2019; Dupontet al., 2012; Puppo et al., 2005; Serova & Tsyganov, 2014). Some investiga-tors have used the initial decline in nitrogenase activity and BNF efficiencyas an indication that nodule senescence has begun (Klucas, 1974). Others au-thors have defined nodule senescence as the period of rapid nodule decaywhich occurs at a plant physiological maturity stage, i.e. at leaf yellowing(Streeter, 1981), after flowering has finished (Serova & Tsyganov, 2014),or at pod filling (Bethlenfalvay & Phillips, 1977; Lawn & Brun, 1974).The nodule senescence represents a strong agronomical interest throughthe possible improvement of N2 fixation efficiency under adverse environ-mental conditions to maintain or even increase the yield of legume crops. Inthis review, we summarize the different modifications of nodule functioningobserved during the set up and the progress of nodule senescence, and wefocus on the role of antioxidants, ROS/RNS, hormones and proteases inregulating this process.
2. The regulation of nodule senescence
2.1 Physiological and structural modifications
Although the lifespan of nodules of most legumes is 10e12 weeks,their ability to fix N starts to decline 3e5 weeks post-inoculation (Puppo
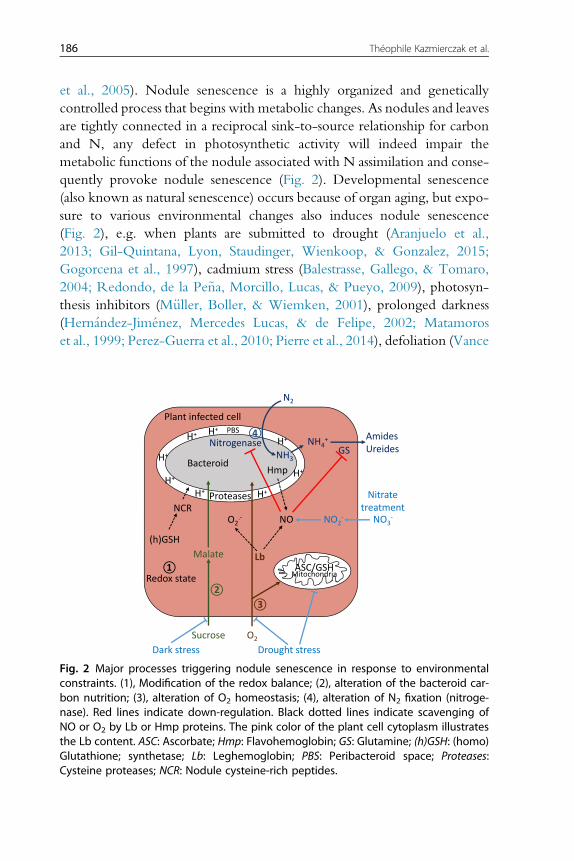
186 Théophile Kazmierczak et al.
et al., 2005). Nodule senescence is a highly organized and geneticallycontrolled process that begins with metabolic changes. As nodules and leavesare tightly connected in a reciprocal sink-to-source relationship for carbonand N, any defect in photosynthetic activity will indeed impair themetabolic functions of the nodule associated with N assimilation and conse-quently provoke nodule senescence (Fig. 2). Developmental senescence(also known as natural senescence) occurs because of organ aging, but expo-sure to various environmental changes also induces nodule senescence(Fig. 2), e.g. when plants are submitted to drought (Aranjuelo et al.,2013; Gil-Quintana, Lyon, Staudinger, Wienkoop, & Gonzalez, 2015;Gogorcena et al., 1997), cadmium stress (Balestrasse, Gallego, & Tomaro,2004; Redondo, de la Pe~na, Morcillo, Lucas, & Pueyo, 2009), photosyn-thesis inhibitors (M€uller, Boller, & Wiemken, 2001), prolonged darkness(Hernandez-Jiménez, Mercedes Lucas, & de Felipe, 2002; Matamoroset al., 1999; Perez-Guerra et al., 2010; Pierre et al., 2014), defoliation (Vance
Plant infected cell
Sucrose O2
Lb
Proteases
Nitrogenase
Hmp
Redox state
N2
NH3
AmidesUreides
O2.-
(h)GSH
①
②③
④PBS
Drought stressDark stress
H+H+
H+
H+
H+H+
H+
H+
NO3-NO2
-NO
NH4+
ASC/GSH
GS
Malate
Bacteroid
Nitrate treatmentNCR
Mitochondria
Fig. 2 Major processes triggering nodule senescence in response to environmentalconstraints. (1), Modification of the redox balance; (2), alteration of the bacteroid car-bon nutrition; (3), alteration of O2 homeostasis; (4), alteration of N2 fixation (nitroge-nase). Red lines indicate down-regulation. Black dotted lines indicate scavenging ofNO or O2 by Lb or Hmp proteins. The pink color of the plant cell cytoplasm illustratesthe Lb content. ASC: Ascorbate; Hmp: Flavohemoglobin; GS: Glutamine; (h)GSH: (homo)Glutathione; synthetase; Lb: Leghemoglobin; PBS: Peribacteroid space; Proteases:Cysteine proteases; NCR: Nodule cysteine-rich peptides.

Legume nodule senescence 187
et al. al., 1979), or treatments with argon (Curioni, Hartwig, Nosberger, &Schuller, 1999), nitrate (Matamoros et al., 1999; Pierre et al., 2014) orabscisic acid (ABA; Gonzalez, Galvez, & Arrese-Igor, 2001). Alternately,senescence can result from a deficient recognition between the two partners(Banba, Siddique, Kouchi, Izui, & Hata, 2001). Depending on the type oftriggering factor, the senescence process is more or less rapid and implementvariable responses (Van de Velde et al., 2006), but a number of commontypical features are found in most nodule senescence processes.
During developmental senescence, the first sign of nodule senescence isthe drop in measurable N2 fixation activity that occurs in parallel with adecrease in Lb content (Dalton, Russell, Hanus, Pascoe, & Evans, 1986;Lahiri, Chattopadhyay, Chatterjee, & Ghosh, 1993). However, duringinduced senescence, such as after dark treatment, drought stress, or ABAtreatment, the rapid decline in N2 fixation occurs well before a decreasein Lb transcripts or amounts is observed (Escuredo et al., 1996; Gogorcenaet al., 1997; Gonzalez et al., 2001; Matamoros et al., 1999). The most visiblesign of senescence is a change of the nodule color, from pink to green,caused by bilirubin and biliverdin accumulation after disruption of the Lbstructure (Lehtovaara & Perttil€a, 1978; Swaraj & Bishnoi, 1996). In senes-cent soybean nodules, the green color comes partially from Lb nitration,which highlights the critical role of ROS and RNS in the senescence process(Meakin et al., 2007; Navascués et al., 2012; Sainz et al., 2015).
In nodules of the determinate type, senescence develops radially (Fig. 1),starting from the center of the nodule and then progressively extending tothe peripheral tissues in a few weeks (Puppo et al., 2005). In nodules ofindeterminate type, the N2 fixation zone is constantly renewed and thesenescence zone (zone IV) grows as the nodule grows (Fig. 1); it increasesfrom the root proximal zone toward the apical zone of the nodule (Montiel,Arthikala, Cardenas, & Quinto, 2016; Perez-Guerra et al., 2010; Puppoet al., 2005). Because this zonation is not strictly defined, a senescence in-ter-zone is sometimes described between zone III and zone IV. Histologi-cally, early nodule senescent cells exhibit a less dense cytoplasmcontaining numerous vacuoles, altered peroxisomes, mitochondria, starchgranules, and late senescent cells ultimately accumulate damaged “ghost”membranes and cell walls (Perez-Guerra et al., 2010; Puppo et al., 2005).In common bean nodule developmental senescence, mitochondria werefound to be early targets of oxidative modifications and a potential sourceof ROS/RNS (Matamoros et al., 2013). A dynamic evolution of both plantand bacteroid cell structures is observed: bacteroids are first degraded

188 Théophile Kazmierczak et al.
whereas plant cells remain alive during the early senescence stage, allowingnutrient remobilization, even though at later stages plant cells are alsodegraded (Van de Velde et al., 2006; Vasse, de Billy, Camut, & Truchet,1990). In senescent cells, the degradation of symbiosomes is correlatedwith the loss of the acidity of the peribacteroid space (Pierre et al., 2013),this acidification being an indicator of the bacteroid functioning in thenodule N fixing zone (Pierre et al., 2013).
The senescence process was analyzed at gene expression level in bothleaves and nodules, particularly in Medicago truncatula (De Michele et al.,2009; Van de Velde et al., 2006). It appeared that nodule senescence has asignificant degree of overlap with leaf senescence, arguing for the recruit-ment of similar pathways. A strong redox regulation, through antioxidantsdecrease and ROS/RNS accumulation, as well as hormonal regulationssuch as abscisic acid, ethylene or jasmonic acid, was evidenced to correlatewith catabolic and remobilization processes during nodule senescence(Puppo et al., 2005; Van de Velde et al., 2006).
2.2 Redox state, ROS and RNSChanges in the redox balance, which modify the antioxidant defense andthe level of ROS and RNS, is directly associated with the transition betweenN2 fixation and senescence in nodules (Fig. 2; Puppo et al., 2005).
Ascorbate (ASC) and glutathione (GSH) are the main reported solubleantioxidants within nodules and their regulation was supposed to controlthe organ lifespan (Becana, Matamoros, Udvardi, & Dalton, 2010; Loscos,Matamoros, & Becana, 2008). In soybean plants, drought-induced nodulesenescence is characterized by a decrease in ASC reduction state and an in-crease in GSH content (Marquez-Garcia et al., 2015). In bean nodules, theprogression of developmental senescence correlates with a decrease in bothASC and GSH concentrations and reduction states (Loscos et al., 2008).Moreover, the decrease in ASC and GSH concentrations in mitochondriacorrelates to an increase of oxidative modifications of lipids and proteins,supporting the hypothesis that the mitochondria is an early target of senes-cence in nodules (Matamoros et al., 2013). In contrast, an increased level ofGSH in the N2-fixing zone is correlated with a higher N2 fixation efficiency(El Msehli et al., 2011). GSH content is notably involved in the control ofglutathione S-transferases (GSTs) activities, which are enzymes involved inthe cell protection from an oxidative damage, by catalyzing the conjugationof GSH with oxidative degradation products. The level of the GST9 pro-tein, the most prevalent glutathione-S-transferase in soybean nodules, is

Legume nodule senescence 189
accordingly increased during senescence (Dalton et al., 2009). A decrease inN2 fixation associated with an increase of the oxidatively damaged proteincontent is observed in GST9-RNAi silenced soybean nodules, illustratingthe role of GSTs in nodule senescence through the regulation of antioxidantdefences (Dalton et al., 2009). In parallel, the importance of GSH duringsymbiosis of Sinorhizobium meliloti and Rhizobium tropici has been alsodemonstrated (Harrison et al., 2005; Muglia, Comai, Spegazzini, Riccillo,& Aguilar, 2008). The S. meliloti and R. tropici strains deficient in gshB,the gene encoding the enzyme catalyzing the second step of GSH synthesis,have a reduced N2 fixation capacity associated with early nodule senescence.Similarly, mutation in the S. meliloti glutathione reductase gene, gor,impaired N2 fixation efficiency (Tang et al., 2018).
During the legume-rhizobium symbiosis, both ROS and RNS, andespecially nitric oxide (NO), appear to play crucial roles in the establishmentand functioning of the interaction (Hichri et al., 2015; Puppo et al., 2013).ROS including superoxide radicals (O2
.) and hydrogen peroxide (H2O2)are produced in high amounts in nodules due to elevated rates of respirationin bacteroids and plant mitochondria (Matamoros et al., 2013). PlantNADPH oxidases, known as Respiratory Burst Oxidase Homologues(RBOHs), are plasma membrane enzymes catalyzing the reduction of O2
to generate O2. using the reducing power of NADPH. Seven Rboh genes
have been identified in legume genomes (Marino et al., 2011) and these ox-idases are involved in legume-rhizobium symbioses (Arthikala et al., 2014;Marino et al., 2011; Montiel et al., 2016). Other possible sources forH2O2 are cell wall peroxidases, germin-like oxalate oxidases, and diamineoxidases (Smirnoff & Arnaud, 2019). On the other hand, NO is a reactivediffusible gas known to have important functions as inter- or intra-cellularsignaling molecules in plant growth, development and defense againstabiotic or biotic stresses (Brouquisse, 2019, and references therein). For adecade, NO has been also identified at all stages of legume-rhizobium sym-biotic interactions (Berger, Boscari, Frendo, & Brouquisse, 2019; Hichriet al., 2016, 2015). The way NO is produced in legumes is not completelyclear. The best characterized sources of NO are the nitrate reductase (NR)and the mitochondrial transfer chain (Horchani et al., 2011). Interestingly, ithas been shown in soybean andM. truncatula nodules that the micro-symbi-ont accounts for 30% up to 90% of NO production thanks to a nitrite reduc-tase (Nir) belonging to the denitrification pathway (Calvo-Begueria et al.,2018; Horchani et al., 2011; Sanchez et al., 2010). Even though ROSand NO are different types of molecules issuing from distinct synthesis

190 Théophile Kazmierczak et al.
pathways, some parallels can be established when looking at their occurrenceduring symbiosis. Strikingly, NO and ROS have comparable productionpatterns during symbiosis such as an early production burst in the root inresponse to rhizobia inoculation, later in the infection thread, then duringnodule organogenesis and finally in mature nodules (Hichri et al., 2016;Montiel et al., 2016).
Interestingly, NO and ROS play a role in nodule senescence. At a latestage of the nodulation process (e.g. 6 weeks post-inoculation), H2O2 isdetected in ultrathin sections of nodules (Puppo et al., 2013). At the ultra-structural level, the senescent nodules of Glycine max and Pisum sativumplants display alterations in the symbiosome structure accompanied by thepresence of H2O2, supporting the idea that ROS have roles also duringsenescence (Montiel et al., 2016). In addition, it has been suggested thatNO is present in higher amounts during the late symbiotic stages than atearlier stages (Bruand & Meilhoc, 2019). Indeed, NO detected by meansof a bacterial biosensor is more abundant in the root-proximal part of M.truncatula nodules, where senescence is known to initiate (Cam et al.,2012). Similarly, NO detected by means of a fluorescent probe accumulatesin old Lotus japonicus nodules and Lbs with nitrated heme groups are found inold soybean nodules, indicating the presence of NO in aging nodules(Chungopast, Duangkhet, Tajima, Ma, & Nomura, 2017; Navascuéset al., 2012; Sainz et al., 2015). Remarkably, exogenous addition of a NOdonor on mature nodules of M. truncatula lead to an accelerated senescenceof the nodules (Cam et al., 2012). Also, several S. meliloti mutant strains,altered on hmp, nnrS1, nnrS2 and norB genes (Table 1) and showing eitherincreased or decreased NO degradation capacity compared to the wild-type, were used to modulate NO levels within nodules and, strikingly,strains deficient in NO degradation induced an early nodule senescence,whereas strains with higher degradation activity led to a delayed nodulesenescence (Blanquet et al., 2015; Cam et al., 2012; Meilhoc, Blanquet,Cam, & Bruand, 2013). NO is also involved in stress-induced M. truncatulanodule senescence (Cam et al., 2012). Comparable results were recentlyobtained in determinate L. japonicus nodules (Fukudome et al., 2019).Indeed, overexpression of a phytoglobin gene LjGlb1-1 (Table 1) decreasedNO levels in nodules and delayed nodule senescence.
By analogy, Redondo et al. (2009, 2012) reported that overexpression inS. meliloti of a cyanobacterial flavodoxin involved in the response tooxidative stress in bacteria and cyanobacteria improves the oxidative balance,delays developmental nodule senescence in Medicago sativa, and enhances
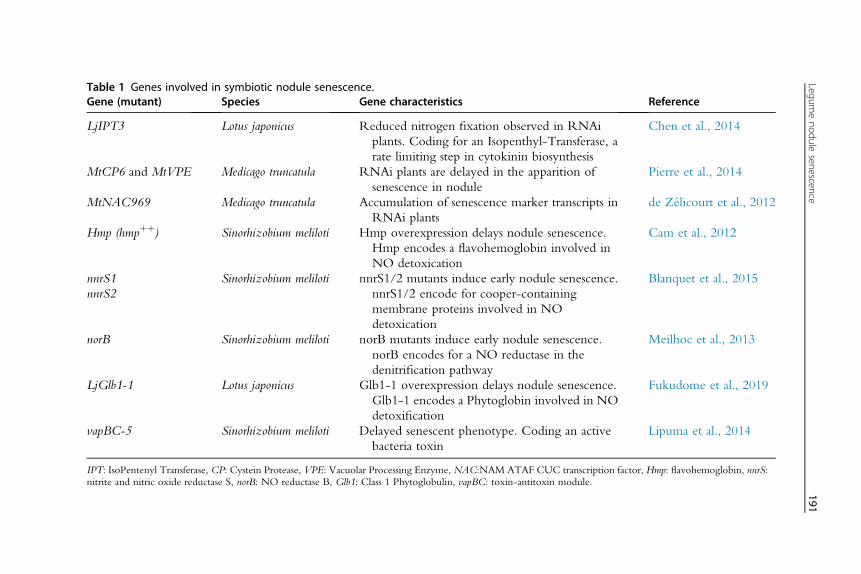
Table 1 Genes involved in symbiotic nodule senescence.Gene (mutant) Species Gene characteristics Reference
LjIPT3 Lotus japonicus Reduced nitrogen fixation observed in RNAiplants. Coding for an Isopenthyl-Transferase, arate limiting step in cytokinin biosynthesis
Chen et al., 2014
MtCP6 and MtVPE Medicago truncatula RNAi plants are delayed in the apparition ofsenescence in nodule
Pierre et al., 2014
MtNAC969 Medicago truncatula Accumulation of senescence marker transcripts inRNAi plants
de Zélicourt et al., 2012
Hmp (hmpþþ) Sinorhizobium meliloti Hmp overexpression delays nodule senescence.Hmp encodes a flavohemoglobin involved inNO detoxication
Cam et al., 2012
nnrS1nnrS2
Sinorhizobium meliloti nnrS1/2 mutants induce early nodule senescence.nnrS1/2 encode for cooper-containingmembrane proteins involved in NOdetoxication
Blanquet et al., 2015
norB Sinorhizobium meliloti norB mutants induce early nodule senescence.norB encodes for a NO reductase in thedenitrification pathway
Meilhoc et al., 2013
LjGlb1-1 Lotus japonicus Glb1-1 overexpression delays nodule senescence.Glb1-1 encodes a Phytoglobin involved in NOdetoxification
Fukudome et al., 2019
vapBC-5 Sinorhizobium meliloti Delayed senescent phenotype. Coding an activebacteria toxin
Lipuma et al., 2014
IPT: IsoPentenyl Transferase, CP: Cystein Protease, VPE: Vacuolar Processing Enzyme, NAC:NAM ATAF CUC transcription factor, Hmp: flavohemoglobin, nnrS:nitrite and nitric oxide reductase S, norB: NO reductase B, Glb1: Class 1 Phytoglobulin, vapBC: toxin-antitoxin module.
Legumenodule
senescence191

192 Théophile Kazmierczak et al.
tolerance to cadmium. Overexpression of this flavodoxin also permitted toavoid the decline in nitrogenase activity associated with a salinity stresswhether the flavodoxin was expressed in S. meliloti or in M. truncatula(Coba de la Pe~na, Redondo, Manrique, Lucas, & Pueyo, 2010). In S. meli-loti, various ROS scavenging enzymes (katA and katC catalases particularly)are expressed in the N2-fixing zone (Roux et al., 2014). The S. meliloti katAkatC double mutant displays a reduced N2 fixation activity and bacteroidsunderwent faster senescence (Puppo et al., 2013). Rhizobium etli and Bradyr-hizobium japonicum possess only one catalase gene and corresponding mutantswere not affected in nodule functioning, but ROS scavenging enzymesother than catalases might reduce H2O2 (Puppo et al., 2013). This mightalso suggest a different role for ROS and antioxidant defense componentsin the senescence of determinate and indeterminate nodules.
A drastic reduction in N2 fixation was observed in Phaseolus vulgaris nod-ules silenced in NADPH oxidase (Rboh) genes probably linked to an earlysenescence phenotype (Arthikala et al., 2014). By contrast, overexpressionof RbohB delays senescence of the nodule. Hence, these elements suggestthat ROS produced by RBOHs may have either a promoting, or an inhib-iting role during nodule senescence in P. vulgaris. Two Rboh genes(MtRbohA and MtRbohB) were found specifically expressed in the N2-fixingzone of M. truncatula nodules (Marino et al., 2011; Montiel et al., 2016;Roux et al., 2014). A decrease in MtRbohA expression via a RNA interfer-ence (RNAi) approach led to a reduction in the N2 fixation capacity,showing again the link between MtRBOHA and nodule function (Marinoet al., 2011). However, none of these genes displays any detectable promoteractivity in the senescence zone of indeterminate nodules, suggesting thatthey do not directly act in nodule senescence. In addition, RBOHs mighthave a more prominent role in determinate nodules than in indeterminatenodules, even though the possibility that they participate in nodule senes-cence should not be eliminated before each gene has been studied in detailin different plants.
Hypotheses on mechanisms by which NO and ROS could triggernodule senescence are multiple. These molecules can both act at the tran-scriptional and post-translational levels. Indeed, NO is known to activatethe expression of bacterial as well as plant genes (Boscari et al., 2013a;Ferrarini et al., 2008; Meilhoc, Cam, Skapski, & Bruand, 2010). However,it is unknown yet which genes or transcription factors (TFs) could beinvolved at the onset of nodule senescence. One of the main downstreameffects of NO is the post-translational modification of proteins such as

Legume nodule senescence 193
S-nitrosation of cysteine residues or nitration of tyrosine residues, whichmight cause conformational changes in the protein with concomitantchanges in their activities, turnover or localization. About 80 proteinswere found to be S-nitrosated in M. truncatula nodules; among them 20and 8 plant and bacterial proteins, respectively, were also found to be sulfe-nylated by ROS (Puppo et al., 2013). These protein targets remain to becharacterized to establish a potential link with the nodule senescence. How-ever, the activity of many of them, involved either in the glycolysis, thetricarboxylic acid cycle, the energy regeneration or the N2 assimilation,were found to be inhibited by NO, suggesting that an increased NO pro-duction could directly down-regulate N, carbon and energy metabolismin senescent nodules (Boscari et al., 2013b). Interestingly, the enzymaticactivity of the plant glutamine synthetases (GS) involved in N assimilationdecreases when GS is subjected to NO-mediated tyrosine nitration (Melo,Silva, Ribeiro, Seabra, & Carvalho, 2011). Nevertheless, methionine sulfox-idation of GS is also important for its regulation in response to ROS,showing again that both NO and ROS can be involved in the regulationof protein activity (Matamoros et al., 2013). It has also been shown thatthe nitrogenase activity is inhibited by NO (Trinchant & Rigaud, 1982).
NO and ROS are generated in cells at the same time and in response tothe same cues, so that it is important to consider their possible interactionand associated consequences. One of the best known interaction betweenNO and ROS is the production of peroxynitrite (ONOO) from NOand superoxide anion (O2
.). This has two potentially important outcomes:first, the reaction removes both O2
. and NO from the cell environment,thereby reducing their bioavailability; and second, there is a new compoundproduced (ONOO) which itself can act as a signaling/regulatory molecule,potentially giving different responses than that resulting from ROS or NOsignaling alone. As an example, peroxynitrite is more potent than NO aloneto trigger tyrosine nitration (Astier & Lindermayr, 2012).
NO/ONOO-dependent modifications may also alter signaling path-ways mediated by other reactive signals. Indeed, Tyr nitration of superoxidedismutase can inhibit its activity and hence alter ROS signaling (Holzmeisteret al., 2015). NO has also been proposed as a negative regulator of RBOHactivity in nodulation (Damiani et al., 2016). Indeed, it has been reportedthat the AtRBOHD enzyme is S-nitrosated which thus reduces its ROSgenerating activity (Yun et al., 2011). Therefore, NO has a role in control-ling ROS levels and hence their associated downstream signaling. In addi-tion, the B. japonicum key regulator of the N2-fixing enzymes FixK2 is

194 Théophile Kazmierczak et al.
reversibly susceptible to oxidation, which might indirectly have an influenceon the expression of bacterial genes involved in the NO metabolism regu-lated by FixK2 (Mesa, Reutimann, Fischer, & Hennecke, 2009). Similarly,LsrB, a TF involved in the nodulation and redox regulation, is also regulatedby its redox state and might be involved in the senescence process (Tanget al., 2017).
2.3 HormonesPlant hormones are other obvious candidate regulatory molecules to controlthe transition between N2 fixation and nodule senescence, some of thesesignals being previously associated with leaf senescence, such as cytokininsthat delay leaf senescence (Zwack & Rashotte, 2013). In white lupin,expression of the cytokinin receptor LaHK1, closely related to AHK4/CRE1 in Arabidopsis thaliana, is increased during nodule senescence (Cobade la Pe~na et al., 2008). In L. japonicus, expression of the LjIPT3 (IsoPen-tenyl-adenine Transferase 3, Table 1) gene encoding a rate-limiting cytokininbiosynthetic enzyme, increases during nodule aging and is associated withvascular bundles of mature nodules. LjIPT3 down-regulation by anRNAi approach triggers a decrease in N2 fixation, consequently associatedwith premature nodule senescence (Chen et al., 2014). In addition, a main-tained N2 fixation associated with delayed nodule senescence was observedin drought-stressed nodules of alfalfa inoculated with an S. meliloti strainengineered to overexpress an IPT cytokinin biosynthesis gene and thereforeproducing increased levels of cytokinins (Xu, Li, & Luo, 2012). Cytokininmay therefore act as a positive regulator of N2 fixation (Boivin et al.,2016; Reid, Heckmann, Novak, Kelly, & Stougaard, 2016), and possiblyas a negative regulator of root nodule senescence as reported during leafsenescence (Zwack, Robinson, Risley, & Rashotte, 2013).
Besides cytokinins, several gene expression analyses revealed putativefunctions for other plant hormones (De Michele et al., 2009; Van de Veldeet al., 2006). Among those, genes encoding enzymes of the ethylene biosyn-thesis pathway, such as an 1-aminocyclopropane-1-carboxylate (ACC)oxidase, are induced in senescent nodules (Van de Velde et al., 2006). Inter-estingly, the lack of an ACC deaminase activity in the Sinorhizobium strain sp.BL3 accelerates nodule senescence in mung bean, suggesting a positive rolefor ethylene in nodule senescence similarly to what is observed during leafsenescence (Tittabutr et al., 2015). Lipoxygenase genes associated withJasmonic Acid (JA) biosynthesis and GA2-oxidase genes associated withGibberellic Acid (GA) degradation are also up-regulated during nodule

Legume nodule senescence 195
senescence, suggesting that JA may positively and GA negatively regulatenodule senescence (Benedito et al., 2008; Van de Velde et al., 2006). Absci-sic acid (ABA) could be also involved in nodule senescence since some ABAresponsive genes such asABR17 (ABA Responsive 17) are induced during ni-trate-induced senescence (Liese, Schulze, & Cabeza, 2017). In addition,ABA, which is associated with various environmental stress responses,improves nodule N2 fixation capacity in alfalfa salt-stressed nodules whereantioxidant defenses mediated by the polyamine spermidine and spermineare induced (Palma, Lopez-Gomez, Tejera, & Lluch, 2014), and mighttherefore regulate senescence progression. Such putative negative effect ofABA on nodule senescence is however not observed in leaves whereABA is a positive regulator of senescence (Lee et al., 2011).
Considered together, if each of these studies suggests that hormones canintervene in the regulation of senescence, in most cases, functional analysesare lacking to clearly identify how these hormones regulate nodule senes-cence and more specifically how they may interact with ROS and RNSsignaling pathways that are the most extensively linked to nodule senes-cence. These limited data may nevertheless suggest that common hormonalregulations between leaf and nodule senescence may exist, which could besurprising considering their differences in carbon versus N metabolic status.
2.4 Proteases associated with bacteroid and plant celldegradation
A hallmark of cell aging is the progressive decline in proteostasis and theaccumulation of misfolded, oxidized and damaged proteins (Lopez-Otín,Blasco, Partridge, Serrano, & Kroemer, 2013). Therefore, proteolytic activ-ities are under strict regulation to ensure protease activation and degradationat appropriate time and space. In plants, proteases have been associated withglobal protein turnover, protein processing, degradation of regulatory pro-teins, cell growth and differentiation, stress response and developmentalsenescence, but to date functional evidence of their mode of action remainsstill limited (Buono, Hudecek, & Nowack, 2019). There are hundreds ofprotease-encoding genes; for example, more than 800 have been identifiedin Arabidopsis (Van der Hoorn, 2008). Such a large number of proteases canbe grouped into families according to their catalytic residues, aspartic-,cysteine-, serine-, threonine-, glutamic-, asparagine-, or metalloproteases(Oda, 2012). A description of the protease classification can be retrievedin the MEROPS database (https://www.ebi.ac.uk/merops/; Rawlings,Barrett, & Bateman, 2010).

196 Théophile Kazmierczak et al.
To date, many reports have elucidated the roles of cysteine proteases(CPs) in nodule senescence and in response to detrimental environments(Cilliers, van Wyk, van Heerden, Kunert, & Vorster, 2018; Perez-Guerraet al. 2010; Pierre et al. 2014; Pladys, Dimitrijevic, & Rigaud, 1991; Pladys& Vance, 1993; Van de Velde et al. 2006). The synthesis of CP precursors isorchestrated by a complex transcriptional regulation network. The post-translational activation of many CPs relies on either self-processing or onthe interaction with other enzymes. Their activity is also modulated by in-hibitors and co-factors. CPs encoding genes are among the most stronglyand specifically induced nodule senescence genes, either developmentallyor environmentally triggered (Cabeza et al., 2014; Chungopast et al.,2014; Seabra, Pereira, Becker, & Carvalho, 2012; Van de Velde et al.,2006). Their activity is directly correlated to the protein degradation occur-ring during senescence, and was documented in various legume plantsincluding soybean (Alesandrini, Mathis, Van de Sype, Hérouart, & Puppo,2003), Astragalus sinicus (Naito, Fujie, Usami, Murooka, & Yamada, 2000),pea (Kardailsky & Brewin, 1996), and M. truncatula (Pierre et al., 2014).
In soybean, 18 cysteine proteases were identified as actively transcribedin nodules during the developmental senescence (Van Wyk, Du Plessis,Cullis, Kunert, & Vorster, 2014). This expression was increased at specifictime points of the senescence process with a majority of CP genes highlyexpressed in 14 week-old nodules. In Trifolium repens, the identified CPencoding genes were specifically expressed in the senescent zone of thenodule as revealed by a spatio-temporal analysis (Asp, Bowra, Borg, &Holm, 2004). In A. sinicus indeterminate nodules, the papain cysteine pro-tease AsNodF32 expressed in senescent tissues of the nodule was identifiedfrom a differential screening of a nodule cDNA library (Naito et al.,2000). An RNAi silencing of AsNodF32 extended the period of the activeN2-fixation, delaying nodule senescence (Li et al., 2008). In M. truncatulanodules, the Cysteine Protease 6 (MtCP6), (MEROPS family C1A;https://www.ebi.ac.uk/merops/), and a Vacuolar Processing Enzyme(MtVPE; MEROPS family C13), belonging respectively to papain andlegumain families, were strongly up-regulated in response to both develop-mental and environmentally-induced nodule senescence (Van de Veldeet al., 2006; Pierre et al., 2014, Table 1). A detailed spatio-temporal analysisof theMtCP6 andMtVPE expression patterns during nodule developmentalsenescence revealed an expression associated with the transition regionbetween the N2-fixing and senescence zones (Fig. 3A and B, Pierre et al.,2014). In nitrate-induced senescent nodules,MtCP6 andMtVPE expression
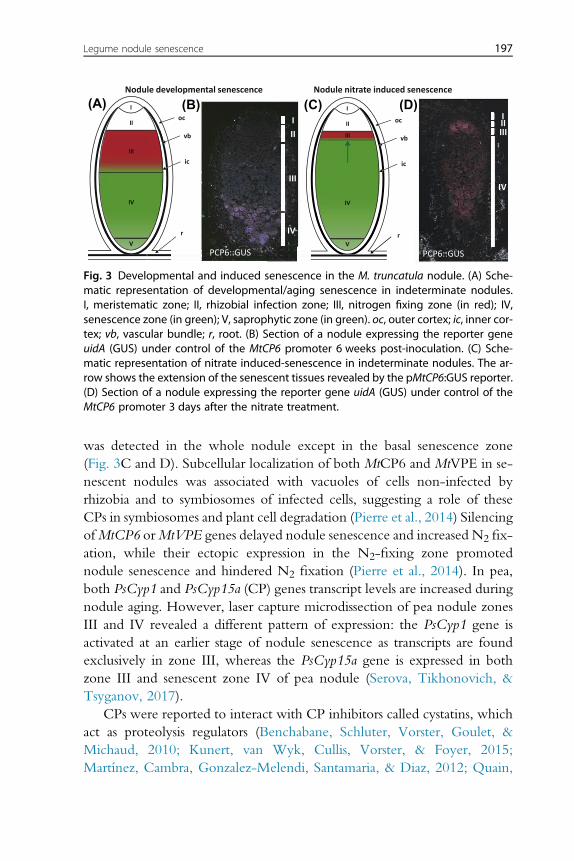
II
III
IV
I
V
II
III
IV
I
V
oc
vb
ic
r
Nodule developmental senescence
PCP6::GUS
III
III
I
IV
Nodule nitrate induced senescence
oc
vb
ic
r
PCP6::GUS
IIIIII
IV
(A) (B) (C) (D)
Fig. 3 Developmental and induced senescence in the M. truncatula nodule. (A) Sche-matic representation of developmental/aging senescence in indeterminate nodules.I, meristematic zone; II, rhizobial infection zone; III, nitrogen fixing zone (in red); IV,senescence zone (in green); V, saprophytic zone (in green). oc, outer cortex; ic, inner cor-tex; vb, vascular bundle; r, root. (B) Section of a nodule expressing the reporter geneuidA (GUS) under control of the MtCP6 promoter 6 weeks post-inoculation. (C) Sche-matic representation of nitrate induced-senescence in indeterminate nodules. The ar-row shows the extension of the senescent tissues revealed by the pMtCP6:GUS reporter.(D) Section of a nodule expressing the reporter gene uidA (GUS) under control of theMtCP6 promoter 3 days after the nitrate treatment.
Legume nodule senescence 197
was detected in the whole nodule except in the basal senescence zone(Fig. 3C and D). Subcellular localization of both MtCP6 and MtVPE in se-nescent nodules was associated with vacuoles of cells non-infected byrhizobia and to symbiosomes of infected cells, suggesting a role of theseCPs in symbiosomes and plant cell degradation (Pierre et al., 2014) SilencingofMtCP6 orMtVPE genes delayed nodule senescence and increased N2 fix-ation, while their ectopic expression in the N2-fixing zone promotednodule senescence and hindered N2 fixation (Pierre et al., 2014). In pea,both PsCyp1 and PsCyp15a (CP) genes transcript levels are increased duringnodule aging. However, laser capture microdissection of pea nodule zonesIII and IV revealed a different pattern of expression: the PsCyp1 gene isactivated at an earlier stage of nodule senescence as transcripts are foundexclusively in zone III, whereas the PsCyp15a gene is expressed in bothzone III and senescent zone IV of pea nodule (Serova, Tikhonovich, &Tsyganov, 2017).
CPs were reported to interact with CP inhibitors called cystatins, whichact as proteolysis regulators (Benchabane, Schluter, Vorster, Goulet, &Michaud, 2010; Kunert, van Wyk, Cullis, Vorster, & Foyer, 2015;Martínez, Cambra, Gonzalez-Melendi, Santamaria, & Diaz, 2012; Quain,

198 Théophile Kazmierczak et al.
Makgopa, Cooper, Kunert, & Foyer, 2015; Van Wyk et al., 2014). In soy-bean developmental senescent nodules, three cystatin genes are co-expressedwith CPs (Van Wyk et al., 2014). Remarkably, despite no direct evidencefor a role of cystatins during nodule senescence was yet reported, an ectopicoverexpression of a rice oryzacystatin in soybean leads to a strong decrease incathepsin L-like CP activities that are linked to nodule senescence (Quainet al., 2015). The coordinated expression of transcripts encoding cystatinsand CPs interacting partners suggests their involvement as a fine-tuning con-trol of proteolysis events associated with nodule senescence. InM. truncatulanodules, serpins (serine protease inhibitors) have also been found to be acti-vated in drought-induced senescence and might play a role as modulators ofpapain-like CPs (Dhanushkodi, Matthew, McManus, & Dijkwel, 2018).
Increasing data show that the symbiotic establishment relies on the sup-pression of plant immunity (Liang et al. 2013; Yu et al. 2018). Transcrip-tomic studies have revealed transient elicitation of legume plant defenserelated genes upon rhizobium inoculation (Libault et al. 2010; Lohar et al.2006; Pucciariello et al., 2009), followed by their suppressions in the subse-quent symbiotic stages (Aslam et al. 2008). Besides being known as anending stage of organ development, nodule senescence is also proposed toinvolve a reversion of the plant innate immunity (Puppo et al. 2005). CPshave also been shown to be involved in salicylic acid induced plant defenseto cope with microbial pathogens (M€ueller, Ziemann, Treitschke, Aßmann,& Doehlemann, 2013; Shindo, Misas-Villamil, H€orger, Song, & van derHoorn, 2012; Van der Linde et al., 2012). Thus, we could hypothesizethat activation of CPs during nodule senescence might be linked toincreasing plant defense responses linked to the declining nodule N2 fixationcapacity.
Genes encoding components of the SCF (skp1-cullin-F-Box protein)complex, also known as the E3 ubiquitin ligase complex, and genes encod-ing 26S proteasome AAA-ATPase subunits and the ubiquitin carrier protein4 are up-regulated during M. truncatula developmental nodule senescence(De Michele et al., 2009; Perez-Guerra et al., 2010; Van de Velde et al.,2006). This suggests that the 26S proteasome pathway may be acting duringnodule senescence.
3. Transcriptomic characterization of nodulesenescence
To get insights into transcriptional regulations associated with symbi-otic nodule senescence, several transcriptomic approaches were performed

Legume nodule senescence 199
initially onM. truncatula nodules. First, a cDNA-AFLP (Amplified FragmentLength Polymorphism) technique was applied on developmental senescentnodules (Van de Velde et al., 2006). A second approach used microarrays onnodules treated for 8h with phosphinotricin, an inhibitor of GS activity andtherefore affecting N2 fixation (Seabra et al., 2012). More recently, anRNAseq was performed on nodules exposed to nitrate for 4 h or 28 h(Cabeza et al., 2014). Similar studies have been also performed in other le-gumes species thanM. truncatula, such as in L. japonicus using microarrays andin soybean using RNAseq, at different time-points during nodule aging(Chungopast et al., 2014; Yuan et al., 2017).
Among the differentially expressed genes identified, the downregulationof specific N2 fixation associated genes, such as Lb, late nodulins and NCRpeptide encoding genes attests the inhibition of the N2 fixation at themolecular level (Cabeza et al., 2014; Seabra et al., 2012; Van de Veldeet al., 2006). Genes related to energy production, such as mitochondrialATP synthase subunit encoding genes, or amino-acid biogenesis encodinggenes, were accordingly repressed (Cabeza et al., 2014; Seabra et al.,2012). An upregulation of genes encoding proteins involved in macromol-ecule degradation, such as the CPs previously mentioned, and nutrientremobilization (phosphate, amino-acid, ion transporters and ABC [ATP-Binding Cassette] transporters) was observed, whereas ammonium, potas-sium and most of the sulfate transporters were down-regulated (Cabezaet al., 2014; Van de Velde et al., 2006). Genes associated with redox stressresponses such as catalases, oxidoreductases, hydrolases, peroxidases, ester-ases, glutathione transferases were also upregulated, showing the relevanceof redox regulations during nodule senescence (Seabra et al., 2012; Vande Velde et al., 2006), as previously described.
In order to distinguish at the molecular level environmentally induced-senescence from developmental senescence, selected genes up-regulatedduring a M. truncatula developmental nodule senescence were tested byqRT-PCR in nodules exposed to darkness for 3e5 days (Perez-Guerraet al., 2010; Van de Velde et al., 2006). Only 50% of the genes selected asup-regulated during developmental senescence were up-regulated duringa dark-induced nodule senescence. Genes not regulated in the dark-inducednodule senescence were notably related to the 26S proteasome-mediatedprotein degradation (Perez-Guerra et al., 2010), suggesting either a less orga-nized, or more probably an accelerated cellular degradation in dark-inducedconditions. To refine the spatio-temporal expression profiles during nodulesenescence, M. truncatula nodules were sectioned into apical versus basal

200 Théophile Kazmierczak et al.
zones and pea nodules were microdissected (Serova et al., 2017; Van deVelde et al., 2006). In this way, theM. truncatula or pea ATB2 genes, encod-ing a bZIP (basic leucine ZIPper) transcription factor (TF), were identified asreaching a peak in fully senescent nodules (Van de Velde et al., 2006;D’haeseleer et al., 2010; Serova et al., 2017).
To evaluate if genes were specifically regulated during the senescence ofnodule organs, a transcriptomic analysis was performed using a cDNA-AFLPapproach in M. truncatula senescent leaves (De Michele et al., 2009). Theoverlap between nodule and leaf senescence associated genes was relativelylow (23%). When excluding N-metabolism related genes, a higher degree ofoverlap was nevertheless observed in the early phase, including specific MAPkinases, ubiquitin cycle components, phosphatases, glutathione transferases,and transcription factors; as well as in the late phase, i.e. several CPs andcomponents of the 26S proteasome (Van de Velde et al., 2006; De Micheleet al., 2009).
Based on transcriptomic analyses previously described, transcription fac-tors (TF) encoding genes differentially regulated during nodule senescencewere identified, mainly from MYB (MYeloBlastosis), bHLH (basic Helix-Loop-Helix), and NAC (NAM ATAF CUC) families (Van de Veldeet al., 2006; Seabra et al., 2012; Cabeza et al., 2014; Chungopast et al.;2014; Yuan et al., 2017). Among those, only few TFs were selected forfurther functional analyses, with the aim of identifying regulatory pathwayscontrolling the transition between N2 fixation and nodule senescence, eitherduring aging or in response to environmental stress conditions. The NACTF family was previously largely associated with leaf senescence based ontranscriptome and genetic studies performed in Arabidopsis (Breeze et al.,2011), and is also enriched in nodule senescence transcriptomes. Based onRNAseq or microarray experiments, approximately one-third of NACgenes expressed in nodules are induced by nitrate (16/59 in Cabeza et al.2014; and 21/57 in Benedito et al., 2008) whereas over 30 out of 90NAC genes are upregulated during Arabidopsis leaf senescence (Breezeet al., 2011). Expression of the M. truncatula TF MtNAC969, closely relatedto the well-known Arabidopsis leaf senescence associated gene AtNAP(NAC-LIKE ACTIVATED BY AP3/PI), is induced by nitrate and associ-ated with the central region of symbiotic nodules including the N2-fixationzone (de Zelicourt et al., 2012). In addition, both AtNAP and MtNAC969genes are induced during Arabidopsis and M. truncatula leaf senescence,respectively (De Michele et al., 2009; Balazadeh et al., 2010). An RNAisilencing of the MtNAC969 TF promotes nodule senescence (Table 1),

Legume nodule senescence 201
based on the increased expression of CP genes used as senescence markers,concomitantly to an accumulation of amyloplasts suggestive of an alterednodule metabolism (de Zelicourt et al., 2012). Expression of the M. trunca-tula MtATB2 bZIP TF is increased during nodule aging senescence, as wellas by sucrose but not by a nitrate treatment (Van de Velde et al., 2006;D’haeseleer et al., 2010). Despite this transcriptional regulation pattern,ectopic expression of MtATB2 reduced the number of nodules formedbut no senescence phenotype could be observed (D’haeseleer et al.,2010). This highlights that an increased expression in senescent nodules isnot a sufficient criteria to identify genes functionally linked to nodule senes-cence. AsMtATB2 is expressed in the peripheral region of nodules contain-ing the vascular bundles and not in the N2 fixation or senescence centralzones, and because of its regulation by sucrose and not by nitrate, this sug-gests that this TF might sense carbon sources availability and regulate remo-bilization processes linked to senescence (D’haeseleer et al., 2010). Finally,no TF was yet directly linked to the upregulation of genes encoding CPsor other cell degradation related enzymes, or to the downregulation of Lbor N-assimilation metabolic enzymes encoding genes, or to componentsof redox pathways. Similarly, a symbiotic bacterial contribution to the regu-lation of nodule senescence associated genes has not been explored even ifsome toxin/antitoxin bacterial systems, such as the vapBC-5 (Table 1),have been showed to alter nodule senescence progression (Olahet al.,2001; Lipuma et al., 2014).
4. Concluding remarks - future prospects
Senescence affecting the legume-rhizobia symbiotic nodule organduring aging or in response to environmental conditions remains poorlydocumented. The best functionally characterized senescence mechanismsare related to the redox dependent inhibition of the N2 fixation and assim-ilation. Physiological and molecular approaches show that nodule senes-cence is followed by a decrease in the level and/or reduction state of themain cellular antioxidants ASC and GSH, and by an increase in the produc-tion of ROS and RNS, such as H2O2 and NO. This results in an increase ofoxidized/denatured proteins and subsequently in an inhibition of N, carbonand energy metabolisms. In addition from being harmful to cellular pro-cesses, both ROS and NO production are also important components ofthe signaling network used by legumes and their symbionts to communicate

202 Théophile Kazmierczak et al.
at the various stages of the symbiotic interaction. The evolution of efficientROS/NO degrading systems has most likely enabled plant cells and bacte-roids inside nodules to overcome ROS and NO toxicity. Diverse bacterialand plant systems contribute to the control of NO and ROS levels innodules. Only when these systems are overcome by NO and ROS, thesemolecules can act as signals to trigger senescence (Puppo et al., 2013). Alter-natively, senescence could be a consequence of NO-induced modificationsof proteins essential for N2 fixation. Further studies are needed to link themodifications of these proteins by NO or ROS to nodule senescence.
An increased proteolysis and the induction of plant vacuolar proteasessuch as CPs and VPEs during senescence are also well documented, andused as a hallmark to follow the senescence progression. Identifying howthese nodule specific proteases are regulated and what are their targets re-mains a challenge to understand better nodule senescence. Both 26S and20S proteasome proteolytic pathways are additionally induced duringnodule senescence, and their contribution should be established. Finally,the nature of bacterial proteases as well as their contribution to the degrada-tion of bacteroid proteins remains to be better documented.
Large-scale transcriptomic analyses from the last decade allowed identi-fying new candidate genes that may act in the senescence processes. Theseapproaches were however focused on the plant transcriptome, and it wouldbe interesting to analyze in parallel the transcriptional response of symbioticbacteria to determine if they could contribute to the regulation of nodulesenescence progression. In the model legumeM. truncatula, partially overlap-ping plant regulatory pathways were identified between environmental,developmental and leaf senescence. This suggests that a core of commongenes is conserved between the senescence of different organs, but alsothat a subset of genes may specifically regulate senescence in nodules, whichmay be notably related to the symbiotic and metabolic features of this organhosting numerous N2 fixing bacteria. Such putative plant-microbe genenetwork interactions remain however to be investigated. Only few regula-tory genes directly regulating nodule senescence have been yet functionallycharacterized, such as TFs and hormone-related genes. A future prospectwould be to integrate how the diversity of bacteria hosted inside the nod-ules, beyond rhizobia, could control senescence progression. Indeed, prom-ising recent data (Rodrigues, Silveira, Bonifacio, & Figueiredo, 2013)reported that other soil bacteria than rhizobia might indeed modify thenodule lifespan, as cowpea nodules co-inoculated with Bradyrhizobium sp.and Paenibacillus graminis or Paenibacillus durus plant growth-promoting

Legume nodule senescence 203
bacteria present an increased N availability and delayed nodule senescence.Finally, an applied perspective of studying nodule senescence would be, asachieved with “staygreen” leaves, to engineer “staypink” nodules thatwould have maintained, and even improved, capacities to fix N2 underenvironmental stress conditions.
AcknowledgmentsThe three laboratories involved (IPS2, ISA and LIPM) are funded by a common “AgenceNationale de la Recherche” (ANR) project (STAYPINK ANR-15-CE20-0005). TheIPS2 institute benefits from the support of the LabEx “Saclay Plant Sciences” (SPS) and ofthe Lidex “Plant Phenotyping Pipeline” (3P). T.K. was supported by Université Paris-Saclayand ANR fellowships. The ISA was supported by the Institut National de la RechercheAgronomique, the Center National de la Recherche Scientifique, the University ofNiceeSophia-Antipolis, and the French Government (National Research Agency, ANR)through the LABEX SIGNALIFE program (ANR-11-LABX-0028-01). L.Y. was supportedby a doctoral fellowship from China Scholarship Council (CSC). The LIPM was supportedby the French Laboratory of Excellence project TULIP (ANR-10-LABX-41). E.M. wassupported by the National Institute for Applied Science (INSA-Toulouse).
ReferencesAlesandrini, F., Mathis, R., Van de Sype, G., Hérouart, D., & Puppo, A. (2003). Possible
roles for a cysteine protease and hydrogen peroxide in soybean nodule developmentand senescence. New Phytologist, 158, 131e138.
Alloing, G., Mandon, K., Boncompagni, E., Montrichard, F., & Frendo, P. (2018). Involve-ment of glutaredoxin and thioredoxin systems in the nitrogen-fixing symbiosis betweenlegumes and rhizobia. Antioxidants (Basel), 7, 182.
Alunni, B., Kevei, Z., Redondo-Nieto, M., Kondorosi, A., Mergaert, P., & Kondorosi, E.(2007). Genomic organization and evolutionary insights on GRP and NCR genes,two large nodule-specific gene families in Medicago truncatula. Molecular Plant-Microbe In-teractions, 20, 1138e1148.
Appleby, C. A. (1992). The origin and functions of haemoglobin in plants. Science Progress,76, 365e398.
Aranjuelo, I., Tcherkez, G., Molero, G., Gilard, F., Avice, J. C., & Nogués, S. (2013).Concerted changes in N and C primary metabolism in alfalfa (Medicago sativa) under wa-ter restriction. Journal of Experimental Botany, 64, 885e897.
Arthikala, M. K., Sanchez-Lopez, R., Nava, N., Santana, O., Cardenas, L., & Quinto, C.(2014). RbohB, a Phaseolus vulgaris NADPH oxidase gene, enhances symbiosomenumber, bacteroid size, and nitrogen fixation in nodules and impairs mycorrhizalcolonization. New Phytologist, 202, 886e900.
Aslam, S. N., Newman, M.-A., Erbs, G., Morrissey, K. L., Chinchilla, D., Boller, T., et al.(2008). Bacterial polysaccharides suppress induced innate immunity by calciumchelation. Current Biology, 18, 1078e1083.
Asp, T., Bowra, S., Borg, S., & Holm, P. B. (2004). Cloning and characterisation of threegroups of cysteine protease genes expressed in the senescing zone of white clover(Trifolium repens) nodules. Plant Science, 167, 825e837.
Astier, J., & Lindermayr, C. (2012). Nitric oxide-dependent posttranslational modification inplants: An update. International Journal of Molecular Sciences, 13, 15193e15208.

204 Théophile Kazmierczak et al.
Balazadeh, S., Siddiqui, H., Allu, A. D., Matallana-Ramirez, L. P., Caldana, C., Mehrnia, M.,et al. (2010). A gene regulatory network controlled by the NAC transcription factorANAC092/AtNAC2/ORE1 during salt-promoted senescence. The Plant Journal, 62,250e264.
Balestrasse, K., Gallego, S., & Tomaro, M. (2004). Cadmium-induced senescence in nodulesof soybean (Glycine max L.) plants. Plant and Soil, 262, 373e381.
Banba, M., Siddique, A. B., Kouchi, H., Izui, K., & Hata, S. (2001). Lotus japonicus formsearly senescent root nodules with Rhizobium etli. Molecular Plant-Microbe Interactions, 14,173e180.
Becana, M., Matamoros, M. A., Udvardi, M., & Dalton, D. A. (2010). Recent insights intoantioxidant defences of legume root nodules. New Phytologist, 188, 960e976.
Benchabane, M., Schluter, U., Vorster, J., Goulet, M. C., & Michaud, D. (2010). Plantcystatins. Biochimie, 92, 1657e1666.
Benedito, V. A., Torres-Jerez, I., Murray, J. D., Andriankaja, A., Allen, S., Kakar, K., et al.(2008). A gene expression atlas of the model legumeMedicago truncatula. The Plant Journal,55, 504e513.
Berger, A., Boscari, A., Frendo, P., & Brouquisse, R. (2019). Nitric oxide signaling, meta-bolism and toxicity in nitrogen-fixing symbiosis. Journal of Experimental Botany, 70.https://doi.org/10.1093/jxb/erz159 (in press).
Bethlenfalvay, G. J., & Phillips, D. A. (1977). Ontogenetic interactions between photosyn-thesis and symbiotic nitrogen fixation in legumes. Plant Physiology, 60, 419e421.
Blanquet, P., Silva, L., Catrice, O., Bruand, C., Carvalho, H., & Meilhoc, E. (2015).Sinorhizobium meliloti controls nitric oxide-mediated post-translational modification ofa Medicago truncatula nodule protein. Molecular Plant-Microbe Interactions, 28, 1353e1363.
Boivin, S., Kazmierczak, T., Brault, M., Wen, J., Gamas, P., et al. (2016). Different cytokininhistidine kinase receptors regulate nodule initiation as well as later nodule developmentalstages in Medicago truncatula. Plant, Cell and Environment, 39, 2198e2209.
Boscari, A., Del Giudice, J., Ferrarini, A., Venturini, L., Zaffini, A.-L., Delledonne, M., et al.(2013a). Expression dynamics of the Medicago truncatula transcriptome during the symbi-otic interaction with Sinorhizobium meliloti: Which role for nitric oxide? Plant physiology,161, 425e439.
Boscari, A., Meilhoc, E., Castella, C., Bruand, C., Puppo, A., & Brouquisse, R. (2013b).Which role for nitric oxide in symbiotic N2-fixing nodules: Toxic by-product or usefulsignaling/metabolic intermediate? Frontiers of Plant Science, 4, 384.
Breeze, E., Harrison, E., McHattie, S., Hughes, L., Hickman, R., Hill, C., et al. (2011).High-resolution temporal profiling of transcripts during Arabidopsis leaf senescencereveals a distinct chronology of processes and regulation. The Plant Cell, 23, 873e894.
Brouquisse, R. (2019). Multifaceted roles of nitric oxide in plants. Journal of Experimental Bot-any, 70. https://doi.org/10.1093/jxb/erz352 (in press).
Bruand, C., & Meilhoc, E. (2019). Nitric oxide in plants: Pro- or anti-senescence. Journal ofExperimental Botany, 70. https://doi.org/10.1093/jxb/erz117 (in press).
Buono, R. A., Hudecek, R., & Nowack, M. K. (2019). Plant proteases during develop-mental programmed cell death. Journal of Experimental Botany, 70, 2097e2112.
Cabeza, R., Koester, B., Liese, R., Lingner, A., Baumgarten, V., Dirks, J., et al. (2014). AnRNA sequencing transcriptome analysis reveals novel insights into molecular aspects ofthe nitrate impact on the nodule activity of Medicago truncatula. Plant Physiology, 164,400e411.
Calvo-Begueria, L., Rubio, M. C., Martinez, J. I., Perez-Rontome, C., Delgado, M. J.,Bedmar, E. J., et al. (2018). Redefining nitric oxide production in legume nodulesthrough complementary insights from electron paramagnetic resonance spectroscopyand specific fluorescent probes. Journal of Experimental Botany, 69, 3703e3714.

Legume nodule senescence 205
Cam, Y., Pierre, O., Boncompagni, E., Herouart, D., Meilhoc, E., & Bruand, C. (2012).Nitric oxide (NO): A key player in the senescence of Medicago truncatula root nodules.New Phytologist, 196, 548e560.
Chen, Y., Chen, W., Li, X., Jiang, H., Wu, P., Xia, K., et al. (2014). Knockdown ofLjIPT3 influences nodule development in Lotus japonicus. Plant and Cell Physiology,55, 183e193.
Christenhunsz, M. J., & Byng, J. W. (2016). The number of known plants species in theworld and its annual increase. Phytotaxa, 261, 201e217.
Chungopast, S., Duangkhet, M., Tajima, S., Ma, J. F., & Nomura, M. (2017). Iron-inducednitric oxide leads to an increase in the expression of ferritin during the senescence of Lotusjaponicus nodules. J Plant Physiology, 208, 40e46.
Chungopast, S., Hirakawa, H., Sato, S., Handa, Y., Saito, K., Kawaguchi, M., et al. (2014).Transcriptomic profiles of nodule senescence in Lotus japonicus and Mesorhizobium lotisymbiosis. Plant Biotechnology, 31, 345e349.
Cilliers, M., van Wyk, S. G., van Heerden, P. D. R., Kunert, K. J., & Vorster, B. J. (2018).Identification and changes of the drought-induced cysteine protease transcriptome insoybean (Glycine max) root nodules. Environmental and Experimental Botany, 148, 59e69.
Coba de la Pe~na, T., Carcamo, C. B., Almonacid, L., Zaballos, A., Lucas, M. M.,Balomenos, D., et al. (2008). A salt stress-responsive cytokinin receptor homologueisolated from Medicago sativa nodules. Planta, 227, 769e779.
Coba de la Pe~na, T., Redondo, F. J., Manrique, E., Lucas, M. M., & Pueyo, J. J. (2010).Nitrogen fixation persists under conditions of salt stress in transgenic Medicago truncatulaplants expressing a cyanobacterial flavodoxin. Plant Biotechnology Journal, 8, 954e965.
Curioni, P. M., Hartwig, U. A., Nosberger, J., & Schuller, K. A. (1999). Glycolytic flux isadjusted to nitrogenase activity in nodules of detopped and argon-treated alfalfa plants.Plant Physiology, 119, 445e454.
Czernic, P., Gully, D., Cartieaux, F., Moulin, L., Guefrachi, I., Patrel, D., et al. (2015).Convergent evolution of endosymbiont differentiation in Dalbergioid and IRLClegumes mediated by nodule-specific cysteine-rich peptides. Plant Physiology, 169,1254e1265.
D’haeseleer, K., De Keyser, A., Goormachtig, S., & Holsters, M. (2010). Transcription factorMtATB2: About nodulation, sucrose and senescence. Plant and Cell Physiology, 51,1416e1424.
Dalton, D. A., Boniface, C., Turner, Z., Lindahl, A., Kim, H. J., Jelinek, L., et al. (2009).Physiological roles of glutathione s-transferases in soybean root nodules. Plant Physiology,150, 521e530.
Dalton, D. A., Russell, S. A., Hanus, F. J., Pascoe, G. A., & Evans, H. J. (1986). Enzymaticreactions of ascorbate & glutathione that prevent peroxide damage in soybean rootnodules. Proceedings of the National Academy of Sciences, 83, 3811e3815.
Damiani, I., Pauly, N., Puppo, A., Brouquisse, R., & Boscari, A. (2016). Reactive oxygenspecies and nitric oxide control early steps of the legume - rhizobium symbioticinteraction. Frontiers of Plant Science, 7, 454.
Day, D. A., Poole, P. S., Tyerman, S. D., & Rosendahl, L. (2001). Ammonia and amino acidtransport across symbiotic membranes in nitrogen-fixing legume nodules. Cellular andMolecular Life Sciences: CMLS, 58, 61e71.
De Michele, R., Formentin, E., Todesco, M., Toppo, S., Carimi, F., Zottini, M., et al.(2009). Transcriptome analysis of Medicago truncatula leaf senescence: Similarities and dif-ferences in metabolic and transcriptional regulations as compared with Arabidopsis,nodule senescence and nitric oxide signalling. New Phytologist, 181, 563e575.
de Zelicourt, A., Diet, A., Marion, J., Laffont, C., Ariel, F., Moison, M., et al. (2012). Dualinvolvement of a Medicago truncatula NAC transcription factor in root abiotic stressresponse and symbiotic nodule senescence. The Plant Journal, 70, 220e230.

206 Théophile Kazmierczak et al.
Dhanushkodi, R., Matthew, C., McManus, M. T., & Dijkwel, P. P. (2018). Drought-induced senescence of Medicago truncatula nodules involves serpin and ferritin to controlproteolytic activity and iron levels. New Phytologist, 220, 196e208.
Dupont, L., Alloing, G., Pierre, O., El Msehli, S., Hopkins, J., Hérouart, D., et al. (2012).The legume root nodule: From symbiotic nitrogen fixation to senescence. InT. Nagata (Ed.), Senescence (Vol. 8, pp. 137e168). Shanghai, China: InTech.
El Msehli, S., Lambert, A., Baldacci-Cresp, F., Hopkins, J., Boncompagni, E., Smiti, S. A.,et al. (2011). Crucial role of (homo)glutathione in nitrogen fixation in Medicago trun-catula nodules. New Phytologist, 192, 496e506.
Escuredo, P. R., Minchin, F. R., Gogorcena, Y., Iturbe-Ormaetxe, I., Klucas, R. V., &Becana, M. (1996). Involvement of activated oxygen in nitrate-induced senescence ofpea root nodules. Plant Physiology, 110, 1187e1195.
Farkas, A., Maroti, G., Durgo, H., Gyorgypal, Z., Lima, R. M., Medzihradszky, K. F., et al.(2014).Medicago truncatula symbiotic peptide NCR247 contributes to bacteroid differen-tiation through multiple mechanisms. Proceedings of the National Academy of Sciences, 111,5183e5188.
Ferguson, B. J., Indrasumunar, A., Hayashi, S., Lin, M.-H., Lin, Y.-H., Reid, D. E., et al.(2010). Molecular analysis of legume nodule development and autoregulation. Journalof Integrative Plant Biology, 52, 61e76.
Ferrarini, A., de Stefano, M., Baudouin, E., Pucciariello, C., Polverari, A., Puppo, A., et al.(2008). Expression of Medicago truncatula genes responsive to nitric oxide in pathogenicand symbiotic conditions. Molecular Plant-Microbe Interactions, 21, 781e790.
Frendo, P., Matamoros, M. A., Alloing, G., & Becana, M. (2013). Thiol-based redoxsignaling in the nitrogen-fixing symbiosis. Frontiers of Plant Science, 4, 376.
Fukudome, M., Watanabe, E., Osuki, K. I., Imaizumi, R., Aoki, T., Becana, M., et al.(2019). Stably-transformed Lotus japonicus plants overexpressing phytoglobin LjGlb1-1show decreased nitric oxide levels in roots and nodules as well as delayed nodulesenescence. Plant and Cell Physiology, 60, 816e825.
Gil-Quintana, E., Lyon, D., Staudinger, C., Wienkoop, S., & Gonzalez, E. M. (2015).Med-icago truncatula and Glycine max: Different drought tolerance and similar local response ofthe root nodule proteome. Journal of Proteome Research, 14, 5240e5251.
Gogorcena, Y., Gordon, A. J., Escuredo, P. R., Minchin, F. R., Witty, J. F., Moran, J. F.,et al. (1997). N2 fixation, carbon metabolism, and oxidative damage in nodules ofdark-stressed common bean plants. Plant Physiology, 113, 1193e1201.
Gonzalez, E. M., Galvez, L., & Arrese-Igor, C. (2001). Abscisic acid induces a decline in ni-trogen fixation that involves leghaemoglobin, but is independent of sucrose synthaseactivity. Journal of Experimental Botany, 52, 285e293.
Harrison, J., Jamet, A., Muglia, C. I., Van de Sype, G., Aguilar, O. M., Puppo, A., et al.(2005). Glutathione plays a fundamental role in growth and symbiotic capacity of Sino-rhizobium meliloti. Journal of Bacteriology, 187, 168e174.
Hernandez-Jiménez, M. J., Mercedes Lucas, M., & de Felipe, M. R. (2002). Antioxidantdefence and damage in senescing lupin nodules. Plant Physiology and Biochemistry, 40,645e657.
Hichri, I., Boscari, A., Castella, C., Rovere, M., Puppo, A., & Brouquisse, R. (2015). Nitricoxide: A multifaceted regulator of the nitrogen-fixing symbiosis. Journal of ExperimentalBotany, 66, 2877e2887.
Hichri, I., Meilhoc, E., Boscari, A., Bruand, C., Frendo, P., & Brouquisse, R. (2016). Nitricoxide: Jack-of-All-Trades of the nitrogen-fixing symbiosis? Advances in Botanical Research,77, 193e218.
Holzmeister, C., Gaupels, F., Geerlof, A., Sarioglu, H., Sattler, M., Durner, J., et al. (2015).Differential inhibition of Arabidopsis superoxide dismutases by peroxynitrite-mediatedtyrosine nitration. Journal of Experimental Botany, 66, 989e999.

Legume nodule senescence 207
Horchani, F., Prevot, M., Boscari, A., Evangelisti, E., Meilhoc, E., Bruand, C., et al. (2011).Both plant and bacterial nitrate reductases contribute to nitric oxide production inMedicago truncatula nitrogen-fixing nodules. Plant Physiology, 155, 1023e1036.
Kardailsky, I. V., & Brewin, N. J. (1996). Expression of cysteine protease genes in pea noduledevelopment and senescence. Molecular Plant-Microbe Interactions, 9, 689e695.
Klucas, R. V. (1974). Studies on soybean nodule senescence. Plant Physiology, 54, 612e616.Kondorosi, E., Mergaert, P., & Kereszt, A. (2013). A paradigm for endosymbiotic life: Cell
differentiation of rhizobium bacteria provoked by host plant factors. Annual Review ofMicrobiology, 67, 611e628.
Kunert, K. J., van Wyk, S. G., Cullis, C. A., Vorster, B. J., & Foyer, C. H. (2015). Potentialuse of phytocystatins in crop improvement, with a particular focus on legumes. Journal ofExperimental Botany, 66, 3559e3570.
Lahiri, K., Chattopadhyay, S., Chatterjee, S., & Ghosh, B. (1993). Biochemical changes innodules of Vigna mungo (L.) during vegetative and reproductive stages of plant growthin the field. Annals of Botany, 71, 485e488.
Lamouche, F., Bonadé-Bottino, N., Peter Mergaert, P., & Alunni, B. (2019). Symbiotic ef-ficiency of spherical and elongated bacteroids in the aeschynomene-bradyrhizobiumsymbiosis. Frontiers of Plant Science, 10, 377.
Lawn, R. J., & Brun, W. A. (1974). Symbiotic nitrogen fixation in soybeans. I. Effect ofphotosynthetic source-sink manipulations1. Crop Science, 14, 11e16.
Lee, I. C., Hong, S. W., Whang, S. S., Lim, P. O., Nam, H. G., & Koo, J. C. (2011).Age-dependent action of an ABA-inducible receptor kinase, RPK1, as a positive regu-lator of senescence in Arabidopsis leaves. Plant & Cell Physiology, 52, 651e662.
Lehtovaara, P., & Perttil€a, U. (1978). Bile-pigment formation from different leghaemoglo-bins. Methine-bridge specificity of coupled oxidation. Biochemical Journal, 176, 359e364.
Li, Y., Zhou, L., Li, Y., Chen, D., Tan, X., Lei, L., et al. (2008). A nodule-specific plantcysteine proteinase, AsNODF32, is involved in nodule senescence and nitrogen fixationactivity of the green manure legume Astragalus sinicus. New Phytologist, 180, 185e192.
Liang, Y., Cao, Y., Tanaka, K., Thibivilliers, S., Wan, J., Choi, J., et al. (2013). Nonlegumesrespond to rhizobial Nod factors by suppressing the innate immune response. Science,341, 1384e1387.
Libault, M., Farmer, A., Brechenmacher, L., Drnevich, J., Langley, R. J., Bilgin, D. D., et al.(2010). Complete transcriptome of the soybean root hair cell, a single-cell model, and itsalteration in response to Bradyrhizobium japonicum infection. Plant Physiology, 152,541e552.
Liese, R., Schulze, J., & Cabeza, R. A. (2017). Nitrate application or P deficiency induce adecline in Medicago truncatula N2-fixation by similar changes in the noduletranscriptome. Scientific Reports, 7, 46264.
Lipuma, J., Cinege, G., Bodogai, M., Olah, B., Kiers, A., Endre, G., et al. (2014). A vapBC-type toxin-antitoxin module of Sinorhizobium meliloti influences symbiotic efficiencyand nodule senescence of Medicago sativa. Environmental Microbiology, 16, 3714e3729.
Lohar, D. P., Sharopova, N., Endre, G., Penuela, S., Samac, D., Town, C., et al. (2006).Transcript analysis of early nodulation events in Medicago truncatula. Plant Physiology,140, 221e234.
Lopez-Otín, C., Blasco, M. A., Partridge, L., Serrano, M., & Kroemer, G. (2013). The hall-marks of aging. Cell, 153, 1194e1217.
Loscos, J., Matamoros, M. A., & Becana, M. (2008). Ascorbate and homoglutathione meta-bolism in common bean nodules under stress conditions and during natural senescence.Plant Physiology, 146, 1282e1292.
Marino, D., Andrio, E., Danchin, E. G., Oger, E., Gucciardo, S., Lambert, A., et al. (2011). AMedicago truncatula NADPH oxidase is involved in symbiotic nodule functioning. NewPhytologist, 189, 580e592.

208 Théophile Kazmierczak et al.
Marquez-Garcia, B., Shaw, D., Cooper, J. W., Karpinska, B., Quain, M. D.,Makgopa, E. M., et al. (2015). Redox markers for drought-induced nodule senescence,a process occurring after drought-induced senescence of the lowest leaves in soybean(Glycine max). Annals of Botany, 116, 497e510.
Martínez, M., Cambra, I., Gonzalez-Melendi, P., Santamaria, M. E., & Diaz, I. (2012). C1Acysteine-proteases and their inhibitors in plants. Physiologia Plantarum, 145, 85e94.
Matamoros, M. A., Baird, L. M., Escuredo, P. R., Dalton, D. A., Minchin, F. R., Iturbe-Ormaetxe, I., et al. (1999). Stress-induced legume root nodule senescence. Physiological,biochemical, and structural alterations. Plant Physiology, 121, 97e112.
Matamoros, M. A., Fernandez-Garcia, N., Wienkoop, S., Loscos, J., Saiz, A., & Becana, M.(2013). Mitochondria are an early target of oxidative modifications in senescing legumenodules. New Phytologist, 197, 873e885.
Meakin, G. E., Bueno, E., Jepson, B., Bedmar, E. J., Richardson, D. J., & Delgado, M. J.(2007). The contribution of bacteroidal nitrate and nitrite reduction to the formationof nitrosylleghaemoglobin complexes in soybean root nodules. Microbiology, 153,411e419.
Meilhoc, E., Blanquet, P., Cam, Y., & Bruand, C. (2013). Control of NO level in rhizo-bium-legume root nodules: Not only a plant globin story. Plant Signaling & Behavior,8. e25923.
Meilhoc, E., Cam, Y., Skapski, A., & Bruand, C. (2010). The response to nitric oxide of thenitrogen-fixing symbiont Sinorhizobium meliloti. Molecular Plant-Microbe Interactions, 23,748e759.
Melo, P. M., Silva, L. S., Ribeiro, I., Seabra, A. R., & Carvalho, H. G. (2011). Glutaminesynthetase is a molecular target of nitric oxide in root nodules of Medicago truncatulaand is regulated by tyrosine nitration. Plant Physiology, 157, 1505e1517.
Mergaert, P., Nikovics, K., Kelemen, Z., Maunoury, N., Vaubert, D., Kondorosi, A., et al.(2003). A novel family inMedicago truncatula consisting of more than 300 nodule-specificgenes coding for small, secreted polypeptides with conserved cysteine motifs. Plant Phys-iology, 132, 161e173.
Mergaert, P., Uchiumi, T., Alunni, B., Evanno, G., Cheron, A., Catrice, O., et al. (2006).Eukaryotic control on bacterial cell cycle and differentiation in the Rhizobiumelegume symbiosis. Proceedings of the National Academy of Sciences, 103, 5230e5235.
Mesa, S., Reutimann, L., Fischer, H. M., &Hennecke, H. (2009). Posttranslational control oftranscription factor FixK2, a key regulator for the Bradyrhizobium japonicum-soybeansymbiosis. Proceedings of the National Academy of Sciences, 106, 21860e21865.
Montiel, J., Arthikala, M. K., Cardenas, L., & Quinto, C. (2016). Legume NADPH oxidaseshave crucial roles at different stages of nodulation. International Journal of Molecular Sciences,17, 680.
M€ueller, A. N., Ziemann, S., Treitschke, S., Aßmann, D., & Doehlemann, G. (2013).Compatibility in the Ustilago maydisemaize interaction requires inhibition of hostcysteine proteases by the fungal effector Pit2. PLoS Pathogens, 9. e1003177.
Muglia, C., Comai, G., Spegazzini, E., Riccillo, P. M., & Aguilar, O. M. (2008). Glutathioneproduced by Rhizobium tropici is important to prevent early senescence in common beannodules. FEMS Microbiology Letters, 286, 191e198.
M€uller, J., Boller, T., & Wiemken, A. (2001). Trehalose becomes the most abundantnon-structural carbohydrate during senescence of soybean nodules. Journal of Experi-mental Botany, 52, 943e947.
Naito, Y., Fujie, M., Usami, S., Murooka, Y., & Yamada, T. (2000). The involvement of acysteine proteinase in the nodule development in Chinese milk vetch infected withMesorhizobium huakuii subsp. rengei. Plant Physiology, 124, 1087e1096.
Navascués, J., Perez-Rontome, C., Gay, M., Marcos, M., Yang, F., Walker, F. A., et al.(2012). Leghemoglobin green derivatives with nitrated hemes evidence production of

Legume nodule senescence 209
highly reactive nitrogen species during aging of legume nodules. Proceedings of the NationalAcademy of Sciences, 109, 2660e2665.
Oda, K. (2012). New families of carboxyl peptidases: Serine-carboxyl peptidases and gluta-mic peptidases. Journal of Biochemistry, 151, 13e25.
Olah, B., Kiss, E., Gy€orgypal, Z., Borzi, J., Cinege, G., Csanadi, G., et al. (2001). Mutation inthe ntrR gene, a member of the vap gene family, increases the symbiotic efficiency ofSinorhizobium meliloti. Molecular Plant-Microbe Interactions, 14, 887e894.
Oldroyd, G. E. (2013). Speak, friend, and enter: Signalling systems that promote beneficialsymbiotic associations in plants. Nature Reviews Microbiology, 11, 252.
Oono, R., Schmitt, I., Sprent, J. I., & Denison, R. F. (2010). Multiple evolutionary origins oflegume traits leading to extreme rhizobial differentiation.New Phytologist, 187, 508e520.
Ott, T., van Dongen, J. T., Gu, C., Krusell, L., Desbrosses, G., Vigeolas, H., et al. (2005).Symbiotic leghemoglobins are crucial for nitrogen fixation in legume root nodules butnot for general plant growth and development. Current Biology, 15, 531e535.
Palma, F., Lopez-Gomez, M., Tejera, N. A., & Lluch, C. (2014). Involvement of abscisic acidin the response ofMedicago sativa plants in symbiosis with Sinorhizobium meliloti to salinity.Plant Science, 223, 16e24.
Perez-Guerra, J. C., Coussens, G., De Keyser, A., De Rycke, R., De Bodt, S., Van DeVelde, W., et al. (2010). Comparison of developmental and stress-induced nodule senes-cence in Medicago truncatula. Plant Physiology, 152, 1574e1584.
Peters, J. W., Fisher, K., & Dean, D. R. (1995). Nitrogenase structure and function: Abiochemical-genetic perspective. Annual Review of Microbiology, 49, 335e366.
Pierre, O., Engler, G., Hopkins, J., Brau, F., Boncompagni, E., & Herouart, D. (2013). Peri-bacteroid space acidification: A marker of mature bacteroid functioning inMedicago trun-catula nodules. Plant, Cell and Environment, 36, 2059e2070.
Pierre, O., Hopkins, J., Combier, M., Baldacci, F., Engler, G., Brouquisse, R., et al. (2014).Involvement of papain and legumain proteinase in the senescence process of Medicagotruncatula nodules. New Phytologist, 202, 849e863.
Pladys, D., Dimitrijevic, L., & Rigaud, J. (1991). Localization of a protease in protoplast prep-arations of infected cells of French bean nodules. Plant Physiology, 97, 1174e1180.
Pladys, D., & Vance, C. P. (1993). Proteolysis during development and senescence of effec-tive and plant gene-controlled ineffective alfalfa nodules. Plant Physiology, 103, 379e384.
Preisig, O., Zufferey, R., Thony-Meyer, L., Appleby, C. A., & Hennecke, H. (1996). Ahigh-affinity cbb3-type cytochrome oxidase terminates the symbiosis-specific respiratorychain of Bradyrhizobium japonicum. Journal of Bacteriology, 178, 1532e1538.
Pucciariello, C., Innocenti, G., Van de Velde, W., Lambert, A., Hopkins, J., Clément, M.,et al. (2009). (Homo)glutathione depletion modulates host gene expression during thesymbiotic interaction between Medicago truncatula and Sinorhizobium meliloti. Plant Physi-ology, 151, 1186e1196.
Puppo, A., Groten, K., Bastian, F., Carzaniga, R., Soussi, M., Lucas, M. M., et al. (2005).Legume nodule senescence: Roles for redox and hormone signalling in the orchestrationof the natural aging process. New Phytologist, 165, 683e701.
Puppo, A., Pauly, N., Boscari, A., Mandon, K., & Brouquisse, R. (2013). Hydrogen peroxideand nitric oxide: Key regulators of the legume-rhizobium and mycorrhizal symbioses.Antioxidants and Redox Signaling, 18, 2202e2219.
Quain, M. D., Makgopa, M. E., Cooper, J. W., Kunert, K. J., & Foyer, C. H. (2015). Ectopicphytocystatin expression increases nodule numbers and influences the responses ofsoybean (Glycine max) to nitrogen deficiency. Phytochemistry, 112, 179e187.
Rawlings, N. D., Barrett, A. J., & Bateman, A. (2010). MEROPS: The peptidase database.Nucleic Acids Research, 38, D227eD233.

210 Théophile Kazmierczak et al.
Redondo, F. J., Coba de la Pena, T., Lucas, M. M., & Pueyo, J. J. (2012). Alfalfa noduleselicited by a flavodoxin-overexpressing Ensifer meliloti strain display nitrogen-fixing activ-ity with enhanced tolerance to salinity stress. Planta, 236, 1687e1700.
Redondo, F. J., de la Pe~na, T. C., Morcillo, C. N., Lucas, M. M., & Pueyo, J. J. (2009).Overexpression of flavodoxin in bacteroids induces changes in antioxidant metabolismleading to delayed senescence and starch accumulation in alfalfa root nodules. Plant Phys-iology, 149, 1166e1178.
Reid, D. E., Heckmann, A. B., Novak, O., Kelly, S., & Stougaard, J. (2016). CYTOKININOXIDASE/DEHYDROGENASE3 maintains cytokinin homeostasis during root andnodule development in Lotus japonicus. Plant Physiology, 170, 1060e1074.
Rodrigues, A. C., Silveira, J. A. G., Bonifacio, A., & Figueiredo, M. D. V. B. (2013). Meta-bolism of nitrogen and carbon: Optimization of biological nitrogen fixation and cowpeadevelopment. Soil Biology and Biochemistry, 67, 226e234.
Roux, B., Rodde, N., Jardinaud, M. F., Timmers, T., Sauviac, L., Cottret, L., et al. (2014).An integrated analysis of plant and bacterial gene expression in symbiotic root nodulesusing laser-capture microdissection coupled to RNA sequencing. The Plant Journal, 77,817e837.
Sainz, M., Calvo-Begueria, L., Perez-Rontome, C., Wienkoop, S., Abian, J., Staudinger, C.,et al. (2015). Leghemoglobin is nitrated in functional legume nodules in a tyrosineresidue within the heme cavity by a nitrite/peroxide-dependent mechanism. The PlantJournal, 81, 723e735.
Sanchez, C., Gates, A. J., Meakin, G. E., Uchiumi, T., Girard, L., Richardson, D. J., et al.(2010). Production of nitric oxide and nitrosylleghemoglobin complexes in soybeannodules in response to flooding. Molecular Plant-Microbe Interactions, 23, 702e711.
Seabra, A. R., Pereira, P. A., Becker, J. D., & Carvalho, H. G. (2012). Inhibition of glutaminesynthetase by phosphinothricin leads to transcriptome reprograming in root nodules ofMedicago truncatula. Molecular Plant-Microbe Interactions, 25, 976e992.
Serova, T., & Tsyganov, V. (2014). Symbiotic nodule senescence in legumes: Molecular-ge-netic and cellular aspects (review). Agricultural Biology, 5, 3e15.
Serova, T. A., Tikhonovich, I. A., & Tsyganov, V. E. (2017). Analysis of nodule senescencein pea (Pisum sativum L.) using laser microdissection, real-time PCR, and ACCimmunolocalization. Journal of Plant Physiology, 212, 29e44.
Shindo, T., Misas-Villamil, J. C., H€orger, A. C., Song, J., & van der Hoorn, R. A. (2012). Arole in immunity for Arabidopsis cysteine protease RD21, the ortholog of the tomatoimmune protease C14. PLoS One, 7. e29317.
Smirnoff, N., & Arnaud, D. (2019). Hydrogen peroxide metabolism and functions in plants.New Phytologist, 221, 1197e1214.
Smith, S. E., & Smith, F. A. (2011). Roles of arbuscular mycorrhizas in plant nutrition andgrowth: New paradigms from cellular to ecosystem scales. Annual Review of Plant Biology,62, 227e250.
Soupene, E., Foussard, M., Boistard, P., Truchet, G., & Batut, J. (1995). Oxygen as a keydevelopmental regulator of Rhizobium meliloti N2-fixation gene expression withinthe alfalfa root nodule. Proceedings of the National Academy of Sciences, 92, 3759e3763.
Streeter, J. G. (1981). Seasonal distribution of carbohydrates in nodules and stem exudatefrom field-grown soya bean plants. Annals of Botany, 48, 441e450.
Swaraj, K., & Bishnoi, N. R. (1996). Physiological and biochemical basis of nodule senes-cence in legumes: A review. Plant Physiology and Biochemistry, 23, 105e116.
Tang, G., Li, N., Liu, Y., Yu, L., Yan, J., & Luo, L. (2018). Sinorhizobium meliloti glutathionereductase is required for both redox homeostasis and symbiosis. Applied and EnvironmentalMicrobiology, 84. e01937-17.

