topicos sobrea y aracnología.pdfpromedio 6 t/ha de grano y superan las 45 t/ha de forraje verde con...
TRANSCRIPT

ii
TOPICOS SOBRE
ACAROLOGIA Y ARACNOLOGIA

2
IDENTIFICACIÓN DE ÁCAROS ASOCIADOS AL ESPÁRRAGO (Asparagus officinalis L.)
EN EL MUNICIPIO DE CABORCA, SONORA
Identification of acaruses associated to the espárrago (Asparagus officinalis l.) in the
municipality of caborca, sonora
Benito Reséndiz García y Marisol Monroy Gutiérrez Departamento de Parasitología Agrícola.
Universidad Autónoma Chapingo. Km. 38.5 carretera México-Texcoco. Chapingo, Texcoco,
Estado de México. C. P. 56230. Correo-e:[email protected].
Palabras Clave: Asparagus officinalis L., Tetranychus cinnabarinus (Boisduval)
Introducción
En México, la producción de hortalizas es una de las actividades económicas más
sobresalientes del sector agrícola. Las hortalizas ocupan un lugar importante dentro de la
alimentación diaria de la población. Por su gran producción, y variedad han sido agrupadas de
acuerdo con la parte comestible de la planta. La mayoría contienen cantidades considerables de
agua, carbohidratos, proteínas, lípidos, vitaminas y minerales (Damián, 1996).
El espárrago es una hortaliza que se encuentra distribuida en todo el mundo (FAO,2005),
los principales países productores son China con aproximadamente el 89% de la producción
mundial, le sigue Perú con una participación equivalente al 3% de la producción mundial,
estando en continuo aumento, debido a que las condiciones climáticas le permiten producir
durante todo el año. Con porcentajes menores, se encuentran Alemania, México, Italia, España y
EEUU aportando el 1% cada uno. China constituye el principal competidor mundial, orientado
fundamentalmente a la industria del congelado y conserva, accediendo al mercado europeo a
menores precios que Perú.
La producción de espárragos en el mundo ha aumentado considerablemente pasando de
4.5 millones de toneladas en el 2000 a 6.6 millones de toneladas en el año 2005, registrándose un
aumento de 45.6% ocasionado principalmente por mayor demanda del mercado europeo de esta
hortaliza.1
En relación a México, se cultivan alrededor de 14,836.50 hectáreas de espárrago; dentro
de las hortalizas de importancia participa con una producción de 59,620.80 toneladas. El
rendimiento promedio es de 4.18 toneladas por hectárea y el valor de la producción es de 1,
651,483.80 miles de pesos. Los Estados con mayor producción en orden de importancia son
Sonora, Guanajuato, Baja California Norte, Baja California Sur, Jalisco, Nuevo León, Querétaro
y Coahuila (Anónimo, 2006)2.
Aunado al auge que esta hortaliza ha tenido hortaliza ha tenido (expansión del cultivo,
producción, consumo, etc.) y considerando que se han observado una serie de problemas
fitosanitarios en los cuales se involucran plagas insectiles y enfermedades fungosas. En la zona
de Caborca, Sonora una de las principales plagas son los ácaros en espárrago, por ello se plantea
el siguiente el objetivo: caracterizar a las especies de ácaros asociados al espárrago en Caborca,
Sonora.

3
Materiales y Método
La colecta del material biológico se llevó a cabo en plantas de espárrago localizadas en
Caborca, Sonora de marzo a junio del 2008, para lo cual se seleccionaron aquellas plantas de
espárrago que presentaron los síntomas típicos del ataque de araña roja como son: plantas
cloróticas, la presencia de telaraña y especímenes. El material colectado se colocó en bolsas de
polietileno previamente humedecidas, las cuales se trasladaron al laboratorio de Acaralogia del
Departamento de Parasitología Agrícola de la UACH, para su posterior revisión.
El material biológico colectado, se revisó tallo por tallo con la ayuda de un microscopio
estereoscópico, los especímenes encontrados se colocaron en un Siracusa que contenía una
solución de aclarado (lactófenol), donde permanecieron por un espacio de 10 a 15 minutos.
Posteriormente se hicieron los montajes permanentes de la siguiente forma: sobre un portaobjetos
de 75 x 25x 1 mm de tamaño, se colocó una gota de liquido de fitoseide en el centro del mismo, y
con ayuda de un pescador el ácaro se trasladó de la Siracusa al centro de la gota, procurando que
el espécimen quedará perfectamente en posición dorsal o ventral hacia arriba según fuera
requerido, en seguida se colocó un cubreobjetos de 6 x 6 mm sobre la gota, lentamente para así
evitar la formación de burbujas y que el ejemplar quedara con sus extremidades extendidas,
enseguida las preparaciones se colocaron en una plancha eléctrica de calentamiento con el fin de
aclarar aun más el ejemplar y eliminar las burbujas que se formaron durante dicho proceso, los
especímenes se examinaron en un microscopio compuesto para comprobar que el ejemplar
estuviera en perfectas condiciones. Las preparaciones se dejaron por un periodo de cinco días
para que se secaran, más tarde se sellaron con barniz transparente, así mismo por medio de un
círculo se marcó la ubicación del ejemplar para facilitar su búsqueda; finalmente se procedió a
limpiarlas perfectamente y se les colocó etiquetas adheribles con los datos de la colecta y
taxonómicos.
La identificación de los ejemplares colectados en el espárrago, fue por medio de claves
dicotómicas: de Krantz, 1986 para familia, las de Tuttle, Baker y Abbatiello, 1979, para género y
especies de tetraniquidos.
Resultados y Discusión
En base a los resultados se identificó a la araña carmín Tetranychus cinnabarinus
(Boisduval) perteneciente a la familia Tetranychidae como plaga del espárrago (Asparagus
officinalis L.) en Caborca, Sonora.
Este ácaro tiene diferentes sinonimias, Acarus cinnabarinus Boisduval, 1867, Tetranychus
telarius Smith & Baker, 1968; conocido comúnmente como ácaro rojo de los trópicos, arañita
roja carmín del clavel, arañita roja del algodonero, araña roja del clavel, araña roja de los cítricos,
arañuela roja común, araña roja del algodón, arañuela del frijol, arañuela roja común.
Esta especie se ha separado del ácaro de dos manchas T. urticae Koch por las diferencias
en morfología, hábitats, preferencias de hospederos, distribución geográfica y reproducción.
Ambos son perjudiciales para una amplia variedad de plantas en invernaderos y en el campo. T.
(T.) urticae, sin embargo, es con frecuencia una plaga seria de árboles frutales, caducifolios de
sombra y arbustos, especialmente en los climas templados, mientras que T. cinnabarinus, es más
común en climas semitropicales, pero su distribución coincide en parte (Jeppson, et al 1975). La
hembra adulta es de color rojo carmín (Fig. 1) de ahí su nombre de araña carmín, produce una

4
gran cantidad de telaraña que puede llegar a cubrir completamente el espárrago (Fig. 2a, b y c),
cuando la densidad de población es alta, es una especie partenogenética arrenotoca,
haplodiploide, polífaga y multivoltina, ya que tiene cerca de 100 plantas hospederas y su ciclo de
vida es de 10 a 12 días, por lo cual se presentan 20 generaciones por año y la hembra con una
longevidad de 20 días durante la cual oviposita 200 huevecillos, que son esféricos y ligeramente
rojizos (Fig.3) y requiere una temperatura óptima de 32 °C, no inverna como es el caso de
Tetranychus urticae Koch que lo hace como hembra. La hembra adulta mide aproximadamente
0.5 mm de largo, son de forma más o menos elíptica (Fig. 4). Estrías dorsales con lóbulos más
altos que anchos (Fig. 5); la seta táctil del tarso I está próxima a las setas dúplex. Empodio
dividido en tres pelos (Fig. 6). En la región anal (Fig. 7) presenta un par de setas paranales y dos
pares de setas anales. Los machos son ligeramente más pequeños y de forma triangular, con
coloración más amarilla (Fig. 8). Su tegumento es finamente más estriado, región anal con setas
normales (Fig.9). El edeago tiene una pequeña protuberancia redondeada; los ángulos son
similares (normalmente) la angulación anterior es redondeada y la posterior es más aguda (Fig.
10), dichas características concuerdan con las reportadas por Jeppson, et al 1975, Tuttle, Baker y
Abbatiello, 1979.
Fig. 1 Hembra adulta de Tetranychus cinnabarinus (Boisduval) de color carmín.
Fig. 2a Tallo con telaraña producida por T.
cinnabarinus.
b.- Tallo con T. cinnabarinus.
c.- Tetranychus cinnabarinus rojo carmín.

5
Fig. 4 Vista ventral de Tetranychus cinnabarinus
(Boisduval) y orificio genital de la hembra. Fig. 3 Huevecillos de Tetranychus
cinnabarinus
Fig. 5.- Estrías dorsales con lóbulos más altos que anchos.
Fig. 6.- Empodio de Tetranychus cinnabarinus
(Boisduval) dividido en tres pelos.
Fig. 7.- Setas anales (a) y paranales (b) de
Tetranychus cinnabarinus (Boisduval), vista
ventral.
a
b

6
Conclusión
Los resultados de esta investigación permiten concluir que la especie que ataca al
espárrago (Asparagus officinalis L.) en el municipio de Caborca, Sonora, fue identificada como
Tetranychus cinnabarinus (Boisduval), perteneciente a la Familia Tetranychidae, lo cual es de
gran importancia ya que con esta identificación puede evitarse el uso desmedido de agroquímicos
que se aplican erróneamente para su control, debido a que a menudo es confundido con el ácaro
de dos manchas Tetranychus urticae Koch.
Literatura Citada
Anaya R. S., Romero N. J. et al. 1999. Hortalizas: plagas y enfermedades. Ed. Trillas, México,
D.F. Pp. 281–284.
Benages, S. S. 1990. El espárrago. Ediciones Mundi-Prensa. Madrid. Pp. 36-41.
Jeepson, L. R.; Keifer, H. H. and Baker E. W. 1975. Mites injurious to economic plants.
University of California Press.UC, Riverside. 218-221 pp.
Fig. 9 Región genital del macho de Tetranychus
cinnabarinus (Boisduval), vista ventral.
Fig. 10.- Edeago de Tetranychus cinnabarinus
(Boisduval).
Fig. 8.-Vista ventral del macho de
Tetranychus cinnabarinus (Boisduval).

7
Tuttle, D.M.; Baker, E.W. and J. Abbatiello. 1974. Spider mites of México (Acari:
Tetranychidae). Intl. J. Acar. Vol. 2, No. 2 Smithsonian Institution Press. Washington,
U.S.A. 104 pp.
IDENTIFICACIÓN DE LOS ÁCAROS ASOCIADOS AL MAÍZ (ZEA MAYS L.) EN LA
COMARCA LAGUNERA.
Identification of the mites associated to the corn (Zea mays L.) in the Lagunera Region.
Benito Reséndiz García y Maria Guadalupe Aguillón Trejo. Departamento de Parasitología
agrícola. Universidad Autónoma Chapingo. Km. 38.5 carretera México-Texcoco. Chapingo,
Texcoco, Estado de México. C. P. 56230. Correo-e:bresendiz21@ hotmail.com
Palabras Clave: maíz, araña roja
Introducción
La Comarca Lagunera es una de las cuencas lecheras más importantes en el ámbito
nacional, con aproximadamente 214 mil cabezas de ganado bovino lechero en producción, que
producen 1.73 millones de litros de leche diarios. La magnitud de este sistema de producción
plantea la necesidad de estrategias para la producción de forraje para su manutención. El maíz se
ha seleccionado como un forraje de importancia, con una superficie de 17,834 ha y cuya
producción es de 712,286 tones a nivel nacional. La Comarca Lagunera donde se produce en
promedio 6 t/ha de grano y superan las 45 t/ha de forraje verde con manejo óptimo, sin embargo,
el potencial productivo del maíz en esta región es superior debido a la alta radiación solar durante
el período libre de heladas y es posible obtener hasta 80 t/ha de forraje fresco y 24 t/ha de forraje
seco (30% de materia seca). En los últimos años, los agricultores han observado daños en sus
cultivos causados por ácaros, principalmente el maíz. Ante esta situación han tratado de combatir
dicha plaga mediante el empleo de productos químicos, sin lograr un control satisfactorio,
desarrollando con ello resistencia a una amplia variedad de acaricidas. Por tal motivo es
importante conocer las especies de ácaros que se encuentran causando daños, para seleccionar la
mejor medida de control. Razón por la cual el objetivo de este trabajo fue: Identificar los ácaros
asociados al maíz (Zea mays L.) en la Comarca Lagunera.
Materiales y Método
La colecta del material biológico se llevó a cabo en parcelas de maíz en la Comarca
Lagunera, en los municipios de de Francisco I. Madero (Rancho 4 Hermanos y propiedad
Buitrón), Coahuila, de mayo a junio del 2008, para lo cual se seleccionaran plantas de maíz al
azar, revisando de 12 a 15 plantas desde hojas inferiores, medias y superiores que presenten los
síntomas típicos de ataques de ácaros como son: hojas cloróticas y la presencia de telarañas (Fig.
1).
El material colectado se colocó en bolsas de polietileno previamente humedecidas, las
cuales se trasladaron al laboratorio de Acaralogia del Departamento de Parasitología Agrícola de
la UACH, para su posterior revisión.

