ziz originagriculturemesoamerica
DESCRIPTION
Origen de agricultura en MesoamericaTRANSCRIPT

7/17/2019 Ziz Originagriculturemesoamerica
http://slidepdf.com/reader/full/ziz-originagriculturemesoamerica 1/13
R E S E A R C H A R T I C L E
Origin of agriculture and plant domestication
in West MesoamericaDaniel Zizumbo-Villarreal • Patricia Colunga-GarcıaMarın
Received: 24 April 2009 / Accepted: 14 December 2009 / Published online: 2 February 2010
Springer Science+Business Media B.V. 2010
Abstract Recent paleoecological, archaeobotanical
and genetic-molecular data are used to develop a
hypothesis on the where, when, how and whom of
plant domestication and the origin of agriculture in
west Mesoamerica, and the formation of the maize-
bean-squash multicrop milpa system and agro-food
system which formed the base for development of
ancient complex societies in this area. It is highly
likely that about 10,000 before present (BP) human
groups specializing in plant gathering and small game
hunting in the dry tropical forest of the Balsas-Jaliscobiotic morphotectonic province began the process of
plant domestication and agriculture, using fire as a
tool. Sympatric distribution of the putative wild
ancestral populations of maize, beans and squash
indicate the extreme northwest Balsas-Jalisco region
as a possible locus of domestication. Diffusion of
these domesticates to the rest of Mesoamerica would
have occurred via existing biological-cultural corri-
dors. The milpa agro-food system would have been
established between 7,000 and 4,400 calendar years
(cal) BP. The complex food technology developed inthe northwest Balsas-Jalisco region between 4,500
and 3,500 BP, much more complex than in other
areas at the time, also suggests this area as the origin
of the milpa agro-food system. Further archaeobo-
tanical research is needed to confirm this hypothesis.
Exploratory, collection and conservation efforts are
needed in these putative source populations, as well
as studies on their adaptation to climatic, edaphic and
biotic factors, before they are displaced by the
African grasses and pesticides forming part of the
region’s growing cattle industry.
Keywords Agriculture Beans Domestication
Maize
Mesoamerica
Squash
Introduction
One of the most significant events in human history
was the transformation from a hunting-gathering
economy to an agricultural economy (Smith 2005).
This change probably occurred independently in at
least six regions in the world, primarily tropical and
subtropical areas with high biological and cultural
diversity (Gepts 2008; Piperno and Pearsall 1998;Sauer 1952). Along with the Middle East and north
China, Mesoamerica is one of the world’s primary
centers of domestication (Harlan 1971, 1995). It was
here that species such as maize ( Zea mays L.), beans
(Phaseolus spp.) and squash (Cucurbita spp.) were
domesticated and integrated into a multi-crop system
known in the region today as milpa. It was this
system’s ecological and nutritional complementarity
that helped to support the development of highly
D. Zizumbo-Villarreal (&) P. Colunga-GarcıaMarın
Unidad de Recursos Naturales, Centro de Investigacion
Cientıfica de Yucatan, Calle 43 No 130. Col. Chuburna de
Hidalgo, 97200 Merida, Yucatan, Mexico
e-mail: [email protected]
1 3
Genet Resour Crop Evol (2010) 57:813–825
DOI 10.1007/s10722-009-9521-4

7/17/2019 Ziz Originagriculturemesoamerica
http://slidepdf.com/reader/full/ziz-originagriculturemesoamerica 2/13
complex societies in Mesoamerica (Gepts 2008).
Mesoamerica is most commonly defined as the
cultural area extending from the Santiago and Panuco
river basins in Mexico, south to the Central Valley
region of Costa Rica (Kirchhoff 1943).
Plant domestication is understood as the continuous
historic evolutionary process, driven by human selec-tion, which produces fixation of a set of alleles that
provides wild populations with favorable human
consumption and cultivation phenotypes, but which
also diminishes or eliminates the capacity for survival
under natural conditions, making a domesticated
population dependent on humans (Harlan 1992). The
set of phenotypic traits determined by these alleles is
known as the domestication syndrome (Hammer
1984). Plants’ reproductive systems and genetic con-
stitution have favored and hindered domestication to
varying degrees. Plants that have responded positivelyto the selection process and agricultural management
have been completely domesticated, while in others
only certain traits of the syndrome have been fixed,
leaving them semi-domesticated or in process (Gepts
2004). In agricultural systems under limiting environ-
mental conditions, humans have managed to maintain
some species in a semi-domesticated state as part of
their productive strategy (Colunga-GarcıaMarın and
Zizumbo-Villarreal 1993).
For the purposes of this discussion, agricultural
management is to be understood as a set of deliberate,environmental modifications made by humans aimed
at increasing survival and biomass production in
selected plant species, and therefore meeting human
needs given prevailing environmental conditions.
Plant domestication and agriculture are temporally
continuous and interdependent processes. Over time,
humans have selected a group of local and introduced
species for subsistence, and developed knowledge
and techniques for their cultivation, transport, stor-
age, transformation, consumption and preservation.
This group of species and the knowledge andtechniques associated with them are known as an
agro-food system.
The present study aim was to integrate current
paleoecological, archaeobotanical and genetic-molec-
ular data to develop hypotheses on the where, when,
how and whom of plant domestication and agricul-
tural development in west Mesoamerica; by what
routes these domesticates and related knowledge and
technology were distributed; and how and where the
milpa multicrop system and associated agro-food
system came into existence. The answers to these
questions will be vital to locating the genetic
diversity nuclei so important to productivity, sustain-
ability and improvement in current agro-food sys-
tems, and to better understanding the material
foundations of complex cultures in Mesoamerica.
Initial human occupation
Human occupation of Mesoamerica began about
11,600 years before present (BP) (Dixon 1999).
These partially sedentary early arrivals survived by
fishing, hunting megafauna and collecting plants in
areas near the highland intermontane lake systems of
Chapala, Sayula-Zacoalco, Zacapu, Cuitzeo, Mexico
and Puebla-Valsequillo, (Aliphat 1980; Irish et al.2000; Lorenzo and Mirabell 1986; Niederberger
1979; Tolstoy et al. 1977). Their principal hunting
implement was initially designed like a harpoon used
for hunting marine mammals, suggesting that they
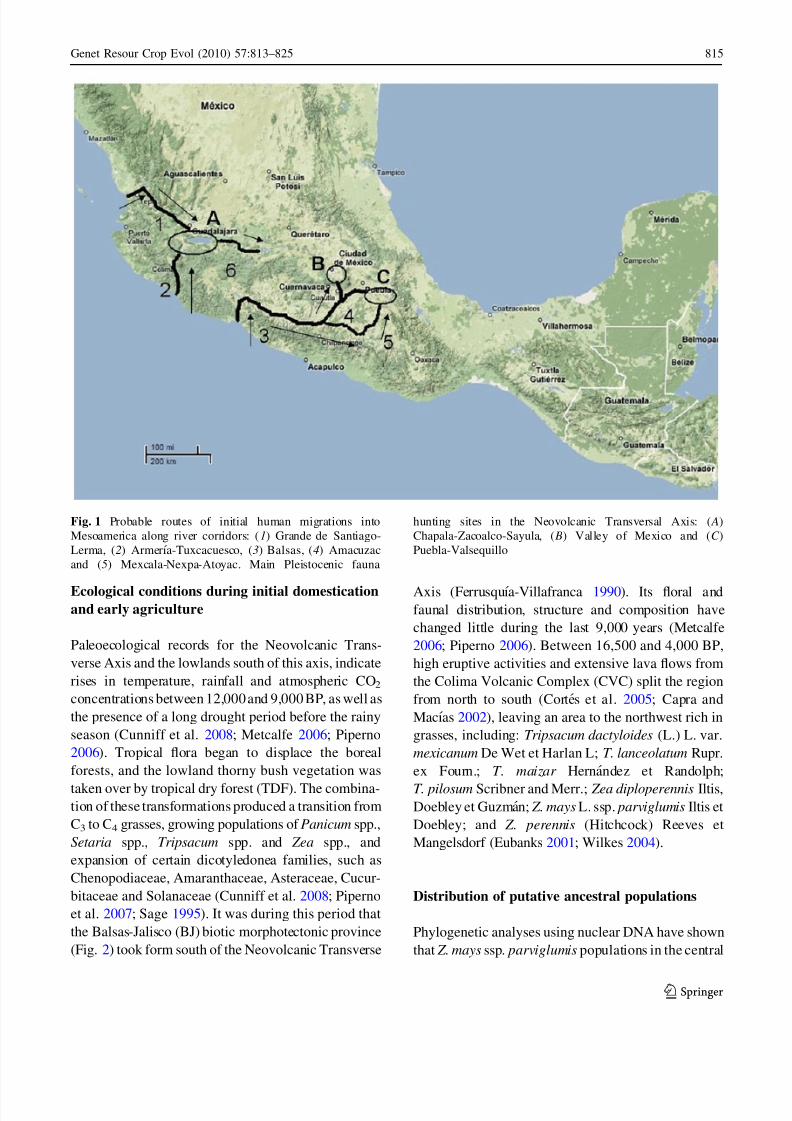
had moved inland from the Pacific coast (Fig. 1).