Legume nodule senescence 211
Tang, G., Xing, S., Wang, S., Yu, L., Li, X., Staehelin, C., et al. (2017). Regulation ofcysteine residues in LsrB proteins from Sinorhizobium meliloti under free-living conditionsand symbiotic oxidative stress. Environmental Microbiology, 19, 5130e5145.
Timmers, A. C. J., Auriac, M. C., & Truchet, G. (1999). Refined analysis of early symbioticsteps of the Rhizobium-Medicago interaction in relationship with microtubular cyto-skeleton rearrangements. Development, 126, 3617e3628.
Tittabutr, P., Sripakdi, S., Boonkerd, N., Tanthanuch, W., Minamisawa, K., &Teaumroong, N. (2015). Possible role of 1-aminocyclopropane-1-carboxylate (ACC)deaminase activity of Sinorhizobium sp. BL3 on symbiosis with mung bean and determi-nate nodule senescence. Microbes and Environments, 30, 310e320.
Tjepkema, J. D., & Yocum, C. S. (1974). Measurement of oxygen partial pressure withinsoybean nodules by oxygen microelectrodes. Planta, 119, 351e360.
Trinchant, J. C., & Rigaud, J. (1982). Nitrite and nitric oxide as inhibitors of nitrogenasefrom soybean bacteroids. Applied and Environmental Microbiology, 44, 1385e1388.
Van Der Heijden, M. G., De Bruin, S., Luckerhoff, L., Van Logtestijn, R. S., & Schlaeppi, K.(2016). A widespread plant-fungal-bacterial symbiosis promotes plant biodiversity, plantnutrition and seedling recruitment. The ISME Journal, 10, 389.
Van Wyk, S. G., Du Plessis, M., Cullis, C. A., Kunert, K. J., & Vorster, B. J. (2014). Cysteineprotease and cystatin expression and activity during soybean nodule development andsenescence. BMC Plant Biology, 14, 294.
Vance, C. P., Heichel, G. H., Barnes, D. K., Bryan, J. W., & Johnson, L. E. (1979). Nitrogenfixation, nodule development, and vegetative regrowth of alfalfa (Medicago sativa L.)following harvest. Plant Physiology, 64, 1e8.
Van der Hoorn, R. A. (2008). Plant proteases: From phenotypes to molecular mechanisms.Annual Review of Plant Biology, 59, 191e223.
Van der Linde, K., Hemetsberger, C., Kastner, C., Kaschani, F., van der Hoorn, R. A.,Kumlehn, J., et al. (2012). A maize cystatin suppresses host immunity by inhibiting apo-plastic cysteine proteases. The Plant Cell, 24, 1285e1300.
Van de Velde, W., Guerra, J. C., De Keyser, A., De Rycke, R., Rombauts, S.,Maunoury, N., et al. (2006). Aging in legume symbiosis. A molecular view on nodulesenescence in Medicago truncatula. Plant Physiology, 141, 711e720.
Van de Velde, W., Zehirov, G., Szatmari, A., Debreczeny, M., Ishihara, H., Kevei, Z., et al.(2010). Plant peptides govern terminal differentiation of bacteria in symbiosis. Science,327, 1122e1126.
Vasse, J., de Billy, F., Camut, S., & Truchet, G. (1990). Correlation between ultrastructuraldifferentiation of bacteroids and nitrogen fixation in alfalfa nodules. Journal of Bacteriology,172, 4295e4306.
Venkateshwaran, M., Volkening, J. D., Sussman, M. R., & Ané, J. M. (2013). Symbiosis andthe social network of higher plants. Current Opinion in Plant Biology, 16, 118e127.
Wang, W., Shi, J., Xie, Q., Jiang, Y., Yu, N., & Wang, E. (2017). Nutrient exchange andregulation in arbuscular mycorrhizal symbiosis. Molecular Plant, 10, 1147e1158.
Xu, J., Li, S.-L., & Luo, L. (2012). Effects of engineered Sinorhizobium meliloti on cytokininsynthesis and tolerance of alfalfa to extreme drought stress. Applied and EnvironmentalMicrobiology, 78, 8056e8061.
Yu, H., Xiao, A., Dong, R., Fan, Y., Zhang, X., Liu, C., et al. (2018). Suppression of innateimmunity mediated by the CDPK-Rboh complex is required for rhizobial colonizationin Medicago truncatula nodules. New Phytologist, 220, 425e434.
Yuan, S. L., Li, R., Chen, H. F., Zhang, C. J., Chen, L. M., Hao, Q. N., et al. (2017). RNA-Seq analysis of nodule development at five different developmental stages of soybean(Glycine max) inoculated with Bradyrhizobium japonicum strain 113-2. Scientific Reports,7, 42248.

212 Théophile Kazmierczak et al.
Yun, B. W., Feechan, A., Yin, M., Saidi, N. B., Le Bihan, T., Yu, M., et al. (2011). S-nitro-sation of NADPH oxidase regulates cell death in plant immunity.Nature, 478, 264e268.
Zwack, P. J., & Rashotte, A. M. (2013). Cytokinin inhibition of leaf senescence. PlantSignaling & Behavior, 8. e24737.
Zwack, P. J., Robinson, B. R., Risley, M. G., & Rashotte, A. M. (2013). Cytokinin responsefactor 6 negatively regulates leaf senescence and is induced in response to cytokinin andnumerous abiotic stresses. Plant and Cell Physiology, 54, 971e981.

GENERAL INTRODUCTION
94
Proteases play vital roles during nodule senescence in leguminous plants
During nodule senescence, proteolytic activities are strongly increased and associated with the final degradation of bacteroid and plant host cells (Van de Velde et al., 2006; Perez Guerra et al., 2010; Kazmierczak et al., 2020). Hence, proteases are found to be the hallmarks of nodule senescence (Van de Velde et al., 2006; Perez Guerra et al., 2010). Proteases are firstly targeting leghaemoglobin, as consequences of declined N fixation, then rupture of bacteriods, eventually eliminating the microbial partner of the symbiotic associations (Pladys & Vance, 1993). Cysteine proteases have been identified in the cytoplasm of infected nodule cells and their activity appears to increase markedly during senescence (Malik et al., 1981; Pladys et al., 1991). However, interestingly, the knockdown of the CP inhibitor gene MtSERPIN6 accelerated nodule aging (Dhanushkodi et al., 2018).
Transcript-profiling of developmental nodule senescence was revealed in the legume M. truncatula via cDNA-amplified fragment length polymorphism (AFLP) (Van de Velde et al., 2006). Functional clustering of 508 gene tags to corresponding genes demonstrated the transition from carbon sink to general source status of the nodule, as well as the activation of a wide variety of defence and stress responses. Three chronologically upregulating and one downregulating cluster were identified with a nodule senescence-associated differential expression pattern (Van de Velde et al., 2006), indicating consecutive cascades of transcriptional regulation. A selective expression pattern shows this clear transcriptional regulation waves of regulators and CPs in Table 5. The cluster 1 genes were found also upregulating in heathy tissues, indicating an early signal transduction of nodule senescence initiation. It is not surprising that most of the transcriptional regulators are found in this group (Table 5), e.g. MYB, AP2/ERF, bHLH, NAC/NAM and bZIP TFs. Among the TFs, ATB2 is involved in balancing carbohydrate demand and supply in A. thaliana (Rook et al., 1998; Wobbes, 2004). In M. truncatula, ectopic expression of MtATB2 diminished nodule formation and affected root growth (D'Haeseleer et al., 2010). MYB4 is suggested to function as repressors in the flavonoid pathway, together with its homologs MYB7 and MYB32 and further interactive with the bHLH transcription factors TT8, GL3 and EGL3 (Wang et al., 2020). Interestingly, in this early cluster, a tag is present that is homologous to genes encoding abscisic acid-insensitive/viviparous related transcription factors and is characterized by an AP2/ERF and B3 DNA-binding domain (Kagaya et al., 1999). The subsequent cluster 2 and 3 are involved in nodule senescence stage I and II (Table 4), corresponding to bacteroid
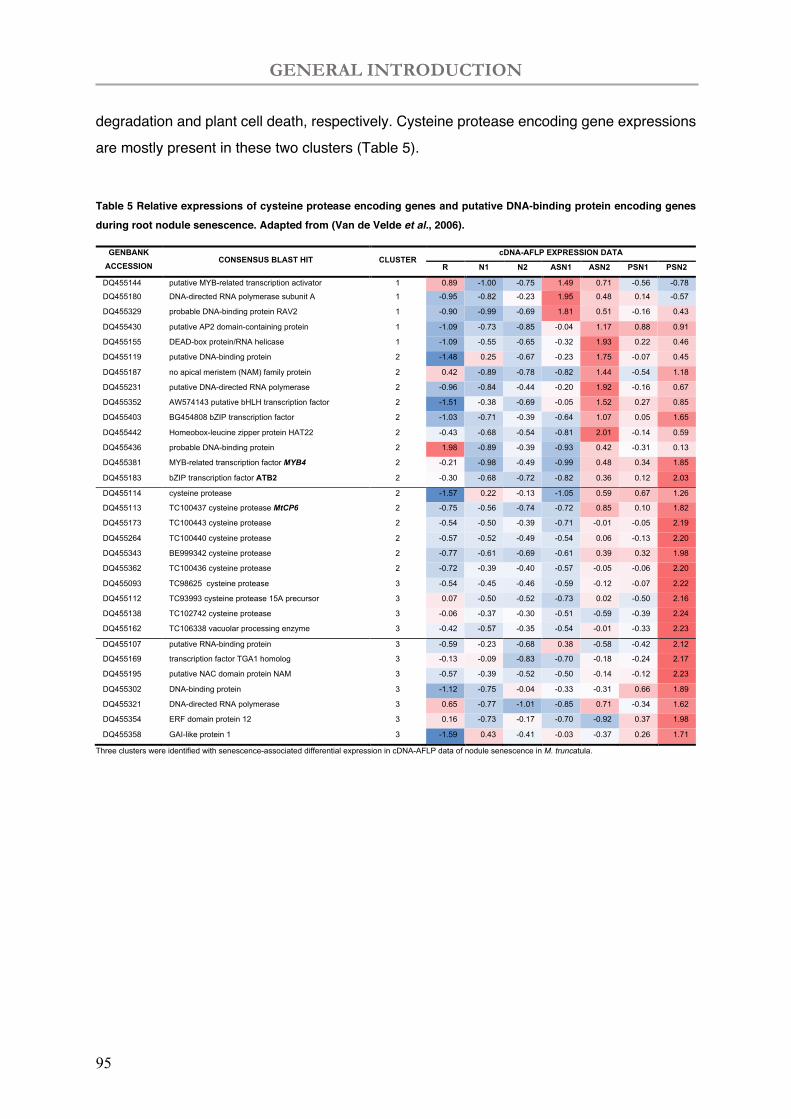
GENERAL INTRODUCTION
95
degradation and plant cell death, respectively. Cysteine protease encoding gene expressions are mostly present in these two clusters (Table 5).
Table 5 Relative expressions of cysteine protease encoding genes and putative DNA-binding protein encoding genes during root nodule senescence. Adapted from (Van de Velde et al., 2006).
GENBANK ACCESSION
NUMBER
CONSENSUS BLAST HIT CLUSTER cDNA-AFLP EXPRESSION DATA
R N1 N2 ASN1 ASN2 PSN1 PSN2
DQ455144 putative MYB-related transcription activator 1 0.89 -1.00 -0.75 1.49 0.71 -0.56 -0.78
DQ455180 DNA-directed RNA polymerase subunit A 1 -0.95 -0.82 -0.23 1.95 0.48 0.14 -0.57
DQ455329 probable DNA-binding protein RAV2 1 -0.90 -0.99 -0.69 1.81 0.51 -0.16 0.43
DQ455430 putative AP2 domain-containing protein 1 -1.09 -0.73 -0.85 -0.04 1.17 0.88 0.91
DQ455155 DEAD-box protein/RNA helicase 1 -1.09 -0.55 -0.65 -0.32 1.93 0.22 0.46
DQ455119 putative DNA-binding protein 2 -1.48 0.25 -0.67 -0.23 1.75 -0.07 0.45
DQ455187 no apical meristem (NAM) family protein 2 0.42 -0.89 -0.78 -0.82 1.44 -0.54 1.18
DQ455231 putative DNA-directed RNA polymerase 2 -0.96 -0.84 -0.44 -0.20 1.92 -0.16 0.67
DQ455352 AW574143 putative bHLH transcription factor 2 -1.51 -0.38 -0.69 -0.05 1.52 0.27 0.85
DQ455403 BG454808 bZIP transcription factor 2 -1.03 -0.71 -0.39 -0.64 1.07 0.05 1.65
DQ455442 Homeobox-leucine zipper protein HAT22 2 -0.43 -0.68 -0.54 -0.81 2.01 -0.14 0.59
DQ455436 probable DNA-binding protein 2 1.98 -0.89 -0.39 -0.93 0.42 -0.31 0.13
DQ455381 MYB-related transcription factor MYB4 2 -0.21 -0.98 -0.49 -0.99 0.48 0.34 1.85
DQ455183 bZIP transcription factor ATB2 2 -0.30 -0.68 -0.72 -0.82 0.36 0.12 2.03
DQ455114 cysteine protease 2 -1.57 0.22 -0.13 -1.05 0.59 0.67 1.26
DQ455113 TC100437 cysteine protease MtCP6 2 -0.75 -0.56 -0.74 -0.72 0.85 0.10 1.82
DQ455173 TC100443 cysteine protease 2 -0.54 -0.50 -0.39 -0.71 -0.01 -0.05 2.19
DQ455264 TC100440 cysteine protease 2 -0.57 -0.52 -0.49 -0.54 0.06 -0.13 2.20
DQ455343 BE999342 cysteine protease 2 -0.77 -0.61 -0.69 -0.61 0.39 0.32 1.98
DQ455362 TC100436 cysteine protease 2 -0.72 -0.39 -0.40 -0.57 -0.05 -0.06 2.20
DQ455093 TC98625 cysteine protease 3 -0.54 -0.45 -0.46 -0.59 -0.12 -0.07 2.22
DQ455112 TC93993 cysteine protease 15A precursor 3 0.07 -0.50 -0.52 -0.73 0.02 -0.50 2.16
DQ455138 TC102742 cysteine protease 3 -0.06 -0.37 -0.30 -0.51 -0.59 -0.39 2.24
DQ455162 TC106338 vacuolar processing enzyme 3 -0.42 -0.57 -0.35 -0.54 -0.01 -0.33 2.23
DQ455107 putative RNA-binding protein 3 -0.59 -0.23 -0.68 0.38 -0.58 -0.42 2.12
DQ455169 transcription factor TGA1 homolog 3 -0.13 -0.09 -0.83 -0.70 -0.18 -0.24 2.17
DQ455195 putative NAC domain protein NAM 3 -0.57 -0.39 -0.52 -0.50 -0.14 -0.12 2.23
DQ455302 DNA-binding protein 3 -1.12 -0.75 -0.04 -0.33 -0.31 0.66 1.89
DQ455321 DNA-directed RNA polymerase 3 0.65 -0.77 -1.01 -0.85 0.71 -0.34 1.62
DQ455354 ERF domain protein 12 3 0.16 -0.73 -0.17 -0.70 -0.92 0.37 1.98
DQ455358 GAI-like protein 1 3 -1.59 0.43 -0.41 -0.03 -0.37 0.26 1.71
Three clusters were identified with senescence-associated differential expression in cDNA-AFLP data of nodule senescence in M. truncatula.
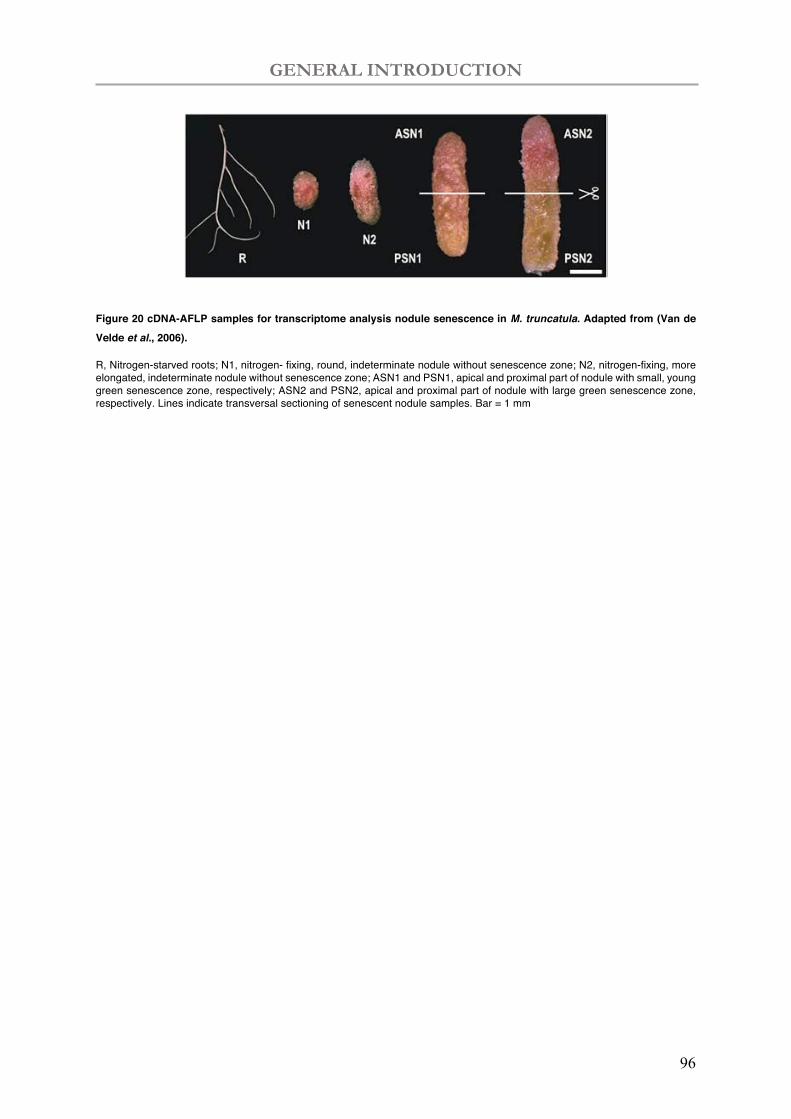
GENERAL INTRODUCTION
96
Figure 20 cDNA-AFLP samples for transcriptome analysis nodule senescence in M. truncatula. Adapted from (Van de Velde et al., 2006).
R, Nitrogen-starved roots; N1, nitrogen- fixing, round, indeterminate nodule without senescence zone; N2, nitrogen-fixing, more elongated, indeterminate nodule without senescence zone; ASN1 and PSN1, apical and proximal part of nodule with small, young green senescence zone, respectively; ASN2 and PSN2, apical and proximal part of nodule with large green senescence zone, respectively. Lines indicate transversal sectioning of senescent nodule samples. Bar = 1 mm

GENERAL INTRODUCTION
97

GENERAL INTRODUCTION
98
The involvement of proteases in the nodule senescence is more described in
the following review (Yang, Li et al., 2020).
Yang L, Syska C, Garcia I, Frendo P, Boncompagni E 2020. Involvement of proteases during nodule senescence in leguminous plants. In: Bruijn Fd ed. The Model Legume Medicago truncatula, 683-693.

GENERAL INTRODUCTION
99

CHAPTER 9.4.1
Involvement of proteases during nodule senescencein leguminous plants
Li Yang, Camille Syska, Isabelle Garcia, Pierre Frendo, and Eric BoncompagniUniversite Cote d’Azur, INRA, CNRS, UMR 1355-7254 ISA, Sophia Antipolis 06900, France
9.4.1.1 Introduction
In plants, as in all other living organisms, proteins are submittedto permanent turnover. The content of a given protein dependson its rate of both synthesis and degradation. The degradation ofdamaged, misfolded, and potentially harmful proteins is associ-ated with the activity of proteolytic enzymes. Beside this regu-latory role in the protein turnover, proteolysis plays numerousroles in the regulation of cellular metabolism, as well as in plantdevelopment and defense (Thomas 2013).
Proteases can be divided into several classes depending on thehydrolysis site of the target protein and on the chemical nature ofthe catalytic site of the protease. Exopeptidases which act on thetermini of (poly)peptide chains can be classified as aminopepti-dases when acting on the N-terminus of the protein and as car-boxypeptidases when acting on the C-terminus of the protein.Endopeptidases, also called proteinases, act on the interior ofpolypeptides. Various amino acids are present in the catalytic siteof proteases. In serine, threonine, and cysteine proteases (CPs),the hydroxyl or sulfhydryl group of the active-site amino acidperforms a nucleophilic attack on the peptide bond to hydrolyzeit. Acidic proteases (aspartic and glutamic proteases) and met-alloproteases, which rely on divalent cations for activity suchas zinc, cobalt, and manganese, activate a water molecule toact as a nucleophile to attack the peptide carbonyl group. Theplant genomes contain around 2% of proteases. According to theMEROPS database (http://merops.sanger.ac.uk; Rawlings et al.2016), the peptidases family contains 847 known and putativemembers in Arabidopsis thaliana and 799 members in Med-icago truncatula of a total of around 40 000 genes per genome.The protease families diverge so far that relationships cannotalways be surely demonstrated by comparison of their primarystructures. Similarity of the three-dimensional structures allowsto classify proteases in clans and families even when primarysequence is not comparable.
The proteases play crucial roles in plant physiology. Theselective breakdown of regulatory proteins via the ubiqui-tin/proteasome pathway controls many aspects of the plantdevelopment. As examples, the ubiquitin-proteasome pathwayregulates auxin, gibberellin, and jasmonate pathways (Wagerand Browse 2012; Salehin et al. 2015) and light perception viathe degradation of regulatory proteins (Huang et al. 2014). Thispathway also controls key aspects of the plant innate immu-nity and general defense (Marino et al. 2012). Proteases are alsoresponsible for the post-translational modifications of proteinsat highly specific sites. This cleavage of proteins is involved inthe maturation of enzymes after the subcellular targeting to vac-uoles, extracellular space, and organelles such as plastids andmitochondria (Kmiec et al. 2014). Proteolysis is also necessaryfor protein assembly and to control the activity of proteins andpeptides, such as systemin (Pearce 2011).
The vacuole is a major compartment involved in proteindegradation. This organelle which can represent 90% of the plantcell volume, harbors in addition to other lytic enzymes, themajority of proteolytic activities (Matile and Martinoia 1982).Numerous proteases have been identified in the vacuole (Carteret al. 2004). The vacuolar proteolysis is involved in many cellu-lar processes such as microautophagy and macroautophagy, reg-ulating multiple aspects of plant physiology from mobilizationof the storage proteins during seed germination, to initiation ofcell death and senescence. The degradation of storage proteinsduring plant germination allows the remobilization of nitrogenfor the formation of new proteins during plantlet development(Tan-Wilson and Wilson 2012).
Protease activity also plays a crucial role in the plant’sadaptation to abiotic stresses. In such cases, proteolysis playsa housekeeping role by removing abnormal, inactivated, ordenatured proteins, the accumulation of which could betoxic for the cell. In general, the plant adaptation to abioticstresses goes together with an increased expression of proteases
The Model Legume Medicago truncatula, First Edition. Edited by Frans J. de Bruijn.© 2020 John Wiley & Sons, Inc. Published 2020 by John Wiley & Sons, Inc.
683

684 Section 9.4: Bacteroid senescence
(Velasco-Arroyo et al. 2016; Mishra and Richa Grover 2016).The stress-induced alteration of photosynthesis also inducesa strong modification in cellular metabolism (Gururani et al.2015). In these stress conditions, the proteases are involvedin the redistribution of amino acids for the synthesis of newenzymes and the basis of carbon backbones for energy produc-tion through respiration (Roberts et al. 2012). Proteases are alsoinvolved in plant–pathogen interactions. Apoplastic proteaseshave been shown to be involved in the plant defense againstmicrobial pathogens such as Pseudomonas syringae (Xia et al.2004). Cysteine proteases have also been shown to be requiredfor plant resistance against the necrotrophic fungus Botrytiscinerea (Shindo et al. 2012) and Ralstonia solanacearum (Zhanget al. 2014a). Cathepsins are also involved in the basal immunityof plants against P. syringae (Ramirez et al. 2013). Similarly, asubtilase protease has been shown to be implicated in defenseagainst bacteria and oomycetes (Ramirez et al. 2013).
Plant proteases participate to the programmed cell death(PCD) associated with developmental processes and plantdefense against abiotic stresses and pathogen invasion (Sueldoand van der Hoorn 2017). In animal systems, the central coreexecution switch for apoptotic PCD is the activation of caspases(cysteine-containing aspartate-specific proteases). In plants, theexistence of caspase-like activity during PCD has been shownto regulate PCD execution (Sueldo and van der Hoorn 2017).Proteasome activity also participates in PCD (Han et al. 2012).Proteases are involved in PCD occurring in embryo suspen-sors (Suarez et al. 2004; Zhao et al. 2013), the degeneration ofthe tapetum during anther development (Zhang et al. 2014b),the inner integument shrinking during seed coat formation(Nakaune et al. 2005), and the development of tracheary ele-ments which constitute the xylem (Avci et al. 2008). Proteasesare also involved in PCD occurring during the hypersensi-tive response induced during plant defense against pathogenmicroorganisms and viruses (McLellan et al. 2009; Coll et al.2010; Chichkova et al. 2010). Likewise, proteases are involved inPCD occurring during abiotic stresses like oxidative and osmoticstresses (Chichkova et al. 2010).
Proteolysis is also associated with plant senescence, which isconsidered as another type of PCD. Senescence is a complex andcontrolled process, essential for (re)mobilization of nutrientsfrom old and stressed tissues to growing or sink organs (Shiet al. 2017; Have et al. 2017). It is connected to the degradationof the cellular structures and macromolecules to allow theformation and the relocation of transportable molecules. Leafsenescence has been particularly well studied (Lim et al. 2007;Otegui et al. 2005). It is a developmental process linked to theorgan’s aging, but it could also be modulated by biotic andabiotic stresses (Jajic et al. 2015; Reinbothe et al. 2009). Pro-teases are involved in senescence with a major role in nutrientrecycling, especially amino acids and nitrogen. Proteases areupregulated during the senescence process and nearly all pro-tease families have been associated with some aspects of plantsenescence (Diaz-Mendoza et al. 2016). They are located in
multiple cell compartments such as chloroplasts, mitochondria,vacuoles, and senescence associated vesicles (Diaz-Mendozaet al. 2016). In addition to their roles in nutrient mobilization,proteases have been shown to regulate the senescence process.Metacaspases, which are cysteine proteases, are positive regula-tors of cell death in the context of pathogen-triggered PCD anda negative regulator of the plant senescence (Coll et al. 2014;Minina et al. 2013).
Within leguminous plant nodules, cysteine proteinases areengaged in response to microbial invasion, protein turnover fortissue remodeling, metabolic homeostasis and nutrient reutiliza-tion, adaptation to environmental stresses, and leaf and nodulesenescence.
Biological nitrogen fixation (BNF) is the biological process bywhich atmospheric nitrogen (N2) is converted to ammonia byan enzyme called nitrogenase. It is the major source of nitrogenin the biosphere and as such has an important ecologicaland agronomical role, accounting for 65% of the nitrogenused in agriculture worldwide. The most important source offixed nitrogen is the symbiotic association between rhizobiaand legumes. The nodule is a de novo formed plant organ inwhich biological nitrogen fixation is achieved by the bacteria(see Chapter 6.1). The bacteria are surrounded by a plant cellmembrane, the peribacteroid membrane (PBM), the wholeforming an organelle-like structure called the symbiosomewhere bacteria differentiate into nitrogen-fixing bacteroids.Symbiosomes completely fill the cytoplasm of infected cells. Asthe bacteria differentiate, infected cells undergo enlargementcoupled to repeated endoreduplication cycles – genomic DNAreplication without mitosis and cytokinesis – and become largepolyploid cells housing thousands of bacteroids (Jones et al.2007; Kondorosi et al. 2000; see Section 9.3). The nodule func-tioning has many particularities, related to the plant–microbecrosstalk and to a metabolism which needs a high energy levelunder micro-oxic conditions. Whereas leaf senescence hasbeen extensively studied, root nodule senescence has been lesscharacterized. Nodule senescence also has specific features(Puppo et al. 2005). In this context, this review focuses on theroles of proteases in root nodule senescence.
9.4.1.2 Involvement of plant proteases inthe nodule senescence process
Developmental and induced nodule senescence are complexprocesses causing a strong decrease of N2 fixation and degra-dation of leghemoglobin (Lb) showing a typical green colorof senescing nodule tissue (Puppo et al. 2005; Lehtovaara andPerttila 1978). Plants keep in homoeostasis by the continuityof cell degradation and synthesis. However, when the rate ofdeterioration exceeds that of novel production, plants proceedto whole senescence. In roots, nodule senescence is accompa-nied with a series of physiological, biochemical, and structuralalterations. Nodule senescence has also been regarded as a
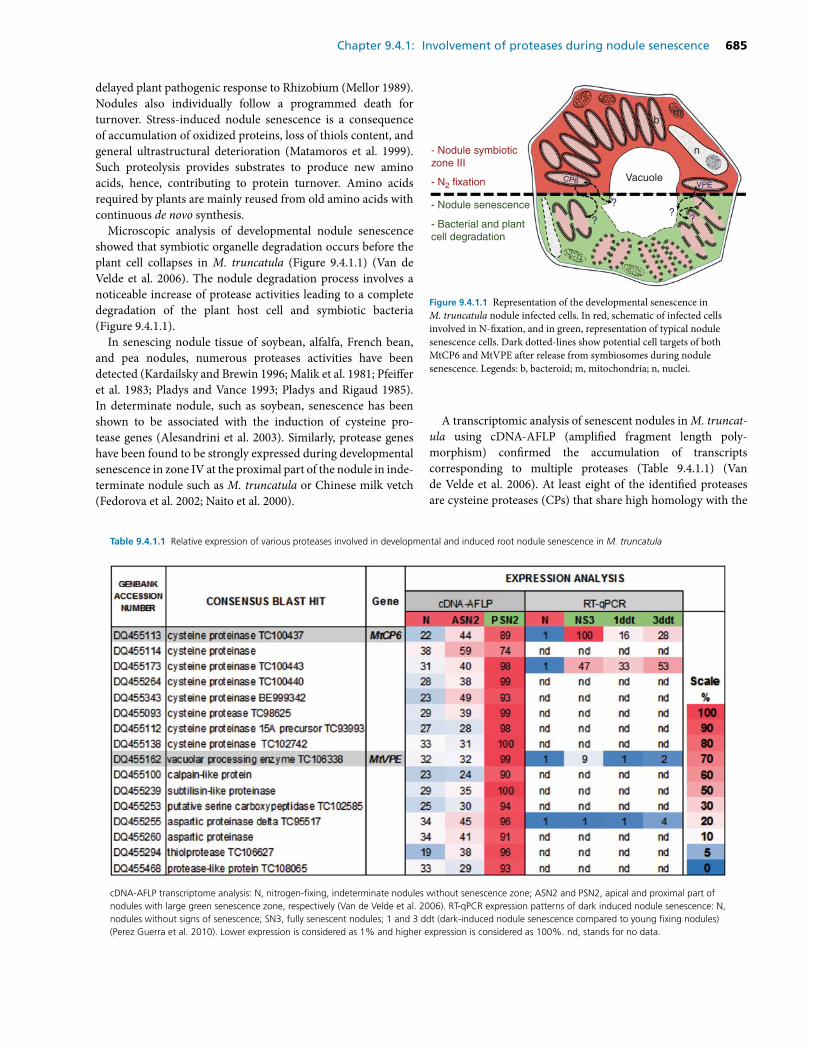
Chapter 9.4.1: Involvement of proteases during nodule senescence 685
delayed plant pathogenic response to Rhizobium (Mellor 1989).Nodules also individually follow a programmed death forturnover. Stress-induced nodule senescence is a consequenceof accumulation of oxidized proteins, loss of thiols content, andgeneral ultrastructural deterioration (Matamoros et al. 1999).Such proteolysis provides substrates to produce new aminoacids, hence, contributing to protein turnover. Amino acidsrequired by plants are mainly reused from old amino acids withcontinuous de novo synthesis.
Microscopic analysis of developmental nodule senescenceshowed that symbiotic organelle degradation occurs before theplant cell collapses in M. truncatula (Figure 9.4.1.1) (Van deVelde et al. 2006). The nodule degradation process involves anoticeable increase of protease activities leading to a completedegradation of the plant host cell and symbiotic bacteria(Figure 9.4.1.1).
In senescing nodule tissue of soybean, alfalfa, French bean,and pea nodules, numerous proteases activities have beendetected (Kardailsky and Brewin 1996; Malik et al. 1981; Pfeifferet al. 1983; Pladys and Vance 1993; Pladys and Rigaud 1985).In determinate nodule, such as soybean, senescence has beenshown to be associated with the induction of cysteine pro-tease genes (Alesandrini et al. 2003). Similarly, protease geneshave been found to be strongly expressed during developmentalsenescence in zone IV at the proximal part of the nodule in inde-terminate nodule such as M. truncatula or Chinese milk vetch(Fedorova et al. 2002; Naito et al. 2000).
- Nodule symbioticzone III
- N2 fixation
- Nodule senescence
- Bacterial and plantcell degradation
VacuoleVPE
??
??
CP6
n
bm
Figure 9.4.1.1 Representation of the developmental senescence inM. truncatula nodule infected cells. In red, schematic of infected cellsinvolved in N-fixation, and in green, representation of typical nodulesenescence cells. Dark dotted-lines show potential cell targets of bothMtCP6 and MtVPE after release from symbiosomes during nodulesenescence. Legends: b, bacteroid; m, mitochondria; n, nuclei.
A transcriptomic analysis of senescent nodules in M. truncat-ula using cDNA-AFLP (amplified fragment length poly-morphism) confirmed the accumulation of transcriptscorresponding to multiple proteases (Table 9.4.1.1) (Vande Velde et al. 2006). At least eight of the identified proteasesare cysteine proteases (CPs) that share high homology with the
Table 9.4.1.1 Relative expression of various proteases involved in developmental and induced root nodule senescence in M. truncatula
cDNA-AFLP transcriptome analysis: N, nitrogen-fixing, indeterminate nodules without senescence zone; ASN2 and PSN2, apical and proximal part ofnodules with large green senescence zone, respectively (Van de Velde et al. 2006). RT-qPCR expression patterns of dark induced nodule senescence: N,nodules without signs of senescence; SN3, fully senescent nodules; 1 and 3 ddt (dark-induced nodule senescence compared to young fixing nodules)(Perez Guerra et al. 2010). Lower expression is considered as 1% and higher expression is considered as 100%. nd, stands for no data.

686 Section 9.4: Bacteroid senescence
senescence-associated gene 12 (AtSAG12), one of the prominentmarkers of leaf senescence in A. thaliana (Lohman et al. 1994).AtSAG12 is a typical papain, a CP member of the peptidases(family C1A and clan CA classification from MEROPS database)typically found as lysosomal or secreted (Otegui et al. 2005;Richau et al. 2012). The overexpression of such CPs suggests animportant role of proteases in the regulation of developmentalnodule senescence. MtCP6 has been mainly studied as a rep-resentative CP from this group of papain in nodule senescence(Van de Velde et al. 2006; Perez Guerra et al. 2010; Pierre et al.2014). In M. truncatula, another cysteine protease, key markerof nodule senescence was found to be a Vacuolar ProcessingEnzyme (MtVPE), a protease belonging to legumain families(Table 9.4.1.1) (Sueldo and van der Hoorn 2017). MtVPE,a cysteine protease (member of family C13 from clan CDclassification from MEROPS database), is evolutionarily relatedto caspases and, found to be located in the vacuole (Pierre et al.2014; Hatsugai et al. 2015). Both MtCP6 and MtVPE have beenfound to be strongly upregulated in response to both develop-mental and environmentally induced nodule senescence such asdark treatment (ddt) (Perez Guerra et al. 2010) (Table 9.4.1.1).Interestingly, only at the onset of nodule senescence, vacuolarSNAREs (soluble N-ethylmaleimide-sensitive factor adaptorprotein receptors) proteins (SYP22 and VTI11) that are typicalmarkers of lytic vacuolar membrane identity are found tobe present on the peribacteroid membrane (Limpens et al.2009). The modification of membrane identity may allow thetargeting of vacuolar proteases to the symbiosomes. In thisscenario, the low pH present in the peribacteroid space wouldinduce the autoactivation of both CPs and VPEs (Pierre et al.2013).
In M. truncatula, five cysteine proteases have been iden-tified as specifically expressed in nodules and share a highdegree of homology with the MtCP6 family (Pierre et al.2014). A spatio-temporal analysis of MtCP6 and MtVPE expres-sion pattern in developmental nodule senescence revealed astrong expression associated to the transition region betweenthe N2-fixing (III in red) and senescence (IV in green) zones(Figure 9.4.1.1) (Pierre et al. 2014). During senescence inducedby three days of ammonitrate treatment, MtCP6 and MtVPEexpression was detected in the entire nodule and correlatedto the extension of the green senescence zone of the nodule(Figure 9.4.1.1).
Subcellular localization of both MtCP6 and MtVPE inM. truncatula senescent nodules has been associated to vacuolesin non-infected cells and to symbiosomes of infected cells, sug-gesting a role of these cysteine proteases in symbiosome degra-dation after the induction of proteases upon developmental orstress-induced senescence (Figure 9.4.1.1) (Pierre et al. 2014).Silencing of MtCP6 or MtVPE genes delayed nodule senescenceand extended the zone III ratio relative to zone IV with aconcomitant increase of 2.5- to 1.5-fold in nitrogen fixation forboth constructions, respectively (Pierre et al. 2014). Conversely,their ectopic expression in the nitrogen-fixing zone promoted
an extension of nodule senescence and a reduction of nodulelength for both constructions (Pierre et al. 2014), showing theinvolvement of these two proteases in the nodule senescenceprocess.
Similarly, cysteine protease genes are induced during nod-ule senescence in other leguminous plants. In soybean, 18 CPsand 6 VPEs were identified as transcriptionally upregulated dur-ing developmental nodule senescence (van Wyk et al. 2014). InTrifolium repens, five CP genes TrCP1, TrCP5, TrCP8, TrCP10,and TrCP12 have been identified as having an increased expres-sion mainly in the nodule senescence zone (Asp et al. 2004)(Table 9.4.1.2). In Astragalus sinicus the transcripts are mostlyaccumulated in senescence zone (IV), as well as in interzone(II-III) under developmental senescence. Interestingly, theexpression of a nodule-specific CP, Asnodf32, appears at 14 dayspost inoculation (dpi) and increases fivefold at 30 dpi (Naitoet al. 2000) (Table 9.4.1.2). The important role of Asnodf32 innodule senescence has been validated by the delayed root nod-ule senescence correlated to a significant extension of the nitro-gen fixation active period obtained by RNAi silencing of theAsnodf32 gene (Li et al. 2008). Furthermore, as in M. truncat-ula, Asnodf32 RNAi leads to the elongation of nodules sug-gesting a modification of the nodule development (Li et al.2008).
Interestingly, using laser microdissection, the analysis of nod-ule senescence in the mutant line E135F (sym13) of Pisumsativum L. showed the increase of mRNA abundance of two cys-teine proteases (PsCyp1 and PsCyp15a) (Table 9.4.1.2) and a thiolprotease (PsTPP) (Serova et al. 2017).
Recently, a large collection of M. truncatula mutants has beenconstructed using ethyl methanesulfonate (EMS) and fast neu-tron mutagenized or Tnt1-insertion, leading to numerous Nod+
Fix− mutants with phenotypes showing signs of early senescence(Domonkos et al. 2013; Pislariu et al. 2012). This collection ofmutants will represent a huge interest for deciphering the molec-ular pathways involving proteases in nodule senescence.
9.4.1.3 Role of cystatin in the nodulesenescence process
To date, regulation of the cysteine proteases in legumes is poorlydocumented. However, recent studies show their interactions invivo with cystatins, which are cysteine protease inhibitors lim-iting proteolytic processes in a reversible manner (for review,Benchabane et al. 2010; Martinez et al. 2012; van Wyk et al.2014; Diaz-Mendoza et al. 2016; Yuan et al. 2016). Cystatinsmainly inhibit protease enzymes belonging to peptidase familiesC1 (papain family) and C13 (legumain family).
In Medicago sativa, expression of the cystatin MsCys1 is devel-opmentally regulated in stems and leaves but not induced byvarious stress signals. Recombinant MsCys1 shows a stronginhibitory effect on alfalfa leaf CPs activities (Rivard et al.2007). However to date, no study concerning the biological
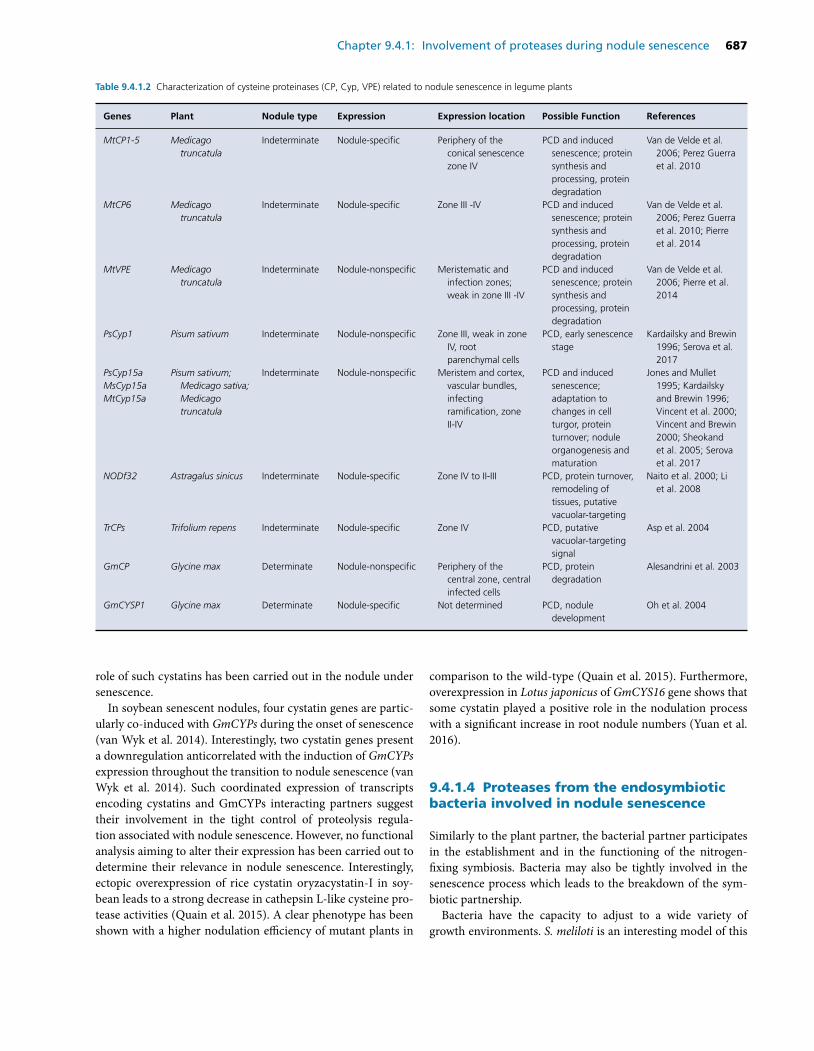
Chapter 9.4.1: Involvement of proteases during nodule senescence 687
Table 9.4.1.2 Characterization of cysteine proteinases (CP, Cyp, VPE) related to nodule senescence in legume plants
Genes Plant Nodule type Expression Expression location Possible Function References
MtCP1-5 Medicagotruncatula
Indeterminate Nodule-specific Periphery of theconical senescencezone IV
PCD and inducedsenescence; proteinsynthesis andprocessing, proteindegradation
Van de Velde et al.2006; Perez Guerraet al. 2010
MtCP6 Medicagotruncatula
Indeterminate Nodule-specific Zone III -IV PCD and inducedsenescence; proteinsynthesis andprocessing, proteindegradation
Van de Velde et al.2006; Perez Guerraet al. 2010; Pierreet al. 2014
MtVPE Medicagotruncatula
Indeterminate Nodule-nonspecific Meristematic andinfection zones;weak in zone III -IV
PCD and inducedsenescence; proteinsynthesis andprocessing, proteindegradation
Van de Velde et al.2006; Pierre et al.2014
PsCyp1 Pisum sativum Indeterminate Nodule-nonspecific Zone III, weak in zoneIV, rootparenchymal cells
PCD, early senescencestage
Kardailsky and Brewin1996; Serova et al.2017
PsCyp15aMsCyp15aMtCyp15a
Pisum sativum;Medicago sativa;Medicagotruncatula
Indeterminate Nodule-nonspecific Meristem and cortex,vascular bundles,infectingramification, zoneII-IV
PCD and inducedsenescence;adaptation tochanges in cellturgor, proteinturnover; noduleorganogenesis andmaturation
Jones and Mullet1995; Kardailskyand Brewin 1996;Vincent et al. 2000;Vincent and Brewin2000; Sheokandet al. 2005; Serovaet al. 2017
NODf32 Astragalus sinicus Indeterminate Nodule-specific Zone IV to II-III PCD, protein turnover,remodeling oftissues, putativevacuolar-targeting
Naito et al. 2000; Liet al. 2008
TrCPs Trifolium repens Indeterminate Nodule-specific Zone IV PCD, putativevacuolar-targetingsignal
Asp et al. 2004
GmCP Glycine max Determinate Nodule-nonspecific Periphery of thecentral zone, centralinfected cells
PCD, proteindegradation
Alesandrini et al. 2003
GmCYSP1 Glycine max Determinate Nodule-specific Not determined PCD, noduledevelopment
Oh et al. 2004
role of such cystatins has been carried out in the nodule undersenescence.
In soybean senescent nodules, four cystatin genes are partic-ularly co-induced with GmCYPs during the onset of senescence(van Wyk et al. 2014). Interestingly, two cystatin genes presenta downregulation anticorrelated with the induction of GmCYPsexpression throughout the transition to nodule senescence (vanWyk et al. 2014). Such coordinated expression of transcriptsencoding cystatins and GmCYPs interacting partners suggesttheir involvement in the tight control of proteolysis regula-tion associated with nodule senescence. However, no functionalanalysis aiming to alter their expression has been carried out todetermine their relevance in nodule senescence. Interestingly,ectopic overexpression of rice cystatin oryzacystatin-I in soy-bean leads to a strong decrease in cathepsin L-like cysteine pro-tease activities (Quain et al. 2015). A clear phenotype has beenshown with a higher nodulation efficiency of mutant plants in
comparison to the wild-type (Quain et al. 2015). Furthermore,overexpression in Lotus japonicus of GmCYS16 gene shows thatsome cystatin played a positive role in the nodulation processwith a significant increase in root nodule numbers (Yuan et al.2016).
9.4.1.4 Proteases from the endosymbioticbacteria involved in nodule senescence
Similarly to the plant partner, the bacterial partner participatesin the establishment and in the functioning of the nitrogen-fixing symbiosis. Bacteria may also be tightly involved in thesenescence process which leads to the breakdown of the sym-biotic partnership.
Bacteria have the capacity to adjust to a wide variety ofgrowth environments. S. meliloti is an interesting model of this
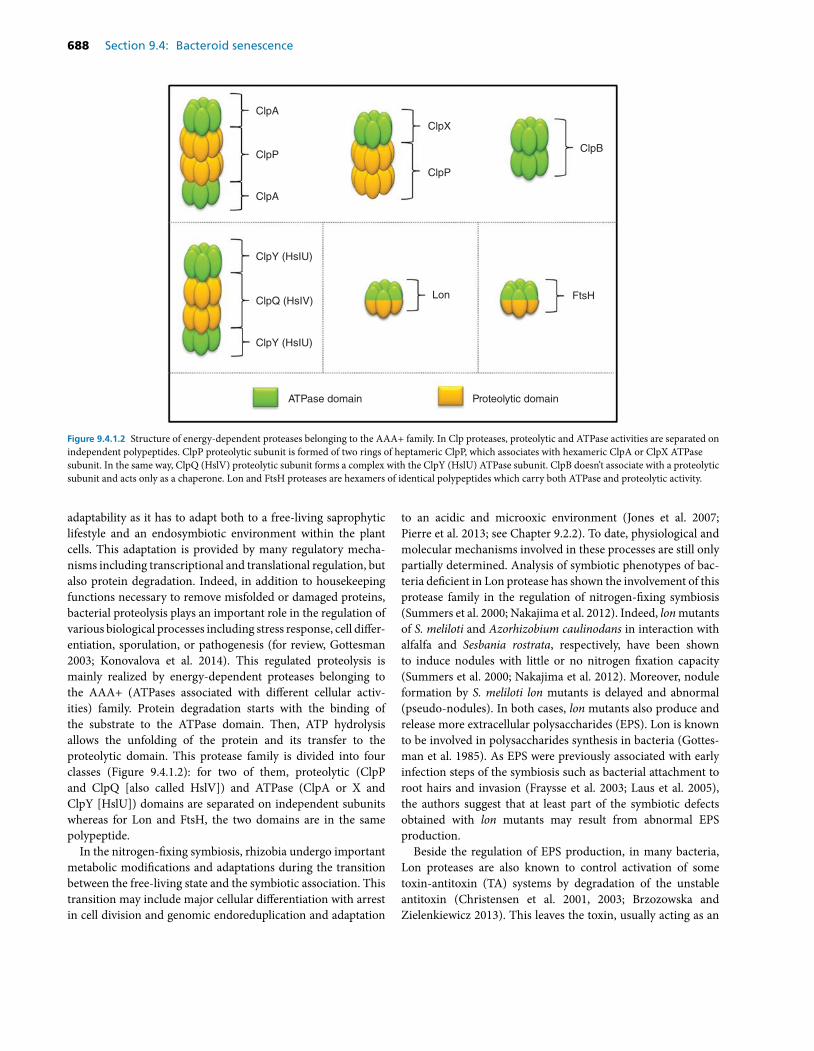
688 Section 9.4: Bacteroid senescence
ClpA
ClpA
ClpY (HsIU)
ClpQ (HsIV)Lon FtsH
Proteolytic domainATPase domain
ClpY (HsIU)
ClpP
ClpP
ClpB
ClpX
Figure 9.4.1.2 Structure of energy-dependent proteases belonging to the AAA+ family. In Clp proteases, proteolytic and ATPase activities are separated onindependent polypeptides. ClpP proteolytic subunit is formed of two rings of heptameric ClpP, which associates with hexameric ClpA or ClpX ATPasesubunit. In the same way, ClpQ (HslV) proteolytic subunit forms a complex with the ClpY (HslU) ATPase subunit. ClpB doesn’t associate with a proteolyticsubunit and acts only as a chaperone. Lon and FtsH proteases are hexamers of identical polypeptides which carry both ATPase and proteolytic activity.
adaptability as it has to adapt both to a free-living saprophyticlifestyle and an endosymbiotic environment within the plantcells. This adaptation is provided by many regulatory mecha-nisms including transcriptional and translational regulation, butalso protein degradation. Indeed, in addition to housekeepingfunctions necessary to remove misfolded or damaged proteins,bacterial proteolysis plays an important role in the regulation ofvarious biological processes including stress response, cell differ-entiation, sporulation, or pathogenesis (for review, Gottesman2003; Konovalova et al. 2014). This regulated proteolysis ismainly realized by energy-dependent proteases belonging tothe AAA+ (ATPases associated with different cellular activ-ities) family. Protein degradation starts with the binding ofthe substrate to the ATPase domain. Then, ATP hydrolysisallows the unfolding of the protein and its transfer to theproteolytic domain. This protease family is divided into fourclasses (Figure 9.4.1.2): for two of them, proteolytic (ClpPand ClpQ [also called HslV]) and ATPase (ClpA or X andClpY [HslU]) domains are separated on independent subunitswhereas for Lon and FtsH, the two domains are in the samepolypeptide.
In the nitrogen-fixing symbiosis, rhizobia undergo importantmetabolic modifications and adaptations during the transitionbetween the free-living state and the symbiotic association. Thistransition may include major cellular differentiation with arrestin cell division and genomic endoreduplication and adaptation
to an acidic and microoxic environment (Jones et al. 2007;Pierre et al. 2013; see Chapter 9.2.2). To date, physiological andmolecular mechanisms involved in these processes are still onlypartially determined. Analysis of symbiotic phenotypes of bac-teria deficient in Lon protease has shown the involvement of thisprotease family in the regulation of nitrogen-fixing symbiosis(Summers et al. 2000; Nakajima et al. 2012). Indeed, lon mutantsof S. meliloti and Azorhizobium caulinodans in interaction withalfalfa and Sesbania rostrata, respectively, have been shownto induce nodules with little or no nitrogen fixation capacity(Summers et al. 2000; Nakajima et al. 2012). Moreover, noduleformation by S. meliloti lon mutants is delayed and abnormal(pseudo-nodules). In both cases, lon mutants also produce andrelease more extracellular polysaccharides (EPS). Lon is knownto be involved in polysaccharides synthesis in bacteria (Gottes-man et al. 1985). As EPS were previously associated with earlyinfection steps of the symbiosis such as bacterial attachment toroot hairs and invasion (Fraysse et al. 2003; Laus et al. 2005),the authors suggest that at least part of the symbiotic defectsobtained with lon mutants may result from abnormal EPSproduction.
Beside the regulation of EPS production, in many bacteria,Lon proteases are also known to control activation of sometoxin-antitoxin (TA) systems by degradation of the unstableantitoxin (Christensen et al. 2001, 2003; Brzozowska andZielenkiewicz 2013). This leaves the toxin, usually acting as an
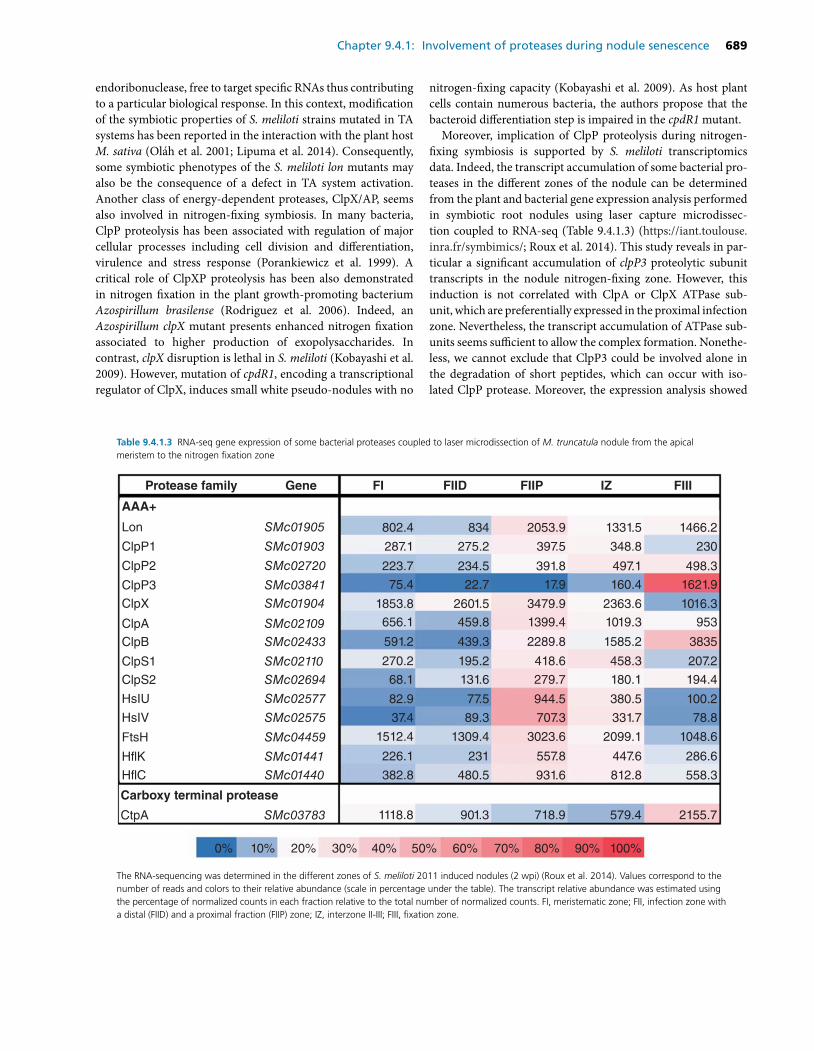
Chapter 9.4.1: Involvement of proteases during nodule senescence 689
endoribonuclease, free to target specific RNAs thus contributingto a particular biological response. In this context, modificationof the symbiotic properties of S. meliloti strains mutated in TAsystems has been reported in the interaction with the plant hostM. sativa (Olah et al. 2001; Lipuma et al. 2014). Consequently,some symbiotic phenotypes of the S. meliloti lon mutants mayalso be the consequence of a defect in TA system activation.Another class of energy-dependent proteases, ClpX/AP, seemsalso involved in nitrogen-fixing symbiosis. In many bacteria,ClpP proteolysis has been associated with regulation of majorcellular processes including cell division and differentiation,virulence and stress response (Porankiewicz et al. 1999). Acritical role of ClpXP proteolysis has been also demonstratedin nitrogen fixation in the plant growth-promoting bacteriumAzospirillum brasilense (Rodriguez et al. 2006). Indeed, anAzospirillum clpX mutant presents enhanced nitrogen fixationassociated to higher production of exopolysaccharides. Incontrast, clpX disruption is lethal in S. meliloti (Kobayashi et al.2009). However, mutation of cpdR1, encoding a transcriptionalregulator of ClpX, induces small white pseudo-nodules with no
nitrogen-fixing capacity (Kobayashi et al. 2009). As host plantcells contain numerous bacteria, the authors propose that thebacteroid differentiation step is impaired in the cpdR1 mutant.
Moreover, implication of ClpP proteolysis during nitrogen-fixing symbiosis is supported by S. meliloti transcriptomicsdata. Indeed, the transcript accumulation of some bacterial pro-teases in the different zones of the nodule can be determinedfrom the plant and bacterial gene expression analysis performedin symbiotic root nodules using laser capture microdissec-tion coupled to RNA-seq (Table 9.4.1.3) (https://iant.toulouse.inra.fr/symbimics/; Roux et al. 2014). This study reveals in par-ticular a significant accumulation of clpP3 proteolytic subunittranscripts in the nodule nitrogen-fixing zone. However, thisinduction is not correlated with ClpA or ClpX ATPase sub-unit, which are preferentially expressed in the proximal infectionzone. Nevertheless, the transcript accumulation of ATPase sub-units seems sufficient to allow the complex formation. Nonethe-less, we cannot exclude that ClpP3 could be involved alone inthe degradation of short peptides, which can occur with iso-lated ClpP protease. Moreover, the expression analysis showed
Table 9.4.1.3 RNA-seq gene expression of some bacterial proteases coupled to laser microdissection of M. truncatula nodule from the apicalmeristem to the nitrogen fixation zone
Protease family
AAA+
Lon 802.4
287.1
223.7
75.4
1853.8
656.1
591.2
270.2
68.1
82.9
37.4
1512.4
226.1
382.8
ClpP1
ClpP2
ClpP3
ClpX
ClpA
ClpB
ClpS1
ClpS2
HsIU
HsIV
FtsH
HflK
HflC
SMc01905
SMc01903
SMc02720
SMc03841
SMc01904
SMc02109SMc02433
SMc02110
SMc02694
SMc02577
SMc02575
SMc04459
SMc01441
SMc01440
CtpA SMc03783 1118.8
834
275.2
234.5
22.7
2601.5
459.8
439.3
195.2
131.6
77.5
89.3
1309.4
231
480.5
901.3
2053.9
397.5
391.8
17.9
3479.9
1399.4
2289.8
418.6
279.7
944.5
707.3
3023.6
557.8
931.6
718.9
1331.5
348.8
497.1
160.4
2363.6
1019.3
1585.2
458.3
180.1
380.5
331.7
2099.1
447.6
812.8
579.4
1466.2
230
498.3
1621.9
1016.3
953
3835
207.2
194.4
100.2
78.8
1048.6
286.6
558.3
2155.7
0% 10% 20% 30% 40% 50% 60% 70% 80% 90% 100%
Carboxy terminal protease
Gene FI FIID FIIP IZ FIII
The RNA-sequencing was determined in the different zones of S. meliloti 2011 induced nodules (2 wpi) (Roux et al. 2014). Values correspond to thenumber of reads and colors to their relative abundance (scale in percentage under the table). The transcript relative abundance was estimated usingthe percentage of normalized counts in each fraction relative to the total number of normalized counts. FI, meristematic zone; FII, infection zone witha distal (FIID) and a proximal fraction (FIIP) zone; IZ, interzone II-III; FIII, fixation zone.

690 Section 9.4: Bacteroid senescence
that clpB, a member of the Clp ATPase family, which acts asa chaperone with the DnaJ, DnaK, and GrpE cellular machin-ery to disaggregate proteins, is also induced in the fixing zone.Therefore, all these data suggest an important role of this pro-tease family in the functioning of the nitrogen-fixing interac-tion in addition to its probable implication in its establishment.Interestingly, the transcriptomic data analysis also reveals theinduction in the nitrogen-fixing zone of a periplasmic carboxy-terminal processing serine protease gene, ctpA, known to be crit-ical for viability of Rhizobium leguminosarum when grown onsemi-solid medium (Gilbert et al. 2007). The role of the expres-sion of ctpA during symbiosis remains to be determined.
Finally, the expression of a S. meliloti metallopeptidase(HrrP) involved in the cleavage of host-encoded nodule-specificcysteine-rich (NCR) peptides may lead to premature bacteroidsdegeneration (Price et al. 2015). This result suggests a role of thisprotease in modifying the host-encoded NCR pool. This con-tributes to the plant-microbe crosstalk in the late developmentstage and demonstrates the role of bacterial proteolysis to main-tain the symbiotic interaction.
The understanding of the implication of bacterium in thenodule senescence is in its infancy. Further analyses have to beperformed to acquire a global vision of this critical process.
9.4.1.5 Conclusion
Many analyses have shown that a large panel of proteases areinvolved in nodule senescence under developmental or stress-induced senescence. One of the main reasons for the activationof the protease pathways in nodule senescence is linked to nutri-ent remobilization and programmed degradation of symbioticpartnership, in order to eliminate the useless organ during devel-opmental senescence or to maintain the survival of the plantduring stress-induced senescence. Nevertheless, the strong andfast induction of proteases may interfere with an efficient yieldproduction of legumes in agricultural conditions. In a worldconfronted with global warming and limited access to water, alarge investment in plant research attempting to moderate ordelay strong responses to abiotic stresses can be seen as an inter-esting goal in order to produce more resilient plants. In this con-text, the analysis of the regulation of protease activation duringthe transition from symbiosis to senescence may allow us to bet-ter understand the key mechanisms involved in the senescenceinitiation. Moreover, such protease genes may also be essentialtools for the plant assisted marker selections in order to delaythe general plant senescence during adverse climate changes.
Acknowledgments
L.Y. is supported by a doctoral fellowship from the China schol-arship council (CSC). C.S. is the recipient of a doctoral grantfrom the “Ministere de l’Education Nationale, de l’Enseignement
superieur et de la Recherche.” This work was supported by theFrench Government (National Research Agency, ANR) throughthe “STAYPINK” project (reference # ANR-15-CE20-0005) and“Investments for the Future” LABEX SIGNALIFE (program ref-erence # ANR-11-LABX-0028-01).
References
Alesandrini, F., Frendo, P., Puppo, A., and Herouart, D. (2003). Isolationof a molecular marker of soybean nodule senescence. Plant Physiol-ogy and Biochemistry 41: 727–732.
Asp, T., Bowra, S., Borg, S., and Holm, P.B. (2004). Cloning and char-acterisation of three groups of cysteine protease genes expressed inthe senescing zone of white clover (Trifolium repens) nodules. PlantScience 167: 825–837.
Avci, U., Petzold, H.E., Ismail, I.O. et al. (2008). Cysteine proteasesXCP1 and XCP2 aid micro-autolysis within the intact central vacuoleduring xylogenesis in Arabidopsis roots. The Plant Journal 56: 303–315.
Benchabane, M., Schluter, U., Vorster, J. et al. (2010). Plant cystatins.Biochimie 92: 1657–1666.
Brzozowska, I. and Zielenkiewicz, U. (2013). Regulation of toxin–antitoxin systems by proteolysis. Plasmid 70: 33–41.
Carter, C., Pan, S., Zouhar, J. et al. (2004). The vegetative vacuole pro-teome of Arabidopsis thaliana reveals predicted and unexpected pro-teins. Plant Cell 16: 3285–3303.
Chichkova, N.V., Shaw, J., Galiullina, R.A. et al. (2010). Phytaspase, arelocalisable cell death promoting plant protease with caspase speci-ficity. The EMBO Journal 29: 1149–1161.
Christensen, S.K., Mikkelsen, M., Pedersen, K., and Gerdes, K. (2001).RelE, a global inhibitor of translation, is activated during nutritionalstress. Proceedings of the National Academy of Sciences of the UnitedStates of America 98: 14328–14333.
Christensen, S.K., Pedersen, K., Hansen, F.G., and Gerdes, K. (2003).Toxin–antitoxin loci as stress-response-elements: ChpAK/MazF andChpBK cleave translated RNAs and are counteracted by tmRNA.Journal of Molecular Biology 332: 809–819.
Coll, N.S., Vercammen, D., Smidler, A. et al. (2010). Arabidopsis type Imetacaspases control cell death. Science 330: 1393–1397.
Coll, N.S., Smidler, A., Puigvert, M. et al. (2014). The plant metacaspaseAtMC1 in pathogen-triggered programmed cell death and aging:functional linkage with autophagy. Cell Death and Differentiation 21:1399–1408.
Diaz-Mendoza, M., Velasco-Arroyo, B., Santamaria, M.E. et al. (2016).Plant senescence and proteolysis: two processes with one destiny.Genetics and Molecular Biology 39: 329–338.
Domonkos, A., Horvath, B., Marsh, J.F. et al. (2013). The identificationof novel loci required for appropriate nodule development in Med-icago truncatula. BMC Plant Biology 13: 157.
Fedorova, M., van de Mortel, J., Matsumoto, P.A. et al. (2002). Genome-wide identification of nodule-specific transcripts in the model legumeMedicago truncatula. Plant Physiology 130: 519–537.
Fraysse, N., Couderc, F., and Poinsot, V. (2003). Surface polysaccharideinvolvement in establishing the rhizobium–legume symbiosis. TheFEBS Journal 270: 1365–1380.
Gilbert, K.B., Vanderlinde, E.M., and Yost, C.K. (2007). Mutagenesis ofthe carboxy terminal protease CtpA decreases desiccation tolerance

Chapter 9.4.1: Involvement of proteases during nodule senescence 691
in Rhizobium leguminosarum. FEMS Microbiology Letters 272: 65–74.
Gottesman, S. (2003). Proteolysis in bacterial regulatory circuits.Annual Review of Cell and Developmental Biology 19: 565–587.
Gottesman, S., Trisler, P., and Torres-Cabassa, A. (1985). Regulation ofcapsular polysaccharide synthesis in Escherichia coli K-12: character-ization of three regulatory genes. Journal of Bacteriology 162: 1111–1119.
Gururani, M.A., Venkatesh, J., and Tran, L.S. (2015). Regulation of pho-tosynthesis during abiotic stress-induced photoinhibition. MolecularPlant 8: 1304–1320.
Han, J.J., Lin, W., Oda, Y. et al. (2012). The proteasome is responsible forcaspase-3-like activity during xylem development. The Plant Journal72: 129–141.
Hatsugai, N., Yamada, K., Goto-Yamada, S., and Hara-Nishimura, I.(2015). Vacuolar processing enzyme in plant programmed cell death.Frontiers in Plant Science 6: 234.
Have, M., Marmagne, A., Chardon, F., and Masclaux-Daubresse, C.(2017). Nitrogen remobilization during leaf senescence: lessons fromArabidopsis to crops. Journal of Experimental Botany 68: 2513–2529.
Huang, X., Ouyang, X., and Deng, X.W. (2014). Beyond repression ofphotomorphogenesis: role switching of COP/DET/FUS in light sig-naling. Current Opinion in Plant Biology 21: 96–103.
Jajic, I., Sarna, T., and Strzalka, K. (2015). Senescence, stress, and reactiveoxygen species. Plants (Basel) 4: 393–411.
Jones, J.T. and Mullet, J.E. (1995). A salt-and dehydration-inducible peagene, Cyp15a, encodes a cell-wall protein with sequence similarity tocysteine proteases. Plant Molecular Biology 28: 1055–1065.
Jones, K.M., Kobayashi, H., Davies, B.W. et al. (2007). How rhizobialsymbionts invade plants: the Sinorhizobium–Medicago model. NatureReviews Microbiology 5: 619–633.
Kardailsky, I.V. and Brewin, N.J. (1996). Expression of cysteine proteasegenes in pea nodule development and senescence. Molecular Plant-Microbe Interactions 9: 689–695.
Kmiec, B., Teixeira, P.F., and Glaser, E. (2014). Shredding the signal: tar-geting peptide degradation in mitochondria and chloroplasts. Trendsin Plant Science 19: 771–778.
Kobayashi, H., De Nisco, N.J., Chien, P. et al. (2009). Sinorhizobiummeliloti CpdR1 is critical for co-ordinating cell cycle progression andthe symbiotic chronic infection. Molecular Microbiology 73: 586–600.
Kondorosi, E., Roudier, F., and Gendreau, E. (2000). Plant cell-size con-trol: growing by ploidy? Current Opinion in Plant Biology 3: 488–492.
Konovalova, A., Søgaard-Andersen, L., and Kroos, L. (2014). Regulatedproteolysis in bacterial development. FEMS Microbiology Reviews38: 493–522.
Laus, M., Van Brussel, A., and Kijne, J. (2005). Role of cellulose fibrilsand exopolysaccharides of Rhizobium leguminosarum in attachmentto and infection of Vicia sativa root hairs. Molecular Plant-MicrobeInteractions 18: 533–538.
Lehtovaara, P. and Perttila, U. (1978). Bile-pigment formation from dif-ferent leghaemoglobins. Methine-bridge specificity of coupled oxida-tion. Biochemical Journal 176: 359–364.
Li, Y., Zhou, L., Chen, D. et al. (2008). A nodule-specific plant cysteineproteinase, AsNODF32, is involved in nodule senescence and nitro-gen fixation activity of the green manure legume Astragalus sinicus.The New Phytologist 180: 185–192.
Lim, P.O., Kim, H.J., and Nam, H.G. (2007). Leaf senescence. AnnualReview of Plant Biology 58: 115–136.
Limpens, E., Ivanov, S., van Esse, W. et al. (2009). Medicago N2-fixing symbiosomes acquire the endocytic identity marker Rab7 butdelay the acquisition of vacuolar identity. The Plant Cell 21: 2811–2828.
Lipuma, J., Cinege, G., Bodogai, M. et al. (2014). A vapBC-type toxin–antitoxin module of Sinorhizobium meliloti influences symbiotic effi-ciency and nodule senescence of Medicago sativa. EnvironmentalMicrobiology 16: 3714–3729.
Lohman, K.N., Gan, S., John, M.C., and Amasino, R.M. (1994). Molec-ular analysis of natural leaf senescence in Arabidopsis thaliana. Phys-iologia Plantarum 92: 322–328.
Malik, N.S., Pfeiffer, N., Williams, D., and Wagner, F.W. (1981). Pep-tidohydrolases of soybean root nodules identification, separation,and partial characterization of enzymes from bacteroid-free extracts.Journal of Plant Physiology 68: 386–392.
Marino, D., Peeters, N., and Rivas, S. (2012). Ubiquitination duringplant immune signaling. Journal of Plant Physiology 160: 15–27.
Martinez, M., Cambra, I., Gonzalez-Melendi, P. et al. (2012). C1Acysteine-proteases and their inhibitors in plants. Physiologia Plan-tarum 145: 85–94.
Matamoros, M.A., Baird, L.M., Escuredo, P.R. et al. (1999). Stress-induced legume root nodule senescence. Physiological, biochemi-cal, and structural alterations. Journal of Plant Physiology 121: 97–112.
Matile, P. and Martinoia, E. (1982). Catabolism of carotenoids: involve-ment of peroxidase? Plant Cell Reports 1: 244–246.
McLellan, H., Gilroy, E.M., Yun, B.W. et al. (2009). Functional redun-dancy in the Arabidopsis Cathepsin B gene family contributes to basaldefence, the hypersensitive response and senescence. The New Phy-tologist 183: 408–418.
Mellor, R.B. (1989). Bacteroids in the rhizobium-legume symbiosisinhabit a plant internal lytic compartment: implications for othermicrobial endosymbioses. Journal of Experimental Botany 40: 831–839.
Minina, E.A., Filonova, L.H., Fukada, K. et al. (2013). Autophagy andmetacaspase determine the mode of cell death in plants. The Journalof Cell Biology 203: 917–927.
Mishra, R.C. and Richa Grover, A. (2016). Constitutive over-expressionof rice ClpD1 protein enhances tolerance to salt and desicca-tion stresses in transgenic Arabidopsis plants. Plant Science 250:69–78.
Naito, Y., Fujie, M., Usami, S. et al. (2000). The involvement of a cys-teine proteinase in the nodule development in Chinese milk vetchinfected with Mesorhizobium huakuii subsp. rengei. Plant Physiology124: 1087–1096.
Nakajima, A., Aono, T., Tsukada, S. et al. (2012). Lon protease ofAzorhizobium caulinodans ORS571 is required for suppression of rebgene expression. Applied and Environmental Microbiology 78: 6251–6261.
Nakaune, S., Yamada, K., Kondo, M. et al. (2005). A vacuolar process-ing enzyme, deltaVPE, is involved in seed coat formation at the earlystage of seed development. Plant Cell 17: 876–887.
Oh, C.J., Lee, H., Kim, H.B., and An, C.S. (2004). Isolation and charac-terization of a root nodule-specific cysteine proteinase cDNA fromsoybean. Journal of Plant Biology 47: 216–220.
Olah, B., Kiss, E., Gyorgypal, Z. et al. (2001). Mutation in the ntrR gene,a member of the vap gene family, increases the symbiotic efficiency

692 Section 9.4: Bacteroid senescence
of Sinorhizobium meliloti. Molecular Plant-Microbe Interactions 14:887–894.
Otegui, M.S., Noh, Y.S., Martınez, D.E. et al. (2005). Senescence-associated vacuoles with intense proteolytic activity develop inleaves of Arabidopsis and soybean. The Plant Journal 41: 831–844.
Pearce, G. (2011). Systemin, hydroxyproline-rich systemin and theinduction of protease inhibitors. Current Protein and Peptide Science12: 399–408.
Perez Guerra, J.C., Coussens, G., De Keyser, A. et al. (2010). Comparisonof developmental and stress-induced nodule senescence in Medicagotruncatula. Plant Physiology 152: 1574–1584.
Pfeiffer, N.E., Malik, N., and Wagner, F.W. (1983). Reversible dark-induced senecence of soybean root nodules. Journal of Plant Phys-iology 71: 393–399.
Pierre, O., Engler, G., Hopkins, J. et al. (2013). Peribacteroid spaceacidification: a marker of mature bacteroid functioning in Med-icago truncatula nodules. Plant, Cell & Environment 36: 2059–2070.
Pierre, O., Hopkins, J., Combier, M. et al. (2014). Involvement of papainand legumain proteinase in the senescence process of Medicago trun-catula nodules. The New Phytologist 202: 849–863.
Pislariu, C.I., Murray, J.D., Wen, J. et al. (2012). A Medicago truncatulatobacco retrotransposon insertion mutant collection with defects innodule development and symbiotic nitrogen fixation. Journal of PlantPhysiology 159: 1686–1699.
Pladys, D. and Rigaud, J. (1985). Senescence in French-bean nodules:occurrence of different proteolytic activities. Physiologia Plantarum63: 43–48.
Pladys, D. and Vance, C.P. (1993). Proteolysis during development andsenescence of effective and plant gene-controlled ineffective alfalfanodules. Journal of Plant Physiology 103: 379–384.
Porankiewicz, J., Wang, J., and Clarke, A.K. (1999). New insights into theATP-dependent Clp protease: Escherichia coli and beyond. MolecularMicrobiology 32: 449–458.
Price, P.A., Tanner, H.R., Dillon, B.A. et al. (2015). Rhizobial peptidaseHrrP cleaves host-encoded signaling peptides and mediates symbi-otic compatibility. Proceedings of the National Academy of Sciencesof the United States of America 112: 15244–15249.
Puppo, A., Groten, K., Bastian, F. et al. (2005). Legume nodule senes-cence: roles for redox and hormone signalling in the orchestra-tion of the natural aging process. The New Phytologist 165: 683–701.
Quain, M.D., Makgopa, M.E., Cooper, J.W. et al. (2015). Ectopic phy-tocystatin expression increases nodule numbers and influences theresponses of soybean (Glycine max) to nitrogen deficiency. Phyto-chemistry 112: 179–187.
Ramirez, V., Lopez, A., Mauch-Mani, B. et al. (2013). An extracellularsubtilase switch for immune priming in Arabidopsis. PLoS Pathogens9: e1003445.
Rawlings, N.D., Barrett, A.J., and Bateman, A. (2016). MEROPS: thedatabase of proteolytic enzymes, their substrates and inhibitors.Nucleic Acids Research 40: D343–D350.
Reinbothe, C., Springer, A., Samol, I., and Reinbothe, S. (2009). Plantoxylipins: role of jasmonic acid during programmed cell death,defence and leaf senescence. The FEBS Journal 276: 4666–4681.
Richau, K.H., Kaschani, F., Verdoes, M. et al. (2012). Subclassifica-tion and biochemical analysis of plant papain-like cysteine proteases
displays subfamily-specific characteristics. Journal of Plant Physiol-ogy 158: 1583–1599.
Rivard, D., Girard, C., Anguenot, R. et al. (2007). MsCYS1, adevelopmentally-regulated cystatin from alfalfa. Plant Physiologyand Biochemistry 45: 508–514.
Roberts, I.N., Caputo, C., Criado, M.V., and Funk, C. (2012).Senescence-associated proteases in plants. Physiologia Plantarum145: 130–139.
Rodriguez, H., Mendoza, A., Cruz, M.A. et al. (2006). Pleiotropic phys-iological effects in the plant growth-promoting bacterium Azospir-illum brasilense following chromosomal labeling in the clpX gene.FEMS Microbiology Ecology 57: 217–225.
Roux, B., Rodde, N., Jardinaud, M.F. et al. (2014). An integrated analysisof plant and bacterial gene expression in symbiotic root nodules usinglaser-capture microdissection coupled to RNA sequencing. The PlantJournal 77: 817–837.
Salehin, M., Bagchi, R., and Estelle, M. (2015). SCFTIR1/AFB-basedauxin perception: mechanism and role in plant growth and develop-ment. Plant Cell 27: 9–19.
Serova, T.A., Tikhonovich, I.A., and Tsyganov, V.E. (2017). Analysis ofnodule senescence in pea (Pisum sativum L.) using laser microdissec-tion, real-time PCR, and ACC immunolocalization. Journal of PlantPhysiology 212: 29–44.
Sheokand, S., Dahiya, P., Vincent, J., and Brewin, N. (2005). Modifiedexpression of cysteine protease affects seed germination, vegetativegrowth and nodule development in transgenic lines of Medicago trun-catula. Plant Science 169: 966–975.
Shi, H., Liu, W., Wei, Y., and Ye, T. (2017). Integration of auxin/indole-3-acetic acid 17 and RGA-LIKE3 confers salt stress resistance throughstabilization by nitric oxide in Arabidopsis. Journal of ExperimentalBotany 68: 1239–1249.
Shindo, T., Misas-Villamil, J.C., Horger, A.C. et al. (2012). A role inimmunity for Arabidopsis cysteine protease RD21, the ortholog of thetomato immune protease C14. PLoS One 7: e29317.
Suarez, M.F., Filonova, L.H., Smertenko, A. et al. (2004). Metacaspase-dependent programmed cell death is essential for plant embryogen-esis. Current Biology 14: R339–R340.
Sueldo, D.J. and van der Hoorn, R.A.L. (2017). Plant life needs cell death,but does plant cell death need Cys proteases? The FEBS Journal 284:1577–1585.
Summers, M.L., Botero, L.M., Busse, S.C., and McDermott, T.R. (2000).The Sinorhizobium meliloti Lon protease is involved in regulatingexopolysaccharide synthesis and is required for nodulation of alfalfa.Journal of Bacteriology 182: 2551–2558.
Tan-Wilson, A.L. and Wilson, K.A. (2012). Mobilization of seed proteinreserves. Physiologia Plantarum 145: 140–153.
Thomas, H. (2013). Senescence, ageing and death of the whole plant.The New Phytologist 197: 696–711.
Van de Velde, W., Guerra, J.C., De Keyser, A. et al. (2006). Aging inlegume symbiosis. A molecular view on nodule senescence in Med-icago truncatula. Journal of Plant Physiology 141: 711–720.
Velasco-Arroyo, B., Diaz-Mendoza, M., Gandullo, J. et al. (2016).HvPap-1 C1A protease actively participates in barley proteolysismediated by abiotic stresses. Journal of Experimental Botany 67:4297–4310.
Vincent, J.L. and Brewin, N.J. (2000). Immunolocalization of a cysteineprotease in vacuoles, vesicles, and symbiosomes of pea nodule cells.Plant Physiology 123: 521–530.