8
Fig. 1.- Hojas de maíz (Zea mays.) con daño y ejemplares de araña roja.
El material biológico colectado, se revisó hoja por hoja con la ayuda de un microscopio
estereoscópico, los especímenes encontrados se colocaron en una Siracusa que contenía una
solución de aclarado (lactófenol), donde permanecieron por un espacio de 10 a 15 minutos.
Posteriormente se hicieron los montajes permanentes de la siguiente forma: sobre un portaobjetos
de 75 x 25x 1 mm de tamaño, se colocó una gota de liquido de fitoseide en el centro del mismo, y
con ayuda de un pescador el ácaro se trasladó de la Siracusa al centro de la gota, procurando que
el espécimen quedará perfectamente en posición dorsal o ventral hacia arriba según fuera
requerido, en seguida se colocó un cubreobjetos de 6 x 6 mm sobre la gota, lentamente para así
evitar la formación de burbujas y que el ejemplar quedara con sus extremidades extendidas,
enseguida las preparaciones se colocaron en una plancha de calentamiento eléctrica con el fin de
aclarar aun más el ejemplar y eliminar las burbujas que se formaron durante dicho proceso, los
especímenes se examinaron en un microscopio compuesto para comprobar que el ejemplar
estuviera en perfectas condiciones. Las preparaciones se dejaron por un periodo de cinco días
para que se secaran, más tarde se sellaron con barniz transparente, así mismo por medio de un
círculo se marcó la ubicación del ejemplar para facilitar su búsqueda; finalmente se procedió a
limpiarlas perfectamente y se les colocó etiquetas adheribles con los datos de la colecta y
taxonómicos (Reséndiz, 1982).
La identificación de los ejemplares colectados en el maíz, fue por medio de claves
dicotómicas: de Krantz, 1986 para familia, las de Tuttle, Baker y Abbatiello, 1979, para género y
especies de tetraníquidos.
Resultados y Discusión
En base a los resultados se identifico a la especie Oligonychus pratensis (Banks)
perteneciente a la familia Tetranychidae como la principal plaga del maíz (Zea mays), en la
Comarca Lagunera. Conocido comúnmente como ácaro del pasto, ácaro del dátil, ácaro de las
bandas u orillas de los pastos. Este ácaro ataca las hojas y los daños se manifiestan por la
presencia de una fina telaraña, las hojas tienden a perder turgencia y presentan manchas
amarillas. Si el ataque es severo, hay clorosis total, necrosis y pérdida de crecimiento (Fig. 2).

9
Lo característico de este género es la modificación del empodio a una uña con pelos
cortos proximoventrales, esto se comparó con las características mencionadas por Tuttle et al.
(1974) y tanto el macho como la hembra presentan características similares.
La hembra es amarilla o verdosa, el gnatosoma y patas con tonos rojizos-anaranjados.
Mide aproximadamente de 0.42 mm, es de forma ovalada, el idiosoma con seis manchas oscuras
en dos filas laterales de tres cada una (Fig. 3).
Tarso I y II con dos pares de setas duplex aproximadas (Fig.4); los empodios I y IV
uncinados con solamente tres pares de pelos proximoventrales (Fig. 5). Dos pares de setas anales
y un par de para-anales (Fig. 6).
El macho adulto es ligeramente más pequeño que la hembra y de forma cónica, su
tegumento es finamente más estriado sus patas proporcionalmente alargadas y su color es más
pálido en comparación con la hembra.
El edeago del macho de O. Pratensis presenta en la parte terminal una curvatura hacia el
dorso que se encuentra casi en ángulo recto en relación con el Histerosoma, además presenta una
protuberancia (joroba) distal fuertemente sigmoidea, con una angulación anterior redondeada y
posterior más aguda (Fig. 7).
Fig. 2.- Síntomas y daños ocasionados por O. pratensis (Banks).
Fig. 3.- Hembra de O. pratensis y vista ventral

10
Fig. 4.- Tarso I con setas dobles. Fig. 5.- Tarso I con uña empodial y con pelos
proximoventrales.
Fig. 6.- Región ventral del Histerosoma. Fig. 7.- Edeago de O. pratensis (Banks).
Conclusión
La especie que ataca al maíz (Zea mays L.) en el municipio de Francisco I. Madero,
Región Laguna del estado de Coahuila, fue identificada como Oligonychus pratensis (Banks),
perteneciente a la Familia Tetranychidae, y conocido comúnmente como ácaro de los pastos.
Literatura Citada
Jeppson, L. R., Keifer, H. H. and Baker, E. W. 1975. Mites in jurious to economic plants.
University of California Press. pp. 212-215.
Krantz, G.W. 1986. A manual of acarology O.S.V. Book Stores. Inc. Corvallis, Oregon. pp. 295-
342.
Reséndiz, G. B. 1982. Nuevas técnicas para el montaje de ácaros de las Familias Veigaiidae,
Parasitidae y Macrochelidae. Tesis de Licenciatura. Universidad Autónoma de Chapingo.
Chapingo, México. pp. 6-11.

11
Tuttle, D. M; Baker, E. W. and M. J. Abbatiello. 1979. Spider mites of Mexico (Acari:
Tetranychidae). Intl. J. Acar. 2 (2): 224-423. Smithsonian Institution Press. Washington,
U.S.A.
ESTADO ACTUAL DE LA FAMILIA THERIDIIDAE SUNDEVALL, 1833 (ARACHNIDA:
ARANEAE) Y SU PRESENCIA EN MÉXICO
Current status of the family Theridiidae Sundevall, 1833 (Arachnida: Araneae) and its presence
in Mexico
César Gabriel Durán-Barrón. Colección Nacional de Arácnidos (CNAN), Departamento de
Zoología, Instituto de Biología, Universidad Nacional Autónoma de México. Ciudad
Universitaria, Apartado Postal 70-153, México, D. F., C. P. 04510. [email protected]
Palabras Clave: Arañas, Theridiidae, México
Introducción
La familia Theridiidae es considerada como una de las más grandes, diversas y
cosmopolitas; a nivel mundial esta conformada por 109 géneros y 2,293 especies (Platnick 2009),
por lo que ocupa el cuarto lugar mundial en diversidad de especies (Coddington y Levi 1991).
Actualmente en México esta familia esta representada por 32 géneros y 227 especies (Durán-
Barrón, 2004b). Los integrantes de esta familia son sedentarios (Levi, 2005), habitan encima o
debajo de rocas, sobre la vegetación, la hojarasca y la corteza de los árboles (Griswold et al.,
1998), debajo de estiércol seco de vaca (Durán-Barrón, obs pers), lugares sombríos, oquedades,
ranuras, grietas (Levi y Levi, 1962), sobre una variedad de telarañas (Palmer, 1961; Eberhard et
al., 2008) o pueden ser sinantrópicas (Robinson 1996, Durán-Barrón, 2004b, Dura-Barrón, 2007).
Además presenta casos de eusocialismo (Agnarsson, 2004; Agnarsson et. al., 2006),
kleptoparasitismo (Foelix, 1996; Agnarsson, 2002; Agnarsson, 2003a; Coddington y Agnarsson,
2006; Koh y Li, 2007), mirmecofagia (Agnarsson, 2004), alta diversidad de arquitecturas en la
red (Eberhard, 1991; Jörger y Eberhard, 2006; Eberhard et al., 2008) y una extrema diversidad
morfológica, ecológica y etológica (Agnarsson, 2004).
Desde su origen esta familia ha sido objeto de severos problemas, tanto taxonómicos
como filogenéticos. Levi y Levi (1962) realizaron la primera revisión y delimitación de la
familia, creando una clasificación más natural, además presentaron diagnosis completas de los
géneros válidos y una primera propuesta filogenética. Forster et al. (1990) reconocen la
monofilia de Hadrotarsinae y Spintharinae como subfamilias de Theridiidae. Griswold et al.
(1998) corroboran la monofilia del clado Theridiidae + Nesticidae, basado en 93 caracteres
morfológico-etológicos. Yoshida (2001) sugiere la unión de las subfamilias Spintharinae y
Moneteae con Theridiinae, basado en dos sinapomorfías putativas. Yoshida (2002) corrobora la
inclusión de la subfamilia Hadrotarsinae dentro de Theridiidae. Análisis morfológicos sugieren
que la familia Nesticidae es grupo hermano de Theridiidae y ambos, a su vez, son grupo
hermano de Cyatholipoidea (incluyendo a Synotaxidae) (Agnarsson, 2003b; Agnarsson, 2004).
En contraste, análisis moleculares sugieren resultados distintos, ya que Arnedo et al. (2004)
prueban la monofilia de la familia Theridiidae y la validez de las subfamilias y géneros
propuestos, mediante una filogenia molecular basada en 2,500 pares de bases de tres genes