A following migration into the area probably
occurred around 10,600 BP (Dixon 2001) from the
west and great plains of North America. These groups
implemented an ecological strategy based on small
game hunting and plant gathering. They processed
some plant species to make them edible or extractcertain elements; for instance, ground seeds from
Panicum spp. and Setaria spp.; cooked stems and leaf
bases from Agave spp.; stems and fresh fruits from
Opuntia spp.; seeds from Quercus spp. and Pinus
spp.; and fruits from Prosopis spp. They used
sharpened scrapers and points for gathering and
hunting, and stone technology for breaking, grinding
and cooking (Doebley 1984; Poinar et al. 2001;
Willis 1995). In addition to their use of fire, they
introduced the dog (Canis familiaris L.), domesti-
cated from multiple lineages in eastern Asia (Leonardet al. 2002; Wayne et al. 2006). Use of dogs allowed
them to form small, highly mobile bands with
defensive capacity. They occupied rock shelters
along the banks of rivers communicating the inter-
montane valleys with the west coast. Technology
among these groups included thin bifaces, flint points,
river pebbles and stone blocks or platforms modified
for grinding (Flannery 1986; MacNeish 1967a; Mac-
Neish and Peterson 1962; Ranere et al. 2009).
814 Genet Resour Crop Evol (2010) 57:813–825
1 3
7/17/2019 Ziz Originagriculturemesoamerica
http://slidepdf.com/reader/full/ziz-originagriculturemesoamerica 3/13
Ecological conditions during initial domesticationand early agriculture
Paleoecological records for the Neovolcanic Trans-
verse Axis and the lowlands south of this axis, indicate
rises in temperature, rainfall and atmospheric CO2
concentrations between 12,000 and 9,000 BP, as well as
the presence of a long drought period before the rainy
season (Cunniff et al. 2008; Metcalfe 2006; Piperno
2006). Tropical flora began to displace the boreal
forests, and the lowland thorny bush vegetation was
taken over by tropical dry forest (TDF). The combina-tion of these transformations produced a transition from
C3 to C4 grasses, growing populations of Panicum spp.,
Setaria spp., Tripsacum spp. and Zea spp., and
expansion of certain dicotyledonea families, such as
Chenopodiaceae, Amaranthaceae, Asteraceae, Cucur-
bitaceae and Solanaceae (Cunniff et al. 2008; Piperno
et al. 2007; Sage 1995). It was during this period that
the Balsas-Jalisco (BJ) biotic morphotectonic province
(Fig. 2) took form south of the Neovolcanic Transverse
Axis (Ferrusquıa-Villafranca 1990). Its floral andfaunal distribution, structure and composition have
changed little during the last 9,000 years (Metcalfe
2006; Piperno 2006). Between 16,500 and 4,000 BP,
high eruptive activities and extensive lava flows from
the Colima Volcanic Complex (CVC) split the region
from north to south (Cortes et al. 2005; Capra and
Macıas 2002), leaving an area to the northwest rich in
grasses, including: Tripsacum dactyloides (L.) L. var.
mexicanum De Wet et Harlan L; T. lanceolatum Rupr.
ex Fourn.; T. maizar Hernandez et Randolph;
T. pilosum Scribner and Merr.; Zea diploperennis Iltis,Doebley et Guzman; Z. mays L. ssp. parviglumis Iltis et
Doebley; and Z. perennis (Hitchcock) Reeves et
Mangelsdorf (Eubanks 2001; Wilkes 2004).
Distribution of putative ancestral populations
Phylogenetic analyses using nuclear DNA have shown
that Z. mays ssp. parviglumis populations in the central
Fig. 1 Probable routes of initial human migrations into
Mesoamerica along river corridors: (1) Grande de Santiago-
Lerma, (2) Armerıa-Tuxcacuesco, (3) Balsas, (4) Amacuzac
and (5) Mexcala-Nexpa-Atoyac. Main Pleistocenic fauna
hunting sites in the Neovolcanic Transversal Axis: ( A)
Chapala-Zacoalco-Sayula, ( B) Valley of Mexico and (C )
Puebla-Valsequillo
Genet Resour Crop Evol (2010) 57:813–825 815
1 3

7/17/2019 Ziz Originagriculturemesoamerica
http://slidepdf.com/reader/full/ziz-originagriculturemesoamerica 4/13
and west BJ region could be the putative progenitors of
domesticated maize (Buckler et al. 2006; Fukunaga
et al. 2005; Matsuoka et al. 2002). Phylogenetic
relationships inferred from mitochondrial genes sug-
gest that wild populations of C. argyrosperma Huber
ssp. sororia Merrick et Bates in this region are the
putative source populations of domesticated varieties
(Sanjur et al. 2002). For the bean Phaseolus vulgaris
L., genetic-molecular research using nuclear DNA
suggests that wild populations of this species in the
lower Lerma-Santiago river basin and in the northwestBJ region are the ancestral populations of domesti-
cated varieties (Gepts 1988; Kwak et al. 2009,
Zizumbo-Villarreal et al. 2009a) (Fig. 3).
The origin of agriculture
A growing carbon accumulation in paleoecological
records from the BJ region to Panama’s southern
coast between 10,000 and 7,000 BP seems to indicate
systematic use of fire by humans. There is also a
simultaneous increase in the presence and accumu-
lation of Zea genus grass pollens in the central-
western BJ (Piperno 2006; Piperno et al. 2007).
Continued use of fire in the TDF of the BJ region led
to a drastic decline in diversity, the dominance of tree
species with the capacity to sprout from the root
crown or stem and the appearance of grass patches
(Miller 1999; Miller and Kauffman 1998; Sanchez-
Velazquez et al. 2002). Eventually, teocintle, beansand squash would begin to grow together in these
patches (Flannery 1986; Wilkes 2004).
The increased density of edible species promoted by
burning could have defined collecting rounds and the
location of seasonal shelters nearby. Rock shelter
distribution, their size and the archaeological remains
inside them in the central BJ region about 8,700 cal-
endar years (cal) BP indicate that they were occupied
for a number of weeks by small groups of humans.