Chapter 9.4.1: Involvement of proteases during nodule senescence 693
Vincent, J.L., Knox, M.R., Ellis, T.N. et al. (2000). Nodule-expressedCyp15a cysteine protease genes map to syntenic genome regions inPisum and Medicago spp. Molecular Plant-Microbe Interactions 13:715–723.
Wager, A. and Browse, J. (2012). Social network: JAZ protein inter-actions expand our knowledge of jasmonate signaling. Frontiers inPlant Science 3: 41.
van Wyk, S.G., Du Plessis, M., Cullis, C.A. et al. (2014). Cysteine pro-tease and cystatin expression and activity during soybean noduledevelopment and senescence. BMC Plant Biology 14: 294.
Xia, Y., Suzuki, H., Borevitz, J. et al. (2004). An extracellular asparticprotease functions in Arabidopsis disease resistance signaling. TheEMBO Journal 23: 980–988.
Yuan, S., Li, R., Wang, L. et al. (2016). Search for nodulation and nod-ule development-related cystatin genes in the genome of soybean(Glycine max). Frontiers in Plant Science 7.
Zhang, B., Tremousaygue, D., Denance, N. et al. (2014a). PIRIN2 stabi-lizes cysteine protease XCP2 and increases susceptibility to the vascu-lar pathogen Ralstonia solanacearum in Arabidopsis. The Plant Jour-nal 79: 1009–1019.
Zhang, D., Liu, D., Lv, X. et al. (2014b). The cysteine protease CEP1,a key executor involved in tapetal programmed cell death, regulatespollen development in Arabidopsis. Plant Cell 26: 2939–2961.
Zhao, P., Zhou, X.M., Zhang, L.Y. et al. (2013). A bipartite molecularmodule controls cell death activation in the basal cell lineage of plantembryos. PLoS Biology 11: e1001655.


111
The flower that smiles to-day
To-morrow dies;
All that we wish to stay
Tempts and then flies.
Percy Bysshe Shelley

112

113
OBJECTIVES To date, it is unclear whether nodule senescence is the consequence of genetic control
by plant or attack by microbes or is a result of systematic interplay. Despite intensive studies that have been conducted on the early stages of the rhizobium-legume interaction for last decades, research data are scarce on signal transduction cascades and regulatory functions in terms of the nodule senescence phase. Our to-date very limited understanding on the later stage of nodule senescence necessitates more research. Here in this thesis, we have investigated the roles of a bacterial redox-related gene and a plant protease gene in root nodule senescence with different approaches.
On the bacterial side, the first part of this thesis is intended to characterize the senescence process induced by the bacterial glutathione deficiency in the symbiotic interactions. Time course analysis was performed to analyse the degradation process occurring in the interaction between Medicago truncatula and the SmghsB, a Sinorhizobium meliloti mutant deficient in GSH synthesis. Our scientific question was whether this deficiency induced an early senescence associated to a lack of bacteroid differentiation or an early senescence associated to an early death of bacteroids
On the plant side, one plant cysteine protease gene, MtCP6, was proven to play a crucial regulatory role in nodule senescence by delaying senescence with RNAi while accelerating the senescence process with ectopic overexpression (Pierre et al., 2014). Therefore, a comprehensive study on the transcriptional regulation of this cell death executioner is promising to elucidate the nodule senescence regulatory network. The second part of the thesis is the identification of cis/trans factors in the complex transcriptional network involved in the MtCP6 regulation at the onset of nodule senescence.

114

115
CHAPTER I
Analysis of bacterial GSH deficiency on nodule functioning

116

CHAPTER I
117
I. Introduction
I.1 Redox homeostasis in nodulation
ROS production in response to bacterial invasion
Reactive oxygen species (ROS) refer to oxygen-derived molecules, including free radicals (e.g. superoxide, hydroxyl radicals) and non-free radicals (e.g., hydrogen peroxide, singlet oxygen) (Matamoros et al., 2003). Main sources for ROS are in plants under physiological conditions are respiration, photosynthesis, and N2 fixation (Matamoros et al., 2003). ROS are highly produced during plant development and in response to stresses. As an example, a burst of ROS is featured in plant hypersensitive reaction responding to pathogen elicitors (Lamb & Dixon, 1997).
An oxidative burst is also elicited in the infection of roots by rhizobia (Santos et al., 2001; Puppo et al., 2005). Rhizobia and Nod factor are proven to induce a low level of ROS production which can be effectively blocked by antioxidants such as SOD (Santos et al., 2000; D’Haeze et al., 2002; Ramu et al., 2002). A peroxidase encoding gene, rip1, is rapidly triggered by Nod factor and coincides with the superoxide accumulation during early symbiotic processes (Ramu et al., 2002). Additionally, interfering with the NADPH oxidase-catalysed ROS production by silencing MtRBOHa impairs symbiosis showing that the ROS production is needed to have an efficient nodulation (Marino et al., 2011).
Antioxidants in maintaining nodule function
On the other hand, overproduction of ROS is detrimental for plant development and metabolism. Antioxidant metabolites and enzymes are produced to protect cells from the oxidative damages. The ROS production is kept at a low level after rhizobia infection to maintain a proper function of nodule metabolism including the oxygen sensitive nitrogenase. However, the nitrogen fixation is high energy-demanding therefore requiring high rates of respiration and thus ROS production. Antioxidants which includes molecules such as glutathione and enzymes such as catalases, superoxide dismutases, and peroxidases are thus required to allow the proper nodule functions (Sigaud et al., 1999; Santos et al., 2000; Jamet et al., 2003; Dombrecht et al., 2005). Catalases can convert H2O2 to water and oxygen without consuming reductants (Scandalios et al., 1997). S. meliloti expresses three genes
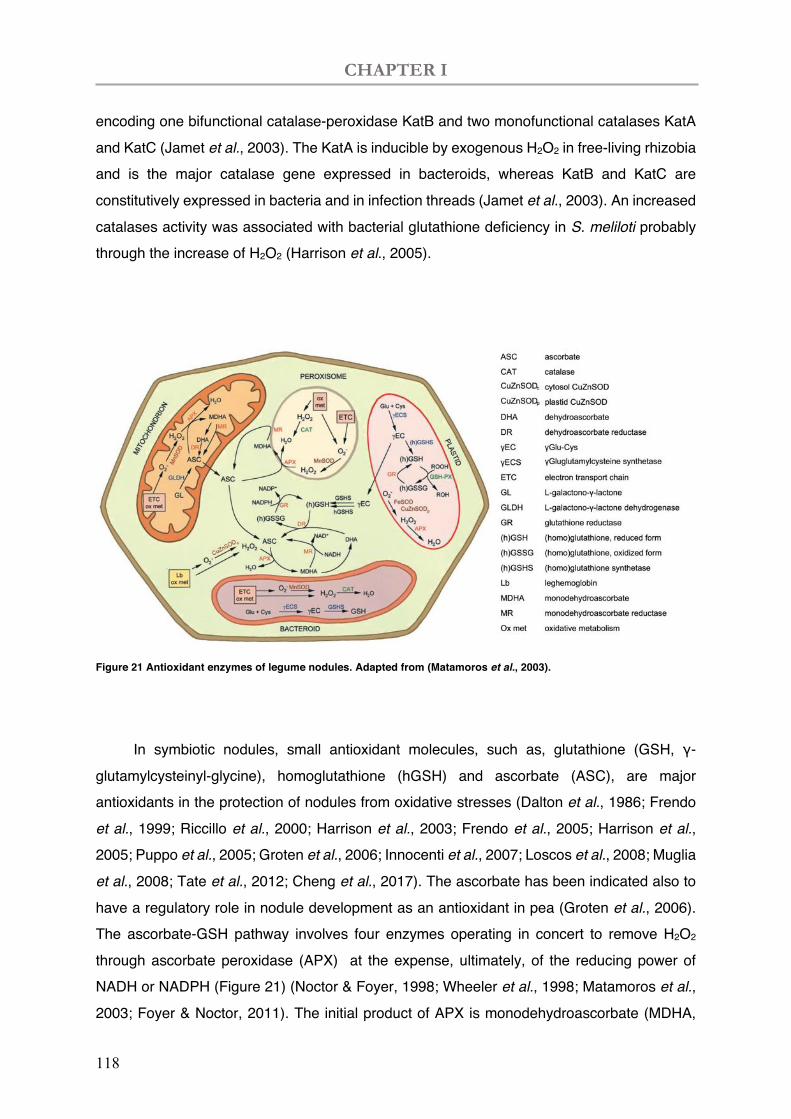
CHAPTER I
118
encoding one bifunctional catalase-peroxidase KatB and two monofunctional catalases KatA and KatC (Jamet et al., 2003). The KatA is inducible by exogenous H2O2 in free-living rhizobia and is the major catalase gene expressed in bacteroids, whereas KatB and KatC are constitutively expressed in bacteria and in infection threads (Jamet et al., 2003). An increased catalases activity was associated with bacterial glutathione deficiency in S. meliloti probably through the increase of H2O2 (Harrison et al., 2005).
Figure 21 Antioxidant enzymes of legume nodules. Adapted from (Matamoros et al., 2003).
In symbiotic nodules, small antioxidant molecules, such as, glutathione (GSH, γ-
glutamylcysteinyl-glycine), homoglutathione (hGSH) and ascorbate (ASC), are major antioxidants in the protection of nodules from oxidative stresses (Dalton et al., 1986; Frendo et al., 1999; Riccillo et al., 2000; Harrison et al., 2003; Frendo et al., 2005; Harrison et al., 2005; Puppo et al., 2005; Groten et al., 2006; Innocenti et al., 2007; Loscos et al., 2008; Muglia et al., 2008; Tate et al., 2012; Cheng et al., 2017). The ascorbate has been indicated also to have a regulatory role in nodule development as an antioxidant in pea (Groten et al., 2006). The ascorbate-GSH pathway involves four enzymes operating in concert to remove H2O2 through ascorbate peroxidase (APX) at the expense, ultimately, of the reducing power of NADH or NADPH (Figure 21) (Noctor & Foyer, 1998; Wheeler et al., 1998; Matamoros et al., 2003; Foyer & Noctor, 2011). The initial product of APX is monodehydroascorbate (MDHA,

CHAPTER I
119
ascorbate free radical), which then disproportionate to ASC and dehydroascorbate (DHA). In the infected cells, APX may participate in the operation of the oxygen diffusion barrier in the nodule parenchyma by adjusting their respiratory activity (Dalton et al., 1998; Matamoros et al., 2003). MDHA and DHA are reduced back to ASC by specific reductases (Monodehydroascorbate reductase-MR, Dihydroascorbate reductase-DR) using NADH and GSH, respectively. Finally, the oxidized GSH (GSSG) formed by DR is reduced to GSH by glutathione reductase (GR) using NADPH (Figure 21) (Matamoros et al., 2003).
ROS production resurges during nodule senescence
During nodule senescence, an increase in ROS productions and a diminish in antioxidants oxidize leghaemoglobin and enhance autolytic processes (Matamoros et al., 1999; Becana et al., 2000; Santos et al., 2001; Hérouart et al., 2002; Alesandrini et al., 2003; Puppo et al., 2005; Pauly et al., 2006; Günther et al., 2007; Muglia et al., 2008; Andrio et al., 2013). Eventually, nodule senescence is accompanied with a reduction in nitrogen fixation capacity and an increase in ROS accumulation (Puppo et al., 2005; Muglia et al., 2008). Therein, oxidative damage of lipids, proteins, and DNA has been observed in nodules during natural and stress-induced senescence (Evans et al., 1999; Matamoros et al., 1999). Large amounts of H2O2 has been shown to accumulate in the cells and apoplast in the central zone of senescing soybean nodules (Alesandrini et al., 2003). These structural and biochemical alterations may result from a switch from a reductive state to an oxidative state, which is a general characteristic of plant senescence (Evans et al., 1999; Puppo et al., 2005; Marquez-Garcia et al., 2015).
I.2 Glutathione in prokaryotes
GSH Synthesis involves a two-step ATP-dependent pathway
The tripeptide GSH is a well-known antioxidant and detoxifier existing in many organisms, with a concentration ranging from 0.1 to 10 mM in bacteria (Penninckx & Elskens, 1993; Carmel-Harel & Storz, 2000; Noctor et al., 2002; Masip et al., 2006). GSH synthesis involves a two-step ATP-dependent pathway (Masip et al., 2006). In the first step, glutamate and cysteine are conjugated by γ-glutamyl cysteine synthetase (γ-ECS; encoding by gshA gene) to form γ-glutamyl cysteine (γ-EC). In the second step, glycine is added to the γ-EC to produce GSH in the reaction catalysed by glutathione synthetase (GSHS; encoding by gshB

CHAPTER I
120
gene). Alternatively, hGSH is formed by adding β-alanine to the γ-EC catalysed by homoglutathione synthetase (hGSHS).
i) L-Glu + L-Cys + ATP ↔ γ-Glu-Cys + ADP + Pi ii) γ-Glu-Cys + Gly + ATP ↔ GSH + ADP + Pi
The first step is the rate-limiting and under control of cysteine concentration and the feedback inhibition of the final product GSH (Lu, 2009). Another possible way to control GSH pool is through its degradation by the γ-glutamyl transpeptidase (GGT) to the γ-linkage between the glutamic acid and cysteine (Masip et al., 2006). However, GGT is localized in the periplasm, which hardly possible to modulate the cytoplasmic GSH level (Suzuki et al., 1986).
GSH play multiple roles in prokaryotes
GSH is involved in multiple biological processes including control of cell redox status, progression of the cell cycle, and detoxification and maintaining intracellular K+ pools (Masip et al., 2006) (Ferguson & Booth, 1998; Matamoros et al., 2003). Most of the GSH functions are investigated in Escherichia coli (Masip et al., 2006). The redox couple GSH:GSSG (reduced and oxidized states of GSH) is carefully controlled in the cytoplasm (Fig 1.2) (Masip et al., 2006). Protein glutathionylation, the reversible formation of mixed disulfides between GSH and cysteinyl residues (Dalle-Donne et al., 2007), is thereafter serving as the protection of reactive thiols under oxidative stresses (Masip et al., 2006). Glutathionylation has been shown to modulate MetE of methionine biosynthesis (Hondorp & Matthews, 2004), 3’-phosphoadenylsulfate (PAPS) reductase of the sulfate assimilation pathway (Lillig et al., 2003), and the transcription factor OxyR of peroxide oxidative stress (Aslund et al., 1999).
During oxidative stress, GSH indirectly reduces the oxyR transcription factor via glutaredoxin 1 (Storz & Tartaglia, 1992; Zheng et al., 1998; Aslund et al., 1999). In E. coli, glutaredoxin is involved in DNA synthesis via providing reduction of Ribonucleotide reductase, which needs at least one of the reducing equivalents NADPH and GSH (Ortenberg et al., 2004; Masip et al., 2006). Other substrates of the glutaredoxin pathway are PAPS reductase, methionine sulfoxide reductase and arsenate reductase (Ritz & Beckwith, 2001; Masip et al., 2006). GSH is required for bacterial anaerobic growth via reduction of fumarate nitrate reductase regulator (FNR). GSH can be the direct reducing power for FNR, which is necessary to control the switch from aerobic to anaerobic conditions (Masip et al., 2006). During anaerobic conditions, GSH is required to activate FNR thus the formed homodimer is with [4Fe-4S] cluster and is able to control bacterial anaerobic growth (Ritz & Beckwith, 2001).

CHAPTER I
121
GSH plays roles in detoxification of xenobiotics. GSH too is known to protect cells from halogens such as chlorine compounds (Chesney et al., 1996; Masip et al., 2006). However, in the glutathione-deficient organisms, resistance can be restored with exogenous GSH caused by chlorine-based oxidants but not by H2O2 (Chesney et al., 1996). GSH is required for the enzymes glyoxalase I and II to detoxify methylglyoxal into D-Lactate (Masip et al., 2006). GSH can negatively regulate KefB and KefC K+ efflux channels, thus depletion of GSH will lead to K+ leakage and counter cytoplasmatic acidity (Ferguson & Booth, 1998; Masip et al., 2006). The decrease in pH will in turn protect cells against toxicity of the electrophile methylglyoxal in absence of GSH (Ferguson & Booth, 1998).
I.3 Role of glutathione in nodule functioning.
Glutathione is an important molecule for the growth of S. meliloti. Previous works on the synthesis of GSH during free-living growth and symbiosis of S. meliloti 1021 with M. sativa were examined by Harrison et al. (Harrison et al., 2005). It was found that a S. meliloti mutant strain which was unable to synthesize GSH because of a gene disruption in gshA, was unable to grow under non-stress conditions, precluding any nodulation. This result showed that GSH plays a crucial role in S. meliloti growth. In contrast, a S. meliloti mutant strain with gshB deleted was able to grow, indicating that γ-EC (di-peptide intermediate) could partially substitute for GSH. However, the SmgshB deficient strain showed a delayed nodulation phenotype coupled with a 75% reduction in the nitrogen fixation capacity of Medicago sativa nodules (Harrison et al., 2005).
Preliminary results obtained during the interaction between M. truncatula and S. meliloti gshB mutant strains suggested that the symbiotic interaction was impaired in the bacterial differentiation step (El Msehli, 2012). The goals of this research project were to analyse in kinetics the symbiotic processes in order to characterize the defective interaction between the S. meliloti 2011 mutant strain (SmgshB) and M. truncatula.

CHAPTER I
122

CHAPTER I
123
II. Results
Yang, L., El Msehli, S., Benyamina, S., Lambert, A., Hopkins, J., Cazareth, J., Pierre, O., Hérouart, D., Achi-Smiti, S., Boncompagni, E., and Frendo, P. (2020). Glutathione Deficiency in Sinorhizobium meliloti Does Not Impair Bacteroid Differentiation But Induces Early Senescence in the Interaction With Medicago truncatula. Front. Plant Sci. 11. doi:10.3389/fpls.2020.00137

CHAPTER I
124

Frontiers in Plant Science | www.frontiersin
Edited by:Benjamin Gourion,
Laboratoire Interactions Plantes-Microorganismes, France
Reviewed by:Maria Jose Soto,
Experimental Station of Zaidín (EEZ),Spain
Viktor E. Tsyganov,All-Russian Research Institute ofAgricultural Microbiology of the
Russian Academy of AgriculturalSciences, Russia
*Correspondence:Pierre Frendo
†These authors share first authorship
Specialty section:This article was submitted toPlant Microbe Interactions,
a section of the journalFrontiers in Plant Science
Received: 17 September 2019Accepted: 29 January 2020Published: 03 March 2020
Citation:Yang L, El Msehli S, Benyamina S,Lambert A, Hopkins J, Cazareth J,Pierre O, Hérouart D, Achi-Smiti S,
Boncompagni E and Frendo P (2020)Glutathione Deficiency in
Sinorhizobium meliloti Does Not ImpairBacteroid Differentiation But InducesEarly Senescence in the Interaction
With Medicago truncatula.Front. Plant Sci. 11:137.
doi: 10.3389/fpls.2020.00137
ORIGINAL RESEARCHpublished: 03 March 2020
doi: 10.3389/fpls.2020.00137
Glutathione Deficiency inSinorhizobium meliloti Does NotImpair Bacteroid Differentiation ButInduces Early Senescence in theInteraction With Medicago truncatulaLi Yang1†, Sarra El Msehli 2†, Sofiane Benyamina1, Annie Lambert1, Julie Hopkins1,Julie Cazareth3, Olivier Pierre1, Didier Hérouart1, Samira Achi-Smiti 2, Eric Boncompagni1
and Pierre Frendo1*
1 Université Côte d'Azur, INRA, CNRS, ISA, Sophia-Antipolis, France, 2 Laboratoire de Physiologie Végétale, Faculté desSciences de Tunis, Campus Universitaire El Manar II, Tunis, Tunisia, 3 Institut de Pharmacologie Moléculaire et Cellulaire,CNRS UMR 7275, Université Côte d'Azur, Valbonne, France
Under nitrogen-limiting conditions, legumes are able to interact symbiotically with bacteriaof the Rhizobiaceae family. This interaction gives rise to a new organ, named a rootnodule. Root nodules are characterized by an increased glutathione (GSH) andhomoglutathione (hGSH) content compared to roots. These low molecular thiols arevery important in the biological nitrogen fixation. In order to characterize the modification ofnodule activity induced by the microsymbiont glutathione deficiency, physiological,biochemical, and gene expression modifications were analyzed in nodules after theinoculation of Medicago truncatula with the SmgshB mutant of Sinorhizobium melilotiwhich is deficient in GSH production. The decline in nitrogen fixation efficiency wascorrelated to the reduction in plant shoot biomass. Flow cytometry analysis showed thatSmgshB bacteroids present a higher DNA content than free living bacteria. Live/deadmicroscopic analysis showed an early bacteroid degradation in SmgshB nodulescompared to control nodules which is correlated to a lower bacteroid content at 20dpi. Finally, the expression of two marker genes involved in nitrogen fixation metabolism,Leghemoglobin and Nodule Cysteine Rich Peptide 001, decreased significantly in mutantnodules at 20 dpi. In contrast, the expression of two marker genes involved in the nodulesenescence, Cysteine Protease 6 and Purple Acid Protease, increased significantly inmutant nodules at 10 dpi strengthening the idea that an early senescence process occursin SmgshB nodules. In conclusion, our results showed that bacterial GSH deficiency doesnot impair bacterial differentiation but induces an early nodule senescence.
Keywords: Medicago truncatula, Sinorhizobium meliloti SmgshB, glutathione, bacteroid differentiation, senescence
.org March 2020 | Volume 11 | Article 1371

Yang et al. S. meliloti GSH Deficiency in Root Nodule
INTRODUCTION
Nitrogen (N) is the major macronutrient needed for plantgrowth. However, whereas N is the most abundant element inthe atmosphere, it is generally the most limiting plant nutrient insoils. Legume plants are able to interact symbiotically withrhizobia to perform nitrogen fixing symbiosis (NFS). In thesemutualistic associations, bacteria deliver nitrogen source to theplants while plants provide bacteria with photosynthates,nutrients required for rhizobium metabolism and a specializedecological niche that favors their growth. Indeed, the nitrogen-fixing symbiosis involves the de novo formation of a new organ,the nodule, associated to a reprogramming of the plantdevelopment and metabolism. This allows the intracellularaccommodation of several thousands of differentiated bacteria,within nodule cells, called bacteroids which reduce atmosphericnitrogen. Amongst the multiple regulatory processes involved inthe setup and the regulation of NFS, a modification of the cellularredox state of the plant partner is observed during the plantinfection and nodule functioning (Puppo et al., 2013). Reactiveoxygen species production is detected in root hairs in response tonodulation factors (Cárdenas et al., 2008), the root infectionprocess (Santos et al., 2001; Jamet et al., 2007) and in the noduledifferentiation zone in Medicago truncatula nodules (Andrioet al., 2013). The implication of the ROS in the symbioticinteraction between legumes and rhizobia has beendemonstrated using genetic approaches (Marino et al., 2011;Arthikala et al., 2014). Genetic alteration of the expression ofrespiratory burst oxidative homologs (Rbohs), proteins involvedin the ROS production, significantly modifies the symbiosisbetween legume plants and the rhizobia (Marino et al., 2011;Marino et al., 2012). Similarly, nitric oxide (NO) production isalso detected in the NFS (Hichri et al., 2015). It is required for theoptimal establishment of the symbiotic interaction (Hichri et al.,2015; Berger et al., 2019). In mature nodules, NO has a beneficialmetabolic function for the maintenance of the energy statusunder hypoxic conditions, but has also been shown to inhibitnitrogen fixation and to trigger nodule senescence. NO has alsobeen shown to regulate the glutathione (GSH) synthesis in roots(Innocenti et al., 2007).
The root nodules contain significantly higher contents of lowmolecular thiols, GSH and homoglutathione (hGSH), incomparison to roots (Frendo et al., 2013a; Frendo et al., 2013b).GSH is the most abundant low-molecular weight thiol in cells andhas several well-known functions. For example, it acts as a redoxbuffer and as a signaling molecule, and it is also involved in thedetoxification of radicals and xenobiotics (Rouhier et al., 2008; Foyerand Noctor, 2011). GSH is synthesized by a two-step process. In thefirst step, glutamate and cysteine are conjugated by g-glutamylcysteine synthetase (g-ECS) to form g-glutamyl cysteine (g-EC). In asecond step, glycine is added to g-EC to form GSH in a reactioncatalyzed by glutathione synthetase (GSHS). Alternatively,homoglutathione synthetase (hGSHS) catalyzes the addition of b-alanine to g-EC to form hGSH. Genetic and pharmacologicalapproaches showed that the GSH pool produced by the plantpartner is important to allow the nodule development and anefficient biological nitrogen fixation (BNF) (Frendo et al., 2005;
Frontiers in Plant Science | www.frontiersin.org 2
El Msehli et al., 2011). On the bacterial side, the use of mutantsdeficient in the GSHS-encoding gene (gshB) impairs the BNFefficiency during the symbiosis between Medicago sativa/S.meliloti strain 1021, Phaseolus vulgaris/Rhizobium tropici, P.vulgaris/Rhizobium etli, and Pisum sativum/Rhizobiumleguminosarum (Harrison et al., 2005; Muglia et al., 2008; Tatéet al., 2012; Cheng et al., 2017). The reduced BNF observed with thegshB mutant strain was correlated with nodule and bacteroid earlysenescence inM. sativa and P. vulgaris (Harrison et al., 2005;Mugliaet al., 2008; Taté et al., 2012). In parallel, recycling of GSH byglutathione reductase was shown to be required to maintain aneffective nitrogen fixation (Tang et al., 2018). Finally, a deficiency inbacteroid differentiation was observed for a S. meliloti (strain 2011)mutant for glutaredoxin 1 (Grx1), a glutaredoxin involved in thedeglutathionylation of proteins using GSH as a cofactor, in thesymbiotic interaction with M. truncatula (Benyamina et al., 2013).Harrison and colleagues (2005) also suggested that glutathionedeficiency in S. meliloti strain 1021 interferes with bacteroiddifferentiation in M. sativa (Harrison et al., 2005) but thishypothesis was not carefully investigated.
In this context, we hypothesized that the genome backgroundof S. meliloti (strain 1021 versus strain 2011) or the genomebackground of the plant (M. sativa versus M. truncatula) couldmodify the outcome of the interaction between the two partners.With this in mind, we intended to characterize the impact of thebacterial GSH deficiency on the bacteroid differentiation and thenodule functioning during the symbiotic interaction between M.truncatula and the strain 2011 of S. meliloti. Physiological,biochemical, cellular and genetic markers were used to describethe nodule functioning at 10 and 20 days after plant inoculation,two early time points of nodule functioning. Our results showthat bacterial GSH deficiency does not affect bacteroiddifferentiation. However, bacterial GSH deficiency induces anearly nodule senescence process in M. truncatula alreadydetectable at 10-d post-infection.
MATERIALS AND METHODS
Plant Material, Bacterial Strainsand Growth ConditionsA mutation in gshB (Smc00419) of S. meliloti 2011 was made byplasmid integration using single-crossover. A 680-bp gshBfragment was PCR-amplified using primers SMc00419-F (5'-AAGAATTCCCATGTTTCGGGCATCACC-3 ' ) andSMc00419-R (5'-AACGATC GCTGCAGTGCATGTTGGAGCGAGAATC-3'). The fragment was cloned into EcoRI-PvuI sitesof pSUP202, resulting in plasmid pSUP202gshB. PlasmidpSup202gshB was transferred from E. coli to S. meliloti 2011and recombined into the genomic gshB region via singlecrossover to give the mutant strain SmgshB in S. meliloti 2011genetic background. The mutants were screen for resistance totetracyclin (10 mg per ml) and streptomycin (100 mg per mL).The SmgshB mutant strain in Sm 2011 genetic background wasvalidated by HPLC for the lack of GSH as described previously(Harrison et al., 2005). The delayed growth phenotype observedfor the mutant strain was complemented by the addition of GSH
March 2020 | Volume 11 | Article 137

Yang et al. S. meliloti GSH Deficiency in Root Nodule
(Figure S1). M. truncatula ecotype Jemalong A17, S. meliloti2011 (wild type), and S. meliloti 2011 gshB (SmgshB) strains weregrown as described (Harrison et al., 2005). M. truncatula seedswere then spread on 0.4% agar gel at 4°C for 2 days forvernalization and germinated at 16°C for 1 day. Seedlings wereafterwards growing in vitro on Fahraeus medium (Boisson-Dernier et al., 2001). Each seedling was inoculated with 200µlof S. meliloti at an O.D.600nm of 0.05. All the plants used in thisstudy were grown in a 16-h light (23°C): 8-h dark (20°C)photoperiod. Nodules were harvested at 10- and 20-days post-inoculation (dpi). At least three independent biologicalrepetitions were analyzed.
Nitrogenase Activity AssayNitrogen fixation activity was determined by C2H2 reductionassay (ARA), using a gas chromatograph. Nodulated roots at 10and 20 dpi were detached from plants and put inside sealed vials.In each vial, 10% of the atmosphere was substituted withacetylene. After 1 h of incubation at 28°C, gas samples wereanalyzed with a gas chromatograph system (Agilent GC 6890N;Agilent Technologies, Massy, France). Twelve plants were testedper replicate and at least three different biological replicateswere analyzed.
Flow CytometryS. meliloti bacteroids were purified as previously described(Trinchant et al., 2004) using a 320 mOsM extraction buffer.Bacterial cells were fixed by heat treatment (70°C, 15 min) andstained with 10 mg/mL propidium iodide (PI). They were thensubjected to flow cytometry analysis with a LSRII Fortessa flowcytometer (BD BioSciences, Rungis, France). Bacterial DNAcontent was determined from PI fluorescence at 585-625 nmafter excitation at 565 nm with a single laser line. Single eventswere recorded and analyzed with BD FACSDiva v6.1.3 (BDBioSciences) and Kaluza v1.2 software (Beckman Coulter,Villepinte, France), respectively. Experiments were performedwith biological triplicates and the recording of more than200,000 events for each sample to ensure the statisticalrobustness of each assay (Ribeiro et al., 2017).
Live/Dead Staining and Microscopic ObservationsNodulated roots and nodules at 10 and 20 dpi, respectively, wereharvested and embedded in 4% agarose. Nodule sections of 70µmwere prepared with a Vibratome HM650V (Thermo FisherScientific, Illkirch-Graffenstaden, France). Afterwards, nodulesections were stained with SYTO™ 9 and propidium iodide(PI) (LIVE/DEAD BacLight Bacterial Viability Kit, ThermoFisher scientific) for 20 min as previously described (Haaget al., 2011). Sections were then mounted on slides andvisualized with a laser scanning confocal microscope LSM880(Carl Zeiss, Marly le Roi, France). The SYTO™ 9 and PI weresequentially excited by 488 nm Argon and 561 nm HeNe laserlines, separately. Both fluorescence signals were recorded onseparated detectors. All images were treated with Fiji software(Schindelin et al., 2012). Samples are from at least 30 nodules of15 plants from three independent replicates.
Frontiers in Plant Science | www.frontiersin.org 3
Quantitative Reverse Transcriptase PCR (RT-qPCR)Young nodules (10 dpi) and mature nodules (20 dpi) wereharvested. Total RNA was extracted from 100 mg of noduleswith Trizol (Invitrogen, Illkirch-Graffenstaden, France) andcDNA was synthesized with the Omniscript RT kit (Qiagen,Les Ulis, France) after a DNAse treatment of total RNA. Absenceof genomic contamination was ensured by control PCR withintron-spanning primers. Quantitative PCR was performedusing AriaMx Real-time PCR system (Agilent, Les Ulis,France). All data were normalized with two constitutivelyexpressed genes, Mtc27 and A38 (M. truncatula) for the M.truncatula genes, and 16S ribosomal RNA and Polyribonucleotidenucleotidyltransferase for the S. meliloti genes. The primersequences used for PCR are described in Table S1. Each RT-qPCR reaction for each of the three biological experiments wasperformed in triplicate. The expression fold change wascalculated as 2-ΔΔCt method (Livak and Schmittgen, 2001).
Statistical AnalysesOur data were reported as mean ± standard error. Thesignificance of the results was assessed using the Mann–Whitney nonparametric test, which allows the comparison ofsmall quantitative samples. When the number of samples wasenough to define a normal distribution (30 samples), the resultswere assessed using Student's t-test.
RESULTS
Analysis of Plant Phenotype and BiologicalNitrogen FixationTo test the phenotype of the S. meliloti SmgshB strain, we haveanalyzed nodule functioning at 10- and 20- dpi which correspondsto young and mature nodules in vitro growth conditions. Thephenotypes of the plants and the root nodules were examined, andthe nitrogen fixation activity was analyzed by ARA (Figure 1). Thenitrogen fixation efficiency was significantly reduced by 53% and21% in SmgshB inoculated plants compared to control plants at 10and 20 dpi, respectively (Figure 1A). A significant lower numberof nodules was observed for SmgshB inoculated plants comparedto the control plants at 10 dpi (Figure S2) and nodule size was notdifferent at 10 or 20 dpi (Figure S2). Phenotypic analysis of rootnodules showed that SmgshB nodules presented a less pronouncedpink color compared to control ones and present an extendedgreen color on the distal part of nodule at 20 dpi (Figure 1B). Asignificant diminution of shoot dry weight biomass and a reducedplant development were observed in SmgshB inoculated plantscompared to the control plants at 20 dpi (Figures 1C and S3). Inconclusion, SmgshB inoculated plants present an altered growthphenotype compared to the control plants.
Analysis of Bacteroid Differentiation UsingFlow CytometryThe bacteroid differentiation is correlated to an increase in size andin DNA content of the bacteroid compared to the bacterium
March 2020 | Volume 11 | Article 137
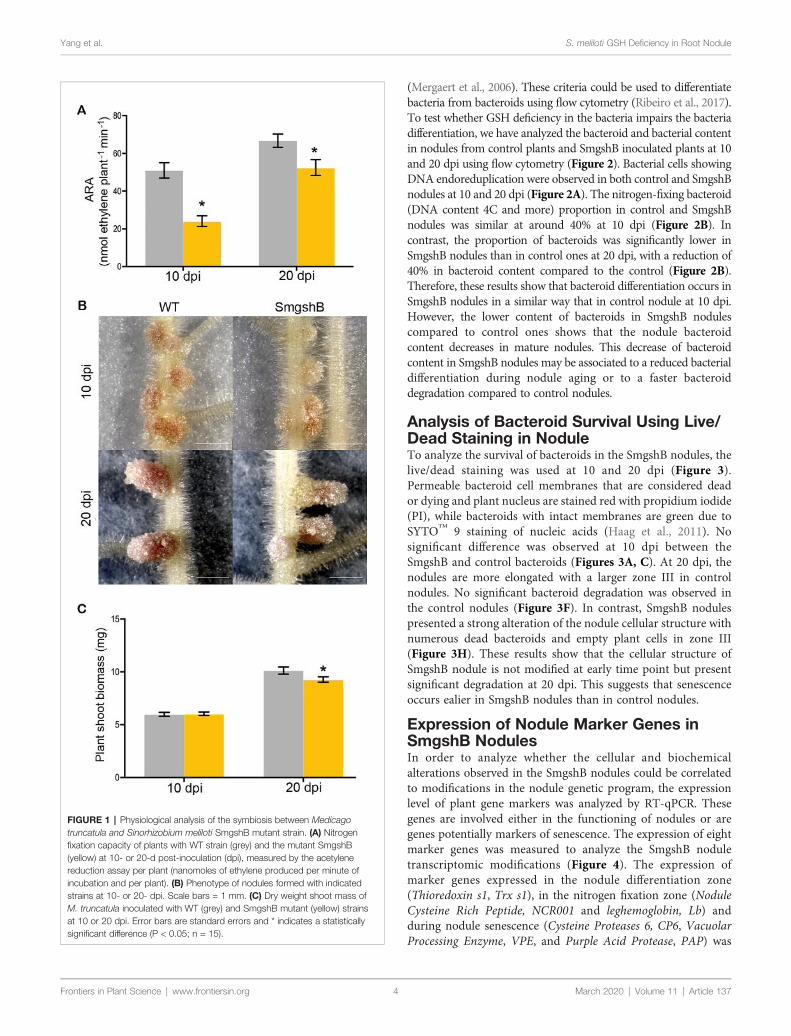
Yang et al. S. meliloti GSH Deficiency in Root Nodule
Frontiers in Plant Science | www.frontiersin.org 4
(Mergaert et al., 2006). These criteria could be used to differentiatebacteria from bacteroids using flow cytometry (Ribeiro et al., 2017).To test whether GSH deficiency in the bacteria impairs the bacteriadifferentiation, we have analyzed the bacteroid and bacterial contentin nodules from control plants and SmgshB inoculated plants at 10and 20 dpi using flow cytometry (Figure 2). Bacterial cells showingDNA endoreduplication were observed in both control and SmgshBnodules at 10 and 20 dpi (Figure 2A). The nitrogen-fixing bacteroid(DNA content 4C and more) proportion in control and SmgshBnodules was similar at around 40% at 10 dpi (Figure 2B). Incontrast, the proportion of bacteroids was significantly lower inSmgshB nodules than in control ones at 20 dpi, with a reduction of40% in bacteroid content compared to the control (Figure 2B).Therefore, these results show that bacteroid differentiation occurs inSmgshB nodules in a similar way that in control nodule at 10 dpi.However, the lower content of bacteroids in SmgshB nodulescompared to control ones shows that the nodule bacteroidcontent decreases in mature nodules. This decrease of bacteroidcontent in SmgshB nodules may be associated to a reduced bacterialdifferentiation during nodule aging or to a faster bacteroiddegradation compared to control nodules.
Analysis of Bacteroid Survival Using Live/Dead Staining in NoduleTo analyze the survival of bacteroids in the SmgshB nodules, thelive/dead staining was used at 10 and 20 dpi (Figure 3).Permeable bacteroid cell membranes that are considered deador dying and plant nucleus are stained red with propidium iodide(PI), while bacteroids with intact membranes are green due toSYTO™ 9 staining of nucleic acids (Haag et al., 2011). Nosignificant difference was observed at 10 dpi between theSmgshB and control bacteroids (Figures 3A, C). At 20 dpi, thenodules are more elongated with a larger zone III in controlnodules. No significant bacteroid degradation was observed inthe control nodules (Figure 3F). In contrast, SmgshB nodulespresented a strong alteration of the nodule cellular structure withnumerous dead bacteroids and empty plant cells in zone III(Figure 3H). These results show that the cellular structure ofSmgshB nodule is not modified at early time point but presentsignificant degradation at 20 dpi. This suggests that senescenceoccurs ealier in SmgshB nodules than in control nodules.
Expression of Nodule Marker Genes inSmgshB NodulesIn order to analyze whether the cellular and biochemicalalterations observed in the SmgshB nodules could be correlatedto modifications in the nodule genetic program, the expressionlevel of plant gene markers was analyzed by RT-qPCR. Thesegenes are involved either in the functioning of nodules or aregenes potentially markers of senescence. The expression of eightmarker genes was measured to analyze the SmgshB noduletranscriptomic modifications (Figure 4). The expression ofmarker genes expressed in the nodule differentiation zone(Thioredoxin s1, Trx s1), in the nitrogen fixation zone (NoduleCysteine Rich Peptide, NCR001 and leghemoglobin, Lb) andduring nodule senescence (Cysteine Proteases 6, CP6, VacuolarProcessing Enzyme, VPE, and Purple Acid Protease, PAP) was
FIGURE 1 | Physiological analysis of the symbiosis between Medicagotruncatula and Sinorhizobium meliloti SmgshB mutant strain. (A) Nitrogenfixation capacity of plants with WT strain (grey) and the mutant SmgshB(yellow) at 10- or 20-d post-inoculation (dpi), measured by the acetylenereduction assay per plant (nanomoles of ethylene produced per minute ofincubation and per plant). (B) Phenotype of nodules formed with indicatedstrains at 10- or 20- dpi. Scale bars = 1 mm. (C) Dry weight shoot mass ofM. truncatula inoculated with WT (grey) and SmgshB mutant (yellow) strainsat 10 or 20 dpi. Error bars are standard errors and * indicates a statisticallysignificant difference (P < 0.05; n = 15).
March 2020 | Volume 11 | Article 137
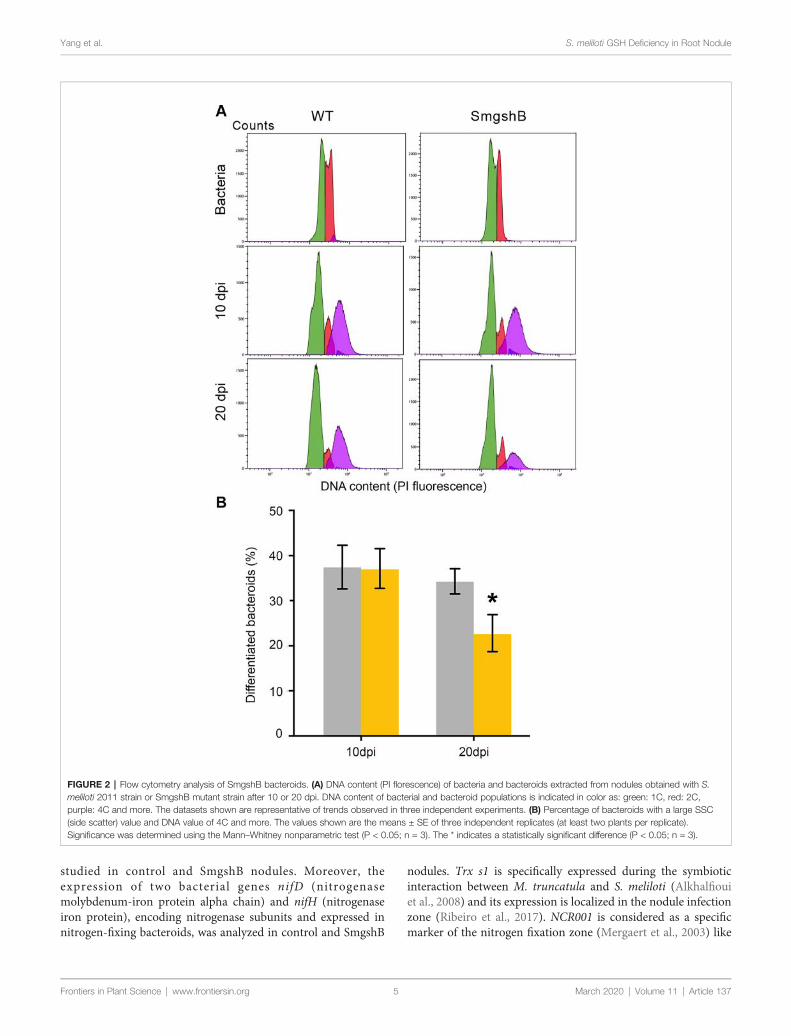
Yang et al. S. meliloti GSH Deficiency in Root Nodule
studied in control and SmgshB nodules. Moreover, theexpression of two bacterial genes nifD (nitrogenasemolybdenum-iron protein alpha chain) and nifH (nitrogenaseiron protein), encoding nitrogenase subunits and expressed innitrogen-fixing bacteroids, was analyzed in control and SmgshB
Frontiers in Plant Science | www.frontiersin.org 5
nodules. Trx s1 is specifically expressed during the symbioticinteraction between M. truncatula and S. meliloti (Alkhalfiouiet al., 2008) and its expression is localized in the nodule infectionzone (Ribeiro et al., 2017). NCR001 is considered as a specificmarker of the nitrogen fixation zone (Mergaert et al., 2003) like
FIGURE 2 | Flow cytometry analysis of SmgshB bacteroids. (A) DNA content (PI florescence) of bacteria and bacteroids extracted from nodules obtained with S.meliloti 2011 strain or SmgshB mutant strain after 10 or 20 dpi. DNA content of bacterial and bacteroid populations is indicated in color as: green: 1C, red: 2C,purple: 4C and more. The datasets shown are representative of trends observed in three independent experiments. (B) Percentage of bacteroids with a large SSC(side scatter) value and DNA value of 4C and more. The values shown are the means ± SE of three independent replicates (at least two plants per replicate).Significance was determined using the Mann–Whitney nonparametric test (P < 0.05; n = 3). The * indicates a statistically significant difference (P < 0.05; n = 3).
March 2020 | Volume 11 | Article 137
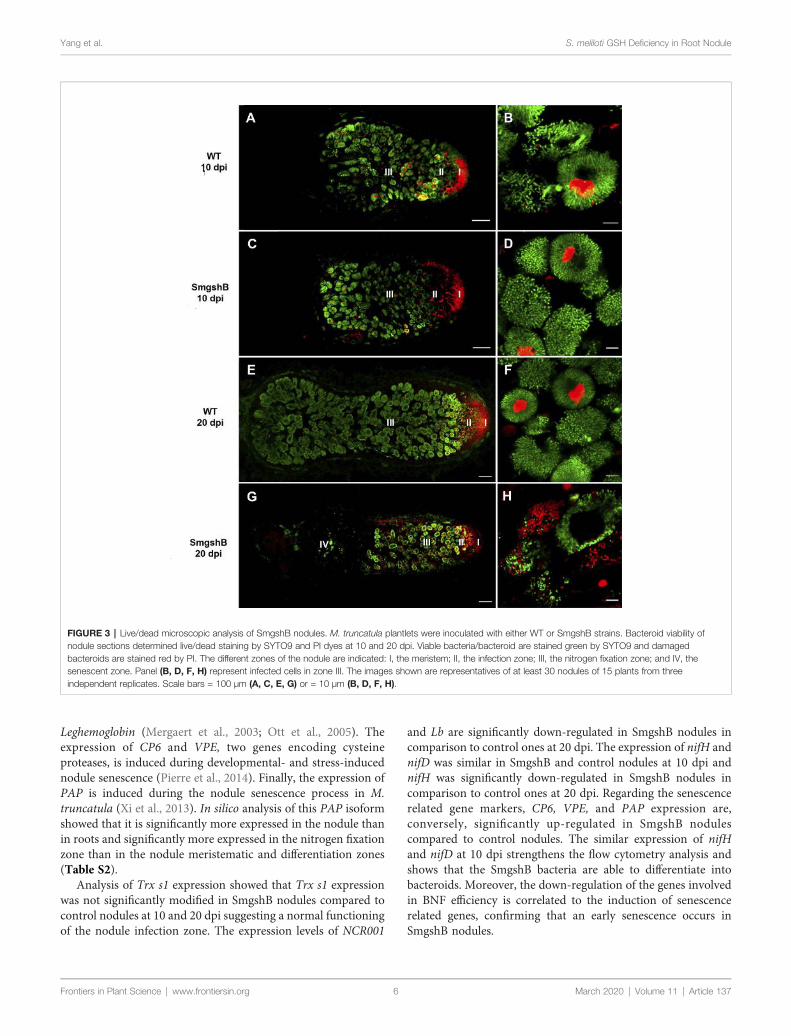
Yang et al. S. meliloti GSH Deficiency in Root Nodule
Leghemoglobin (Mergaert et al., 2003; Ott et al., 2005). Theexpression of CP6 and VPE, two genes encoding cysteineproteases, is induced during developmental- and stress-inducednodule senescence (Pierre et al., 2014). Finally, the expression ofPAP is induced during the nodule senescence process in M.truncatula (Xi et al., 2013). In silico analysis of this PAP isoformshowed that it is significantly more expressed in the nodule thanin roots and significantly more expressed in the nitrogen fixationzone than in the nodule meristematic and differentiation zones(Table S2).
Analysis of Trx s1 expression showed that Trx s1 expressionwas not significantly modified in SmgshB nodules compared tocontrol nodules at 10 and 20 dpi suggesting a normal functioningof the nodule infection zone. The expression levels of NCR001
Frontiers in Plant Science | www.frontiersin.org 6
and Lb are significantly down-regulated in SmgshB nodules incomparison to control ones at 20 dpi. The expression of nifH andnifD was similar in SmgshB and control nodules at 10 dpi andnifH was significantly down-regulated in SmgshB nodules incomparison to control ones at 20 dpi. Regarding the senescencerelated gene markers, CP6, VPE, and PAP expression are,conversely, significantly up-regulated in SmgshB nodulescompared to control nodules. The similar expression of nifHand nifD at 10 dpi strengthens the flow cytometry analysis andshows that the SmgshB bacteria are able to differentiate intobacteroids. Moreover, the down-regulation of the genes involvedin BNF efficiency is correlated to the induction of senescencerelated genes, confirming that an early senescence occurs inSmgshB nodules.
FIGURE 3 | Live/dead microscopic analysis of SmgshB nodules. M. truncatula plantlets were inoculated with either WT or SmgshB strains. Bacteroid viability ofnodule sections determined live/dead staining by SYTO9 and PI dyes at 10 and 20 dpi. Viable bacteria/bacteroid are stained green by SYTO9 and damagedbacteroids are stained red by PI. The different zones of the nodule are indicated: I, the meristem; II, the infection zone; III, the nitrogen fixation zone; and IV, thesenescent zone. Panel (B, D, F, H) represent infected cells in zone III. The images shown are representatives of at least 30 nodules of 15 plants from threeindependent replicates. Scale bars = 100 µm (A, C, E, G) or = 10 µm (B, D, F, H).
March 2020 | Volume 11 | Article 137
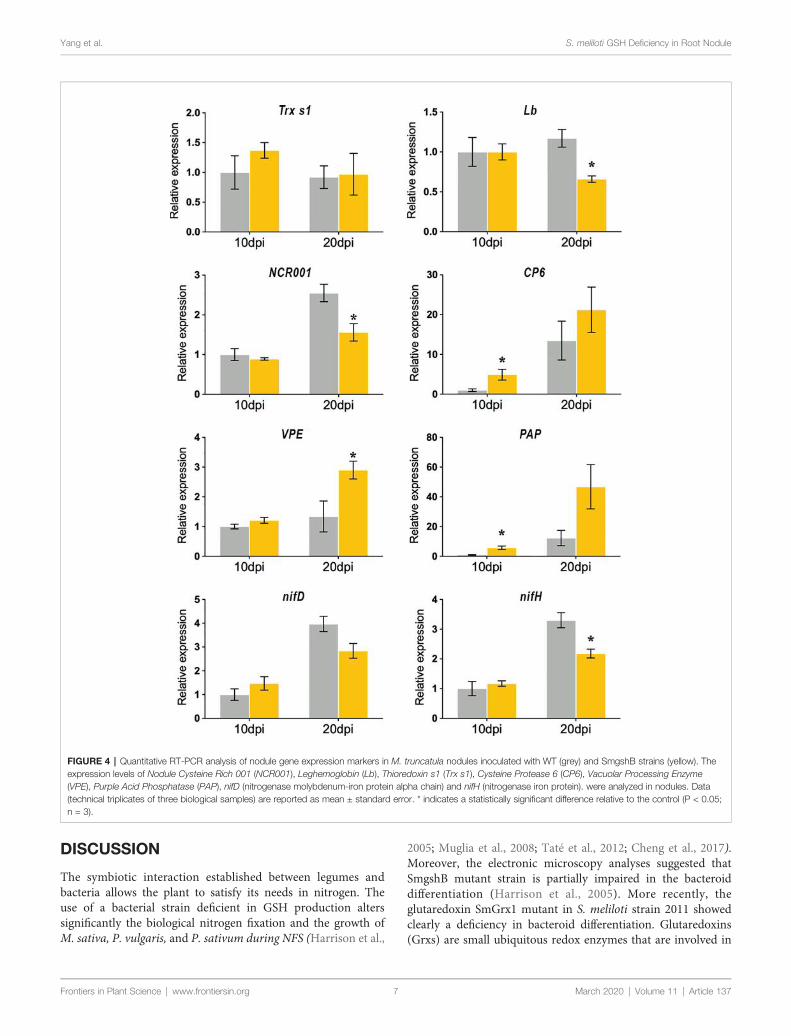
Yang et al. S. meliloti GSH Deficiency in Root Nodule
DISCUSSION
The symbiotic interaction established between legumes andbacteria allows the plant to satisfy its needs in nitrogen. Theuse of a bacterial strain deficient in GSH production alterssignificantly the biological nitrogen fixation and the growth ofM. sativa, P. vulgaris, and P. sativum during NFS (Harrison et al.,
Frontiers in Plant Science | www.frontiersin.org 7
2005; Muglia et al., 2008; Taté et al., 2012; Cheng et al., 2017).Moreover, the electronic microscopy analyses suggested thatSmgshB mutant strain is partially impaired in the bacteroiddifferentiation (Harrison et al., 2005). More recently, theglutaredoxin SmGrx1 mutant in S. meliloti strain 2011 showedclearly a deficiency in bacteroid differentiation. Glutaredoxins(Grxs) are small ubiquitous redox enzymes that are involved in
FIGURE 4 | Quantitative RT-PCR analysis of nodule gene expression markers in M. truncatula nodules inoculated with WT (grey) and SmgshB strains (yellow). Theexpression levels of Nodule Cysteine Rich 001 (NCR001), Leghemoglobin (Lb), Thioredoxin s1 (Trx s1), Cysteine Protease 6 (CP6), Vacuolar Processing Enzyme(VPE), Purple Acid Phosphatase (PAP), nifD (nitrogenase molybdenum-iron protein alpha chain) and nifH (nitrogenase iron protein). were analyzed in nodules. Data(technical triplicates of three biological samples) are reported as mean ± standard error. * indicates a statistically significant difference relative to the control (P < 0.05;n = 3).
March 2020 | Volume 11 | Article 137

Yang et al. S. meliloti GSH Deficiency in Root Nodule
the reduction of oxidative modifications using GSH (Benyaminaet al., 2013). Thus, our hypothesis is that GSH produced by thestrain 2011 is necessary to allow normal bacteroid differentiationin nodules formed on M. truncatula. In this context, we testedthe symbiotic interaction between M. truncatula and the GSHdeficient-SmgshB mutant constructed in S. meliloti strain 2011 at10 and 20 dpi, two early time points of nodule nitrogen fixation,using a multidisciplinary approach.
The nitrogen fixation efficiency was significantly impaired inplants inoculated with SmgshB strain (Figure 1). However, thisreduction is less noticeable than in plants inoculated with SmGrx1(Benyamina et al., 2013). Moreover, at 20 dpi, elongated nodulesare observed suggesting that the nodule meristem is more active inplants inoculated with SmgshB strain than in plants inoculatedwith SmGrx1 strain (Benyamina et al., 2013). InM. truncatula, thebacteroids present a terminal differentiation accompanied with aDNA endoreduplication and an arrest in cell division coupled toan increase in cell size (Ribeiro et al., 2015). The flow cytometryanalysis of SmgshB bacteroids shows that DNA endoreduplicationoccurs in SmgshB bacteroid and no significant difference wasobserved between SmgshB nodules and control ones at 10 dpi(Figure 2). In contrast, the number of terminally differentiatedbacteroids is significantly lower in SmgshB nodules than in controlones at 20 dpi suggesting that there is a faster degradation or aslower formation of bacteroids in SmgshB nodules than in controlones. The live/dead experiments allow us to analyze the viability ofthe bacteroids in the root nodules. Such live/dead labelling showsan early death of bacteroids in SmgshB nodules compared tocontrol ones at 20 dpi (Figure 3).
Gene expression analysis showed that at 10 dpi, genesexpressed in the nitrogen fixing zone, NCR001 and Lb, aresimilarly expressed in SmgshB and control nodules suggesting
Frontiers in Plant Science | www.frontiersin.org 8
that the gene expression in the nitrogen fixing zone is notsignificantly affected (Figure 4). A similar result is observed forTrx s1, a gene marker for the nodule infection zone, and VPE.Similarly, expression of nifH and nifD genes was similar inSmgshB and control nodules strengthening that SmgshBbacteroid differentiation occurs properly at 10 dpi. In contrast,significant higher expression of CP6 and PAP is already observedin SmgshB nodules compared to control ones at 10 dpi. Thisresult showed that in our plant growth conditions, CP6 and PAPare the earlier senescence marker genes whose expression issignificantly modified, suggesting that there is an earlydevelopment of senescence in SmgshB nodules. At 20 dpi,there is a significant modification in the expression of differentgenes. NCR001, Lb, and nifH are less expressed in SmgshBnodules compared to control ones showing a correlationbetween the diminution of the nitrogen fixation efficiency andthe reduction of the expression of genes expressed in the nitrogenfixation zone. Despite a strong increase of CP6 expression in 20dpi mature SmgshB nodule, there is no more a significantdifference compared to the control suggesting that a senescenceprocess begins in control nodules in our conditions. In contrast,VPE expression is significantly induced in SmgshB noduleindicating the progression of the senescence at 20 dpi asobserved in the live/dead analysis.
Taken together, our data show that, in contrast to SmGrx1bacterial mutant (strain 2011), SmgshB mutant (strain 2011) isable to differentiate in bacteroids inside M. truncatula cells. Thephenotype observed is similar to the one obtained during theinteraction of SmgshB mutant (strain 1021) with M. sativa(Harrison et al., 2005). In this context, SmGrx1 should useanother reducing power than glutathione to perform itsactivity. We hypothesize that the Grx1 could use the g-ECaccumulated in high quantity in the SmgshB mutant strain ascofactor for the reductive power instead of GSH. Differentphenotypes were observed for Arabidopsis thaliana g-ECS(gsh1) and GSHS (gsh2) plant mutants. The ghs1 mutantpresents an embryo lethal phenotype whereas the gsh2 mutanthas a seedling lethal phenotype suggesting a partial replacementof GSH functions by g-EC (Cairns et al., 2006; Pasternak et al.,2008). A similar observation is also true for S. meliloti strain 1021in which the SmgshA mutant strain cannot divide while theSmgshB mutant strain shows a growth delay under free livingconditions (Harrison et al., 2005). g-EC is also used as a cofactorby glutathione peroxidase-1 to detoxify ROS in mice (Quintana-Cabrera et al., 2012).
The correlation between the GSH content and the BNF invarious legumes was the object of several works (Dalton et al.,1986; Evans et al., 1999; Matamoros et al., 1999; Moran et al.,2000; Groten et al., 2005). The plant GSH content is crucial forthe bacterial infection effectiveness and the nitrogen fixationefficiency. GSH produced in the plant does not compensate thegshB mutation suggesting that it is not able to be transported tothe bacteroid (Becana et al., 2018). In this context, it should bementioned that sulfate transport from the plant cell to thebacteroids is very active (Schneider et al., 2019) and thatmutation in the sulfate transporter SST1 in Lotus japonicus
FIGURE 5 | General overview of GSH role in S. meliloti bacteroid. GSH playsa central role in the redox networks of glutaredoxin (SmGrx1) and SmLsrB.Defect in GSH accumulation lead to an early nodule senescence whereasdeletion in Grx1 impairs bacteroid differentiation. The green color indicatesgeneral effects on the root nodule and the orange color indicates directeffects on the bacteroid. Dashed arrow indicates the potential role of g-EC onSmGrx1 in absence of GSH.
March 2020 | Volume 11 | Article 137

Yang et al. S. meliloti GSH Deficiency in Root Nodule
impairs symbiotic nitrogen fixation (Krusell et al., 2005). Thesedata suggest that the sulfur metabolism in bacteroid is dependingon sulfate transport and that other sulfur containing moleculessuch as methionine, cysteine or GSH are not transported throughthe symbiosome membrane. In this sense, the R. etli metZ geneinvolved in methionine synthesis is required for the efficientinteraction with P. vulgaris suggesting that methionine isproduced by R. etli bacteroids during symbiosis, and notsupplied by the host cells (Taté et al., 1999). In contrast, thecysGmutant strain of R. etli, unable to grow on sulfate as the solesulfur source, was able to induce nitrogen-fixing nodules on P.vulgaris suggesting that cysteine or glutathione is supplied by theplant to bacteria (Taté et al., 1997). Moreover, the nodulesinduced by cysteine auxotrophs of S. meliloti Rmd201 werefully effective like those of the parental strain-induced nodulessuggesting that the alfalfa host provides cysteine to the bacteroid(Abbas et al., 2002). However, even if gshB mutant strains showan impaired symbiotic phenotype, it cannot be excluded that asmall fraction of GSH produced in the plant may pass throughthe bacteroid membrane, partially compensating the bacterialdeficiency. The bacterial GSH pool plays also a crucial role fornitrogen fixation efficiency (Figure 5). In the gshB mutant of S.meliloti strain 1021, the GSH deficiency was associated with ahigher catalase activity in the bacterial mutant in free-livingconditions and a reduction of growth efficiency (Harrison et al.,2005). A decrease in the BNF was observed during the symbiosiswithM. sativa correlated to an early bacteroid death. In R. tropicigshBmutant strain, the lower symbiotic efficiency was associatedto an early senescence pattern connected to increased levels ofsuperoxide accumulation (Muglia et al., 2008). For Rhizobiumetli gshB mutant, the decrease in BNF was correlated to a strongreduction in nodule number and a deficiency of glutaminetransport through Aap and Bra transporter (Taté et al., 2012).This deficiency in nutrient transport also detected in R.leguminosarum 3841, which fails to efficiently use variouscarbon sources such as glucose, succinate, glutamine andhistidine, may trigger an early nodule senescence. Thistransport deficiency in R. leguminosarum 3841 gshB mutantstrain is also associated to a lower BNF (Cheng et al., 2017).The LysR-type transcriptional regulator LsrB, required fornodulation, also regulates gshA gene expression throughmodification of cysteine residues (Luo et al., 2005; Tang et al.,2018) and gshB gene expression (Lu et al., 2013).
In conclusion, the inoculation ofM. truncatulawith the SmgshBbacterial mutant triggers the development of nodules with a reducedcapacity to fix nitrogen. This lower BNF does not seem correlated toa reduced bacteroid differentiation efficiency. However, it is strongly
Frontiers in Plant Science | www.frontiersin.org 9
correlated to an early senescence process and the alteration of theexpression of multiple nodule gene markers showing thefundamental role of the bacterial GSH pool in nodule functioning.
DATA AVAILABILITY STATEMENT
The datasets generated for this study are available on request tothe corresponding author.
AUTHOR CONTRIBUTIONS
Conceived and designed the experiments: LY, SE, EB, PF.Performed the experiments: LY, SE, SB, AL, JH, JC, OP, EB.Analyzed the data: LY, SE, EB, PF. Wrote the paper: LY, SE, EB,PF. Amended the paper: SAS, DH.
FUNDING
LY is supported by a doctoral fellowship from the Chinascholarship council (CSC). SE was financed by IMAGEENprogram from the European Communities and the Tunisiangovernment (bourse d'alternance). This work was supported bythe “Institut National de la Recherche Agronomique”, the“Centre National de la Recherche Scientifique”, the Universityof Nice-Sophia Antipolis and the French Government (NationalResearch Agency, ANR) through the “STAYPINK” project(reference # ANR-15-CE20-0005) and the “Investments for theFuture” LABEX SIGNALIFE: program reference # ANR-11-LABX-0028-01.
ACKNOWLEDGMENTS
We are largely indebted to the symbiosis team members fornumerous nodule harvests. We thank Renaud Brouquisse forcritical reading of the manuscript.
SUPPLEMENTARY MATERIAL
The Supplementary Material for this article can be found onlineat: https://www.frontiersin.org/articles/10.3389/fpls.2020.00137/full#supplementary-material
REFERENCES
Abbas, B. A., Vineetha, K. E., Prasad, C. K., Vij, N., Hassani, R., and Randhawa, G. S.(2002). Symbiotic characteristics of cysteine and methionine auxotrophs ofSinorhizobium meliloti. Indian J. Exp. Biol. 40, 1121–1130.
Alkhalfioui, F., Renard, M., Frendo, P., Keichinger, C., Meyer, Y., Gelhaye, E., et al.(2008). A novel type of thioredoxin dedicated to symbiosis in legumes. PlantPhysiol. 148, 424–435. doi: 10.1104/pp.108.123778
Andrio, E., Marino, D., Marmeys, A., De Segonzac, M. D., Damiani, I., Genre, A.,et al. (2013). Hydrogen peroxide-regulated genes in the Medicago truncatula–Sinorhizobium meliloti symbiosis. New Phytol. 198, 179–189. doi: 10.1111/nph.12120
Arthikala,M. K., Sanchez-Lopez, R., Nava, N., Santana, O., Cardenas, L., andQuinto, C.(2014). RbohB, a Phaseolus vulgaris NADPH oxidase gene, enhances symbiosomenumber, bacteroid size, and nitrogen fixation in nodules and impairs mycorrhizalcolonization. New Phytol. 202, 886–900. doi: 10.1111/nph.12714
March 2020 | Volume 11 | Article 137

Yang et al. S. meliloti GSH Deficiency in Root Nodule
Becana, M., Wienkoop, S., and Matamoros, M. (2018). Sulfur transport andmetabolism in legume root nodules. Front. Plant Sci. 9, 1434. doi: 10.3389/fpls.2018.01434
Benyamina, S. M., Baldacci-Cresp, F., Couturier, J., Chibani, K., Hopkins, J.,Bekki, A., et al. (2013). Two Sinorhizobium meliloti glutaredoxins regulate ironmetabolism and symbiotic bacteroid differentiation. Environ. Microbiol. 15,795–810. doi: 10.1111/j.1462-2920.2012.02835.x
Berger, A., Boscari, A., Frendo, P., and Brouquisse, R. (2019). Nitric oxidesignaling, metabolism and toxicity in nitrogen-fixing symbiosis. J. Exp. Bot.70, 4505–4520. doi: 10.1093/jxb/erz159
Boisson-Dernier, A., Chabaud, M., Garcia, F., Becard, G., Rosenberg, C., and Barker,D. G. (2001). Agrobacterium rhizogenes-transformed roots of Medicago truncatulafor the study of nitrogen-fixing and endomycorrhizal symbiotic associations.Mol. Plant Microbe Interact. 14, 695–700. doi: 10.1094/MPMI.2001.14.6.695
Cárdenas, L., Martínez, A., Sánchez, F., and Quinto, C. (2008). Fast, transient andspecific intracellular ROS changes in living root hair cells responding to Nodfactors (NFs). Plant J. 56, 802–813. doi: 10.1111/j.1365-313X.2008.03644.x
Cairns, N. G., Pasternak, M., Wachter, A., Cobbett, C. S., and Meyer, A. J. (2006).Maturation of Arabidopsis seeds is dependent on glutathione biosynthesiswithin the embryo. Plant Physiol. 141, 446–455. doi: 10.1104/pp.106.077982
Cheng, G., Karunakaran, R., East, A. K., Munoz-Azcarate, O., and Poole, P. S.(2017). Glutathione affects the transport activity of Rhizobium leguminosarum3841 and is essential for efficient nodulation. FEMS Microbiol. Lett. 364,fnx045. doi: 10.1093/femsle/fnx045
Dalton, D. A., Russell, S. A., Hanus, F., Pascoe, G. A., and Evans, H. J. (1986).Enzymatic reactions of ascorbate and glutathione that prevent peroxidedamage in soybean root nodules. Proc. Natl. Acad. Sci. U.S.A. 83, 3811–3815. doi: 10.1073/pnas.83.11.3811
El Msehli, S., Lambert, A., Baldacci-Cresp, F., Hopkins, J., Boncompagni, E., Smiti,S. A., et al. (2011). Crucial role of (homo)glutathione in nitrogen fixation inMedicago truncatula nodules. New Phytol. 192, 496–506. doi: 10.1111/j.1469-8137.2011.03810.x
Evans, P. J., Gallesi, D., Mathieu, C., Hernandez, M. J., De Felipe, M., Halliwell, B.,et al. (1999). Oxidative stress occurs during soybean nodule senescence. Planta208, 73–79. doi: 10.1007/s004250050536
Foyer, C. H., and Noctor, G. (2011). Ascorbate and glutathione: the heart of theredox hub. Plant Physiol. 155, 2–18. doi: 10.1104/pp.110.167569
Frendo, P., Harrison, J., Norman, C., Hernandez Jimenez, M. J., Van De Sype, G.,Gilabert, A., et al. (2005). Glutathione and homoglutathione play a critical rolein the nodulation process ofMedicago truncatula.Mol. Plant Microbe Interact.18, 254–259. doi: 10.1094/MPMI-18-0254
Frendo, P., Baldacci-Cresp, F., Benyamina, S. M., and Puppo, A. (2013a).Glutathione and plant response to the biotic environment. Free. Radical. Bio.Med. 65, 724–730. doi: 10.1016/j.freeradbiomed.2013.07.035
Frendo, P., Matamoros, M. A., Alloing, G., and Becana, M. (2013b). Thiol-basedredox signaling in the nitrogen-fixing symbiosis. Front. Plant Sci. 4, 376. doi:10.3389/fpls.2013.00376
Groten, K., Vanacker, H., Dutilleul, C., Bastian, F., Bernard, S., Carzaniga, R., et al.(2005). The roles of redox processes in pea nodule development andsenescence. Plant Cell Environ. 28, 1293–1304. doi: 10.1111/j.1365-3040.2005.01376.x
Haag, A. F., Baloban, M., Sani, M., Kerscher, B., Pierre, O., Farkas, A., et al. (2011).Protection of Sinorhizobium against host cysteine-rich antimicrobial peptidesis critical for symbiosis. PloS Biol. 9, e1001169. doi: 10.1371/journal.pbio.1001169
Harrison, J., Jamet, A., Muglia, C. I., Van De Sype, G., Aguilar, O. M., Puppo, A.,et al. (2005). Glutathione plays a fundamental role in growth and symbioticcapacity of Sinorhizobium meliloti. J. Bacteriol. 187, 168–174. doi: 10.1128/JB.187.1.168-174.2005
Hichri, I., Boscari, A., Castella, C., Rovere, M., Puppo, A., and Brouquisse, R.(2015). Nitric oxide: a multifaceted regulator of the nitrogen-fixing symbiosis.J. Exp. Bot. 66, 2877–2887. doi: 10.1093/jxb/erv051
Innocenti, G., Pucciariello, C., Le Gleuher, M., Hopkins, J., De Stefano, M., Delledonne,M., et al. (2007). Glutathione synthesis is regulated by nitric oxide in Medicagotruncatula roots. Planta 225, 1597–1602. doi: 10.1007/s00425-006-0461-3
Jamet, A., Mandon, K., Puppo, A., and Hérouart, D. (2007). H2O2 is required foroptimal establishment of the Medicago sativa/Sinorhizobium melilotisymbiosis. J. Bacteriol. 189, 8741–8745. doi: 10.1128/JB.01130-07
Frontiers in Plant Science | www.frontiersin.org 10
Krusell, L., Krause, K., Ott, T., Desbrosses, G., Krämer, U., Sato, S., et al. (2005).The sulfate transporter SST1 is crucial for symbiotic nitrogen fixation in Lotusjaponicus root nodules. Plant Cell. 17, 1625–1636. doi: 10.1105/tpc.104.030106
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression datausing real-time quantitative PCR and the 2–DDCT method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Lu, D., Tang, G., and Luo, L. (2013). The Sinorhizobium meliloti LysR familytranscriptional factor LsrB is involved in the regulation of glutathionebiosynthesis. Acta Biochim. Biophys. Sin. (Shanghai) 45, 882–888. doi:10.1093/abbs/gmt083
Luo, L., Yao, S. Y., Becker, A., Rüberg, S., Yu, G. Q., Zhu, J. B., et al. (2005). Twonew Sinorhizobium meliloti LysR-type transcriptional regulators required fornodulation. J. Bacteriol. 187, 4562–4572. doi: 10.1128/JB.187.13.4562-4572.2005
Marino, D., Andrio, E., Danchin, E. G., Oger, E., Gucciardo, S., Lambert, A., et al.(2011). A Medicago truncatula NADPH oxidase is involved in symbioticnodule functioning. New Phytol. 189, 580–592. doi: 10.1111/j.1469-8137.2010.03509.x
Marino, D., Dunand, C., Puppo, A., and Pauly, N. (2012). A burst of plantNADPH oxidases . Trends Plant Sc i . 17, 9–15. doi : 10 .1016/j.tplants.2011.10.001
Matamoros, M. A., Moran, J. F., Iturbe-Ormaetxe, I., Rubio, M. C., and Becana, M.(1999). Glutathione and homoglutathione synthesis in legume root nodules.Plant Physiol. 121, 879–888. doi: 10.1104/pp.121.3.879
Mergaert, P., Nikovics, K., Kelemen, Z., Maunoury, N., Vaubert, D., Kondorosi, A.,et al. (2003). A novel family in Medicago truncatula consisting of more than300 nodule-specific genes coding for small, secreted polypeptides withconserved cysteine motifs. Plant Physiol. 132, 161–173. doi: 10.1104/pp.102.018192
Mergaert, P., Uchiumi, T., Alunni, B., Evanno, G., Cheron, A., Catrice, O., et al.(2006). Eukaryotic control on bacterial cell cycle and differentiation in theRhizobium–legume symbiosis. Proc. Natl. Acad. Sci. U.S.A. 103, 5230–5235.doi: 10.1073/pnas.0600912103
Moran, J. F., Iturbe-Ormaetxe, I., Matamoros, M. A., Rubio, M. C., Clemente, M. R.,Brewin, N. J., et al. (2000). Glutathione and homoglutathione synthetases of legumenodules. cloning, expression, and subcellular localization. Plant Physiol. 124, 1381–1392. doi: 10.1104/pp.124.3.1381
Muglia, C., Comai, G., Spegazzini, E., Riccillo, P. M., and Aguilar, O. M. (2008).Glutathione produced by Rhizobium tropici is important to prevent earlysenescence in common bean nodules. FEMS Microbiol. Lett. 286, 191–198. doi:10.1111/j.1574-6968.2008.01285.x
Ott, T., Van Dongen, J. T., Gu, C., Krusell, L., Desbrosses, G., Vigeolas, H., et al.(2005). Symbiotic leghemoglobins are crucial for nitrogen fixation in legumeroot nodules but not for general plant growth and development. Curr. Biol. 15,531–535. doi: 10.1016/j.cub.2005.01.042
Pasternak, M., Lim, B., Wirtz, M., Hell, R., Cobbett, C. S., and Meyer, A. J.(2008). Restricting glutathione biosynthesis to the cytosol is sufficient fornormal plant development. Plant J. 53, 999–1012. doi: 10.1111/j.1365-313X.2007.03389.x
Pierre, O., Hopkins, J., Combier, M., Baldacci, F., Engler, G., Brouquisse, R., et al.(2014). Involvement of papain and legumain proteinase in the senescenceprocess of Medicago truncatula nodules. New Phytol. 202, 849–863. doi:10.1111/nph.12717
Puppo, A., Pauly, N., Boscari, A., Mandon, K., and Brouquisse, R. (2013).Hydrogen peroxide and nitric oxide: key regulators of the Legume-Rhizobium and mycorrhizal symbioses. Antioxid. Redox Sign. 18, 2202–2219.doi: 10.1089/ars.2012.5136
Quintana-Cabrera, R., Fernandez-Fernandez, S., Bobo-Jimenez, V., Escobar, J.,Sastre, J., Almeida, A., et al. (2012). g-Glutamylcysteine detoxifies reactiveoxygen species by acting as glutathione peroxidase-1 cofactor. Nat. Commun.3, 718. doi: 10.1038/ncomms1722
Ribeiro, C. W., Alloing, G., Mandon, K., and Frendo, P. (2015). Redox regulationof differentiation in symbiotic nitrogen fixation. Biochim. Biophys. Acta 1850,1469–1478. doi: 10.1016/j.bbagen.2014.11.018
Ribeiro, C. W., Baldacci-Cresp, F., Pierre, O., Larousse, M., Benyamina, S.,Lambert, A., et al. (2017). Regulation of differentiation of nitrogen-fixingbacteria by microsymbiont targeting of plant thioredoxin s1. Curr. Biol. 27,250–256. doi: 10.1016/j.cub.2016.11.013
March 2020 | Volume 11 | Article 137