12
nucleares y dos genes mitocondriales. Recientemente Agnarsson (2004) realiza un análisis
filogenético de 32 géneros, basado en morfología y comportamiento y propone que la familia
debería de estar conformada por siete subfamilias. En el World Spider Catalog (Platnick, 2009)
se presenta una pequeña reseña de los movimientos taxonómicos de la familia Theridiidae.
Materiales y Método Se llevo a cabo una revisión bibliográfica exhaustiva, con la finalidad de recabar la mayor
cantidad de información sobre la fauna de teridiidos mexicanos, con el objetivo de analizar el
estado del conocimiento de la familia Theridiidae en México. La información se obtuvo a partir
de la revisión de listados de especies incluidos en tesis de Licenciatura y Maestría, publicaciones
científicas especializadas (capítulos en libros y artículos) y trabajos en Congresos. Para obtener la
relación de géneros y especies de teridiidos mexicanos se tomo como base el listado de especies
publicado por Hoffmann (1976) y se cotejo con los trabajos de Durán-Barrón y Pérez-Ortiz
(2000), Durán-Barrón (2000, 2004a y 2006) y Desales-Lara, et al. (2008), para finalmente crear
el listado actualizado de géneros y especies que se incluye en el presente trabajo. La
corroboración del estatus taxonómico de las especies se realizó con ayuda del World Spider
Catalog (Platnick, 2009).
Resultados
Se encontró un total de 31 trabajos sobre teridiidos para México, agrupados en 6
categorías (Estudios faunísticos y de diversidad 33%, Taxonómicos 29%, Ecológicos 19%,
Venenos 10%, Listados de especies 6% y Biogeográficos 3%). De estos trabajos se tomaron las
especies nominales y se analizaron en conjunto con los trabajos antes citados, encontrándose que
el estado de Aguascalientes es el único que no presenta registro alguno de teridiidos; mientras
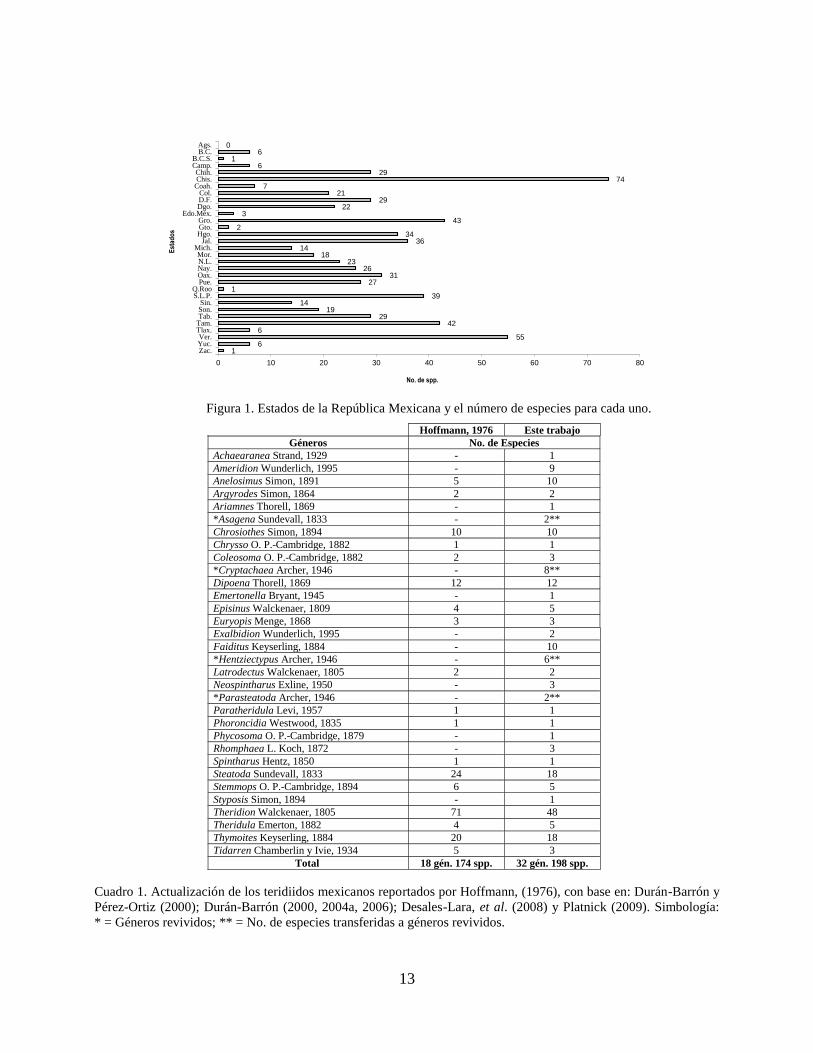
que el resto de los estados presentan como mínimo una especie y como máximo 74 (Fig. 1). Así
mismo, se cotejaron los cambios taxonómicos recientes y se encontró que 4 géneros fueron
revividos y 18 especies fueron reubicadas en otros géneros. También se registraron 7 especies sin
una distribución nacional específica y 32 endemismos (Durán-Barrón en prensa). En resumen los
resultados para la republica mexicana indican que actualmente existen 32 géneros y 198 especies
de teridiidos (Cuadro 1).
Discusión y Conclusiones Actualmente la taxonomía y sistemática de la familia Theridiidae sigue siendo sujeto de
cambios (Forster et al., 1990; Griswold et al., 1998; Yoshida, 2001, 2002; Agnarsson, 2003b,
2004; Arnedo et al., 2004; Platnick, 2009). Prueba de ello son los diferentes géneros revividos
por varios autores. Como por ejemplo el género Asagena que fue creado inicialmente por
Sundevall en 1833, pero Levi en 1957 lo sinonimizo con el género Steatoda y subsecuentemente
Wunderlich lo revive en el 2008 y transfiere un par de especies. Para el caso del género
Cryptachaea, este fue descrito inicialmente como un subgénero de Theridion Walckenaer, 1805,
pero elevado a nivel de género por Archer en 1950 y Yoshida en el 2008 transfiere ocho especies
a este género. El género Hentziectypus Archer, 1946, también es revivido por Yoshida en el 2008
y actualmente lo conforman seis especies trasferidas del género Achaearanea Strand, 1929. El
género Parasteatoda inicialmente se manejo como un subgénero de Theridion Walckenaer, 1805
y fue elevado a género por Archer en 1950 y actualmente se conforma de dos especies
transferidas del género Steatoda Sundevall, 1833.
13
Figura 1. Estados de la República Mexicana y el número de especies para cada uno.
Cuadro 1. Actualización de los teridiidos mexicanos reportados por Hoffmann, (1976), con base en: Durán-Barrón y
Pérez-Ortiz (2000); Durán-Barrón (2000, 2004a, 2006); Desales-Lara, et al. (2008) y Platnick (2009). Simbología:
* = Géneros revividos; ** = No. de especies transferidas a géneros revividos.
1 6
55 6
42 29
19 14
39 1
27 31
26 23
18 14
36 34
2 43
3 22
29 21
7 74
29 6
1 6
0
0 10 20 30 40 50 60 70 80
Zac.Yuc.Ver.
Tlax.Tam.Tab.Son.Sin.
S.L.P.Q.Roo
Pue.Oax.Nay.N.L.Mor.
Mich.Jal.
Hgo.Gto.Gro.
Edo.Méx.Dgo.D.F.Col.
Coah.Chis.Chih.
Camp.B.C.S.
B.C.Ags.
No. de spp.
Est
ado
s
Hoffmann, 1976 Este trabajo
Géneros No. de Especies
Achaearanea Strand, 1929 - 1
Ameridion Wunderlich, 1995 - 9
Anelosimus Simon, 1891 5 10
Argyrodes Simon, 1864 2 2
Ariamnes Thorell, 1869 - 1
*Asagena Sundevall, 1833 - 2**
Chrosiothes Simon, 1894 10 10
Chrysso O. P.-Cambridge, 1882 1 1
Coleosoma O. P.-Cambridge, 1882 2 3
*Cryptachaea Archer, 1946 - 8**
Dipoena Thorell, 1869 12 12
Emertonella Bryant, 1945 - 1
Episinus Walckenaer, 1809 4 5
Euryopis Menge, 1868 3 3
Exalbidion Wunderlich, 1995 - 2
Faiditus Keyserling, 1884 - 10
*Hentziectypus Archer, 1946 - 6**
Latrodectus Walckenaer, 1805 2 2
Neospintharus Exline, 1950 - 3
*Parasteatoda Archer, 1946 - 2**
Paratheridula Levi, 1957 1 1
Phoroncidia Westwood, 1835 1 1
Phycosoma O. P.-Cambridge, 1879 - 1
Rhomphaea L. Koch, 1872 - 3
Spintharus Hentz, 1850 1 1
Steatoda Sundevall, 1833 24 18
Stemmops O. P.-Cambridge, 1894 6 5
Styposis Simon, 1894 - 1
Theridion Walckenaer, 1805 71 48
Theridula Emerton, 1882 4 5
Thymoites Keyserling, 1884 20 18
Tidarren Chamberlin y Ivie, 1934 5 3
Total 18 gén. 174 spp. 32 gén. 198 spp.

14
En general la familia Theridiidae es poco estudiada en México y prueba de ello son los
escasos 31 trabajos de los que se tiene conocimiento; la razón principal estriba en la gran
dificultad taxonómica que representan (Durán-Barrón, 2004a) y la extrema diversidad
morfológica, ecológica y etológica (Agnarsson, 2004). Este desconocimiento se ve reflejado en
el número de géneros y especies que Hoffmann reporta para México (Cuadro 1). Sin embargo la
revisión de los trabajos de Durán-Barrón y Pérez-Ortiz (2000); Durán-Barrón (2000, 2004a y
2006) y Desales-Lara, et al. (2008) permitieron ampliar el panorama de los teridiidos mexicanos;
ya que se incremento en 14 el número de géneros (78%) y en 23 el número de especies (13%).
Agradecimientos
Al Programa de Doctorado en Ciencias del Posgrado en Ciencias Biológicas del Instituto
de Biología de la UNAM, por el apoyo económico otorgado. Al Dr. Oscar Francke y la M. en C.
Griselda Montiel-Parra, por los comentarios y correcciones al presente trabajo.
Literatura Citada Agnarsson, I. 2002. Sharing a web-on the relation of sociality and kleptoparasitism in Theridiid spiders
(Theridiidae, Araneae). The Journal of Arachnology, 30: 181-188.
Agnarsson, I. 2003a. Spider webs as habitat patches –the distribution of kleptoparasites (Argyrodes, heridiidae)
among host webs (Nephila, Tetragnathidae). The Journal of Arachnology, 31: 344-349.
Agnarsson, I. 2003b. The phylogenetic placement and circumscription of the genus Synotaxus (Araneae:
Synotaxidae) with a description of a new species from Guyana, and notes on theridioid phylogeny.
Invertebrate Systematics, 17: 719-734.
Agnarsson, I. 2004. Morphological phylogeny of cobweb spiders and their relatives (Araneae, Araneoidea,
Theridiidae). Zoological Journal of the Linnean Society, 141447-626.
Agnarsson, I., L. Avíles, J. A., Coddington, y W. P. Maddison. 2006. Sociality in Theridiid spiders: Repeated
origins of an evolutionary dead end. Evolution, 60 (11): 2342-2351.
Archer, A. F. 1950. A study of theridiid and mimetid spiders with descriptions of new genera and species.
Museum Paper Alabama Museum of Natural History, 30: 1-40.
Arnedo, M. A., J. Coddington, I. Aganarsson y R. Gillespi. 2004. From a comb to a tree: phylogenetic
relationships of the comb-footed spiders (Araneae, Theridiidae) inferred from nuclear and
mitochondrial genes. Molecular phylogenetics and evolution, 31: 225-245.
Coddington, J. A. y H. W. Levi. 1991. Systematics and evolution of spiders (Araneae). Annual Review of
Ecology and Systematics, 22 : 565–592.
Coddington, J. A. y I. Agnarsson. 2006. Subsociality in Helvibis thorelli Keyserling 1884 (Araneae,
Theridiidae, Theridiinae) from French Guiana. The Journal of Arachnology, 34: 642-645.
Desales-Lara, M. A., C. G., Durán-Barrón y C. E., Mulia. 2008. Nuevos registros de aranéidos y terídiidos
(Araneae: Araneidae, Theridiidae) del Edo. Méx. Dugesiana, 15 (1): 25-30.
Durán-Barrón, C. G. 2000. Estudio faunístico de la familia Theridiidae (Arachnida: Araneae); en la selva baja
caducifolia del sur de Jalisco (Mpio. “El Limón”), México. Tesis de Licenciatura, Escuela Nacional de
Estudios Profesionales Iztacala, Universidad Nacional Autónoma de México, 112 pp.
Durán-Barrón, C. G. 2004a. Arañas de la familia Theridiidae Sundevall 1833 (Arachnida: Araneae) de la
Estación de Biología Chamela. Pp. 1-14. En: Artrópodos de Chamela. (Alfonso N. García Aldrete y
Ricardo Ayala Barajas, eds.). Instituto de Biología, UNAM, México, 227 pp.
Durán-Barrón, C. G. 2004b. Diversidad de arañas (Arachnida: Araneae) asociadas a viviendas de la ciudad de
México (Área metropolitana). Tesis de Maestría en Ciencias Biológicas (Sistemática). Posgrado en
Ciencias Biológicas, Instituto de Biología, Universidad Nacional Autónoma de México, 229 pp.

15
Durán-Barrón, C. G. 2006. Arañas de la Familia Theridiidae (Arachnida: Araneae) asociadas a la selva
mediana perennifolia de Frontera Corozal, Ocosingo, Chiapas. Pp. 25-30. En: Entomología Mexicana,
Vol. 5 Tomo 1 (Estrada Venegas, E. G., J. Romero, A. Equihua, C. Luna León, J. L. Rosas Acevedo,
eds). Manzanillo; Colima, México. 634 pp.
Durán-Barrón, C. G. 2007. El sinantropismo en las arañas. Pp. 9-13. En: Entomología Mexicana, Vol. 6
Tomo 1, (Estrada Venegas, E. G., A. Equihua, C. Luna León, J. L. Rosas Acevedo, eds) Acapulco;
Guerrero, México. 728 pp.
Durán-Barrón, C. G. y T. M. Pérez-Ortiz. 2000. Actualización en el conocimiento de las arañas de la familia
Theridiidae (Arachnida: Araneae) en México. Pp. 580-586. En: Memorias del XXXV Congreso
Nacional de Entomología, (Stanford, C. S., A. Morales M., J. R. Padilla R., M. P. Ibarra G., eds).
Acapulco, Guerrero, 852 pp.
Eberhard, W. G. 1991. Chrosiothes tonala (Araneae, Theridiidae): A web-bulding spider specializing on
termites. Psyche, 98 (1): 7-20.
Eberhard, W. G., I. Agnarsson y H. W. Levi. 2008. Web forms and the phylogeny of theridiid spiders
(Araneae: Theridiidae): chaos from order. Systematics and Biodiversity, 6 (4): 415-475.
Foelix, R. F. 1996. Biology of spiders. Harvard University Press, New York, 306 pp.
Forster, R. R., N. I. Platnick y J. A. Coddington. 1990. A proposal and review of the spider family Synotaxidae
(Araneae, Araneoidea), with notes on theridiid interrelationships. Bulletin of the American Museum of
Natural History, 193: 1-106.
Griswold, C. E., J. A. Coddington, G. Hormiga y N. Scharff. 1998. Phylogeny of the orb-web building spiders
(Araneae, Orbiculariae: Deinopoidea, Araneoidea). Zoological Journal of the Linnean Society, 123 (1)
: 1-99.
Hoffmann, A. 1976. Relación bibliográfica preliminar de las arañas de México (Arachnida: Araneae).
Publicaciones Especiales No. 3. Instituto de Biología, Universidad Nacional Autónoma de México;
México, D. F., 117 pp.
Jörger, M. K. y W. G. Eberhard. 2006. Web construction and modification by Achaearanea tesselata (Araneae,
Theridiidae). The Journal of Arachnology, 34: 511-523.
Koh, T. H. y D. Li, D. 2007. State-dependent prey type preferentes of a kleptoparasitic spider Argyrodes
flavescens (Araneae: Theridiidae). The Zoological Society of London, 260: 227-233.
Levi, H. W. 1957. The spider genera Crustulina and Steatoda in North America, Central America, and the
West Indies (Araneae, Theridiidae). Bulletin of the Museum of Comparative Zoology. Harvard.
117: 367-424.
Levi, H. W. 2005. Theridiidae. P. 235 En: Ubick, D., P. Paquin, P. E. Cushing y V. Roth (eds.) Spiders of
North America: an identification manual. American Arachnological Society. 377 pp.
Levi, H. W. y L. R. Levi. 1962. The genera of spider family Theridiidae. Bulletin of the Museum of
Comparative Zoology 127 (1): 1-71.
Palmer, E. L. 1961. Spiders and webs: various types of spiders webs afford rich area of special study. Natural
History, 1-11.
Platnick, N. I. 2009. The world spider catalog, version 9.5. American Museum of Natural History, online at
http://research.amnh.org/entomology/spiders/catalog/index.html
Robinson, W. H. 1996. Spiders and bugs. P. 205-229. En: Urban Entomology (insects and mites pests in the
human environment). Chapman & Hall (Eds.). 430 pp.
Ubick, D., P. Paquin, P. E. Cushing y V. Roth. 2005. Spiders of North America: an identification manual.
American Arachnological Society, 337 pp.
Wunderlich, J. 2008. On extant and fossil (Eocene) European comb-footed spiders (Araneae: Theridiidae),
with notes on their subfamilies, and with descriptions of new taxa. Beiträge zur Araneologie 5: 140-
469.
Yoshida, H. 2001. The genus Rhomphaea (Aranea: Theridiidae) from Japan, with notes on the subfamily
Argyrodinae. Acta Arachnologica, 50 (2): 183-192.
Yoshida, H. 2002. A revision of the Japanese genera and species of the subfamily Hadrotarsinae (Araneae:
Theridiidae). Acta Arachnologica, 51 (1): 7-18.