Fig. 2 Biotic provinces (Ferrusquıa-Villafranca 1990), and archaeological sites (Flannery 1986; Kelly 1980; MacNeish 1964;
Mountjoy 2006; Ranere et al. 2009)
816 Genet Resour Crop Evol (2010) 57:813–825
1 3

7/17/2019 Ziz Originagriculturemesoamerica
http://slidepdf.com/reader/full/ziz-originagriculturemesoamerica 5/13
These groups made use of the surrounding resources bygathering plants, hunting deer and smaller fauna, and
cultivating of maize and squash using tools such as fire,
bifacial scrapers and choppers, as well as handstones
and base stones for grinding (Ranere et al. 2009).
Plant domestication
Based on molecular clock results, the initial separa-
tion between wild and domesticated maize popula-
tions probably occurred &9,000 BP (Matsuoka et al.2002). This coincides with accelerator mass spec-
trography (AMS) results for maize and squash starch
grains domesticated about 8,700 cal BP found in
Xihuatoxtla in the central BJ region (Ranere et al.
2009). Results reported by Piperno et al. (2009)
support a scenario in which maize was domesticated
in the low, humid portions of the Balsas river
watershed, where Z. mays ssp. parviglumis is native.
The size and morphology of maize starch grains and
phytoliths is indicative of human selection for theteocintle glume architecture gene (tga1), a major
domestication gene in Zea which controls phenotypic
attributes key to efficient utilization of maize as a
grain (Doebley 2004; Piperno et al. 2009). This
selection increased starch quantity and quality, while
reducing grain glume size and hardness, leading to
naked grains with soft glumes (Wang et al. 2005). For
squash, phytolith size and morphology is indicative
of human selection pressure on the Hr gene, a major
domestication gene in Cucurbita which controls
crucial phenotypic attributes such as fruit hardnessand phytolith quantity (Piperno et al. 2002; 2009).
Leguminous starch grains found at Xihuatoxtla in
the central BJ region may correspond to beans
(P. vulgaris or P. lunatus) (Piperno et al. 2009:
supporting data) and suggest that bean may have been
domesticated in this region as early as 9,000 cal BP.
However, AMS-dated macro-remains of beans found
at Tehuacan suggest domestication by 2,285 cal BP,
while bean remains from Guila Naquitz have been
Fig. 3 Location of wild populations of putative ancestors to
plant species domesticated in Mesoamerica: (Cu) Cucurbita
argyrosperma Huber; (Pv) Phaseolus vulgaris L; ( Zm) Zea
mays L. (Fukunaga et al. 2005; Gepts 1988; Kwak et al. 2009;
Matsuoka et al. 2002; Sanjur et al. 2002; Zizumbo-Villarreal
et al. 2009a)
Genet Resour Crop Evol (2010) 57:813–825 817
1 3
7/17/2019 Ziz Originagriculturemesoamerica
http://slidepdf.com/reader/full/ziz-originagriculturemesoamerica 6/13
dated to 2,098 cal BP (Kaplan and Lynch 1999). In
beans, the domestication syndrome includes traits
that limit natural dispersion, such as loss of fruit
dehiscence, seed latency and photoperiod sensitivity
(Koinange et al. 1996).
Diffusion of domesticated species
In the early Holocene, the distribution of human
shelters in Mesoamerica and the similar technological
development of human groups in the area (MacNeish
1967a; MacNeish and Peterson 1962; Flannery 1986;
Ranere et al. 2009) suggest that rivers, which would
have been regular water sources during the long dry
season, functioned as biological and cultural disper-
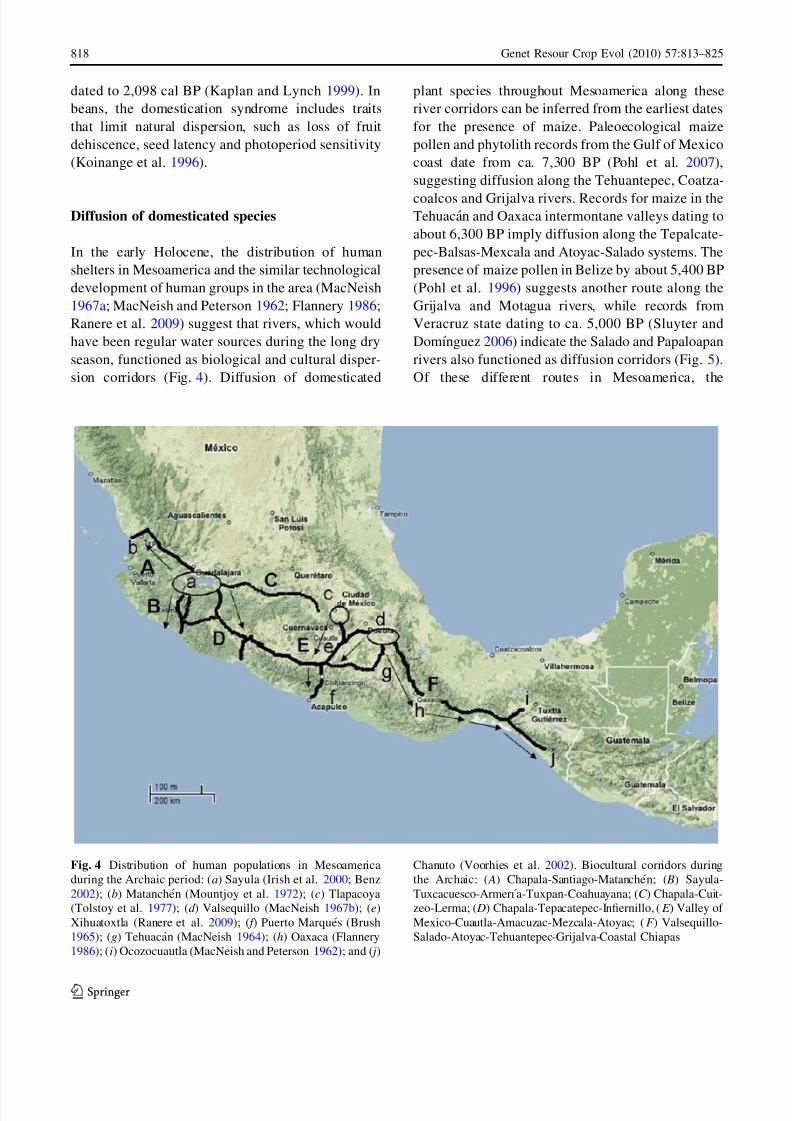
sion corridors (Fig. 4). Diffusion of domesticated
plant species throughout Mesoamerica along these
river corridors can be inferred from the earliest dates
for the presence of maize. Paleoecological maize
pollen and phytolith records from the Gulf of Mexico
coast date from ca. 7,300 BP (Pohl et al. 2007),
suggesting diffusion along the Tehuantepec, Coatza-
coalcos and Grijalva rivers. Records for maize in theTehuacan and Oaxaca intermontane valleys dating to
about 6,300 BP imply diffusion along the Tepalcate-
pec-Balsas-Mexcala and Atoyac-Salado systems. The
presence of maize pollen in Belize by about 5,400 BP
(Pohl et al. 1996) suggests another route along the
Grijalva and Motagua rivers, while records from
Veracruz state dating to ca. 5,000 BP (Sluyter and
Domınguez 2006) indicate the Salado and Papaloapan
rivers also functioned as diffusion corridors (Fig. 5).