Yang et al. S. meliloti GSH Deficiency in Root Nodule
Rouhier, N., Lemaire, S. D., and Jacquot, J.-P. (2008). The role of glutathione inphotosynthetic organisms: emerging functions for glutaredoxins andglutathionylation. Annu. Rev. Plant Biol. 59, 143–166. doi: 10.1146/annurev.arplant.59.032607.092811
Santos, R., Hérouart, D., Sigaud, S., Touati, D., and Puppo, A. (2001). Oxidativeburst in alfalfa-Sinorhizobium meliloti symbiotic interaction. Mol. PlantMicrobe Interact. 14, 86–89. doi: 10.1094/MPMI.2001.14.1.86
Schindelin, J., Arganda-Carreras, I., Frise, E., Kaynig, V., Longair, M., Pietzsch, T.,et al. (2012). Fiji: an open-source platform for biological-image analysis. Nat.Methods 9, 676–682. doi: 10.1038/nmeth.2019
Schneider, S., Schintlmeister, A., Becana, M., Wagner, M., Woebken, ,. D., andWienkoop, S. (2019). Sulfate is transported at significant rates through thesymbiosome membrane and is crucial for nitrogenase biosynthesis. Plant CellEnviron. 42, 1180–1189. doi: 10.3389/fpls.2018.01434
Tang, G., Li, N., Liu, Y., Yu, L., Yan, J., and Luo, L. (2018). Sinorhizobium melilotiglutathione reductase is required for both redox homeostasis and symbiosis.Appl. Environ. Microb. 84, e01937–e01917. doi: 10.1128/aem.01937-17
Taté, R., Riccio, A., Iaccarino, M., and Patriarca, E. J. (1997). A cysG mutant strainof Rhizobium etli pleiotropically defective in sulfate and nitrate assimilation. J.Bacteriol. 179, 7343–7350. doi: 10.1128/jb.179.23.7343-7350.1997
Taté, R., Riccio, A., Caputo, E., Iaccarino, M., and Patriarca, E. J. (1999). TheRhizobium etli metZ gene is essential for methionine biosynthesis andnodulation of Phaseolus vulgaris. Mol. Plant Microbe Interact. 1999 12, 24–34. doi: 10.1094/MPMI.1999.12.1.24
Frontiers in Plant Science | www.frontiersin.org 11
Taté, R., Cermola, M., Riccio, A., Diez-Roux, G., and Patriarca, E. J. (2012).Glutathione is required by Rhizobium etli for glutamine utilization andsymbiotic effectiveness. Mol. Plant Microbe Interact. 25, 331–340. doi:10.1094/MPMI-06-11-0163
Trinchant, J.-C., Boscari, A., Spennato, G., Van de Sype, G., and Le Rudelier, D.(2004). Proline betaine accumulation in alfalfa plants under sodium chloridestress. Exploring its compartmentalization in nodules. Plant Physiol. 135,1583–1594. doi: 10.1104/pp.103.037556
Xi, J., Chen, Y., Nakashima, J., Wang, S. M., and Chen, R. (2013). Medicagotruncatula esn1 defines a genetic locus involved in nodule senescence andsymbiotic nitrogen fixation. Mol. Plant Microbe Interact. 26, 893–902. doi:10.1094/MPMI-02-13-0043-R
Conflict of Interest: The authors declare that the research was conducted in theabsence of any commercial or financial relationships that could be construed as apotential conflict of interest.
Copyright © 2020 Yang, El Msehli, Benyamina, Lambert, Hopkins, Cazareth, Pierre,Hérouart, Achi-Smiti, Boncompagni and Frendo. This is an open-access articledistributed under the terms of the Creative Commons Attribution License (CC BY).The use, distribution or reproduction in other forums is permitted, provided theoriginal author(s) and the copyright owner(s) are credited and that the originalpublication in this journal is cited, in accordance with accepted academic practice. Nouse, distribution or reproduction is permitted which does not comply with these terms.
March 2020 | Volume 11 | Article 137
1
Table S1: List of primers used to quantify gene expression levels by qRT-PCR
Genes Forward primers Reverse primers
Mtc27 TGAGGGAGCAACCAAATACC GCCAAAACCAAGCTACCATC
A38 TCGTGGTGGTGGTTATCAAA TTCAGACCTTCCCATTGACA
16S GATAAGCCGAGAGGAAGGTG GTGTAGCCCAGCCCGTAAG
16S ribosomal RNA
PNP
GGACGGCTACAAGAACGACAC TCATGCTGCATGACCGTGAT (Smc00324)
Polyribonucleotide nucleotidyl transferase
nifD TCCGACTTTCAGGAGAAGGA CGATGTCGTCACCAATCAAG Nitrogenase molybdenum-iron protein
alpha chain nifH
GTTATCACCTCGATCAACTTCCT CGATGTAGATTTCCTGAGCCT Nitrogenase iron protein
Thrxs1 TGTGTTGGTGGAATTCTTCG AGCCACGAGTTTGTTTCGAC
Thioredoxin s1
Lb GAGCGAAGAATTGAGCACTGCT TGCCTTCTTAATTGCAGTTGCC
Leghemoglobin
NCR001 CACTTCTGACGCTAACTG TGAACCTTCAATATACCACC
Nodule-specific Cysteine-Rich peptide 001
CP6 CCTGCTGCTACTATTGCTGGATATG CACTCGCATCAATGGCTACGG
Cysteine Protease 6
VPE CCAGGGGTTCTTGGTATGCCCG ACTGCCAGATTCACATGCCTCCA
Vascular Processing Enzyme
PAP TTTTGCACCGGATTCTTCAC CCAACACCACTACCCAAGGA
Purple Acid Phosphatase
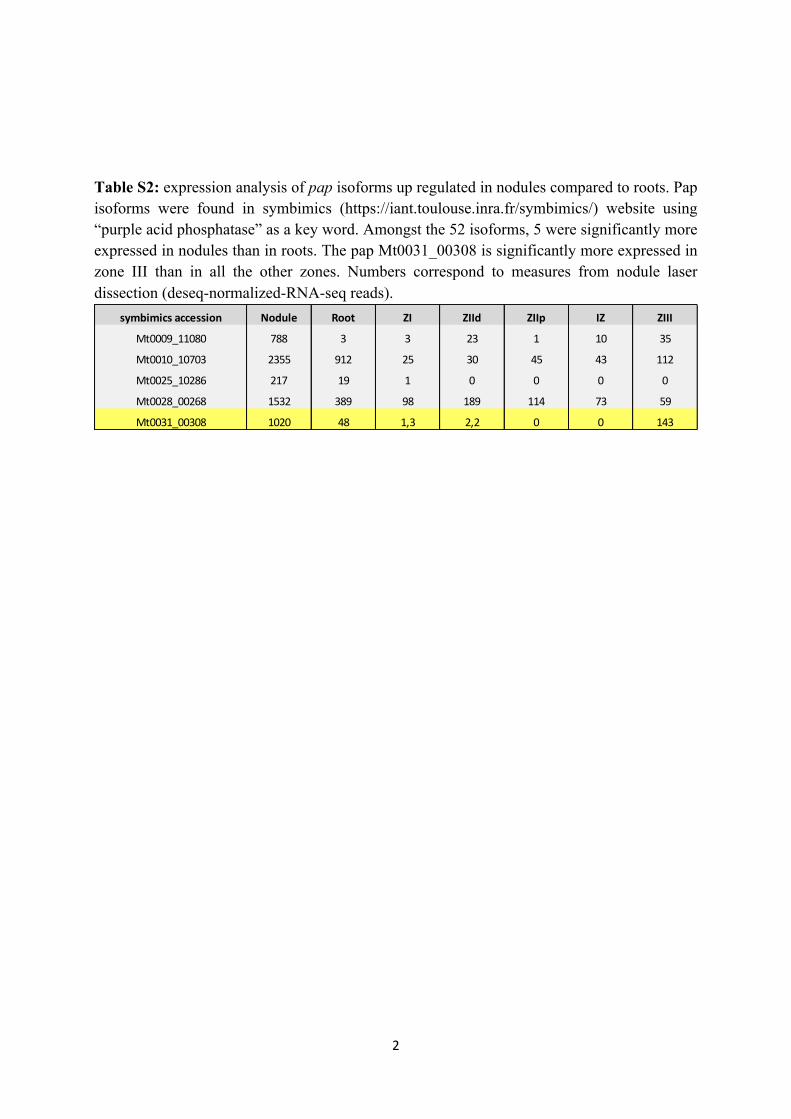
2
Table S2: expression analysis of pap isoforms up regulated in nodules compared to roots. Pap isoforms were found in symbimics (https://iant.toulouse.inra.fr/symbimics/) website using “purple acid phosphatase” as a key word. Amongst the 52 isoforms, 5 were significantly more expressed in nodules than in roots. The pap Mt0031_00308 is significantly more expressed in zone III than in all the other zones. Numbers correspond to measures from nodule laser dissection (deseq-normalized-RNA-seq reads).
symbimics accession Nodule Root ZI ZIId ZIIp IZ ZIII
Mt0009_11080 788 3 3 23 1 10 35
Mt0010_10703 2355 912 25 30 45 43 112
Mt0025_10286 217 19 1 0 0 0 0
Mt0028_00268 1532 389 98 189 114 73 59
Mt0031_00308 1020 48 1,3 2,2 0 0 143
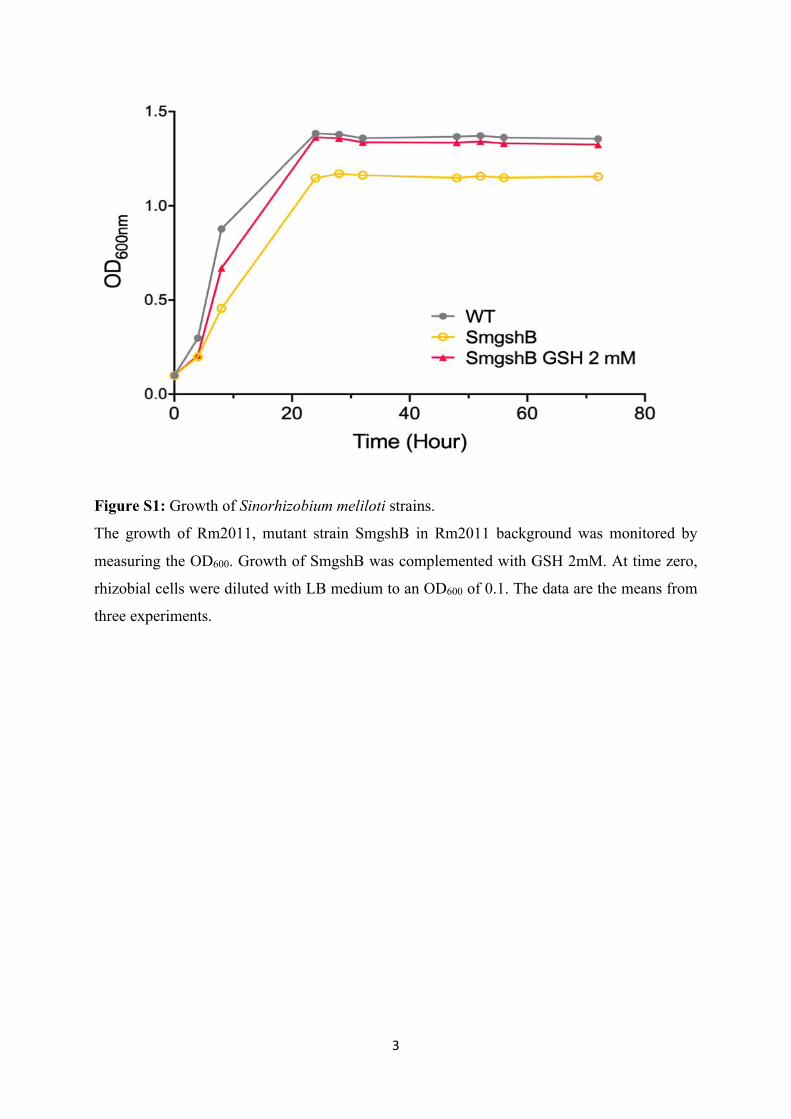
3
Figure S1: Growth of Sinorhizobium meliloti strains.
The growth of Rm2011, mutant strain SmgshB in Rm2011 background was monitored by
measuring the OD600. Growth of SmgshB was complemented with GSH 2mM. At time zero,
rhizobial cells were diluted with LB medium to an OD600 of 0.1. The data are the means from
three experiments.
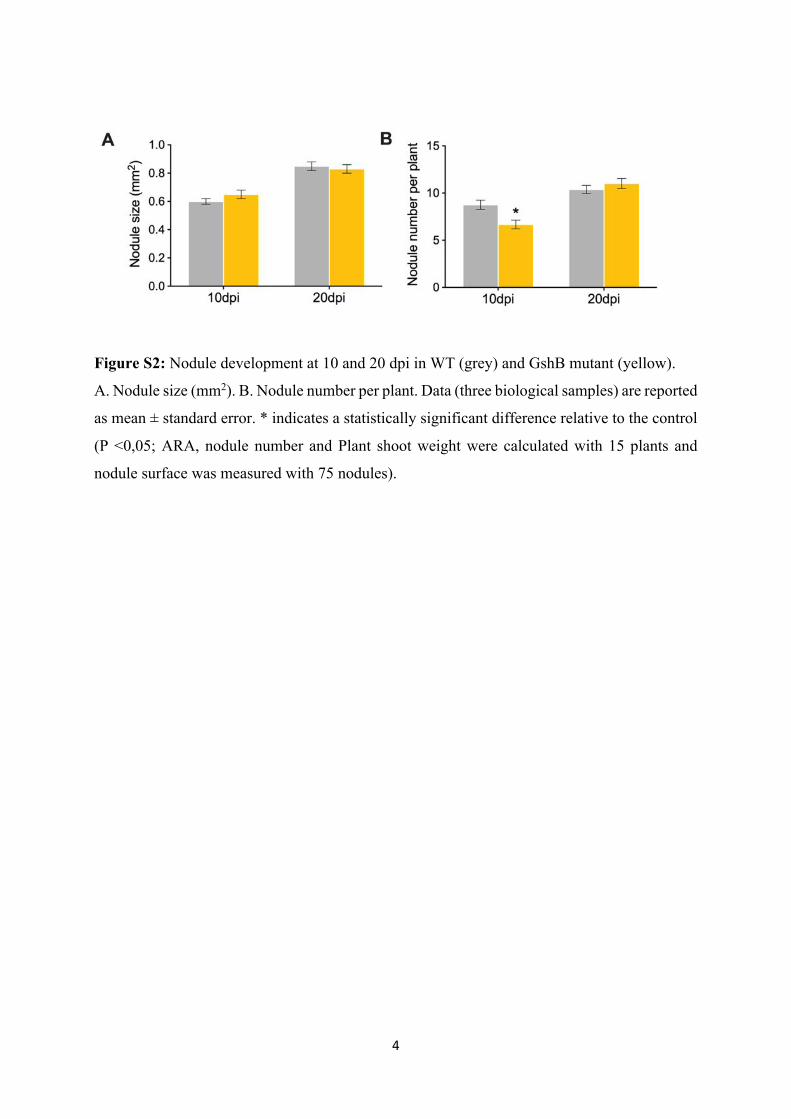
4
Figure S2: Nodule development at 10 and 20 dpi in WT (grey) and GshB mutant (yellow).
A. Nodule size (mm2). B. Nodule number per plant. Data (three biological samples) are reported
as mean ± standard error. * indicates a statistically significant difference relative to the control
(P <0,05; ARA, nodule number and Plant shoot weight were calculated with 15 plants and
nodule surface was measured with 75 nodules).
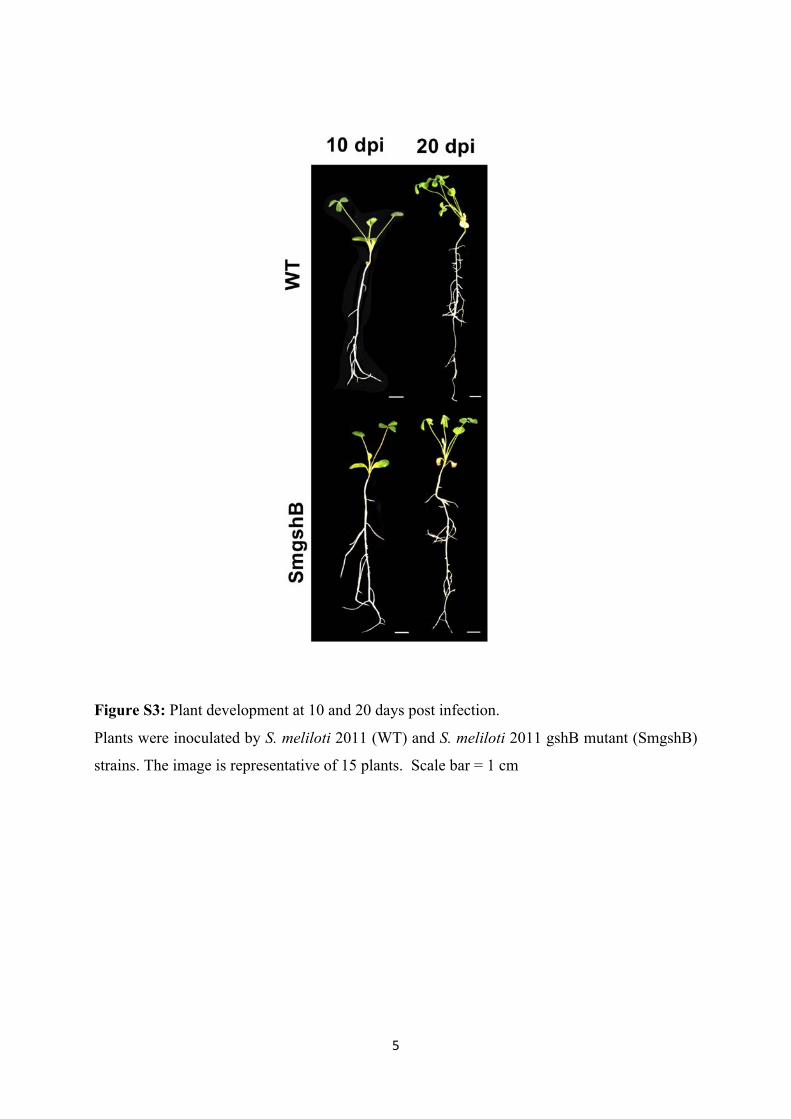
5
Figure S3: Plant development at 10 and 20 days post infection.
Plants were inoculated by S. meliloti 2011 (WT) and S. meliloti 2011 gshB mutant (SmgshB)
strains. The image is representative of 15 plants. Scale bar = 1 cm

CHAPTER I
141
III. Discussion and perspectives
Bacteroid redox homeostasis ensures symbiotic nitrogen fixation
We have characterized a S. meliloti 2011 mutant strain SmgshB which was generated via integration of a plasmid into the genome by single cross-over recombination in the gene gshB, encoding the enzyme glutathione synthase. The SmgshB strain exhibits a significant reduction in the symbiotic nitrogen fixation capacity, consequently diminishing biomass accumulation in plant aerial parts and accelerating the nodule senescence. Similar evidences have been observed with many other rhizobial strain that the GSH deficiency impairs symbiotic nitrogen fixation (Cheng et al., 2017) (Tate et al., 2012) (Muglia et al., 2008) (Harrison et al., 2005). The symbiotic nitrogen fixation is highly sensitive to the redox status at both transcriptional and post-translational level. The nitrogenase is with high redox potential and prone to be damaged rapidly under exposure to oxygen. Besides, the four regulatory proteins of symbiotic nitrogen fixation, NifA, NifL, FixL and RegB, are also oxygen/redox sensors (Dixon & Kahn, 2004). The anaerobic condition is essential for symbiotic nitrogen fixation, whereas GSH is required to activate FNR transcription regulator to form dimer of [4Fe-4S] cluster subunit to control gene expression under anaerobic conditions (Ritz & Beckwith, 2001). It is therefore plausible that the altered redox homeostasis in bacteroid caused by GSH deficiency is detrimental for FNR regulation, thereafter triggering a premature nodule senescence.
GSH deficiency hinders nitrogen-fixing symbiosis
Our study also demonstrates an early senescence phenotype of nodules generated by SmgshB, and we have observed a delayed nodulation phenotype before 10dpi. The symbiotic experiments of the R. tropici CIAT899-13T2 mutant strain show that the mutant strain can induce similar effective nodules albeit outcompeted in co-inoculation with WT strain (Riccillo et al., 2000). The authors later on examined the importance of GSH in symbiosis with the R. tropici gshB mutant which presented a delayed nodulation, reduced weight in aerial part and early senescence phenotype (Muglia et al., 2007; Muglia et al., 2008). The reminiscent phenotype of gshB mutant in R. tropici and S. meliloti implying a conserved functional role of GSH in rhizobia (Riccillo et al., 2000) (Harrison et al., 2005).
The expression of SmgshB culminates at the interzone (IZ; Figure 22), also indicating a function of GSH during bacteroid terminal differentiation step which involved DNA endoreduplication. It has been proven in E. coli that the GSH/glutaredoxin system is involved in DNA synthesis ultimately using the reducing power from NADPH (Ortenberg et al., 2004;
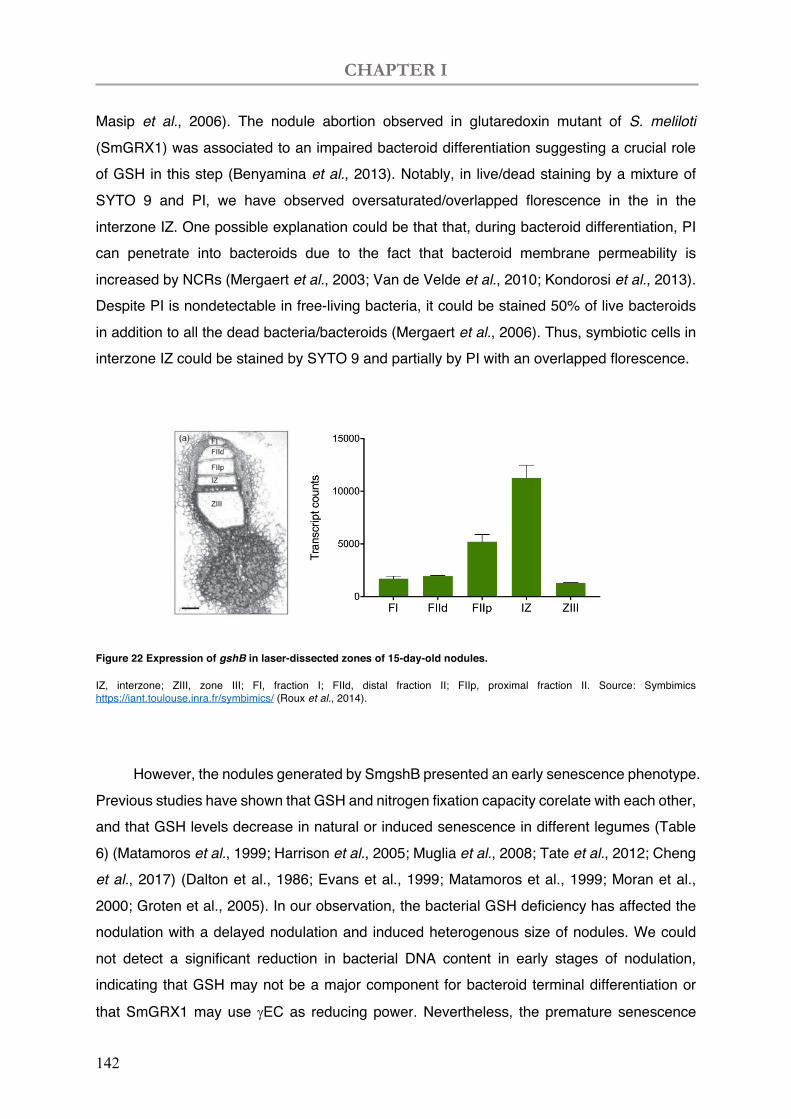
CHAPTER I
142
Masip et al., 2006). The nodule abortion observed in glutaredoxin mutant of S. meliloti (SmGRX1) was associated to an impaired bacteroid differentiation suggesting a crucial role of GSH in this step (Benyamina et al., 2013). Notably, in live/dead staining by a mixture of SYTO 9 and PI, we have observed oversaturated/overlapped florescence in the in the interzone IZ. One possible explanation could be that that, during bacteroid differentiation, PI can penetrate into bacteroids due to the fact that bacteroid membrane permeability is increased by NCRs (Mergaert et al., 2003; Van de Velde et al., 2010; Kondorosi et al., 2013). Despite PI is nondetectable in free-living bacteria, it could be stained 50% of live bacteroids in addition to all the dead bacteria/bacteroids (Mergaert et al., 2006). Thus, symbiotic cells in interzone IZ could be stained by SYTO 9 and partially by PI with an overlapped florescence.
Figure 22 Expression of gshB in laser-dissected zones of 15-day-old nodules.
IZ, interzone; ZIII, zone III; FI, fraction I; FIId, distal fraction II; FIIp, proximal fraction II. Source: Symbimics https://iant.toulouse.inra.fr/symbimics/ (Roux et al., 2014).
However, the nodules generated by SmgshB presented an early senescence phenotype. Previous studies have shown that GSH and nitrogen fixation capacity corelate with each other, and that GSH levels decrease in natural or induced senescence in different legumes (Table 6) (Matamoros et al., 1999; Harrison et al., 2005; Muglia et al., 2008; Tate et al., 2012; Cheng et al., 2017) (Dalton et al., 1986; Evans et al., 1999; Matamoros et al., 1999; Moran et al., 2000; Groten et al., 2005). In our observation, the bacterial GSH deficiency has affected the nodulation with a delayed nodulation and induced heterogenous size of nodules. We could not detect a significant reduction in bacterial DNA content in early stages of nodulation, indicating that GSH may not be a major component for bacteroid terminal differentiation or
that SmGRX1 may use gEC as reducing power. Nevertheless, the premature senescence

CHAPTER I
143
pattern of the SmgshB infected nodules is proven with enhanced green colour in nodules, reduced nitrogen fixation capacity with less biomass accumulated in plants, early senescent bacteroid in live/dead staining and fast degradation in DNA content, as well as elevated gene expression of nodule senescence markers already seen at 10dpi.
With these gshB mutants showing an early senescence phenotype (Table 6), we therefore conclude that bacterial GSH is important for efficient nitrogen fixation and maintaining nodule function in time. Developmental nodule senescence has been proposed to result from a decrease in the ascorbate/glutathione antioxidant pool coupled to decreasing carbon-to-nitrogen ratios inside the nodule (Puppo et al., 2005). In this context, analyse of carbon and nitrogen metabolism in nodule obtained with gshB mutant strain could be very interesting to analyse the potential redox regulatory process of carbon and nitrogen metabolism.
GSH endows bacterial stress tolerance
GSH was reported to protect a diversity of bacteria against environmental stresses (Chesney et al., 1996; Crockford et al., 1996; Masip et al., 2006), therefore another evidenced feature of GSH deficient strains is that they are prone to be susceptible to stresses and uncompetitive in nodulation. For example, one R. tropici CIAT899 strain is with an intrinsic tolerance of acidity. One of its mutant strains CIAT899-13T2 is unable to produce GSH and incapable to grow under acidic condition, due to a single Tn5-luxAB insertion in a gene with high similarity of E. coli gshB gene. The mutant presented a sensitivity to low pH levels, high osmolarity, hydrogen peroxide, methylglyoxal toxicity (Riccillo et al., 2000). The acid tolerance can be restored in this mutant by exogenous GSH at 80µM (Riccillo et al., 2000) and by plasmid-based complementation of the gshB gene (Muglia et al., 2007). In terms of the transcription of gshB, it was induced by acidity and UV radiation, but showed no response to environmental constraints, such as osmotic and oxidative stresses, which is known to be mediated by GSH (Muglia et al., 2007).
These results suggest that metabolic production of GSH is important in stress tolerance of bacteria under adverse conditions, with peculiar regulation of potassium channels, and involvement in antioxidation and detoxification. In our study, we have also observed an accidental sudden senescence of SmgshB inoculated nodules when growth chamber light was partially off for two days (over the weekend), while the WT type seems more tolerant under weak lumination. However, more experimentations are needed to reveal the role of GSH in stress tolerance of S. meliloti than just a single occurrence.

144
Table 6 Phenotypes of GSH deficient rhizobia strains
Strains Free-living phenotype
Symbiotic phenotype compared to WT strain
Ref. Nodulation
Nitrogen fixation & plant
biomass Senescence
R. tropici CIAT899-
13T2
Growth restored with 80µM GSH or plasmidic
complementation;
intracellular potassium accumulation;
reduced tolerance to acid and osmotic stresses;
requires exogenous GSH to detoxify methylglycol;
outcompeted in co-
inoculation;
higher number of nodules but
half in size at 2wpi;
reduced dry matter of
plant aerial parts at 2- and
4- wpi;
Similar with WT at 2wpi;
Premature nodule
senescence at 4wpi;
(Riccillo et al., 2000)
(Muglia et al., 2007)
(Muglia et al., 2008)
S. meliloti 1021
gshB
Growth restored with 100µM GSH;
enhanced catalase activity;
delayed nodulation with lower
number of nodules at 1wpi;
reduced nitrogen fixation
capacity at 4wpi;
Premature nodule
senescence;
(Harrison et al.,
2005)
R. etli gshB
impaired glutamine utilization;
susceptible to acidity and electrophilic chemicals;
Growth restored with 100µM GSH or plasmidic
complementation;
delayed nodulation;
lower number of nodules at
3wpi;
heterogeneous nodule size;
reduced nitrogen fixation
capacity at 3wpi; premature senescence; (Tate et al., 2012)
R. leguminosarum
biovar viciae 3841
gshB
Growth restored with 100µM GSH or plasmidic
complementation;
outcompeted in co-
inoculation;
similar nodule number;
heterogeneous nodule size;
reduced plant dry weight
and nitrogen fixation
ability;
(Cheng et al., 2017)
S. meliloti 2011
gshB (SmgshB) Growth restored with 2M GSH;
delayed nodulation;
lower number of nodules at
10dpi but higher at 28dpi ;
heterogeneous nodule size;
reduced nitrogen fixation
capacity at 10-, 20- and
28- dpi;
less accumulated plant
dry weight at 20- and 28-
dpi;
premature senescence;
similar bacterial DNA
content at 10- and 20- dpi,
but diminished at 28dpi;
this study
(Figure S 2)

145
"%'
+)&+)

146

147
CHAPTER II
The Nodule Senescence (NS) related cis-regulatory elements

148

CHAPTER II
149
I. Introduction
I.1 Transcriptional regulatory system in eukaryotes
I.1.1 Transcriptional regulation
Transcriptional regulation is a fundamental biological principle in eukaryotes. It determines divergent cell types and multiple biological functions, nevertheless governed by invariant gene sets. Multiple biological processes of cell life, such as cell proliferation, specification, differentiation, and the final cell death, are initially under the strict control of transcriptional mechanism. Expression of a single gene can be influenced by internal or external signals, as consequences of different spatial localizations in plant tissue, temporal expression under given developmental stages, or responses to environmental stimuli.
Transcriptional machinery controls gene expression
Cellular levels of RNA are under control of the complex mixture of transcriptional regulatory mechanisms (Figure 23) (Wasserman & Sandelin, 2004; Noonan & McCallion, 2010). Multiple events are included in transcription in eukaryotes, such as decondensation of the locus, nucleosome remodelling, histone modifications, recruitment of the basal transcription machinery to the core promoter, and binding of transcriptional activators and coactivators to enhancers and promoters (Smale & Kadonaga, 2003). The basal transcription is determined with the RNA polymerase binding to the sequence-specific core promoter which is present on all of the genes. The dynamic gene expression is under control of transcriptional regulation network, which is composed of the binding of Transcription Factors (TFs) to the Transcription-Factor Binding Sites (TFBS; the cis-regulatory DNA elements) that are either proximal or distal to a Transcription Start Site (TSS), in addition to co-factors and chromatin structure (Wasserman & Sandelin, 2004). Sets of numerous TFs proteins are capable to bind to specifically TFBS thus coordinately activating or repressing gene transcription. Therefore, the transcriptional binding is not only determined by the sequence-specificity of TFBS, but also influenced by the adjacent sequences and the proximity of other proteins. TFs have strict spatial requirements in their binding sites that preclude variable spacing. For some TFs, such as a subset of the nuclear receptor family, variable spacing is allowed (Wasserman & Sandelin, 2004). The bound TFs can interact with cofactors to stabilize the transcription-initiation
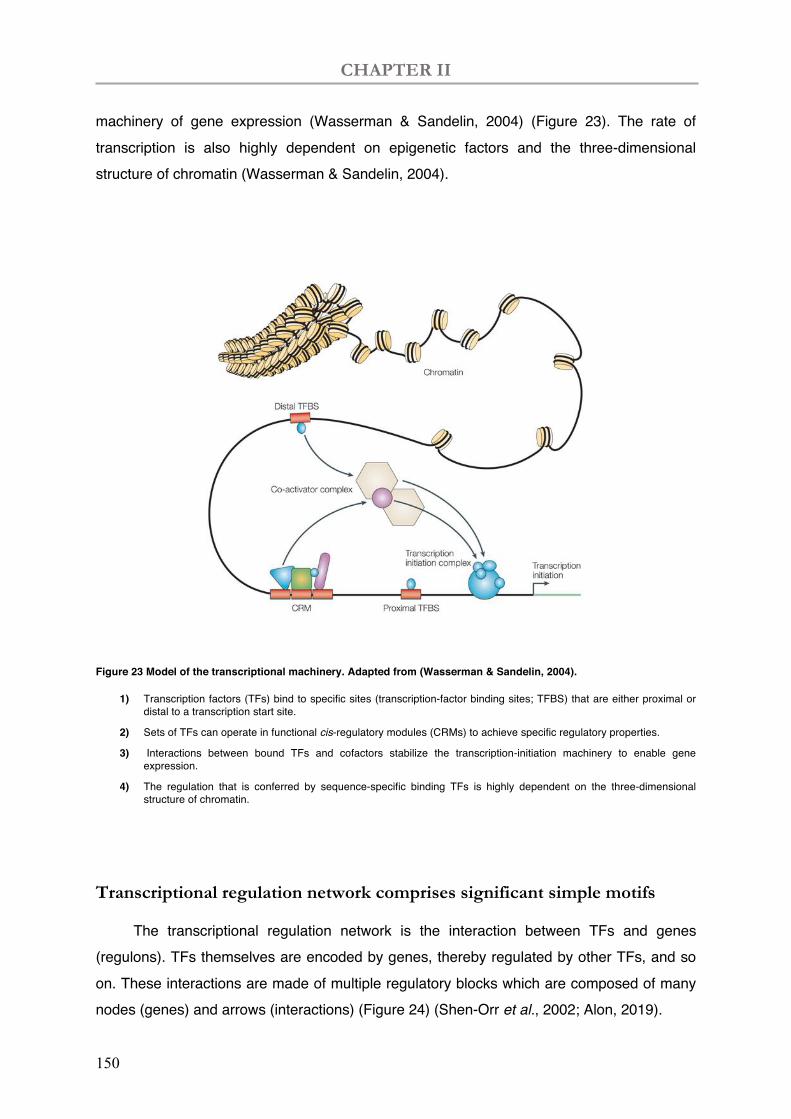
CHAPTER II
150
machinery of gene expression (Wasserman & Sandelin, 2004) (Figure 23). The rate of transcription is also highly dependent on epigenetic factors and the three-dimensional structure of chromatin (Wasserman & Sandelin, 2004).
Figure 23 Model of the transcriptional machinery. Adapted from (Wasserman & Sandelin, 2004).
1) Transcription factors (TFs) bind to specific sites (transcription-factor binding sites; TFBS) that are either proximal or distal to a transcription start site.
2) Sets of TFs can operate in functional cis-regulatory modules (CRMs) to achieve specific regulatory properties.
3) Interactions between bound TFs and cofactors stabilize the transcription-initiation machinery to enable gene expression.
4) The regulation that is conferred by sequence-specific binding TFs is highly dependent on the three-dimensional structure of chromatin.
Transcriptional regulation network comprises significant simple motifs
The transcriptional regulation network is the interaction between TFs and genes (regulons). TFs themselves are encoded by genes, thereby regulated by other TFs, and so on. These interactions are made of multiple regulatory blocks which are composed of many nodes (genes) and arrows (interactions) (Figure 24) (Shen-Orr et al., 2002; Alon, 2019).

CHAPTER II
151
1) Autoregulation is the autogenous control that a gene is regulated by itself (Alon, 2019). 2) The feedforward loop (FFL) is defined by a TF X that regulates a second TF Y, such
that both X and Y jointly regulate a regulon Z (Figure 24) (Shen-Orr et al., 2002). 3) The single-input module (SIM) is defined by a set of target genes that are controlled
by a master TF (Figure 24) (Shen-Orr et al., 2002), also known as Multi-Input Motif (MIM) (Lee et al., 2002). The genes in SIM always have a common biological function (Alon, 2019).
4) The ‘Dense Overlapping Regulons (DOR) is a layer of overlapping interactions between regulons and a group of input transcription that is much more dense than corresponding structures in randomized networks (Figure 24) (Shen-Orr et al., 2002).
Thereof a complex transcription network is formed by these basic regulatory motifs
which are significantly frequent in real transcription. Each network motif has a specific function in governing specific gene expressions (Figure 24)(Milo et al., 2002; Shen-Orr et al., 2002). Altogether, gene expression is the net effect of the dynamical transcription network. Many genes, as nodes in the network, can be regulated by multiple TFs. Their promoter activity is thus a multi-dimensional input function of the different input transcription factors (Alon, 2019). Additionally, TFs in plants can bind to hundreds or thousands of genomic sequences, but with only a small portion of targets responding (Franco-Zorrilla et al., 2014).
Figure 24 The feedforward loop, single-input module (SIM) and dense-overlapping regulons (DOR) found in transcription networks. Adapted from (Shen-Orr et al., 2002).
Each arrow represents for the direct regulation of a TF on its target genes, as activation or repression.

CHAPTER II
152
I.1.2 Cis-regulatory elements
Cis-regulatory elements involved in transcriptional regulation in eukaryotes
Non-coding genomic DNA were long-time regarded as junk DNA as without a protein-coding function. For the last decades, due to boosted knowledge on phylogenetic footprinting between orthologous genes, nucleotide composition and the assessment of available transcript data, there are increasing evidences revealing the roles of non-coding sequences in transcriptional regulation and a prominent contribution in evolution (Wasserman & Sandelin, 2004). The abundant cis-regulatory sequences distribute discretely in cluster in genome. The cis-regulatory elements act across divergent distances in genome to control the spatial and temporal expression of genes. The biological significance of cis-regulatory elements is increasingly uncovered mainly on sequence-conservation-based analyses and application of transgenic approaches in model organisms.
Cis-regulatory elements play different roles in transcriptional control
The cis-elements are recognized with different roles in transcriptional regulation (Figure 25) (Noonan & McCallion, 2010). Transcriptional promoters are in the centre of the basal transcription machinery and located at the 5’ ends of genes. Promoters defines the orientation and origin of transcription (Maston et al., 2006). Mostly, functional regulatory sequences are more likely to be near the TSS according to the current laboratory annotation (Wasserman & Sandelin, 2004). But many genes have multiple TSSs. Thus, promoters, that contain one or more TSS(s) are more and more investigated for identification of regulatory sequences. Promoter regions can be identified with transcript data via recurrent alignment of the 5′ ESTs and/or full-length cDNAs (Wasserman & Sandelin, 2004), which is generally accessible in genome browsers.
The cis-regulatory elements have also been revealed with important regulatory functions for gene expression positively (as enhancers) or negatively (as repressors/silencers). The regulatory output will be the net effects of positive and negative inputs, often as consequences of environmental stimuli and developmental cues (Noonan & McCallion, 2010). Some enhancers can also behave like repressors dependent on the differential transcription factor binding (Noonan & McCallion, 2010). Insulators possess two kinds of activities as enhancer-blocking or barrier activities (Figure 25, d & e) (Noonan & McCallion, 2010). Enhancer-blocking activity refers to the inhibition of activating effects when placed between enhancers, whereas

CHAPTER II
153
the barrier activities is the ability to set boundary between active and repressive chromatin (Noonan & McCallion, 2010).
Figure 25 Classes of known regulatory elements. Adapted from (Noonan & McCallion, 2010).
(a) Promoter sequence shown binding the general transcriptional machinery to mediate basal transcriptional control of a transcribed sequence. (b) Enhancer and (c) repressor sequences are shown to mediate positive and negative effects of transcription through interaction with the promoter sequence. (d) Enhancer blocking and (e) barrier insulator elements block promiscuous activation of gene B by regulatory elements of gene A (enhancer blocking, d) or prevent the spread of chromatin condensation from encroaching on the regulatory sequences controlling a hypothetical gene (barrier, e).
Regulatory DNA sequences are vital in evolutionary processes
Evolutionary pressures lead to the retention of conserved genetic modules such as the regulatory sequences (Rubin et al., 2000). Genetic mutations on one hand boost genetic diversity, whereas on the other hand mutations with certain functions will be more retained allowing population to adapt and species to evolve. Mutations in functional sequences, which mostly negative effects for fitness, will be lost from the population (Noonan & McCallion, 2010). On the contrary, non-functional mutations are free to be fixed by drift as sequences of high

CHAPTER II
154
genetic variations (Kimura, 1968). For example, the movement of transposable elements are the major originators to promote mutations and chromosome rearrangements, which is thereafter possible to become heritable after the phenotypic modification and adaptation (Biémont & Vieira, 2006). Waves of invasion or loss of transposable elements shape the genome and control the activity of genes during the evolution of populations and species (Biémont & Vieira, 2006). The assumption that regulatory mechanisms are conserved in orthologous genes of different species, enables the prediction of conserved regulatory genomic sequences. An establishment of datasets of regulatory sequences is necessary including the protein binding specificity, epigenetic marks, as well as the validated data of regulatory sequences and tissue specificity of characterized promoters.
Increasing evidence enrich the databases of cis-regulatory elements
During the last decades, advances in the identification of the TF binding-sequences have been helpful to decipher the transcriptional regulatory codes (Harbison et al., 2004; Badis et al., 2009). The general cis-elements in eukaryotes boxes, such TATA, CCAAT and GC (Mitchell, P. J. & Tjian, R., 1989). In eukaryotes, one of the best described core promoters are the TATA-contained core promoters in RNA polymerase II transcribed genes. TATA box can associate with the basal transcription factors with direct binding with the TATA-binding protein (TBP) (Lifton et al., 1978; Smale & Kadonaga, 2003; Molina & Grotewold, 2005). The consensus site of TATA box is TATAWAW, while a wide variety of AT rich sequences can be recognized by TBP (Smale & Kadonaga, 2003). It is often located 25-30 bp upstream of a TSS (Smale & Kadonaga, 2003). The orientation of the TATA box would have minimal contribution to the direction of transcription (Smale & Kadonaga, 2003). TATA box is essential in TATA-contained core promoters, but it is not prevalent in the majority of gene promoters. For example, in Arabidopsis, only about 29% of the promoters are TATA-contained, harbouring the TATA box around -32bp to the TSS, while a large proportion of promoters were identified as TATA-less (Molina & Grotewold, 2005). The RNA polymerase II, together with other basal TFs, gives only low level of transcription. Other factors, such as regulatory enhancers, are necessary to increase the transcription activity. For example, CAAT box, sometimes also called CCAAT box or CAT box, is the binding sites for the general TFs, i.e., nuclear factor Y (NF-Y; or core binding factors, CBF), with a consensus sequence that is CAAT (Shirsat et al., 1989) or CCAAT (Laloum et al., 2013).
In plant, (Franco-Zorrilla et al., 2014) et al. have identified DNA-binding specificities of 63 TFs, which are mostly consistent with previous data in corresponding TF families. More

CHAPTER II
155
than 80% of these binding sites have a biological relevance based on the analysis of coregulated genes, transcriptomic data, and chromatin hypersensitive regions (Franco-Zorrilla et al., 2014). Approximately half of the tested TFs recognized a secondary DNA element differing from its primary motifs. The sequence enrichment is higher in upstream than in downstream regions in terms of enhancers, whereas the spatial restriction is less strict when it comes to the repressors. The sequence frequency is found to be higher in upstream (1kb) in case of positively co-regulated genes, while analogous in negatively co-regulated genes (Franco-Zorrilla et al., 2014). These data enriched the datasets for mapping cis-regulatory elements in genome and contributed to decipher transcriptional regulatory, which in return allows the prediction of functions of particular TFs or DNA fragments code.
Several Cis-regulatory elements have been identified in leguminous plants
Functional studies on transcriptional regulators has uncovered numerous cis-regulatory elements in leguminous plants, particularly during the course of symbiosis. In bacterial the infection process, the promoter of MtENOD11 located -411bp upstream of the translation start codon is proven to be sufficient to induce the expression in response to Nod factor and bacterial infection (Boisson-Dernier et al., 2005). Then three TFs ERN1, -2, and -3 were identified to directly regulate the gene expression of MtENOD11 via specific binding to a Nod factor responsive cis-element, the NF-box (between -411bp to -358bp to ATG) (Andriankaja et al., 2007). During rhizobial infection and organogenesis, certain cis-regulatory sequences within ~7kb upstream of NIN are required to regulate bacterial infection but are insufficient for the activation of cortical cell divisions (Yoro et al., 2014). More recently, the 5 kb upstream region of NIN is identified to be sufficient for root hair curling and bacterial infection, but not for nodule primordium formation (Liu et al., 2019). Interestingly, a remote cis-regulatory region located 18kb upstream of the start codon is identified for MtNIN expression initiate nodule primordium formation, containing putative cytokinin response elements (Liu et al., 2019). However, no cis-regulatory element has been found to-date related to nodule senescence regulation.
I.2 Cysteine proteases (CPs) in nodule senescence
There are approximately 140 cysteine proteases (CPs, also cysteine peptidases, cysteine proteinases) among the about 800 proteases encoded by plant genomes (Barrett et al., 2012). CPs belong to 15 families in five clans according to the MEROPS classification (http://www.ebi.ac.uk/merops/) (Rawlings et al., 2018). The papain-like proteases C1A (family

CHAPTER II
156
C1, clan CA) have been described as active proteolytic enzymes induced in plants during multiple physiological processes such as germination, senescence processes and plant defence (Sheokand & Brewin, 2003; Gilroy et al., 2007; Barrett et al., 2012). Within leguminous plant nodules, CPs are engaged in response to the microbial invasion; protein turnover for tissue remodelling; metabolic homeostasis and nutrient reutilization; adaptation to environmental stresses; leaf and nodule senescence (Sheokand & Brewin, 2003). In indeterminate nodules, senescence begins from the proximal to the apical part of the nodule. CP gene expression have been widely associated to nodule senescence of M. truncatula (Fedorova et al., 2002; Van de Velde et al., 2006), soybean (Alesandrini et al., 2003a,b), Chinese milk vetch (Naito et al., 2000), Pisum sativum (Kardailsky & Brewin, 1996) and Astragalus sinicus (Li et al., 2008).
The upregulation of cysteine protease genes features the onset of nodule
senescence
Enhanced activity of proteolytic enzymes is a hallmark of senescence (Pladys & Vance, 1993; Lopez-Otin et al., 2013). Nodule senescence is featured with lytic-related processes such as rupture of the peribacteroid membranes, lysis of bacteroids, and disorganization of host-cell cytosol (Pladys & Vance, 1993; Van de Velde et al., 2006). Thereof, CPs have been evidenced to activate nodule senescence by multiple groups (Pladys & Vance, 1993; Kardailsky & Brewin, 1996; Naito et al., 2000; Vincent & Brewin, 2000; Alesandrini et al., 2003; Asp et al., 2004; Van de Velde et al., 2006; Li et al., 2008; Perez Guerra et al., 2010; Pierre et al., 2014; van Wyk et al., 2014; Cilliers et al., 2018; Deng et al., 2019). In 2010, a cluster of six M. truncatula CP genes (MtCP1 to MtCP6) were shown to be in relation with senescence process (Perez Guerra et al., 2010). In the same group, other nodule senescence related CPs were found such as TrCP1,5,8,10 (Asp et al., 2004) and AsNODf32 (Naito et al., 2000). The members of this cluster contains an N-terminal signal peptide and a putative tetrapeptide vacuolar targeting signal (LQRD; Chrispeels, 1991), while an HDEL or KDEL endoplasmic reticulum retention signal was absent (Perez Guerra et al., 2010). The protein structure indicates a secretion pathway destined for vacuole-like structures, while later on MtCP6 protease was found to accumulate in the vacuole of uninfected cells or in the symbiosomes of infected cells (Pierre et al., 2014). Considering the discrepancies of gene annotation in genomic and transcriptomic databases, a list of MtCPs involved in nodule senescence are shown in Table 7.
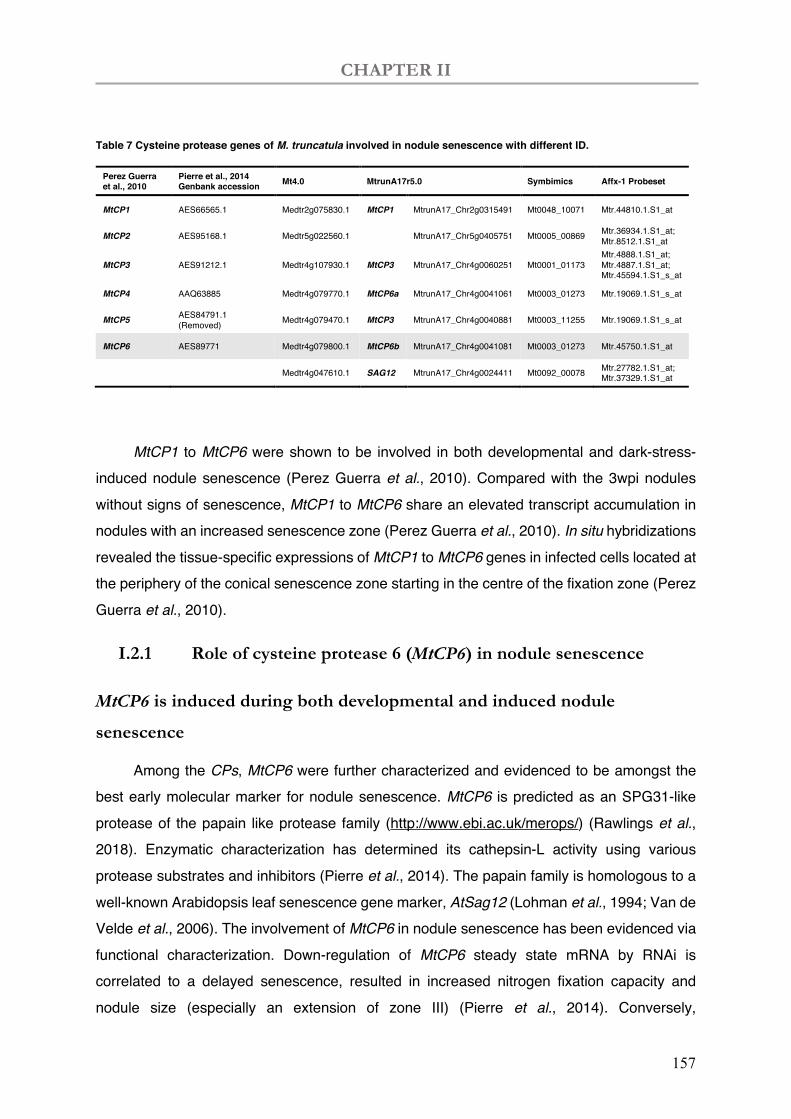
CHAPTER II
157
Table 7 Cysteine protease genes of M. truncatula involved in nodule senescence with different ID.
Perez Guerra et al., 2010
Pierre et al., 2014 Genbank accession Mt4.0 MtrunA17r5.0 Symbimics Affx-1 Probeset
MtCP1 AES66565.1 Medtr2g075830.1 MtCP1 MtrunA17_Chr2g0315491 Mt0048_10071 Mtr.44810.1.S1_at
MtCP2 AES95168.1 Medtr5g022560.1 MtrunA17_Chr5g0405751 Mt0005_00869 Mtr.36934.1.S1_at; Mtr.8512.1.S1_at
MtCP3 AES91212.1 Medtr4g107930.1 MtCP3 MtrunA17_Chr4g0060251 Mt0001_01173 Mtr.4888.1.S1_at; Mtr.4887.1.S1_at; Mtr.45594.1.S1_s_at
MtCP4 AAQ63885 Medtr4g079770.1 MtCP6a MtrunA17_Chr4g0041061 Mt0003_01273 Mtr.19069.1.S1_s_at
MtCP5 AES84791.1 (Removed) Medtr4g079470.1 MtCP3 MtrunA17_Chr4g0040881 Mt0003_11255 Mtr.19069.1.S1_s_at
MtCP6 AES89771 Medtr4g079800.1 MtCP6b MtrunA17_Chr4g0041081 Mt0003_01273 Mtr.45750.1.S1_at
Medtr4g047610.1 SAG12 MtrunA17_Chr4g0024411 Mt0092_00078 Mtr.27782.1.S1_at; Mtr.37329.1.S1_at
MtCP1 to MtCP6 were shown to be involved in both developmental and dark-stress-
induced nodule senescence (Perez Guerra et al., 2010). Compared with the 3wpi nodules without signs of senescence, MtCP1 to MtCP6 share an elevated transcript accumulation in nodules with an increased senescence zone (Perez Guerra et al., 2010). In situ hybridizations revealed the tissue-specific expressions of MtCP1 to MtCP6 genes in infected cells located at the periphery of the conical senescence zone starting in the centre of the fixation zone (Perez Guerra et al., 2010).
I.2.1 Role of cysteine protease 6 (MtCP6) in nodule senescence
MtCP6 is induced during both developmental and induced nodule
senescence
Among the CPs, MtCP6 were further characterized and evidenced to be amongst the best early molecular marker for nodule senescence. MtCP6 is predicted as an SPG31-like protease of the papain like protease family (http://www.ebi.ac.uk/merops/) (Rawlings et al., 2018). Enzymatic characterization has determined its cathepsin-L activity using various protease substrates and inhibitors (Pierre et al., 2014). The papain family is homologous to a well-known Arabidopsis leaf senescence gene marker, AtSag12 (Lohman et al., 1994; Van de Velde et al., 2006). The involvement of MtCP6 in nodule senescence has been evidenced via functional characterization. Down-regulation of MtCP6 steady state mRNA by RNAi is correlated to a delayed senescence, resulted in increased nitrogen fixation capacity and nodule size (especially an extension of zone III) (Pierre et al., 2014). Conversely,

CHAPTER II
158
overexpression of MtCP6 leads to a significant increase in N fixation capacity and in size of the zone III in nodule (Pierre et al., 2014).
MtCP6 has been used as a molecular marker for the onset of nodule senescence in numerous studies. Expression of MtCP6 is induced during both developmental senescence and induced nodule senescence, such as dark stress and nitrate treatment (Perez Guerra et al., 2010; Pierre et al., 2014). Using the β-glucuronidase (GUS) reporter gene, the expression of MtCP6 was localized in the basal part of 5wpi nodules, but absent in young nodules (Figure 26) (Perez Guerra et al., 2010). Later on, GUS-promoter analysis of MtCP6 shows a specific expression at the end of N fixation zone (zone III) to the beginning of senescence zone (zone IV), and sharply extended to the whole nodules under abiotic stresses like dark and nitrate treatment (Figure 26) (Pierre et al., 2014). In addition, an accelerated nodule senescence are observed with inefficient symbiosis, with enhanced the development of nodule zone IV and the extension of spatial MtCP6 expression (Cam et al., 2012; Pierre et al., 2014; Yang, L. et al., 2020). The findings of a spatiotemporal expression of MtCP6 at transcriptional level that correlated with proteolytic activities involved in nodule senescence indicate a specific transcriptional regulation of nodule senescence related signals.
To date, functional promoter studies have led to the identification of several nodule-specific cis-elements most in early steps of nodulation, e.g. CYC-box (TGCCATGTGGCA) (Singh et al., 2014), NSP1 (cis-element AATTT) (Hirsch et al., 2009), NFbox (Andriankaja et al., 2007), NIN (AAGMT; a cytokinin-responsive cis-regulatory region) (Marsh et al., 2007 ; Vernié et al., 2015; Liu et al., 2019). However, no regulatory sequences have been identified directly related to nodule senescence. To decipher the early steps of transcriptional regulation of nodule senescence, we looked for in this chapter cis-regulatory elements regulating the transcription of the early senescence marker MtCP6 in transgenic root nodules generated by Agrobacterium rhizogenes.
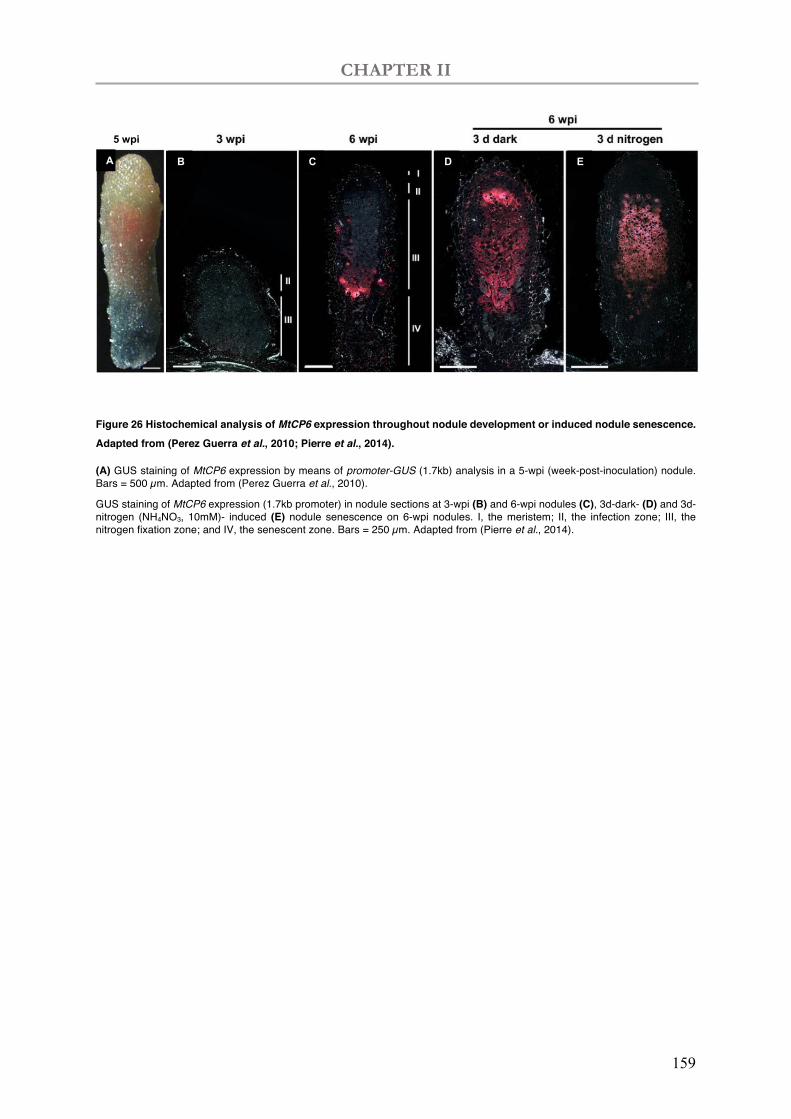
CHAPTER II
159
Figure 26 Histochemical analysis of MtCP6 expression throughout nodule development or induced nodule senescence. Adapted from (Perez Guerra et al., 2010; Pierre et al., 2014).
(A) GUS staining of MtCP6 expression by means of promoter-GUS (1.7kb) analysis in a 5-wpi (week-post-inoculation) nodule. Bars = 500 µm. Adapted from (Perez Guerra et al., 2010).
GUS staining of MtCP6 expression (1.7kb promoter) in nodule sections at 3-wpi (B) and 6-wpi nodules (C), 3d-dark- (D) and 3d-nitrogen (NH4NO3, 10mM)- induced (E) nodule senescence on 6-wpi nodules. I, the meristem; II, the infection zone; III, the nitrogen fixation zone; and IV, the senescent zone. Bars = 250 µm. Adapted from (Pierre et al., 2014).
A B C D E 5 wpi

CHAPTER II
160

CHAPTER II
161
II. Results (manuscript)
Yang L, Frances L, de Carvalho-Niebel F, Boncompagni E, Frendo P. 2020. Identification of cis-acting elements involved in the transcriptional regulation of the cysteine protease gene MtCP6 in nodule senescence of Medicago truncatula. In preparation.

CHAPTER II
162

1
Identification of cis-regulatory elements involved in the 1
transcriptional regulation of the cysteine protease gene 2
MtCP6 during nodule senescence of Medicago truncatula 3
Li Yang1, Lisa Frances2, Fernanda De Carvalho-Niebel2, Pierre Frendo1, Éric 4
Boncompagni1 5 1 Institut Sophia Agrobiotech, UMR INRAE 1355, Université Côte d’Azur, CNRS 7254, 400 6
Route de Chappes, BP167, 06903 Sophia Antipolis, France. 7 2 LIPM, Université de Toulouse, INRA, CNRS, 31326 Castanet-Tolosan, France 8
Short title: Transcriptional Regulation of MtCP6 9
One-sentence summary: 10
Identification of cis-acting regulatory elements of MtCP6 during nodule senescence 11
12

2
ABSTRACT 13
Leguminous plants associate with rhizobia to establish nitrogen-fixing symbiosis, leading to 14
the formation of new root organs called nodules. However, lifespan of nodules is limited due 15
to the senescence process, which ends with a complete degradation of bacteroids and host plant 16
cells. The increased proteolytic activity is one of the hallmarks of nodule senescence. In the 17
nodule senescence initiation process of Medicago truncatula, a papain cysteine protease, 18
MtCP6, is found to be involved. The corresponding gene MtCP6 is specifically induced in the 19
nodule transition zone from nitrogen fixation to senescence, both under developmental and 20
induced nodule senescence. In order to decipher the senescence-signalling pathway in 21
symbiotic nodules, we focus on the analysis of the cis-regulatory elements present on the 22
proximal MtCP6 gene promoter. First, progressive deletions of this promoter have permitted 23
to identify, in vivo, a minimal promoter sequence sufficient for a specific spatiotemporal 24
transcriptional activation in nodule. Furthermore, functional analysis has validated the cis 25
elements containing promoter fragment by gain- or loss-of-function. This work will lead to a 26
better comprehension of the transcriptional regulatory network involved in the nodule 27
senescence process. 28
29
Keywords: Medicago truncatula; nodule senescence; cysteine proteases; cis-regulatory 30
elements; transcriptional regulation 31
32

3
Introduction 33
Leguminous plants play an important role in sustainable agriculture due to the establishment 34
of a symbiotic association with soil nitrogen-fixing bacteria of the Rhizobiacea family. This 35
symbiosis leads to the development of de novo root organ, the root nodule, in which the bacteria 36
differentiated in bacteroids are able to reduce atmospheric (Ferguson et al., 2010; Syska et al., 37
2019). Nodules are developmentally classified into determinate and indeterminate nodules 38
according to the persistence of nodule meristem. In determinate nodules, meristem is 39
nonpersistent, and a radial gradient of senescent cells occurs from the centre and extend to the 40
periphery along with nodule aging (Puppo et al., 2005; Kazmierczak et al., 2020). Conversely, 41
in indeterminate nodules, the presence of a continually active apical meristem (zone I) ensures 42
the continuous growth of the nodule. Cells from the meristematic zone are continuously 43
infected by Sinorhizobium meliloti in the infection zone (zone II). Therein, bacteria reside in 44
plant cells and are surrounded by the plant-originated peribacteroid membrane, collectively 45
known as symbiosome. Thereafter, host plant and bacterial cells initiate a terminal 46
differentiation process in the interzone II–III to form the nitrogen-fixing symbiotic cells (zone 47
III). In zone III, the bacteroids express nitrogenase and reduce atmospheric nitrogen into 48
ammonia. However, the lifespan of root nodule is time-limited and extremely sensitive to 49
abiotic stresses, leading to the development of the senescence zone (zone IV) (Kazmierczak et 50
al., 2020; Puppo et al., 2005). The nodule senescence is characterized with a decline of nitrogen 51
fixation capacity and a coordinated death of both bacteria and plant cells (Puppo et al., 2005; 52
Van de Velde et al., 2006; Perez Guerra et al., 2010; Kazmierczak et al., 2020). 53
The symbiosis between legumes and rhizobia are mainly based on nutrient exchange of 54
photosynthates and symbiotically fixed nitrogen. Senescence mostly occurs when the nutrient 55
trading is out of equilibrium and the associated organ, the root nodule, will turn to general 56
nutrient sources instead of carbon sink, for nutrient reutilization in other developing organs of 57
the host plants (Puppo et al., 2005; Van de Velde et al., 2006; Perez Guerra et al., 2010; 58
Kazmierczak et al., 2020). Thus, nodule senescence occurs not only during the organ aging 59
(developmental senescence), but it is also triggered by various environmental changes (induced 60
senescence), such as drought (Aranjuelo et al., 2013; Gil-Quintana et al., 2015), dark stress 61
(Perez Guerra et al., 2010; Pierre et al., 2014), and nitrate treatment (Matamoros et al., 1999; 62
Pierre et al., 2014). Accelerated nodule senescence was also observed by interruption of plant 63
genes required for effective symbiosis, e.g. MtDNF2 (Bourcy et al., 2013), MtNAC969 (de 64
Zelicourt et al., 2012), MtNPDs (Trujillo et al., 2019), or in symbiotic interaction with an 65

4
inefficient bacteria partner, e.g. S. meliloti gshB (Yang et al., 2020a) and hmp (Cam et al., 66
2012). 67
A key feature of nodule senescence in legume is the triggering of proteolytic activities, 68
involved in cellular degradation and nutrient remobilisation (Alesandrini et al., 2003; Puppo et 69
al., 2005; Groten et al., 2006; Kazmierczak et al., 2020; Yang et al., 2020b). Although many 70
factors can affect protease activities, transcriptomic analysis has emphasized the induction of 71
the transcription of a group of cysteine protease encoding genes at the onset of nodule 72
senescence in M. truncatula - S. meliloti symbiosis (Pladys & Vance, 1993; Kardailsky & 73
Brewin, 1996; Fedorova et al., 2002; Alesandrini et al., 2003; Van de Velde et al., 2006; Perez 74
Guerra et al., 2010). In situ hybridizations revealed the tissue-specific expressions of MtCP1 75
to MtCP6 genes in infected cells located at the periphery of the conical senescence zone starting 76
in the center of the fixation zone (Perez Guerra et al., 2010). A further functional 77
characterization of MtCP6 gene expression has shown its significant role as an early gene 78
marker of developmental and induced nodule senescence (Pierre et al., 2014). Down-regulation 79
of MtCP6 steady state mRNA by RNAi is correlated to a delayed senescence, and conversely, 80
overexpression of MtCP6 leads to a significant increase in nitrogen fixation capacity and in 81
size of the symbiotic zone in nodule (Pierre et al., 2014). Using the β-glucuronidase (GUS) 82
reporter gene, MtCP6 was found to be specifically expressed at the transition between the 83
nitrogen fixing zone (zone III) and the senescence zone (Zone IV) during developmental nodule 84
senescence (Perez Guerra et al., 2010; Pierre et al., 2014). By contrast, under induced nodule 85
senescence conditions of dark stress or nitrate treatment, the spatial expression of MtCP6 86
changes drastically and extends to the nodule distal part (Pierre et al., 2014). These results 87
suggest a specific spatiotemporal transcriptional regulation of MtCP6 during developmental 88
and induced nodule senescence. 89
To date, functional promoter studies have led to the identification of several nodule-specific 90
cis-regulatory elements, mostly in early steps of nodule organogenesis (Vijn et al., 1995; 91
Andriankaja et al., 2007; Liu et al., 2019). However, no cis-regulatory sequences have been 92
identified directly related to nodule senescence. Nevertheless, a few transcription factors (TFs) 93
have been identified via differential expression analysis of transcriptomic data, mainly from 94
MYeloBlastosis (MYB), basic Helix-Loop-Helix (bHLH), basic region leucine ZIPper (bZIP), 95
and NAM/ATAF/CUC (NAC) families (Van de Velde et al., 2006; Seabra et al., 2012; Cabeza 96
et al., 2014; Chungopast et al., 2014; Serova et al., 2017; Yuan et al., 2017). For instance, a 97
TF MtATB2 (bZIP family) is regulated by sucrose and is enhanced during nodule senescence 98
(D'Haeseleer et al., 2010). Recently, the TF MtbHLH2 is reported to be a transcriptional 99

5
repressor of a cysteine protease encoding gene (MtCP77) and down-regulated accordingly in 100
nodule senescence (Deng et al., 2019). 101
In this study, in order to decipher transcriptional regulation at the early steps of nodule 102
senescence, we looked for cis-regulatory elements involved in the transcriptional regulation of 103
MtCP6 in transgenic root nodule under developmental and induced nodule senescence. Serial 104
promoter deletion analysis of MtCP6 allowed us to determine the minimal MtCP6 promoter 105
(ProCP6) down to −242bp upstream of the transcription starting site (TSS), which was 106
sufficient to drive a robust tissue-specific expression related to root nodule senescence. 107
Furthermore, a 67 bases pair sequence of the promoter region, termed as Nodule Senescence 108
(NS) element or NS-box, was defined to drive the tissue-specific expression related to both 109
developmental and nitrate induced nodule senescence using both gain and loss of function 110
experiments. Mutagenesis of putative CAAT/WRKY cis-regulatory elements in the NS-box of 111
MtCP6 promoter impairs the NS sequence regulatory activity. Taken together, these results 112
showed that we have identified the first cis-regulatory elements involved in the early regulation 113
of root nodule senescence. 114
115

6
Materials and Methods 116
Plant material, root transformation and growth condition 117
M. truncatula Jemalong A17 seeds were scarified, surface-sterilized and germinated as 118
described by Boisson-Dernier et al. (Boisson-Dernier et al., 2001). Germinated seedlings were 119
stabbed at hypocotyl with Agrobacterium rhizogenes suspension using a modified protocol 120
from Kereszt et al. (Kereszt et al., 2007). Plantlets were thereafter transferred into a mixture 121
substrate of perlite and sand (3:1) with a basic nitrogen supply of 1mM NH4NO3. Plants were 122
grown at 20 for one week for optimal transformation and then grown at 23 for 2 weeks. 123
Then transgenic plant roots were selected by cutting off the non-green-fluorescent roots (GFP) 124
under a Leica MZ FLIII fluorescence stereomicroscope (Leica, Wetzlar, Germany). After one-125
week-adaptation from the selective cutting, transgenic composite plants were inoculated with 126
10mL of Sinorhizobium meliloti 2011 pXLGD4 (Grefen et al., 2010) at optical density of 127
OD600nm=0.1. Nodules were harvested at 4 week-post-inoculation (wpi). Nitrogen-treated 128
nodules were from plants applied with 10ml KNO3 (10mM) for 2 days before 4wpi. 129
Escherichia coli DH5α was cultivated in LB Broth at 37 with appropriate antibiotics. S. 130
meliloti 2011 pXLGD4 was growing in LBMC (LB Broth, added with 2.5mM CaCl2 and 2.5 131
mM MgSO4) with 100μg/mL streptomycin and 10μg/mL tetracycline at 30. Agrobacterium 132
rhizogenes Arqua1 was grown in TY medium (5g/L bacto-tryptone, 3g/L yeast extract, 6mM 133
CaCl2) with 100μg/ml streptomycin at 28(Quandt, 1993). 134
135
Plasmid constructions 136
Fifteen promoter fragments (−1,720bp, −1,467bp, −1,278bp, −1,088bp, −599bp, −511bp, 137
−356bp, −303bp, −273bp, −242bp, −211bp, −175bp, −141bp, and −80bp) were cloned into 138
entry vector pDONR™ P4-P1r (Invitrogen). Empty entry vector was built by recombination of 139
attB4::empty cassette::attB1 into pDONR™ P4-P1r. Expression vector was constructed by 140
multi-site Gateway recombination of promoter entry vectors with pENTR-GUS and 141
pENTL2L3-T35S into the destination vector pKm43GWD-RolD:eGFP (Cam et al., 2012). 142
Primer sequences are listed in Table S2. 143
For gain of function experiments, seamless tetramers of NS region (−242bp to −175bp), NS1 144
(−242bp to −182bp) and NS2 (−210bp to −175bp) were synthesized into pUC53-Kan (Genewiz 145
corporation, Takeley, United Kingdom). Tetramers were then inserted into EcoRI/HindIII sites 146
upstream of a minimal CaMV 35S promoter (47bp) in a binary vector pLP100 (Szabados et al., 147

7
1995). Binary pLP100 constructs were then introduced into A. rhizogenes Arqua1 by electro-148
transformation (MicroPulser, Bio-Rad). 149
150
Internal and Block Deletion Constructs 151
Block deletion of the NS region was generated with complementary primers (Table S1) from 152
pDONR™ P4-P1r-ProCP6 (−1,720bp). Similarly, two other promoter deletion constructs 153
(ΔCW, ΔDof) were generated from pDONR™ P4-P1r:ProCP6 (−242bp). Then the deleted 154
promoters (ΔNS, ΔCW and ΔDof) was recombined into pKm43GWD-RolD:eGFP as 155
described in promoter deletion construction. 156
157
Histological analyses and microscope observations 158
Nodules with attached root fragments were harvested at 4wpi, and at 4wpi with a 2d-nitrate 159
treatment. Samples were stained with X-Gluc (5-bromo-4-chloro-3-indolyl-β-D glucuronide, 160
cyclohexylammonium salt; Euromedex) with a modified protocol from Jefferson et al. 161
(Jefferson, 1987). Nodules were incubated at 37°C overnight. Alternatively, short-time staining 162
(4h) were applied to better differentiate staining intensity of promoters less than -303bp. Whole 163
stained nodule images were taken with a Leica MZFLIII stereomicroscope (Leica 164
Microsystems). The nodules were thereafter embedded in 6% (w/v) agarose and sectioned 165
(70µm) with HM650V vibratome (Thermo Scientific). Nodule section images were taken 166
under dark field using Axioplan 2 microscope (Carl Zeiss). From all the histological 167
experiments, more than 100 nodules were analysed from at least 15 independent plants. All 168
experiments were conducted with three biological replicates. 169
170
Fluorimetric GUS assays 171
To quantify enzymatic GUS activity, 20g of nodules were grounded in liquid nitrogen. Total 172
proteins were extracted with GUS buffer (50 mM sodium phosphate, pH 7.5, 10 mM 2-173
mercaptoethanol, 10 mM Na2EDTA, 0.1% Triton X-100, and 0.1% sodium lauryl-sarcosine). 174
Quantitative GUS activities were measured as previously described (Boisson-Dernier et al., 175
2005). Total protein (10µg) extraction was measured with 4-methylumbelliferyl-β-D-176
glucuronide (MUG) as substrate (Biosynth AG). GUS activities were measured using a 177
microtiter fluorimeter (FL600, Bio-Tek). Standard curves were prepared with a range of 178
increasing concentrations of 4-methylumbelliferone (4-MU) (Sigma-Aldrich). 179
180
In silico analysis 181

8
In silico analysis of MtCP6 promoter was performed in multiple research-based databases 182
(PLACE, PlantPAN and Plantcare) to identify conserved cis-element motifs (Higo et al., 1999; 183
Lescot et al., 2002; Chow et al., 2016). To visualize cis-regulatory element homolog patterns, 184
analysis was carried out on promoters of Cysteine Protease 1 (MtCP1, 1,701bp), MtCP2 185
(1,700bp), MtCP3 (513bp), MtCP4 (1,700bp), MtCP5 (1,701bp), MtCP6 (1,732bp) which are 186
co-expression genes in nodule senescence with bioinformatic tools (de Pater et al., 1996; Bailey 187
et al., 2009). Analysis was conducted with MEME algorithm (p-value>0.05) on MEME Suite 188
web server (http://meme-suite.org) with the following parameters: maximum number of motifs 189
(3), minimum motif width (6), maximum motif width (50), minimum sites per motif (2), 190
maximum sites per motif (600). 191
192

9
RESULTS 193
The truncated −242bp MtCP6 promoter is sufficient to confer specific spatiotemporal 194
GUS expression under developmental and nitrate-induced nodule senescence 195
The MtCP6 promoter-GUS fusion is specifically expressed in the transition between the nodule 196
nitrogen-fixing zone III and the nodule senescence zone IV (Fig. 1A) (Perez Guerra et al., 2010; 197
Pierre et al., 2014). Furthermore, the expression is dramatically upregulated under abiotic 198
stresses like nitrogen treatment and dark stress (Perez Guerra et al., 2010; Pierre et al., 2014). 199
In order to identify cis-regulatory elements in the promoter region responsive to developmental 200
and induced nodule senescence, fifteen 5’ progressive MtCP6 promoter deletions were 201
constructed and fused to a GUS reporter gene (Fig. 1B). The promoter fragments were upstream 202
of TSS, ranging from −1,720bp to −80bp. The position of TSS is determined at the start of 5’ 203
UTR (35bp) via the genomic data (ID: Medtr4g079800.1; website: 204
https://legumeinfo.org/genomes/jbrowse/?data=Mt4.0 ) (Tang et al., 2014). The multiple 205
constructions were introduced in M. truncatula via A. rhizogenes transformation. A promoter 206
less construct (Empty vector) was generated for negative control. Transgenic roots and nodules 207
were harvested at 4wpi from composite transgenic M. truncatula plants and stained for 208
detecting GUS activities. 209
To analyse GUS activity spatiotemporal localization, stained nodules were classified according 210
to the control nodules with a complete MtCP6 promoter expression (Fig. 1B; Fig. S1). Class A 211
corresponded to nodules showing GUS staining tissue at the interzone III-IV. Class B 212
corresponded to those presenting abnormal spatiotemporal expressions. While, Class C 213
corresponded to nodules with no detectable GUS staining. Detectable GUS activity for the full 214
promoter construct (−1,720bp) was present in 87% of the harvested nodules, with 79% of Class 215
A. Detectable Gus activity was present in at least 50% of the harvested nodules, with 40% of 216
Class A for the promoter constructs ranging from -1,467bp to -242 bp. Additional 5′ deletion 217
down to −211bp reduced drastically the percentage of stained nodule to 10%, albeit 9% of 218
nodules were in class A (Fig. 1B). Non-specific GUS staining (Class B) mostly appears in 219
shorter promoters under −211bp, at −175bp, 46% of nodules hold a detectable GUS staining 220
but 43% were from Class B nodules. Interestingly, the −242bp promoter allowed 79% of 221
detectable GUS activity with 97% of Class A whereas the −211bp promoter region allowed 10% 222
of detectable activity with 90% of Class A (Fig. 1B). GUS activity was not detectable in any 223
nodule with the negative control. 224

10
To unravel the cis-elements of MtCP6 promoter responding to nitrate, treatment was applied 2 225
days before harvesting at 4wpi and GUS activity was compared in treated and untreated 226
nodulated roots (Fig. 1C). To evaluate GUS expression intensities, nodule staining was 227
classified into strong (+++), moderate (++), weak (+) and undetectable (−) staining. In total, 228
for all promoter deletion, more than 200 nodules were analyzed from at least 15 independent 229
plants of at least three biological replicates. 230
Transgenic nodules of composite plants harboring the different MtCP6 promoter deletion 231
constructs were stained for GUS activity overnight and sectioned before microscopic 232
observation, which showed a strong blue precipitation (+++). To obtain a better gradient 233
detection (+; +/−; −), staining period was reduced to 4h in the proximal promoter -303bp 234
upstream of the TSS (Transcriptional Starting Site) (Fig. 1C). Sliced nodules were used to 235
define the different zones of the nodules. Representative images of GUS activity in sliced 236
nodules are presented for each promoter deletion construct (Fig. 1C). Under nitrogen treatment, 237
the GUS staining for the full MtCP6 promoter construct was enhanced in transgenic nodules 238
and the expression zone expanded from proximal to distal part of nodules spreading into the 239
nitrogen fixing zone (zone III). Similarly, to the developmental senescence, a threshold of GUS 240
activity detection was observed between the construction under the control of the promoter 241
constructs at −242bp and −211bp. Whereas the GUS activity was correctly localize in nodule, 242
the promoter deletion at −211bp led to a strong reduction (+/−) of GUS activity under nitrogen 243
treatment (Fig. 1B, 1C). 244
Taken together, these results suggest that a promoter region of 254bp (between −242bp and 245
+12bp) is sufficient to maintain the tissue-specific transcription of MtCP6 during both 246
developmental and nitrate-induced senescence. Specific cis-regulatory elements allowing gene 247
expression during senescence may be present in the promoter region between −242bp and 248
−175bp and bear specific cis-regulatory elements related to this specific regulation. Thus, the 249
MtCP6 promoter region between −242bp and −175bp was termed as the nodule senescence 250
(NS) related box or NS-box. 251
252
The NS-box is sufficient to endow nodule senescence expression patterns to GUS reporter 253
gene 254
Our data suggest that the NS-box corresponding to a 67bp sequence is involved in the 255
regulation of the specific spatiotemporal expression related to nodule senescence pattern. To 256
validate this hypothesis, gain of function analysis was performed with tetramers of complete 257
NS-box (NS; 67bp), of 5’-part of NS-box sequence (NS1, 50bp) and of 3’-part of NS- box 258

11
sequence (NS2, 45bp) added to a minimal CaMV 35S promoter (Pmin35S) and fused to GUS 259
reporter gene in pLP100 binary vector (Fig. 2A). The expression pattern of the resulting 260
chimeric genes was analysed in transgenic root nodules at 4wpi with or without nitrogen 261
application. Representative images of transgenic nodules expressing chimeric gene constructs 262
are presented in Fig. 2B. GUS activity was observed in the basal part of the nodule (Class A) 263
when GUS was under the control of NS tetramer (Fig. 2B, 2C). GUS staining was not detected 264
when the gene was under the control of NS1 and NS2 tetramers. Nitrate treatment induced a 265
stronger GUS staining in nodule with NS tetramer which was localized in zone III and in the 266
beginning of zone IV (Fig. 2C). As during developmental senescence, GUS staining was not 267
detected when the gene was under the control of NS1 and NS2 tetramers. The GUS activity of 268
4xNS construct was detected in 12% of the analysed samples with an appropriate localisation 269
of the expression in 79% of the positive samples (Fig. 2B). Under nitrate treatment, 20% of 270
nodules were stained and all the GUS positive nodules presented an appropriate GUS staining 271
localization (Fig. 2B). Detectable GUS activity related to 4xNS1 construct at 4wpi appeared 272
only in one biological experiment with only 8% of nodules (77% in Class A) and the nitrate 273
treatment did not increase the GUS activity in nodules. No GUS activity was observed for 274
4xNS2 construct. The control (Pmin35S) with an empty pLP100 vector does not showed any 275
stained nodules in the same treatment conditions. 276
Quantification of GUS activity using fluorometric assay was performed to compare the 277
transcriptional activity of the different constructs (Fig. 2D). 4xNS construct allowed a 278
significant higher GUS activity (124±13 pmol MU/min/mg protein) than 4xNS1 construct 279
(41±2 pmol MU/min/mg protein), 4xNS2 construct (17±2 pmol MU/min/mg protein). After 280
nitrate treatment, GUS activity of 4xNS construct increased 6-fold to 786±110 pmol 281
MU/min/mg protein, while the increase was less than 1.8-fold for 4xNS1 construct and 4xNS2 282
construct with 73±2 and 31±2 pmol MU/min/mg protein, respectively. The GUS activities for 283
the Pmin35S construct were 18±2 and 20±2 pmol MU/min/mg protein at 4wpi PI or 4wpi PI 284
with nitrate treatment, respectively (Fig. 2D). 285
Taken together, these data show that the tetramer of the NS-box fused to minimal 35S promoter 286
is able to provide the nodule senescence tissue-transcription specificity to GUS supporting the 287
hypothesis that the NS-box contains cis-regulatory elements involved in the gene regulation 288
during nodule senescence process. 289
290
Loss of the NS-box impairs the promoter MtCP6 transcriptional activity. 291