16
Yoshida, H. 2008. A revision of the genus Achaearanea (Araneae: Theridiidae). Acta Arachnologica of Tokyo
57: 37-40.
DIVERSIDAD EN LA COMUNIDAD DE ÁCAROS EN DOS SITIOS DE LA RESERVA
ECOLÓGICA DEL PEDREGAL DE SAN ÁNGEL, DISTRITO FEDERAL, MÉXICO
Mites diversity in two sites of Pedregal de San Ángel Ecological Reserve, Distrito Federal,
México
Daniela Pérez-Velázquez1 y Alicia Callejas-Chavero
1,2.
1Ecología y Sistemática de
Microartrópodos, Departamento de Ecología y Recursos Naturales, Facultad de Ciencias,
Universidad Nacional Autónoma de México, Coyoacán 04510, México, D. F., 2
Laboratorio de
Ecología Vegetal, Departamento de Botánica, Escuela Nacional de Ciencias Biológicas, Instituto
Politécnico Nacional, Prolongación de Carpio y Plan de Ayala, Casco de Santo Tomás, Miguel
Hidalgo 11340, México, D.F. E: mail: [email protected], [email protected]
Palabras Clave: Suelo, Mesostigmata, Prostigmata, Astigmata, Cryptostigmata, variación
espacial.
Introducción Dentro de los microartrópodos del suelo, los ácaros representan un grupo de gran
diversidad y abundancia, cuya función repercute de manera esencial en diversos procesos
biogeoquímicos del mismo (Heneghan et al., 1998, 1999). Se considera, por ejemplo, que los
ácaros sinergizan las actividades de los microorganismos descomponedores y, en particular,
participan en el desmenuzamiento de la materia orgánica más compleja y difícil de romper así
como en el movimiento de los nutrimentos y microorganismos en los estratos del suelo (Estrada-
Venegas, 2008). Por su diversidad de hábitos alimenticios, participan en distintos niveles de las
cadenas tróficas, tanto como detritívoros, actuando en los procesos de descomposición e
integración de la materia orgánica al suelo, como depredadores activos de distintos grupos de
invertebrados (Coleman, et al., 2004).
Debido a ello, son un grupo estrechamente relacionado con los procesos del suelo, por lo
que cambios en este ecosistema se ven reflejados en diversos parámetros de las comunidades de
ácaros, lo que ha sido utilizado para evaluar la alteración y de calidad del suelo (Parisi et al.
2005; Van Straalen, 1998). Diversas especies de ácaros, principalmente oribátidos,
mesostigamados y astigmados, han sido propuestas como bioindicadores en suelos agrícolas y
forestales (Behan-Pelletier, 1999; Benton et al, 2004; Cameron & Benton, 2004; Koehler, 1999;
Ruf, 1998).
La Reserva Ecológica del Pedregal de San Ángel (REPSA) se ubica al Sureste de la
Ciudad de México (19º 14’ - 19º 25’ N - 99º 08’ - 99º 15’ W), dentro del campus central de la
Universidad Nacional Autónoma de México (UNAM), y abarca aproximadamente 232.3 ha,
donde se encuentra una alta diversidad de plantas y animales con un gran número de endemismos
(Valiente-Baunet y Luna, 1994). La comunidad vegetacional más extendida y característica del
Pedregal, es la que Rzedowski (1994) denominó Senecionetum praecosis, misma que está
constituida por un matorral abierto donde están representados los estrato arbustivo, herbáceo y
rasante, sin que exista un estrato arbóreo como tal.

17
Esta área fue declara zona protegida en 1983 con le objeto de preservar la vegetación
endémica de la zona y su extraordinaria biodiversidad. Desafortunadamente, el rápido
crecimiento urbano ha ocasionado una reducción de sus dimensiones, así como un cambio
importante en la biodiversidad del sitio. Los cambios más relevantes han consistido
principalmente, en una disminución en el número de especies nativas y un aumento de especies
introducidas con hábitos ruderales y asociadas a sitios con alta frecuencia de perturbaciones
(Rzedowski, 1994; Valiente-Baunet y Luna, 1994). Originalmente la comunidad de Senecionetum
praecosis ocupaba cerca de 40 Km², lo cual representaba un 50% de la superficie total que resultó
del derrame de lava del Xitle. En la actualidad, como resultado del avance en los procesos
urbanos, ésta se ha reducido a sólo 2.9 Km², lo que representa una pérdida del 90% del área
original (Álvarez et al., 1994).
En el presente trabajo se realiza la comparación entre dos sitos con distinto grado de
alteración dentro de la REPSA, estudiando el efecto de tal alteración en la diversidad y
composición de la comunidad de ácaros edáficos, a fin de determinar su papel como indicadores
de dicha alteración.
Materiales y Método.
Los suelos de la REPSA son principalmente de origen eólico y orgánico, la acumulación
ocurre en grietas, fisuras y depresiones del terreno, su espesor es de pocos centímetros, por lo que
es difícil distinguir horizontes edafológicos típicos. Son suelos arenosos, limosos,
moderadamente ácidos; poseen gran cantidad de materia orgánica, de potasio y de calcio y son
pobres en nitrógeno y fósforo aprovechables (Rzedowski, 1994). La temperatura media anual es
de 14-15.5 ˚C y presenta variaciones extremas que van de 6 a 34.6 ˚C, mientras que la
precipitación fluctúa entre 700 y 950 mm por año (Valiente- Baunet y Luna,1994). El clima,
según la clasificación de Köppen, es templado sub-húmedo con régimen de lluvias de verano
(Soberón et al., 1994).
En el área del Jardín Botánico Exterior, se seleccionaron dos sitios con distinto grado de
alteración, uno denominado Cerrado, en donde la cobertura vegetal es mayor y el acceso al área
es más limitado, y otro denominado Abierto, ubicado a la orilla del camino frecuentemente usado
por los visitantes. En cada uno de los sitios se seleccionaron al azar tres árboles de Senecio
praecox, con una altura superior a 1.50. m En cada uno de ellos se tomaron muestras de suelo de
95 cm2 a 10 cm del tronco, en dirección norte, sur, este y oeste, las cuales fueron colocadas en
recipientes de plástico (10x10 cm). La fauna se extrajo mediante embudos de Berlesse–Tullgren
durante seis días (tres a temperatura ambiente y tres días con un foco de 25 watts). Una vez
obtenida la fauna, los ácaros fueron separados y cuantificados a nivel de Órdenes y
morfoespecies con ayuda de un microscopio estereoscópico.

18
Fig. 1. Sitio Cerrado y Abierto en la Reserva Ecológica del Pedregal de San Ángel.
Como parámetros para conocer la estructura de la comunidad de los ácaros, se calculó la
diversidad (con el inverso del índice de Simpson) y la Equitatividad, con la finalidad de conocer
la variación especial de la comunidad de ácaros durante el mes de Abril en ambos sitios. También
se calculó el coeficiente de Similitud de Sörensen para determinar que tan parecidas o diferentes
son entre si la comunidad de ácaros en ambos lugares.
Resultados y Discusión
El total de ácaros obtenidos de las muestras de suelo fueron 1,871 individuos, de los
cuales 1,030 corresponden al Sitio Abierto, donde se encontraros 27 morfoespecies diferentes (11
de Prostigmata con 629 individuos, 6 de Cryptostigmata con 166 individuos, 2 de Astigmata con
19 individuos y 8 de Mesostigmata con 216 individuos), y 841 al Sitio Cerrado en un total de 26
morfoespecies (Prostigmata: 12 con 754 individuos; Cryptostigmata: 9 con 36 individuos ;
Astigmata 1 con 9 individuos; Mesostigmata: 4 con 42). El número de morfoespecies
compartidas por cada Órden respecto a ambos sitios es el siguiente: Prostigmata, 11 de 12
morfos, Cryptostigmata, 6 de 9 morfos, Astigmata, 1 de 2 morfos y Mesostigmata, 3 de 9 morfos.
El mayor porcentaje de individuos encontrados pertenecen al orden Prostigmata, y los
menos representados pertenecen al Orden Astigmata, mientras que los Mesostigmata y
Cryptostigmata se encuentran en mayor abundancia en el Sitio Abierto (Fig. 2.).
Fig. 2. Representación en porcentajes de los individuos obtenidos de cada Órden por sitio: A. Abierto y B. Cerrado.
De acuerdo con lo obtenido con el inverso del índice de Simpson para ambos Sitios
(Cuadro 1) se obtuvo que el sitio Abierto resulto ser más diverso que el Cerrado, en cuanto a la
Equitativilidad, encontramos que la distribución de las morfoespecies es más homogénea en el
sitio Cerrado, y heterogénea en el sitio Abierto. La similitud entre sitios, de acuerdo con el
coeficiente de Sörensen fue de 0.79; lo que significa que ambos sitios comparten el 79% de las
morfoespecies encontradas.
Cuadro 1. Índice de Diversidad de Simpson por Sitio.
Sitio Índice de Diversidad Equitatividad
Abierto 8.43 0.12
Cerrado 3.86 0.26
Numero de
individuos,
Prosti…
Numero de
individuos,
Crypt…
Numero de
individuos,
Astig…
Numero de
individuos,
Meso…
A
Numero de
individuos,
Prosti…
Numero de
individuos,
Crypt…
Numero de
individuos,
Astig…
Numero de
individuos,
Meso…
B

19
Estudios previos han demostrado que Cryptostigmata constituye el grupo más diverso y
abundante en el horizonte orgánico del suelo, seguido de Prostigmata y Mesostigmata, mientras
que los Astigmata son poco comunes (Vásquez. 2007). De acuerdo con los resultados obtenidos
en nuestro trabajo, los individuos del Orden Prostigmata se encontraron en mayor abundancia y
con mayor cantidad de morfoespecies, difiere de la afirmación de que los Cryptostigmata son los
más diversos tal vez por que este estudio se realizó en la época de secas en la Reserva del
Pedregal de San Ángel, correspondiente al mes de Abril, aunque si coincide con lo que menciona
Sánchez-Rocha (2008), que dentro del grupo de los ácaros, los Prostigmados son los más
abundantes en diversidad como en morfología, y que se encuentran en mayor cantidad en suelos
con poca humedad, lo cual corresponde con las circunstancias del medio abiótico en el mes de
colecta, además de la escasa materia orgánica acumulada en el suelo de la reserva, el que es de
origen volcánico y por lo tanto dificulta la abundancia de los criptostigmados; así mismo los
Mesostigmata ocupan el tercer lugar en cuanto a número de individuos y morfoespecies y los
Astigmata son los menos comunes con apenas tres morfoespecies y 28 individuos, lo que
representa sólo el 2% del total obtenido.
Se sabe también que los patrones climáticos influyen grandemente en la descomposición y
movimiento de los nutrientes, lo cual afecta directamente las fluctuaciones estacionales de las
densidades de prostigmados, lo mismo que de mesostigmados y criptostigmados (Sánchez-Rocha
2008).
En cuanto a la diversidad encontrada, al comparar los dos sitios de muestreo, se observó
que el sitio más diverso es el Abierto, aunque con una distribución menos equitativa de las
morfoespecies, mientras que para el sitio Cerrado, la diversidad fue menor, pero con una
distribución más uniforme de los individuos, con lo cual podemos concluir que el grado de
perturbación influye sobre la diversidad y la estructura de la comunidad de ácaros en el suelo. Al
calcular el coeficiente de similitud de Sörensen, encontramos que el 79% de las morfoespecies
están presentes en ambos lugares, lo que nos dice que la composición en morfoespecies es muy
parecida entre ellos, lo cual puede estar relacionado con la temporada de muestreo (época de
secas) mas que con el grado de perturbación del sitio.
Conclusiones.
Se obtuvieron 1,871 ácaros, de los cuales 1,383 son Prostigmata, 202 Cryptostigmata, 28
Astigmata y 58 Mesostigmata, con un total de 27 morfoespecies.
El sitio Abierto tiene mayor diversidad y una distribución menos equitativa de los
individuos que el Sitio Cerrado.Ambos sitios comparten el 79% de las morfoespecies, lo que las
hace muy similares en su composición.
Agradecimientos.
El trabajo de campo se realizó gracias a la ayuda de Leopoldo Cutz-Pool, Miguel Blanco
Becerril, Arturo García Gómez y Sandra Gómez Acevedo. Se agradece a la Dra. Gabriela
Castaño Meneses, por su apoyo en el análisis estadístico y la estructuración y revisión de este
trabajo, así como al Dr. José G. Palacios-Vargas por la revisión del escrito. Este estudio es parte
del proyecto PAPIIT-IN208508, financiado por la Dirección General de Asuntos del Personal
Académico, UNAM