Of these different routes in Mesoamerica, the
Fig. 4 Distribution of human populations in Mesoamerica
during the Archaic period: (a) Sayula (Irish et al. 2000; Benz
2002); (b) Matanchen (Mountjoy et al. 1972); (c) Tlapacoya
(Tolstoy et al. 1977); (d ) Valsequillo (MacNeish 1967b); (e)
Xihuatoxtla (Ranere et al. 2009); ( f ) Puerto Marques (Brush
1965); (g) Tehuacan (MacNeish 1964); (h) Oaxaca (Flannery
1986); (i) Ocozocuautla (MacNeish and Peterson 1962); and ( j)
Chanuto (Voorhies et al. 2002). Biocultural corridors during
the Archaic: ( A) Chapala-Santiago-Matanchen; ( B) Sayula-
Tuxcacuesco-Armerı a-Tuxpan-Coahuayana; (C ) Chapala-Cuit-
zeo-Lerma; ( D) Chapala-Tepacatepec-Infiernillo, (E ) Valley of
Mexico-Cuautla-Amacuzac-Mezcala-Atoyac; (F ) Valsequillo-
Salado-Atoyac-Tehuantepec-Grijalva-Coastal Chiapas
818 Genet Resour Crop Evol (2010) 57:813–825
1 3

7/17/2019 Ziz Originagriculturemesoamerica
http://slidepdf.com/reader/full/ziz-originagriculturemesoamerica 7/13
Tepalcatepec-Balsas-Mexcala corridor was probably
a major route for human movement beginning in the
early Holocene (Benz 1999).
Domesticated maize also moved quickly into
Central and South America, as shown by its presence
in Panama by about 7,800–7,000 BP (Dickau et al.
2007), suggesting diffusion routes along the Grijalva,
Motagua and Chamelecon rivers and along the south
coast of Central America. Paleoecological data for
the presence of maize pollen, starch grains and
phytoliths in the Cauca and Ponce valleys of Colombia by about 7,500 BP (Aceituno and Castillo
2005), and on the southwest coast of Ecuador around
the 6,200 cal. BP (Zarrillo et al. 2008) imply another
route along the western and central mountain ranges
of Colombia. Finally, maize and chili phytoliths and
starch grain records from grinding artifacts in the
Andean region by 6,000 BP indicate these two
domesticated species were probably diffused simul-
taneously (Perry et al. 2007).
Agricultural intensification
Paleoecological records for the BJ region between
7,000 and 5,550 BP contain evidence of increased
levels of Asteraceae family weed species (typical of
short fallow systems), greater maize pollen accumu-
lation and decreases in carbon deposits (Piperno
2006; Piperno et al. 2007), suggesting agricultural
intensification. Between 7,000 and 6,000 cal BP, a
new tool kit begins to appear in the archaeological
record, including large, open stone ovens, woodentools such as levers, contracting stems and traps,
bifacial knives, plano-convex scrapers, bifacial stone
choppers, mortars, pestles and grinding handstones
and bases. Human shelter and camp distribution, size
and occupation show that macrobands (15–20 per-
sons) began to form during the rainy season to
cultivate and harvest plants (Flannery 1986; MacNe-
ish 1964), further suggesting agricultural intensifica-
tion. Cultivation areas were in the piedmont, on
Fig. 5 Possible routes for early diffusion of plant species
domesticated in Mesoamerica: ( A) Santiago-Lerma; ( B) Tepal-
catepec-Balsas; (C ) Mexcala-Amacuzac; ( D) Atoyac1-Salado-
Atoyac2; (E ) Mixteco-Verde-Atoyac2; (F ) Tehuantepec;
(G) Grijalva; ( H ) South coast; ( I ) Motagua; ( J ) Caribbean;
(K ) Chamalecon; ( L ) Belize; ( M ) Coatzacoalcos; ( N ) Papaluapan;
and (O) Maya lowland
Genet Resour Crop Evol (2010) 57:813–825 819
1 3

7/17/2019 Ziz Originagriculturemesoamerica
http://slidepdf.com/reader/full/ziz-originagriculturemesoamerica 8/13
terraces close to valleys and plains near lakes
(MacNeish 1964; Ranere et al. 2009).
Morphological analyses of maize cobs dating to
6,300 BP indicate the presence of cobs with two to
four rows of grains, grains with reduced glumes and
total retention of seeds, meaning this crop had
become completely reliant on humans as a replace-ment seed dispersal mechanism (Benz 2001; Piperno
and Flannery 2001). Molecular analyses using
archaic DNA show fixing of Teocintle branched 1
(Tb1), a gene that largely controls short branches
tipped with ears in domesticated maize (Jaenicke-
Despres et al. 2003; Jaenicke-Despres and Smith
2006).
Milpa multicropping
Between 6,000 and 5,000 BP in Mesoamerica,
semi-sedentary macrobands lived on valley terraces
in small circular or oval units, with small ovens for
cooking food, and common storage of agricultural
surplus (MacNeish 1964; Flannery 1986). Stone
bases are replaced by manos and metates for
grinding, and stone cups and pots appear in the
record (Flannery 1986; MacNeish 1964, 1967b).
These tools broadened the ability to transform
foods, involving their human users in the applica-
tion of new selection pressures on grain species.During this period, cultivated plots may have been
established near habitation units by transport of
domesticated perennials such as red mombin and
agaves and the spontaneous arrival of weed species
like chilies, tomatoes (Physalis spp.), amaranth
( Amaranthus spp.) and cotton (Gossypium hirsutum
L.). Dogs were raised as food, and archaeological
and molecular studies suggest it was at this time
that the Xoloitzcuintle dog breed arose and was
raised as a food resource in the BJ region (Wayne
et al. 2006).By 5,500 BP, the alleles for four rows of grains on
maize cobs had fixed; archaeobotanical remains
indicate the presence of cobs with 8–12 rows,
although the alleles for this trait had not yet fixed.
In-field selection for higher numbers of rows sub-
stantially increased maize’s genetic productivity
(Jaenicke-Despres et al. 2003; Jaenicke-Despres and
Smith 2006).
Development of the agro-food system
The traits involved in the compact architecture of the
maize plant had fixed by about 4,400 BP, including
apical dominance, fewer stalks, one to two cobs on
the central axis and cobs with 12–16 rows (Jaenicke-
Despres et al. 2003; Jaenicke-Despres and Smith2006). Ceramics appear in the archaeological record
in the form of comals, cups and simple pots (Brush
1965; Mac Neish 1976b; Flannery 1994), which
would further open the possibilities for food trans-
formation and introduce new selection pressures. It is
during this period that human selection begins to
change the alleles determining protein and starch
quality ( pbf and SuI ) (Jaenicke-Despres et al. 2003;
Jaenicke-Despres and Smith 2006), suggesting a
focus on cooking traits. By 4,000 BP, inhabitants of
the intermontane valleys cultivated crops in agrohab-itats including piedmont terraces, valley terraces,
plains along lakes and rivers and house gardens
(Flannery 1986; MacNeish 1964). A utilitarian
ceramic tradition developed and diversified between
4,500 and 3,500 BP that was related to food harvest,
transport, storage, transformation and consumption.
By 3,500 BP, the Capacha culture had developed in
Colima state and Mascota in southern Jalisco, with a
ceramic corpus that included food processing tools
for soaking, cooking and steaming, as well as
fermentation and possibly distillation of alcoholicbeverages (Kelly 1980; Mountjoy 2006; Zizumbo-
Villarreal et al. 2009b) (Fig. 2). This ceramic diver-
sification reflects greater complexity in the methods
used for food transformation and in the selection
pressures on domesticated species. Between 3,000
and 2,000 BP, maize with a high frequency of the suI-
M2 allele, implied in the amount and quality of floury
starch, appears in the archaeological record (Jae-
nicke-Despres and Smith 2006), suggesting selection
linked to tortilla production.