12
The transcriptional regulation of MtCP6 during nodule senescence may involve multiple cis-292
regulatory sequences. To determine the significance of NS-box regulatory element in the full 293
promoter (−1,710bp), loss of function analysis was conducted with a ΔNS deletion construct 294
(Fig. 3A). Qualitative analysis of GUS activity suggests that transgenic nodules carrying the 295
ΔNS construct expressed GUS similarly to the full promoter (Fig. 3B). The spatial expression 296
localization of GUS was also similar for the construct ΔNS and the complete promoter (Fig. 297
3C). However, the GUS activity was performed overnight, and the signal seemed to be 298
saturated in these staining conditions. The quantitative analysis of nodule GUS staining 299
performed by fluorometric assay showed that GUS activity in ProCP6 ΔNS nodule (1,879±267 300
pmol MU/min/mg) was strongly reduced by more than 80% compared to the activity in full 301
promoter control nodule (11,544±965 pmol MU/min/mg) (Fig. 3D). 302
Taken together, the results showed the significant role of NS-box in MtCP6 promoter activity 303
and suggest the presence of other cis-regulatory sequence(s) allowing the expression of MtCP6 304
during nodule senescence. 305
306
A CW motif is crucial for the NS box activity 307
The bioinformatic analysis of the full promoter gives abundant putative cis-regulatory elements, 308
which hinders the possible laboratory validation. Hence, we decided to focus on 309
characterization the NS-box which allows the GUS expression in the nodule senescence zone. 310
To identify the putative cis-regulatory elements present in the minimal sufficient promoter 311
from −242bp to -175bp, we analyze the sequence with Plant cis-regulatory element databases 312
(Higo et al., 1999; Lescot et al., 2002; Chow et al., 2016). Into the NS-box, putative regulatory 313
cis-elements have been found and might be involved in the specific regulation of MtCP6 314
expression in nodule (Fig. 4). The core promoter element TATA-box (CTATAAATAC) 315
(Shirsat et al., 1989; Grace et al., 2004) is located at −32bp upstream of TSS in MtCP6 316
promoter. Two CAAT-box are found in the NS-box (Shirsat et al., 1989) which are widely 317
recognized as common sequences found in the 5’-non-coding regions of eukaryotic genes were 318
identified. The core sequence of W-box (TGAC) has been found twice within NS-box (Eulgem 319
et al., 1999; Zhang et al., 2004; Xie et al., 2005). The core site required for binding of Dof 320
(DNA binding with one finger) proteins (AAAG) was also observed (Yanagisawa & Schmidt, 321
1999; Yanagisawa, 2000; Yanagisawa, 2002). The consensus motif (AATTT) of a GRAS 322
domain transcriptional regulator, Nodulation Signalling Pathway1 (NSP1) (Hirsch et al., 2009), 323
is also present in inside NS-box. 324
325

13
Considering that NS1 was found to be the more important part of the NS-box involved in 326
regulation of MtCP6 spatiotemporal gene expression, selected motifs between −242bp and 327
−211bp, but not found in NS2 (CAAT/WRKY motifs and a tandem of Dof motifs) were 328
validated by loss of function experiments. Thus, we deleted CAAT-WRKY sequence (ΔCW) 329
and tandem Dof sequence (ΔDof) in the -242pb promoter which allows GUS expression in the 330
nodule senescence zone (Fig. 5A). Constructs were fused to GUS reporter gene, introduced 331
into transgenic roots and qualitative analysis of GUS staining was performed (Fig. 5B). 332
Histological staining showed a visible but weaker GUS staining in ΔCW nodules in comparison 333
to −242bp MtCP6 promoter. In contrast, no significant difference in GUS activity was observed 334
between the ΔCW nodules and control nodules. 46% and 74% of nodules were found to be 335
from Class A for ΔCW and in ΔDof constructs, respectively (Fig. 5C), while 64% of nodules 336
were stained blue in the −242bp MtCP6 promoter construct. Using fluorometric assay, the 337
lower GUS expression level in nodules was confirmed for ΔCW construct (292±49 pmol 338
MU/min/mg protein) compared to the −242bp MtCP6 promoter construct (as 3,436±378) with 339
the −242bp MtCP6 promoter construct (Fig. 5D). 340
In conclusion, the CAAT-WRKY motif present in the NS-box, is involved in the transcriptional 341
activity of the MtCP6 promoter (−242bp) during nodule senescence. 342
343

14
DISCUSSION 344
345
Promoter activity of MtCP6 is related to the tissue-specific gene expression at the onset of 346
nodule senescence. 347
Expression of MtCP6 has been used as a specific marker for the breakdown of nodule 348
symbiosis under developmental and induced senescence (Perez Guerra et al., 2010; Cam et al., 349
2012; de Zelicourt et al., 2012; Pierre et al., 2014; Yang et al., 2020a). Multiple transcriptomic 350
analyses have shown the co-expression of MtCP6 with five other CP genes during nodule 351
senescence (Fig. S2) (Benedito et al., 2008; Perez Guerra et al., 2010). Eight probesets 352
corresponding to CPs (Fig. S2) were highlighted with induction in nodules and with nitrate 353
treatment (Fig. S2) (https://mtgea.noble.org/v3/) (Benedito et al., 2008). Additionally, 354
expression of CPs was triggered in nodules treated with phosphinothricin (PPT), which 355
inhibited the Glutamine synthetase and induced during premature nodule senescence (Fig. S2) 356
(Seabra et al., 2012). These data indicate that CPs are associated to nodule senescence, and 357
they are highly sensitive to plant nitrogen status. 358
To define cis-regulatory elements involved in the specific spatiotemporal expression at the 359
onset of nodule senescence, we conducted a serial deletion analysis initiated from a 1,732bp 360
MtCP6 promoter characterized in previous studies (Perez Guerra et al., 2010; Pierre et al., 361
2014). The spatiotemporal specificity and nitrate response of MtCP6 promoter are preserved 362
in promoter deletions until −242bp to TSS, which is identified as the minimal and sufficient 363
promoter of MtCP6. However, the intensity of the GUS activity decreased gradually 364
concomitantly to the gradual shortening of MtCP6 promoter. Thus, other cis-regulatory motifs, 365
probably functioning as enhancers, might be present upstream the nucleotide -242 in the CP6 366
promoter. The significant GUS staining obtained with the deletion of the NS-box in the 1,732bp 367
CP6 promoter supports this idea. We could thus assume that other cis-regulatory elements may 368
be involved in the transcriptional regulation, in addition to the NS-box that controls the tissue-369
specific transcription in nodule senescence. 370
371
NS-box mediates a transcriptional activation at the onset of developmental nodule 372
senescence and nodule response to nitrate 373
We have determined a specific DNA sequence, the NS-box, that drives a similar transcriptional 374
activity to the −1,720bp CP6 promoter. Interestingly, all MtCPs presenting a correlated 375
expression pattern with MtCP6 (Fig. S2) are sharing large similarities between NS-box element 376

15
(Fig. S3A&B). To date, no transcriptional regulatory factors (TFs) has been demonstrated to 377
switch on the transcription during nodule senescence. In contrast, few TFs has been evidenced 378
as repressors for CP gene expressions (de Zelicourt et al., 2012; Deng et al., 2019). 379
Transcriptomic analysis of the MtbHLH2 mutant enabled the identification of a target gene, 380
MtCP77, and the protein-DNA binding was proved by electrophoretic mobility shift assays and 381
chromatin immunoprecipitation analysis (Deng et al., 2019). Gene expression of MtCP77 is 382
positively related to nodule senescence with a decreased nitrogenase activity, accelerated 383
programmed cell death and reactive oxygen species accumulation (Deng et al., 2019). However, 384
analysis of the MtCP6 promoter sequence down to −1,720bp did not allow the identification of 385
the TF binding sequence known as G-box (5’-CACGTG-3’) (Menkens et al., 1995). 386
Nevertheless, it is not found in, however, the less canonical CANNTG motif (Kawagoe et al., 387
1994), also known as E-box (Hartmann et al., 2005), can be found multiple times (−1236, −204 388
and −156 bp) in the MtCP6 promoter. However, no direct evidence has been found concerning 389
the regulation of MtCP6 by the bHLH repressor as the promoter deletion analysis did not show 390
a detectable shift of promoter activity between −175 and −146 bp which harbour one of the E-391
box. Finally, the general expression profile of MtCP77 is different from the MtCP1 to MtCP6 392
suggesting a rather different regulation of expression. The identification of multiple 393
transcriptional repressors strengthens this idea that multiple regulatory elements are involved 394
in the regulation of CP expression in nodule senescence. 395
The synthetic promoter of NS-box-based tetramers restores partially the transcriptional activity 396
of the -242 bp promoter (Figure 2D). This result showed that positive transcriptional regulators 397
may be involved in MtCP6 transcriptional regulation in nodule senescence. The partial 398
restoration of the transcriptional regulation is quite common as for NF-box that was proven to 399
be sufficient to mediate the nod factor-elicited gene activation, for which the corresponding 400
synthetic promoter constituted of NF-box tetramers recovered less than half of the GUS activity 401
in fluorometric assays (Boisson-Dernier et al., 2005; Andriankaja et al., 2007). Moreover, 402
repetitive number of the same element within a synthetic promoter could have a silencing effect 403
on transcriptional activation (Müller & Sheen, 2008; Zürcher et al., 2016). 404
The gain of function experiments using the synthetic NS-box promoter showed that the NS-405
box activity is sensitive to the nodule nitrate treatment. This suggests that the NS-box plays a 406
regulatory role during developmental and nitrate induced senescence. Henceforth a potential 407
shared transcriptional regulation between developmental and nitrate induced nodule 408
senescence is plausible. However, transcriptional regulators for nitrate response are generally 409
specific. In Arabidopsis, a Nitrate-Responsive cis-regulatory Element (NRE) of 43bp, 410

16
containing conserved sequence 5’-tGACcCTTN10AAGagtcc-3’, was identified and conserved 411
in nitrite reductase (NIR) gene promoters (Konishi & Yanagisawa, 2010; Konishi & 412
Yanagisawa, 2011). NRE is binding to NIN-Like Proteins (NLPs) (Konishi & Yanagisawa, 413
2013) and activate NRE-dependent transcription in plant response to nitrate (Konishi & 414
Yanagisawa, 2013; Lin et al., 2018; Nishida et al., 2018). In M. truncatula, in the NAC family 415
TF, MtNAC969 was reported to be up-regulated with nitrate treatment, albeit negatively 416
regulate developmental nodule senescence (de Zelicourt et al., 2012). 417
418
Putative CAAT and WRKY cis-regulatory elements orchestrate the specific 419
transcriptional role of NS-box 420
Gene transcription is the net effect of subtle equilibrium conferred by multiple regulators, so 421
that multiple cis-regulatory elements are generally required to determine promoter activity. In 422
this work, site specific deletion have been carried out to validate the potential TF binding sites, 423
onto the CAAT-box and the core sequence of consensus WRKY binding site together (ΔCW), 424
as well as a tandem Dof binding site (ΔDof) (Fig. 4; Fig. 5) Whereas, no significant effect of 425
Dof deletion was observed in the promoter activity, a decrease of GUS activity with the ΔCW 426
promoter construct strongly suggests an activating transcriptional role for CAAT/WRKY 427
sequence (Fig. 5C). In plants, CAAT-box are binding the transcription factor family (NF-Y) 428
(Laloum et al., 2013). NF-Y regulatory element is composed of three subunits: NF-YA, NF-429
YB and NF-YC, where NF-YA is the subunit that directly interact with the CAAT-box. 430
Interestingly, MtNF-YA1 gene is specifically expressed in nodule meristematic zone and in 431
distal infection, and gene silencing leads to strong alterations in nodule meristem function and 432
poor nodule development (Combier et al., 2006). MtNF-YA1 belongs to a subgroup of group I 433
where gene are mostly expressed in root and nodule tissues and others MtNF-YA are indeed 434
expected to play key roles in nodule development processes (Laloum et al., 2013). W-box is 435
the consensus sequence for binding sites of WRKY TFs (Eulgem et al., 1999; Zhang et al., 436
2004; Xie et al., 2005). WRKY TFs have been identified as predominantly involved in 437
transcriptional modulators during abiotic stresses. In M. truncatula, WRKY genes 438
Medtr8g005750 and Medtr3g093830 are responding to nod factor and S. meliloti. Interestingly, 439
in tobacco plants, an overexpression of chickpea CaWRKY50 lead to an early plant senescence 440
(Kumar et al., 2016) and in Arabidopsis thaliana, WRKY genes (AtWRKY53, 54, and 80) are 441
found to be either positive or negative regulators of plant senescence (Ülker et al., 2007; 442
Potschin et al., 2014; Kumar et al., 2016). The WRKY binding-site has also been detected in 443
the Nitrate Responsive Element (NRE) in the Arabidopsis NIR1 promoter (Konishi & 444

17
Yanagisawa, 2010). 445
Overall, this study identified a specific DNA sequence (NS-box) positively associated to 446
nodule senescence from the MtCP6 promoter. The fact that NS-box responds to both 447
developmental and induced nodule senescence strongly suggest that nodule senescence and 448
nodule response to environmental changes share, at least partially, similar transcriptional 449
regulatory processes. In the future, transactivation activities between NS-box and some 450
transcription proteins could be identified with yeast-one-hybrid approach. Alternatively, a 451
targeted approach using NF-Y and WRKY to analyse NS-box- TFs interaction could be also 452
considered. 453

18
FUNDING 454
LY is supported by a doctoral fellowship from the China Scholarship Council (CSC). This 455
work was supported by the “Institut National de la Recherche Agronomique”, the “Université 456
Nice Côte d’azur”, and the French Government (National Research Agency, ANR) through the 457
“STAYPINK” project reference # ANR-15-CE20-0005) and the “Investments for the Future” 458
LABEX SIGNALIFE: program reference # ANR-11- LABX-0028-01. 459
460
ACKNOWLEDGEMENTS 461
We are largely indebted to the symbiosis team members for numerous nodule harvests. 462
463
FIGURE LEGENDS 464
Figure 1: Schematic graph of MtCP6 5’ promoter deletion constructs, and corresponding 465
GUS expression spatial specificity analysis. 466
A. Schematic depiction of progressive 5’-deletion of MtCP6 promoter constructs (−1,720, 467
−1,467, −1,278, −1,088, −955, −655, −599, −511, −356, −303, −273, −242,−201,−175, −141, 468
−96, −80 and empty vector). Deleted promoter fragments are fused to β-glucuronidase (GUS) 469
gene. Transcription starting site (TSS) is designated with arrow. The position of promoter is 470
indicated in bp relation to TSS. B. nodules were harvested at 4wpi (week-post-inoculation) 471
from A. rhizogenes mediated transgenic M. truncatula roots inoculated with S. meliloti 2011. 472
Overall spatial GUS expression pattern is shown in percentages and classed in specific 473
expression in root nodule zone III-IV (blue, Class A), unspecific expression in other nodule 474
zones (teal blue, Class B), and non-detectable (GUS-, grey, Class C). Total number of nodules 475
analysed is indicated on the right (n=110 to 460). GUS stained nodules are from more than 476
three independent experiments. C. Histochemical localization of GUS activity of MtCP6 477
truncated promoters in nodules at 4wpi, 2d after nitrate treatment (4wpi). The images are from 478
nodule vibrosections (70µm). Nodules sections were stained overnight with X-Gluc (−1,710, 479
−1,450, −1,268, −1,078, −955, −655, −511, −599, −511, −356), or during 4h (−303, −273, 480
−242,−201,−175, −141, −96, −80 and empty vector). Promoter positions are indicated in bp 481
relative to TSS. Overall, GUS staining intensity is denoted as: strong (+++), detectable (+; +/−) 482
or non-detectable (−). Scale bar = 100µm. 483
484
Figure 2: Gain of function analysis of ProCP6 cis elements. 485
A. Schematic design of gain of function analysis. NS-box (Nodule Senescence) was designed 486
to decipher promoter regulatory region from −242 to −175bp. NS1 (50bp) and NS2 (45bp) are 487

19
5’ or 3’ part of NS-box. Tetramers of NS (4x NS), NS1 (4x NS1) and NS2 (4x NS2) are 488
seamlessly synthesized and separately constructed in front of a minimal CaMV 35S promoter 489
(Pmin35S, 47bp). B. Histochemical localization of β-glucuronidase (GUS) activity of synthetic 490
promoters in A. rhizogenes mediated M. truncatula transgenic root nodules, with the empty 491
vector transformed nodules as negative control. Nodules were harvest at 4wpi and 10mM of 492
KNO3 were applied on nodulated roots for 2d. At least 340 nodules were sampled from more 493
than three independent biological replicates. C. The images show entire nodule at 4wpi after 494
GUS staining for 4x NS, 4x NS1, 4x NS2 and control Pmin35S. GUS reporter in blue show the 495
expression observed in the transition from nitrogen-fixing zone III to senescent zone IV. D. 496
Fluorometric analyses of GUS activity driven by synthetic promoters in nodules at 4wpi with 497
(blue) or without nitrate treatment (yellow). Datapoints stand for independent test from three 498
biological replicates. 499
500
Figure 3: Loss of function of cis-regulatory elements on MtCP6 promoter. 501
A. Design of ΔNS deletion onto CP6 promoter. B. relative distribution of transgenic root 502
nodules with spatiotemporal distribution of zone III/IV, non-specific or undetectable (GUS-). 503
C. Representative nodule sample of GUS histochemical localization of expression from 504
ProCP6 (−1,710bp), ΔNS, and control Pmin35S. D. Fluorometric assay of GUS activity driven 505
by synthetic promoters in nodules at 4wpi from ProCP6 (−1,710bp), ΔNS, and control 506
Pmin35S constructs. Results are obtained from nine independent experimental data points from 507
three biological replicates. 508
509
510
Figure 4: In silico analysis of putative cis-regulatory elements present onto NS-box 511
(−242~−175bp). 512
Previously identified cis-regulatory elements area indicated as: TATA-box with double 513
underline (CTATAAATAC) (Shirsat et al., 1989; Grace et al., 2004), ARR1-binding-site 514
(NGATT) in dashed line, NSP1 consensus motif (consensus motif AATTT) underlined (Hirsh 515
et al., 2009); or annotated with name: CAAT-box (Shirsat et al., 1989), CCAAT-box (Rieping 516
& Schöffl, 1992; Haralampidis et al., 2002; Wenkel et al., 2006; Laloum et al., 2013), WRKY 517
(W-box) (Eulgem et al., 1999; Zhang et al., 2004; Xie et al., 2005); core binding site of Dof 518
proteins (AAAG) (Yanagisawa & Schmidt, 1999; Yanagisawa, 2000). 519
520
Figure 5: Loss of function of cis-elements on minimal MtCP6 promoter (−242bp). 521

20
A. Design of ΔCW and ΔDof deletions onto MtCP6 (−242bp) promoter. B. relative distribution 522
of transgenic root nodules with spatiotemporal distribution of zone III/IV (Class A), non-523
specific (Class B) or undetectable (Class C). C. Representative nodule sample of the 524
histochemical GUS localization of expression from MtCP6 (−242bp), ΔCW, ΔDof and control 525
Pmin35S. D. Fluorometric assay of GUS activity driven by synthetic promoters, MtCP6 526
(−242bp), ΔCW, ΔDof and control Pmin35S, in nodules at 4wpi. Results are obtained from 527
nine independent experimental data points from three biological replicates. 528
529
SUPPLEMENTARY DATA: 530
531
Table S1: List of primers used in this study. 532
533
Figure S1: Overall spatial GUS expression pattern is shown in percentages and classed in 534
specific expression in root nodule zone III-IV (Class A), unspecific expression in other nodule 535
zones (Class B), and non-detectable (Class C). Total number of nodules analysed is indicated 536
on the right (n=110 to 460). GUS stained nodules are from more than three independent 537
experiments. 538
539
Figure S2: Expression profile of MtCP1 to MtCP6 on MtGEA: M. truncatula Gene Expression 540
Atlas / Noble Foundation (https://mtgea.noble.org/v3). 541
542
Figure S3: NS box enrichment in co-expressed CPs (500bp promoter fragments). 543
A. Research of motif similarities over MtCP1 to MtCP6 using as Bioinformatic tool MEME 544
Suite web server (http://meme-suite.org). 545
IUPAC nucleotide code (A: Adenine, C: Cytosine, G: Guanine, T: Thymine, R: A /G, Y: C/T, 546
S: G/C, W: A /T, K: G/T, M: A/C, B: C/G/T, D: A/G/T, H: A/C/T, V: A/C/G, N: any base) 547
B. Conservation of Motif 2. Parameters of motif 3: E-value (5.6e-028); Site Count (7); Width 548
(37). 549
550

21
REFERENCES 551
Alesandrini F, Frendo P, Puppo A, Herouart D. 2003. Isolation of a molecular marker of soybean 552 nodule senescence. Plant Physiology and Biochemistry 41(8): 727-732. 553
Andriankaja A, Boisson-Dernier A, Frances L, Sauviac L, Jauneau A, Barker DG, de Carvalho-Niebel F. 554 2007. AP2-ERF transcription factors mediate Nod factor dependent Mt ENOD11 activation in 555 root hairs via a novel cis-regulatory motif. Plant Cell 19(9): 2866-2885. 556
Aranjuelo I, Tcherkez G, Molero G, Gilard F, Avice JC, Nogues S. 2013. Concerted changes in N and C 557 primary metabolism in alfalfa (Medicago sativa) under water restriction. Journal Of 558 Experimental Botany 64(4): 885-897. 559
Bailey TL, Boden M, Buske FA, Frith M, Grant CE, Clementi L, Ren J, Li WW, Noble WS. 2009. MEME 560 SUITE: tools for motif discovery and searching. Nucleic Acids Research 37(Web Server issue): 561 W202-208. 562
Benedito VA, Torres-Jerez I, Murray JD, Andriankaja A, Allen S, Kakar K, Wandrey M, Verdier J, Zuber 563 H, Ott T, et al. 2008. A gene expression atlas of the model legume Medicago truncatula. Plant 564 Journal 55(3): 504-513. 565
Boisson-Dernier A, Andriankaja A, Chabaud M, Niebel A, Journet EP, Barker DG, de Carvalho-Niebel 566 F. 2005. MtENOD11 gene activation during rhizobial infection and mycorrhizal arbuscule 567 development requires a common AT-rich-containing regulatory sequence. Molecular Plant-568 microbe Interactions 18(12): 1269-1276. 569
Boisson-Dernier A, Chabaud M, Garcia F, Becard G, Rosenberg C, Barker DG. 2001. Agrobacterium 570 rhizogenes-transformed roots of Medicago truncatula for the study of nitrogen-fixing and 571 endomycorrhizal symbiotic associations. Molecular Plant-microbe Interactions 14(6): 695-700. 572
Bourcy M, Brocard L, Pislariu CI, Cosson V, Mergaert P, Tadege M, Mysore KS, Udvardi MK, Gourion 573 B, Ratet P. 2013. Medicago truncatula DNF2 is a PI-PLC-XD-containing protein required for 574 bacteroid persistence and prevention of nodule early senescence and defense-like reactions. 575 New Phytologist 197(4): 1250-1261. 576
Cabeza R, Koester B, Liese R, Lingner A, Baumgarten V, Dirks J, Salinas-Riester G, Pommerenke C, 577 Dittert K, Schulze J. 2014. An RNA sequencing transcriptome analysis reveals novel insights 578 into molecular aspects of the nitrate impact on the nodule activity of Medicago truncatula. 579 Plant Physiology 164(1): 400-411. 580
Cam Y, Pierre O, Boncompagni E, Herouart D, Meilhoc E, Bruand C. 2012. Nitric oxide (NO): a key 581 player in the senescence of Medicago truncatula root nodules. New Phytologist 196(2): 548-582 560. 583
Chow CN, Zheng HQ, Wu NY, Chien CH, Huang HD, Lee TY, Chiang-Hsieh YF, Hou PF, Yang TY, Chang 584 WC. 2016. PlantPAN 2.0: an update of plant promoter analysis navigator for reconstructing 585 transcriptional regulatory networks in plants. Nucleic Acids Research 44(D1): D1154-1160. 586
Chungopast S, Hirakawa H, Sato S, Handa Y, Saito K, Kawaguchi M, Tajima S, Nomura M. 2014. 587 Transcriptomic profiles of nodule senescence in Lotus japonicus and Mesorhizobium loti 588 symbiosis. Plant Biotechnology 31(4): 345-U115. 589
Combier JP, Frugier F, de Billy F, Boualem A, El-Yahyaoui F, Moreau S, Vernie T, Ott T, Gamas P, 590 Crespi M, et al. 2006. MtHAP2-1 is a key transcriptional regulator of symbiotic nodule 591 development regulated by microRNA169 in Medicago truncatula. Genes & Development 592 20(22): 3084-3088. 593
D'Haeseleer K, De Keyser A, Goormachtig S, Holsters M. 2010. Transcription factor MtATB2: about 594 nodulation, sucrose and senescence. Plant and Cell Physiology 51(9): 1416-1424. 595
de Pater S, Greco V, Pham K, Memelink J, Kijne J. 1996. Characterization of a zinc-dependent 596 transcriptional activator from Arabidopsis. Nucleic Acids Research 24(23): 4624-4631. 597
de Zelicourt A, Diet A, Marion J, Laffont C, Ariel F, Moison M, Zahaf O, Crespi M, Gruber V, Frugier F. 598 2012. Dual involvement of a Medicago truncatula NAC transcription factor in root abiotic 599 stress response and symbiotic nodule senescence. Plant Journal 70(2): 220-230. 600

22
Deng J, Zhu F, Liu J, Zhao Y, Wen J, Wang T, Dong J. 2019. Transcription Factor bHLH2 Represses 601 CYSTEINE PROTEASE77 to Negatively Regulate Nodule Senescence. Plant Physiology 181(4): 602 1683-1703. 603
Diaz I, Martinez M, Isabel-LaMoneda I, Rubio-Somoza I, Carbonero P. 2005. The DOF protein, SAD, 604 interacts with GAMYB in plant nuclei and activates transcription of endosperm-specific genes 605 during barley seed development. The Plant Journal 42(5): 652-662. 606
Eulgem T, Rushton PJ, Schmelzer E, Hahlbrock K, Somssich IE. 1999. Early nuclear events in plant 607 defence signalling: rapid gene activation by WRKY transcription factors. The EMBO journal 608 18(17): 4689-4699. 609
Fedorova M, van de Mortel J, Matsumoto PA, Cho J, Town CD, VandenBosch KA, Gantt JS, Vance CP. 610 2002. Genome-wide identification of nodule-specific transcripts in the model legume 611 Medicago truncatula. Plant Physiology 130(2): 519-537. 612
Ferguson BJ, Indrasumunar A, Hayashi S, Lin MH, Lin YH, Reid DE, Gresshoff PM. 2010. Molecular 613 analysis of legume nodule development and autoregulation. Journal of integrative plant 614 biology 52(1): 61-76. 615
Gil-Quintana E, Lyon D, Staudinger C, Wienkoop S, Gonzalez EM. 2015. Medicago truncatula and 616 Glycine max: Different Drought Tolerance and Similar Local Response of the Root Nodule 617 Proteome. Journal of Proteome Research 14(12): 5240-5251. 618
Gomez SK, Javot H, Deewatthanawong P, Torres-Jerez I, Tang Y, Blancaflor EB, Udvardi MK, Harrison 619 MJ. 2009. Medicago truncatula and Glomus intraradices gene expression in cortical cells 620 harboring arbuscules in the arbuscular mycorrhizal symbiosis. BMC Plant Biology 9(1): 10. 621
Gourion B, Berrabah F, Ratet P, Stacey G. 2015. Rhizobium-legume symbioses: the crucial role of plant 622 immunity. Trends In Plant Science 20(3): 186-194. 623
Grace ML, Chandrasekharan MB, Hall TC, Crowe AJ. 2004. Sequence and spacing of TATA box 624 elements are critical for accurate initiation from the β-phaseolin promoter. Journal Of 625 Biological Chemistry 279(9): 8102-8110. 626
Grefen C, Donald N, Hashimoto K, Kudla J, Schumacher K, Blatt MR. 2010. A ubiquitin-10 promoter-627 based vector set for fluorescent protein tagging facilitates temporal stability and native 628 protein distribution in transient and stable expression studies. The Plant Journal 64(2): 355-629 365. 630
Groten K, Dutilleul C, van Heerden PD, Vanacker H, Bernard S, Finkemeier I, Dietz KJ, Foyer CH. 2006. 631 Redox regulation of peroxiredoxin and proteinases by ascorbate and thiols during pea root 632 nodule senescence. FEBS Letters 580(5): 1269-1276. 633
Haralampidis K, Milioni D, Rigas S, Hatzopoulos P. 2002. Combinatorial interaction of cis elements 634 specifies the expression of the Arabidopsis AtHsp90-1gene. Plant Physiology 129(3): 1138-635 1149. 636
Harrison M, Dixon R. 1993. Isoflavonoid accumulation and expression of defense gene transcripts 637 during the establishment of vesicular-arbuscular mycorrhizal associations in roots of 638 Medicago truncatula. Molecular Plant-microbe Interactions 6(5): 643-654. 639
Hartmann U, Sagasser M, Mehrtens F, Stracke R, Weisshaar B. 2005. Differential combinatorial 640 interactions of cis-acting elements recognized by R2R3-MYB, BZIP, and BHLH factors control 641 light-responsive and tissue-specific activation of phenylpropanoid biosynthesis genes. Plant 642 Molecular Biology 57(2): 155-171. 643
Higo K, Ugawa Y, Iwamoto M, Korenaga T. 1999. Plant cis-acting regulatory DNA elements (PLACE) 644 database: 1999. Nucleic Acids Research 27(1): 297-300. 645
Hirsch S, Kim J, Muñoz A, Heckmann AB, Downie JA, Oldroyd GED. 2009. GRAS proteins form a DNA 646 binding complex to induce gene expression during nodulation signaling in Medicago 647 truncatula. The Plant Cell 21(2): 545-557. 648
Hogekamp C, Arndt D, Pereira PA, Becker JD, Hohnjec N, Küster H. 2011. Laser microdissection 649 unravels cell-type-specific transcription in arbuscular mycorrhizal roots, including CAAT-box 650

23
transcription factor gene expression correlating with fungal contact and spread. Plant 651 Physiology 157(4): 2023-2043. 652
Jefferson RA. 1987. Assaying chimeric genes in plants: the GUS gene fusion system. Plant Molecular 653 Biology Reporter 5(4): 387-405. 654
Jin Z, Chandrasekaran U, Liu A. 2014. Genome-wide analysis of the Dof transcription factors in castor 655 bean (Ricinus communis L.). Genes & genomics 36(4): 527-537. 656
Kardailsky IV, Brewin NJ. 1996. Expression of cysteine protease genes in pea nodule development and 657 senescence. Molecular Plant-microbe Interactions 9(8): 689-695. 658
Karmarkar V. 2014. Transcriptional regulation of nodule development and senescence in Medicago 659 truncatula: Wageningen University. 660
Kawagoe Y, Campell BR, Murai N. 1994. Synergism between CACGTG (G-box) and CACCTG cis-661 elements is required for activation of the bean seed storage protein β-phaseolin gene. The 662 Plant Journal 5(6): 885-890. 663
Kazmierczak T, Yang L, Boncompagni E, Meilhoc E, Frugier F, Frendo P, Bruand C, Gruber V, 664 Brouquisse R 2020. Chapter Seven - Legume nodule senescence: a coordinated death 665 mechanism between bacteria and plant cells. In: Frendo P, Frugier F, Masson-Boivin C eds. 666 Advances in Botanical Research: Academic Press, 181-212. 667
Kereszt A, Li D, Indrasumunar A, D T Nguyen C, Nontachaiyapoom S, Kinkema M, M Gresshoff P. 668 2007. Agrobacterium rhizogenes-mediated transformation of soybean to study root biology. 669
Konishi M, Yanagisawa S. 2010. Identification of a nitrate-responsive cis-element in the Arabidopsis 670 NIR1 promoter defines the presence of multiple cis - regulatory elements for nitrogen 671 response. The Plant Journal 63(2): 269-282. 672
Konishi M, Yanagisawa S. 2011. Roles of the transcriptional regulation mediated by the nitrate-673 responsive cis-element in higher plants. Biochemical and Biophysical Research 674 Communications 411(4): 708-713. 675
Konishi M, Yanagisawa S. 2013. Arabidopsis NIN-like transcription factors have a central role in nitrate 676 signalling. Nature Communications 4(1): 1-9. 677
Kumar K, Srivastava V, Purayannur S, Kaladhar VC, Cheruvu PJ, Verma PK. 2016. WRKY domain-678 encoding genes of a crop legume chickpea (Cicer arietinum): comparative analysis with 679 Medicago truncatula WRKY family and characterization of group-III gene(s). DNA Research 680 23(3): 225-239. 681
Laloum T, De Mita S, Gamas P, Baudin M, Niebel A. 2013. CCAAT-box binding transcription factors in 682 plants: Y so many? Trends In Plant Science 18(3): 157-166. 683
Lescot M, Dehais P, Thijs G, Marchal K, Moreau Y, Van de Peer Y, Rouze P, Rombauts S. 2002. 684 PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico 685 analysis of promoter sequences. Nucleic Acids Research 30(1): 325-327. 686
Lin J-s, Li X, Luo Z, Mysore KS, Wen J, Xie F. 2018. Author Correction: NIN interacts with NLPs to 687 mediate nitrate inhibition of nodulation in Medicago truncatula. Nature Plants 4(12): 1125-688 1125. 689
Liu J, Rutten L, Limpens E, van der Molen T, van Velzen R, Chen R, Chen Y, Geurts R, Kohlen W, 690 Kulikova O, et al. 2019. A Remote cis-Regulatory Region Is Required for NIN Expression in the 691 Pericycle to Initiate Nodule Primordium Formation in Medicago truncatula. The Plant Cell 692 31(1): 68-83. 693
Matamoros MA, Baird LM, Escuredo PR, Dalton DA, Minchin FR, Iturbe-Ormaetxe I, Rubio MC, 694 Moran JF, Gordon AJ, Becana M. 1999. Stress-induced legume root nodule senescence. 695 Physiological, biochemical, and structural alterations. Plant Physiology 121(1): 97-112. 696
Menkens AE, Schindler U, Cashmore AR. 1995. The G-box: a ubiquitous regulatory DNA element in 697 plants bound by the GBF family of bZIP proteins. Trends In Biochemical Sciences 20(12): 506-698 510. 699
Müller B, Sheen J. 2008. Cytokinin and auxin interaction in root stem-cell specification during early 700 embryogenesis. Nature 453(7198): 1094-1097. 701

24
Nishida H, Tanaka S, Handa Y, Ito M, Sakamoto Y, Matsunaga S, Betsuyaku S, Miura K, Soyano T, 702 Kawaguchi M, et al. 2018. A NIN-LIKE PROTEIN mediates nitrate-induced control of root 703 nodule symbiosis in Lotus japonicus. Nature Communications 9(1): 499. 704
Perez Guerra JC, Coussens G, De Keyser A, De Rycke R, De Bodt S, Van De Velde W, Goormachtig S, 705 Holsters M. 2010. Comparison of developmental and stress-induced nodule senescence in 706 Medicago truncatula. Plant Physiology 152(3): 1574-1584. 707
Pierre O, Hopkins J, Combier M, Baldacci F, Engler G, Brouquisse R, Herouart D, Boncompagni E. 708 2014. Involvement of papain and legumain proteinase in the senescence process of Medicago 709 truncatula nodules. New Phytologist 202(3): 849-863. 710
Pladys D, Vance CP. 1993. Proteolysis during Development and Senescence of Effective and Plant 711 Gene-Controlled Ineffective Alfalfa Nodules. Plant Physiology 103(2): 379-384. 712
Potschin M, Schlienger S, Bieker S, Zentgraf U. 2014. Senescence networking: WRKY18 is an upstream 713 regulator, a downstream target gene, and a protein interaction partner of WRKY53. Journal 714 Of Plant Growth Regulation 33(1): 106-118. 715
Puppo A, Groten K, Bastian F, Carzaniga R, Soussi M, Lucas MM, de Felipe MR, Harrison J, Vanacker 716 H, Foyer CH. 2005. Legume nodule senescence: roles for redox and hormone signalling in the 717 orchestration of the natural aging process. New Phytologist 165(3): 683-701. 718
Quandt HJ. 1993. Transgenic Root Nodules of Vicia hirsuta:A Fast and Efficient System for the Study 719 of Gene Expression in Indeterminate-Type Nodules. Molecular Plant-microbe Interactions 6(6): 720 699-706. 721
Rieping M, Schöffl F. 1992. Synergistic effect of upstream sequences, CCAAT box elements, and HSE 722 sequences for enhanced expression of chimaeric heat shock genes in transgenic tobacco. 723 Molecular and General Genetics MGG 231(2): 226-232. 724
Ross EJ, Stone JM, Elowsky CG, Arredondo-Peter R, Klucas RV, Sarath G. 2004. Activation of the Oryza 725 sativa non-symbiotic haemoglobin-2 promoter by the cytokinin-regulated transcription factor, 726 ARR1. Journal Of Experimental Botany 55(403): 1721-1731. 727
Roux B, Rodde N, Jardinaud MF, Timmers T, Sauviac L, Cottret L, Carrère S, Sallet E, Courcelle E, 728 Moreau S. 2014. An integrated analysis of plant and bacterial gene expression in symbiotic 729 root nodules using laser-capture microdissection coupled to RNA sequencing. Plant Journal 730 77(6): 817-837. 731
Sakai H, Aoyama T, Oka A. 2000. Arabidopsis ARR1 and ARR2 response regulators operate as 732 transcriptional activators. The Plant Journal 24(6): 703-711. 733
Seabra AR, Pereira PA, Becker JD, Carvalho HG. 2012. Inhibition of glutamine synthetase by 734 phosphinothricin leads to transcriptome reprograming in root nodules of Medicago truncatula. 735 Molecular Plant-microbe Interactions 25(7): 976-992. 736
Serova TA, Tikhonovich IA, Tsyganov VE. 2017. Analysis of nodule senescence in pea (Pisum sativum 737 L.) using laser microdissection, real-time PCR, and ACC immunolocalization. Journal Of Plant 738 Physiology 212: 29-44. 739
Shirsat A, Wilford N, Croy R, Boulter D. 1989. Sequences responsible for the tissue specific promoter 740 activity of a pea legumin gene in tobacco. Molecular and General Genetics MGG 215(2): 326-741 331. 742
Shu YJ, Song LL, Zhang J, Liu Y, Guo CH. 2015. Genome-wide identification and characterization of the 743 Dof gene family in Medicago truncatula. Genet Mol Res 14(3): 10645-10657. 744
Syska C, Brouquisse R, Alloing G, Pauly N, Frendo P, Bosseno M, Dupont L, Boscari A. 2019. Molecular 745 weapons contribute to intracellular rhizobia accommodation within legume host cell. 746 Frontiers in Plant Science 10: 1496. 747
Szabados L, Charrier B, Kondorosi A, de Bruijn FJ, Ratet P. 1995. New plant promoter and enhancer 748 testing vectors. Molecular Breeding 1(4): 419-423. 749
Tang H, Krishnakumar V, Bidwell S, Rosen B, Chan A, Zhou S, Gentzbittel L, Childs KL, Yandell M, 750 Gundlach H. 2014. An improved genome release (version Mt4. 0) for the model legume 751 Medicago truncatula. BMC genomics 15(1): 312. 752

25
Trujillo DI, Silverstein KA, Young ND. 2019. Nodule-specific PLAT domain proteins are expanded in 753 the Medicago lineage and required for nodulation. New Phytologist 222(3): 1538-1550. 754
Ülker B, Mukhtar MS, Somssich IE. 2007. The WRKY70 transcription factor of Arabidopsis influences 755 both the plant senescence and defense signaling pathways. Planta 226(1): 125-137. 756
Van de Velde W, Guerra JC, De Keyser A, De Rycke R, Rombauts S, Maunoury N, Mergaert P, 757 Kondorosi E, Holsters M, Goormachtig S. 2006. Aging in legume symbiosis. A molecular view 758 on nodule senescence in Medicago truncatula. Plant Physiology 141(2): 711-720. 759
Vijn I, Christiansen H, Lauridsen P, Kardailsky I, Quandt H-J, Broer I, Drenth J, Jensen EØ, van 760 Kammen A, Bisseling T. 1995. A 200 bp region of the pea ENOD12 promoter is sufficient for 761 nodule-specific and Nod factor induced expression. Plant Molecular Biology 28(6): 1103-1110. 762
Wenkel S, Turck F, Singer K, Gissot L, Le Gourrierec J, Samach A, Coupland G. 2006. CONSTANS and 763 the CCAAT box binding complex share a functionally important domain and interact to 764 regulate flowering of Arabidopsis. The Plant Cell 18(11): 2971-2984. 765
Xie Z, Zhang Z-L, Zou X, Huang J, Ruas P, Thompson D, Shen QJ. 2005. Annotations and functional 766 analyses of the rice WRKY gene superfamily reveal positive and negative regulators of abscisic 767 acid signaling in aleurone cells. Plant Physiology 137(1): 176-189. 768
Yanagisawa S. 2000. Dof1 and Dof2 transcription factors are associated with expression of multiple 769 genes involved in carbon metabolism in maize. The Plant Journal 21(3): 281-288. 770
Yanagisawa S. 2002. The Dof family of plant transcription factors. Trends In Plant Science 7(12): 555-771 560. 772
Yanagisawa S, Schmidt RJ. 1999. Diversity and similarity among recognition sequences of Dof 773 transcription factors. The Plant Journal 17(2): 209-214. 774
Yang L, El Msehli S, Benyamina S, Lambert A, Hopkins J, Cazareth J, Pierre O, Hérouart D, Achi-Smiti 775 S, Boncompagni E, et al. 2020a. Glutathione Deficiency in Sinorhizobium meliloti Does Not 776 Impair Bacteroid Differentiation But Induces Early Senescence in the Interaction With 777 Medicago truncatula. Frontiers in Plant Science 11(137). 778
Yang L, Syska C, Garcia I, Frendo P, Boncompagni E 2020b. Involvement of proteases during nodule 779 senescence in leguminous plants. In: Bruijn Fd ed. The Model Legume Medicago truncatula, 780 683-693. 781
Yuan SL, Li R, Chen HF, Zhang CJ, Chen LM, Hao QN, Chen SL, Shan ZH, Yang ZL, Zhang XJ, et al. 2017. 782 RNA-Seq analysis of nodule development at five different developmental stages of soybean 783 (Glycine max) inoculated with Bradyrhizobium japonicum strain 113-2. Scientific Reports 7: 784 42248. 785
Zhang ZL, Xie Z, Zou X, Casaretto J, Ho TH, Shen QJ. 2004. A rice WRKY gene encodes a transcriptional 786 repressor of the gibberellin signaling pathway in aleurone cells. Plant Physiology 134(4): 1500-787 1513. 788
Zürcher E, Liu J, di Donato M, Geisler M, Müller B. 2016. Plant development regulated by cytokinin 789 sinks. Science 353(6303): 1027-1030. 790
791
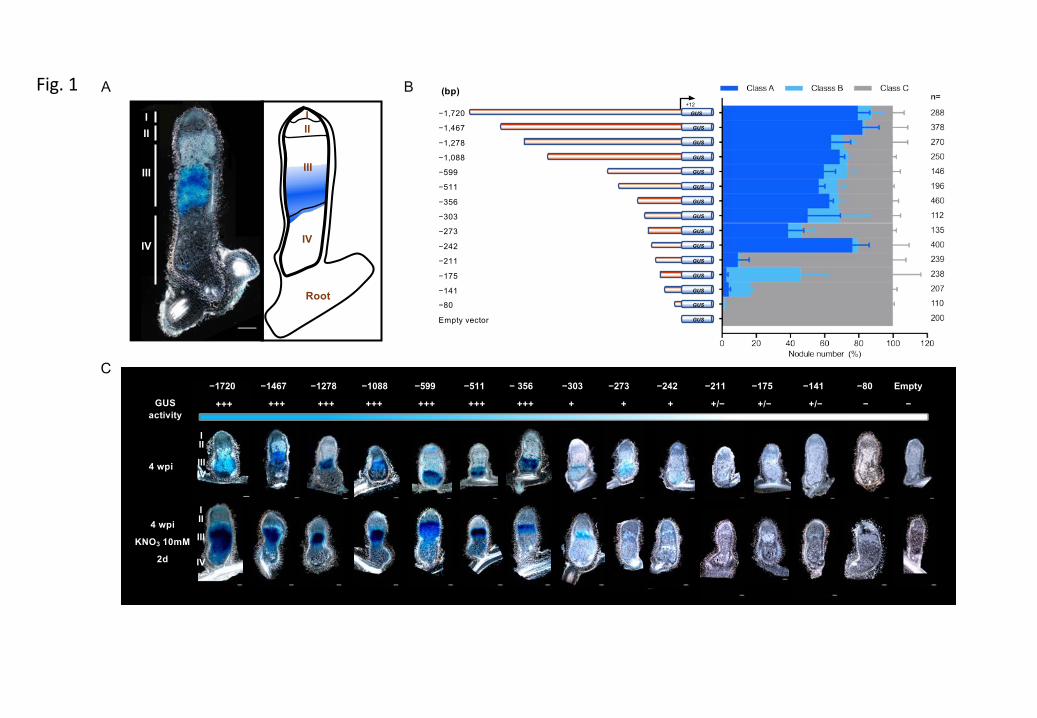
+12GUS−1,720
GUS−1,467
GUS−1,278
GUS−1,088GUS−599
GUS−511
GUS−356GUS−303
GUS−273
GUS−242
GUS−211
GUS−175
GUS−141GUS−80
GUSEmpty vector
(bp)
vz
−1720 −175−211−273−303 Empty−80−141− 356−511−1088 −599−1278−1467
4 wpi
4 wpi
KNO3 10mM
2d
+++GUS activity
+++ +++ +++ +++ +++ +++ + + + +/− +/− +/− − −
GUS activity: +++ strong + detectable − non-detectable
−242
A B
C
III
IIIIV
III
III
IV
III
III
IV
II
III
IV
I
Root
Fig. 1
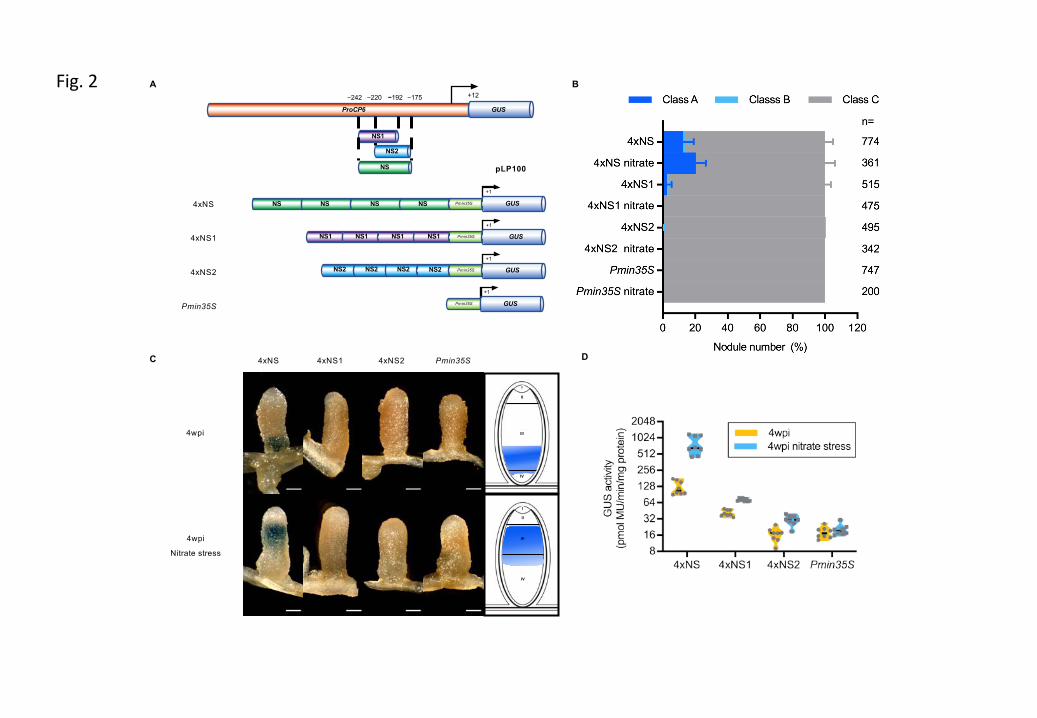
Fig. 2
NS
GUSProCP6
−242 −175
NS1
NS2
−192 +12−220
A B
C D
NS2 NS2 NS2 NS2 GUS
+1
Pmin35S
NS1 NS1 NS1 NS1 GUS
+1
Pmin35S
NS NS NS NS GUS
+1
Pmin35S
GUS
+1
Pmin35S
4xNS
4xNS1
4xNS2
Pmin35S
pLP100
4xNS1 4xNS24xNS Pmin35S
4wpi
4wpi
Nitrate stress
I
II
III
IV
I
II
III
IV
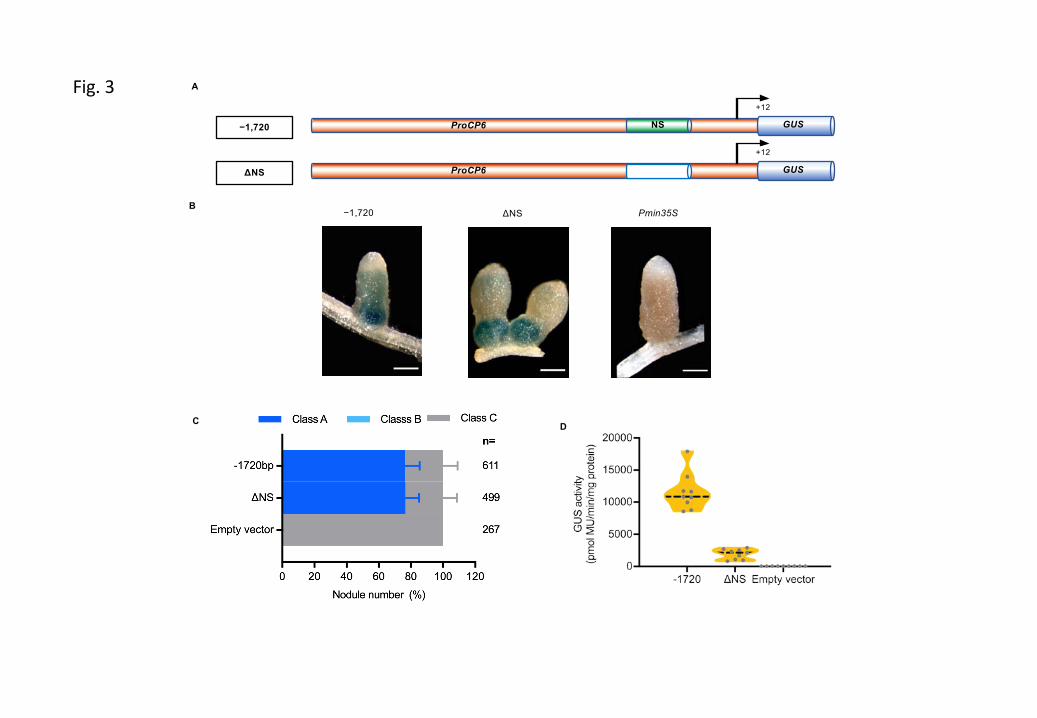
Fig. 3
NS GUSProCP6
+12
−1,720
GUSProCP6
+12
ΔNS
−1,720 ΔNS Pmin35S
A
B
C D
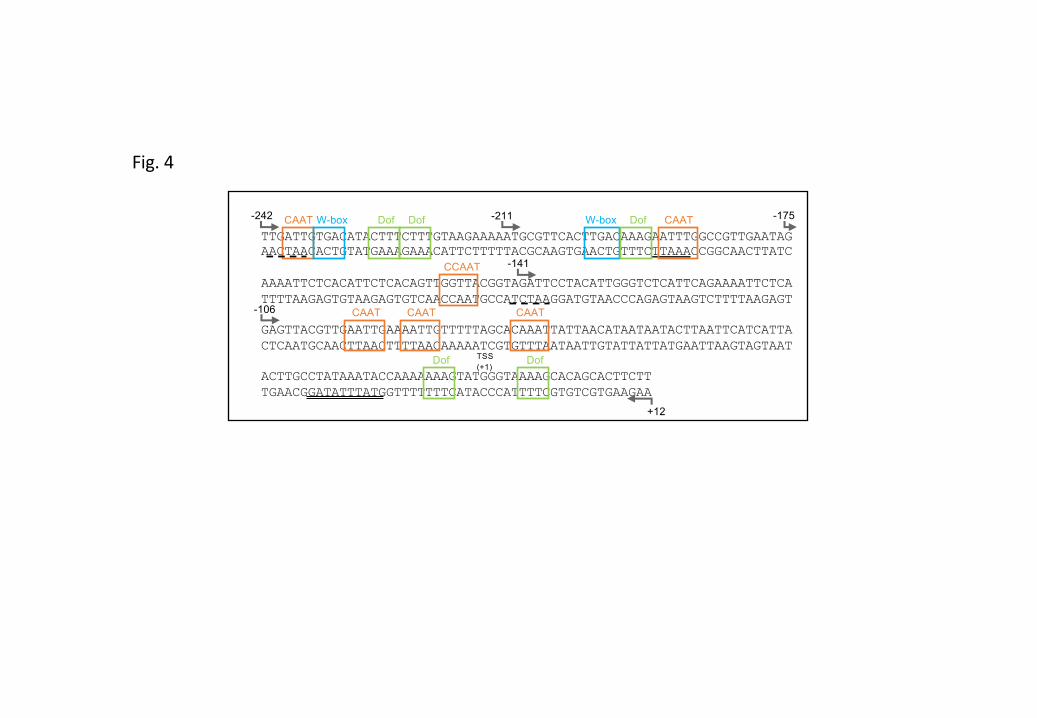
TTGATTGTGACATACTTTCTTTGTAAGAAAAATGCGTTCACTTGACAAAGAATTTGGCCGTTGAATAGAACTAACACTGTATGAAAGAAACATTCTTTTTACGCAAGTGAACTGTTTCTTAAACCGGCAACTTATC
AAAATTCTCACATTCTCACAGTTGGTTACGGTAGATTCCTACATTGGGTCTCATTCAGAAAATTCTCATTTTAAGAGTGTAAGAGTGTCAACCAATGCCATCTAAGGATGTAACCCAGAGTAAGTCTTTTAAGAGT
GAGTTACGTTGAATTGAAAATTGTTTTTAGCACAAATTATTAACATAATAATACTTAATTCATCATTACTCAATGCAACTTAACTTTTAACAAAAATCGTGTTTAATAATTGTATTATTATGAATTAAGTAGTAAT
ACTTGCCTATAAATACCAAAAAAAGTATGGGTAAAAGCACAGCACTTCTTTGAACGGATATTTATGGTTTTTTTCATACCCATTTTCGTGTCGTGAAGAA
+12
CAAT W-box W-box
-141
-242 -175Dof Dof -211
-106
Dof
CCAAT
CAAT CAAT
TSS (+1)
DofDof
CAAT
CAAT
Fig. 4
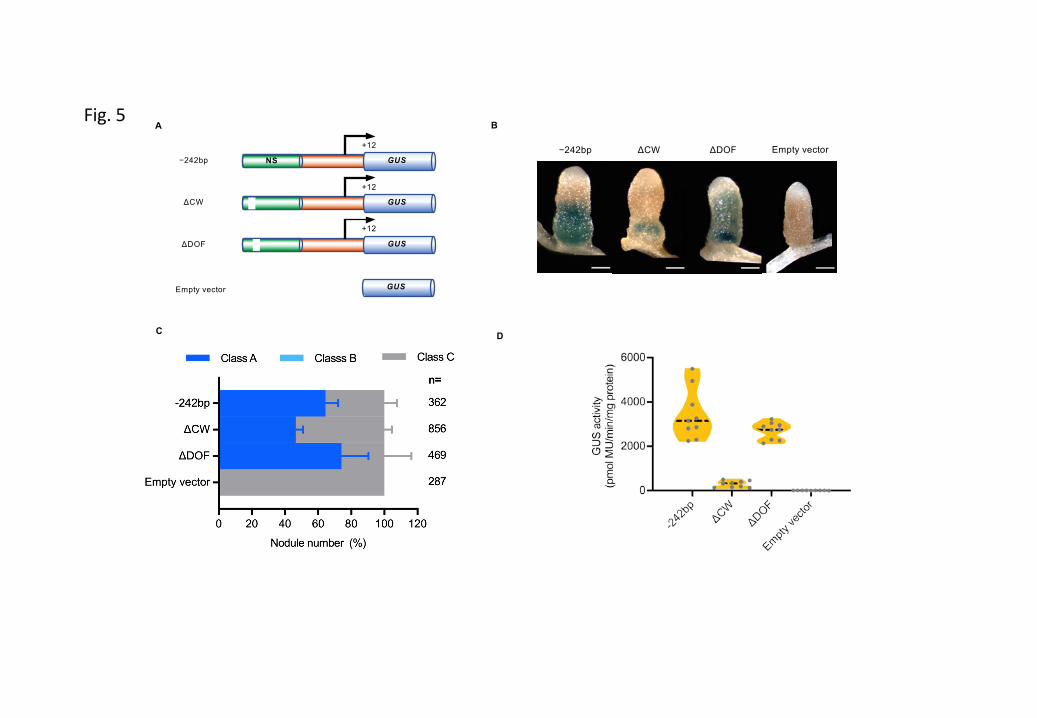
Fig. 5
ΔCW−242bp ΔDOF Empty vector
A
C D
B
ΔCW
NS GUS
+12
−242bp
GUS
+12
GUS
+12
ΔDOF
GUSEmpty vector

Table S1: Primer sequence list
Primers Sequences (5’ – 3’)−1720 F (attB4) GGGGACAACTTTGTATAGAAAAGTTGTCCGCCATCTATCTTCATTCGTCA−1467 F (attB4) GGGGACAACTTTGTATAGAAAAGTTGTCTGAGAGAGATGAATGGTACTAT −1278 F (attB4) GGGGACAACTTTGTATAGAAAAGTTGTCATGAGATGAGATGAGATGAGATGA−1088 F (attB4) GGGGACAACTTTGTATAGAAAAGTTGTCGATAGTGATTGAGGACCGCT −599 F (attB4) GGGGACAACTTTGTATAGAAAAGTTGTCTGAAATAAAGAGCTATTTGTTG −511 F (attB4) GGGGACAACTTTGTATAGAAAAGTTGTCTTGTAAAAAATAATCAAAATTATCTA −356 F (attB4) GGGGACAACTTTGTATAGAAAAGTTGTCCCATAGAATTTGCCATCACAAT −303 F (attB4) GGGGACAACTTTGTATAGAAAAGTTGTCCATCACTTAGTTACGTGAAGCA−273 F (attB4) GGGGACAACTTTGTATAGAAAAGTTGTCACACGTTAGTTAACGTGAAACA−242 F (attB4) GGGGACAACTTTGTATAGAAAAGTTGTCTTGATTGTGACATACTTTCTTT−211 F (attB4) GGGGACAACTTTGTATAGAAAAGTTGTCATGCGTTCACTTGACAAAGAAT−175 F (attB4) GGGGACAACTTTGTATAGAAAAGTTGTCGAAAATTCTCACATTCTCACAG−141 F (attB4) GGGGACAACTTTGTATAGAAAAGTTGTCGATTCCTACATTGGGTCTCATT−80 F (attB4) GGGGACAACTTTGTATAGAAAAGTTGTCTTAGCACAAATTATTAACATAATAattB1R-ProCP6 GGGGACTGCTTTTTTGTACAAACTTGGAAGAAGTGCTGTGCTTTTACCC empty vecoter F (attB4-attB3) GGGGACAACTTTGTATAGAAAAGTTGTCCAAGTTTGTACAAAAAAGCAGTCCCCempty vecoter R (attB4-attB3) GGGGACTGCTTTTTTGTACAAACTTGGACAACTTTTCTATACAAAGTTGTCCCCM13F GTAAAACGACGGCCAG M13R CAGGAAACAGCTATGAC pLP100-MSC F TGCCACCTGACGTCTAAGAAACGUS R AAGACTTCGCGCTGATACCΔNS 5' R CTGTGAGAATGTGAGAATTTTCTAACTCAGTTTGTTTCACGTTAACΔNS 3' F GTTAACGTGAAACAAACTGAGTTAGAAAATTCTCACATTCTCACAGΔCW F CTTTGTATAGAAAAGTTGTCTTGTACTTTCTTTGTAAGAAAAATGC ΔCW R GCATTTTTCTTACAAAGAAAGTACAAGACAACTTTTCTATACAAAG ΔDOF F GAAAAGTTGTCTTGATTGTGACATAGTAAGAAAAATGCGTTCACTTGACAΔDOF R TGTCAAGTGAACGCATTTTTCTTACTATGTCACAATCAAGACAACTTTTC
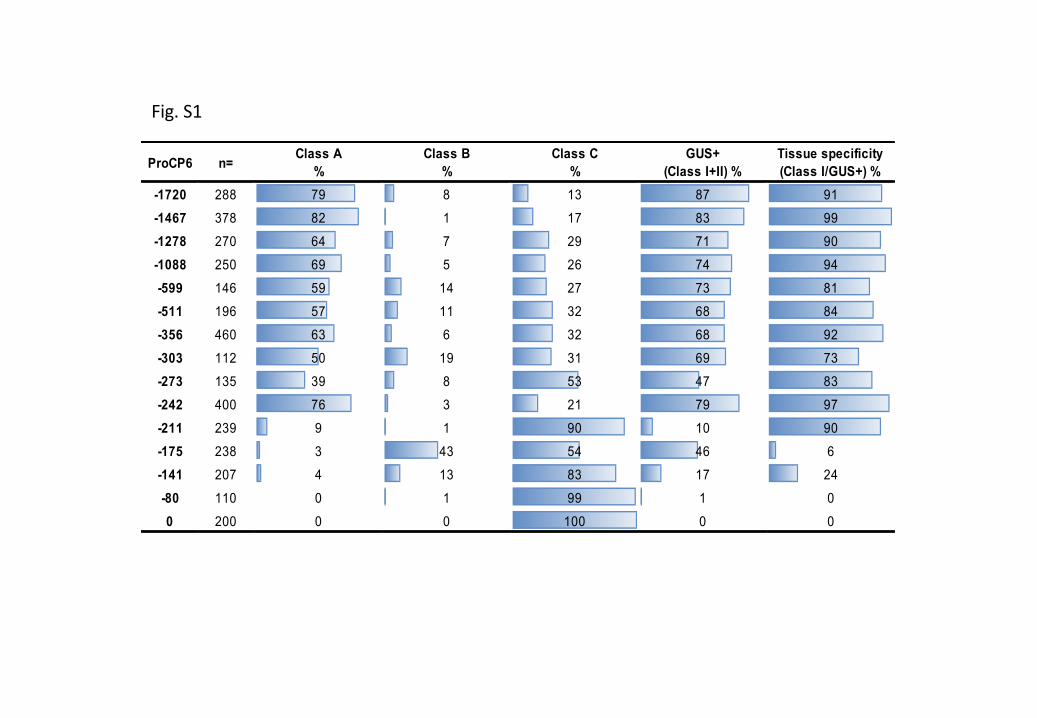
ProCP6 n=Class A
%Class B
%Class C
%GUS+
(Class I+II) %Tissue specificity (Class I/GUS+) %
-1720 288 79 8 13 87 91-1467 378 82 1 17 83 99-1278 270 64 7 29 71 90-1088 250 69 5 26 74 94-599 146 59 14 27 73 81-511 196 57 11 32 68 84-356 460 63 6 32 68 92-303 112 50 19 31 69 73-273 135 39 8 53 47 83-242 400 76 3 21 79 97-211 239 9 1 90 10 90-175 238 3 43 54 46 6-141 207 4 13 83 17 24-80 110 0 1 99 1 00 200 0 0 100 0 0
Fig. S1
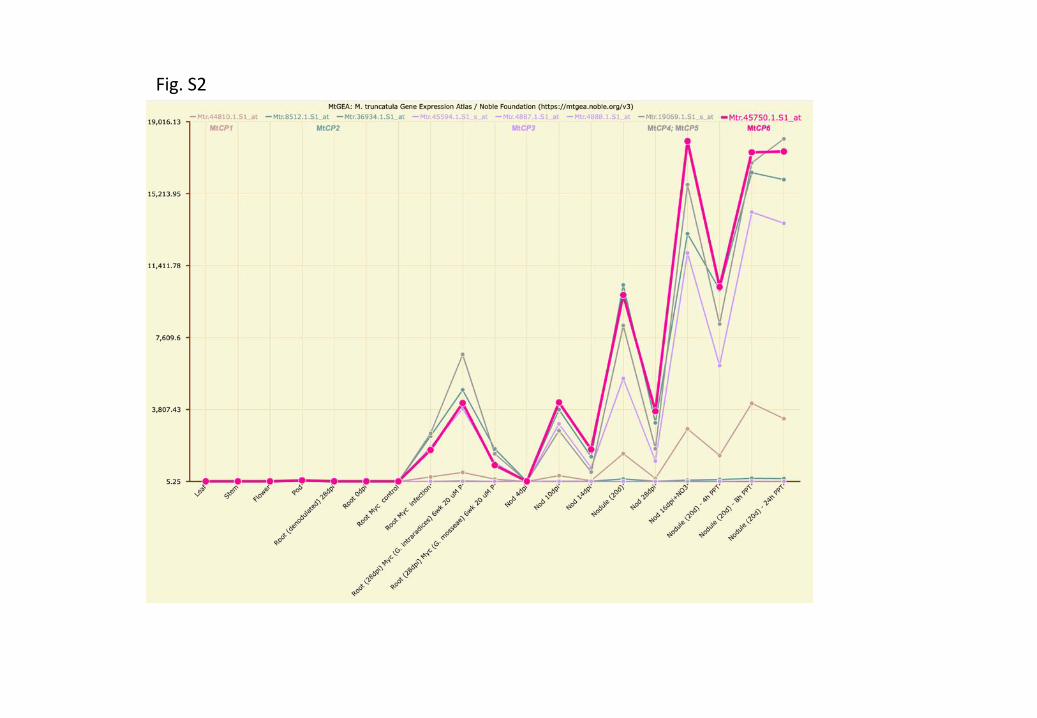
Fig. S2
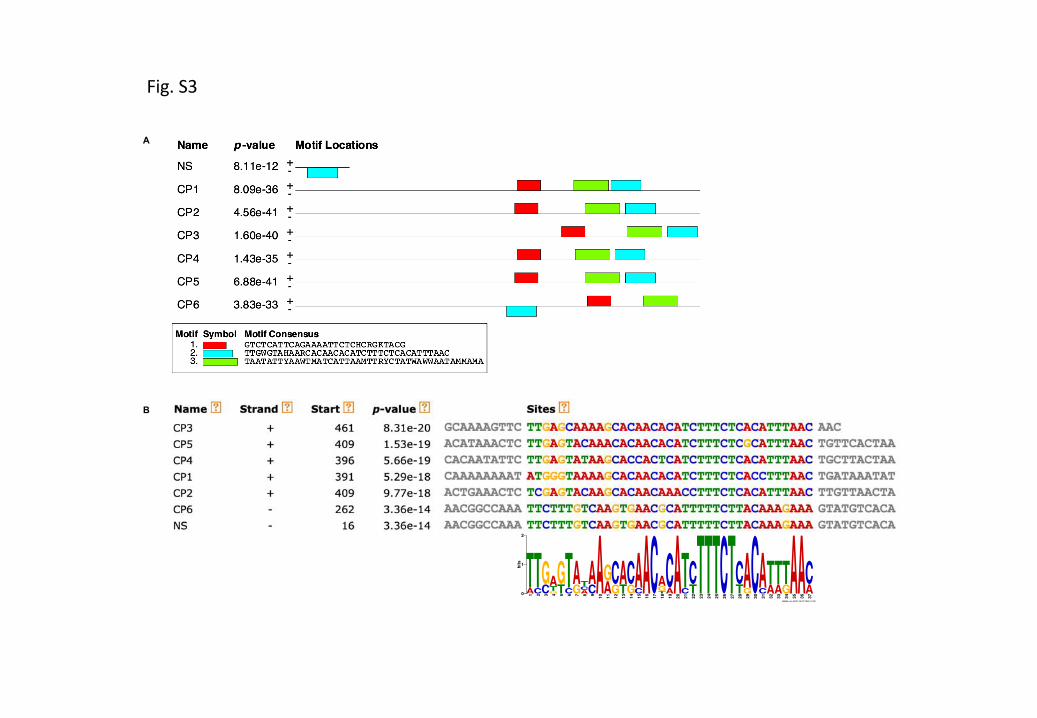
A
B
MEME (no SSC) 20.07.2020 12:35
0
1
2
bits
1AT
2CT
3CG
4GTA
5TG
6CT
7GA
8ACT
9CA
10
A11
AG
12
GC
13
TA
14
GC
15
CA
16
A17
C18
TGA
19
AC
20
A21
CT
22
TC
23
T24
T25
T26
C
27
T
28
TC
29
GA
30
C
31
CA
32
AT
33
AT
34
GT
35
A
36
A
37
AC
Fig. S3
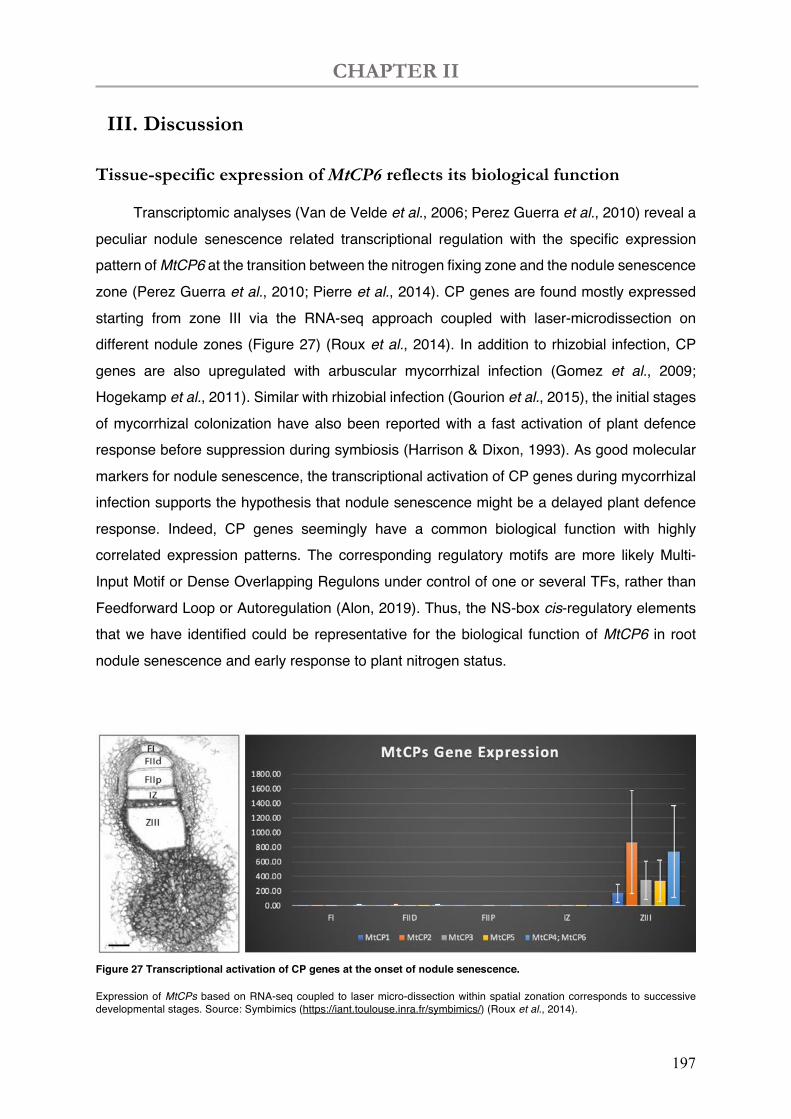
CHAPTER II
197
III. Discussion
Tissue-specific expression of MtCP6 reflects its biological function
Transcriptomic analyses (Van de Velde et al., 2006; Perez Guerra et al., 2010) reveal a peculiar nodule senescence related transcriptional regulation with the specific expression pattern of MtCP6 at the transition between the nitrogen fixing zone and the nodule senescence zone (Perez Guerra et al., 2010; Pierre et al., 2014). CP genes are found mostly expressed starting from zone III via the RNA-seq approach coupled with laser-microdissection on different nodule zones (Figure 27) (Roux et al., 2014). In addition to rhizobial infection, CP genes are also upregulated with arbuscular mycorrhizal infection (Gomez et al., 2009; Hogekamp et al., 2011). Similar with rhizobial infection (Gourion et al., 2015), the initial stages of mycorrhizal colonization have also been reported with a fast activation of plant defence response before suppression during symbiosis (Harrison & Dixon, 1993). As good molecular markers for nodule senescence, the transcriptional activation of CP genes during mycorrhizal infection supports the hypothesis that nodule senescence might be a delayed plant defence response. Indeed, CP genes seemingly have a common biological function with highly correlated expression patterns. The corresponding regulatory motifs are more likely Multi-Input Motif or Dense Overlapping Regulons under control of one or several TFs, rather than Feedforward Loop or Autoregulation (Alon, 2019). Thus, the NS-box cis-regulatory elements that we have identified could be representative for the biological function of MtCP6 in root nodule senescence and early response to plant nitrogen status.
Figure 27 Transcriptional activation of CP genes at the onset of nodule senescence.
Expression of MtCPs based on RNA-seq coupled to laser micro-dissection within spatial zonation corresponds to successive developmental stages. Source: Symbimics (https://iant.toulouse.inra.fr/symbimics/) (Roux et al., 2014).

CHAPTER II
198
In this study, serial promoter deletion analysis of MtCP6 allowed us to decipher the transcriptional regulation of the tissue specific expression manner of MtCP6. To further clarify the temporal pattern of MtCP6 gene expression, we carried out kinetic study at 1, 2, 4 and 6wpi to show temporal expression on transgenic nodules carrying MtCP6 promoters (–1720bp and –242bp), as well as empty vector. (Annex IV, Figure S 3). The induction of tested MtCP6 promoter expressions has started in young nodules since 1wpi. This result is comparable to microarray data of MtCP6 expression (MtGEA), in which MtCP6 induction was detected as early as 10dpi. However, only a small proportion of nodules carrying MtCP6 promoters were stained blue in young nodules, whilst more than 94% of nodules were detected as GUS positive. Hence, we confirmed the temporal transcriptional activity MtCP6 is induced in later stage of nodule development. It appears that spatial and temporal expressions are correlated in determinate nodules. In addition, we have also observed occasional expression in nodule meristems and roots. This might be due to rare expression of MtCP6 in roots or its response to A. rhizogenes, or it could result from the promoter leakage.
IV. Perspectives
NS-box-based promoters could serve for tissue-specific and transactivation
studies at the onset of nodule senescence
The NS-box contains the minimal regulatory elements required for the transcriptional activation of MtCP6 which is specifically expressed at the onset of nodule senescence. For example, it would be possible to apply the synthetic promoter of the NS tetramer in nodule senescence-related studies. For instance, the tissue-specific feature might be used for regulation studies using a GAL4-VP16/UAS transactivation system (Sevin-Pujol et al., 2017). Alternatively, the NS-box synthetic promoter might also be used as sensor to monitor plant nitrogen status if we consider its high sensitivity to nitrate treatment. However, the transcriptional output of the NS-box is not restricted in nitrate response, as it exhibits a dynamic spatiotemporal pattern. It is more likely that the NS-box would be able to be used as a more general sensor of the nodule fitness status. In this context, more evidences are necessary to prove a multi-response of the NS-synthetic promoter to biotic and abiotic factors. Additionally, the activity of a synthetic promoter is determined by the sequence conservation, number of binding sites, identity of flanking, orientation and distance between the core motifs (Zurcher et al., 2013; Wu et al., 2018). An improvement on NS-based promoter might be done

CHAPTER II
199
to robustly monitor the fitness status of the nodules and the transition from symbiotic interaction to senescence.
A novel cis-regulatory element might shed light on the transcriptional
network initiating nodule senescence
We have defined in our study the NS-box from 5’ flanking sequences of MtCP6 that is related in the early transcription of root nodule senescence. The synthetic promoter composed of an NS tetramer can independently restore the tissue-specific spatiotemporal regulation similar to the promoter of MtCP6. The NS-box is presenting various characteristics with responses to developmental senescence to timescale and induced senescence under changes of abiotic conditions. Due to the fact that developmental and induced senescence might be regulated by distinct cis-elements via multiple pathways, it would be more promising to identify the interacting proteins of the NS-box under different conditions. Thus, this experimentally validated cis-regulatory element might be used to capture the dynamic chromatin interactions orchestrating the nodule senescence-related regulation. Under this perspective, gene-centered approaches could be used to search for interacting proteins, such as the yeast-one-hybrid assay (Fuxman Bass et al., 2016) which we have applied in the next Chapter. Our study should help to decipher the complex biological network that regulates nodule senescence from a detailed description on particular regulators.

200

201
($!$
.$
$
*$#
,-

202

203
CHAPTER III
Searching for NS-box-interacting transcription factors

204

CHAPTER III
205
I. Introduction
I.1 Searching for NS-box-interacting factors
The tetramer of the nodule senescence related cis-regulatory element (4x NS-box), has shown in this study to mediate the expression of MtCP6 during developmental nodule senescence and nodule response to nitrogen status. It is still questioned thereafter which transcription factors (TFs) are interacting with the NS-box. TFs are regulatory proteins that modulate the transcription activity of a given gene by binding to DNA-regulatory sequences and/or via the interaction with other transcriptional regulators, including chromatin remodelling/modifying proteins, to recruit or block the access of RNA polymerases to the DNA template (Udvardi et al., 2007). TF proteins are generally recognized by the presence of a conserved DNA-binding domain (DB) (Mitchell, Pamela J & Tjian, Robert, 1989; Ptashne & Gann, 1997) which may be conserved across eukaryotic organisms or plant specific (Libault et al., 2009). TFs can act as activators (enhancers) or repressors to change mRNA production of RNA polymerase II (Figure 28) (Alon, 2019). Expression of transcriptional activators are expected to have overlapped expression patterns to the corresponding target genes, while the repressors might display opposite expression patterns (Franco-Zorrilla et al., 2014).
Figure 28 Schematic representation of the transcriptional machinery under control of activator (A) and repressor (B)
TFs, respectively. Adapted from (Alon, 2019).
A B

CHAPTER III
206
TF-coding genes accounts for a large number of genes in plant genomes, ranging from 6% to 10%, with 2000 TF encoding genes in Arabidopsis (Riechmann et al., 2000; Riechmann, 2018). In M. truncatula, 1298 putative TF genes, mostly (1169) falling in 45 known plant TF families, were identified on the Affymetrix Medicago GeneChip (Udvardi et al., 2007). In the gene expression atlas of M. truncatula, key regulators such as TFs are revealed to orchestrate genetic reprograming during development and differentiation among organs (Benedito et al., 2008). Among these putative TF genes, 532 are differentially expressed during nodule development and may therefore play important roles in symbiotic nitrogen fixation (Benedito et al., 2008).
I.2 TFs associated with nodule senescence processes in legume plants
To date, many TFs have been functionally characterized to play key roles during nodulation, for example, GRAS family proteins (Kalo et al., 2005; Smit et al., 2005; Hirsch et al., 2009), CYCLOPS (Yano et al., 2008; Singh et al., 2014), DELLA,(Fonouni-Farde et al., 2016b; Jin et al., 2016; Fonouni-Farde et al., 2017), ERN (Andriankaja et al., 2007; Cerri et al., 2012; Cerri et al., 2017) and NF-Y (Combier et al., 2006; Laloum et al., 2014; Laporte et al., 2014; Baudin et al., 2015). The first TF found in legume plants, NIN, has been intensively studied in both cis- and trans- regulation with prominent roles in rhizobial infection and nodule organogenesis (Schauser et al., 1999; Marsh et al., 2007; Vernié et al., 2015; Liu et al., 2019). ZPT2-1 (Frugier et al., 2000) and EFD (Vernié et al., 2008) were related to nodule differentiation process.
During nodule senescence stage, only a few of TFs have been identified and essentially only by correlative transcriptomic analyses, e.g. MYB (MYeloBlastosis), bHLH (basic Helix-Loop-Helix), and NAC (NAM ATAF CUC) families (Kazmierczak et al., 2020; Van de Veldeet al., 2006; Seabra et al., 2012; Cabeza et al., 2014; Chungopast et al., 2014; Yuan et al., 2017). Nevertheless, a few of them have been functional characterized and directly linked to the transition between N-fixation to nodule senescence. MtATB2 (bZIP family) is regulated by sucrose and is enhanced during nodule senescence (D'Haeseleer et al., 2010), while the TF MtbHLH2 was recently reported to be a transcriptional repressor of a cysteine protease encoding gene (MtCP77) and down-regulated accordingly in nodule senescence (Deng et al., 2019). Regarding to TFs regulating induced nodule senescence, MtNAC969 was evidenced to be upregulated during nitrate treatment, but it was negatively related to nodule developmental senescence (de Zelicourt et al., 2012).