20
Literatura Citada Alvarez, F., J. Carabias., J. Meave., P. Moreno ., D. Nava., C.Tovar y A Valiente–Baunet. 1994. Proyecto
para la creación de una reserva en el Pedregal de San Angel. Reserva Ecológica “el Pedregal de
San Angel”. Ecología e historia natural y manejo (ed. A. Rojo) Universidad Nacional Autónoma
de México. México, D.F.
Behan-Pelletier, V.M. 1999. Oribatid mite biodivesity in agroecoystems: role for bioindication.
Agriculture, Ecosystems & Environment, 74: 411-423.
Benton, T.G., T.C. Camaron & A. Grant. 2004. Population responses to perturbations: perdictions and
responses from laboratory mite populations. Journal of Animal Ecology, 73: 983-995.
Cameron, T.C. & T.G. Benton. 2004. Stage-structured harvesting and its effects: an empirical
investigation using soil mites. Journal of Animal Ecology, 73: 996-1006.
Coleman, D.C., D.A. Jr. Crossley & P.F. Hendrix. 2004. Fundamentals of soil ecology. 2nd
edition. Elsevier Academia Press, Nueva York. 386 pp.
Estrada-Venegas G. & A.R. Moldenke. 2008. Importancia del Suelo como Ecosistema. Tomado
de Fauna del Suelo I: micro, meso y macrofauna. Colegio de Postgraduados.
Heneghan, L., D.C.Coleman, X. Zou, D.A. Jr. Crossley & B.L. Haines. 1998. Soil microar-
thropod community structure and litter decomposition dynamics: a study of tropical and
temperate sites. Applied Soil Ecology 9: 33–38.
Heneghan, L., D.C. Coleman, X. Zou, D.A. Jr. Crossley & B.L. Haines. 1999. Soil micro-
arthropods contributions to decomposition dynamics: tropical and temperate comparisons
of a single substrate (Quercus prinus L.). Ecology 80: 1873–1882.
Koehler, H.H. 1999. Predatory mites (Gamasina, Mesostigmata). Agriculture, Ecosystems &
Environment, 74: 395-410. Parisi, V., C. Menta, C. Gardi, C. Jacomini & E. Mozzanica. 2005. Microarthropod communities as a tool
to assess soil quality and biodiversity: a new approach in Italy. Agriculture, Ecosystems and
Environment, 105: 323-333.
Ruf, A. 1998. A maturity index for predatory soil mites (Mesostigmata: Gamasina) as an indicator of
environmental impacts of pollution on forest soils. Applied Soil Ecology, 9: 447-452.
Rzedowski, J. 1994. Vegetación del Pedregal de San Angel. Reserva ecológica “El Pedregal de San
Angel”. Ecología e historia natural y manejo (ed. A. Rojo) Universidad Nacional Autónoma de
México. México, D.F.
Sánchez-Rocha, I. 2008. Estudio de los Ácaros Prostigmata del suelo de México. Tomado de
Fauna del Suelo I: micro, meso y macrofauna. Colegio de Postgraduados. Soberón, J., M. Rosas y G. Jiménez. 1994. Ecología hipotética de la reserva del Pedregal de San Angel.
Reserva ecológica “El Pedregal de San Angel”. Ecología e historia natural y manejo (ed. A. Rojo)
Universidad Nacional Autónoma de México. México, D.F.
Valiente-Baunet, A y E. Luna. 1994. Una lista florística para la reserva del Pedregal de San Angel.
Reserva ecológica “El Pedregal de San Angel”. Ecología e historia natural y manejo (ed. A. Rojo)
Universidad Nacional Autónoma de México. México, D.F.
Van Straalen, N.M. 1998. Evaluation of bioindicador systems derived from soil arthropod communities.
Applied Soil Ecology, 9: 429-437.
Vásquez, C., C. Sánchez & N. Valera. 2007. Diversidad de ácaros (Acari: Prostigmata, Mesostigmata,,
Astigmata) asociados a la hojarasca de formaciones vegetales del parque universitario de la
UCLA, Venezuela. Iheringia, Sér. Zool. vol.97 no.4 Porto Alegre Dec.

21
EVALUACION DE CUATRO ACARICIDAS PARA EL CONTROL DE LA ARAÑA DE DOS
MANCHAS (Tetranychus urticae KOCH) (ACARI: TETRANYCHIDAE)
Four acaricides evaluation for the control two spotted mite (Tetranychus urticae Koch) (Acari:
Tetranychidae)
Ernesto Cerna1, Yisa Ochoa
2, Rigoberto Jiménez
3, Santiago Pérez
3, Carlos Ail
3 y Omar García
3
1Universidad Autónoma Agraria Antonio Narro. Buenavista, Saltillo, Coahuila. CP 25315.
2Universidad Autónoma de Aguascalientes. Posta Zootécnica. Aguascalientes. CP 25530.
3Estudiantes
de Postgrado de la Universidad Autónoma Agraria Antonio Narro. Buenavista,
Saltillo, Coahuila. CP 25315. [email protected]
Palabras Clave: Acaricidas, CL50, Grupos Toxicológicos
Introducción
Los ácaros juegan un papel importante en la vida del hombre, ya que las plantas
cultivadas se ven fuertemente atacadas por especies pertenecientes a la familia Tetranychidae; tal
es el caso de la especie Tetranychus urticae Koch, por su alto potencial reproductivo, que en
poco tiempo puede rebasar el umbral económico si no se toman medidas de control pertinentes,
ocasionando la muerte de las plantas por secado del follaje (Goodwin et al. 1995).
Una de las herramientas más utilizadas para el control de ésta arañita es el control
químico, por ser una herramienta barata y de fácil manejo, dando buenos resultados en las
primeras aplicaciones, lo que permite que se tenga una mayor producción por unidad de
superficie (Gould, 1987). Sin embargo, la importancia radica en el uso inapropiado de acaricidas
sintéticos para su control, lo que ha ocasionado un elevado desarrollo de la resistencia (Granham
y Helle, 1985).
El uso irracional de agroquímicos presenta desventajas, como la destrucción de la fauna
silvestre y la inducción de la resistencia a la mayoría de los productos utilizados al paso del
tiempo (Jeppson et al., 1975). T. urticae es una de las especies que más casos de resistencia a
presentado, debido al uso irracional de acaricidas con el afán de obtener mejores utilidades
(Cone, 1979).
El manejo deficiente de acaricidas contra T. urticae data de más de 70 años, iniciando con
los primeros registros en la década de los 30’s (Georghiou y Saito, 1983).
En años anteriores, poblaciones de T. urticae muestran resistencia a acaricidas más
recientes como es el caso de las avermectinas (Campos et al., 1995). Considerando entonces, que
desde la década de los 50’s y hasta finales de los 80’s, la rotación de acaricidas ha sido mínima.
Ante esta circunstancia y por el grado de importancia que presenta esta plaga, es necesario
conocer el nivel de tolerancia o resistencia a los acaricidas, lo que obliga a realizar estudios
toxicológicos para determinar líneas de respuesta dosis-mortalidad, y con ello establecer cuáles
son los productos más eficientes y poder realizar un mejor manejo. Por lo que el objetivo del
presente estudio fue determinar la CL50 de cuatro acaricidas pertenecientes a diferente grupo
toxicológico en el ácaro de dos manchas T. urticae.

22
Materiales y Método Para el desarrollo de esta investigación fue necesario realizar una serie de colectas de T.
urticae en invernaderos comerciales del estado de Guanajuato. El material recolectado se
trasportó al laboratorio de acarología de la Universidad Autónoma Agraria Antonio Narro y se
multiplicó en plántulas de frijol a temperatura de 25 + 2 °C, de 60 a 70 % de humedad relativa y
en condiciones de 12:12 luz: oscuridad, respectivamente, e inmediatamente se procedió al
desarrollo de los bioensayos.
Los bioensayos se realizaron de acuerdo con la técnica de inmersión en hoja (FAO, 1979),
para ello se seleccionaron foliolos de frijol con al menos 30 ácaros adultos hembra por
concentración, los foliolos tratados se depositaron sobre esponjas saturadas de agua en charolas
de plástico (Abou-Setta, 1987). Los acaricidas utilizados fueron; Avermectina (Agrimec CE 1.8
%), Dicofol (AK 20 CE 18.5 %), Oxido de fenbutatin (Torque 500 SC 44.64 %) y Naled (Naled
60 CE 60 %). Para la preparación de las diferentes concentraciones se utilizó agua destilada y el
producto Bionex como dispersante, en una proporción 1mL: 1L de agua; el intervalo de
concentraciones utilizadas fue de 500 ppm a 6500 ppm excepto para la Avermectina que osciló
de 0.5 ppm a 8.0 ppm; Para el testigo solo se usó agua destilada con adherente. Se tomó como
criterio de muerte la inmovilidad total, síntomas de ataxia o el desplazamiento menor al tamaño
de su cuerpo al recibir un estímulo. Las lecturas de mortalidad se realizaron a las 24 h excepto
para la Avermectina que se obtuvo a las 48 h.
Los resultados obtenidos de mortalidad se corrigieron con la formula de Abbott (1925), y
se realizó un análisis Probit, mediante el método de máxima verosimilitud (Finney, 1971).
Utilizando el programa SAS system for Windows ver 9.0 (2002).
Resultados y Discusión
En el cuadro 1, se muestran los resultados obtenidos de la respuesta de la población
colectada de T. urticae, en relación a cuatro acaricidas de diferente grupo toxicológico. Como se
puede observar la CL50 fue de 1.8, 1109.5, 52.9, 1014.7 ppm para los acaricidas Avermectina,
Dicofol, Oxido de Fenbutatin y Naled respectivamente.
Cuadro 1. Concentración letal y limites fiduciales de acaricidas aplicados a hembras adultas de Tetranychus urticae
Koch.
ppm
Acaricida n CL50 Límites fiduciales 95% CL95 g.l.
Avermectina 760 1.8 (1.634 – 2.083) 15.28 4
Dicofol 952 1109.5 (1008.56-1215.11) 3493.32 4
Oxido de
fenbutatin
769 52.9 (31.362 – 55.433) 143.65 4
Naled 816 1014.7 (911.18–1569.25) 3529.65 4 n: Número de hembras adultas de T. urticae, g.l.: Grados de libertad
La CL50 registrada para la Avermectina (1.8 ppm) en esta investigación es superior a la
reportada para otros trabajos, Campos et al. (1995) reportan para líneas de campo provenientes de