Discussion
Archaeological evidence suggests that agriculture and
plant domestication in west Mesoamerica were
probably initiated by small, highly mobile groups of
humans from a Clovis cultural tradition who gathered
plants, hunted small animals and seasonally inhabited
820 Genet Resour Crop Evol (2010) 57:813–825
1 3

7/17/2019 Ziz Originagriculturemesoamerica
http://slidepdf.com/reader/full/ziz-originagriculturemesoamerica 9/13
small rock shelters near rivers between inland lake
systems and the Pacific coast (Flannery 1986; Mac-
Neish 1967a; MacNeish and Peterson 1962; Ranere
et al. 2009). These archaeological data do not support
the hypothesis of Sauer (1952) and Harlan (1995) that
domestication was initiated by semi-sedentary fish-
erfolk living near lake systems who initiated agricul-ture and domestication by transporting weedy
ancestral populations of domesticated maize, squash
and beans to plots near habitation sites, where they
would care for and harvest them. Evidence for more
permanent settlements with homegardens in the BJ
region, Oaxaca and Tehuacan appear rather later than
domestication and agriculture (Flannery 1986; Rane-
re et al. 2009).
Paleoecological, ecophysiological and molecular
genetic data suggest that plant domestication and
agriculture began in the TDF of the BJ region underconditions of warm temperatures and annual rainfall
near 1,000 mm, and from selection and management
of the tropical grass Zea mays ssp. parviglumis. It is
unlikely that domestication and agriculture arose in
the intermontane valleys under temperate, semi-dry
conditions in xerophytic bush vegetation through
cultivation of the hybrid Zea diploperennis 9
Tripsacum dactyloides, as suggested by MacNeish
and Eubanks (2000) and Eubanks (2001, 2002).
The autoecological traits and altitude distribution
of putative wild ancestral populations for maize andbeans suggest that agriculture could have begun at
intermediate elevations (600–1,600 masl), including
at the oak forest ecotone (Buckler et al. 2006; Kwak
et al. 2009). This would have enabled human groups
to cultivate plants, while making relatively short
excursions to the coast or intermontane lakes (&40–
80 km), thus reinforcing the biocultural corridors
developed during the early Holocene.
The sympatric geographic distribution of the
putative ancestral populations of maize, beans and
squash in the northwest BJ region suggest that theircultivation and domestication could have begun in
that area. Early dates for maize and squash place the
beginning of this process around 10,000 BP, making
it roughly contemporaneous with the beginning of
domestication in the Old World (Piperno et al. 2009).
The presence of starch grains from domesticated
maize and squash, as well as the presumed presence
of beans, in grinding stones by 9,000 BP (Piperno
et al. 2009) indicates that these species were
simultaneously harvested and consumed and may
have been contemporaneously domesticated. This
possibility is reinforced by records of early and
simultaneous diffusion of maize and chili to South
America (Perry et al. 2007), although a number of
authors have suggested that maize, beans and squash
were domesticated in different regions and periods(Harlan 1995; Kwak et al. 2009).
According to paleoecological data, clearing and
fire were commonly used between 10,000 and
7,000 cal BP. Initially, fire was probably a tool for
hunting, but probably also became the main strategy
to produce harvests and a strong selective force for
plant species that would become domesticated, a
scenario suggested for other regions in the world
(Lewis 1972; Zong et al. 2007). During the Archaic
period, at least 220 hunter-gatherer groups in what is
today west Mexico and the United Stated used fire toestablish grasslands, guide and enclose animals and
augment forage and grain species production (Parker
2002; Stewart et al. 2002; Williams 2003). Zea,
Phaseolus and Cucurbita species are preadapted to
fire. This element favors colonization and establish-
ment in Zea species, and cyclical fire disturbance
helps them maintain populations (Sanchez-Velazquez
et al. 2002). Fire also helps Phaseolus, Cucurbita and
Capsicum species to recolonize fire-disturbed areas
since their seeds exit latency when exposed to
temperatures greater than 60C (Rolston 1978;Degreef et al. 2002). Arboreal species such as
Leucaena spp.; Psidium spp., Prosopis spp. and
Spondias purpurea L. are also preadapted to fire
because of their ability to sprout from the stem base
and root crown. The same holds true for agaves with
the capacity for vegetative propagation because they
can emit root-shoots. Under these circumstances,
humans may have taken advantage of partial removal
of vegetation by fire to cultivate desired grains and
seeds, while the sprouting ability of wild bush species
and humans’ technological inability to eliminateperennial plant roots dictated use of an agricultural
system with a long fallow period in which both fire-
resistant annuals and perennials were domesticated
and incorporated.
Changes in the maize tga 1 gene were initially
focused on the cob and grain, in an effort to produce
naked, detachable grains (Piperno et al. 2009).
Selection for infructescences that retain ripe grains
increased the likelihood of harvest in the field, while
Genet Resour Crop Evol (2010) 57:813–825 821
1 3

7/17/2019 Ziz Originagriculturemesoamerica
http://slidepdf.com/reader/full/ziz-originagriculturemesoamerica 10/13
selection for naked grains facilitated processing by
grinding. Archaeological dates for maize from
Xihuatoxtla (Piperno et al. 2009) do not support the
hypothesis of Harlan (1995), Iltis (2000) or Smalley
and Blake (2003) that initial selection of maize was
linked to use of the stalk and unripe cobs as greens
and as a sugar source in alcoholic beverage produc-tion. Archaeological records also indicate frequent
use of cooked agave leaf bases and Spondias fruit
about 9,000 BP (Callen 1967; Flannery 1986; Smith
1986), suggesting selection for high sugar content
variants in both plants. These sugar sources became
extremely important in alcoholic beverage production
in the BJ region (Bruman 2000; Colunga-Gar-
cıaMarın and Zizumbo-Villarreal 2007).
Initial domestication of grasses was therefore
linked to ease of harvest and processing. Given that
technological development was similar among humangroups at the time, domestication could have
occurred over a wide area, both in the intermontane
valleys where Setaria parviflora (Poiert) Kerguelen
could have been domesticated (Austin 2006; Callen
1967), and in the lowlands where Z. parviglumis was
domesticated (Ranere et al. 2009). Fixation of
maize’s Tb1 and tga1 genes by about 6,400 BP
suggests that selection continued to be linked to ease
of harvest (e.g. total retention of seeds on the cob,
rachis with two spikelets with two rows of grains) and
processing (e.g. naked grains with short glumes)(Doebley 2006; Dorweiler et al. 1993; Wang et al.
2005). Simultaneously, the high eruptive activities in
CVC between 7,300 and 6,300 BP might have caused
environmental changes related to the metal content of
local soils that may have been important in maize
domestication (Cortes et al. 2005; Ville-Calzada et al.
2009).
Diffusion of the maize-beans-squash domesticates
suite most likely occurred via existing biocultural
corridors, i.e. rivers, since these would have been
dependable year round food and water sources forhumans and animals, particularly during the long dry
season. The diffusion of domesticated maize and
squash began when they still exhibited low genetic
differentiation with wild varieties. As a result, their
transport to areas lacking wild populations could have
accelerated the fixation of domestic traits and the
disappearance of wild traits. Environmental and
cultural conditions in each region would have
promoted initial diversification.