CHAPTER III
207
I.3 Search for trans-regulatory elements interacting with NS-box
To search for interactive trans-proteins that might interact with the cis-regulatory NS-box sequence identified in this study , we chose to use the DNA-centered Yeast-One-Hybrid (Y1H) screening approach (Li & Herskowitz, 1993) (Fuxman Bass et al., 2016), derived from the yeast-two-hybrid approach used to identify protein-protein interactors (Fields & Song, 1989). The Y1H strategy, firstly developed to detect yeast proteins that can bind to specific DNA sequences (Li & Herskowitz, 1993; Wang & Reed, 1993). The approach is based on the generation of a ‘bait’ DNA sequence that is inserted in front of a reporter gene that is then integrated by recombination into yeast genome. Auxotrophic gene reporters (HIS3, URA3, TRP1, and LEU2), which allow yeast growth on corresponding amino acid deficient selective medium are generally used here. While the prey protein (DNA-BP) is fused to an activation domain (AD) of yeast TF Gal4 in a ‘prey’ hybrid vector (thus named as one-hybrid). High affinity interaction of the cis- element and the trans-factor would bring the AD close to the transcription initiation site of the reporter gene, thereby enabling yeast growth in auxotrophic conditions and selection of a positive yeast clone. However, false-positive yeast candidates can grow because of a basal level of transcription of the reporter gene (e.g. histidine) by non-specific interaction of endogenous yeast proteins to the cis-binding sites. Thus, a proper concentration of histidine biosynthesis inhibitor 3-amino-1,2,4-trizol (3-AT) is often used in these screens in order to limit the background.
One hybrid screen has been successfully used to identify several plant TFs, e.g. during nodulation (Andriankaja et al., 2007), lipid transfer during arbuscular mycorrhiza symbiosis (Jiang et al., 2018) and plant cold responses (Zhang et al., 2016). Indeed, this approach has led to the identification of ERF transcription factors (ERNs) that bind to the NF-responsive box. We therefore decided to use a similar approach to identify TF-interacting factors of the NS-box, which is required and sufficient for driving MtCP6-related nodule senescence-associated transcription. By using a tetramer of the NS-box, this chapter describe the yeast one-hybrid screen of a nodule cDNA library. After screening 1.1 X 105 yeast clones and confirmation of positive candidates by yeast retransformation, this screen has resulted to date in the identification of 7 ERF transcription factors, among them 5 belonging to the same ERF subgroup IX of ERF transcription factors, which are potential interesting candidates likely involved in the regulation of nodule-associated expression of MtCP6.

CHAPTER III
208
II. Material and Methods
Yeast-One-Hybrid screening
The Yeast-One-Hybrid screening of the tetramer of NS-box (4x NS-box) for interactive TF candidates was done as previously described (Andriankaja et al., 2007). The tetramer of 4x tandem NS-box sequences was inserted into the pHISi vector (Clontech) via the EcoRI/MluI restriction sites to generate the 4xNS-box-HIS3 construct, where the NFbox cis-sequences are placed in front of the Histidine reporter. The yeast strain YM4271 carrying the bait tetramer NS-box was generated and validated by L. Frances in the context of a collaboration with F. de Carvalho-Niebel’s team (LIPM, Toulouse). Briefly, she linearized the 4xNS-box pHISi plasmid by XhoI digestion and transformed it to be integrated into the genome of the YM4271 yeast strain. The positive yeast clones were selected on their ability to complement the HIS- auxotrophic phenotype and yeast colony PCR and sequencing has subsequently confirmed the integration of the 4xNS-box-HIS3 fusion in the yeast genome. A positive yeast recombinant (YM4271-4NS strain) showing a relatively lower background growth in selective 10 to 15mM 3-Amino-1,2,4-triazole (3-AT) conditions, was selected to be used in the further one-hybrid screen. The One-hybrid screen was done using a nodule AD fusion cDNA library, generated before by the Dualsystems company (http://www.dualsystems.com/) from RNA samples extracted from 4 and 19 days-old nodules (and kindly provided by A. Niebel) (Baudin et al., 2015). This screen used a concentration of 10mM 3-AT in the yeast media. Yeast transformation methods and media composition are all detailed in the Method section (ANNEX III) of this PhD thesis.
His+ yeast colonies were recovered in 10µl of water and 2-3 µl were re-spotted on SD/−Leu/−His plates with 15mM 3-AT to confirm growth in selective conditions. Positive candidates were analysed by PCR on 2µl of yeast suspension using primers (ANNEX III Table S3). After sequencing using the same primers, clones exhibiting ORFs in frame with the AD activation domain were further analysed by BLASTN and BLASTX algorithms (Altschul et al., 1997) using Medicago Mt4.0 (http://blast.jcvi.org/Medicago-Blast/index.cgi) and MtrunA17r5.0-ANR (https://medicago.toulouse.inra.fr/MtrunA17r5.0-ANR/) genomic databases. Among the 29 in frame clones (Table 8), eight encode for putative TFs mostly of the ERF TF family. We therefore decided to try to validate these candidates by retransformation of the yeast 4xNS-box strain.
Plasmid DNAs from selected His+ candidate clones were prepared and used to
transform Escherichia coli DH5a. Plasmids of the NS-interacting ERF candidates and ERN1

CHAPTER III
209
(Andrinakaja), extracted from E. coli, were then used for sequencing and/or retransformation of yeast bait strains 4x 4NS-box (this study) and 4x NF-box (Andriankaja et al., 2007). Dilution series (DO= 1, 0,1 and 0,01) of the yeast transformants was spotted on SD-L, SD-H-L, and SD-H-L + 15 mM and their growth was then monitored for 3-4 days, for evaluating the specificity of the interaction towards the 4x NS-box or 4x NF-box regulatory sequences.
III. Results
Yeast-one-hybrid (Y1H) screening for NS-box interactive proteins
Before initiating the one-hybrid screen, it was important to first (i) generate the yeast YM4271 bait strain carrying the 4x NS-box-HIS3 reporter and to (ii) select the best yeast bait candidate showing low HIS3 expression levels that can be inhibited by 3-AT (Histidine enzyme inhibitor) for performing the screen. The generation and selection of such strain, named YM4271-4xNF-box-His3 was done before at the LIPM as described in the Method sections. When I arrived in Toulouse, we have then used this strain to test the optimal concentration of 3-AT to be added to the media for inhibiting the HIS3 background levels. The YM4271-4xNS-His3 strain was grown on SD/−His plates with increasing concentrations of 3-AT (0, 1, 2, 4, 6, 8 and 10mM). At 3 days post-transformation, the concentration of 10mM 3-AT was effective to reduce the background levels of the bait yeast strain. We therefore decided to perform the one-hybrid screen on SD/-Leu/-His media supplemented with 10 mM 3-AT. The Y1H screening strategy used for the identification of NS-box-interacting TFs is schematically represented in Figure 29.
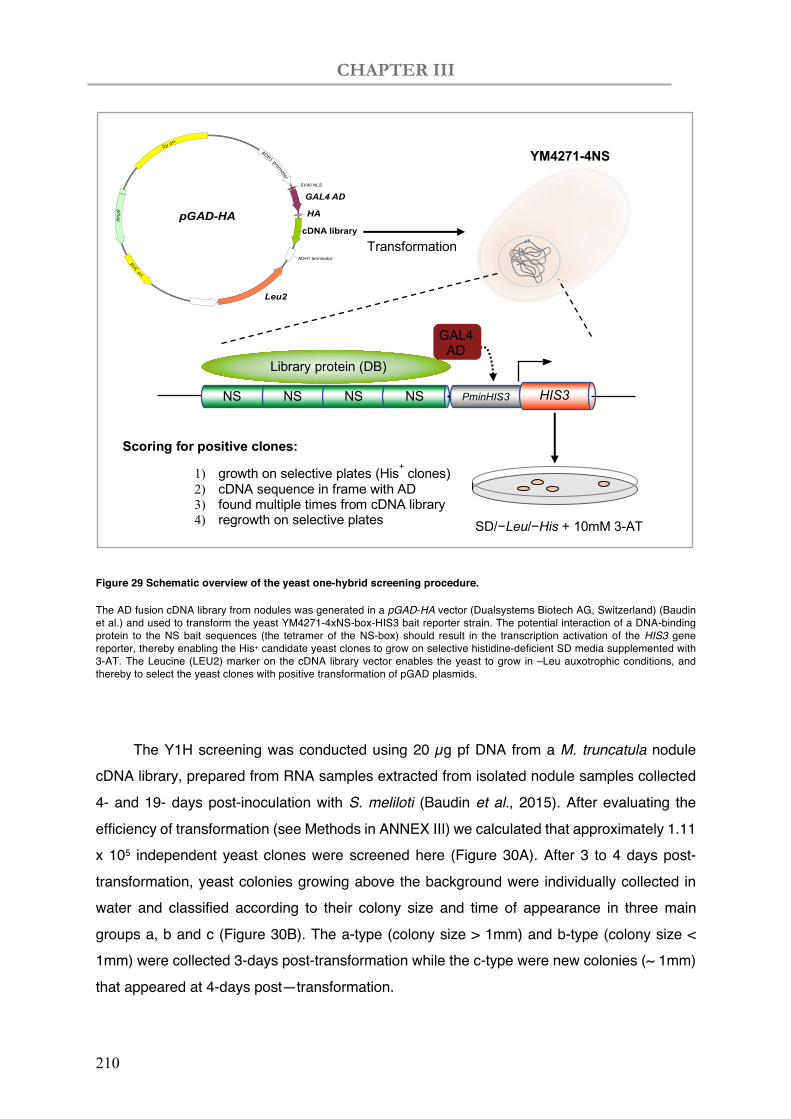
CHAPTER III
210
Figure 29 Schematic overview of the yeast one-hybrid screening procedure.
The AD fusion cDNA library from nodules was generated in a pGAD-HA vector (Dualsystems Biotech AG, Switzerland) (Baudin et al.) and used to transform the yeast YM4271-4xNS-box-HIS3 bait reporter strain. The potential interaction of a DNA-binding protein to the NS bait sequences (the tetramer of the NS-box) should result in the transcription activation of the HIS3 gene reporter, thereby enabling the His+ candidate yeast clones to grow on selective histidine-deficient SD media supplemented with 3-AT. The Leucine (LEU2) marker on the cDNA library vector enables the yeast to grow in –Leu auxotrophic conditions, and thereby to select the yeast clones with positive transformation of pGAD plasmids.
The Y1H screening was conducted using 20 µg pf DNA from a M. truncatula nodule
cDNA library, prepared from RNA samples extracted from isolated nodule samples collected 4- and 19- days post-inoculation with S. meliloti (Baudin et al., 2015). After evaluating the efficiency of transformation (see Methods in ANNEX III) we calculated that approximately 1.11 x 105 independent yeast clones were screened here (Figure 30A). After 3 to 4 days post-transformation, yeast colonies growing above the background were individually collected in water and classified according to their colony size and time of appearance in three main groups a, b and c (Figure 30B). The a-type (colony size > 1mm) and b-type (colony size < 1mm) were collected 3-days post-transformation while the c-type were new colonies (~ 1mm) that appeared at 4-days post—transformation.
Transformation
YM4271-4NS
NS NS NS NS HIS3 PminHIS3
GAL4 AD
SD/−Leu/−His + 10mM 3-AT
Scoring for positive clones:
1) growth on selective plates (His+ clones) 2) cDNA sequence in frame with AD 3) found multiple times from cDNA library 4) regrowth on selective plates
SV40 NLS
HA
ADH1 terminator
pGAD-HAAmpR
GAL4 AD
cDNA library
Leu2
Library protein (DB)
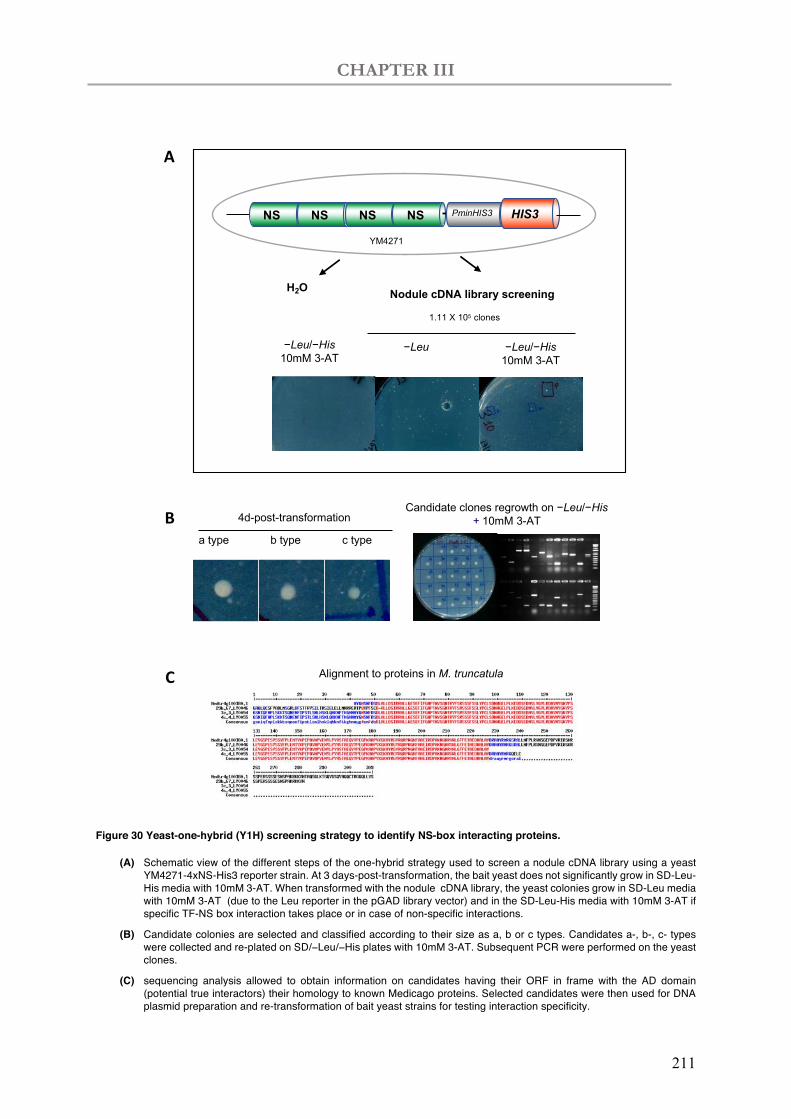
CHAPTER III
211
Figure 30 Yeast-one-hybrid (Y1H) screening strategy to identify NS-box interacting proteins.
(A) Schematic view of the different steps of the one-hybrid strategy used to screen a nodule cDNA library using a yeast YM4271-4xNS-His3 reporter strain. At 3 days-post-transformation, the bait yeast does not significantly grow in SD-Leu-His media with 10mM 3-AT. When transformed with the nodule cDNA library, the yeast colonies grow in SD-Leu media with 10mM 3-AT (due to the Leu reporter in the pGAD library vector) and in the SD-Leu-His media with 10mM 3-AT if specific TF-NS box interaction takes place or in case of non-specific interactions.
(B) Candidate colonies are selected and classified according to their size as a, b or c types. Candidates a-, b-, c- types were collected and re-plated on SD/−Leu/−His plates with 10mM 3-AT. Subsequent PCR were performed on the yeast clones.
(C) sequencing analysis allowed to obtain information on candidates having their ORF in frame with the AD domain (potential true interactors) their homology to known Medicago proteins. Selected candidates were then used for DNA plasmid preparation and re-transformation of bait yeast strains for testing interaction specificity.
A
B
C Alignment to proteins in M. truncatula
NSNSNS NS HIS3PminHIS3
−Leu −Leu/−His10mM 3-AT
H2O Nodule cDNA library screening
1.11 X 105 clones
−Leu/−His10mM 3-AT
YM4271
Candidate clones regrowth on −Leu/−His+ 10mM 3-AT
c typea type b type
4d-post-transformation

CHAPTER III
212
In total, 88 clones were selected as primary candidates (His+ clones) with distinguished colony size above the background. These 88 clones were confirmed to grow on selective conditions after re-dropping on SD/−Leu/−His plates with 10mM 3-AT. Among the 88 PCR-amplified clones that were analysed by sequencing most of them (52) contained either artefactual vector sequences or were not in frame with the AD activation domain. Despite this large proportion of apparent false positives, 29 clones were in frame with the AD domain in the gene fusion, and this included 15 clones encoding proteins with DNA regulatory functions (in grey shade, Table 8). Interestingly, seven of them code for ERF-type transcription factors belonging to the APETALA2/ethylene responsive factor (AP2/ERF) superfamily (Shu et al., 2015; Phukan et al., 2017). These positive clones, which are termed here ERF091 (MtERF1-1, Medtr4g100380.1), ERF092 (Medtr4g100420.1), ERF069 (MtERFB2.3, Medtr2g435590.1) and ERF073 (Medtr4g078710.1), are represented four ERF genes coding for entire (ERF091, ERF092, ERF069 and ERF073) or truncated (ERF092) proteins. However, in all cases the ERF DNA binding domain is present in the encoded proteins (a figure with an alignment in ANNEX IV Figure S 6). The four ERF genes belong to three main groups of ERF TFs whose members have been involved in: (i) plant defence responses or hormone signalling in Arabidopsis or in legumes (group IX, ERF091 and ERF092) (Penmetsa & Cook, 2000; Varma Penmetsa et al., 2008; Anderson et al., 2010)), (ii) to be induced by abiotic stresses conditions (group VIII, MtERF073) (Shu et al., 2015) or (iii) to be involved in the regulation of gene expression in anaerobic conditions, including during nodulation (Rovere, 2018) (group VII, MtERF069/MtERFB2.3). The overrepresentation of members of the ERF group IX (Five out of the seven clones), expected for “true positives in this type of approach, is encouraging and suggest a potential regulatory role of ERFs of this group for driving CP6 expression in nodules.
Additionally, another positive clone codes for a TCP (TEOSINTE BRANCHED / CYCLOIDEA / PROLIFERATING CELL FACTORS) family TF MtTCP20 (Medtr7g028160.1) was identified before by an Y1H screens as a nitrate-mediated regulator of AtNIA1 (Wang et al., 2010). MtTCP20 is homologous to AtTCP20 (At3g27010) of A. thaliana., which is recently reported to interact with the NIN-like protein (NLP) transcription factors AtNLP6&7 to orchestrate plant response to nitrate availability (Guan et al., 2014; Guan et al., 2017). Additionally, the mutant tcp20 showed an accelerated leaf senescence, indicating a repressive role in transcriptional regulation (Danisman et al., 2012). In M. truncatula, 21 putative TCPs were identified and subjected into class I (also known as PCF or TCP-P) and class II (also known as TCP-C, with subclasses: CIN and CYC/TB1) (Wang et al., 2018). But there is not so much functional characterization of TCP proteins in symbiosis. Although it is tempting to
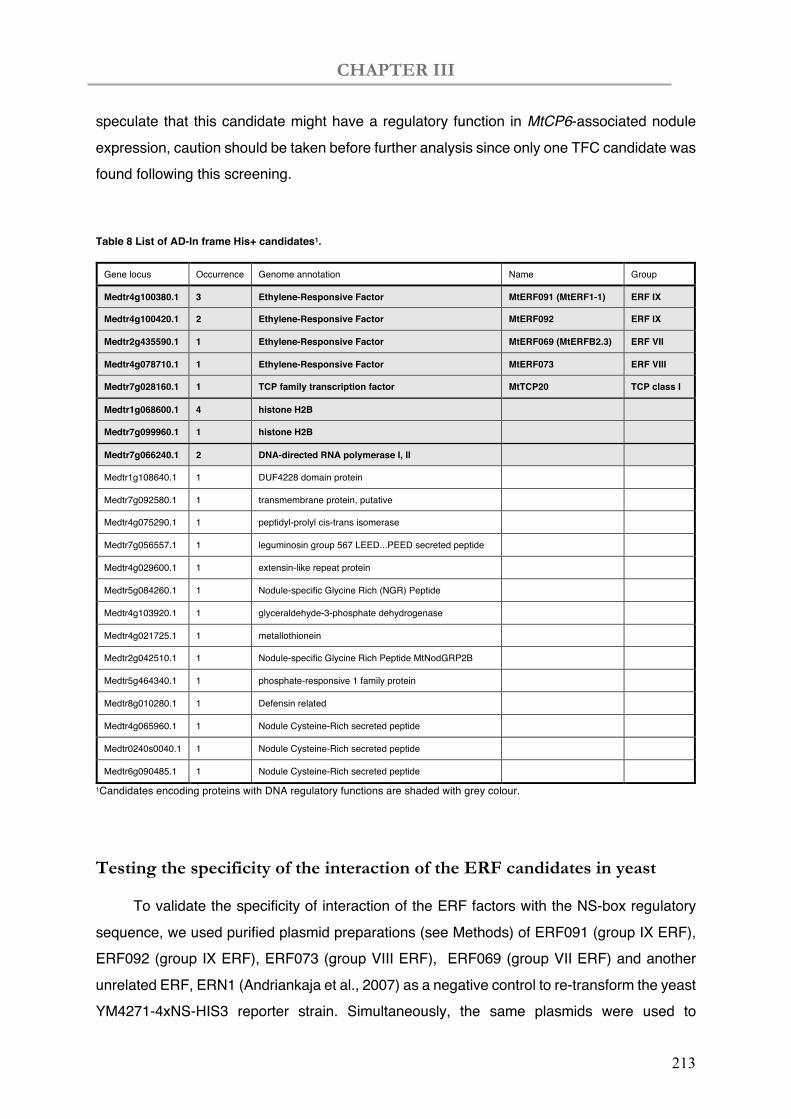
CHAPTER III
213
speculate that this candidate might have a regulatory function in MtCP6-associated nodule expression, caution should be taken before further analysis since only one TFC candidate was found following this screening.
Table 8 List of AD-In frame His+ candidates1.
Gene locus Occurrence Genome annotation Name Group
Medtr4g100380.1 3 Ethylene-Responsive Factor MtERF091 (MtERF1-1) ERF IX
Medtr4g100420.1 2 Ethylene-Responsive Factor MtERF092 ERF IX
Medtr2g435590.1 1 Ethylene-Responsive Factor MtERF069 (MtERFB2.3) ERF VII
Medtr4g078710.1 1 Ethylene-Responsive Factor MtERF073 ERF VIII
Medtr7g028160.1 1 TCP family transcription factor MtTCP20 TCP class I
Medtr1g068600.1 4 histone H2B
Medtr7g099960.1 1 histone H2B
Medtr7g066240.1 2 DNA-directed RNA polymerase I, II
Medtr1g108640.1 1 DUF4228 domain protein
Medtr7g092580.1 1 transmembrane protein, putative
Medtr4g075290.1 1 peptidyl-prolyl cis-trans isomerase
Medtr7g056557.1 1 leguminosin group 567 LEED...PEED secreted peptide
Medtr4g029600.1 1 extensin-like repeat protein
Medtr5g084260.1 1 Nodule-specific Glycine Rich (NGR) Peptide
Medtr4g103920.1 1 glyceraldehyde-3-phosphate dehydrogenase
Medtr4g021725.1 1 metallothionein
Medtr2g042510.1 1 Nodule-specific Glycine Rich Peptide MtNodGRP2B
Medtr5g464340.1 1 phosphate-responsive 1 family protein
Medtr8g010280.1 1 Defensin related
Medtr4g065960.1 1 Nodule Cysteine-Rich secreted peptide
Medtr0240s0040.1 1 Nodule Cysteine-Rich secreted peptide
Medtr6g090485.1 1 Nodule Cysteine-Rich secreted peptide
1Candidates encoding proteins with DNA regulatory functions are shaded with grey colour.
Testing the specificity of the interaction of the ERF candidates in yeast
To validate the specificity of interaction of the ERF factors with the NS-box regulatory sequence, we used purified plasmid preparations (see Methods) of ERF091 (group IX ERF), ERF092 (group IX ERF), ERF073 (group VIII ERF), ERF069 (group VII ERF) and another unrelated ERF, ERN1 (Andriankaja et al., 2007) as a negative control to re-transform the yeast YM4271-4xNS-HIS3 reporter strain. Simultaneously, the same plasmids were used to

CHAPTER III
214
transform the yeast bait strain YM4271-4x NF-box strain as a control. NFbox was the cis-regulatory elements identified preciously to be the binding site for the TF ERN1 (Andriankaja et al., 2007). The specific interaction of the positive control ERN1 to the NF-box results in prominent yeast growth in selective SD-Leu-His+ 3-AT conditions at 2.5-days after transformation (Figure 31). In contrast, no significant yeast growth is observed when the same strain was transformed with the ERF091, -092, -073 or-069 plasmids. This indicate that these other ERF factors are unable to specifically interact with the NF-box. On the other hand, these ERF were able to interact with the NS-box resulting in growth of the YM4271-4X NS-box strain in selective conditions (Figure 31). Although ERN1 can to a certain extent interact non-specifically to the NS-box, this interaction is significantly less strong compared to the group IX ERF factors. In conclusion, these results validate the specific interaction of the ERF candidates (ERF091, -092, -073 and -069) to the NS-box regulatory sequences in yeast.
Figure 31 Evaluating the specificity of the interaction of the ERF candidates to the 4xNS reporter in yeast.
Purified plasmids (pGAD-ERF091, -ERF092, -ERF073, -ERF0069 and ERN1) or water (-) were used to transform YM4271-4xNF-box (containing a tetramer of the Nod factor-responsive cis-regulatory element, Andrianjkaja) or 4xNS-box (this study) yeast reporter strains. Transformed yeast were diluted (OD600nm=0.1 and 0.01) and spot inoculated on SD/L–, SD/H–/L– and SD/H–/L– plates with 15 mM 3-AT. The images were taken 2.5 days after the LiAc-mediated yeast transformation.
YM4271-pHISi-4XNS-box
YM4271-pHISi-4xNF-box
O.D.
0.1
0.01
0.1
0.01
SD L- SD H-L- SD H-L- 15mM 3-AT
- ERF0
91
ERF0
92
ERF0
73
ERF0
69
ERN
1
VIII VII VIX IX
- ERF0
91
ERF0
92
ERF0
73
ERF0
69
ERN
1 VIII VII VIX IX
- ERF0
91
ERF0
92
ERF0
73
ERF0
69
ERN
1
VIII VII VIX IX

CHAPTER III
215
IV. Discussion
ERF regulators as new possible regulators of nodule senescence
We have conducted a yeast-one-hybrid screening using the NS-box tetramer as a bait sequence and identified among 29 positive candidates, 7 clones encoding ERF TFs. These clones code for four ERF TFs belonging to three ERF family groups (IX, VII and VIII). The most represented clones (five out of the seven ERFs) belong to the ERF group IX. Members of this group have been widely reported to participate in defence responses or in hormone signalling in plants. MtERF091 (MtERF1-1) was identified previously in the ethylene-sensitive sickle mutant (Penmetsa & Cook, 2000; Varma Penmetsa et al., 2008). The defence response to R. solani was also proven with an early induction of promoter activity containing the 4×GCC::luciferase construct in transgenic hairy roots of the skl mutant (Anderson et al., 2010). MtSkl encodes the ortholog of the Arabidopsis ethylene signalling protein Ethylene-Insensitive2 (EIN2) (Varma Penmetsa et al., 2008). The skl mutant is presenting a hypernodulation phenotype (Penmetsa & Cook, 2000; Varma Penmetsa et al., 2008). Previous work demonstrated that ectopic overexpression of MtERF091 driven by the CaMV 35S promoter did not interfere with root nodule symbiosis (Figure 32A) (Anderson et al., 2010). However, if MtERF091 plays a major role during later stages of nodule senescence, a phenotype would not be observed with a CaMV 35S promoter which is known to be non-fully functional in conditions of low oxygen pressure, as expected to occur in mature nodules. The homologue of MtERF091 in L. japonicus, LjERF1, has been shown up-regulated in roots upon ethylene and jasmonic acid and activating downstream defence genes (Figure 32B) (Asamizu et al., 2008), which is similar to the Arabidopsis homologue AtERF1 (Lorenzo et al., 2003). Interestingly, LjERF1 was induced as early as 3h after inoculation with M. loti, whereas repressed at a later stage of nodulation (4 dpi) (Asamizu et al., 2008), implying a transient plant defence response upon rhizobia infection followed by genetic suppression in the later nodulation process.

CHAPTER III
216
Figure 32 MtERF091 (MtERF1-1) homologues function in leguminous plants.
(A) MtERF1-1 (MtERF091) does not interfere root symbiosis effectively with ectopic overexpression driven by the CaMV
35S promoter. Composite A17 plants overexpressing MtERF1-1 (MtERF091) and GFP 14 weeks after
transformation and transgenic roots inoculated with S. meliloti. The hash mark indicates non-transgenic root,
asterisks indicate transgenic roots. Adapted from (Anderson et al., 2010).
(B) Effects of ethylene (ET) and/or JA on the expression of LjERF1 in shoots and root. The relative expression of the
indicated LjERF1 in shoots (light gray) and root (dark gray) are shown relative to the nontreated controls (shoots or
root). The shoots and roots of hormone-treated seedlings were collected at 3, 6, and 24 h after treatment. Adapted
from (Asamizu et al., 2008).
However, the expression pattern of these ERF candidates appears not to be analogous
to MtCP6 expression according to Medicago gene expression atlas (Figure S 7). It might be therefore necessary to test if these ERFs (or ERFs from the same group) are upregulated under developmental and stress induced senescence conditions, e.g. via RNAseq analysis of senescing nodules. Nevertheless, TF proteins might still function at low protein/transcript levels. It remains to be clarified the regulation activities of a TF protein and the potential feedback regulation on its own gene expression. Another possibility is that a TF could have distinct target genes in different developmental stages or is dynamically regulated under environmental conditions. For example, the oxygen sensitive TF RAP2.12 (group VII ERF) has been shown to migrate from the plasma membrane to the nucleus upon hypoxia, thereby activating expression of genes adaptative to low-oxygen conditions (Kosmacz et al., 2015). Thus, it would be hard to predict its function according to the expression pattern of a single TF protein, considering multiple transcriptional actors may coordinate together for the transcriptional outputs in situ.
B A

CHAPTER III
217
Our findings suggest a potential role of this ERF factor in the regulation of senescence genes in nodules. Provided with a validated interaction of these ERF candidates with NS-box, our results could support the hypothesis that nodule senescence is a recovery of plant defence responses. Upon nodule senescence, the symbiosis between legume and rhizobia is lost because of perturbed nutrient trading, wherein the beneficial bacteria could be regarded by the host plant similarly to a detrimental pathogen. Consequently, the activation of plant defence related regulators (e.g. MtERF091/MtERF1-1) might mark the initiation of a signal transduction pathway for the loss of the legume-rhizobia mutualism. ABA and ethylene have been suggested to play roles in nodule senescence process based on the transcriptomic profiles of senescing nodules, which demonstrates the up-regulation of ethylene-related genes (Van de Velde et al., 2006). Interestingly, homologs of the ERF factors characterized here were shown to be under the control of ethylene, JA or ABA signalling pathways (Muller & Munne-Bosch, 2015; Shu et al., 2015; Phukan et al., 2017; Shu et al., 2018; Xie et al., 2019). It is therefore plausible that these ERF regulators might represent downstream responses of these hormone regulated pathways to promote nodule senescence.
DNA-binding domain are conserved among the ERF candidates
ERF TFs comprise a DB domain of about 60aa, typically binding to the canonical GCC-box with high affinity in its target gene promoters (Ohme-Takagi & Shinshi, 1995; Fujimoto et al., 2000). All the TF candidates ERF091, ERF092, ERF069 and ERF073 are found to be conserved in this DB domain (Figure 33A), and homologous to the Arabidopsis ERFs, AtERF1A (Lorenzo et al., 2003), AtERF5 (Fujimoto et al., 2000), AtRAP2.3 (Related To APETALA2 PROTEIN3) (León et al., 2020) and AtERF9 (Maruyama et al., 2013), respectively. The phylogenetic analysis of the homologs is shown in Figure 33B. Moreover, the ERF domains are highly conserved in ERF candidates (ERF091, ERF092, ERF069 and ERF073) and their Arabidopsis homologs (Figure 33C). However, these ERF candidates (ERF091, ERF092, ERF069 and ERF073) do not show similar amino acid residues outside the ERF
domain. ERF domain of AtERF1A consists of a three-stranded anti-parallel b-sheet and an a-
helix packed approximately parallel to the b-sheet (Figure 33D) (Allen et al., 1998). This ERF domain binds to the target DNA which bends slightly at the central CG step (Figure 33D) (Allen et al., 1998).
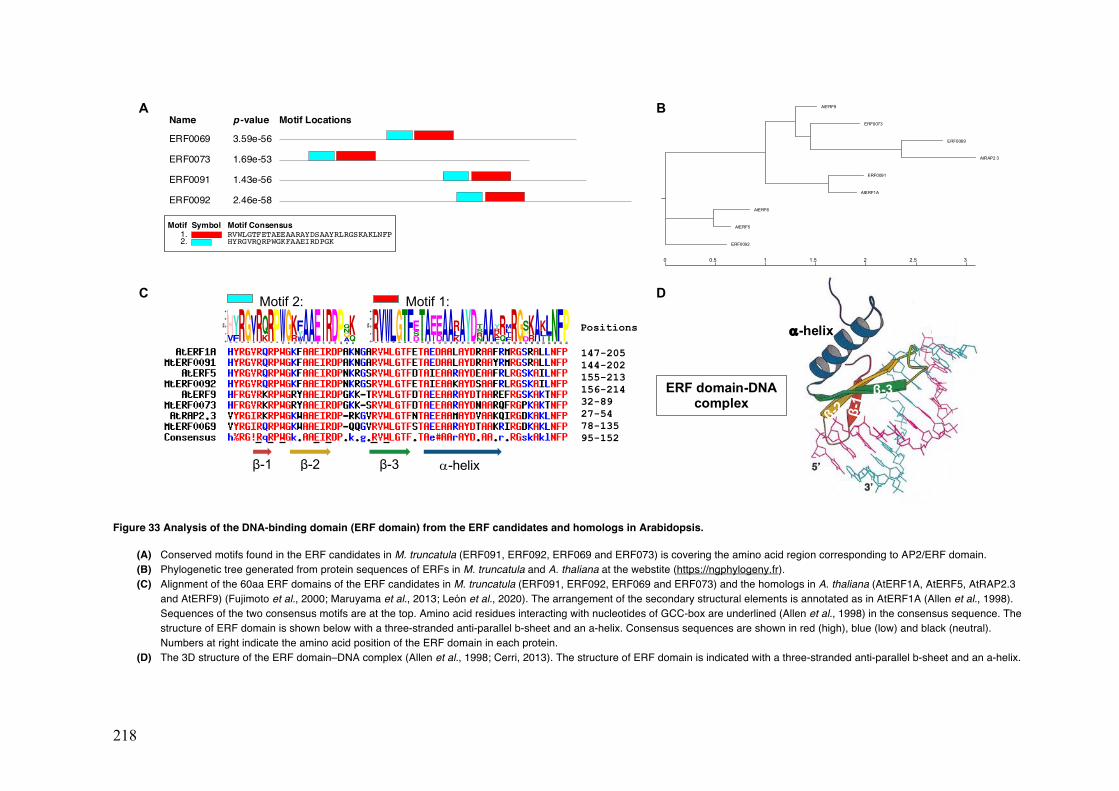
218
Figure 33 Analysis of the DNA-binding domain (ERF domain) from the ERF candidates and homologs in Arabidopsis.
(A) Conserved motifs found in the ERF candidates in M. truncatula (ERF091, ERF092, ERF069 and ERF073) is covering the amino acid region corresponding to AP2/ERF domain. (B) Phylogenetic tree generated from protein sequences of ERFs in M. truncatula and A. thaliana at the webstite (https://ngphylogeny.fr). (C) Alignment of the 60aa ERF domains of the ERF candidates in M. truncatula (ERF091, ERF092, ERF069 and ERF073) and the homologs in A. thaliana (AtERF1A, AtERF5, AtRAP2.3
and AtERF9) (Fujimoto et al., 2000; Maruyama et al., 2013; León et al., 2020). The arrangement of the secondary structural elements is annotated as in AtERF1A (Allen et al., 1998). Sequences of the two consensus motifs are at the top. Amino acid residues interacting with nucleotides of GCC-box are underlined (Allen et al., 1998) in the consensus sequence. The structure of ERF domain is shown below with a three-stranded anti-parallel b-sheet and an a-helix. Consensus sequences are shown in red (high), blue (low) and black (neutral). Numbers at right indicate the amino acid position of the ERF domain in each protein.
(D) The 3D structure of the ERF domain–DNA complex (Allen et al., 1998; Cerri, 2013). The structure of ERF domain is indicated with a three-stranded anti-parallel b-sheet and an a-helix.
A
D
Name p -value Motif Locations
ERF0069 3.59e-56
ERF0073 1.69e-53
ERF0091 1.43e-56
ERF0092 2.46e-58
Motif Symbol Motif Consensus1. RVWLGTFETAEEAARAYDSAAYRLRGSKAKLNFP2. HYRGVRQRPWGKFAAEIRDPGK
C
a-helix
β-3
β-1
β-2 ERF domain-DNA
complex
Motif 2: Motif 1:
MEME (no SSC) 20.07.2020 17:33
0
1
2
3
4
bits
1R 2V 3W 4L 5
G6T 7F 8D
SE
9T 10
A
11
IE
12
DE
13
A
14
A
15
KLR
16
A
17
Y
18
D
19
NRST
20
A
21
A
22
FKRY
23
QR
24
FILM
25
R
26
G
27
DPS
28
RK
29
A
30
ILK
31
TL
32
N
33
F
34
PMEME (no SSC) 20.07.2020 17:34
0
1
2
3
4
bits
1VH
2FY
3R 4
G
5IV
6R 7KQ
8R 9P 10
W11
G12
RK
13
WYF
14
A15
A16
E17
I18
R19
D20
P21
AGNQ
22
QK
β-1 β-2 β-3 a-helix
147-205 144-202 155-213
Positions
156-214 32-89 27-54 78-135 95-152
AtERF9
ERF0073
ERF0069
AtRAP2 3
ERF0091
AtERF1A
AtERF6
AtERF5
ERF0092
0 0.5 1 1.5 2 2.5 3
B

CHAPTER III
219
ERF domain binds to GCC-related cis-regulatory elements
The DNA binding domain of ERF1-like transcription factors to the typical GCC-box motif (Ohme-Takagi & Shinshi, 1990; Ohme-Takagi & Shinshi, 1995). The GCC-box was firstly identified from the 228-bp-long cis-regulatory elements known as the Ethylene-Responsive Element (ERE) (Broglie et al., 1989) from ethylene inducible pathogenesis-related (PR) genes in Nicotiana spp., as the 11-bp sequence (TAAGAGCCGCC) (Ohme-Takagi & Shinshi, 1990; Ohme-Takagi & Shinshi, 1995). AtERF1 has been shown to recognize GCC-box (Franco-Zorrilla et al., 2014), but with only marginal binding to other GCC-like variants (Godoy et al., 2011). A GCC-like sequence (TGCAGGCCT) has been identified from the upstream region of MtENOD11 to mediate gene activation upon Nod factor treatment (Andriankaja et al., 2007) and this sequence is recognized by group V ERF TFs. C-repeat/DRE that contains similar sequences has been found in promoters of COR (cold-regulated) genes in A. thaliana, such as COR15a (Baker et al., 1994), COR78 (Yamaguchi-Shinozaki & Shinozaki, 1994) and C-repeat/DRE Binding Factor 1 (CBF1) (Stockinger et al., 1997). Inside NS-box, a 11-bp GC-rich sequence (TTTGGCCGTT) could be found in addition to other cis-regulatory elements discussed in Chapter I (CAAT-box, W-box and Dof, etc.) (Figure 34A). This sequence region is here termed as GCCcore-like, which is similar to previously identified GCC-like or C-repeat/DREB sequences (Figure 34B) (Ohme-Takagi & Shinshi, 1990; Ohme-Takagi & Shinshi, 1995; Stockinger et al., 1997; Andriankaja et al., 2007).
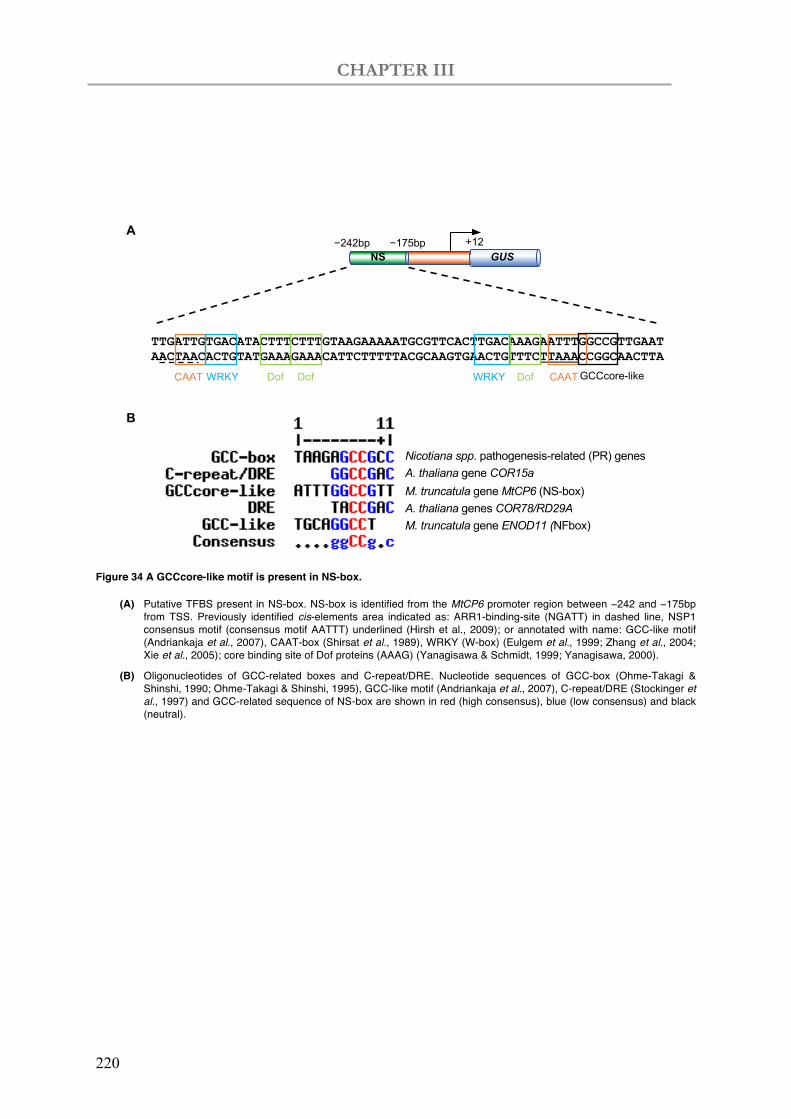
CHAPTER III
220
Figure 34 A GCCcore-like motif is present in NS-box.
(A) Putative TFBS present in NS-box. NS-box is identified from the MtCP6 promoter region between −242 and −175bp from TSS. Previously identified cis-elements area indicated as: ARR1-binding-site (NGATT) in dashed line, NSP1 consensus motif (consensus motif AATTT) underlined (Hirsh et al., 2009); or annotated with name: GCC-like motif (Andriankaja et al., 2007), CAAT-box (Shirsat et al., 1989), WRKY (W-box) (Eulgem et al., 1999; Zhang et al., 2004; Xie et al., 2005); core binding site of Dof proteins (AAAG) (Yanagisawa & Schmidt, 1999; Yanagisawa, 2000).
(B) Oligonucleotides of GCC-related boxes and C-repeat/DRE. Nucleotide sequences of GCC-box (Ohme-Takagi & Shinshi, 1990; Ohme-Takagi & Shinshi, 1995), GCC-like motif (Andriankaja et al., 2007), C-repeat/DRE (Stockinger et al., 1997) and GCC-related sequence of NS-box are shown in red (high consensus), blue (low consensus) and black (neutral).
TTGATTGTGACATACTTTCTTTGTAAGAAAAATGCGTTCACTTGACAAAGAATTTGGCCGTTGAATAACTAACACTGTATGAAAGAAACATTCTTTTTACGCAAGTGAACTGTTTCTTAAACCGGCAACTTA
CAAT WRKY WRKYDof Dof Dof CAATGCCcore-like
NS GUS+12−242bp −175bp
A
B
Nicotiana spp. pathogenesis-related (PR) genes
M. truncatula gene ENOD11 (NFbox)
M. truncatula gene MtCP6 (NS-box)A. thaliana gene COR15a
A. thaliana genes COR78/RD29A

CHAPTER III
221
TCP20 is involved in plant responses to nitrate availability and jasmonic
acid (JA) signalling pathways
It was interesting that one positive clone is identical to TCP20 which might be involved in nodule response to nitrate status and other environmental stresses. However, caution should be taken here by the fact that only one TCP clone was found out of this screening. Nonetheless, it is necessary to keep in mind that the cDNA library we used in the Y1H screening were not specifically designed to study nodule senescence. It is therefore possible that the interactive TFs of NS-box are not yet highly abundant in the cDNA library from nodules of 4- and 19- days. The output of Y1H screenings might be affected by different reasons: 1) protein toxicity in yeast, 2) the TF may be absent from the prey source, 3) the TF might only bind to DNA as a heterodimer (and the current Y1H system expresses only one TF at a time, 4) the TF needs posttranslational modifications (Fuxman Bass et al., 2016), 5) the TF may have an efficient DNA binding domain but not be expressed in the senescence conditions. Thus, complementary approaches are necessary to validate the interactions observed in Y1H. The use of transient expression in heterologous system such as Nicotiana bentamiana and in vitro analysis of protein/DNA interaction could be used to validate the interactions.
In Arabidopsis, TCP20 has been reported to activate nitrate assimilatory genes via interacting with NIN-Like Proteins (NLPs) (Guan et al., 2014; Guan et al., 2017). TCP20 is also negatively involved in jasmonic acid (JA) biosynthesis and in this context it repress its target gene LIPOXYGENASE2 (AtLOX2) (Danisman et al., 2012). JA has been involved in plant responses to environmental stresses and leaf senescence (Creelman & Mullet, 1997; He et al., 2002). The role of JA in nodule senescence has been suggested based on transcriptomes of senescing nodules, wherein gene targets of JA were shown to be upregulated (Van de Velde et al., 2006). The stress-related role of JA was tested on nodules of common bean, particularly in the ascorbate-glutathione pathway, which contributes to antioxidative protection in nodules (Dalton et al., 1986; Becana et al., 2010). The authors found that JA treatment had some major effects on nodules, with an accumulation of hGSH, inhibition
of g-glutamylcysteine synthetase (gECS) activity, transcriptional induction of ascorbate oxidase (AO) and posttranslational suppression in dehydroascorbate reductase (DR) activity (Loscos et al., 2008). TCP20 is revealed to response nitrate and interact with NLPs (Guan et al., 2014; Guan et al., 2017). Nevertheless, TCP20 is not expressed in an organ-specific manner neither in A. thaliana nor in M. truncatula. It is thus hardly predictable that TCP20 could interact alone with the NS-box, which directs tissue-specific expression in root nodules.

CHAPTER III
222
Other His+ clones not encoding typical TFs with recognized DNA binding
domains
With regard to other positive clones in addition to TF candidates, two histone H2B, Medtr1g068600.1 and Medtr7g099960, are found 4 and 1 times, respectively (Table 8). H2B is one of the fundamental histone proteins (H3, H4, H2A, H2B and H1) that wrap the genomic DNA to form nucleosomes of chromatin in eukaryotic cells. Histone modifications on amino-termini, including methylation, acetylation, phosphorylation and ubiquitination are thought to carry epigenetic information regulating chromatin dynamics and gene expressions (Strahl & Allis, 2000; Jenuwein & Allis, 2001; Zhang, 2003). However, histones are widely distributed in eukaryotic cells, it is not very well-understood if histone modifications are related to genomic DNA sequence specificity.
Apart from that, the output of Y1H screen also resulted in three NCR peptides (Table 8). NCRs are defensin-like peptides possessing antimicrobial activity and therefore are classified as AMPs (Van de Velde et al., 2010; Maróti et al., 2015). NCRs are abundant and with more than 500 members in M. truncatula (Maróti et al., 2015), which are essential for bacteroid terminal differentiation (Mergaert et al., 2003; Mergaert et al., 2006; Van de Velde et al., 2010; Farkas et al., 2014). During terminal differentiation of bacteria, NCRs were shown to suppress bacterial reproduction and increase bacterial membrane permeability, also indicating their positive roles during nodule senescence, when the fragilized bacteroid membranes can be efficiently degraded by the plant cell proteases (Vasse et al., 1990; Van de Velde et al., 2006; Van de Velde et al., 2010). The NCR MtNFS2 was shown to functions as a negative regulator of symbiotic persistence, suggesting a host control on the success of the symbiotic partnership (Wang et al., 2017). However, there are currently no evidence of NCRs in transcriptional regulation or relationship to CP gene expressions.

CHAPTER III
223
V. Summary of potential transcriptional actors for MtCP6 expression
Overall, the 67-bp-long cis-regulatory elements (NS) is sufficient to drive transcriptional gene activation during developmental nodule senescence and in response to nitrate. A summary of the potential regulation for MtCP6 gene expression is in Figure 35. In the Y1H screening, the mostly represented NS-interacting TFs belong to ERFs involved in plant defence responses. ERF091 (group IX) has been found to be pathogen related and involved in ethylene/jasmonate signalling pathway in M. truncatula (Anderson et al., 2010). Gene expression of ERF073 (group VIII) appears to be sensitive to many environmental stimuli (Shu et al., 2015). Arabidopsis ERF genes in groups VIII and IX have often been implicated with plant response to disease and stresses in ethylene and/or JA signalling pathways (Nakano et al., 2006). The group VII ERF members, including ERF069, might be regulators under hypoxia/anaerobic conditions via perception of gaseous signals (nitric oxide: NO) (Gibbs et al., 2014; Rovere, 2018). NO was shown to be produced at interzone III–IV by using a NO-biosensor S. meliloti strain, thus proposed to be a signal in developmental and induced nodule senescence. (Cam et al., 2012). In M. truncatula nodules, NO3– (nitrate) is reduced to NO2–, which is subsequently reduced to NO via the mitochondrial and bacteroid electron-transport chains (Horchani et al., 2011; Hichri et al., 2015). In presence of NO, the group VII ERFs were shown to be degraded in a N-end rule pathway of targeted proteolysis (Gibbs et al., 2014). ERF069 is thus more likely to play a role under the anaerobic conditions inside nodules. Another potential nitrate response candidate is TCP20, which has been proven to interact with NLPs in nitrate response of Arabidopsis (Guan et al., 2014; Guan et al., 2017).
Finally, no transcription factors putatively binding to the WRKY/CAAT box identified in Chapter II was isolated from Y1H. Further experiments might be needed to understand the roles of these DNA regulatory sequences in gene regulation. In Chapter II, we have shown a dramatic decline of GUS activity driven by the tetramer of NS-box in synthetic promoter and the minimal functional MtCP6 promoter down to −242bp. A potential transcriptional activator is thus presumably present in the minimal MtCP6 promoter besides NS-box.
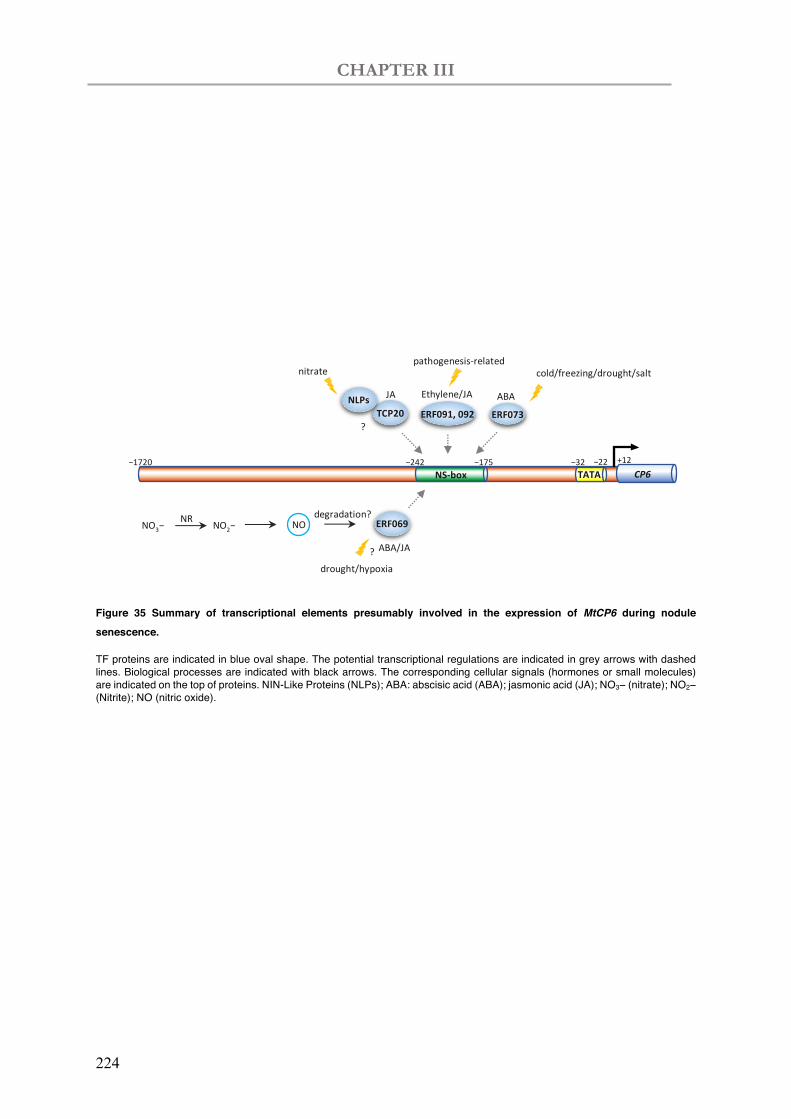
CHAPTER III
224
Figure 35 Summary of transcriptional elements presumably involved in the expression of MtCP6 during nodule senescence.
TF proteins are indicated in blue oval shape. The potential transcriptional regulations are indicated in grey arrows with dashed lines. Biological processes are indicated with black arrows. The corresponding cellular signals (hormones or small molecules) are indicated on the top of proteins. NIN-Like Proteins (NLPs); ABA: abscisic acid (ABA); jasmonic acid (JA); NO3− (nitrate); NO2− (Nitrite); NO (nitric oxide).
NS-box CP6
ERF069
JA TCP20
NLPs
nitrate
ERF091, 092 Ethylene/JA
pathogenesis-related
?
+12 −1720 −242 −175
ERF073 ABA
cold/freezing/drought/salt
drought/hypoxia ?
NO NO2− NO3− ABA/JA
degradation? NR
TATA −32 −22

CHAPTER III
225
VI. Perspectives
Based on the NS-box from Chapter II, we searched the interacting TFs via Y1H. However, this preliminary result requires further validation or potentially characterization to prove roles of the candidates in nodule senescence (Figure 36). Firstly, to validate the interaction of TF candidate and the NS-box, another spot assay experiment in yeast is necessary to be done with the yeast strains transformed with corresponding plasmids. In this context, a higher concentration (up to 45mM according to the Clontech protocol) of 3-AT may be considered to validate the interaction of TF candidates and NS-box. Moreover, considering that the TFs are fused to the GAL4-AD (activation domain) which activates in situ the PminHIS3 for HIS3 expression, the transactivation in yeast hybrid may only provide evidence for the function of the TF BDs (DNA-binding domains). We therefore require further characterization to determine if the TF candidates act as activators or repressors. Although the ERF factors characterized in this study are supposed to act as activators, we need anyhow to show it experimentally. For that we propose to use a N. benthamiana (Andriankaja et al., 2007) transactivation system that allow us to examine the regulatory activity of a TF towards NS-containing promoter-GUS gene fusions.
In this system co-infiltration of A. tumefaciens strains containing respective TF constructs (under the 35S promoter) and the 4xNS-GUS constructs will allow us to evaluate by histochemical and fluorimetric (quantitative assays) the relative transcription activity of these transcription factors. In preliminary experiments with L. Frances (LIPM, Toulouse), we demonstrated the low basal of a 4x NS-box-GUS fusion in this system and we now aim to use it in the presence of the ERF TFs and other important controls. This includes the use of mutated NS versions at the GCC-rich motif, mutated versions of the ERFs without the DNA-binding domain and other unrelated ERFs (such as ERN1) to test specificity in the transcription assays.. If they act as activators, we can also subsequently evaluate if their overexpression is sufficient to activate the transcription of their target (CP6) in M. truncatula in non-inoculated plants, as previously done for other symbiotic regulators (Laloum et al., 2014; Singh et al., 2014; Cerri et al., 2017). Complementary analyses of GFP-protein fusions will provide information on the subcellular localization of these factors.
If the candidates are validated by yeast drop test and transactivation in N. benthamiana, it will be important to validate the TF-DNA interaction by complementary approaches such as DNA pull-down (Wu, 2006; Kliszczak et al., 2011), EMSA (electrophoretic mobility shift assay) (Hellman & Fried, 2007) or in vitro translation DNA-binding assay (Nomoto et al., 2019). To identify other targets of TF proteins in the genome, it might also be performed with an and

CHAPTER III
226
CHIP-PCR (Ren et al., 2000; Nelson et al., 2006) to determine a genome-wide binding positions in vivo. The CHIP-PCR strategy has been successfully used both in N. benthamiana and M. truncatula to study the DNA-protein binding activity of other interaction as previously done for the ERN ERF regulators (Andriankaja et al., 2007; Cerri et al., 2017).
Figure 36 Flow chart of perspectives to characterize TF candidates.
If ERF and TCP candidates are validated to bind to the NS-box, it might be interesting
to carry out functional studies on its mutants via reverse genetic approaches. From the Nobel Medicago mutant bank, several mutants are available for MtCERF092 (e.g. Solexa-NF5874-6), MtERF069 (e.g. NF9447_high_9, NF7237-high-3, and NF19360_high_33) and MtERF073 (e.g. NF1441_high_58, NF5110_high_25). However, functional redundancy can be an issue here, we can then envisage to use alternative strategies to knockout (CRISPR-Cas) or knockdown (RNAi) closely related ERF members and complementary ectopic expression of TF encoding genes to understand their roles in the regulation of CPs expression. With regards to TCP20, we can envisage testing its potential roles in developmental nodule senescence and in nitrate signalling. The mutant line (NF0759_high_45) is available from the Nobel Medicago mutant bank. From the gene expression atlas, an up-regulation of MtTCP20 can be seen in 16-day-old nodules treated with nitrate (10mM, 2 day) (Benedito et al., 2008).
To search effective phenotypes of TFs in terms of nodule senescence, an example can be found in Chapter III, wherein a premature nodule senescence phenotype has been found to be induced by ineffective bacterial partner as a glutathione deficient S. meliloti mutant
Transactivation • Drop test in yeast
• Transient expression in N. benthamiana.
(TF nuclear localization; co-infiltration with 4NS::GUS)
Protein-DNA binding
Phenotyping
• DNA pull down, EMSA, CHIP, etc.
• Mutation in GCC-related-sequence of NS-box
• Mutation in DNA-binding domain of TFs
• Phenotyping of TF candidates
• Promoter activity (4NS::Pmin35S) in TF mutants

CHAPTER III
227
(SmgshB). Under this perspective, it might be better to carry out a preliminary experiment to observe nodule phenotypes (nodule number, size, colour, etc.) in comparison to the wild-type nodules (WT). Given by effective nodule phenotypes caused by genetic perturbation, some additional conventional studies on nodule symbiosis might be carried out: 1) Acetylene Reduction Assay (ARA) to quantify the nitrogen fixation capacity; 2) RT-qPCR to investigate expression of nodule function marker genes or related transcriptional regulating genes (e.g. CPs); 3) histological analysis such as Toluidine blue- or live/dead- staining; 4) spatial localization of transcripts and proteins; 5) plant phenotyping provided with homozygous mutants; etc. Finally, it might be interesting to evaluate the 4NS-box-Pmin35S synthetic promoter activities with provided TF mutants. Possibly, it could be interesting to investigate promoter activity (using the luciferase reporter) in M. truncatula protoplasts for various purposes (e.g. nitrate response, transactivation with TF candidates, hormone responses).

228

229
Men say they know many things;
But lo! they have taken wings, —
The arts and sciences,
And a thousand appliances;
The wind that blows
Is all that anybody knows.
Henry David Thoreau

230

231
GENERAL DISCUSSION & PERSPECTIVES

232

GENERAL DISCUSSION & PERSPECTIVES
233
I. Disruption of symbiotic partnership during root nodule senescence
Mutualistic processes of plant and rhizobial partners determinate the fate of
symbiosis
Symbiotic nodules are the consequence of the reciprocal nutrient exchange between legume plants and rhizobia. The success or failure of a symbiotic relationship is under the control of both partners. In this thesis, the involvement of the plant host genetic regulation and the rhizobial performance was investigated in the nodule senescence process.
On the plant side, a cysteine protease encoding gene MtCP6 was firstly identified from a group of CP genes up-regulated during nodule senescence (Van de Velde et al., 2006; Perez Guerra et al., 2010). MtCP6 has been proven to modulate nodule senescence positively and negatively via ectopic overexpression and RNA interference approaches, respectively (Pierre et al., 2014). Expression of MtCP6 in nodules is specifically at interzone III-IV and sensitively response to environmental stresses such as dark- and nitrate- treatments (Perez Guerra et al., 2010; Pierre et al., 2014). In this study, a 67bp-long nodule-senescence related cis-regulatory element (NS-box) is therein identified from the 1.7kb-long 5’ flanking sequence of MtCP6. A tetramer of the NS-box was able to drive expression pattern at the onset of nodule senescence and was also responsive to nitrate, which is reminiscent of the native promoter activity of MtCP6. Using the NS-box, numerous transcription factors have been trapped from nodule cDNA library and were found to be identical to plant defence related genes. Our study has uncovered several actors of the root nodule senescence regulation mechanism. We have also provided a vision to decipher a complex 3D transcriptional network, which could be triggered by multiple extrinsic and intrinsic stimulus while terminating at activating the executors of the coordinated bacteroid and plant cell death.
In addition to the plant host, the bacterial partner is also involved in the participatory decision-making process in the departure of symbiotic friendship. Inside nodules, rhizobia reside in plant cells to fix dinitrogen necessitating an anaerobic condition mostly due to the extreme oxygen-sensitivity of nitrogenases. We have shown that a S. meliloti 2011 mutant SmgshB, in which the glutathione synthetase encoding gene gshB is disrupted, induced a premature nodule senescence phenotype. In association with such a rhizobium strain deficient with the antioxidant GSH, a series of genetic and phenotypic changes have occurred in

GENERAL DISCUSSION & PERSPECTIVES
234
nodules probably caused by accumulated oxidative stresses inside bacteroids. The ascorbate-GSH or “Foyer-Halliwell-Asada” pathway is recognized as a key player in H2O2 metabolism (Noctor & Foyer, 1998; Matamoros et al., 2003; Foyer & Noctor, 2011). H2O2 accumulation, together with CP gene expressions, have been observed in all of the central infected tissue at the onset of the nodule senescence process (Alesandrini et al., 2003). Notably, expression of MtCP6 was the most elevated gene in RT-qPCR gene expression analysis of WT S. meliloti 2011 and SmgshB inoculated nodules. This result indicated an early and sensitive transcriptional response to induce nodule senescence in case of the ineffective bacteria with impaired nitrogen fixation capacity.
In the future, analysis of plant and bacterial actors regulating the developmental and stress senescence will be an interesting scientific topic to develop. The expression of NCR peptides has been already shown to play a major regulatory role in the interaction between the two partners (Mergaert et al., 2003; Alunni et al., 2007; Van de Velde et al., 2010; Haag et al., 2011; Farkas et al., 2014). However, the NCR peptides are not present in legumes producing determined nodules suggesting that these peptides are only a fraction of the plant proteins regulating the interaction with the bacteria. In developmental senescence, microscopy analysis suggests that the symbiosome is firstly degraded in the symbiotic plant cells. We could predict that this degraded organelle could produce molecular signals which could induce senescence of the plant cell. However, this scenario is maybe not identical in stress induced senescence in which a faster degradation of bacteroid content undergoes in the absence of cellular signs for nutrient remobilization. In this context, the identification of communicating signals from the host and the bacteria involved in the senescence processes represent a major interest to decipher the similar but distinct nodule senescence processes triggered by intrinsic cellular signals or by environmental perturbations.
Is nodule senescence a delayed plant defence response?
Upon symbiotic recognition, symbiotic microorganisms like rhizobia can accommodate into plant cells with the host-specific Nod factors binding to the plant kinase-like receptors. Nod factors are actually derivatives of chitin oligosaccharides which are potent inducers of plant immune responses, but the lipid modification on its chitin-oligosaccharide backbone may enable a compatible recognition by host plants (Zipfel & Oldroyd, 2017). Nod factor-dependent nodulation is not the only strategy to enter symbiosis. Some Bradyrhizobium strains were found to be able to elicit root and stem nodules even lacking nod genes (Giraud et al., 2007; Masson-Boivin et al., 2009; Okazaki et al., 2013). Therein, rhizobia type III (T3SS), or type IV

GENERAL DISCUSSION & PERSPECTIVES
235
(T4SS) secretion systems are required for nodule organogenesis (Deakin & Broughton, 2009; Okazaki et al., 2013). Evidence is emerging that symbiotic colonization needs a suppression in plant immunity, for reviews (Gourion et al., 2015; Tóth & Stacey, 2015). Transcriptomic analysis has also shed light on the suppression of plant defence genes during nodulation (Van de Velde et al., 2006). The DNF2 and SymCRK plant genes, encoding a phosphoinositide phospholipase C (PI-PLC)-like protein and a cysteine-rich, non RD receptor-like kinase, respectively, can suppress defence reactions by an unknown mechanism (Oldroyd, 2013).
It was proposed in 2005 that nodule senescence may be a delayed response on the part of the plant to rhizobium as a potential pathogen (Puppo et al., 2005). Upon rhizobial infection, a plant defence response is transiently activated with an oxidative burst, similar to that observed in plant pathogen response (Santos et al., 2001; Puppo et al., 2005). While accepting infection by the bacterial partner, the plant keeps reactive oxygen species (ROS) low, which is indeed necessary for the proper functioning of the rhizobial nitrogenase enzyme complex (Puppo et al., 2005; Muglia et al., 2008). Finally, during nodule senescence, a decrease in N fixation levels and an increase in ROS occur (Puppo et al., 2005; Muglia et al., 2008). The transcriptomics of nodule senescence (Van de Velde et al., 2006) revealed a wide variety of activation in plant defence (e.g., glutathione transferase) and stress responses (e.g. wound-inducible P450 hydroxylase). We have seen that CP genes are elevated during nodule and multiple plant defence relate proteins were trapped in the Y1H screening using the nodule senescence related element we identified. Indeed, papain-like cysteine proteases (PLCPs) have been considered as central hubs in plant immunity, which are targeted by secreted pathogen effectors to suppress immune responses (Misas‐Villamil et al., 2016). Arabidopsis null mutants for the PLCP RD21 are significantly more susceptible for the necrotrophic fungal pathogen Botrytis cinereal (Shindo et al., 2012). The study in this thesis has been dedicated to deciphering the transcriptional regulation signalling upstream in activation of the PLCP-like enzymes (CPs) in symbiotic organ. In comparison, the investigations on plant immunity in symbiosis and pathogenesis, including the roles of CPs, might shed light on the distinct evolution of host-rhizobia or host-pathogen relationships. In the future, it might be interesting to compare symbiosis and pathogenesis in parallel to uncover the similar but distinct signalling pathways.

GENERAL DISCUSSION & PERSPECTIVES
236
II. Deciphering nodule senescence mechanism at transcriptional level
Transcriptional actors are involved in determination of nodule success
To date, many researches have contributed to decipher the transcriptional cascades during early stages of nodulation. Numerous identified TFs have been revealed to be vital in symbiosis establishment between legume and rhizobia, e.g. IPD3 (known as CYCLOPS in L. japonicus) (Messinese et al., 2007; Yano et al., 2008; Singh et al., 2014), a gibberellin (GA) induced repressor DELLA (Fonouni-Farde et al., 2016a; Fonouni-Farde et al., 2016b; Jin et al., 2016; Fonouni-Farde et al., 2017), the GRAS protein NSP1,2 (Kalo et al., 2005; Smit et al., 2005 ; Hirsch et al., 2009; Hirsch & Oldroyd, 2009; Jin et al., 2016 ; Hartmann, 2018), ERNs (ERF group V) (Andriankaja et al., 2007; Cerri et al., 2012; Cerri et al., 2017), NIN (Schauser et al., 1999; Marsh et al., 2007; Vernié et al., 2015), cytokinin Response Regulators RRs (Gonzalez-Rizzo et al., 2006), and the CCAAT-binding TF NF-Ys (Soyano et al., 2013; Laloum et al., 2014; Laporte et al., 2014; Baudin et al., 2015; Zanetti et al., 2017). Some of the interacting cis-regulatory DNA elements have therefrom been uncovered, e.g. CYC-box (TGCCATGTGGCA) (Singh et al., 2014), NSP1 (cis-element AATTT) (Hirsch et al., 2009), NFbox (Andriankaja et al., 2007), NIN (AAGMT; a cytokinin-responsive cis-regulatory region) (Marsh et al., 2007 ; Vernié et al., 2015; Liu et al., 2019), cytokinin sensor TCS (or TCSn) (Muller & Sheen, 2008; Zurcher et al., 2013; Liu & Müller, 2017). Several TFs have been related to maintain nodule function, e.g. EFD (Ethylene response factor required For nodule Differentiation) (ERF group V) (Vernié et al., 2008), ZPT2-1(Frugier et al., 2000), Regulator of Symbiosome Differentiation (RSD) (Sinharoy et al., 2013).
However, it is not well understood about the transcriptional regulation of nodule senescence process. The TF MtATB2 (bZIP family), which was identified from transcriptomic profile of senescent nodules (Van de Velde et al., 2006), is shown later by the authors under the regulation of sucrose (D'Haeseleer et al., 2010). Cysteine proteinase gene CP77 (GenBank: QFO38998.1; in M. truncatula A17: Medtr4g065077.1) was identified as one of the highly upregulated gene of the transcriptional repressor bHLH2 (Medtr5g030430.1) from the transcriptome of bhlh mutants and was related to nodule senescence (Deng et al., 2019). bHLH2 was shown to be expressed antagonistically to the nodule senescence processing, while CP77 is dramatically upregulated at 28dpi in the bhlh2 mutants. The transcriptional binding of bHLH2 to G-box on the promoter of CP77 was confirmed by Electrophoretic Mobility Shift Assay (EMSA) and chromatin immunoprecipitation (Korkuc et al., 2014) assay, thereof

GENERAL DISCUSSION & PERSPECTIVES
237
suggesting that bHLH2 is the transcriptional repressor of CP77 (Deng et al., 2019). However, the authors suggested bHLH2 as a repressor deduced from an enhancing transcriptional activity of CP77 in bhlh2 mutants via RT-qPCR and GUS staining. It might be interesting to exam if there is a down-regulation of CP77 expression by overexpressing bHLH2. Other than that, expression of CP77 (Medtr4g065077.1) is upregulated in zone IZ and zone III according to the RNA-seq data of laser-dissected nodules (Roux et al., 2014), and prominently expressed in seeds according to the M. truncatula Gene Expression Atlas (Benedito et al., 2008). The expression pattern of CP77 is not reminiscent of that in the MtCPs (MtCP1 to MtCP6) identified from nodule senescence transcriptome (Van de Velde et al., 2006; Perez Guerra et al., 2010). In addition, numerous TFs were characterized to mediate nodule senescence under various environmental changes. NAC969 was evidenced to be up-regulated upon abiotic stresses with nitrate, but it was negatively related to nodule developmental senescence (de Zelicourt et al., 2012). The divergent transcriptional activities of NAC969 imply a distinct regulation pathway of developmental nodule senescence and nodule response to abiotic stresses. NIN-Like Protein 1 (NLP1) appears to compete with the nodulation gene NIN to bind the nitrate responsive cis-regulatory element NRE, and thus modulates nodulation or nitrate-triggered nodulation inhibition (Lin et al., 2018). Notably, NLPs in Arabidopsis are well known to bind this nitrate responsive cis-regulatory elements NRE which is conserved in NIR promoters (Konishi & Yanagisawa, 2013). ERFB2.1/B2.11, which are in the same group (ERF VII) of our candidate ERF069, were related to O2 sensing with a potential role in maintaining anaerobic condition in nodules, whereas silencing these two genes did not affect MtCP6 expression (Rovere, 2018).
These data and our results suggest that multiple transcription factors and thus cis-regulatory elements are involved in the senescence process which includes a degradation process of the plant and the bacterial cells and a recovery process of nutrients by the plant host. Moreover, there is also a saprophytic zone where bacteria are still present in the nodule. Therefore, it is likely that modulators of defence response in plant immunity, cell death and recovery of nutrients are likely to be found in nodule senescence regulation. Overall, it might be thus explained that some TFs such as NAC969 exhibited divergent regulation patterns in case of developmental and induced nodule senescence. On the contrary, the NS-box cis-regulatory element, which we identified from the proximal 5’ flanking sequence of MtCP6, is possessing transcriptional information positively responsive to both developmental and induced nodule senescence. These results indicate that nodule senescence is likely to culminate with proteolytic activities initiated by the co-elevation of cysteine protease genes.

GENERAL DISCUSSION & PERSPECTIVES
238
Hence, transcriptional regulation of nodule senescence, e.g. NS-box mediated expressions, is in the downstream of both developmental and induced nodule senescence pathways. We assume that multiple transcriptional pathways may orchestrate the activation of senescence proteases and eventually trigger the nodule senescence terminating the final death of the symbiotic organ.
In this context, the identification of other TFs involved in different steps of the developmental or induced nodule senescence is an interesting scientific topic. The analysis of the transcriptional regulation of MtCP6 has allowed to show that the MtCP6 promoter includes multiple regulatory elements which modulate the transcription of the gene. In this context, identification of the transcription factors which regulates positively or negatively MtCP6 is crucial to initiate the definition of the transcriptional network regulating the nodule senescence process. A better definition of the gene targets of the already identified TFs is also necessary to implement our knowledge of the senescence transcription network. Nevertheless, the transcriptional efficiency might be also conferred via other transcriptional factors such as DNA methylation and chromatin conformation. Analysis of TF interacting factors via tap-tag experiments may allow to identify these factors. Other approaches have been developed to capture interacting proteins with a priori knowledge of the target loci. Using a bait nucleotide probe, its associated proteins could be isolated and identified via a protocol called Proteomics of Isolated CHromatin segments (PICh) (Dejardin & Kingston, 2009). More recently, an in situ approach has been developed to capture the locus-specific chromatin events with biotinylated nuclease-deficient Cas9 and sequence-specific guide RNAs (Liu et al., 2017).
Nevertheless, we have already identified the NS-box and multiple TFs that may be involved in the nodule senescence regulation. Concerning the NS-box, we did not find TF that putatively bind to the CW motif. As these sequences seem to play an important role in the NS-box activity, it will be important to test whether the TF candidates bind to DNA using this sequence or if other candidates have been missed. To answer to this question, the screening of a senescence specific cDNA library by Y1H could be performed to increase the number of putative senescence related TFs. We could also construct two different cDNA library deriving from RNAs extracted from developmentally senescent nodules or stress induced senescent nodules. However, a short-term perspective might still depend on the validation of the transcription factor candidates from the Y1H experiments. Multiple approaches could be used validate the target genes. As an example, ChIP is a powerful tool to search for the target DNA locus distributed in chromatin (Nelson et al., 2006). Identification of transcriptional regulators

GENERAL DISCUSSION & PERSPECTIVES
239
Forward and reverse approaches can be used to decipher a transcription
network
In our study, we intended to decipher to some extent the transcriptional regulation of nodule senescence in a DNA-centered strategy. Generally, gene transcriptional networks can be explored following two approaches: (1) given a TF, one could identify the target genes subjected to the transcriptional control with binding activity (forward approach), or (2) given a gene of interest (GOI), one could search for the upstream TFs (reverse approach). In both conditions, it would be more efficient to initiate from a central player (protein/gene) with evidenced biological functions.
The TF-centered strategy includes a priori characterization of a TF and the identification of the molecular targets. Firstly, functional characterizations are required with phenotyping via mutagenesis, subcellular localization, in addition to interaction with other proteins. An evidenced functional TF could be thereby used to identify its target genes and the specific binding sites. However, many years of research may be spent to screen a functional TF considering genomic abundance, functional compensation and technical limitations. The following identification of target genes can also be challenging. In this context, chromatin immunoprecipitation (Korkuc et al., 2014) is a powerful assay which provided crucial insights into the protein-DNA interactions distributed in chromatin (Nelson et al., 2006). Sometimes, the targets have been screened/selected from genes with perturbated expression based on the transcriptomic profile of a TF mutant, following by transactivation validations. Nevertheless, the screening approach is easy to be flooded with too many candidates, for the reason that hundreds of genes can be modified in the mutant. Furthermore, the transcriptional binding of certain TF family is conserved to specific sequences with marginal variations, which is abundant in non-coding genomic sequences. Thus, a transactivation analysis in vivo is more likely to reflect a biological context spatially and temporally than in vitro assays.
In contrast, promoter of the GOI can be used for fishing the interactive proteins, as we have conducted. In this case, it is often necessary to conduct the classical promoter deletion analysis, which is often labour-intensive particular in planta/vivo. Moreover, the cis-regulatory elements of GOI are not necessarily to be binding sites of certain TFs. The function might be also conferred via other transcriptional factors such as DNA methylation and chromatin conformation. In this context, it would be complicated to fish the interactive TFs, as well as the subsequent validation in terms of transcriptional binding and biological function of the TF candidates. In addition to Y1H (Li & Herskowitz, 1993), numerous approaches have been developed to capture interacting proteins with a priori knowledge of the target loci. Using a

GENERAL DISCUSSION & PERSPECTIVES
240
bait nucleotide probe, its associated proteins could be isolated and identified via a protocol called Proteomics of Isolated CHromatin segments (PICh) (Dejardin & Kingston, 2009). More recently, an in situ approach has been developed to capture the locus-specific chromatin events with biotinylated nuclease-deficient Cas9 and sequence-specific guide RNAs (Liu et al., 2017).
To date, many bioinformatician are also dedicated to decoding the genome sequences via continuously updating algorithms. Even though to master the comprehensive transcription networks remains to be a distant aspiration and ambitious hope, it has been developed to refine bioinformatic prediction of the regulatory sequences based on flooded data from genome sequencing and large-scale expression data (Wasserman & Sandelin, 2004). Even so, current bioinformatic predication of conserved sequences alone is of limited usage which could hardly represent the highly dynamic chromatin status in situ. Nonetheless, it still provides valuable guides and previous evidences in prior to experimental validations. In return, laboratory data can be used to refine the prediction models, which are generally generated from promoter deletion mapping, and mutagenesis of regulatory sequences (Wasserman & Sandelin, 2004). Overall, despite of the current protein-coding system, we still lack sufficient knowledge to understand how the regulatory information is encoded in genomic DNA sequences. A systematic combination of multiple transcriptional analyses from different perspectives would contribute to the decoding of regulatory sequences.
III. The future of symbiotic nitrogen fixation
Is it beneficial to delay nodule senescence in agricultural production?
For the sake of sustainable agriculture and global equity, an increasing attention has been paid on the substitution of industrial fertilizer with biological N fixers. In the nitrogen-fixing symbiotic nodules, plant and bacterial partners associate together with a nutrient trading agreement, therein nodules are kept in an appropriate number, size and nitrogen-fixing efficiency. A prominent feature of nodule senescence is the declining symbiotic nitrogen-fixation capacity, which shuts down the nutrient exchange. A strict genetic control is therein involved to determine the fate of a nitrogen-fixing symbiosis. Several genes have been related nodule senescence, mostly due to the early senescence phenotype observed in its mutant, such as LjIPT3, MtNAC969, MtCP6 and MtVPE in plants (de Zelicourt et al., 2012; Chen et al., 2014; Pierre et al., 2014), as well as nnrS, norB and and vapBC in bacteria (Meilhoc et al., 2013; Lipuma et al., 2014; Blanquet et al., 2015). Nevertheless, many of such studies showed

GENERAL DISCUSSION & PERSPECTIVES
241
early senescence phenotype with reduction in N fixation capacity, via gene knock-down. Thus, the essential role of these genes to maintain N-fixing instead of directly triggering nodule senescence is deductive. Nonetheless, a delayed senescence phenotype has been found by overexpression of two genes involved in NO detoxification, LjGlb1-1 and hmp (Cam et al., 2012; Fukudome et al., 2019), and a vapB toxin mutant (Lipuma et al., 2014). Accumulated toxic molecules thereof have a direct impact on nodule senescence. In terms of the degradation stage, the delayed nodule senescence phenotype has been observed via silencing two cysteine protease genes MtCP6 (Pierre et al., 2014) and MtCP77 (Deng et al., 2019). It appears feasible to keep the pink nodules fixing nitrogen beyond the timing – ‘StayPink’, which entitled the ANR funding project. Thus, it might be considered to apply in agriculture the senescence-delay strategies to promote plant growth or the protein content of seed production.
Nevertheless, it is still not sure if a prolonged nitrogen fixation period would benefit plant growth. To verify a positive role on plant production by hindering nodule senescence, it necessitates more related experiments or evidences in field conditions. There is still a long way to go from the basic scientific research to industrial/agronomic implementation, let alone the disputes on genetically modified organisms. From an evolutionary point of view, all plastids, such as chloroplast and mitochondria, are derived from the uptake of a prokaryotic ancestor that gradually evolved into a plant organelle (McFadden & van Dooren, 2004). Symbiosome may also be considered as an N-fixing organelle and might represent an intermediate phase in an analogous bacteria-to-plant organelle evolution (Van de Velde et al., 2006). It might be possible in the future to repress plant defence genes that function in nodule senescence so as to accelerate the evolution of the nitrogen-fixing symbiosome evolve to organelle in root cells. Ultimately, it could probably facilitate the engineering of symbiosis into cereal crops to increase yield and protein content. Additionally, a possible solution might be still hidden in the natural plant and rhizobium populations, in which efficient plant-rhizobium partners are trained in response to diverse environments. Substitutivity, compatible and efficient symbiotic strains might be selected from the training of repeated inoculation processes.