23
ornamentales una CL50 para Avermectina que van de 0.5 ppm a 8.8 ppm; mientras que James
(2000) reporta una CL50 para líneas de campo sobre lúpulo de 2.5 a 5 ppm. Lo anterior nos
muestra que al comparar los resultados de otras líneas de campo, con los obtenidos en nuestra
investigación, no presenta elevados niveles de tolerancia; sin embargo al realizar la comparación
con una línea susceptible, encontramos que, James (2000) reporta una CL50 de 0.019 ppm para la
línea susceptible Kennewick-s, lo que muestra que nuestra población en estudio tiene 94 veces
más tolerancia que esta. Al respecto Lagunes y Villanueva (1994), mencionan que al comparar
una línea susceptible y una de campo, existen problemas de resistencia, si la población de campo
presenta una tolerancia 10 veces mayor que la susceptible. La razón de encontrar una alta
tolerancia a la Avermectina se debe al número repetido de aplicaciones, de acuerdo a información
de los técnicos de los invernaderos donde se realizó la recolecta, mencionando que al menos se
realizan de 3 a 6 aplicaciones por temporada. Así mismo, Campos et al. (1995) mencionan que en
poblaciones de araña de dos manchas donde se realizan dos aplicaciones por temporada la
tolerancia permanece estable, sin embargo en poblaciones donde se realizan 12 o más
aplicaciones por temporada, la tolerancia se puede incrementar hasta 150 veces.
Para el caso del Dicofol (1109.5 ppm), los resultados son menores a los reportados por
Cotero et al. (1989) quienes reportan una CL50 de 2804 ppm para una línea de campo sobre
ornamentales, y Dennehy et al. (1984) determinaron una CL50 de 8590 ppm para una línea de
campo sobre algodón. Al comparar nuestros resultados con una línea susceptible para Dicofol,
Cerna et al. (2005), reportan una CL50 para una línea susceptible a Dicofol de 177 ppm, lo que
nos arroja una proporción de tolerancia de 6.28 veces.
En lo que respecta al acaricida Oxido de fenbutatin, nuestra línea muestra una CL50 de
52.9 ppm. Estos resultados son menores a los reportados por Tian et al. (1992), quienes indican
una CL50 para una línea de campo sobre perales de 1177 ppm. Al comparar nuestros resultados
con una línea susceptible para el Oxido de fenbutatin, encontramos que el resultado obtenido en
nuestra investigación, fue superior al comportamiento de una línea reportada por Tian et al.
(1992) con una CL50 de 30 ppm, con una proporción de resistencia de 1.76.
Finalmente para el acaricida Naled (1014.7) nuestros resultados son mayores a los
reportador por Sato et al. (2000) quienes muestran una CL50 de 586 ppm; al comparar nuestros
resultados con una línea susceptible reportada por los mismos autores determinaron una CL50 de
137 ppm, con una proporción de resistencia de 7.40 veces. Por otro lado, podemos mencionar que
la razón de encontrar niveles por debajo del umbral (10 veces) para los productos Dicofol, Oxido
de fenbutain y Naled, se debe a la poca aplicación de productos organoclorados (grupo
toxicológico al que pertenece el Dicofol) y a los organoestanosos (grupo al que pertenece el
Oxido de fenbutatin). Sin embargo, el acaricida Naled, que está por debajo del umbral es el más
próximo a superarlo, debido a que la aplicación de productos fosforados son más comunes para el
control de plagas en invernadero.
En el cuadro 2 se presentan los coeficientes de determinación (r2), chi-cuadrada (x
2),
pendiente y probabilidad, para líneas de regresión dosis/mortalidad para los acaricidas
Avermectina, Dicofol, Oxido de fenbutatin y Naled. Donde se puede observar que los valores
estimados para r2
oscilan entre 0.90 y 0.98. Estos resultados de acuerdo a Romahn et al. (1994)
indican que se obtuvo una correlación alta; así mismo el bajo valor de chi-cuadrada (x2) obtenido
en esta investigación, indica poca separación entre los puntos y la línea final de la dosis-
mortalidad observada, por tal motivo, los valores de probabilidad son altos. Por último, se puede
observar que los valores de la pendiente bajos; lo cual nos indica que la población en estudio,

24
presenta poca variabilidad genética en relación a la respuesta con los acaricidas (Lagunes y
Villanueva, 1994). Cuadro 2.- Coeficientes de determinación (r
2), chi-cuadrada (x
2), pendiente y probabilidad de ocurrencia del evento
de los diferentes acaricidas en hembras adultas de Tetranychus urticae.
Acaricida r2
x2
Pendiente Probabilidad
Avermectina 0.96 0.027 1.424 + 0.671 0.99
Dicofol 0.98 0.092 2.209 + 1.108 0.99
Oxido de fenbutatin 0.90 0.025 2.625 + 0.983 0.99
Naled 0.95 0.064 2.016 + 0.824 0.99
En la figura 1 se presentan las líneas de respuesta dosis-mortalidad, en referencia a la
recta correspondiente los acaricidas Avermectina, Dicofol, Oxido de fenbutatin y Naled. Donde
se obtuvo una CL50 de 1.8, 1109.5, 52.9, 1014.7 ppm respectivamente, así como sus límites
fiduciales.
Figura 1. - Representación grafica de límites fiduciales obtenidos a nivel de CL50 de hembras adultas de Tetranychus
urticae expuestas a los diferentes acaricidas.
Al respecto, podemos mencionar que los acaricidas Avermectina y Oxido de fenbutatin
estadísticamente son diferentes, al presentar limites fiduciales diferentes; sin embargo, para los
acaricidas Dicofol y Naled podemos mencionar que estadísticamente presentan la misma
respuesta sobre las hembras adultas de T. urticae de nuestro estudio, debido a que los limites
fiduciales de estos productos se traslapan. Georghiuo y Lagunes (1991) mencionan que los
límites fiduciales son un parámetro importante para relacionar los insecticidas que presentan
diferentes valores de CL50 pero similar limite fiducial.
Conclusiones La arañita de dos manchas presentó una alta tolerancia a la Avermectina y en menor
proporción a Oxido de fenbutatin.
El producto Naled presentó valores por debajo del umbral de resistencia sin embargo, se
considera importante hacer rotación de productos fosforados para evitar un incremento en la
tolerancia. Debido a que son productos baratos, de amplio espectro y de media vida residual, por
lo que son los más utilizados por los productores.
95
5
50
M
o
r t a
l i d
a
d
0.1 1 10 100 1000 10000
A
D N O
A: Avermectina D: Dicofol O: Oxido de fenbutatin
N: Naled
: Límites fiduciales
DOSIS ppm

25
El producto Dicofol presentó también valores por debajo del umbral, sin embargo estos
productos organoclorados, están restringidos en su uso en México, por lo que es difícil
conseguirlos. Así como ser muy dañinos para el hombre y el ambiente.
Literatura Citada Aboott, W. S. 1925. A method for computing the effectiveness of an insecticide. J. Econ. Entomol. 18:
265-267.
Abou-Setta, M. M.; Childers, C. C. 1987. A modified leaf arena technique for rearing phytoseiid or
tetranychid mites for biological studies. Florida Entomol. 70: 245-248.
Campos, F.; Dybas, R.A.;Krupa, D.A. 1995. Susceptibility of Tetranychus urticae (Acari: Tetranychidae)
populations in California to abamectin. J. Econ. Entomol. 88(2) : 225-231.
Cerna E., J. Landeros, E. Guerrero, A. E. Flores y M. H. Badii. 2005. Detección de resistencia enzimática
por productos sinergistas en una línea de campo de Tetranychus urticae Koch (Acari:
Tetranychidae). Folia Entomol. Mex. 44 (3): 287-295.
Cone, W. W. 1979. Pheromones of Tetranychidae. In Recents advances of Acarology. Vol. II. G.J.
Rodriguez, ed. Academic Press Inc. New York.
Cotero E. S.; Sánchez, G. M. 1989. Niveles de susceptibilidad de Tetranychus urticae Koch ( Acarida:
Tetranychidae) a ocho acaricidas en el cultivo del clavel (Dianthus caryophyllus L) en la región de
Villa Guerrero México. Rev. Chapingo.14 (6): 145-148.
Dennehy. T. J.;Granett, J. 1984. Spider mite resistance to dicofol in San Joaquin Valley cotton: Inter and
intraspecific variability in susceptibility of three species of Tetranychus. J. Econ. Entomol. 77(6):
1381-1385.
FAO. 1979. Recommended methods for the detection and measurement of resistance of agricultural pests
to pesticides. FAO. Plant Protection Bulletin, 27: 29 - 32.
Finney, D. J. 1971. Probit Analysis. Cambridge at the Univ. Press. 3rd
Ed. 50-80 pp.
Georghiou, P. G.; Saito, T. 1983. Resistance to pesticides. Plenun Press. New York, USA. 809 pp.
Georghiou, G.P.; Lagunes A. 1991. The ocurrence of resistance to pesticides in arthropods. FAO. Rome,
Italy. 318 pp.
Goodwin, S., G. Herron, N. Gough, T. Wellham, J. Rophail, and R. Parker. 1995. Relationship Between
Insecticide – Acaricide Resistance and Field Control in Tetranychus urticae (Tetranychidae)
Infesting Roses. Journal of Economic Entomology. 88 (5):1106 – 1112.
Gould, M. J. 1987. Integrated Pest. Management. In Protected Crops. Burn A.J., T.H Croaker and P.C.
Jepson. Academic Pres. Pp 404-405.
Granham, J. E. and W. Helle. 1985. Pesticide resistance in Tetranychidae. pp.405-420. In: W. Helle, and
M. W. Sabelis (Eds.). Spider mites. Their biology, natural enemies and control. ElSevier.
Amsterdan.
James, D. G. 2000. Abamectin resistance in spider mites on hops. Agrichemical & Environmental News.
170:4-6.
Jepson, L. R., H.H. Keifer, y E.W. Baker. 1975. Mites Injurious to Economic Plants. University of
California Press. p 614.
Lagunes T. A. y Villanueva J. J. 1994. Toxicología y manejo de insecticidas. Colegio de Posgraduados en
Ciencias Agrícolas. Montecillo, Estado de México. 264 pp.
Romanh C. V., Ramírez H. M. y Treviño J. G. Dendrometría. Universidad Autónoma Chapingo, México.
Pp 161-164.
Sato, M. E.; Passerotti, C. M.; Takematsu, A. P.; De Souza M .F.; Potenza, M. R. 2000. Resistência de
Tetranychus urticae (koch, 1836) a acaricidas, em Pessegueiro (Prunus persica (l.) batsch) em
Paranapanema e Jundiaí, sp. Arquivos Instituto Biológico. 67 (1): 20-24.
SAS Institute Inc. 2000. Ghide for personal computers. SAS institute, Cary, N.C.

26
Tian, T., E. .E. Gtafton-Cardwell and J. Granett. 1992. Resistance of Tetranychus urticae Koch (Acari:
Tetranychidae) to cyhexatin and fenbutatin-oxide in California pears. J. Econ. Entomol., V. 85:
2088-2095.
PSEUDOESCORPIONES DE CUEVAS DE COAHUILA, MÉXICO
Pseudoscorpions of caves from Coahuila, Mexico
Gabriel A. Villegas-Guzmán. Colección Nacional de Ácaros, Depto. Zoología, Instituto de
Bología, UNAM. 3er circuito exterior, Apto. P. 70-153, C. P. 04510. Ciudad Universitaria,
México, D. F. [email protected]
Palabras Clave: Pseudoescorpiones, cuevas, nuevos registros, Coahuila
Introducción
Por la presencia de la Sierra Madre Oriental y Occidental, en el norte del país se
desarrollan diversos sistemas de cuevas. En algunos casos estos están ampliamente estudiados
como son los sistemas cavernarios de los estados de Tamaulipas, San Luis Potosí y Nuevo León,
en los cuales se han colectado diez, seis y cuatro especies de pseudoescorpiones cavernícolas,
respectivamente (Ceballos, 2004). Sin embargo, en otros estados los estudios son escasos, como
en el estado de Estado de Coahuila, donde se han realizado pocos estudios faunísticos de los
artrópodos encontrados en este tipo de hábitat (Reddell, 1981).
Los organismos registrados en nueve cuevas del estado de Coahuila son: las arañas de los
géneros Physocyclus y Filistatoides (Reddell, 1981); los alacranes Pseudouroctonus sprousei
(Francke y Savary, 2006) y Pseudouroctonus savvasi (Francke, 2009), los coleópteros de las
familias Dermestidae y Tenebrionidae, los grillos troglófilos, algunos colémbolos y psocópteros
(Reddell, 1981) (Cuadro 1).
Cuadro 1. Artrópodos reportados de las cuevas de Coahuila, México.
Cueva Arácnidos Insectos
Cueva de la Herradura Physocyclus hoogstraali Dermestes carnivorus
Psilochorus sp. Alphotobius laevigatus
Filistatoides sp. Cryptoglossa mexicana
Bocas del Carmen Ceuthophilus sp.
Cueva de las Animas Physocyclus hoogstraali Paracophus subapterus
Filistatoides sp.
Loxosceles belli
Gruta de Cuevecillas Tegenaria gertschi Pseudosinella reddelli
Filistatinella crassipalpis Psyllipsocus ramburii
Metagonia coahuila Rhadine rotgeri
Sierra de Mayrán Filistatoides sp. Centrioptera sp.
Physocyclus sp. Cryptoglossa sp.
Eleodes sp.
Cueva de León Physocyclus enaulus