Agricultural intensification began to occur
between 7,000 and 5,000 BP as humans actively
worked soils using wooden tools such as levers,
which helped to remove perennial plants, and extract
and arrange rocks to improve soil conditions. This
technique produces a heterogeneous land surface in
which maize, squash and bean seeds are sowntogether simulating their natural growth after fire
disturbance in the BJ region. The structuring of the
milpa multi-crop agricultural system occurred
through joint sowing of one or various seeds of these
three species in the same ‘‘microsite’’ within a burned
area, which would have been previously prepared by
removal of roots and rocks using wooden levers.
Additional structuring would have been accom-
plished through individual care and harvest of each
plant. Under these circumstances, these three species
would have been subject to concurrent natural andhuman selection pressures that could have led them to
a state of ecological adaptation and complementarity.
This does not coincide with the scenarios proposed
for other regions such as the Middle East, where
preparation of homogeneous planting areas using
draft animals and crop rows allowed use of broadcast
sowing and harvest without human selection of
individual plants. Fixing of the alleles involved in
maize plant architecture by 4,400 cal. BP suggests
that this multicrop system was established by this
time.Grinding of maize, squash and possibly beans
under domestic conditions suggest the presence of a
selective process that could have produced food
complementarity. Fixation of the alleles involved in
maize protein and starch quality by 4,400 BP may
indicate that this complementarity had been attained
by this time. In the BJ region, agro-food technology
had become highly complex between 4,500 and 3,500
BP, as reflected in the use of pots for steaming, and
probably also distilling, not found in other regions of
Mesoamerica at this time (Zizumbo-Villarreal et al.2009b). In other words, in this region at this time, the
agro-food system was complete, suggesting that the
milpa system originated here.
Despite the highly complex cultural development
in the BJ region during the rise of plant domestication
and agriculture in Mesoamerica, very few archaeo-
botanical studies have been done there (Benz 2002,
Ranere et al. 2009). New research will be vital to
better understanding the initial development of
822 Genet Resour Crop Evol (2010) 57:813–825
1 3

7/17/2019 Ziz Originagriculturemesoamerica
http://slidepdf.com/reader/full/ziz-originagriculturemesoamerica 11/13
human societies in Mesoamerica. Study of the TDF in
the BJ region is particularly urgent since almost 80%
of this vegetation type has disappeared to date, and,
of the TDF zones in Mesoamerica, it suffers the
highest deforestation rate and is at the highest risk of
disappearing (Janzen 1988; Kauffman et al. 2003;
Trejo and Dirzo 2000). Native grasses in the regionhave experienced strong selective pressure since the
introduction of cattle in the 16th Century. This has
accelerated during the last 30 years as cattle ranching
has grown rapidly and African grasses have been
introduced and managed with pesticides, putting
many of the putative wild source populations,
particularly those of Z. parviglumis, at serious risk
(Houghton et al. 1991; Wilkes 2007). In situ conser-
vation programs at reserves such as Manantlan-Cerro
Grande and Zicuiran-Infiernillo need to be reinforced
and broadened to include nearby areas and humancommunities, collection programs need to be stepped
up and adaptive studies of climatic, edaphic and
biotic factors done before these wild ancestral of
domesticated species disappear.
Acknowledgments The authors thank the CONACYT and
CICY for sabbatical scholarships and P. Gepts for his
hospitality at UC-Davis.
References
Aceituno FJ, Castillo EN (2005) Mobility strategies in
Colombia’s middle mountain range between the early and
middle Holocene. Before farming 2005/2, Article 2.
Available at www.waspress.co.uk/journals/beforefarming/
journal_20052/abstracts/index.php
Aliphat FM (1980) La cuenca Zacoalco-Sayula: Ocupacion
humana durante el pleistoceno final en el occidente de
Mexico. In: Gonzalez-Jacome A (ed) Orıgenes del hom-
bre americano. SEP, Mexico, pp 145–176
Austin DF (2006) Fox-tail millets (Setaria:Poaceae). Aban-
doned food in two hemispheres. Econ Bot 60(2):143–158
Benz B (1999) On origin, evolution and dispersal of maize. In:
Blake M (ed) Pacific Latin America in prehistory: the
evolution of Archaic and Formative cultures. Washington
State University Press, Pullman, pp 25–38
Benz B (2001) Archaeological evidence of teocintle domesti-
cation from Guila Naquitz, Oaxaca. Proc Nat Acad Sci
USA 98:2104–2106
Benz B (2002) The origins of Mesoamerican agriculture: recon-
naissance and testing in the Sayula-Zacoalco lake basin.
www.famsi.org/reports/99074/section11.htm. Accessed 10
Nov 2009
Bruman HJ (2000) Alcohol in ancient Mexico. Univ of Utah
Press, Salt Lake City
Brush CF (1965) Pox pottery: earliest identified Mexican
ceramic. Science 149:194–195
Buckler ES, Goodman MM, Holstford TP, Doebley JF, San-
chez GJ (2006) Phylogeography of the wild subspecies of
Zea mays. Maydica 51:123–134
Callen EO (1967) Food habits of some Pre-Columbian Mexi-
can Indians. Econ Bot 19(4):335–343
Capra L, Macıas JL (2002) The cohesive Naranjo debris-flow
deposit (10 km3): a dam breakout floor derived from the
Pleistocene debris-avalanche deposit of Nevado de Coli-
ma Volcano (Mexico). J Volcan Geotherm Res 117:
213–235
Colunga-GarcıaMarın P, Zizumbo-Villarreal D (1993) La
evolucion de las plantas bajo seleccion artificial y manejo
agrıcola. In: Leff E, Carabias J (eds) Cultura y manejo
sustentable de los recursos naturales. CIIH-UNAM-
Miguel Angel Porrua, Mexico, pp 123–163
Colunga-GarcıaMarın P, Zizumbo-Villarreal D (2007) Tequila
and other Agave spirits from west-central Mexico: current
germplasm diversity, conservation and origin. Biodivers
and Conserv 16:1653–1667
Cortes A, Garduno-Monroy VH, Navarro-Ochoa C, Komo-rowski JC, Saucedo R, Macıas JL, Gavilanes JC (2005)
Geologıa del complejo volcanico Colima. Carta Geolog-
ica y Minera No. 10. Instituto de Geologıa UNAM.
Mexico
Cunniff J, Osborne CP, Ripley BS, Charles M, Jones G (2008)
Response of wild C4 crop progenitors to subambient CO2
highlights a possible role in the origin of agriculture.