GENERAL DISCUSSION & PERSPECTIVES
242

243
ANNEX I
Materials & Methods
244

245
I. Plant and bacteria materials
I.1 Medicago truncatula
I.1.1 Seed sterilization and germination
The M. truncatula seed coat, and especially its hydrophobic, waxy outer tegument, must be scarified in order to allow the penetration of water and O2 that trigger germination (Garcia et al., 2006). Seed coat can be scarified mechanically by sandpaper or chemically by sulfuric acid. Here in this thesis we used only the chemical method. M. truncatula Jemalong A17 seeds (0.4g ≈ 100 seeds) were immersed in Sulphuric acid (H2SO4, 96%) for 6 minutes and sufficiently rinsed with cold sterile water. Then the seeds are sterilized with bleach 6° for 3min and rinsed thoroughly with sufficient sterile water. The seeds were afterwards soaked in water for more than 3h. The scarified seeds were thereafter spread on 0.4% agar and the plates were put inverted 2d at 4°C for vernalization and 1d at 16°C for gemination.
Plant growth conditions on soil
Germinated seedlings were transferred into pots containing a mixture of sand with perlite (1:3). Each pot was planted with 2 planets and covered for 3 days to keep humidity. Plants were regularly watered with nitrogen-free fertilizer (Plant-Prod: 0-15-40, Boulogne Billancourt, France) or reverse osmosis water. Plants were growing in growth chamber with a photoperiod of 16/8h and thermoperiod of 23/20°C respectively for day and night.
In vitro plant growth conditions
Seedlings were afterwards transferred on Fahraeus medium (Barker et al., 2006), with 6 plants for each petri dish. Top part of the gel (approx. 1/3) was removed to allow the growth of the plant aerial parts, whilst plant roots were protected from light by black plastic paper. Plants were growing in growth chamber with a photoperiod of 16/8h and thermoperiod of 23/20°C, respectively for day and night. To observe nodule phenotypes (nodule number, size, colour, etc.) in comparison to the wild-type nodules (WT). However, it may need to bear in mind that plant may not be grown more than 20 days in vitro, as this culture condition is not a good choice for mature nodules undergoing nodule senescence. In addition, exposure of the root system to light had a detrimental effect on nodule development, which necessities an addition of the ethylene biosynthesis inhibitor AVG to agar plates (Gourion et al., 2013).

246
I.2 Nicotiana benthamiana plant growth conditions
Seed of N. benthamiana should be vernalized 4°C for one week and geminated in growth chamber for one week. The pots were filled with growth media as sand:peat at ratio 1:1, and complete fertilizer was used in N. benthamiana plant growth. The growth chamber is with a photoperiod of 16/8h and thermoperiod of 25/20°C, respectively for day and night. The plantlets were transferred into new pots and grown for 3 weeks before infiltration.
Table S 1 Strains and plasmids used during the course of this thesis
Strains/plasmids Characteristics References
Strains
A. rhizogenes ARqua1 SmR, derivative of R1000 strain, moderate virulence (Quandt, 1993)
A. rhizogenes A4RS RifR, high virulence, originally used to transform coffee roots (Alpizar et al., 2006)
A. tumefaciens GV3101 RifR GenR (Van Larebeke et al., 1974)
A. tumefaciens GV3103 SmR RifR GenR (Holsters et al., 1980)
E. coli DB3.1 SmR, F– gyrA462, for propagating plasmids containing ccdB Invitrogen
E. coli DH5a F– Invitrogen
E. coli DH10b SmR, F– Invitrogen
E. coli XL1_Blue TetR, F’, the F plasmid contains ccdB (toxin) and ccdA (antidote) Stratagene
S. cerevisiae YM4271 Yeast reporter strain (Clontech) Fernanda de Carvalho-Niebel
S. meliloti 2011 SmR (Rosenberg et al., 1981)
S. meliloti 2011-gshB SmR TetR, gshB::pSup202gshB this study
Plasmids
pDONR207 GenR, entry vector for Gateway cloning Invritogen
pKGWFS7 SpR, destination vector for Gateway cloning (GOI::GUS::GFP) Invritogen
pDONR P4-P1R KmR, 5' entry vector for three entry multisite Gateway cloning Invritogen (Bruno Favery)
pENTR-GUS KmR, gene entry vector contains uidA (GUS) gene Invritogen
pENTR-T35S KmR, 3' entry vecteur contains the 35S terminator Invritogen (Bruno Favery)
pKM43-rolD::GFP SpR, destination vector for three entry multisite Gateway cloning (Karimi et al., 2002)
pLP100 KmR, –47bp Camv35S (pmin35S)::GUS Fernanda de Carvalho-Niebel
pHISi AmpR, HIS3, reporter vector for Yeast-One-Hybrid (Clontech) Fernanda de Carvalho-Niebel
pGAD-HA AmpR, Leu, prey vector for Yeast-One-Hybrid (Clontech) Fernanda de Carvalho-Niebel
I.3 Bacterial materials
Table S 1 is the list for microbe strains and plasmids used in the course of this thesis. Sinorhizobium meliloti 2011 was growing in LBMC (LB Broth, added with 2.5mM CaCl2 and 2.5 mM MgSO4) with 100μg/ml streptomycin at 30°C. Bacteria strains wild type Sinorhizobium meliloti 2011 and S. meliloti 2011 gshB was growing with LBMC medium with appropriate

247
antibiotics at 30°C. Escherichia coli DH5α was cultivated in LB Broth (Miller) at 37°C with appropriate antibiotics. Agrobacterium rhizogenes ARqua1was grown at 28°C in TY medium (pH = 7.2, 5g/L bacto-tryptone, 3g/L yeast extract, 6mM CaCl2) added with 100μg/ml streptomycin. Agrobacterium tumefaciens was grown in LB Broth (Miller) at 28°C with appropriate antibiotics. For making solid medium, agar (1.5%, 15g/L) was added in the liquid medium before autoclaving.
Inoculation of rhizobia to M. truncatula plants for nodulation
For nodulation studies, strains are recovered from –80°C on LBMC agar plates with appropriate antibiotics and incubate at 30°C for 3d. Pre-culture (5ml) was prepared in LBMC with appropriate antibiotics, and shaken overnight at 30°C. Inoculate 100ml LBMC with appropriate antibiotics, shake overnight at 30°C. Harvest the overnight culture by centrifuge (8000rpm, 5min) and rinse two times with sterile water. The inoculum is obtained by resuspending the pellet in sterile water, with O.D. 600nm adjust to 0.05-0.1. Inoculation at the bottom of each plant with 10ml inoculum with 50ml syringe in pots. In case of in vivo culture on plates of Fahraeus medium, bacteria suspension was prepared with optical density at O.D.
600nm =0.05. Seedlings were inoculated with 100μl S. meliloti inoculum for each plant two days after planting.
II. Molecular biology methods
II.1 Cloning
The polymerase chain reaction (PCR)
To generate promoter fragments, a 50µl PCR reaction was done with the Q5® High-Fidelity DNA Polymerase (ref. M0491, NEB) according to the manufacture’s guide. In primer design, site-specific sequences could be directly flanked by attB sites. In this case, a higher annealing temperature might be considered, but still depending on the primers. Alternatively, in order to avoid the long primers at around 40bp, attB adapters can be add with a two-PCR procedure: 1) 1st PCR using primers with site-specific sequences flanked by partial attB sequences; 2) PCR purification to remove the first step primers and primer-dimers; 3) 2nd PCR with attB adapters using 10µl of the purified 1st step PCR products (annealing: 55 ). An efficient amplification of DNA fragments is necessary to be observed with a gel electrophoresis before continuing the procedure. Finally, it is recommended to purify the PCR products with
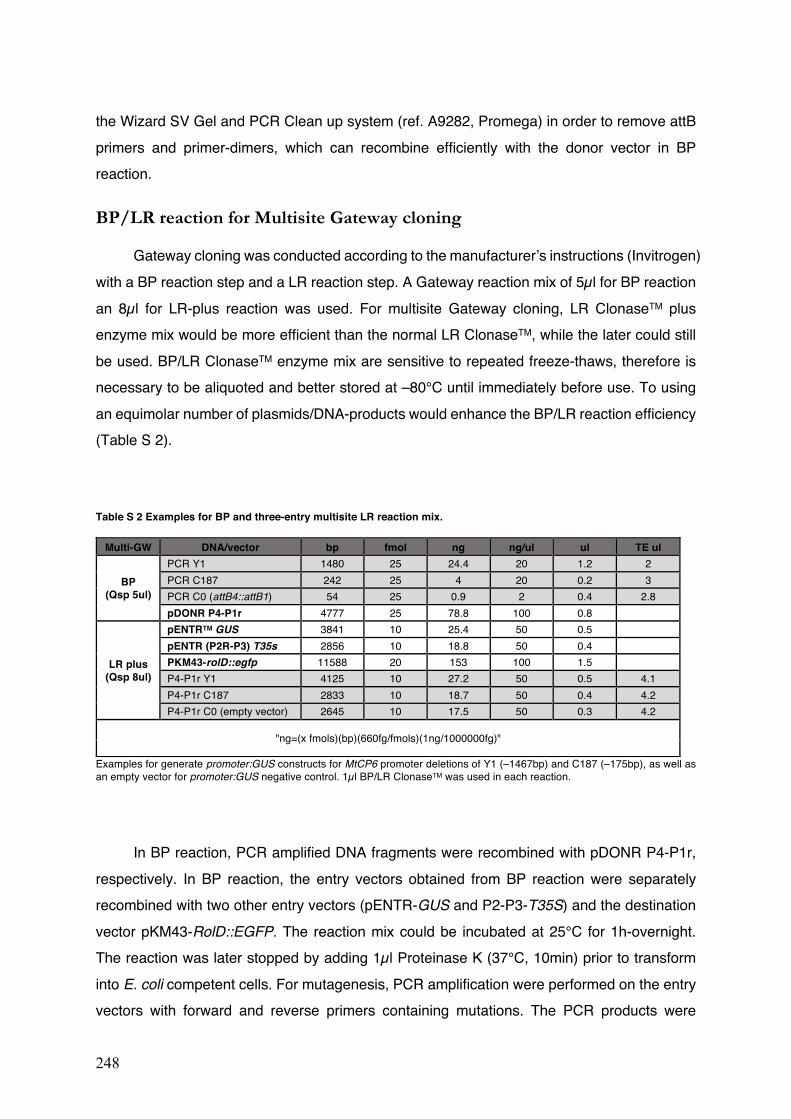
248
the Wizard SV Gel and PCR Clean up system (ref. A9282, Promega) in order to remove attB primers and primer-dimers, which can recombine efficiently with the donor vector in BP reaction.
BP/LR reaction for Multisite Gateway cloning
Gateway cloning was conducted according to the manufacturer’s instructions (Invitrogen) with a BP reaction step and a LR reaction step. A Gateway reaction mix of 5µl for BP reaction an 8µl for LR-plus reaction was used. For multisite Gateway cloning, LR ClonaseTM plus enzyme mix would be more efficient than the normal LR ClonaseTM, while the later could still be used. BP/LR ClonaseTM enzyme mix are sensitive to repeated freeze-thaws, therefore is necessary to be aliquoted and better stored at –80°C until immediately before use. To using an equimolar number of plasmids/DNA-products would enhance the BP/LR reaction efficiency (Table S 2).
Table S 2 Examples for BP and three-entry multisite LR reaction mix.
Multi-GW DNA/vector bp fmol ng ng/ul ul TE ul
BP (Qsp 5ul)
PCR Y1 1480 25 24.4 20 1.2 2 PCR C187 242 25 4 20 0.2 3 PCR C0 (attB4::attB1) 54 25 0.9 2 0.4 2.8 pDONR P4-P1r 4777 25 78.8 100 0.8
LR plus (Qsp 8ul)
pENTRTM GUS 3841 10 25.4 50 0.5 pENTR (P2R-P3) T35s 2856 10 18.8 50 0.4 PKM43-rolD::egfp 11588 20 153 100 1.5 P4-P1r Y1 4125 10 27.2 50 0.5 4.1 P4-P1r C187 2833 10 18.7 50 0.4 4.2 P4-P1r C0 (empty vector) 2645 10 17.5 50 0.3 4.2
"ng=(x fmols)(bp)(660fg/fmols)(1ng/1000000fg)"
Examples for generate promoter:GUS constructs for MtCP6 promoter deletions of Y1 (–1467bp) and C187 (–175bp), as well as an empty vector for promoter:GUS negative control. 1µl BP/LR ClonaseTM was used in each reaction.
In BP reaction, PCR amplified DNA fragments were recombined with pDONR P4-P1r,
respectively. In BP reaction, the entry vectors obtained from BP reaction were separately recombined with two other entry vectors (pENTR-GUS and P2-P3-T35S) and the destination vector pKM43-RolD::EGFP. The reaction mix could be incubated at 25°C for 1h-overnight. The reaction was later stopped by adding 1µl Proteinase K (37°C, 10min) prior to transform into E. coli competent cells. For mutagenesis, PCR amplification were performed on the entry vectors with forward and reverse primers containing mutations. The PCR products were

249
transformed into E. coli DH5a, and confirmed with subsequent colony PCR, plasmids isolation and sanger sequencing (Genewiz). The entry vectors carrying the wanted DNA mutations were recombined into destination vector by the multisite LR reactions.
Restriction cloning
As for restriction cloning, PCR products and vector plasmids were digested with corresponding restriction endonucleases (NEB) to the manufacture’s guide in a final volume of 50µl, respectively. The PCR-DNA/vector-backbone was gel purified and ligated with a molar ratio 6:1 with T4 DNA Ligase (ref M1801, Promega) in a final volume of 20µl (22°C, ~16h). The reaction was inactivated at 70°C for 5min. Considering the presence of salts, the ligation
mix is transformed into E. coli DH5a using the chemical transformation method (42°C, 30s) (Inoue et al., 1990).
II.2 Transforming bacteria competent cells
E. coli transformation
Electro-competent cells of E. coli were prepared for plasmid transformation. E. coli
strains DH5a and DH10b can be used for Gateway cloning. In addition to these two strains, E. coli XL1_blue could be used in restriction cloning (not for Gateway). Strains should be recovered from glycerol stock at –80°C 3-4 days in advance. An overnight pre-culture (5ml in LB) was prepared 37°C, 225 rpm. 1ml (O.D.600nm at ~0.2) was used to inoculate in 500ml 2xTY medium (Tryptone16 g/L, Yeast Extract10 g/L, NaCl 5.0 g/L, pH=6.8) or LB broth. When O.D.600nm reached 0.5-0.6 (for ~6h shaking at 30°C), culture was harvested by centrifugation (4000g, 15min, 4°C). The E. coli cells is necessary to be kept at low temperature after harvesting (on ice or in cold room 4°C). The pellets were rinsed two times with cold sterile water by the same centrifugation. Then the pellet was resuspended in 25ml cold sterile glycerol 10% and centrifuged again. After discarding the supernatant, the pellet was finally suspended in 2ml cold sterile glycerol 10% and the cell concentration was verified with 1µl cells in 1ml glycerol 10% resulted in O.D.600nm at ~0.2-0.3. The competent cells were aliquoted in 50µl (one for one reaction) and frozen in liquid nitrogen before stored at –80°C. Transformation efficiency was evaluated before used in subsequent transformation.
For transformation, 1-5µl Gateway-recombination-mix or ~50ng plasmids could be added into ice-thawed competent cells (50µl). The mix was transferred into the pre-cold electroporation cuvettes (0.1 or 0.2cm) and tapped down to the bottom. The cells were pulsed

250
(set at ‘Ec1’ or ‘Ec2’ for 0.1cm or 0.2 cm cuvettes, respectively) using the MicroPulserTM Electroporation Apparatus (Bio-Rad) with a constant ~5millisecond. Immediately, 950ml S.O.C. medium was added into the cuvettes and the cells were gently resuspended. [S.O.C. medium: 1) S.O.B. medium: Tryptone 20g/L, Yeast extract 5g/L, NaCl 0.5g/L, KCl 2.5mM; pH=7.0, autoclave; sterile magnesium solution: MgSO4 to 10mM and MgCl2 to 10mM; 2) add before use glucose (1M; 0.22µm filter-sterile) to a final concentration of 20mM]. The transformants were then transferred in a 10ml polypropylene tube and shaken for 1h at 37°C and 100µl were spread on LB plates with appropriate antibiotics. Colony PCR was performed using specific primers to select the positive clones. Plasmids were purified with Wizard Plus SV Miniprep kit (ref A1460, Promega). The cloning results were finally confirmed by sequencing. Similar methods were used to prepare competent cells for Agrobacterium Sp. and subsequent transformations with the medium (LB for A. tumefaciens, TY/Ca for A. rhizogenes) at 28°C.
II.3 Composite transgenic plants
A. rhizogenes ARqua1 strain is growth from glycerol –80°C stock on TY/CaCl2 plates with appropriate antibiotics at 28°C one week in advance. An overnight preculture (1.5ml) was then prepared 2d in advance. The preculture (400µl) was used to inoculate the TY/CaCl2 liquid medium (10ml) the day before plant transformation. On the day of transformation, bacteria were harvested (5000rpm, 10min) and rinsed before used. The bacteria pellet was finally resuspended in PS medium (peptone and sucrose, 1%) for stab-injecting into the hypocotyl part of plantlets.
After germination, seedlings of M. truncatula ecotype A17 were under limited growth at 4°C for 5 days in dark, in order to obtain thick shoots at hypocotyl region. Chimeric M. truncatula roots were produced by stabbing ARqua1 suspension at hypocotyl region. Plantlets were then settled in a mixture of perlite and sand (3:1). For an optimal plant growth, NH4NO3
(~5mM) of is recommended to provide at this step. However, after the 4-week transgenic root growth period, nitrogen nutrient should be washed away to avoid any nitrogen-mediated inhibition of the following inoculation. Plants were under growth condition with a photoperiod of 16h:8h, light:dark, and thermoperiod of 20/20°C, respectively. After one-week growth at 20°C (which allows efficient T-DNA insertion), plants were transferred at thermoperiod of 23/20°C. Three weeks later, transgenic roots were selected by hypocotyl localization (adventitious roots) (Figure S 1) and green fluorescence using a stereo-microscope (Leica MZ FL III). Non-transgenic roots were discarded, and chimeric plants were re-planted in nitrogen-

251
free pots. At this time, it could be better to allow around one-week of recovery time for the disturbed plants. Then plants were inoculated with S. meliloti inoculum (O.D. 600nm=0.1). Nodules were harvested for at 4 or 6 week-post-inoculation. Parts of the plants were used for nitrate treatment.
Figure S 1 Example of transgenic roots of M. truncatula generated by A. rhizogenes Arqua1.
The two images show before and after cutting off the non-transgenic root parts from the chimeric plant. The non-transgenic roots were cut and discarded, while the transgenic were confirmed with green florescence (EGFP marker) and used for further studies. The plant was 5 weeks old, including one-week limited growth under 4°C, one-week transformation under 20/20°C and three-week limited growth under 23/20°C.
Transient Expression in N. benthamiana leaves
The A. tumefaciens GV3101/GV3103 strains harbouring the constructs PAM-PAT35S-3xHA-GTW were growing in LB medium under antibiotic selection at 28. The overnight cultures were harvested and resuspended in Agro-buffer [10 mM MgCl2, 10mM MES, with 100 mM acetosyringone (No. D13440-6; Sigma-Aldrich), pH=5.6], subsequently incubated at room temperature for more than 3h. To check the interactions of TF candidates and promoter fragments, the corresponding resuspensions were adjusted to O.D.=0.5 and mixed prior to co-infiltration into leaves of 3-week-old N. benthamiana. The leaf discs were collected 36h after infiltration for histochemical GUS staining or frozen in liquid nitrogen and stored at −80 for further processes.
Before After
Non-transgenic roots
Transgenic roots

252
Plant total RNA extraction
Total RNA was extracted from 100mg plant materials harvested and grounded in liquid N2 using RNAzol® RT Reagent (Sigma-Aldrich) following the manufacturer’s instructions. The grounded nodule powder (100mg for each sample) was added with 1ml RNAzol® RT and 400µl DEPC H2O, then vortexed 15s and incubated at room temperature for 15min. The reaction mix was centrifuged (12 000g, 15min, 4°C) and the supernatant was transferred into a new tube. then added 500µL of isopropanol, mixed by inverting and incubate at room temperature for 10min. The RNA was then precipitated by centrifuge for 10min and rinsed two times with 75% ethanol (500µl and 400µl, respectively) by centrifuge. After elimination of the supernatant, RNA was finally eluted in 50µl Nuclease-free H2O and stored at –80°C. RNA quantity and quality were checked with Nanodrop 2000 (Thermo Scientific). After RQ1 RNase-Free DNase (ref M6101, Promega) treatment (37°C, 30min) in a final volume of 10µl (stop with 1µl RQ1 DNase Stop Solution, 65°C, 10min), gel electromigration was performed to confirm the RNA integrity and the absence of genomic contamination.
cDNA synthesis by reverse transcription (RT)
The DNase treated total RNA (1µg in 11µl) was used to synthesize cDNA with GoScriptTM Reverse Transcriptase (ref A6001, Promega): 1) pre-heating (70°C, 5min) with oligo (dT)15 primer (1µl); 2) reverse transcription (25°C, 5min; 42°C, 1h; 70°C, 15min) in a total volume of 20µl (1µl Nuclease-Free H2O, 4µl GoScript™ 5X Reaction Buffer, 0.5µl MgCl2 25mM, 1µl PCR Nucleotide Mix, 0.5µl Recombinant RNasin® Ribonuclease Inhibitor and 1µl GoScript™ Reverse Transcriptase). A control PCR was carried out on cDNA with a pair of intron-spanning primers of Leghaemoglobin 1 gene (Hopkins et al., 2014) to ensure the absence of genomic DNA contamination.
Quantitative PCR (qPCR)
Then cDNA was then diluted 40 times and 5µl cDNA 40X was firstly deposited in the 96-well qPCR plate. Then 10µl reaction mix (0.45µl Specific forward and reverse primer 10µM, 7.5µl master Mix, 1.9µl Nuclease-Free H2O qPCR, 0.15µl CXR Reference Dye) was added as described in the user manual of Go-Taq qPCR master Mix (ref A6002, Promega). The reaction was carried out in triplicates with AriaMx Real-time PCR system (Agilent) with qPCR cycle as: an initial denaturation (95°C, 3min), 40 thermal cycles (95°C 3s, 60°C 30s) and a melting cycle (95°C 30s, 65°C 30s, 95°C 30s). Housekeeping genes Mtc27 (Favery et al., 2002) and MtA38 (del Giudice et al., 2011) were used as endogenous control. Stability of the reference genes

253
were evaluated with an R package, RqPCRBase (Hilliou & Tran). Relative expressions were analysed with 2-ΔΔCt methods (Livak & Schmittgen, 2001) and confirmed with the output analysed with the R package.
II.4 Yeast-One-Hybrid screening (Y1H)
The Yeast-One-Hybrid screening of the tetramer of NS-box sequence for interacting TF candidates was done at the LIPM in Toulouse, under the supervision of F. de Carvalho-Niebel (DR CNRS) and the practical assistance of Lisa Frances (TR INRA). For performing such screen, we need first to generate and validate the yeast strain carrying the bait sequence (4x NS-box) before performing the screen of the cDNA library for NS-box interacting factors. We used the yeast reporter YM4271 strain (Clontech, Palo Alto, CA) for generating the bait 4x NS-box yeast and a nodule cDNA library (kindly provided by A. Niebel, LIPM) for performing the One-Hybrid screen following protocols adapted in F. de Carvalho-Niebel’ team (Andriankaja et al., 2007) on the basis of the “Clontech Matchmaker One-Hybrid system.
II.4.1 Yeast strain, culture conditions and transformation.
The yeast YM4271 strain (MATa, ura3-52, his3-200, ade2-101, lys2-801, leu2-3, 112, trp1-901, tyr1-501, gal4-Δ512, gal80-Δ538, ade5::hisG)(Liu et al., 1993) was grown at 28-30 on YPD [Difco peptone (BD 211677) 20g/L, yeast extract (BD 212750) 10g/L, pH=5.8, glucose 2%]. The sterilization of the media requires a special autoclave condition (temperature at 110, 20min) to avoid glucose degradation. Alternatively, a filter (0.2μm)-sterilized glucose mother stock solution (40%) is prepared and added to a final concentration of 2% to the autoclaved media (without glucose). For the One hybrid screen, the yeast was grown in the selective “synthetically defined” (SD) media, composed of yeast nitrogen base (Sigma, 6.7g/L), yeast synthetic Drop-out (Clontech, 0.62g/L) without Histidine (-His), without Histidine and Leucine (-His/-Leu) or without Leucine (Zhao et al.) supplemented with 2% glucose and an adjusted pH of 5.8. 2% Bacto-Agar (BD 214010) was added to media for the preparation of solid YPD or SD plates.
Transformation of yeast was done according to a PEG/Lithium acetate transformation protocol (according to the Clontech protocol handbook and minor modifications by F. de Carvalho-Niebel, LIPM). Firstly, in advance of 3-5 days before transformation, yeast YM4271 strains were recovered from the glycerol stock at -70 and streaked on YPD plates and grown overnight at 28. A liquid pre-culture in YPD (50ml) is initiated in YPD in the next day for about 5-6 hours until the culture reaches a O.D.600nm = 0.5-0.6 (Note: DO were measured with

254
a 10X dilution of the yeast culture). The culture is pelleted by using a swing centrifuge rotor at 3500rpm (5min) and resuspended in water before being centrifuged again and resuspended in 0.1 M Lithium Acetate (1ml freshly made). These “competent yeast cells” are then transferred to a 2 mL Eppendorf before centrifugation at maximum speed for 15 seconds. The pelleted yeast cells are then resuspended in 400μl of 0,1 M Lithium Acetate (enough for ~8 transformations) and aliquoted in 2 mL Eppendorf tubes (50 µl/tube). For each tube (50μl yeast competent cells aliquot) add 360μl of the Transformation Mix [containing 20μl of DNA (1μg cDNA library or prey plasmids), and a 340µl solution comprising 240µl PEG 50%, 36μl LiAc 1M, 14μl sterile dH2O, 50μl carrier DNA (ref 1467140; Roche, Basel, Switzerland) 2mg/ml (boiled for 5min in advance)]. After vortexing vigorously for 30s, the transformation mix were incubated in two steps at 30°C for 30 min (invert mix every 10min) and then at 42°C for 45min. After centrifugation at 7000 rpm for 15s, the pelleted yeast cells were recovered in 200µl sterile dH2O and incubated at room temperature for 10-30 min. (Note: yeast competent cells should be tenderly treated and homogenized by flicking the tube). Finally, 100μl of transformed yeast was plated on selective plates by using glass beads. For the generation of the bait 4xNS-box strain, they were plated on SD/-His while for selective prey screen they were plated on SD/-Leu/-His plates with or without different concentrations of 3-AT (ref A-8056, Sigma-Aldrich; stock solution (1M) was filter sterilized and stored at 4). For evaluating the yeast transformation efficiency, 1μl of the transformed yeast culture was diluted with 99μl sterile of dH2O and plated on SD/-Leu plates. The plates were thereafter incubated at 28 and yeast colony growth was monitored 3 to 5 days post-transformation.
II.4.2 Generation and selection of a 4x NS-box YM4271 yeast reporter
strain
Integration of the 4x NS-box bait sequence into the yeast YM4271 genome
The tetramer of tandem NS-box sequences (4x NS-box) was synthesised (Genewiz corporation, Takeley, United Kingdom) and inserted into the EcoRI/MluI restriction sites of the pHISi vector (Clontech) to generate the 4xNS-box-HIS3 construct. This construct was sent to Toulouse and L. Frances (LIPM) has linearized it by XhoI digestion and used it to transform and select YM4271 strains carrying the construct integrated in the yeast genome. Yeast positive colonies growing in selective SD/-His plates and carrying the 4xNS-box sequence (after validation by PCR and DNA sequencing) were selected to test their aptitude to grow in selective screen conditions.

255
Testing the ability of the 4x NS-box YM4271 reporter strain to grow in
selective screen conditions
The non-specific activation of the HIS3 reporter can results in yeast growth in the absence of Histidine. It is therefore important, before any cDNA library screen, to select the growth conditions that minimize this non-specific background level. The 3-aminotriazole (3-AT), a competitive inhibitor of the His3 enzyme, can be used to inhibit growth driven by background HIS3 expression. In this study, we first randomly selected 5 bait 4xNS-box strains and tested their ability to grow (in SD/-His media supplemented with increased concentrations of 3-AT (0mM, 1mM, 2mM, 4mM, 6mM, 8mM, 10mM and 15 mM). A relative inhibition of growth was obtained at 10 to 15mM of 3-AT, we therefore selected one of these yeast 4xNS-box strains for testing their ability to grow in selective 3-AT conditions after DNA transformation. To this end, we transformed the 4xNS-box (this study) and the 4x NF-box (Andriankaja et al., 2007) bait yeast strains with respective water control and a prey plasmid for expressing the NF-box-interactor ERN1 ERF transcription factor (Andriankaja et al., 2007). Serial dilutions (O.D.600nm = 1, 0.1, 0.01) of the transformed strains were generated in 96-well plates and spotted (3-5μl) on selective media (SD/Leu−, SD/His−/Leu− and SD/His−/Leu− + 10mM or 15 mM 3-AT). It is very important here to well homogenize the yeast solution before pipetting and make sure that the volume pipetted inside the tips are the same. Wait until the plates are dry and incubate at 28-30 for 2 days. Scan the plates when a differential growth is observed. The 4x NS-box yeast showed a higher growth background level then the 4xNF-box strain, nevertheless we could control it by the addition of 3-AT to the media at a given time frame.
II.4.3 Yeast-one hybrid screen of a nodule cDNA library
For performing the yeast-one hybrid screen using our 4xNS-box yeast bait strain, we decided to use a nodule cDNA library prepared in pGAD-HA vectors (Dualsystems Biotech AG, Switzerland, (http://www.dualsystems.com/) from RNA samples extracted from 4 and 19 days-old nodules (Baudin et al., 2015). The screening of the library was done under 10 mM 3-AT selective conditions.
For the Y1H screening, the yeast YM4271 4x NS-box strain was transformed with 20μg of DNA of the nodule cDNA library and the transformed yeast were selected on SD/-Leu/-His with 10 mM 3-AT. Transformation efficiency of the library was evaluated by plating 1µl (1µl/200µl) of the transformed yeast on SD/-Leu. The appearance of the yeast colonies was monitored over time (at 28°C) and yeast clones were classified according with their time of

256
appearance and their colony sizes (that measured from 1 to >2mm in diameter at 3 to 4 days after transformation).
Selection and validation of His+ yeast candidates
Candidate yeast clones grown in selective conditions (His+) were recovered from the screening plates in 10μl sterile H2O with a toothpick. To recover clones and confirm their growth in selective conditions, 2-5μl of the yeast suspension was plated on SD/-Leu/-His, SD/-Leu, or SD/-Leu/-His + 15mM 3-AT plates. After growth, yeast colony PCR was performed using 2μl of the yeast suspension and using the primer pairs listed in Table S 3. Each PCR reaction was performed with 2μl yeast suspension, with 0.5mM of each primer in a final volume of 20µl. The cycling conditions were as follows: 95°C for 10min followed by 35 cycles of denaturation at 95°C for 30s, annealing at 55°C for 30s, and extension at 72°C for 3min. 15μl of the PCR reaction were analysed on a 1.5% agarose gel. Positive clones comprising amplified bands were analysed by sequencing using 5μl of the PCR reaction. If the colony PCR resulted in more than one bands, DNA bands were PCR purified (Promega) and sent for sequencing.
After carefully analysing the DNA sequencing chromatograms, the candidate sequences were analysed for open reading frames that were in frame with the N-terminal Activation Domain (AD). Sequences and translated proteins of His+ candidates were compared by blastn, blastx and blastp algorithms (Altschul et al., 1997) to Medicago Mt4.0 and MtrunA17r5.0-ANR genomic databases. Plasmids of the candidates were thereafter prepared from a 3ml overnight culture of the candidate yeast clones grown in YPD, using the Miniprep kit (Promega) and eluted in 50μl dH2O. The extracted plasmid DNA preparations were used for E. coli DH5ɑ transformation for subsequently extraction of plasmid DNA of higher quality and yield for subsequent DNA sequencing and retransformation of yeast. 1 µg of plasmid preparations of ERF091, ERF092, ERF073 and ERF069 clones belonging to the ERF-IV, VII and VIII groups (Table 8) and of ERN1 (Andriankaja et al., 2007) were then used to respectively transform 4x NF-box and 4x NS yeast bait strains. The interaction of respective TF candidates and bait tetramer sequences were evaluated by spot inoculation of 1, 0,1 and 0,01 dilution series of yeast in SD/-Leu; SD/-Leu-His; and SD/-Leu-His + 15 mM 3-AT.

257
Table S 3 List of primers used in yeast-one-hybird screen strategy
Name Sequence Source Comments
4NSHindIII-MluI R GATCACGCGTATCCCAAGCTTATTCAACGG Li Y. multiple binding in tetramers
pHIS fw (LS) (Lisa F) TTCCCAGTCACGACGTTG Lisa F.
pHIS rev (LS) (Lisa R) ATATTCTTCGAAGAAATCAC Lisa F.
SacIEcoRI4NS F CAGGAGCTCAGCCGGAATTCTTGATTGTG Li Y. multiple binding in tetramers
pGAD10-R1 CAC AGT TGA AGT GAA CTT GC Lisa F. pGAD Sfi vecteur AD-levure
pGADT7-For1 AGT ACC CAT ACG ACG TAC CAG Lisa F. pGAD Sfi vecteur AD-levure
Bioinformatic methods in identification of regulatory elements
In silico analysis of MtCP6 promoter was performed in multiple research-based databases (PLACE, PlantPAN and Plantcare) to identify conserved cis-element motifs (Higo et al., 1999; Lescot et al., 2002; Chow et al., 2016). The used datasets of plant regulatory binding sites are in Table S2.
Table S 4 Web-based resources for plant cis-regulatory sequence analysis used in the course of this thesis.
Resource name URL
PLACE https://www.dna.affrc.go.jp/PLACE/?action=newplace
PlantCARE http://bioinformatics.psb.ugent.be/webtools/plantcare/html/
PlantPAN http://plantpan2.itps.ncku.edu.tw/index.html
TSSP http://www.softberry.com/berry.phtml?topic=tssp&group=programs&subgroup=promoter
TRANSFAC http://gene-regulation.com
III. Phenotyping
III.1 Plant phenotyping
Acetylene reduction assay.
Nitrogen fixation capacity was determined by Acetylene Reduction Assay (ARA) (Halliday & Pate, 1976), using a gas chromatograph system (Agilent GC 6890N; Agilent Technologies, Massy, France). Nodulated roots were detached from plants and put inside

258
sealed vials (10ml). In each vial, 10% of the atmosphere (1ml) was substituted with fresh prepared acetylene (1ml). After 1h of incubation at 28°C, gas samples were collected with 1ml syringes and needles were blocked with rubber before use. The gas samples (200µl) were analysed with a gas chromatograph system. Twelve plants were tested per replicate and at least three different biological replicates were analysed.
Nodule imaging
All the images of nodules were treated with the software Fiji (Schindelin et al., 2012). The brightness/contrast was adjusted in the post-processing of images. Confocal images of each channel were treated independently and then merged. As for nodule size analysis, nodule images were transformed in 8-bit (‘image’ –> ‘Type’) and converted to black and white (‘make binary’). Th parameters of selected particles (nodules) can be analysed under the menu ‘Analyze’.
Flow cytometry
Nodules (equal amount) were crushed in 1.5ml Eppendorf in phosphate buffer (pH=7.5) with 4% mannitol (m/v, 320~400 mOSM/L). The mixture was centrifuged 3min at 1500 rpm at 4°C to separate bacteria/bacteroids from plant tissue by differential density gradients. 500μl of supernatant was transferred into a new Eppendorf tube and the debris was discarded. Bacteroids were then harvested from the pellets after centrifugation (10min, 8000rpm, 4°C). Resuspended bacteroids were heat-killed (70°C, 15min) to allow the penetration of PI and stored at 4°C for up to one week. Bacteroids were stained with propidium iodide (PI; ref P4170, Sigma-Aldrich) (10μg/ml) and measured with flow cytometry analysis with a LSRII Fortessa flow cytometer (BD BioSciences, Rungis, France). Overnight cultures of free-living bacteria were prepared in the same way and used as control for DNA content. Totally 200 000 bacteria/bacteroids were measured with flow cytometry. Bacterial DNA content was determined from PI fluorescence at 585-625nm after excitation at 561nm with a single laser line. Single events were recorded and analyzed with BD FACSDiva v6.1.3 (BD BioSciences) and Kaluza v1.2 software (Beckman Coulter, Villepinte, France), respectively.

259
III.2 Histology and Microscopic observation
GUS staining
Transgenic roots were harvested via the EGFP selection at 4-wpi, 6-wpi, as well as 4-wpi with ammonium nitrate stress (3d). Nodules with root fragments were stained with X-Gluc (5-bromo-4-chloro-3-indolyl-β-Dglucuronide, cyclohexylammonium salt) (Euromedex) as described (Jefferson, 1987). Nodules with attached root fragments were pre-fixed in cold acetone 90% for 1h at –20°C. After three times rinse with phosphate buffer (pH=7.4, 77.4% NaH2PO4 100mM, 22.6% Na2HPO4 100mM), nodules were submerged in the GUS-staining solution (X-Gluc 0.5mg/ml, potassium ferricyanide 0.5mM, Potassium Ferrocyanide 0.5mM, prepared solution with phosphate buffer) after 20min of vacuum. Nodules were then incubated at 37°C either overnight or 4h in order to see a different staining intensity. (Note: overnight staining nodules would be over-stained for the native promoter of MtCP6, whereas it is necessary for the synthetic promoters containing 4xNS. Samples might be kept at 4°C in phosphate buffer and the staining time could be under strict control in the next morning to be stopped at a visible blue colour). Stained nodules were embedded in 4-6% (w/v) agarose and sections (70µm) were prepared with HM650V vibratome (Thermo Scientific). Images were taken in dark-field using Axioplan 2 microscope (Carl Zeiss). From all the experiments, more than 100 nodules were analysed from at least 16 plants. The experiments were conducted with three biological replicates.
GUS fluorimetric assay
In order to obtain a better estimation of population (nodule GUS activities) means, we have increased the sample size by harvesting transgenic nodules of every 5 plants into one tube. As results, we measured the GUS activity with three independent samples (15 plants) for each construction each biological replicate. In total, 9 samples were analysed with three biological replicates conducted for each construction, which should have increased the power of experiment. The harvested nodule samples were immediately frozen in liquid nitrogen and stored at –80°C. The florimetric assays was performed under the practical assistance of Lisa Frances (TR, INRA).
To quantify enzymatic GUS activity, each nodule sample were grounded independently in liquid nitrogen with mortar and pestle. Nodule powder (50mg) was mixed with 400µl GUS buffer [sodium phosphate 50 mM, pH 7.5; Na2EDTA 10mM, Triton X-100 0.1% (ref T8787, Sigma-Aldrich); 2-mercaptoethanol 10 mM (ref 63689, FLUKA), and sodium lauryl-sarcosine

260
0.1% (ref 61747, Sigma-Aldrich)], and vortexed until dissolved. The supernatant was taken after two times centrifugation (maximum speed at 14 000rpm, 15min, 4°C). Protein concentration of each supernatant was determined via the Bradford protein assay (Bradford reagent, Bio-Rad) (Bradford, 1976). Samples were either directly used for the following fluorometric assay or stored at –70°C until use.
As previously described (Boisson-Dernier et al., 2005), total protein (10µg) extraction was incubating shaking at 37°C, 400rpm for a kinetics of 4h in presence of 1mM substrate [4-methylumbelliferyl-β-D-glucuronide (4-MUG) (Biosynth AG)]. Upon hydrolysis of β-glucuronidase enzyme (GUS), the fluorescent compound 4-methylumbelliferone (4-MU) is produced, thus the detected florescent intensity (FI) are used in this assay. The excitation/emission maxima for 4-MU was about 355/460nm. GUS activities were measured using a microtiter fluorimeter (FLUOstar OMEGA, BMG Labtech, Germany) in 96-well microplates (COSTAR). Standard curves were prepared with a range of increasing concentrations of 4-MU (Sigma-Aldrich). The GUS activity is calculated as follows:
Live/Dead Staining
Molecular Probes’ LIVE/DEAD® BacLightTM Bacterial Viability Kits (ref. L7007, Invitrogen) was used to test bacterial viability with a two-color fluorescence assay. All bacteria (with/without intact membranes) will be labelled by the green-fluorescent nucleic acid stain SYTO® 9. In contrast, bacteria with damaged membranes are penetrated by the red-fluorescent nucleic acid stain propidium iodide (PI), causing a reduction in the SYTO 9 stain fluorescence when both dyes are present. Thus, bacteria with intact cell membranes stain fluorescent green, whereas bacteria with damaged membranes stain fluorescent red.
This method was then applied to test bacteria/bacteroid viability on fresh nodule sections (Haag et al., 2011). Nodules were harvested and kept in phosphate buffer (pH=7.2, 100mM) on ice, then embedded in 4-6% agarose and sectioned longitudinally with Vibratome HM650V (Thermo Scientific) into 70μm slices. Afterwards nodule sections were stained with 1.5µl/ml Component A (SYTO 9 and PI) and 1.5µl/ml Component B (SYTO 9 and PI) for 20 min. It would improve the staining with gently shaking if possible, and the staining can be stopped via rinse with phosphate buffer. Sample sections were then mounted on slides and observed
Activity of extract (pmol MU/min/μg)
FI/min
FI/pmol MU
reaction volume (μl)
sample volume (μl) Volume per test (μl)
Extract concentration (µg protein/μl)
= x x 1 1
x

261
with a laser scanning confocal microscope LSM880 (Carl Zeiss, Oberkochen, Germany). The excitation/emission maxima used for SYTO 9 stain is 488nm (Argon laser) and 506-545nm, respectively. As for PI, the fluorescence is excited at 561nm (DPSS laser) and its emission was detected at 650-684nm.

262

263
ANNEX II
Supplementary Figures

264
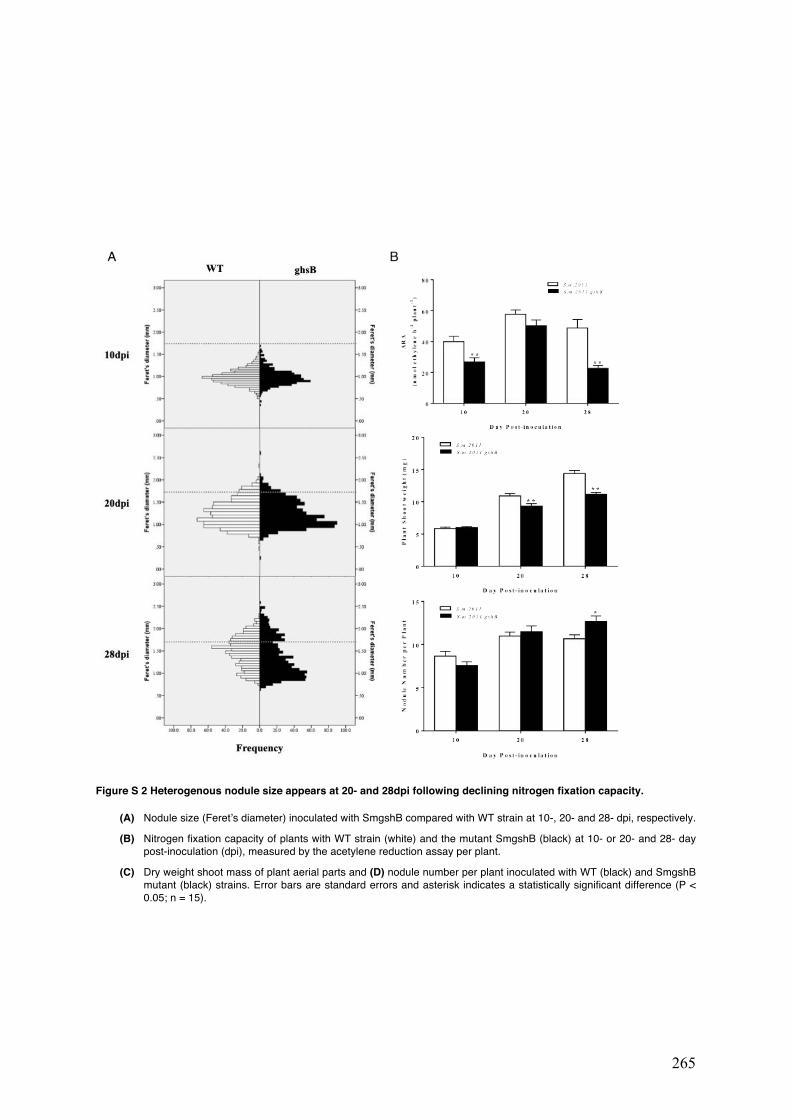
265
Figure S 2 Heterogenous nodule size appears at 20- and 28dpi following declining nitrogen fixation capacity.
(A) Nodule size (Feret’s diameter) inoculated with SmgshB compared with WT strain at 10-, 20- and 28- dpi, respectively.
(B) Nitrogen fixation capacity of plants with WT strain (white) and the mutant SmgshB (black) at 10- or 20- and 28- day post-inoculation (dpi), measured by the acetylene reduction assay per plant.
(C) Dry weight shoot mass of plant aerial parts and (D) nodule number per plant inoculated with WT (black) and SmgshB mutant (black) strains. Error bars are standard errors and asterisk indicates a statistically significant difference (P < 0.05; n = 15).

266
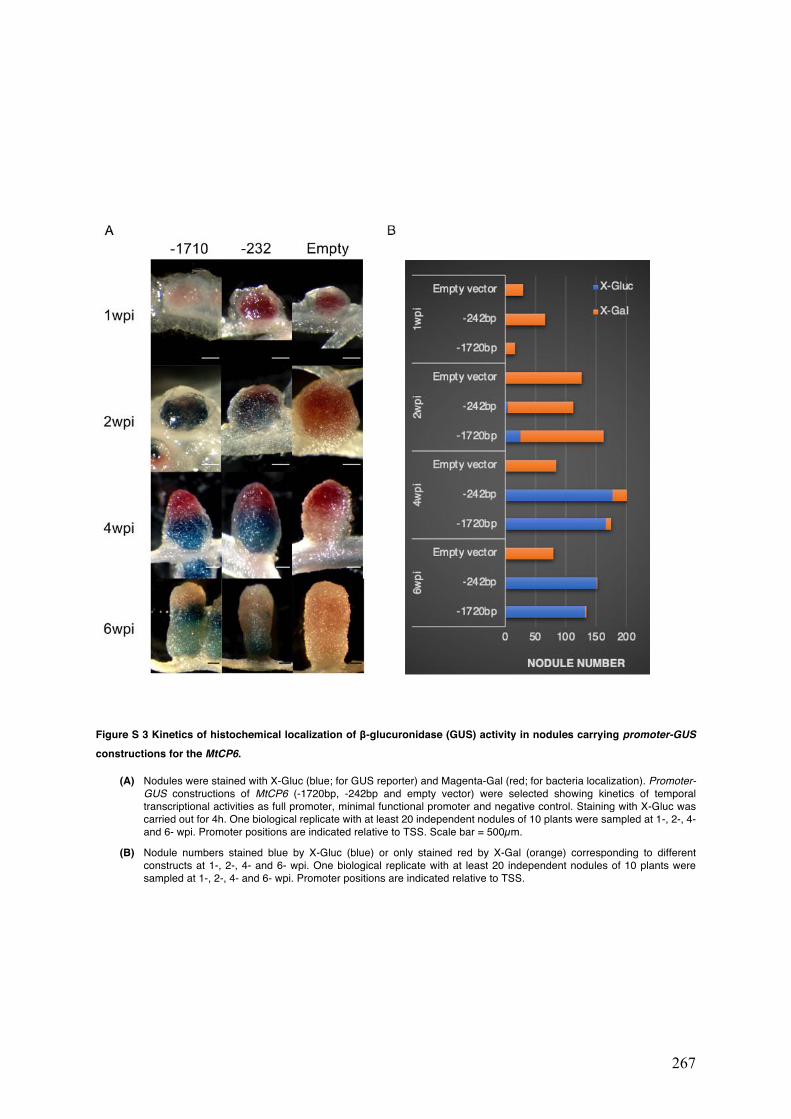
267
Figure S 3 Kinetics of histochemical localization of β-glucuronidase (GUS) activity in nodules carrying promoter-GUS constructions for the MtCP6.
(A) Nodules were stained with X-Gluc (blue; for GUS reporter) and Magenta-Gal (red; for bacteria localization). Promoter-GUS constructions of MtCP6 (-1720bp, -242bp and empty vector) were selected showing kinetics of temporal transcriptional activities as full promoter, minimal functional promoter and negative control. Staining with X-Gluc was carried out for 4h. One biological replicate with at least 20 independent nodules of 10 plants were sampled at 1-, 2-, 4- and 6- wpi. Promoter positions are indicated relative to TSS. Scale bar = 500µm.
(B) Nodule numbers stained blue by X-Gluc (blue) or only stained red by X-Gal (orange) corresponding to different constructs at 1-, 2-, 4- and 6- wpi. One biological replicate with at least 20 independent nodules of 10 plants were sampled at 1-, 2-, 4- and 6- wpi. Promoter positions are indicated relative to TSS.

268

269
Figure S 4 Putative cis-regulatory elements present in the minimal functional MtCP6 promoter (–242bp).
5’ flanking sequences of MtCP6 (–242bp to the TSS) was analysed using plant cis-regulatory element databases PLACE, PlantCare and PlantPAN 2.0 (Higo et al., 1999; Lescot et al., 2002; Chow et al., 2016).

270

271
Figure S 5 Re-growth and yeast colony PCR of Y1H candidates.
(C) Regrowth for 2 days of yeast candidates on selective medium SD H-/L- added with 10mM 3-AT. 88 yeast clones were recovered in 10µl H2O and 2µl was spotted for each clone.
(D) Yeast colony PCR. Green colored candidates were gel-purified and sequenced. Blue colored candidates contain only pGAD vector backbone sequences with PCR products at ~250bp.

272
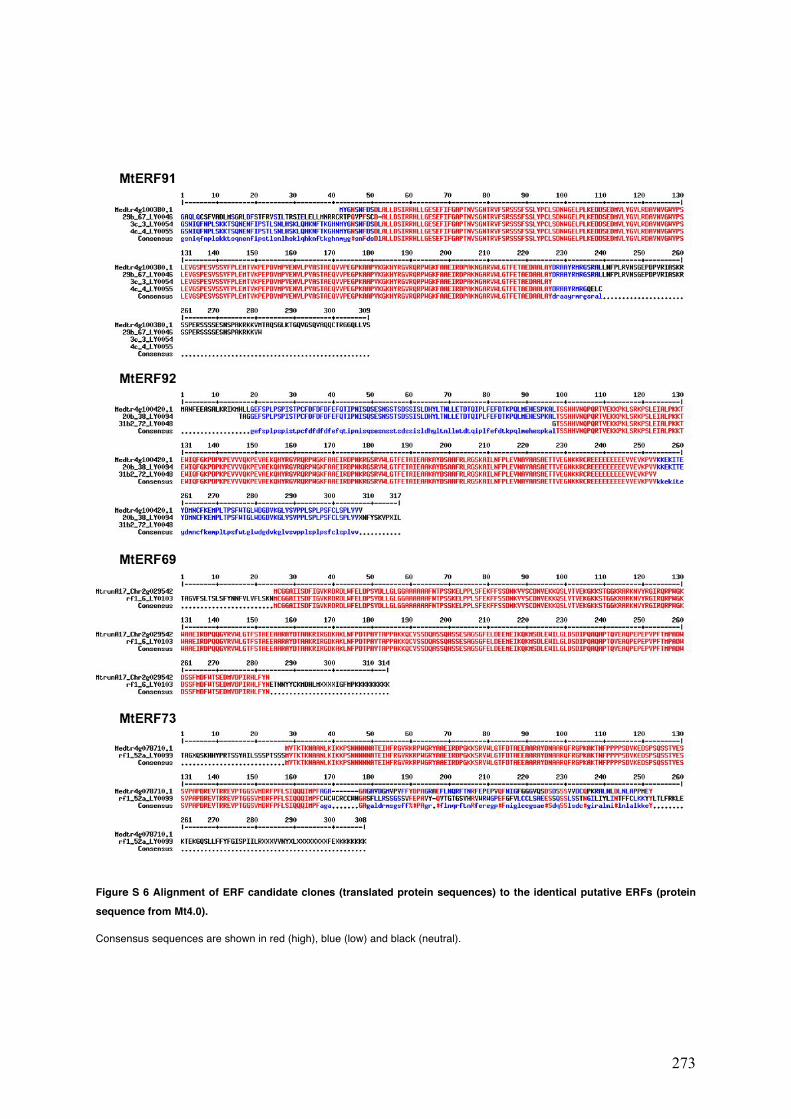
273
Figure S 6 Alignment of ERF candidate clones (translated protein sequences) to the identical putative ERFs (protein sequence from Mt4.0).
Consensus sequences are shown in red (high), blue (low) and black (neutral).

274
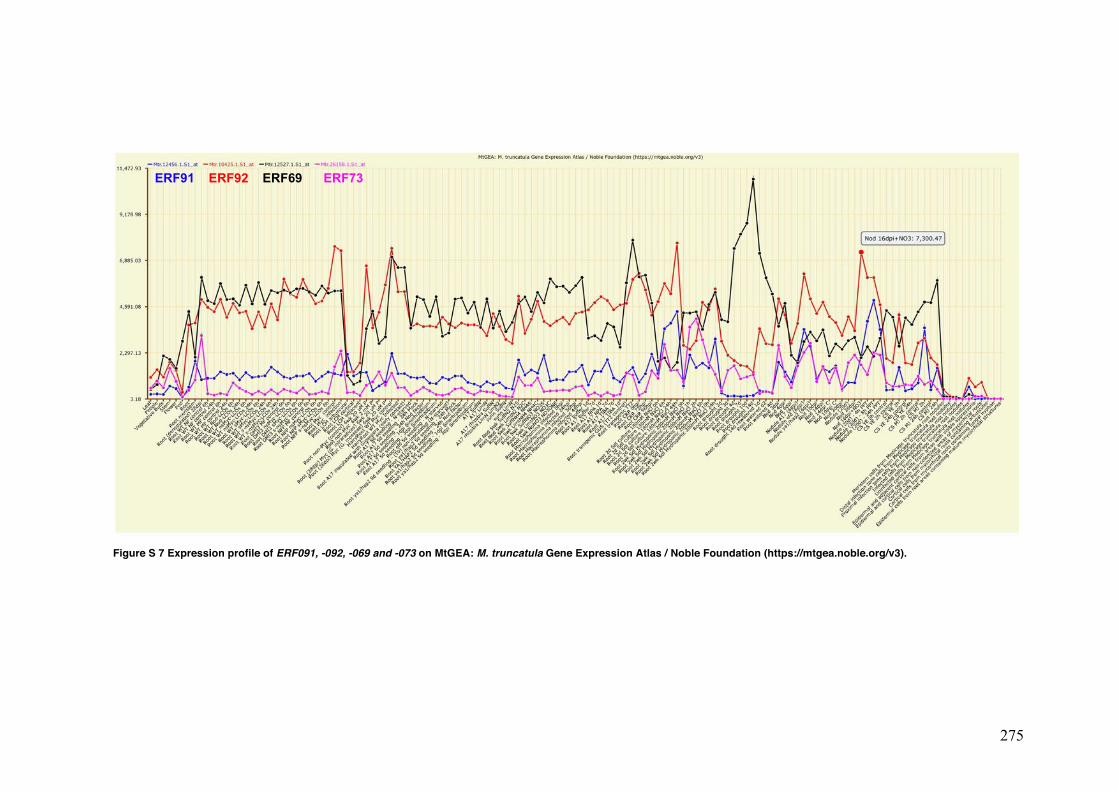
275
Figure S 7 Expression profile of ERF091, -092, -069 and -073 on MtGEA: M. truncatula Gene Expression Atlas / Noble Foundation (https://mtgea.noble.org/v3).
ERF91 ERF92 ERF69 ERF73

276

277
REFERENCES

278

279
A
Alesandrini F, Mathis R, Van de Sype G, Hérouart D, Puppo A. 2003. Possible roles for a cysteine
protease and hydrogen peroxide in soybean nodule development and senescence. New Phytologist 158(1): 131-138.
Allen MD, Yamasaki K, Ohme-Takagi M, Tateno M, Suzuki M. 1998. A novel mode of DNA recognition by a beta-sheet revealed by the solution structure of the GCC-box binding domain in complex with DNA. The EMBO Journal 17(18): 5484-5496.
Alon U. 2019. An introduction to systems biology: design principles of biological circuits: CRC press. Alpizar E, Dechamp E, Espeout S, Royer M, Lecouls AC, Nicole M, Bertrand B, Lashermes P,
Etienne H. 2006. Efficient production of Agrobacterium rhizogenes-transformed roots and composite plants for studying gene expression in coffee roots. Plant Cell Reports 25(9): 959-967.
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ. 1997. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Research 25(17): 3389-3402.
Alunni B, Kevei Z, Redondo-Nieto M, Kondorosi A, Mergaert P, Kondorosi E. 2007. Genomic organization and evolutionary insights on GRP and NCR genes, two large nodule-specific gene families in Medicago truncatula. Molecular Plant-Microbe Interactions 20(9): 1138-1148.
Anderson JP, Lichtenzveig J, Gleason C, Oliver RP, Singh KB. 2010. The B-3 ethylene response factor MtERF1-1 mediates resistance to a subset of root pathogens in Medicago truncatula without adversely affecting symbiosis with rhizobia. Plant Physiology 154(2): 861-873.
Andriankaja A, Boisson-Dernier A, Frances L, Sauviac L, Jauneau A, Barker DG, de Carvalho-Niebel F. 2007. AP2-ERF transcription factors mediate Nod factor dependent Mt ENOD11 activation in root hairs via a novel cis-regulatory motif. The Plant Cell 19(9): 2866-2885.
Andrio E, Marino D, Marmeys A, de Segonzac MD, Damiani I, Genre A, Huguet S, Frendo P, Puppo A, Pauly N. 2013. Hydrogen peroxide-regulated genes in the Medicago truncatula-Sinorhizobium meliloti symbiosis. New Phytologist 198(1): 179-189.
Arrighi J-F, Barre A, Amor BB, Bersoult A, Soriano LC, Mirabella R, de Carvalho-Niebel F, Journet E-P, Ghérardi M, Huguet T. 2006. The Medicago truncatula lysine motif-receptor-like kinase gene family includes NFP and new nodule-expressed genes. Plant Physiology 142(1): 265-279.
Arumuganathan K, Earle E. 1991. Nuclear DNA content of some important plant species. Plant Molecular Biology Reporter 9(3): 208-218.
Asamizu E, Shimoda Y, Kouchi H, Tabata S, Sato S. 2008. A positive regulatory role for LjERF1 in the nodulation process is revealed by systematic analysis of nodule-associated transcription factors of Lotus japonicus. Plant Physiology 147(4): 2030-2040.
Aslund F, Zheng M, Beckwith J, Storz G. 1999. Regulation of the OxyR transcription factor by hydrogen peroxide and the cellular thiol-disulfide status. Proceedings of the National Academy of Sciences of the United States of America 96(11): 6161-6165.
Asp T, Bowra S, Borg S, Holm PB. 2004. Cloning and characterisation of three groups of cysteine protease genes expressed in the senescing zone of white clover (Trifolium repens) nodules. Plant Science 167(4): 825-837.
Avila-Ospina L, Moison M, Yoshimoto K, Masclaux-Daubresse C. 2014. Autophagy, plant senescence, and nutrient recycling. Journal of Experimental Botany 65(14): 3799-3811.

280
Azani N, Babineau M, Bailey CD, Banks H, Barbosa AR, Pinto RB, Boatwright JS, Borges LM, Brown GK, Bruneau A. 2017. A new subfamily classification of the Leguminosae based on a taxonomically comprehensive phylogeny: The Legume Phylogeny Working Group (LPWG). Taxon 66(1): 44-77.
B Badis G, Berger MF, Philippakis AA, Talukder S, Gehrke AR, Jaeger SA, Chan ET, Metzler G,
Vedenko A, Chen X, et al. 2009. Diversity and complexity in DNA recognition by transcription factors. Science 324(5935): 1720-1723.
Baena-González E, Hanson J. 2017. Shaping plant development through the SnRK1–TOR metabolic regulators. Current Opinion In Plant Biology 35: 152-157.
Baker SS, Wilhelm KS, Thomashow MF. 1994. The 5′-region of Arabidopsis thaliana cor15a has cis-acting elements that confer cold-, drought-and ABA-regulated gene expression. Plant Molecular Biology 24(5): 701-713.
Balint-Kurti P. 2019. The plant hypersensitive response: concepts, control and consequences. Molecular plant pathology 20(8): 1163-1178.
Barker DG, Pfaff T, Moreau D, Groves E, Ruffel S, Lepetit M, Whitehand S, Maillet F, Nair RM, Journet E-P 2006. Growing M. truncatula: choice of substrates and growth conditions. In: The Medicago truncatula handbook. Mathesius U, Journet EP, Sumner LW (eds). ISBN 0-9754303-1-9. http://www.noble.org/MedicagoHandbook/
Barrett AJ, Woessner JF, Rawlings ND. 2012. Handbook of proteolytic enzymes: Elsevier. Baudin M, Laloum T, Lepage A, Rípodas C, Ariel F, Frances L, Crespi M, Gamas PC, Blanco FA,
Zanetti ME. 2015. A phylogenetically conserved group of NF-Y transcription factors interact to control nodulation in legumes. Plant Physiology: pp. 01144.02015.
Becana M, Dalton DA, Moran JF, Iturbe-Ormaetxe I, Matamoros MA, C. Rubio M. 2000. Reactive oxygen species and antioxidants in legume nodules. Physiologia Plantarum 109(4): 372-381.
Becana M, Matamoros MA, Udvardi M, Dalton DA. 2010. Recent insights into antioxidant defences of legume root nodules. New Phytologist 188(4): 960-976.
Becker A, Berges H, Krol E, Bruand C, Ruberg S, Capela D, Lauber E, Meilhoc E, Ampe F, de Bruijn FJ, et al. 2004. Global changes in gene expression in Sinorhizobium meliloti 1021 under microoxic and symbiotic conditions. Molecular Plant-Microbe Interactions 17(3): 292-303.
Benedito VA, Torres-Jerez I, Murray JD, Andriankaja A, Allen S, Kakar K, Wandrey M, Verdier J, Zuber H, Ott T, et al. 2008. A gene expression atlas of the model legume Medicago truncatula. The Plant Journal 55(3): 504-513.
Benyamina SM, Baldacci-Cresp F, Couturier J, Chibani K, Hopkins J, Bekki A, de Lajudie P, Rouhier N, Jacquot JP, Alloing G, et al. 2013. Two Sinorhizobium meliloti glutaredoxins regulate iron metabolism and symbiotic bacteroid differentiation. Environmental Microbiology 15(3): 795-810.
Bethlenfalvay GJ, Phillips DA. 1977. Ontogenetic Interactions between Photosynthesis and Symbiotic Nitrogen Fixation in Legumes. Plant Physiology 60(3): 419-421.
Biémont C, Vieira C. 2006. Junk DNA as an evolutionary force. Nature 443(7111): 521-524. Blahnik KR, Dou L, O'Geen H, McPhillips T, Xu X, Cao AR, Iyengar S, Nicolet CM, Ludascher B,
Korf I, et al. 2010. Sole-Search: an integrated analysis program for peak detection and functional annotation using ChIP-seq data. Nucleic Acids Research 38(3): e13.

281
Blanquet P, Silva L, Catrice O, Bruand C, Carvalho H, Meilhoc E. 2015. Sinorhizobium meliloti Controls Nitric Oxide-Mediated Post-Translational Modification of a Medicago truncatula Nodule Protein. Molecular Plant-Microbe Interactions 28(12): 1353-1363.
Boisson-Dernier A, Andriankaja A, Chabaud M, Niebel A, Journet EP, Barker DG, de Carvalho-Niebel F. 2005. MtENOD11 gene activation during rhizobial infection and mycorrhizal arbuscule development requires a common AT-rich-containing regulatory sequence. Molecular Plant-Microbe Interactions 18(12): 1269-1276.
Bradford MM. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry 72(1-2): 248-254.
Breeze E, Harrison E, McHattie S, Hughes L, Hickman R, Hill C, Kiddle S, Kim YS, Penfold CA, Jenkins D, et al. 2011. High-resolution temporal profiling of transcripts during Arabidopsis leaf senescence reveals a distinct chronology of processes and regulation. The Plant Cell 23(3): 873-894.
Brewin NJ. 2004. Plant Cell Wall Remodelling in the Rhizobium–Legume Symbiosis. Critical Reviews In Plant Sciences 23(4): 293-316.
Brilli M, Fondi M, Fani R, Mengoni A, Ferri L, Bazzicalupo M, Biondi EG. 2010. The diversity and evolution of cell cycle regulation in alpha-proteobacteria: a comparative genomic analysis. BMC systems biology 4(1): 52.
Broghammer A, Krusell L, Blaise M, Sauer J, Sullivan JT, Maolanon N, Vinther M, Lorentzen A, Madsen EB, Jensen KJ, et al. 2012. Legume receptors perceive the rhizobial lipochitin oligosaccharide signal molecules by direct binding. Proceedings of the National Academy of Sciences of the United States of America 109(34): 13859-13864.
Broglie KE, Biddle P, Cressman R, Broglie R. 1989. Functional analysis of DNA sequences responsible for ethylene regulation of a bean chitinase gene in transgenic tobacco. The Plant Cell 1(6): 599-607.
Buono RA, Hudecek R, Nowack MK. 2019. Plant proteases during developmental programmed cell death. Journal of Experimental Botany 70(7): 2097-2112.
Burgess BK, Lowe DJ. 1996. Mechanism of Molybdenum Nitrogenase. Chemical Reviews 96(7): 2983-3012.
Burris RH, Roberts GP. 1993. Biological nitrogen fixation. Annual Review of Nutrition 13(1): 317-335.
C Cabeza R, Koester B, Liese R, Lingner A, Baumgarten V, Dirks J, Salinas-Riester G, Pommerenke
C, Dittert K, Schulze J. 2014. An RNA sequencing transcriptome analysis reveals novel insights into molecular aspects of the nitrate impact on the nodule activity of Medicago truncatula. Plant Physiology 164(1): 400-411.
Cam Y, Pierre O, Boncompagni E, Herouart D, Meilhoc E, Bruand C. 2012. Nitric oxide (NO): a key player in the senescence of Medicago truncatula root nodules. New Phytologist 196(2): 548-560.
Canfield DE, Glazer AN, Falkowski PG. 2010. The evolution and future of Earth's nitrogen cycle. Science 330(6001): 192-196.
Carmel-Harel O, Storz G. 2000. Roles of the glutathione- and thioredoxin-dependent reduction systems in the Escherichia coli and Saccharomyces cerevisiae responses to oxidative stress. Annual Review of Microbiology 54(1): 439-461.

282
Carpenter CD, Kreps JA, Simon AE. 1994. Genes encoding glycine-rich Arabidopsis thaliana proteins with RNA-binding motifs are influenced by cold treatment and an endogenous circadian rhythm. Plant Physiology 104(3): 1015-1025.
Carri Re SB, Verdenaud M, Gough C, Gouzy JRM, Gamas P. 2020. LeGOO: An Expertized Knowledge Database for the Model Legume Medicago truncatula. Plant and Cell Physiology 61(1): 203-211.
Cebolla A, Vinardell JM, Kiss E, Olah B, Roudier F, Kondorosi A, Kondorosi E. 1999. The mitotic inhibitor ccs52 is required for endoreduplication and ploidy‐dependent cell enlargement in plants. The EMBO Journal 18(16): 4476-4484.
Cerri M. 2013. Régulation et fonctions de facteurs de transcription ERF ERN au cours de la symbiose entre Medicago truncatula et Sinorhizobium meliloti. Doctoral dissertation, Université de Toulouse.
Cerri MR, Frances L, Laloum T, Auriac MC, Niebel A, Oldroyd GE, Barker DG, Fournier J, de Carvalho-Niebel F. 2012. Medicago truncatula ERN transcription factors: regulatory interplay with NSP1/NSP2 GRAS factors and expression dynamics throughout rhizobial infection. Plant Physiology 160(4): 2155-2172.
Cerri MR, Wang Q, Stolz P, Folgmann J, Frances L, Katzer K, Li X, Heckmann AB, Wang TL, Downie JA, et al. 2017. The ERN1 transcription factor gene is a target of the CCaMK/CYCLOPS complex and controls rhizobial infection in Lotus japonicus. New Phytologist 215(1): 323-337.
Charpentier M, Oldroyd GE. 2013. Nuclear calcium signaling in plants. Plant Physiology 163(2): 496-503.
Charpentier M, Sun J, Vaz Martins T, Radhakrishnan GV, Findlay K, Soumpourou E, Thouin J, Very AA, Sanders D, Morris RJ, et al. 2016. Nuclear-localized cyclic nucleotide-gated channels mediate symbiotic calcium oscillations. Science 352(6289): 1102-1105.
Chen Y, Chen W, Li X, Jiang H, Wu P, Xia K, Yang Y, Wu G. 2014. Knockdown of LjIPT3 influences nodule development in Lotus japonicus. Plant and Cell Physiology 55(1): 183-193.
Cheng G, Karunakaran R, East AK, Munoz-Azcarate O, Poole PS. 2017. Glutathione affects the transport activity of Rhizobium leguminosarum 3841 and is essential for efficient nodulation. Fems Microbiology Letters 364(8).
Cheng J, Sibley CD, Zaheer R, Finan TM. 2007. A Sinorhizobium meliloti minE mutant has an altered morphology and exhibits defects in legume symbiosis. Microbiology 153(2): 375-387.
Chesney JA, Eaton JW, Mahoney JR, Jr. 1996. Bacterial glutathione: a sacrificial defence against chlorine compounds. Journal of Bacteriology 178(7): 2131-2135.
Cho YH, Yoo SD, Sheen J. 2006. Regulatory functions of nuclear hexokinase1 complex in glucose signaling. Cell 127(3): 579-589.
Chow CN, Zheng HQ, Wu NY, Chien CH, Huang HD, Lee TY, Chiang-Hsieh YF, Hou PF, Yang TY, Chang WC. 2016. PlantPAN 2.0: an update of plant promoter analysis navigator for reconstructing transcriptional regulatory networks in plants. Nucleic Acids Research 44(D1): D1154-1160.
Cilliers M, van Wyk SG, van Heerden PDR, Kunert KJ, Vorster BJ. 2018. Identification and changes of the drought-induced cysteine protease transcriptome in soybean (Glycine max) root nodules. Environmental and Experimental Botany 148: 59-69.
Colebatch G, Desbrosses G, Ott T, Krusell L, Montanari O, Kloska S, Kopka J, Udvardi MK. 2004. Global changes in transcription orchestrate metabolic differentiation during symbiotic nitrogen fixation in Lotus japonicus. The Plant Journal 39(4): 487-512.
Coll NS, Smidler A, Puigvert M, Popa C, Valls M, Dangl JL. 2014. The plant metacaspase AtMC1 in pathogen-triggered programmed cell death and aging: functional linkage with autophagy. Cell Death and Differentiation 21(9): 1399-1408.

283
Combier JP, Frugier F, de Billy F, Boualem A, El-Yahyaoui F, Moreau S, Vernie T, Ott T, Gamas P, Crespi M, et al. 2006. MtHAP2-1 is a key transcriptional regulator of symbiotic nodule development regulated by microRNA169 in Medicago truncatula. Genes & Development 20(22): 3084-3088.
Cook D, Dreyer D, Bonnet D, Howell M, Nony E, VandenBosch K. 1995. Transient induction of a peroxidase gene in Medicago truncatula precedes infection by Rhizobium meliloti. The Plant Cell 7(1): 43-55.
Craine JM, Elmore AJ, Wang L, Aranibar J, Bauters M, Boeckx P, Crowley BE, Dawes MA, Delzon S, Fajardo A, et al. 2018. Isotopic evidence for oligotrophication of terrestrial ecosystems. Nature Ecology and Evolution 2(11): 1735-1744.
Creelman RA, Mullet JE. 1997. Biosynthesis and Action of Jasmonates in Plants. Annual Review Of Plant Physiology and Plant Molecular Biology 48(1): 355-381.
Crepin N, Rolland F. 2019. SnRK1 activation, signaling, and networking for energy homeostasis. Current Opinion In Plant Biology 51: 29-36.
Crockford AJ, Behncke C, Williams HD. 1996. The adaptation of Rhizobium leguminosarum bv. phaseoli to oxidative stress and its overlap with other environmental stress responses. Microbiology 142(2): 331-336.
Cunningham F, Amode MR, Barrell D, Beal K, Billis K, Brent S, Carvalho-Silva D, Clapham P, Coates G, Fitzgerald S, et al. 2015. Ensembl 2015. Nucleic Acids Research 43(Database issue): D662-669.
D D'Haeseleer K, De Keyser A, Goormachtig S, Holsters M. 2010. Transcription factor MtATB2: about
nodulation, sucrose and senescence. Plant and Cell Physiology 51(9): 1416-1424. D’Haeze W, Chaparro C, De Keyser A, Deleu S, De Rycke R, Goormachtig S, Lievens S, Mathis
R, Schroeyers K, Van De Velde W. 2002. Role of reactive oxygen species and ethylene in programmed cell death during nodule initiation on Sesbania rostrata. Nitrogen Fixation: Global Perspectives. CAB International, Oxon, UK: 137-141.
Dalle-Donne I, Rossi R, Giustarini D, Colombo R, Milzani A. 2007. S-glutathionylation in protein redox regulation. Free Radical Biology and Medicine 43(6): 883-898.
Dalton DA, Joyner SL, Becana M, Iturbe-Ormaetxe I, Chatfield JM. 1998. Antioxidant defences in the peripheral cell layers of legume root nodules. Plant Physiology 116(1): 37-43.
Dalton DA, Russell SA, Hanus FJ, Pascoe GA, Evans HJ. 1986. Enzymatic reactions of ascorbate and glutathione that prevent peroxide damage in soybean root nodules. Proceedings of the National Academy of Sciences of the United States of America 83(11): 3811-3815.
Dangl JL, Dietrich RA, Richberg MH. 1996. Death don't have no mercy: cell death programs in plant-microbe interactions. The Plant Cell 8(10): 1793.
Danisman S, van der Wal F, Dhondt S, Waites R, de Folter S, Bimbo A, van Dijk AD, Muino JM, Cutri L, Dornelas MC, et al. 2012. Arabidopsis class I and class II TCP transcription factors regulate jasmonic acid metabolism and leaf development antagonistically. Plant Physiology 159(4): 1511-1523.
David M, Daveran ML, Batut J, Dedieu A, Domergue O, Ghai J, Hertig C, Boistard P, Kahn D. 1988. Cascade regulation of nif gene expression in Rhizobium meliloti. Cell 54(5): 671-683.
de Zelicourt A, Diet A, Marion J, Laffont C, Ariel F, Moison M, Zahaf O, Crespi M, Gruber V, Frugier F. 2012. Dual involvement of a Medicago truncatula NAC transcription factor in root abiotic stress response and symbiotic nodule senescence. The Plant Journal 70(2): 220-230.

284
Deakin WJ, Broughton WJ. 2009. Symbiotic use of pathogenic strategies: rhizobial protein secretion systems. Nature Reviews Microbiology 7(4): 312-320.
Dechorgnat J, Nguyen CT, Armengaud P, Jossier M, Diatloff E, Filleur S, Daniel-Vedele F. 2011. From the soil to the seeds: the long journey of nitrate in plants. Journal of Experimental Botany 62(4): 1349-1359.
Dejardin J, Kingston RE. 2009. Purification of proteins associated with specific genomic Loci. Cell 136(1): 175-186.
del Giudice J, Cam Y, Damiani I, Fung-Chat F, Meilhoc E, Bruand C, Brouquisse R, Puppo A, Boscari A. 2011. Nitric oxide is required for an optimal establishment of the Medicago truncatula-Sinorhizobium meliloti symbiosis. New Phytologist 191(2): 405-417.
Deng J, Zhu F, Liu J, Zhao Y, Wen J, Wang T, Dong J. 2019. Transcription Factor bHLH2 Represses CYSTEINE PROTEASE77 to Negatively Regulate Nodule Senescence. Plant Physiology 181(4): 1683-1703.
Desclos M, Etienne P, Coquet L, Jouenne T, Bonnefoy J, Segura R, Reze S, Ourry A, Avice JC. 2009. A combined 15N tracing/proteomics study in Brassica napus reveals the chronology of proteomics events associated with N remobilisation during leaf senescence induced by nitrate limitation or starvation. Proteomics 9(13): 3580-3608.
Dhanushkodi R, Matthew C, McManus MT, Dijkwel PP. 2018. Drought-induced senescence of Medicago truncatula nodules involves serpin and ferritin to control proteolytic activity and iron levels. New Phytologist 220(1): 196-208.
Diaz-Mendoza M, Velasco-Arroyo B, Santamaria ME, Gonzalez-Melendi P, Martinez M, Diaz I. 2016. Plant senescence and proteolysis: two processes with one destiny. Genetics and Molecular Biology 39(3): 329-338.
Dill A, Thomas SG, Hu J, Steber CM, Sun TP. 2004. The Arabidopsis F-box protein SLEEPY1 targets gibberellin signaling repressors for gibberellin-induced degradation. The Plant Cell 16(6): 1392-1405.
Dixon R, Kahn D. 2004. Genetic regulation of biological nitrogen fixation. Nature Reviews Microbiology 2(8): 621-631.
Dombrecht B, Heusdens C, Beullens S, Verreth C, Mulkers E, Proost P, Vanderleyden J, Michiels J. 2005. Defence of Rhizobium etli bacteroids against oxidative stress involves a complexly regulated atypical 2-Cys peroxiredoxin. Molecular Microbiology 55(4): 1207-1221.
Doyle JJ. 2011. Phylogenetic perspectives on the origins of nodulation. Molecular Plant-Microbe Interactions 24(11): 1289-1295.
E El Msehli S. 2012. Etude de l’interaction symbiotique Légumineuses-Rhizobium [Medicago truncatula-
Sinorhizobium meliloti] Sénescence nodositaire et glutathion. El Yahyaoui F, Küster H, Amor BB, Hohnjec N, Pühler A, Becker A, Gouzy J, Vernié T, Gough C,
Niebel A. 2004. Expression profiling in Medicago truncatula identifies more than 750 genes differentially expressed during nodulation, including many potential regulators of the symbiotic program. Plant Physiology 136(2): 3159-3176.
Engardt M, Simpson D, Schwikowski M, Granat L. 2017. Deposition of sulphur and nitrogen in Europe 1900–2050. Model calculations and comparison to historical observations. Tellus B: Chemical and Physical Meteorology 69(1): 1328945.
Espinosa‐Victoria D, Vance CP, Graham PH. 2000. Host variation in traits associated with crown nodule senescence in soybean. Crop Science 40(1): 103-109.