27
Pozo del Potrero Metagonia sp.
Cueva Casa Blanca Pseudouroctonus savvasi
Cueva El Abra Pseudouroctonus sprousei
No se conocen registros de pseudoescorpiones colectados en cuevas de Coahuila, sin
embargo, existe un reporte de la especie Albiorix conodentatus Hoff, 1945, colectado 5 millas al
oeste de Saltillo (Hoff, 1945), de hecho es el único registro conocido de esta especie. El objetivo
de este trabajo es el de dar a conocer los primeros registros de pseudoescorpiones de cuevas del
estado de Coahuila.
Materiales y Método
El material biológico fue recolectado en seis cuevas: Gruta Consuelo, Candela, NAD27
14 331290 2964113 (26º 47,3987’ N, 100º 41.827’ O) , 566 msnm, 26 enero, 2006 y 5 enero
2007, cols Peter Sprouse y Charley Savvas. Cueva el Tulillo, San Buena Ventura, NAD27 14
242097 3032607 (27º 23.639’ N, 101º 36.481 O), 789 msnm, 24 enero 2006; cols. Peter Sprouse,
B. Shade, J. Krejca, C. Savvas, J. Kennedy, C. Kennedy. Gruta de la Llanta, Torreón, UTM
WGS84 704263 2769481 (25º 01605’ N, 96º 58.520’ O), msnm 1593, 7 julio 2007, cols.
Kathleen O’Connor y Mark Sanders. Cueva Popo de Oso, Zaragoza, NAD27 14 269602 3169357
(28º 37.944’ N, 101º 21.407 O), 699 mnsm, cols. Patrick Rhoades y Peter Sprause. Pozo
Cokendolpher, Ramos Arizpe, NAD27 14 286662 2898284 (26º 11.382’ N, 101º 08.089’ O),
msnm 1177, cols. B. Shade, J. Krejca y C. Savvas. Cueva del detector perdido, Monclova,
NAD27 14 295710 2985074 (26° 58.451’ N, 101° 03.497’ O), 2 agosto 2008, col. Peter Sprouse.
Los organismos fueron colocados en alcohol al 80% y transportados al laboratorio donde
se procesaron por la técnica de Hoff (1949) y las modificaciones de Wirth y Marston (1968). Los
pseudoescorpiones están depositados en la Colección Nacional de Arácnidos del Instituto de
Biología de la Universidad Nacional Autónoma de México.
Resultados
Se recolectaron 61 pseudoescorpiones (47 ♂♂, 14♀♀) de seis cuevas del estado de
Coahuila, que pertenecen a tres familias, cuatro géneros y cuatro especies (Cuadro 2). Las
especies encontrados fueron Neoallochernes stercoreus (Turk), Illinichernes stephensis Benedict
y Malcolm, 1982, Albiorix conodentatus Hoff, 1945 y Cheiridium sp.
Los ejemplares de N. stercoreus se hallaron en tres cuevas (Cuadro 2) y con un mayor
número (43 ♂♂, 13♀♀); mientras que de las otras tres especies, I. stephensis, A. conodentatus y
Cheiridium sp. se localizaron en una cueva cada una. Se obtuvieron sólo organismos machos, las
dos primeras especies con dos ejemplares cada una con dos ejemplares y la tercera con un sólo
ejemplar.
Cuadro 2. Pseudoescorpiones encontrados en cuevas de Coahuila, México
Familia Especie Cueva
Chernetidae Neoallochernes stercoreus 10♂♂, 3♀♀ Gruta del Consuelo
N. stercoreus 32 ♂♂, 9♀♀ Cueva del Tulillo
N. stercoreus 1♂, 1♀ Pozo Cokendolpher
Illichernes stephensis 2♂♂ Cueva Popo de Oso
Ideoroncidae Albiorix conodentatus 2♂♂ Gruta de la Llanta

28
Cheiridiidae Cheiridium sp. 1♂ Cueva del detector
perdido
Discusión y Conclusiones
Dos de las cuatro especies encontradas ya han sido registradas en cuevas, como es el caso
de N. stercoreus e I. stephensis, en Sinaloa y Jalisco, respectivamente (Villegas-Guzmán, 2001;
Villegas-Guzmán et al., 2000), ambas especies están asociadas al guano de los murciélagos
Tadarida brasiliensis mexicana (Saussure) y Mormoops megalophylla Peters, 1864.
En el caso de A. conodentatus este el primer registro de esta especie en cuevas,
desafortunadamente se desconocen los datos del sitio de colecta del registro previo, sólo se sabe
que fue recolectado cerca de Saltillo (Hoff, 1945).
La presencia de Cheiridium en cuevas, representa el primer registro del género en este
tipo de hábitat en el país, previamente sólo se conoce a la especie Cheiridium insperatum Hoff y
Clawon, 1952, en nidos de Neotoma (Villegas-Guzmán y Pérez, 2005). Sin embargo, existen
registros de la especie Cheiridium reyesi Muchmore, 1992, en cuevas de Texas (Muchmore,
1992), por lo que su existencia en la cueva no es extraña. El ejemplar encontrado tiene cierta
similitud con Cheiridium firmum Hoff, 1952, la cual fue recolectada en una oquedad de un árbol
(Hoff, 1952), pero se requiere de una revisión más detallada para poder determinar la especie del
ejemplar recolectado.
Las cuatro especies de pseudoescorpiones encontradas son trogofilas, ya que no presentan
adaptaciones morfológicas propias de los organismos troglobios, como son los apéndices
alargados y en general mayor tamaño del cuerpo, si no que presentan características de
organismos epigeos. Sin embargo, N. stercoreus únicamente se ha encontrado en hábitats
cavernícolas en los refugios de murciélagos (Muchmore, 1990; Villegas-Guzmán, 2001). La
presencia de los pseudoescorpiones en este tipo de hábitat se debe a que aquí encuentran las
condiciones adecuadas (temperatura, humedad y alimentación) para su desarrollo.
Con los pseudoescorpiones aquí registrados se dan a conocer las primeras especies
asociadas a cuevas de Coahuila, además se aumenta a cuatro especies la diversidad de
pseudoescorpiones en el estado. Se considera necesario incrementar estudios sobre estos
arácnidos en el norte de México, debido al escaso número de registros que actualmente se
conocen.
Agradecimientos
A Peter Sprouse, Charley Savvas, Kathleen O’Connor, Mark Sanders, Patrick Rhoades, B.
Shade, J. Krejca, J. Kennedy, C. Kennedy por la recolecta de material biológico y el envió del
mismo. Al Dr. Oscar F. Francke y a la M. en C. Griselda Montiel por sus comentarios y
sugerencias al manuscrito.
Literatura Citada
Ceballos, A. 2004. Pseudoscorpionida. En: Llorente J., Morrone J. J., Yánez O. y Vargas I. (eds)
Biodiversidad, taxonomía y biogeografía de artrópodos de México, hacía una síntesis de
su conocimiento. Vol. 4. CONABIO-UNAM, México, pp. 417-429.
Hoff, C. C. 1945. The pseudoscorpion genus Albiorix Chamberlin. American Museum Novitates,
1277:1-12.

29
Hoff, C. C. 1949. The pseudoscorpions of Illinois. Bulletin of the Illinois Natural History Survey,
24:409-498.
Hoff, C. C. 1952. Two new species of pseudoscorpions from Illinois. Academy of Science
Transactions, 45:189-195.
Francke, O. F. 2009. Description of a new species of troglophile Pseudouroctonus (Scorpiones:
Vaejovidae) from Coahuila, Mexico [Descripción de una nueva especie Pseudouroctonus
de troglófilo (Scorpions: Vaejovidae) de Coahuila, México]. Texas Memorial Museum
Speleological Monographs, 7. Studies on the cave and endogean fauna of North America,
V. Pp.11-18.
Francke, O. F. y W. E. Savary. 2006. A new troglobitic Pseudouroctonus Stahnke (Scorpiones:
Vaejovidae) from northern México. Zootaxa, 1302:21-30.
Muchmore, W.M. 1992. Cavernicolous pseudoscorpions form Texas and New Mexico
(Arachnida: Pseudoscorpionida). Texas Mmemorial Museum, Speleological Monographs,
3:127-153.
Muchmore, W.M. 1990. Pseuoescorpionida. En Dindal, D. L. (ed), Soil biology guide. John
Wiley and Sons. New York. pp. 503-527.
Reddell, J.R. 1981. A rewiev of the cavernicole fauna of Mexico, Guatemala and Belize. Texas
Memorial Museum, the University of Texas at Austin, 327pp.
Villegas-Guzmán, G. A. 2001. Pseudoescorpiones (Arachnida: Pseudoscorpionida) de las cuevas
“Las Garrochas”, Jalisco, México. En: Memorias del XXXVI Congreso Nacional de
Entomología, Sociedad Mexicana de Entomología, Querétaro, Querétaro. Pp. E-33.
Villegas-Guzmán, G. A. y T. M. Pérez. 2005. Pseudoescorpiones (Arachnida:
Pseudoscorpionida) asociados a nidos de ratas del género Neotoma (Mammalia: Rodentia)
del Altiplano mexicano. Acta Zoológica Mexicana (n. s.), 21:63-77.
Villegas-Guzmán, G. A., G. Montiel-Parra y O. J. Polaco 2000. Artropodofauna de la cueva “La
Chinacatera”, Sinaloa, México. En: Memorias del XXXV Congreso Nacional de
Entomología, Sociedad Mexicana de Entomología, Acapulco, Guerrero. Pp. 559-563.
Wirth, W.W. y N. Marston.1968. A method for mounting small insects on microscope slides in
Canada balsam. Annals of the Entomological Society of America, 61:783-784.

30
EFECTO EN CONCENTRACIONES FOLIARES DE N, P Y K EN DURAZNERO
POR LA ALIMENTACIÓN DE Eotetranychus lewisi (ACARI: TETRANYCHIDAE)
Effects on foliar contents of NPK on peach leaves by mite feeding Eotetranychus lewisi
(Acari.Tetranychidae)
Gerardo Pérez-Santiago1, Gabriel Otero-Colina
2 y Víctor A. González-Hernández
2. 1
CIIDIR-
COFAA. IPN. Sigma s/n. Fracc. 20 de Nov. II, Durango, 34220 México. 2Colegio de
Postgraduados, Campus Montecillo. 56230, Montecillo, Edo. de México, México.
Palabras Clave: NPK, ácaros-día acumulados, peso específico.
Introducción
Estudios efectuados en los 70s por Herbert y Butler (1973) mostraron que en árboles de
manzano (Malus pumila Mill.) infestados con el ácaro Panonychus ulmi (Koch), las
concentraciones de minerales como el N disminuyen en forma creciente al elevarse las
poblaciones de ácaros. Por su parte, Marscher (2002) considera que el daño en plantas por
artrópodos chupadores está relacionado con la cantidad de nitrógeno soluble en hojas, más que
con la cantidad de proteínas. Además de evaluar las concentraciones nutrimentales de las hojas,
otra forma de medir el daño causado por ácaros es mediante la acumulación de materia seca por
unidad de superficie foliar; índice denominado peso específico de la hoja (PEH), que está
estrechamente asociado con la fotosíntesis y el metabolismo foliar. Kappel y Flore (1983) señalan
que el PEH del duraznero `Redhaven' disminuyó al reducirse la intensidad de la luz.
En estudios realizados para comparar el efecto de los ácaros T. urticae y P. ulmi, Mobley
y Marini (1990) encontraron que en duraznero el PEH no resultó afectado por la infestación
mientras que en manzano se incrementó, lo cual mostró que el duraznero es más tolerante al daño
por ácaros. En México se carece de trabajos que cuantifiquen el efecto de las infestaciones de las
especies de ácaros presentes en los cultivos. Por ello, el objetivo de este estudio fue determinar el
efecto de diferentes densidades del ácaro E. lewisi en los contenidos foliares de N, P y K y en el
peso específico de la hoja, en plantas de duraznero de dos años de edad creciendo en condiciones
de invernadero.
Materiales y Método
Durante 2003 se mantuvo en forma permanente una colonia de ácaros E. lewisi
procedentes de hojas de duraznero colectadas en El Álamo, Sombrerete, Zacatecas, México. Los
ácaros se llevaron al laboratorio de Acarología del CP, se establecieron en plantas de frijol var.
Flor de Mayo. Estas plantas se mantuvieron en una jaula de aluminio de 0.6 x 0.6 x 1.20 m,
cubierta con malla antiáfidos, a temperatura de 25 4º C, humedad relativa de 40-50% y