Global Change Biol 14:576–587
Degreef J, Rocha OJ, Vanderborght T, Baudoin JP (2002) Soil
seed bank and seed dormancy in wild populations of lima
bean (Fabaceae): considerations for in situ and ex situ
conservation. Am J Bot 89(10):1644–1650
Dickau R, Ranere AJ, Cooke RG (2007) Starch grain evidence
for the preceramic dispersals of maize and root crops intotropical dry and humid forest of Panama. Proc Nat Acad
Sci USA 104:3651–3656
Dixon EJ (1999) Bones, boats and bison. Archaeology and the
first colonization of western North America. Univ of Utah
Press, Salt Lake City
Dixon EJ (2001) Human colonization of the Americas: timing,
technology and process. Quarter Sci Rev 20:277–299
Doebley JF (1984) ‘‘Seeds’’ of wild grasses: a major food for
Southwestern Indians. Econ Bot 38(1):52–64
Doebley JF (2004) The genetics of maize evolution. Ann Rev
Genet 38:37–59
Doebley JF (2006) Unfallen grains: how ancient farmers turned
weeds into crops. Science 312:1318–1319
Dorweiler J, Stec A, Kermicle J, Doebley JF (1993) Teocintle
glume architecture 1: a genetic locus controlling a key
step in maize evolution. Science 262:233–235
Eubanks M (2001) The origin of maize: evidence for Tripsa-
cum ancestry. Plant Breed Rev 20:15–66
Eubanks M (2002) An interdisciplinary perspective on the
origin of maize. Latin Am Antiquity 12:91–98
Ferrusquıa-Villafranca I (1990) Provincias biogeograficas con
base en rasgos morfotectonicos. Mapa IV8.10. Atlas
Nacional de Mexico, vol III. Instituto de Geografıa,
UNAM, Mexico
Flannery KV (1986) Guila Naquitz. Academic Press, Orlando
Genet Resour Crop Evol (2010) 57:813–825 823
1 3

7/17/2019 Ziz Originagriculturemesoamerica
http://slidepdf.com/reader/full/ziz-originagriculturemesoamerica 12/13
Flannery KV (1994) Early formative pottery of the Valley of
Oaxaca, Mexico. Memoir No. 27. Museum of Anthro-
pology, University of Michigan, Ann Arbor
Fukunaga K, Hill J, Vigouroux Y, Matsuoka Y, Sanchez J, Liu
K, Buckler ES, Doebley J (2005) Genetic diversity and
population structure of teocintle. Genetics 169:2241–2254
Gepts P (1988) Phaseolin as an evolutionary marker. In: Gepts
P (ed) Genetic resources of Phaseolus beans. Kluwer,
Dordrecht, pp 215–241
Gepts P (2004) Crop domestication as a long-term selection
experiment. Plant Breed Rev 24(Part 2):1–44
Gepts P (2008) Tropical environments, biodiversity, and the
origin of crops. In: Moore P, Ming R (eds) Genomics of
tropical crop plants. Springer, New York, pp 1–20
Hammer K (1984) Das Domestikationssyndrom. Kulturpflanze
32:11–34
Harlan JR (1971) Agricultural origins: centers and non centers.
Science 174:468–474
Harlan JR (1992) Crops and man, 2nd edn. American Society
of Agronomy and Crop Science Society, Madison
Harlan JR (1995) The living fields. Cambridge Univ Press,
CambridgeHoughton RA, Lefkowitz DS, Skole DL (1991) Changes in the
landscape of Latin America between 1850–1985. I. Pro-
gressive loss of forest. For Ecol Manag 38:173–199
Iltis HH (2000) Homeotic sexual translocations and the origin
of maize ( Zea mays, Poaceae): a new look at an old
problem. Econ Bot 54:7–42
Irish DJ, Davis SD, Lobdell JE, Solorzano FA (2000) Prehis-
toric human remains from Jalisco, Mexico. Curr Res in
the Pleistocene 17:95–96
Jaenicke-Despres V, Smith BD (2006) Ancient DNA and
integration of archaeological and genetic approaches to
study of maize domestication. In: Staller J, Tykot R, Benz
B (eds) Histories of maize. Academic Press, San Diego,
pp 83–95Jaenicke-Despres V, Buckler ES, Smith BD, Gilbert TM,
Cooper A, Doebley J, Paabo S (2003) Early allelic
selection in maize as revealed by ancient DNA. Science
302:1206–1208
Janzen DH (1988) Tropical dry forest. The most endangered
major tropical ecosystem. In: Wilson EO (ed) Biodiver-
sity. National Academy Press, Washington, pp 130–137
Kaplan L, Lynch TE (1999) Phaseolus (Fabaceae) in archae-
ology: AMS radiocarbon dates and their significance for
pre-Colombian agriculture. Econ Bot 53:261–272
Kauffman JB, Steele MD, Cummings DL, Jaramillo VJ (2003)
Biomass dynamic associated with deforestation, fire, and
conversion to cattle pasture in Mexican tropical dry forest.
For Ecol Manag 176:1–12
Kelly I (1980) Ceramic sequence in Colima: capacha, an early
phase. Anthropological papers No 37. Univ of Arizona
Press, Tucson
Kirchhoff P (1943) Mesoamerica. Acta Americana. Revista de
la Sociedad de Antropologıa y Geografıa 1:92–107
Koinange EMK, Singh SP, Gepts P (1996) Genetic control of
the domestication syndrome in common bean. Crop Sci
36:1037–1045
Kwak M, Kami J, Gepts P (2009) The putative Mesoamerican
center of domestication of Phaseolus vulgaris L. is
located in the Rio Lerma-Santiago basin of Mexico. Crop
Sci 49:554–563
Leonard JA, Wayne RK, Wheeler J, Valadez R, Gullen S, Vila
C (2002) Ancient DNA evidence for Old World origin of
New World dogs. Science 298:1613–1616
Lewis HT (1972) The role of fire in the domestication of plants
and animals in Southwest Asia: a hypothesis. Man (Lond)
7:195–222
Lorenzo JL, Mirabell L (1986) Tlapacoya: 35, 000 anos de
historia en el Lago de Chalco, Coleccion Cientıfica, Serie
Prehistoria. Instituto Nacional de Antropologıa e Historia,
Mexico
MacNeish RS (1964) Ancient Mesoamerica civilization. Sci-
ence 143:531–537
MacNeish RS (1967a) A summary of the subsistence. In: Byers
S (ed) Environment and subsistence, Vol. 1: the prehistory
of the Tehuacan Valley. Univ Texas Press, Austin, pp
290–309
MacNeish RS (1967b) Mesoamerican archaeology. Bienn Rev
Anthropol 5:306–331
MacNeish RS, Eubanks EM (2000) Comparative analysis of
the Rio Balsas and Tehuacan models for the origin of maize. Latin Am Antiq 11(1):3–20
MacNeish RS, Peterson FA (1962) The Santa Marta Rock
Shelter, Ocozocoautla, Chiapas, Mexico. Papers of the
New World Archaeological Foundation 14. Brigham
Young Univ, Provo
Matsuoka Y, Vigouroux Y, Goodman MM, Sanchez GJ,
Buckler E, Doebley J (2002) A single domestication for
maize shown by multilocus microsatellite genotyping.
Proc Nat Acad Sci USA 99:6080–6084
Metcalfe SE (2006) Late quaternary environments of the
northern deserts and central transvolcanic belt of Mexico.
Ann Missouri Bot Gar 93:258–273
Miller PM (1999) Effects of deforestation on seed banks in a
tropical deciduous forest of western Mexico. J Trop Ecol15:179–188
Miller PM, Kauffman JB (1998) Seedling and sprout response
to slash and burn agriculture in tropical deciduous forest.
Biotropica 30:538–546
Mountjoy JM (2006) Excavation of two middle formative
cemeteries in the Mascota Valley of Jalisco, Mexico.
Available at www.famsi.org/reports/03009/index.html.
Accessed 12 Nov 2009
Mountjoy JB, Taylor RE, Feldman LH (1972) Matanchen
complex: new radiocarbon dates on early coastal adapta-
tion in west Mexico. Science 175:1242–1243
Niederberger C (1979) Early sedentary economy in the basin of
Mexico. Science 203:131–142
Parker C (2002) Fire in Pre-European lowlands of the Ameri-
can Southwest. In: Vale TR (ed) Western wilderness: fire,
native peoples, and the natural landscape. Island Press,
Washington, pp 101–141
Perry L, Dickau R, Zarrillo S, Holst I, Persall DM, Piperno D,
Berman JM, Cooke GR, Rademaker K et al (2007) Starch
fossils and the domestication and dispersal of chili peppers
(Capsicum spp. L.) in the Americas. Science 315:986–988
Piperno DR (2006) Quaternary environmental history and
agricultural impact on vegetation in Central America. Ann
Missouri Bot Gar 93:274–296
824 Genet Resour Crop Evol (2010) 57:813–825
1 3

7/17/2019 Ziz Originagriculturemesoamerica
http://slidepdf.com/reader/full/ziz-originagriculturemesoamerica 13/13
Piperno DR, Flannery KV (2001) The earliest archaeological
maize ( Zea mays L.) from highland Mexico: new accel-
erator mass spectrometry dates and their implications.