285
Eulgem T, Rushton PJ, Schmelzer E, Hahlbrock K, Somssich IE. 1999. Early nuclear events in plant defence signalling: rapid gene activation by WRKY transcription factors. The EMBO Journal 18(17): 4689-4699.
Evans PJ, Gallesi D, Mathieu C, Hernandez MJ, de Felipe M, Halliwell B, Puppo A. 1999. Oxidative stress occurs during soybean nodule senescence. Planta 208(1): 73-79.
F Farkas A, Maroti G, Durgo H, Gyorgypal Z, Lima RM, Medzihradszky KF, Kereszt A, Mergaert P,
Kondorosi E. 2014. Medicago truncatula symbiotic peptide NCR247 contributes to bacteroid differentiation through multiple mechanisms. Proceedings of the National Academy of Sciences of the United States of America 111(14): 5183-5188.
Favery B, Complainville A, Vinardell JM, Lecomte P, Vaubert D, Mergaert P, Kondorosi A, Kondorosi E, Crespi M, Abad P. 2002. The endosymbiosis-induced genes ENOD40 and CCS52a are involved in endoparasitic-nematode interactions in Medicago truncatula. Molecular Plant-Microbe Interactions 15(10): 1008-1013.
Fedorova M, van de Mortel J, Matsumoto PA, Cho J, Town CD, VandenBosch KA, Gantt JS, Vance CP. 2002. Genome-wide identification of nodule-specific transcripts in the model legume Medicago truncatula. Plant Physiology 130(2): 519-537.
Ferguson GP, Booth IR. 1998. Importance of glutathione for growth and survival of Escherichia coli cells: detoxification of methylglyoxal and maintenance of intracellular K+. Journal of Bacteriology 180(16): 4314-4318.
Fields S, Song O. 1989. A novel genetic system to detect protein-protein interactions. Nature 340(6230): 245-246.
Foley JA, Ramankutty N, Brauman KA, Cassidy ES, Gerber JS, Johnston M, Mueller ND, O'Connell C, Ray DK, West PC, et al. 2011. Solutions for a cultivated planet. Nature 478(7369): 337-342.
Fonouni-Farde C, Diet A, Frugier F. 2016a. Root Development and Endosymbioses: DELLAs Lead the Orchestra. Trends In Plant Science 21(11): 898-900.
Fonouni-Farde C, Kisiala A, Brault M, Emery RN, Diet A, Frugier F. 2017. DELLA1-gibberellin signaling regulates cytokinin-dependent symbiotic nodulation. Plant Physiology: pp. 00919.02017.
Fonouni-Farde C, Tan S, Baudin M, Brault M, Wen J, Mysore KS, Niebel A, Frugier F, Diet A. 2016b. DELLA-mediated gibberellin signalling regulates Nod factor signalling and rhizobial infection. Nature Communications 7: 12636.
Fowler D, Coyle M, Skiba U, Sutton MA, Cape JN, Reis S, Sheppard LJ, Jenkins A, Grizzetti B, Galloway JN. 2013. The global nitrogen cycle in the twenty-first century. Philosophical Transactions of the Royal Society B: Biological Sciences 368(1621): 20130164.
Foyer CH, Noctor G. 2011. Ascorbate and glutathione: the heart of the redox hub. Plant Physiology 155(1): 2-18.
Franche C, Lindström K, Elmerich C. 2009. Nitrogen-fixing bacteria associated with leguminous and non-leguminous plants. Plant and Soil 321(1-2): 35-59.
Franco-Zorrilla JM, Lopez-Vidriero I, Carrasco JL, Godoy M, Vera P, Solano R. 2014. DNA-binding specificities of plant transcription factors and their potential to define target genes. Proceedings of the National Academy of Sciences of the United States of America 111(6): 2367-2372.

286
Frendo P, Gallesi D, Turnbull R, Van de Sype G, Hérouart D, Puppo A. 1999. Localisation of glutathione and homoglutathione in Medicago truncatula is correlated to a differential expression of genes involved in their synthesis. The Plant Journal 17(2): 215-219.
Frendo P, Harrison J, Norman C, Hernandez Jimenez MJ, Van de Sype G, Gilabert A, Puppo A. 2005. Glutathione and homoglutathione play a critical role in the nodulation process of Medicago truncatula. Molecular Plant-Microbe Interactions 18(3): 254-259.
Frugier F, Poirier S, Satiat-Jeunemaître B, Kondorosi A, Crespi M. 2000. A Krüppel-like zinc finger protein is involved in nitrogen-fixing root nodule organogenesis. Genes & Development 14(4): 475-482.
Fujimoto SY, Ohta M, Usui A, Shinshi H, Ohme-Takagi M. 2000. Arabidopsis ethylene-responsive element binding factors act as transcriptional activators or repressors of GCC box–mediated gene expression. The Plant Cell 12(3): 393-404.
Fukudome M, Watanabe E, Osuki KI, Imaizumi R, Aoki T, Becana M, Uchiumi T. 2019. Stably Transformed Lotus japonicus Plants Overexpressing Phytoglobin LjGlb1-1 Show Decreased Nitric Oxide Levels in Roots and Nodules as Well as Delayed Nodule Senescence. Plant and Cell Physiology 60(4): 816-825.
Fuxman Bass JI, Reece-Hoyes JS, Walhout AJ. 2016. Gene-Centered Yeast One-Hybrid Assays. Cold Spring Harbor Protocols 2016(12): pdb.top077669.
G Galloway JN, Cowling EB. 2002. Reactive nitrogen and the world: 200 years of change. AMBIO: A
Journal of the Human Environment 31(2): 64-71. Galloway JN, Townsend AR, Erisman JW, Bekunda M, Cai Z, Freney JR, Martinelli LA, Seitzinger
SP, Sutton MA. 2008. Transformation of the nitrogen cycle: recent trends, questions, and potential solutions. Science 320(5878): 889-892.
Garcia J, Barker DG, Journet E-P. 2006. Seed storage and germination. In: The Medicago truncatula handbook. Mathesius U, Journet EP, Sumner LW (eds). ISBN 0-9754303-1-9. http://www.noble.org/MedicagoHandbook/
Gibbs DJ, Isa NM, Movahedi M, Lozano-Juste J, Mendiondo GM, Berckhan S, Marín-de la Rosa N, Conde JV, Correia CS, Pearce SP. 2014. Nitric oxide sensing in plants is mediated by proteolytic control of group VII ERF transcription factors. Molecular Cell 53(3): 369-379.
Gilroy EM, Hein I, van der Hoorn R, Boevink PC, Venter E, McLellan H, Kaffarnik F, Hrubikova K, Shaw J, Holeva M, et al. 2007. Involvement of cathepsin B in the plant disease resistance hypersensitive response. The Plant Journal 52(1): 1-13.
Giraud E, Moulin L, Vallenet D, Barbe V, Cytryn E, Avarre JC, Jaubert M, Simon D, Cartieaux F, Prin Y, et al. 2007. Legumes symbioses: absence of Nod genes in photosynthetic bradyrhizobia. Science 316(5829): 1307-1312.
Godiard L, Lepage A, Moreau S, Laporte D, Verdenaud M, Timmers T, Gamas P. 2011. MtbHLH1, a bHLH transcription factor involved in Medicago truncatula nodule vascular patterning and nodule to plant metabolic exchanges. New Phytologist 191(2): 391-404.
Godoy M, Franco-Zorrilla JM, Perez-Perez J, Oliveros JC, Lorenzo O, Solano R. 2011. Improven protein-binding microarrays for the identification of DNA-binding specificities of transcription factors. The Plant Journal 66(4): 700-711.
Gómez MD, Vera-Sirera F, Pérez-Amador MA. 2013. Molecular programme of senescence in dry and fleshy fruits. Journal of Experimental Botany 65(16): 4515-4526.

287
Gomez SK, Javot H, Deewatthanawong P, Torres-Jerez I, Tang Y, Blancaflor EB, Udvardi MK, Harrison MJ. 2009. Medicago truncatula and Glomus intraradices gene expression in cortical cells harboring arbuscules in the arbuscular mycorrhizal symbiosis. BMC Plant Biology 9(1): 10.
Gomi K, Sasaki A, Itoh H, Ueguchi-Tanaka M, Ashikari M, Kitano H, Matsuoka M. 2004. GID2, an F-box subunit of the SCF E3 complex, specifically interacts with phosphorylated SLR1 protein and regulates the gibberellin-dependent degradation of SLR1 in rice. The Plant Journal 37(4): 626-634.
Gonzalez-Rizzo S, Crespi M, Frugier F. 2006. The Medicago truncatula CRE1 cytokinin receptor regulates lateral root development and early symbiotic interaction with Sinorhizobium meliloti. The Plant Cell 18(10): 2680-2693.
Gourion B, Berrabah F, Ratet P, Stacey G. 2015. Rhizobium-legume symbioses: the crucial role of plant immunity. Trends In Plant Science 20(3): 186-194.
Gourion B, Bourcy M, Cosson V, Ratet P 2013. Protocols for growing plant symbioses; rhizobia. Plant Mineral Nutrients: Springer, 61-75.
Griffiths J, Murase K, Rieu I, Zentella R, Zhang ZL, Powers SJ, Gong F, Phillips AL, Hedden P, Sun TP, et al. 2006. Genetic characterization and functional analysis of the GID1 gibberellin receptors in Arabidopsis. The Plant Cell 18(12): 3399-3414.
Groten K, Dutilleul C, van Heerden PD, Vanacker H, Bernard S, Finkemeier I, Dietz KJ, Foyer CH. 2006. Redox regulation of peroxiredoxin and proteinases by ascorbate and thiols during pea root nodule senescence. FEBS Letters 580(5): 1269-1276.
Guan P, Ripoll JJ, Wang R, Vuong L, Bailey-Steinitz LJ, Ye D, Crawford NM. 2017. Interacting TCP and NLP transcription factors control plant responses to nitrate availability. Proceedings of the National Academy of Sciences of the United States of America 114(9): 2419-2424.
Guan P, Wang R, Nacry P, Breton G, Kay SA, Pruneda-Paz JL, Davani A, Crawford NM. 2014. Nitrate foraging by Arabidopsis roots is mediated by the transcription factor TCP20 through the systemic signaling pathway. Proceedings of the National Academy of Sciences of the United States of America 111(42): 15267-15272.
Guiboileau A, Sormani R, Meyer C, Masclaux-Daubresse C. 2010. Senescence and death of plant organs: nutrient recycling and developmental regulation. Comptes rendus biologies 333(4): 382-391.
Günther C, Schlereth A, Udvardi M, Ott T. 2007. Metabolism of reactive oxygen species is attenuated in leghaemoglobin-deficient nodules of Lotus japonicus. Molecular Plant-Microbe Interactions 20(12): 1596-1603.
Guo Y, Gan S. 2011. AtMYB2 regulates whole plant senescence by inhibiting cytokinin-mediated branching at late stages of development in Arabidopsis. Plant Physiology 156(3): 1612-1619.
H Haag AF, Baloban M, Sani M, Kerscher B, Pierre O, Farkas A, Longhi R, Boncompagni E,
Herouart D, Dall'angelo S, et al. 2011. Protection of Sinorhizobium against host cysteine-rich antimicrobial peptides is critical for symbiosis. PLOS Biology 9(10): e1001169.
Halliday J, Pate J. 1976. The acetylene reduction assay as a means of studying nitrogen fixation in white clover under sward and laboratory conditions. Grass and Forage Science 31(1): 29-35.
Hanaoka H, Noda T, Shirano Y, Kato T, Hayashi H, Shibata D, Tabata S, Ohsumi Y. 2002. Leaf senescence and starvation-induced chlorosis are accelerated by the disruption of an Arabidopsis autophagy gene. Plant Physiology 129(3): 1181-1193.

288
Harbison CT, Gordon DB, Lee TI, Rinaldi NJ, Macisaac KD, Danford TW, Hannett NM, Tagne JB, Reynolds DB, Yoo J, et al. 2004. Transcriptional regulatory code of a eukaryotic genome. Nature 431(7004): 99-104.
Harrison J, Jamet A, Muglia CI, Van de Sype G, Aguilar OM, Puppo A, Frendo P. 2005. Glutathione plays a fundamental role in growth and symbiotic capacity of Sinorhizobium meliloti. Journal of Bacteriology 187(1): 168-174.
Harrison J, Puppo A, Frendo P. 2003. The synthesis and the roles of glutathione and homoglutathione in legumes. Adv Plant Physiol 6: 385-412.
Harrison M, Dixon R. 1993. Isoflavonoid accumulation and expression of defence gene transcripts during the establishment of vesicular-arbuscular mycorrhizal associations in roots of Medicago truncatula. Molecular Plant-Microbe Interactions 6(5): 643-654.
Hartmann RM. 2018. Functional analysis of arbuscular mycorrhiza-related GRAS transcription factor genes of Medicago truncatula. (Doctoral dissertation, Hannover: Institutionelles Repositorium der Leibniz Universität Hannover).
He Y, Fukushige H, Hildebrand DF, Gan S. 2002. Evidence supporting a role of jasmonic acid in Arabidopsis leaf senescence. Plant Physiology 128(3): 876-884.
Heath MC 2000. Hypersensitive response-related death. Programmed cell death in higher plants: Springer, 77-90.
Hellman LM, Fried MG. 2007. Electrophoretic mobility shift assay (EMSA) for detecting protein-nucleic acid interactions. Nature Protocols 2(8): 1849-1861.
Hernández-Jiménez MJ, Mercedes Lucas M, de Felipe MR. 2002. Antioxidant defence and damage in senescing lupin nodules. Plant Physiology and Biochemistry 40(6-8): 645-657.
Hérouart D, Baudouin E, Frendo P, Harrison J, Santos R, Jamet A, Van de Sype G, Touati D, Puppo A. 2002. Reactive oxygen species, nitric oxide and glutathione: a key role in the establishment of the legume–Rhizobium symbiosis? Plant Physiology and Biochemistry 40(6-8): 619-624.
Herridge DF, Peoples MB, Boddey RM. 2008. Global inputs of biological nitrogen fixation in agricultural systems. Plant and Soil 311(1-2): 1-18.
Hichri I, Boscari A, Castella C, Rovere M, Puppo A, Brouquisse R. 2015. Nitric oxide: a multifaceted regulator of the nitrogen-fixing symbiosis. Journal of Experimental Botany 66(10): 2877-2887.
Higo K, Ugawa Y, Iwamoto M, Korenaga T. 1999. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Research 27(1): 297-300.
Hilliou F, Tran T. RqPCRBase: R package for analysis of quantitative real-time PCR data. Hirsch S, Kim J, Munoz A, Heckmann AB, Downie JA, Oldroyd GE. 2009. GRAS proteins form a
DNA binding complex to induce gene expression during nodulation signaling in Medicago truncatula. The Plant Cell 21(2): 545-557.
Hirsch S, Oldroyd GE. 2009. GRAS-domain transcription factors that regulate plant development. Plant signaling & behavior 4(8): 698-700.
Ho C-H, Lin S-H, Hu H-C, Tsay Y-F. 2009. CHL1 functions as a nitrate sensor in plants. Cell 138(6): 1184-1194.
Hogekamp C, Arndt D, Pereira PA, Becker JD, Hohnjec N, Kuster H. 2011. Laser microdissection unravels cell-type-specific transcription in arbuscular mycorrhizal roots, including CAAT-box transcription factor gene expression correlating with fungal contact and spread. Plant Physiology 157(4): 2023-2043.
Holsters M, Silva B, Van Vliet F, Genetello C, De Block M, Dhaese P, Depicker A, Inze D, Engler G, Villarroel R, et al. 1980. The functional organization of the nopaline A. tumefaciens plasmid pTiC58. Plasmid 3(2): 212-230.

289
Hondorp ER, Matthews RG. 2004. Oxidative stress inactivates cobalamin-independent methionine synthase (MetE) in Escherichia coli. PLOS Biology 2(11): e336.
Hopkins J, Pierre O, Kazmierczak T, Gruber V, Frugier F, Clement M, Frendo P, Herouart D, Boncompagni E. 2014. MtZR1, a PRAF protein, is involved in the development of roots and symbiotic root nodules in Medicago truncatula. Plant, Cell & Environment 37(3): 658-669.
Horchani F, Prévot M, Boscari A, Evangelisti E, Meilhoc E, Bruand C, Raymond P, Boncompagni E, Aschi-Smiti S, Puppo A. 2011. Both plant and bacterial nitrate reductases contribute to nitric oxide production in Medicago truncatula nitrogen-fixing nodules. Plant Physiology 155(2): 1023-1036.
I Innocenti G, Pucciariello C, Le Gleuher M, Hopkins J, de Stefano M, Delledonne M, Puppo A,
Baudouin E, Frendo P. 2007. Glutathione synthesis is regulated by nitric oxide in Medicago truncatula roots. Planta 225(6): 1597-1602.
Inoue H, Nojima H, Okayama H. 1990. High efficiency transformation of Escherichia coli with plasmids. Gene 96(1): 23-28.
J Jamet A, Sigaud S, Van de Sype G, Puppo A, Hérouart D. 2003. Expression of the bacterial catalase
genes during Sinorhizobium meliloti-Medicago sativa symbiosis and their crucial role during the infection process. Molecular Plant-Microbe Interactions 16(3): 217-225.
Jansson S, Thomas H. 2008. Senescence: developmental program or timetable? New Phytologist 179(3): 575-579.
Jefferson RA. 1987. Assaying chimeric genes in plants: the GUS gene fusion system. Plant Molecular Biology Reporter 5(4): 387-405.
Jenuwein T, Allis CD. 2001. Translating the histone code. Science 293(5532): 1074-1080. Jiang Y, Xie Q, Wang W, Yang J, Zhang X, Yu N, Zhou Y, Wang E. 2018. Medicago AP2-domain
transcription factor WRI5a is a master regulator of lipid biosynthesis and transfer during mycorrhizal symbiosis. Molecular Plant 11(11): 1344-1359.
Jin Y, Liu H, Luo D, Yu N, Dong W, Wang C, Zhang X, Dai H, Yang J, Wang E. 2016. DELLA proteins are common components of symbiotic rhizobial and mycorrhizal signalling pathways. Nature Communications 7(1): 12433.
Johnson DS, Mortazavi A, Myers RM, Wold B. 2007. Genome-wide mapping of in vivo protein-DNA interactions. Science 316(5830): 1497-1502.
Jongebloed U, Szederkenyi J, Hartig K, Schobert C, Komor E. 2004. Sequence of morphological and physiological events during natural ageing and senescence of a castor bean leaf: sieve tube occlusion and carbohydrate back-up precede chlorophyll degradation. Physiologia Plantarum 120(2): 338-346.
Journet EP, El-Gachtouli N, Vernoud V, de Billy F, Pichon M, Dedieu A, Arnould C, Morandi D, Barker DG, Gianinazzi-Pearson V. 2001. Medicago truncatula ENOD11: a novel RPRP-encoding early nodulin gene expressed during mycorrhization in arbuscule-containing cells. Molecular Plant-Microbe Interactions 14(6): 737-748.

290
K Kagaya Y, Ohmiya K, Hattori T. 1999. Regulation by external and developmental signals of the RAV2
gene encoding a DNA binding protein containing AP2 and B3-like domains. Plant Biotechnology 16(2): 135-140.
Kalo P, Gleason C, Edwards A, Marsh J, Mitra RM, Hirsch S, Jakab J, Sims S, Long SR, Rogers J, et al. 2005. Nodulation signaling in legumes requires NSP2, a member of the GRAS family of transcriptional regulators. Science 308(5729): 1786-1789.
Kardailsky IV, Brewin NJ. 1996. Expression of cysteine protease genes in pea nodule development and senescence. Molecular Plant-Microbe Interactions 9(8): 689-695.
Karimi M, Inzé D, Depicker A. 2002. GATEWAY™ vectors for Agrobacterium-mediated plant transformation. Trends In Plant Science 7(5): 193-195.
Kazmierczak T, Yang L, Boncompagni E, Meilhoc E, Frugier F, Frendo P, Bruand C, Gruber V, Brouquisse R 2020. Legume nodule senescence: a coordinated death mechanism between bacteria and plant cells. In: Frendo P, Frugier F, Masson-Boivin C eds. Regulation of Nitrogen-Fixing Symbioses in Legumes: Academic Press, 181-212.
Kevei Z, Lougnon G, Mergaert P, Horvath GV, Kereszt A, Jayaraman D, Zaman N, Marcel F, Regulski K, Kiss GB, et al. 2007. 3-hydroxy-3-methylglutaryl coenzyme a reductase 1 interacts with NORK and is crucial for nodulation in Medicago truncatula. The Plant Cell 19(12): 3974-3989.
Kevei Z, Vinardell JM, Kiss GB, Kondorosi A, Kondorosi E. 2002. Glycine-rich proteins encoded by a nodule-specific gene family are implicated in different stages of symbiotic nodule development in Medicago spp. Molecular Plant-Microbe Interactions 15(9): 922-931.
Khademi S, O'Connell J, Remis J, Robles-Colmenares Y, Miercke LJ, Stroud RM. 2004. Mechanism of ammonia transport by Amt/MEP/Rh: structure of AmtB at 1.35 Å. Science 305(5690): 1587-1594.
Kim JY, Park SJ, Jang B, Jung CH, Ahn SJ, Goh CH, Cho K, Han O, Kang H. 2007. Functional characterization of a glycine-rich RNA-binding protein 2 in Arabidopsis thaliana under abiotic stress conditions. The Plant Journal 50(3): 439-451.
Kimura M. 1968. Evolutionary rate at the molecular level. Nature 217(5129): 624-626. Kirkwood TB, Melov S. 2011. On the programmed/non-programmed nature of ageing within the life
history. Current Biology 21(18): R701-R707. Kliszczak AE, Rainey MD, Harhen B, Boisvert FM, Santocanale C. 2011. DNA mediated chromatin
pull-down for the study of chromatin replication. Scientific Reports 1: 95. Kobayashi H, Naciri-Graven Y, Broughton WJ, Perret X. 2004. Flavonoids induce temporal shifts in
gene-expression of nod-box controlled loci in Rhizobium sp. NGR234. Molecular Microbiology 51(2): 335-347.
Kondorosi A, Kiss G, Dusha I 1984. Plasmids governing symbiotic nitrogen fixation. Current Developments in Biological Nitrogen Fixation: Oxford/IBH New Delhi/Bombay/Calcutta, 135-171.
Kondorosi E, Gyuris J, Schmidt J, John M, Duda E, Hoffmann B, Schell J, Kondorosi A. 1989. Positive and negative control of nod gene expression in Rhizobium meliloti is required for optimal nodulation. The EMBO Journal 8(5): 1331-1340.
Kondorosi E, Kondorosi A. 1986. Nodule induction on plant roots by Rhizobium. Trends In Biochemical Sciences 11(7): 296-299.
Kondorosi E, Mergaert P, Kereszt A. 2013. A paradigm for endosymbiotic life: cell differentiation of Rhizobium bacteria provoked by host plant factors. Annual Review of Microbiology 67: 611-628.

291
Konishi M, Yanagisawa S. 2013. Arabidopsis NIN-like transcription factors have a central role in nitrate signalling. Nature Communications 4(1): 1-9.
Korkuc P, Schippers JH, Walther D. 2014. Characterization and identification of cis-regulatory elements in Arabidopsis based on single-nucleotide polymorphism information. Plant Physiology 164(1): 181-200.
Kosmacz M, Parlanti S, Schwarzländer M, Kragler F, Licausi F, Van Dongen JT. 2015. The stability and nuclear localization of the transcription factor RAP 2.12 are dynamically regulated by oxygen concentration. Plant, Cell & Environment 38(6): 1094-1103.
Koyama T. 2014. The roles of ethylene and transcription factors in the regulation of onset of leaf senescence. Frontiers in Plant Science 5: 650.
Kundu M, Thompson CB. 2005. Macroautophagy versus mitochondrial autophagy: a question of fate? Cell Death and Differentiation 12: 1484-1489.
L
Laloum T, Baudin M, Frances L, Lepage A, Billault-Penneteau B, Cerri MR, Ariel F, Jardinaud MF,
Gamas P, de Carvalho-Niebel F, et al. 2014. Two CCAAT-box-binding transcription factors redundantly regulate early steps of the legume-rhizobia endosymbiosis. The Plant Journal 79(5): 757-768.
Laloum T, De Mita S, Gamas P, Baudin M, Niebel A. 2013. CCAAT-box binding transcription factors in plants: Y so many? Trends In Plant Science 18(3): 157-166.
Lamb C, Dixon RA. 1997. The Oxidative Burst in Plant Disease Resistance. Annual Review Of Plant Physiology and Plant Molecular Biology 48(1): 251-275.
Lang C, Long SR. 2015. Transcriptomic Analysis of Sinorhizobium meliloti and Medicago truncatula Symbiosis Using Nitrogen Fixation-Deficient Nodules. Molecular Plant-Microbe Interactions 28(8): 856-868.
Laporte P, Lepage A, Fournier J, Catrice O, Moreau S, Jardinaud MF, Mun JH, Larrainzar E, Cook DR, Gamas P, et al. 2014. The CCAAT box-binding transcription factor NF-YA1 controls rhizobial infection. Journal of Experimental Botany 65(2): 481-494.
Laub MT, Chen SL, Shapiro L, McAdams HH. 2002. Genes directly controlled by CtrA, a master regulator of the Caulobacter cell cycle. Proceedings of the National Academy of Sciences of the United States of America 99(7): 4632-4637.
Laub MT, McAdams HH, Feldblyum T, Fraser CM, Shapiro L. 2000. Global analysis of the genetic network controlling a bacterial cell cycle. Science 290(5499): 2144-2148.
Lawn RJ, Brun WA. 1974. Symbiotic Nitrogen Fixation in Soybeans. I. Effect of Photosynthetic Source‐Sink Manipulations 1. Crop Science 14(1): 11-16.
Lee TI, Rinaldi NJ, Robert F, Odom DT, Bar-Joseph Z, Gerber GK, Hannett NM, Harbison CT, Thompson CM, Simon I, et al. 2002. Transcriptional regulatory networks in Saccharomyces cerevisiae. Science 298(5594): 799-804.
Legocki RP, Verma DP. 1979. A nodule-specific plant protein (nodulin-35) from soybean. Science 205(4402): 190-193.
Legocki RP, Verma DPS. 1980. Identification of “nodule-specific” host proteins (nodulins) involved in the development of Rhizobium-Legume symbiosis. Cell 20(1): 153-163.
León J, Costa-Broseta Á, Castillo MC. 2020. RAP2. 3 negatively regulates nitric oxide biosynthesis and related responses through a rheostat-like mechanism in Arabidopsis. Journal of Experimental Botany 71(10): 3157-3171.

292
Lescot M, Dehais P, Thijs G, Marchal K, Moreau Y, Van de Peer Y, Rouze P, Rombauts S. 2002. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Research 30(1): 325-327.
Li JJ, Herskowitz I. 1993. Isolation of ORC6, a component of the yeast origin recognition complex by a one-hybrid system. Science 262(5141): 1870-1874.
Li JY, Fu YL, Pike SM, Bao J, Tian W, Zhang Y, Chen CZ, Zhang Y, Li HM, Huang J, et al. 2010. The Arabidopsis nitrate transporter NRT1.8 functions in nitrate removal from the xylem sap and mediates cadmium tolerance. The Plant Cell 22(5): 1633-1646.
Li X, Feng H, Wen J, Dong J, Wang T. 2018. MtCAS31 Aids Symbiotic Nitrogen Fixation by Protecting the Leghemoglobin MtLb120-1 Under Drought Stress in Medicago truncatula. Frontiers in Plant Science 9: 633.
Li Y, Zhou L, Li Y, Chen D, Tan X, Lei L, Zhou J. 2008. A nodule‐specific plant cysteine proteinase, AsNODF32, is involved in nodule senescence and nitrogen fixation activity of the green manure legume Astragalus sinicus. New Phytologist 180(1): 185-192.
Libault M, Joshi T, Benedito VA, Xu D, Udvardi MK, Stacey G. 2009. Legume transcription factor genes: what makes legumes so special? Plant Physiology 151(3): 991-1001.
Lifton R, Goldberg M, Karp R, Hogness D 1978. The organization of the histone genes in Drosophila melanogaster: functional and evolutionary implications. Cold Spring Harbor Symposia On Quantitative Biology: Cold Spring Harbor Laboratory Press. 1047-1051.
Lillig CH, Potamitou A, Schwenn JD, Vlamis-Gardikas A, Holmgren A. 2003. Redox regulation of 3'-phosphoadenylylsulfate reductase from Escherichia coli by glutathione and glutaredoxins. Journal Of Biological Chemistry 278(25): 22325-22330.
Lim PO, Kim HJ, Nam HG. 2007. Leaf senescence. Annual Review of Plant Biology 58: 115-136. Lima JE, Kojima S, Takahashi H, von Wiren N. 2010. Ammonium triggers lateral root branching in
Arabidopsis in an AMMONIUM TRANSPORTER1;3-dependent manner. The Plant Cell 22(11): 3621-3633.
Lin JS, Li X, Luo Z, Mysore KS, Wen J, Xie F. 2018. Author Correction: NIN interacts with NLPs to mediate nitrate inhibition of nodulation in Medicago truncatula. Nature Plants 4(12): 1125.
Lin SH, Kuo HF, Canivenc G, Lin CS, Lepetit M, Hsu PK, Tillard P, Lin HL, Wang YY, Tsai CB, et al. 2008. Mutation of the Arabidopsis NRT1.5 nitrate transporter causes defective root-to-shoot nitrate transport. The Plant Cell 20(9): 2514-2528.
Lipuma J, Cinege G, Bodogai M, Olah B, Kiers A, Endre G, Dupont L, Dusha I. 2014. A vapBC-type toxin-antitoxin module of Sinorhizobium meliloti influences symbiotic efficiency and nodule senescence of Medicago sativa. Environmental Microbiology 16(12): 3714-3729.
Liu H, Hu M, Wang Q, Cheng L, Zhang Z. 2018. Role of Papain-Like Cysteine Proteases in Plant Development. Frontiers in Plant Science 9: 1717.
Liu J, Müller B 2017. Imaging TCSn:: GFP, a synthetic cytokinin reporter, in Arabidopsis thaliana. Plant Hormones: Springer, 81-90.
Liu J, Rutten L, Limpens E, van der Molen T, van Velzen R, Chen R, Chen Y, Geurts R, Kohlen W, Kulikova O, et al. 2019. A Remote cis-Regulatory Region Is Required for NIN Expression in the Pericycle to Initiate Nodule Primordium Formation in Medicago truncatula. The Plant Cell 31(1): 68-83.
Liu J, Wilson TE, Milbrandt J, Johnston M. 1993. Identifying DNA-binding sites and analyzing DNA-binding domains using a yeast selection system. Methods 5(2): 125-137.
Liu X, Zhang Y, Chen Y, Li M, Zhou F, Li K, Cao H, Ni M, Liu Y, Gu Z, et al. 2017. In Situ Capture of Chromatin Interactions by Biotinylated dCas9. Cell 170(5): 1028-1043 e1019.
Livak KJ, Schmittgen TD. 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25(4): 402-408.

293
Lohman KN, Gan S, John MC, Amasino RM. 1994. Molecular analysis of natural leaf senescence in Arabidopsis thaliana. Physiologia Plantarum 92(2): 322-328.
Lopez-Otin C, Blasco MA, Partridge L, Serrano M, Kroemer G. 2013. The hallmarks of aging. Cell 153(6): 1194-1217.
Loque D, Lalonde S, Looger LL, von Wiren N, Frommer WB. 2007. A cytosolic trans-activation domain essential for ammonium uptake. Nature 446(7132): 195-198.
Lorenzo O, Piqueras R, Sánchez-Serrano JJ, Solano R. 2003. ETHYLENE RESPONSE FACTOR1 integrates signals from ethylene and jasmonate pathways in plant defence. The Plant Cell 15(1): 165-178.
Loscos J, Matamoros MA, Becana M. 2008. Ascorbate and homoglutathione metabolism in common bean nodules under stress conditions and during natural senescence. Plant Physiology 146(3): 1282-1292.
Lu SC. 2009. Regulation of glutathione synthesis. Molecular aspects of medicine 30(1-2): 42-59.
M
Madsen EB, Madsen LH, Radutoiu S, Olbryt M, Rakwalska M, Szczyglowski K, Sato S, Kaneko T,
Tabata S, Sandal N. 2003. A receptor kinase gene of the LysM type is involved in legume perception of rhizobial signals. Nature 425(6958): 637-640.
Malik NS, Pfeiffer NE, Williams DR, Wagner FW. 1981. Peptidohydrolases of Soybean Root Nodules : IDENTIFICATION, SEPARATION, AND PARTIAL CHARACTERIZATION OF ENZYMES FROM BACTEROID-FREE EXTRACTS. Plant Physiology 68(2): 386-392.
Mangeon A, Junqueira RM, Sachetto-Martins G. 2010. Functional diversity of the plant glycine-rich proteins superfamily. Plant Signaling & Behavior 5(2): 99-104.
Marino D, Andrio E, Danchin EG, Oger E, Gucciardo S, Lambert A, Puppo A, Pauly N. 2011. A Medicago truncatula NADPH oxidase is involved in symbiotic nodule functioning. New Phytologist 189(2): 580-592.
Maróti G, Downie JA, Kondorosi É. 2015. Plant cysteine-rich peptides that inhibit pathogen growth and control rhizobial differentiation in legume nodules. Current Opinion In Plant Biology 26: 57-63.
Marquez-Garcia B, Shaw D, Cooper JW, Karpinska B, Quain MD, Makgopa EM, Kunert K, Foyer CH. 2015. Redox markers for drought-induced nodule senescence, a process occurring after drought-induced senescence of the lowest leaves in soybean (Glycine max). Annals of Botany 116(4): 497-510.
Marschner H. 2011. Marschner's mineral nutrition of higher plants: Academic press. Marsh JF, Rakocevic A, Mitra RM, Brocard L, Sun J, Eschstruth A, Long SR, Schultze M, Ratet
P, Oldroyd GE. 2007. Medicago truncatula NIN is essential for rhizobial-independent nodule organogenesis induced by autoactive calcium/calmodulin-dependent protein kinase. Plant Physiology 144(1): 324-335.
Martínez DE, Costa L, Guiamét JJ 2018. Activities of vacuolar cysteine proteases in plant senescence. Plant Senescence: Springer, 283-297.
Maruyama Y, Yamoto N, Suzuki Y, Chiba Y, Yamazaki K, Sato T, Yamaguchi J. 2013. The Arabidopsis transcriptional repressor ERF9 participates in resistance against necrotrophic fungi. Plant Science 213: 79-87.

294
Masclaux-Daubresse C, Daniel-Vedele F, Dechorgnat J, Chardon F, Gaufichon L, Suzuki A. 2010. Nitrogen uptake, assimilation and remobilization in plants: challenges for sustainable and productive agriculture. Annals of Botany 105(7): 1141-1157.
Masip L, Veeravalli K, Georgiou G. 2006. The many faces of glutathione in bacteria. Antioxidants & redox signaling 8(5-6): 753-762.
Masson-Boivin C, Giraud E, Perret X, Batut J. 2009. Establishing nitrogen-fixing symbiosis with legumes: how many rhizobium recipes? Trends In Microbiology 17(10): 458-466.
Maston GA, Evans SK, Green MR. 2006. Transcriptional regulatory elements in the human genome. Annual Review of Genomics and Human Genetics 7: 29-59.
Matamoros MA, Baird LM, Escuredo PR, Dalton DA, Minchin FR, Iturbe-Ormaetxe I, Rubio MC, Moran JF, Gordon AJ, Becana M. 1999. Stress-induced legume root nodule senescence. Physiological, biochemical, and structural alterations. Plant Physiology 121(1): 97-112.
Matamoros MA, Dalton DA, Ramos J, Clemente MR, Rubio MC, Becana M. 2003. Biochemistry and molecular biology of antioxidants in the rhizobia-legume symbiosis. Plant Physiology 133(2): 499-509.
Maunoury N, Redondo-Nieto M, Bourcy M, Van de Velde W, Alunni B, Laporte P, Durand P, Agier N, Marisa L, Vaubert D, et al. 2010. Differentiation of symbiotic cells and endosymbionts in Medicago truncatula nodulation are coupled to two transcriptome-switches. PLOS One 5(3): e9519.
McFadden GI, van Dooren GG. 2004. Evolution: red algal genome affirms a common origin of all plastids. Current Biology 14(13): R514-R516.
McGinnis KM, Thomas SG, Soule JD, Strader LC, Zale JM, Sun TP, Steber CM. 2003. The Arabidopsis SLEEPY1 gene encodes a putative F-box subunit of an SCF E3 ubiquitin ligase. The Plant Cell 15(5): 1120-1130.
Meilhoc E, Blanquet P, Cam Y, Bruand C. 2013. Control of NO level in rhizobium-legume root nodules: not only a plant globin story. Plant Signaling & Behavior 8(10): doi: 10 4161/psb 25923.
Merbach W, Schilling G. 1980. Effectiveness of symbiotic N2-fixation in leguminous plants, as affected by inoculation with rhizobia, by substrate, n-fertilizing, and 14c-sucrose application (author's transl). Zentralblatt fur Bakteriologie, Parasitenkunde, Infektionskrankheiten und Hygiene. Zweite Naturwissenschaftliche Abteilung: Mikrobiologie der Landwirtschaft der Technologie und des Umweltschutzes 135(2): 99-118.
Mergaert P, Nikovics K, Kelemen Z, Maunoury N, Vaubert D, Kondorosi A, Kondorosi E. 2003. A novel family in Medicago truncatula consisting of more than 300 nodule-specific genes coding for small, secreted polypeptides with conserved cysteine motifs. Plant Physiology 132(1): 161-173.
Mergaert P, Uchiumi T, Alunni B, Evanno G, Cheron A, Catrice O, Mausset AE, Barloy-Hubler F, Galibert F, Kondorosi A, et al. 2006. Eukaryotic control on bacterial cell cycle and differentiation in the Rhizobium-legume symbiosis. Proceedings of the National Academy of Sciences of the United States of America 103(13): 5230-5235.
Messinese E, Mun JH, Yeun LH, Jayaraman D, Rouge P, Barre A, Lougnon G, Schornack S, Bono JJ, Cook DR, et al. 2007. A novel nuclear protein interacts with the symbiotic DMI3 calcium- and calmodulin-dependent protein kinase of Medicago truncatula. Molecular Plant-Microbe Interactions 20(8): 912-921.
Miller AJ, Fan X, Orsel M, Smith SJ, Wells DM. 2007. Nitrate transport and signalling. Journal of Experimental Botany 58(9): 2297-2306.
Milo R, Shen-Orr S, Itzkovitz S, Kashtan N, Chklovskii D, Alon U. 2002. Network motifs: simple building blocks of complex networks. Science 298(5594): 824-827.

295
Minina EA, Filonova LH, Fukada K, Savenkov EI, Gogvadze V, Clapham D, Sanchez-Vera V, Suarez MF, Zhivotovsky B, Daniel G, et al. 2013. Autophagy and metacaspase determine the mode of cell death in plants. Journal Of Cell Biology 203(6): 917-927.
Misas‐Villamil JC, van der Hoorn RA, Doehlemann G. 2016. Papain‐like cysteine proteases as hubs in plant immunity. New Phytologist 212(4): 902-907.
Mitchell PJ, Tjian R. 1989. Transcriptional regulation in mammalian cells by sequence-specific DNA binding proteins. Science 245(4916): 371-378.
Mitra RM, Shaw SL, Long SR. 2004. Six nonnodulating plant mutants defective for Nod factor-induced transcriptional changes associated with the legume-rhizobia symbiosis. Proceedings of the National Academy of Sciences of the United States of America 101(27): 10217-10222.
Molina A, Mena M, Carbonero P, Garcia-Olmedo F. 1997. Differential expression of pathogen-responsive genes encoding two types of glycine-rich proteins in barley. Plant Molecular Biology 33(5): 803-810.
Molina C, Grotewold E. 2005. Genome wide analysis of Arabidopsis core promoters. BMC genomics 6(1): 25.
Moling S, Pietraszewska-Bogiel A, Postma M, Fedorova E, Hink MA, Limpens E, Gadella TW, Bisseling T. 2014. Nod factor receptors form heteromeric complexes and are essential for intracellular infection in Medicago nodules. The Plant Cell 26(10): 4188-4199.
Moore B, Zhou L, Rolland F, Hall Q, Cheng WH, Liu YX, Hwang I, Jones T, Sheen J. 2003. Role of the Arabidopsis glucose sensor HXK1 in nutrient, light, and hormonal signaling. Science 300(5617): 332-336.
Mueller ND, Gerber JS, Johnston M, Ray DK, Ramankutty N, Foley JA. 2012. Closing yield gaps through nutrient and water management. Nature 490(7419): 254-257.
Muglia C, Comai G, Spegazzini E, Riccillo PM, Aguilar OM. 2008. Glutathione produced by Rhizobium tropici is important to prevent early senescence in common bean nodules. Fems Microbiology Letters 286(2): 191-198.
Muglia CI, Grasso DH, Aguilar OM. 2007. Rhizobium tropici response to acidity involves activation of glutathione synthesis. Microbiology 153(Pt 4): 1286-1296.
Muller B, Sheen J. 2008. Cytokinin and auxin interaction in root stem-cell specification during early embryogenesis. Nature 453(7198): 1094-1097.
Muller M, Munne-Bosch S. 2015. Ethylene Response Factors: A Key Regulatory Hub in Hormone and Stress Signaling. Plant Physiology 169(1): 32-41.
N Naito Y, Fujie M, Usami S, Murooka Y, Yamada T. 2000. The involvement of a cysteine proteinase in
the nodule development in Chinese milk vetch infected with Mesorhizobium huakuii subsp. rengei. Plant Physiology 124(3): 1087-1096.
Nakajima A, Aono T, Tsukada S, Siarot L, Ogawa T, Oyaizu H. 2012. Lon protease of Azorhizobium caulinodans ORS571 is required for suppression of reb gene expression. Appl Environ Microbiol 78(17): 6251-6261.
Nakano T, Suzuki K, Fujimura T, Shinshi H. 2006. Genome-Wide Analysis of the ERF Gene Family in Arabidopsis and Rice. Plant Physiology 140(2): 411-432.
Nakaune S, Yamada K, Kondo M, Kato T, Tabata S, Nishimura M, Hara-Nishimura I. 2005. A vacuolar processing enzyme, δVPE, is involved in seed coat formation at the early stage of seed development. The Plant Cell 17(3): 876-887.

296
Nelson JD, Denisenko O, Bomsztyk K. 2006. Protocol for the fast chromatin immunoprecipitation (ChIP) method. Nature Protocols 1(1): 179-185.
Neuhäuser B, Dynowski M, Mayer M, Ludewig U. 2007. Regulation of NH4+ transport by essential cross talk between AMT monomers through the carboxyl tails. Plant Physiology 143(4): 1651-1659.
Nishida H, Tanaka S, Handa Y, Ito M, Sakamoto Y, Matsunaga S, Betsuyaku S, Miura K, Soyano T, Kawaguchi M, et al. 2018. A NIN-LIKE PROTEIN mediates nitrate-induced control of root nodule symbiosis in Lotus japonicus. Nature Communications 9(1): 499.
Noctor G, Foyer CH. 1998. ASCORBATE AND GLUTATHIONE: Keeping Active Oxygen Under Control. Annual Review Of Plant Physiology and Plant Molecular Biology 49(1): 249-279.
Noctor G, Gomez L, Vanacker H, Foyer CH. 2002. Interactions between biosynthesis, compartmentation and transport in the control of glutathione homeostasis and signalling. Journal of Experimental Botany 53(372): 1283-1304.
Nomoto M, Tada Y, Tsukagoshi H. 2019. In vitro Protein-DNA Binding Assay (AlphaScreen® Technology). BIO-PROTOCOL 9(3).
Noonan JP, McCallion AS. 2010. Genomics of long-range regulatory elements. Annual Review of Genomics and Human Genetics 11: 1-23.
O O'Brien JA, Vega A, Bouguyon E, Krouk G, Gojon A, Coruzzi G, Gutierrez RA. 2016. Nitrate
Transport, Sensing, and Responses in Plants. Molecular Plant 9(6): 837-856. Ohme-Takagi M, Shinshi H. 1990. Structure and expression of a tobacco beta-1,3-glucanase gene.
Plant Molecular Biology 15(6): 941-946. Ohme-Takagi M, Shinshi H. 1995. Ethylene-inducible DNA binding proteins that interact with an
ethylene-responsive element. The Plant Cell 7(2): 173-182. Okazaki S, Kaneko T, Sato S, Saeki K. 2013. Hijacking of leguminous nodulation signaling by the
rhizobial type III secretion system. Proceedings of the National Academy of Sciences of the United States of America 110(42): 17131-17136.
Oldroyd GE. 2013. Speak, friend, and enter: signalling systems that promote beneficial symbiotic associations in plants. Nature Reviews Microbiology 11(4): 252-263.
Oldroyd GE, Dixon R. 2014. Biotechnological solutions to the nitrogen problem. Current Opinion In Biotechnology 26: 19-24.
Oldroyd GE, Downie JA. 2004. Calcium, kinases and nodulation signalling in legumes. Nature Reviews Molecular Cell Biology 5(7): 566-576.
Oldroyd GE, Downie JA. 2006. Nuclear calcium changes at the core of symbiosis signalling. Current Opinion In Plant Biology 9(4): 351-357.
Oldroyd GE, Murray JD, Poole PS, Downie JA. 2011. The rules of engagement in the legume-rhizobial symbiosis. Annual Review of Genetics 45: 119-144.
Oldroyd GED, Leyser O. 2020. A plant's diet, surviving in a variable nutrient environment. Science 368(6486): eaba0196.
Olvera-Carrillo Y, Van Bel M, Van Hautegem T, Fendrych M, Huysmans M, Simaskova M, van Durme M, Buscaill P, Rivas S, Coll NS, et al. 2015. A Conserved Core of Programmed Cell Death Indicator Genes Discriminates Developmentally and Environmentally Induced Programmed Cell Death in Plants. Plant Physiology 169(4): 2684-2699.

297
Ortenberg R, Gon S, Porat A, Beckwith J. 2004. Interactions of glutaredoxins, ribonucleotide reductase, and components of the DNA replication system of Escherichia coli. Proceedings of the National Academy of Sciences of the United States of America 101(19): 7439-7444.
Otegui MS, Noh YS, Martinez DE, Vila Petroff MG, Staehelin LA, Amasino RM, Guiamet JJ. 2005. Senescence-associated vacuoles with intense proteolytic activity develop in leaves of Arabidopsis and soybean. The Plant Journal 41(6): 831-844.
Ott T, van Dongen JT, Gunther C, Krusell L, Desbrosses G, Vigeolas H, Bock V, Czechowski T, Geigenberger P, Udvardi MK. 2005. Symbiotic leghemoglobins are crucial for nitrogen fixation in legume root nodules but not for general plant growth and development. Current Biology 15(6): 531-535.
P Park SH, Jeong JS, Seo JS, Park BS, Chua NH. 2019. Arabidopsis ubiquitin-specific proteases
UBP12 and UBP13 shape ORE1 levels during leaf senescence induced by nitrogen deficiency. New Phytologist 223(3): 1447-1460.
Pate JS. 1973. Uptake, assimilation and transport of nitrogen compounds by plants. Soil Biology and Biochemistry 5(1): 109-119.
Pauly N, Pucciariello C, Mandon K, Innocenti G, Jamet A, Baudouin E, Herouart D, Frendo P, Puppo A. 2006. Reactive oxygen and nitrogen species and glutathione: key players in the legume-Rhizobium symbiosis. Journal of Experimental Botany 57(8): 1769-1776.
Pecrix Y, Staton SE, Sallet E, Lelandais-Briere C, Moreau S, Carrere S, Blein T, Jardinaud MF, Latrasse D, Zouine M, et al. 2018. Whole-genome landscape of Medicago truncatula symbiotic genes. Nature Plants 4(12): 1017-1025.
Peiter E, Schubert S. 2003. Sugar uptake and proton release by protoplasts from the infected zone of Vicia faba L. nodules: evidence against apoplastic sugar supply of infected cells. Journal of Experimental Botany 54(388): 1691-1700.
Penmetsa RV, Cook DR. 2000. Production and characterization of diverse developmental mutants of Medicago truncatula. Plant Physiology 123(4): 1387-1398.
Penninckx MJ, Elskens MT 1993. Metabolism and functions of glutathione in micro-organisms. Advances In Microbial Physiology: Elsevier, 239-301.
Perez Guerra JC, Coussens G, De Keyser A, De Rycke R, De Bodt S, Van De Velde W, Goormachtig S, Holsters M. 2010. Comparison of developmental and stress-induced nodule senescence in Medicago truncatula. Plant Physiology 152(3): 1574-1584.
Perez-Llorca M, Munoz P, Muller M, Munne-Bosch S. 2019. Biosynthesis, Metabolism and Function of Auxin, Salicylic Acid and Melatonin in Climacteric and Non-climacteric Fruits. Frontiers in Plant Science 10: 136.
Phukan UJ, Jeena GS, Tripathi V, Shukla RK. 2017. Regulation of Apetala2/Ethylene Response Factors in Plants. Frontiers in Plant Science 8: 150.
Pierre O, Hopkins J, Combier M, Baldacci F, Engler G, Brouquisse R, Herouart D, Boncompagni E. 2014. Involvement of papain and legumain proteinase in the senescence process of Medicago truncatula nodules. New Phytologist 202(3): 849-863.
Pini F, De Nisco NJ, Ferri L, Penterman J, Fioravanti A, Brilli M, Mengoni A, Bazzicalupo M, Viollier PH, Walker GC, et al. 2015. Cell Cycle Control by the Master Regulator CtrA in Sinorhizobium meliloti. PLOS Genetics 11(5): e1005232.
Pislariu CI, Murray JD, Wen J, Cosson V, Muni RR, Wang M, Benedito VA, Andriankaja A, Cheng X, Jerez IT, et al. 2012. A Medicago truncatula tobacco retrotransposon insertion mutant

298
collection with defects in nodule development and symbiotic nitrogen fixation. Plant Physiology 159(4): 1686-1699.
Pladys D, Dimitrijevic L, Rigaud J. 1991. Localization of a protease in protoplast preparations in infected cells of French bean nodules. Plant Physiology 97(3): 1174-1180.
Pladys D, Vance CP. 1993. Proteolysis during Development and Senescence of Effective and Plant Gene-Controlled Ineffective Alfalfa Nodules. Plant Physiology 103(2): 379-384.
Poole P, Ramachandran V, Terpolilli J. 2018. Rhizobia: from saprophytes to endosymbionts. Nature Reviews Microbiology 16(5): 291.
Ptashne M, Gann A. 1997. Transcriptional activation by recruitment. Nature 386(6625): 569-577. Puppo A, Groten K, Bastian F, Carzaniga R, Soussi M, Lucas MM, de Felipe MR, Harrison J,
Vanacker H, Foyer CH. 2005. Legume nodule senescence: roles for redox and hormone signalling in the orchestration of the natural aging process. New Phytologist 165(3): 683-701.
Q Qin C, Qian W, Wang W, Wu Y, Yu C, Jiang X, Wang D, Wu P. 2008. GDP-mannose
pyrophosphorylase is a genetic determinant of ammonium sensitivity in Arabidopsis thaliana. Proceedings of the National Academy of Sciences 105(47): 18308-18313.
Quandt HJ. 1993. Transgenic Root Nodules of Vicia hirsuta: A Fast and Efficient System for the Study of Gene Expression in Indeterminate-Type Nodules. Molecular Plant-Microbe Interactions 6(6): 699-706.
R Radutoiu S, Madsen LH, Madsen EB, Felle HH, Umehara Y, Gronlund M, Sato S, Nakamura Y,
Tabata S, Sandal N, et al. 2003. Plant recognition of symbiotic bacteria requires two LysM receptor-like kinases. Nature 425(6958): 585-592.
Radutoiu S, Madsen LH, Madsen EB, Jurkiewicz A, Fukai E, Quistgaard EM, Albrektsen AS, James EK, Thirup S, Stougaard J. 2007. LysM domains mediate lipochitin-oligosaccharide recognition and Nfr genes extend the symbiotic host range. The EMBO Journal 26(17): 3923-3935.
Ramu SK, Peng H-M, Cook DR. 2002. Nod factor induction of reactive oxygen species production is correlated with expression of the early nodulin gene rip1 in Medicago truncatula. Molecular Plant-Microbe Interactions 15(6): 522-528.
Rawlings ND, Barrett AJ, Thomas PD, Huang X, Bateman A, Finn RD. 2018. The MEROPS database of proteolytic enzymes, their substrates and inhibitors in 2017 and a comparison with peptidases in the PANTHER database. Nucleic Acids Research 46(D1): D624-D632.
Reay DS, Dentener F, Smith P, Grace J, Feely RA. 2008. Global nitrogen deposition and carbon sinks. Nature Geoscience 1(7): 430-437.
Remans T, Nacry P, Pervent M, Filleur S, Diatloff E, Mounier E, Tillard P, Forde BG, Gojon A. 2006. The Arabidopsis NRT1.1 transporter participates in the signaling pathway triggering root colonization of nitrate-rich patches. Proceedings of the National Academy of Sciences of the United States of America 103(50): 19206-19211.

299
Ren B, Robert F, Wyrick JJ, Aparicio O, Jennings EG, Simon I, Zeitlinger J, Schreiber J, Hannett N, Kanin E, et al. 2000. Genome-wide location and function of DNA binding proteins. Science 290(5500): 2306-2309.
Riccillo PM, Muglia CI, de Bruijn FJ, Roe AJ, Booth IR, Aguilar OM. 2000. Glutathione is involved in environmental stress responses in Rhizobium tropici, including acid tolerance. Journal of Bacteriology 182(6): 1748-1753.
Riechmann JL. 2018. Transcription factors of Arabidopsis and rice: a genomic perspective. Annual Plant Reviews online: 28-53.
Riechmann JL, Heard J, Martin G, Reuber L, Jiang C, Keddie J, Adam L, Pineda O, Ratcliffe OJ, Samaha RR, et al. 2000. Arabidopsis transcription factors: genome-wide comparative analysis among eukaryotes. Science 290(5499): 2105-2110.
Ritz D, Beckwith J. 2001. Roles of thiol-redox pathways in bacteria. Annual Review of Microbiology 55(1): 21-48.
Rolland F, Baena-Gonzalez E, Sheen J. 2006. Sugar sensing and signaling in plants: conserved and novel mechanisms. Annual Review of Plant Biology 57: 675-709.
Rook F, Gerrits N, Kortstee A, van Kampen M, Borrias M, Weisbeek P, Smeekens S. 1998. Sucrose-specific signalling represses translation of the Arabidopsis ATB2 bZIP transcription factor gene. The Plant Journal 15(2): 253-263.
Roponen I. 1970. The Effect of Darkness on the Leghemoglobin Content and Amino Acid Levels in the Root Nodules of Pea Plants. Physiologia Plantarum 23(3): 452-460.
Rosenberg C, Boistard P, Denarie J, Casse-Delbart F. 1981. Genes controlling early and late functions in symbiosis are located on a megaplasmid in Rhizobium meliloti. Molecular and General Genetics 184(2): 326-333.
Rostas K, Kondorosi E, Horvath B, Simoncsits A, Kondorosi A. 1986. Conservation of extended promoter regions of nodulation genes in Rhizobium. Proceedings of the National Academy of Sciences of the United States of America 83(6): 1757-1761.
Roux B, Rodde N, Jardinaud MF, Timmers T, Sauviac L, Cottret L, Carrère S, Sallet E, Courcelle E, Moreau S. 2014. An integrated analysis of plant and bacterial gene expression in symbiotic root nodules using laser‐capture microdissection coupled to RNA sequencing. The Plant Journal 77(6): 817-837.
Rovere M. 2018. ERF-VII family as key players in hypoxic signaling and adaptation in Medicago truncatula. Doctoral dissertation, Université Côte d'Azur.
Roy S, Liu W, Nandety RS, Crook A, Mysore KS, Pislariu CI, Frugoli J, Dickstein R, Udvardi MK. 2020. Celebrating 20 Years of Genetic Discoveries in Legume Nodulation and Symbiotic Nitrogen Fixation. The Plant Cell 32(1): 15-41.
Rubin GM, Yandell MD, Wortman JR, Gabor Miklos GL, Nelson CR, Hariharan IK, Fortini ME, Li PW, Apweiler R, Fleischmann W, et al. 2000. Comparative genomics of the eukaryotes. Science 287(5461): 2204-2215.
Ruvkun GB, Sundaresan V, Ausubel FM. 1982. Directed transposon Tn5 mutagenesis and complementation analysis of Rhizobium meliloti symbiotic nitrogen fixation genes. Cell 29(2): 551-559.
S Sachetto-Martins G, Franco LO, de Oliveira DE. 2000. Plant glycine-rich proteins: a family or just
proteins with a common motif? Biochimica et Biophysica Acta 1492(1): 1-14.

300
Santos R, Herouart D, Puppo A, Touati D. 2000. Critical protective role of bacterial superoxide dismutase in rhizobium-legume symbiosis. Molecular Microbiology 38(4): 750-759.
Santos R, Herouart D, Sigaud S, Touati D, Puppo A. 2001. Oxidative burst in alfalfa-Sinorhizobium meliloti symbiotic interaction. Molecular Plant-Microbe Interactions 14(1): 86-89.
Sarwat M, Naqvi AR, Ahmad P, Ashraf M, Akram NA. 2013. Phytohormones and microRNAs as sensors and regulators of leaf senescence: assigning macro roles to small molecules. Biotechnology Advances 31(8): 1153-1171.
Scandalios JG, Guan L, Polidoros AN. 1997. Catalases in plants: gene structure, properties, regulation, and expression. Cold Spring Harbor Monograph Series 34: 343-406.
Schauser L, Roussis A, Stiller J, Stougaard J. 1999. A plant regulator controlling development of symbiotic root nodules. Nature 402(6758): 191-195.
Schindelin J, Arganda-Carreras I, Frise E, Kaynig V, Longair M, Pietzsch T, Preibisch S, Rueden C, Saalfeld S, Schmid B, et al. 2012. Fiji: an open-source platform for biological-image analysis. Nature Methods 9(7): 676-682.
Schultze M, Kondorosi A. 1998. Regulation of symbiotic root nodule development. Annual Review of Genetics 32(1): 33-57.
Seabra AR, Pereira PA, Becker JD, Carvalho HG. 2012. Inhibition of glutamine synthetase by phosphinothricin leads to transcriptome reprograming in root nodules of Medicago truncatula. Molecular Plant-Microbe Interactions 25(7): 976-992.
Seefeldt LC, Hoffman BM, Dean DR. 2009. Mechanism of Mo-dependent nitrogenase. Annual Review of Biochemistry 78(1): 701-722.
Serova TA, Tikhonovich IA, Tsyganov VE. 2017. Analysis of nodule senescence in pea (Pisum sativum L.) using laser microdissection, real-time PCR, and ACC immunolocalization. Journal of Plant Physiology 212: 29-44.
Sevin-Pujol A, Sicard M, Rosenberg C, Auriac MC, Lepage A, Niebel A, Gough C, Bensmihen S. 2017. Development of a GAL4-VP16/UAS trans-activation system for tissue specific expression in Medicago truncatula. PLOS One 12(11): e0188923.
Seymour GB, Ostergaard L, Chapman NH, Knapp S, Martin C. 2013. Fruit development and ripening. Annual Review of Plant Biology 64: 219-241.
Shen-Orr SS, Milo R, Mangan S, Alon U. 2002. Network motifs in the transcriptional regulation network of Escherichia coli. Nature Genetics 31(1): 64-68.
Sheokand S, Brewin NJ. 2003. Cysteine proteases in nodulation and nitrogen fixation. Indian Journal of Experimental Biology 41(10): 1124-1132.
Shindo T, Misas-Villamil JC, Horger AC, Song J, van der Hoorn RA. 2012. A role in immunity for Arabidopsis cysteine protease RD21, the ortholog of the tomato immune protease C14. PLOS One 7(1): e29317.
Shirsat A, Wilford N, Croy R, Boulter D. 1989. Sequences responsible for the tissue specific promoter activity of a pea legumin gene in tobacco. Molecular and General Genetics 215(2): 326-331.
Showalter AM, Zhou J, Rumeau D, Worst SG, Varner JE. 1991. Tomato extensin and extensin-like cDNAs: structure and expression in response to wounding. Plant Molecular Biology 16(4): 547-565.
Shu K, Zhou W, Yang W. 2018. APETALA 2-domain-containing transcription factors: focusing on abscisic acid and gibberellins antagonism. New Phytologist 217(3): 977-983.
Shu Y, Liu Y, Zhang J, Song L, Guo C. 2015. Genome-Wide Analysis of the AP2/ERF Superfamily Genes and their Responses to Abiotic Stress in Medicago truncatula. Frontiers in Plant Science 6: 1247.

301
Sigaud S, Becquet V, Frendo P, Puppo A, Hérouart D. 1999. Differential regulation of two divergent Sinorhizobium meliloti genes for HPII-like catalases during free-living growth and protective role of both catalases during symbiosis. Journal of Bacteriology 181(8): 2634-2639.
Singh S, Giri MK, Singh PK, Siddiqui A, Nandi AK. 2013. Down-regulation of OsSAG12-1 results in enhanced senescence and pathogen-induced cell death in transgenic rice plants. Journal Of Biosciences 38(3): 583-592.
Singh S, Katzer K, Lambert J, Cerri M, Parniske M. 2014. CYCLOPS, a DNA-binding transcriptional activator, orchestrates symbiotic root nodule development. Cell Host Microbe 15(2): 139-152.
Singh S, Parniske M. 2012. Activation of calcium- and calmodulin-dependent protein kinase (CCaMK), the central regulator of plant root endosymbiosis. Current Opinion In Plant Biology 15(4): 444-453.
Sinharoy S, Torres-Jerez I, Bandyopadhyay K, Kereszt A, Pislariu CI, Nakashima J, Benedito VA, Kondorosi E, Udvardi MK. 2013. The C2H2 transcription factor regulator of symbiosome differentiation represses transcription of the secretory pathway gene VAMP721a and promotes symbiosome development in Medicago truncatula. The Plant Cell 25(9): 3584-3601.
Smale ST, Kadonaga JT. 2003. The RNA polymerase II core promoter. Annual Review of Biochemistry 72(1): 449-479.
Smeekens S, Ma J, Hanson J, Rolland F. 2010. Sugar signals and molecular networks controlling plant growth. Current Opinion In Plant Biology 13(3): 274-279.
Smit P, Raedts J, Portyanko V, Debelle F, Gough C, Bisseling T, Geurts R. 2005. NSP1 of the GRAS protein family is essential for rhizobial Nod factor-induced transcription. Science 308(5729): 1789-1791.
Somssich IE, Schmelzer E, Kawalleck P, Hahlbrock K. 1988. Gene structure and in situ transcript localization of pathogenesis-related protein 1 in parsley. Molecular and General Genetics 213(1): 93-98.
Soyano T, Hirakawa H, Sato S, Hayashi M, Kawaguchi M. 2014. Nodule Inception creates a long-distance negative feedback loop involved in homeostatic regulation of nodule organ production. Proceedings of the National Academy of Sciences of the United States of America 111(40): 14607-14612.
Soyano T, Kouchi H, Hirota A, Hayashi M. 2013. Nodule inception directly targets NF-Y subunit genes to regulate essential processes of root nodule development in Lotus japonicus. PLOS Genetics 9(3): e1003352.
Stein LY, Klotz MG. 2016. The nitrogen cycle. Current Biology 26(3): R94-98. Stevens CJ. 2019. Nitrogen in the environment. Science 363(6427): 578-580. Stockinger EJ, Gilmour SJ, Thomashow MF. 1997. Arabidopsis thaliana CBF1 encodes an AP2
domain-containing transcriptional activator that binds to the C-repeat/DRE, a cis-acting DNA regulatory element that stimulates transcription in response to low temperature and water deficit. Proceedings of the National Academy of Sciences 94(3): 1035-1040.
Storz G, Tartaglia LA. 1992. OxyR: a regulator of antioxidant genes. Journal of Nutrition 122(3 Suppl): 627-630.
Strahl BD, Allis CD. 2000. The language of covalent histone modifications. Nature 403(6765): 41-45. Suzuki H, Kumagai H, Tochikura T. 1986. gamma-Glutamyltranspeptidase from Escherichia coli K-
12: purification and properties. Journal of Bacteriology 168(3): 1325-1331. Swaraj K, Sheokand S, Fernandez-Pascual MM, de Felipe MR. 2001. Dark-induced changes in
legume nodule functioning. Functional Plant Biology 28(6): 429-438.

302
T Tadege M, Wen J, He J, Tu H, Kwak Y, Eschstruth A, Cayrel A, Endre G, Zhao PX, Chabaud M, et
al. 2008. Large-scale insertional mutagenesis using the Tnt1 retrotransposon in the model legume Medicago truncatula. The Plant Journal 54(2): 335-347.
Taiz L, Zeiger E, Møller IM, Murphy A. 2015. Plant physiology and development. Tang H, Krishnakumar V, Bidwell S, Rosen B, Chan A, Zhou S, Gentzbittel L, Childs KL, Yandell
M, Gundlach H, et al. 2014. An improven genome release (version Mt4.0) for the model legume Medicago truncatula. BMC genomics 15(1): 312.
Tate R, Cermola M, Riccio A, Diez-Roux G, Patriarca EJ. 2012. Glutathione is required by Rhizobium etli for glutamine utilization and symbiotic effectiveness. Molecular Plant-Microbe Interactions 25(3): 331-340.
Thomas H. 2013. Senescence, ageing and death of the whole plant. New Phytologist 197(3): 696-711. Thomas H, Huang L, Young M, Ougham H. 2009. Evolution of plant senescence. BMC Evolutionary
Biology 9(1): 163. Timmers AC, Soupene E, Auriac MC, de Billy F, Vasse J, Boistard P, Truchet G. 2000. Saprophytic
intracellular rhizobia in alfalfa nodules. Molecular Plant-Microbe Interactions 13(11): 1204-1213. Tjepkema JD, Yocum CS. 1974. Measurement of oxygen partial pressure within soybean nodules by
oxygen microelectrodes. Planta 119(4): 351-360. Tóth K, Stacey G. 2015. Does plant immunity play a critical role during initiation of the legume-
rhizobium symbiosis? Frontiers in Plant Science 6: 401. Tsay YF, Chiu CC, Tsai CB, Ho CH, Hsu PK. 2007. Nitrate transporters and peptide transporters.
FEBS Letters 581(12): 2290-2300.
U Uchiumi T, Ohwada T, Itakura M, Mitsui H, Nukui N, Dawadi P, Kaneko T, Tabata S, Yokoyama T,
Tejima K, et al. 2004. Expression islands clustered on the symbiosis island of the Mesorhizobium loti genome. Journal of Bacteriology 186(8): 2439-2448.
Udvardi MK, Kakar K, Wandrey M, Montanari O, Murray J, Andriankaja A, Zhang JY, Benedito V, Hofer JM, Chueng F, et al. 2007. Legume transcription factors: global regulators of plant development and response to the environment. Plant Physiology 144(2): 538-549.
V Van de Velde W, Guerra JC, De Keyser A, De Rycke R, Rombauts S, Maunoury N, Mergaert P,
Kondorosi E, Holsters M, Goormachtig S. 2006. Aging in legume symbiosis. A molecular view on nodule senescence in Medicago truncatula. Plant Physiology 141(2): 711-720.

303
Van de Velde W, Zehirov G, Szatmari A, Debreczeny M, Ishihara H, Kevei Z, Farkas A, Mikulass K, Nagy A, Tiricz H, et al. 2010. Plant peptides govern terminal differentiation of bacteria in symbiosis. Science 327(5969): 1122-1126.
van Doorn WG, Woltering EJ. 2004. Senescence and programmed cell death: substance or semantics? Journal of Experimental Botany 55(406): 2147-2153.
Van Larebeke N, Engler G, Holsters M, Van den Elsacker S, Zaenen I, Schilperoort RA, Schell J. 1974. Large plasmid in Agrobacterium tumefaciens essential for crown gall-inducing ability. Nature 252(5479): 169-170.
van Wyk SG, Du Plessis M, Cullis CA, Kunert KJ, Vorster BJ. 2014. Cysteine protease and cystatin expression and activity during soybean nodule development and senescence. BMC Plant Biology 14(1): 294.
Varma Penmetsa R, Uribe P, Anderson J, Lichtenzveig J, Gish JC, Nam YW, Engstrom E, Xu K, Sckisel G, Pereira M. 2008. The Medicago truncatula ortholog of Arabidopsis EIN2, sickle, is a negative regulator of symbiotic and pathogenic microbial associations. The Plant Journal 55(4): 580-595.
Vasse J, De Billy F, Camut S, Truchet G. 1990. Correlation between ultrastructural differentiation of bacteroids and nitrogen fixation in alfalfa nodules. Journal of Bacteriology 172(8): 4295-4306.
Vauclare P, Bligny R, Gout E, De Meuron V, Widmer F. 2010. Metabolic and structural rearrangement during dark-induced autophagy in soybean (Glycine max L.) nodules: an electron microscopy and 31P and 13C nuclear magnetic resonance study. Planta 231(6): 1495-1504.
Vernié T, Kim J, Frances L, Ding Y, Sun J, Guan D, Niebel A, Gifford ML, de Carvalho-Niebel F, Oldroyd GE. 2015. The NIN Transcription Factor Coordinates Diverse Nodulation Programs in Different Tissues of the Medicago truncatula Root. The Plant Cell 27(12): 3410-3424.
Vernié T, Moreau S, De Billy F, Plet J, Combier J-P, Rogers C, Oldroyd G, Frugier F, Niebel A, Gamas P. 2008. EFD is an ERF transcription factor involved in the control of nodule number and differentiation in Medicago truncatula. The Plant Cell Online 20(10): 2696-2713.
Vinardell JM, Fedorova E, Cebolla A, Kevei Z, Horvath G, Kelemen Z, Tarayre S, Roudier F,
Mergaert P, Kondorosi A. 2003. Endoreduplication mediated by the anaphase-promoting complex activator CCS52A is required for symbiotic cell differentiation in Medicago truncatula nodules. The Plant Cell 15(9): 2093-2105.
Vincent JL, Brewin NJ. 2000. Immunolocalization of a cysteine protease in vacuoles, vesicles, and symbiosomes of pea nodule cells. Plant Physiology 123(2): 521-530.
W Wang D, Griffitts J, Starker C, Fedorova E, Limpens E, Ivanov S, Bisseling T, Long S. 2010. A
nodule-specific protein secretory pathway required for nitrogen-fixing symbiosis. Science 327(5969): 1126-1129.
Wang H, Wang H, Liu R, Xu Y, Lu Z, Zhou C. 2018. Genome-Wide Identification of TCP Family Transcription Factors in Medicago truncatula Reveals Significant Roles of miR319-Targeted TCPs in Nodule Development. Frontiers in Plant Science 9(774): 774.
Wang MM, Reed RR. 1993. Molecular cloning of the olfactory neuronal transcription factor Olf-1 by genetic selection in yeast. Nature 364(6433): 121-126.
Wang Q, Yang S, Liu J, Terecskei K, Abraham E, Gombar A, Domonkos A, Szucs A, Kormoczi P, Wang T, et al. 2017. Host-secreted antimicrobial peptide enforces symbiotic selectivity in

304
Medicago truncatula. Proceedings of the National Academy of Sciences of the United States of America 114(26): 6854-6859.
Wang R, Guan P, Chen M, Xing X, Zhang Y, Crawford NM. 2010. Multiple regulatory elements in the Arabidopsis NIA1 promoter act synergistically to form a nitrate enhancer. Plant Physiology 154(1): 423-432.
Wang XC, Wu J, Guan ML, Zhao CH, Geng P, Zhao Q. 2020. Arabidopsis MYB4 plays dual roles in flavonoid biosynthesis. The Plant Journal 101(3): 637-652.
Wang Y-Y, Tsay Y-F. 2011. Arabidopsis nitrate transporter NRT1. 9 is important in phloem nitrate transport. The Plant Cell 23(5): 1945-1957.
Wasserman WW, Sandelin A. 2004. Applied bioinformatics for the identification of regulatory elements. Nature Reviews Genetics 5(4): 276-287.
Wheeler GL, Jones MA, Smirnoff N. 1998. The biosynthetic pathway of vitamin C in higher plants. Nature 393(6683): 365-369.
Wingler A, Roitsch T. 2008. Metabolic regulation of leaf senescence: interactions of sugar signalling with biotic and abiotic stress responses. Plant Biology 10: 50-62.
Wobbes B. 2004. Control of plant carbohydrate partitioning by the Arabidopsis thaliana ATB2 bZIP transcription factor gene. Dissertation, Utrecht University Repository.
Wu KK 2006. Analysis of protein-DNA binding by streptavidin-agarose pulldown. Gene Mapping, Discovery, and Expression: Springer, 281-290.
Wu R, Duan L, Pruneda-Paz JL, Oh DH, Pound M, Kay S, Dinneny JR. 2018. The 6xABRE Synthetic Promoter Enables the Spatiotemporal Analysis of ABA-Mediated Transcriptional Regulation. Plant Physiology 177(4): 1650-1665.
X Xie F, Murray JD, Kim J, Heckmann AB, Edwards A, Oldroyd GE, Downie JA. 2012. Legume
pectate lyase required for root infection by rhizobia. Proceedings of the National Academy of Sciences of the United States of America 109(2): 633-638.
Xie Z, Nolan TM, Jiang H, Yin Y. 2019. AP2/ERF Transcription Factor Regulatory Networks in Hormone and Abiotic Stress Responses in Arabidopsis. Frontiers in Plant Science 10: 228.
Xie Z, Zhang ZL, Zou X, Huang J, Ruas P, Thompson D, Shen QJ. 2005. Annotations and functional analyses of the rice WRKY gene superfamily reveal positive and negative regulators of abscisic acid signaling in aleurone cells. Plant Physiology 137(1): 176-189.
Xu G, Fan X, Miller AJ. 2012. Plant nitrogen assimilation and use efficiency. Annual Review of Plant Biology 63: 153-182.
Y Yamaguchi-Shinozaki K, Shinozaki K. 1994. A novel cis-acting element in an Arabidopsis gene is
involved in responsiveness to drought, low-temperature, or high-salt stress. The Plant Cell 6(2): 251-264.
Yanagisawa S. 2000. Dof1 and Dof2 transcription factors are associated with expression of multiple genes involved in carbon metabolism in maize. The Plant Journal 21(3): 281-288.

305
Yanagisawa S, Schmidt RJ. 1999. Diversity and similarity among recognition sequences of Dof transcription factors. The Plant Journal 17(2): 209-214.
Yang L, El Msehli S, Benyamina S, Lambert A, Hopkins J, Cazareth J, Pierre O, Herouart D, Achi-Smiti S, Boncompagni E, et al. 2020. Glutathione Deficiency in Sinorhizobium meliloti Does Not Impair Bacteroid Differentiation But Induces Early Senescence in the Interaction With Medicago truncatula. Frontiers in Plant Science 11(137): 137.
Yang L, Syska C, Garcia I, Frendo P, Boncompagni E 2020. Involvement of proteases during nodule senescence in leguminous plants. In: Bruijn Fd ed. The Model Legume Medicago truncatula, 683-693.
Yano K, Yoshida S, Muller J, Singh S, Banba M, Vickers K, Markmann K, White C, Schuller B, Sato S, et al. 2008. CYCLOPS, a mediator of symbiotic intracellular accommodation. Proceedings of the National Academy of Sciences of the United States of America 105(51): 20540-20545.
Yoro E, Suzaki T, Toyokura K, Miyazawa H, Fukaki H, Kawaguchi M. 2014. A positive regulator of nodule organogenesis, NODULE INCEPTION, acts as a negative regulator of rhizobial infection in Lotus japonicus. Plant Physiology 165(2): 747-758.
Young ND, Debelle F, Oldroyd GE, Geurts R, Cannon SB, Udvardi MK, Benedito VA, Mayer KF, Gouzy J, Schoof H, et al. 2011. The Medicago genome provides insight into the evolution of rhizobial symbioses. Nature 480(7378): 520-524.
Z Zahran HH. 1999. Rhizobium-legume symbiosis and nitrogen fixation under severe conditions and in
an arid climate. Microbiology and Molecular Biology Reviews 63(4): 968-989. Zanetti ME, Ripodas C, Niebel A. 2017. Plant NF-Y transcription factors: Key players in plant-microbe
interactions, root development and adaptation to stress. Biochimica et Biophysica Acta 1860(5): 645-654.
Zhang H, Forde BG. 1998. An Arabidopsis MADS box gene that controls nutrient-induced changes in root architecture. Science 279(5349): 407-409.
Zhang Y. 2003. Transcriptional regulation by histone ubiquitination and deubiquitination. Genes & Development 17(22): 2733-2740.
Zhang Z, Hu X, Zhang Y, Miao Z, Xie C, Meng X, Deng J, Wen J, Mysore KS, Frugier F, et al. 2016. Opposing Control by Transcription Factors MYB61 and MYB3 Increases Freezing Tolerance by Relieving C-Repeat Binding Factor Suppression. Plant Physiology 172(2): 1306-1323.
Zhang ZL, Xie Z, Zou X, Casaretto J, Ho TH, Shen QJ. 2004. A rice WRKY gene encodes a transcriptional repressor of the gibberellin signaling pathway in aleurone cells. Plant Physiology 134(4): 1500-1513.
Zhao S, Fung-Leung WP, Bittner A, Ngo K, Liu X. 2014. Comparison of RNA-Seq and microarray in transcriptome profiling of activated T cells. PLOS One 9(1): e78644.
Zheng M, Aslund F, Storz G. 1998. Activation of the OxyR transcription factor by reversible disulfide bond formation. Science 279(5357): 1718-1721.
Zhou C, Han L, Pislariu C, Nakashima J, Fu C, Jiang Q, Quan L, Blancaflor EB, Tang Y, Bouton JH. 2011. From model to crop: functional analysis of a STAY-GREEN gene in the model legume Medicago truncatula and effective use of the gene for alfalfa improvement. Plant Physiology 157(3): 1483-1496.
Zhou H, Zhao J, Cai J, Patil SB. 2017. UBIQUITIN-SPECIFIC PROTEASES function in plant development and stress responses. Plant Molecular Biology 94(6): 565-576.

306
Zipfel C, Oldroyd GE. 2017. Plant signalling in symbiosis and immunity. Nature 543(7645): 328-336. Zurcher E, Tavor-Deslex D, Lituiev D, Enkerli K, Tarr PT, Muller B. 2013. A robust and sensitive
synthetic sensor to monitor the transcriptional output of the cytokinin signaling network in planta. Plant Physiology 161(3): 1066-1075.

307
The first of the line is tied to a tree
and the last is being eaten by the ants.
Gabriel García Márquez

308