Proc Nat Acad Sci USA 98:2101–2103
Piperno DR, Pearsall DM (1998) The origins of agriculture in
the lowland neotropics. Academic Press, San Diego
Piperno DR, Holst I, Wessel-Beaver L, Andres TC (2002)
Evidence for the control of phytolith formation in Cu-
curbita fruits by the hard rind ( Hr ) genetic locus:
archaeological and ecological implications. Proc Nat
Acad Sci USA 99:10923–10928
Piperno DR, Moreno JE, Iriarte J, Holst I, Lachniet M, Jones G,
Ranere AJ, Castazo R (2007) Late pleistocene and holo-
cene environmental history of the Iguala Valley, central
Balsas watershed of Mexico. Proc Nat Acad Sci USA
104:11874–11881
Piperno DR, Ranere AJ, Holst I, Dickau R, Iriarte J (2009)
Starch grain and phytolith evidence for early ninth mil-
lennium B.P. maize in the central Balsas River valley,
Mexico. Proc Nat Acad Sci USA 106:5019–5024
Pohl ME, Pope KO, Jones JG, Jacob JS, Piperno DR, France
SD, Lentz DL, Gifford JA, Danford ME, Josserand JK (1996) Early agriculture in the Maya Lowlands. Lat Am
Antiq 7:355–372
Pohl ME, Piperno DR, Pope KO, Jones JG (2007) Microfossil
evidence for pre-Columbian maize dispersal in the Neo-
tropics from San Andres, Tabasco, Mexico. Proc Nat
Acad Sci USA 104:6870–6875
Poinar HK, Kuch M, Sobolik DK, Barnes I, Stankiewicz AB,
Kuder T, Spaulding WG, Bryent VM, Cooper A, Paabo S
(2001) A molecular analysis of dietary diversity for three
archaic Native Americans. Proc Nat Acad Sci USA
98:4317–4322
Ranere AJ, Piperno DR, Holst I, Dickau R, Iriarte J (2009) The
cultural and chronological context of early Holocene
maize and squash domestication in the central Balsas RiverValley, Mexico. Proc Nat Acad Sci USA 106:5014–5018
Rolston P (1978) Water impermeable seed dormancy. The Bot
Rev 44(3):365–396
Sage SR (1995) Was low atmospheric CO2 during the Pleis-
tocene a limiting factor for the origin of agriculture? Glob
Change Biol 1:93–106
Sanchez-Velazquez LR, Ezcurra E, Martınez-Ramos M,
Alvarez Buylla E, Lorente R (2002) Population dynamics
of Zea diploperennis, an endangered perennial herb: effect
of slash and burn practice. J Ecol 90:684–692
Sanjur OI, Piperno DR, Andres TC, Wessel-Beaver (2002)
Phylogenetic relationships among domesticated and wild
species of Cucurbita (Cucurbitaceae) inferred from a
mitochondrial gene: implications for crop plant evolution
and areas of origin. Proc Natl Acad Sci USA 99:535–540
Sauer CO (1952) Agricultural origins and dispersal. American
Geographical Society, New York
Sluyter A, Domınguez G (2006) Early maize ( Zea mays L.) cul-
tivation in Mexico: dating sedimentary pollen records and its
implications. Proc Nat Acad Sci USA 103:1147–1151
Smalley S, Blake M (2003) Sweet beginnings: stalk sugar and
domestication of maize. Curr Antropol 44:675–703
Smith CE (1986) Preceramic plant remains from Guila Na-
quitz. In: Flannery KV (ed) Guila Naquitz. Academic
Press, Orlando, pp 265–274
Smith BD (2005) Reassessing Coxcatlan cave and the early
history of domesticated plants in Mesoamerica. Proc Nat
Acad Sci USA 102:9438–9445
Stewart OC, Lewis TH, Anderson MK (2002) Forgotten fires:
Native Americans and the transient wilderness. Univ of
Oklahoma Press, Norman
Tolstoy P, Fish SK, Boksenbaum MW, Vaughan KB, Smith
EC (1977) Early sedentary communities of the Basin of
Mexico. J Field Archaeol 4:91–106
Trejo I, Dirzo R (2000) Deforestation of seasonally dry tropical
forest: a national and local analysis in Mexico. Biol
Conserv 94:133–142
Ville-Calzada JP, Martınez de la Vega O, Hernandez-Guzman
G, Ibarra-Laclette E, Alvarez-Mejıa C, Vega-Arreguın JC,
Jimenez-Moraila B, Fernandez Cortes A, Corona-Ara-
menta G, Herrera-Estrella L, Herrera-Estrella A (2009)
The palomero genome suggests metal effects on domes-
tication. Science 326:1078
Voorhies B, Kennet DJ, Jones GJ, Wake TA (2002) A middle
archaic archaeological site on the west coast of Mexico.
Latin Am Antiq 13:179–200
Wang H, Nussbaum-Waller B, Li Q, Zhao Q, Vigouroux Y,Faller M, Bomblies K, Lunkens L, Doebley J (2005) The
origin of the naked grains of maize. Nature 436:714–719
Wayne R, Leonard JA, Vila C (2006) Genetic analysis of dog
domestication. In: Zeder MA et al (eds) Documenting
domestication: new genetic and archaeological paradigms.
Univ of California Press, Berkley, pp 279–293
Wilkes G (2004) Maize, strange and marvelous: but is a defin-
itive origin known? In: Smith C (ed) Maize: origin, history,
technology and production. Willey, New York, pp 3–63
Wilkes G (2007) Urgent notice to all maize researchers: dis-
appearance and extinction of the last wild teocintle pop-
ulation is more than half completed. A modest proposal
for teocintle evolution and conservation in situ: The
Balsas, Guerrero, Mexico. Maydica 52:49–58Williams GW (2003) References on the American Indian use
of fire in ecosystems. USDA Forest Service. Washington,
DC. Available at www.wildlandfire.com/docs/biblio_
indianfire.htm Accessed 12 Nov 2009
Willis WH (1995) Archaic foraging and the beginning of food
production in the American Southwest. In: Price TD,
Gebauer AB (eds) Last hunters, first farmers. School of
American Research, Santa Fe, pp 215–242
Zarrillo S, Pearsall DM, Raymond JS, Tisdale MA, Quon DJ
(2008) Directly dated starch residues document early
formative maize ( Zea mays L.) in tropical Ecuador. Proc
Nat Acad Sci USA 105:5006–5011
Zizumbo-Villarreal D, Papa R, Hufford M, Repinski S, Gepts P
(2009a) Identification of a new wild population of
Phaseolus vulgaris L. in western Jalisco, Mexico, near the
Mesoamerican domestication center of common bean.
BIC 52:24-25. (Bean Improvement Cooperative USA)
Zizumbo-Villarreal D, Gonzalez-Zozaya F, Olay-Barrientos A,
Almendros-Lopez L, Colunga-GarcıaMarın P (2009b)
Distillation in western Mesoamerica before European
contact. Econ Bot 63(4):413–426
Zong Y, Chen Z, Innes JB, Chen C, Wang Z, Wang H (2007)
Fire and flood management of coastal swamps enabled
first rice paddy cultivation in east China. Nature 449:
459–462
Genet Resour Crop Evol (2010) 57:813–825 825
1 3