31
fotoperiodo de 12:12 horas de luz: oscuridad. Se realizaron siembras escalonadas de frijol para
remplazar al hospedante y revitalizar la colonia cuando fue necesario.
Las plantas infestadas con ácaros fueron árboles de duraznero cv. ‘Diamante Mejorado’
de dos años de edad, con una altura promedio de 1.6 m, establecidos en bolsas de plástico negro
con capacidad de 20 L y llenas con un sustrato compuesto de tierra de monte y tezontle en una
proporción de 1:2, respectivamente, en un invernadero de policarbonato ubicado en
Tequexquináhuac, Estado de México. Los árboles se regaron dos veces por semana, con un
volumen de 5 L por planta y aplicó fertilizante a una concentración de 900 mg L-1
(Hakaphos®
rojo, BASF) (NPK; 18-18-18) cada tres semanas.
Tratamientos de infestación
Los tratamientos consistieron de cuatro niveles de infestación de ácaros: Tratamiento 1 (0
ácaros por hoja –aph-, testigo), Tratamiento 2 (10-20 aph), Tratamiento 3 (21-40 aph) y
Tratamiento 4 (41-80 aph). Para aplicarlos, sobre las cuatro hojas maduras más jóvenes
localizadas en la parte superior del tallo principal de los árboles de duraznero se colocaron
foliolos de frijol infestados de la colonia previamente descrita y que contenían las cantidades de
ácaros necesarias, lo cual se verificó mediante revisión al microscopio estereoscópico. Se infestó
así un total de 24 árboles por tratamiento en la última semana del mes de agosto, en el otoño
2003.
Se monitorearon semanalmente las poblaciones de E. lewisi en cada tratamiento; el
muestreo inició en septiembre, 15 días después de la infestación (ddi), y terminó la segunda
semana del mes de noviembre, a los 81 DDI. Para el muestreo de ácaros se tomaron dos hojas
próximas a los sitios de infestación en cada árbol, de ocho árboles por tratamiento, se guardaron
en bolsas de papel y se trasladaron al laboratorio en una hielera portátil, donde al microscopio se
contaron los estados activos de los ácaros. Con los datos obtenidos se calculó la cantidad de
ácaros-día acumulados en cada temporada (ADA/Ft), con el método propuesto por Hoy (1985):
ADA/Ft = (0.5d) {xa + xp} + ADA/Fa
Donde, d = días desde el anterior muestreo; xa = promedio actual de ácaros por hoja, xp =
promedio anterior de ácaros por hoja del muestreo anterior; y ADA/Fa= ácaros-día acumulados
en el muestreo anterior.
Con esta infestación inducida se partió del supuesto de que el efecto de los ácaros sobre
las plantas es resultado acumulativo de la alimentación por ellos, más que el efecto puntual de las
poblaciones en un momento específico. Se hicieron dos muestreos de hojas: a los 31 ddi y a los
81 ddi. El material se lavó con agua jabonosa para eliminar a los ácaros, se enjuagó con agua
destilada, se secó a temperatura ambiente y posteriormente se secó en estufa a 70 ºC por 3 días.
Los análisis químicos de N, P y K se hicieron en 1 g de materia seca del tejido foliar seco y
molido. La determinación de N se hizo por el método de Micro-Kjeldahl. El contenido de P se
determinó por colorimetría. El K se determinó por flamometría y se cuantificó mediante
espectrometría de absorción atómica (Alcántar y Sandoval, 1999). Estas tres determinaciones se
hicieron por triplicado.
Adicionalmente se determinó el peso específico de la hoja (PEH) de los tratamientos, a
los 81 ddi; como muestras se seleccionaron ocho hojas maduras de la parte media del brote,
próximas a las que se infestaron y a las que se tomaron para la determinación de los nutrimentos.
Dichas hojas se lavaron para eliminar los ácaros, se secaron a temperatura ambiente, se midió su
área (AF) con un integrador de área foliar (LI 3100), se secaron en estufa a 70 oC por 72 h y se
determinó el peso seco (PS). El peso específico de las hojas (PEH) se calculó al dividir PS entre

32
AF. Los datos de contenidos de N, P, K y PEH se sometieron a análisis de varianza conforme a
un diseño de bloques completos al azar con dos muestras compuestas por hojas de cuatro árboles
de cada bloque, para cada fecha de muestreo; para la comparación de medias se usó la prueba de
Tukey con un nivel de significación de 0.05. Los análisis estadísticos se hicieron con el paquete
SAS versión 8.1 (SAS Institute, 2000).
Resultados y Discusión
Ácaros-día acumulados (ADA). Durante el periodo de estudio las infestaciones de
ácaros se mantuvieron artificialmente dentro de límites preestablecidos. El hecho de que las
infestaciones se mantuvieron relativamente constantes en cada uno de los dos ciclos de estudio,
como se mostró en otra publicación previa (Pérez et al., 2007). Como era de esperarse, la
cantidad de ADA total varió entre tratamientos, desde 28 a 772 a los 31 ddi y de 152 a 4771 a los
81 ddi (Cuadro 1); esta diferencia entre estaciones se atribuye a las diferencias climáticas
inherentes que afectaron el desarrollo biológico de las plantas y de los ácaros. Es necesario
señalar que si bien en los tratamientos testigo los árboles debían mantenerse sin ácaros, por la
barrera física instalada y por el constante lavado de hojas con agua jabonosa a presión, la
proximidad con los árboles infestados facilitó la presencia de ácaros pero en bajas poblaciones.
Cuadro 1. Ácaros-día acumulados (ADA) por tratamiento y concentración de N, P y K, en duraznero Prunus persica
cv. ‘Diamante mejorado’, en 2003 a 31 y 81 días después de la infestación (DDI).
Tratamiento
Ácaros-día
acumulados
Concentración (mg g-1
)
N P K
31 días después de la infestación
1 (testigo)
2
3
4
28
195
516
772
23.0az
21.9a
22.4a
22.5a
2.0a
2.3a
2.0a
2.1a
25.1a
26.3a
24.9a
26.2a
81 días después de la infestación
1 (testigo)
2
3
4
152
1313
2844
4771
26.7a
24.2b
22.8c
20.3d
2.5a
2.4a
2.2a
2.2a
27.4a
22.4b
22.0bc
21.7c zValores con la misma letra dentro de cada columna son iguales de acuerdo a la prueba de Tukey con P 0
Concentraciones de N, P y K. A los 31 ddi con sus correspondientes 772 ADA, la
cantidad de daño causado al follaje por la alimentación de E. lewisi no fue suficiente para
ocasionar diferencia estadística (P<0.05) en los contenidos de N, P y K, entre los árboles
sometidos a infestación de ácaros y el testigo (Cuadro 1). En contraste, Ferree y Hall (1980)
encontraron que con la alimentación de 758 individuos inmaduros y adultos de T. urticae se
redujo en 6.5% el contenido de N en hojas de manzano. Tal vez el contraste en resultados se deba
a que el en duraznero es más tolerante al daño por E. lewisi que el manzano al daño por T.
urticae. La concentración de NPK a los 81 DDI se muestran también en el Cuadro 1, en el se
indican los ADA a ese tiempo transcurrido. A consecuencia de una mayor acumulación de ácaros
día se presentaron diferencias estadísticas (P<0.05) en el contenido de N y K en los árboles
infestados con respecto al testigo. La concentración de N se redujo con respecto al testigo 9.3,

33
14.6 y 23.9 % en los tratamientos A, B y C, respectivamente, mientras que la concentración de K
se redujo en 18.2, 19.7 y 20.8 % en los tratamientos A, B y C, respectivamente.
Peso específico de la hoja (PEH). En esta característica no se encontró diferencia
estadística entre los tratamientos infestados con ácaros y el testigo. Una posible explicación a este
comportamiento es que la aplicación de los tratamientos de ácaros se efectuó cuando las hojas ya
habían alcanzado su expansión total, de tal modo que el periodo de crecimiento y el peso de la
hoja no fueron afectados por la alimentación de los ácaros. Este resultado muestra que las
infestaciones aplicadas no fueron suficientemente severas para afectar la acumulación de reservas
en la hoja, uno de los componentes del PEH. Mobley y Marini (1990) encontraron que en
manzano cv. `Imperial Delicious´ infestado con P. ulmi con valores de 0, 738 y 1741 ADA no se
afectó el PEH, mientras que con infestaciones de T. urticae que alcanzaron valores de 0, 776 y
1755 ADA, se incrementó el PEH; en contraste en duraznero cv. `Redhaven´ con infestaciones de
P. ulmi con valores de 0, 3116 y 4309 ADA y valores de 0, 3058 y 4206 ADA de T. urticae, no
se afectó el PEH en ningún caso, lo que indica que las hojas de duraznero fueron más tolerantes a
la alimentación de ambas especies de ácaros. Los resultados encontrados en esta investigación
coinciden con los observados por estos autores.
Cuadro 2. Peso específico de la hoja (PEH) y ácaros-día acumulados por tratamiento en duraznero Prunus persica
cv diamante mejorado, a 81 días después de la infestación
Tratamiento
Ácaros-día
acumulados
PEH
(mg cm-2
)
1Testigo
2
3
4
57
1043
2426
3996
7.49a
7.47a
7.10a
7.42a
Conclusiones
Se encontró una reducción en el contenido de N y K causada por la infestación de ácaros
Eotetranychus lewisi (McGregor) en diversos niveles a los 81 ddi, la reducción fue proporcional a
la cantidad de ADA. No obstante, la alimentación de los ácaros no afectó la concentración de P
en las hojas. La reducción más notable fue de N, lo que llevó a este nutrimento por debajo de los
niveles normales u óptimos para duraznero. No se observó efecto en el PEH en duraznero cv.
`diamante mejorado´ por la alimentación E. lewisi en los diferentes tratamientos.
Literatura Citada
Alcántar, G. M. Sandoval V. 1999. Manual de análisis químico de tejido vegetal. Publicación
Especial 10. Sociedad Mexicana de la Ciencia del Suelo, A. C. Chapingo, México.
Ferree, D. C, F. R. Hall. 1980. Effects of soil water stress and twospotted spider mites on net
photosynthesis and transpiration of apple leaves. Photosynthesis Research: 189-197.
Herbert, H. J. y K. P. Butler. 1973. The effect of European red mite Panonychus ulmi (Acarina:
Tetranychidae) infestations on N, P and K concentrations in apple foliage throughout the
season. Canadian Entomologist 105: 263-269

34
Hoy, M. A. 1985. Almonds (California). In: Spider Mites their Biology, Natural Enemies and
Control. W. Helle and M. W. Sabelis (eds). Elsevier. The Netherlands, Vol. 1B. pp: 299-
310.
Kappel, F. y J. A. Flore. 1983. Effect of shade on photosynthesis, specific leaf weigh, leaf
chlorophyll content, and morphology of young peach trees. J. Amer. Soc. Hort. Sci.
108(4): 541-544.
Marschner, H. 2002. Mineral nutrition of higher plants. Academic Press. 2ed
Great Britain,
London, UK.pp:229-312.
Mobley, K. N. y R. P. Marini 1990. Gas exchange characteristics of apple and peach leaves
infested by European red mite and twospotted spider mite. J. Amer. Soc. Hort Sci. 115(5):
757-761.
Pérez-Santiago, G., G. Otero-Colina, V. A. González H., A. López J. y M. E. Ramírez G. 2007.
Efecto en fotosíntesis y clorofila en duraznero por diferentes densidades de Eotetranychus
lewisi (Acari:Tetranychidae). Entomología mexicana (E. Estrada V., A. Equihua M., C.
Luna L. y J. L. Rosas A. Eds) 6: 35-39.
SAS Institute. 2000. Statistical Analysis System SAS User’s guide. Statistics version 8.1. SAS
Inc. Cary, North Carolina, USA.

35
NUEVOS REGISTROS DE ÁCAROS DE LAS FAMILIAS TROMBIDIIDAE Y
ERYTHRAEIDAE PARÁSITOS DE ARÁCNIDOS EN MÉXICO
New records of mites of the families Trombidiidae and Eryhtraeidae, parasites of arachnids in
Mexico
Griselda Montiel-Parra y Tila Ma. Pérez-Ortiz. Colección Nacional de Ácaros (CNAC),
Departamento de Zoología, Instituto de Biología, Universidad Nacional Autónoma de México.
3er Circuito exterior s/n, anexo al Jardín Botánico exterior, Ciudad Universitaria, México, D. F.,
04510. [email protected]
Palabras Clave: Ácaros, parásitos, Araneae, Amblypygi, México
Introducción
La colonización repetida y la diversificación de los linajes de ácaros simbióticos ha dado
como resultado la asociación con grupos muy diversos, tales como las avispas, hormigas,
escarabajos, termitas, ortópteros, miriápodos, arácnidos, reptiles, aves, mamíferos, coníferas y
angiospermas, etc. Muchos de estos ácaros son parásitos, otros comensales o mutualistas.
Independientemente de la naturaleza de la asociación, la tendencia hacia la especificidad, parece
ser común en aquellos ácaros que pasan toda su vida sobre el huésped (Walter y Proctor, 1999).
Los artrópodos, son los que proveen la mayor parte de los huéspedes para los ácaros
(Walter y Proctor, 1999). En el caso de los ácaros asociados a otros arácnidos, se han registrado
especies parásitas y foréticas de 10 familias, 21 géneros y 88 especies; asociados a 8 de los 11
ordenes de la Clase Arachnida (André, 1949; Baker y Selden, 1997; Cokendolpher, 1993;
Cokendolpher y Mitov, 2007; De Armas y Prieto, 2003; Ebermann y Palacios-Vargas, 1988;
Haitlinger, 2000; Hoffman y López-Campos, 2000; Montiel-Parra y González, 2001; Montiel-
Parra, et al., 2006; Southcott, 1992; Welbourn y Young, 1988; Weygoldt, 2000; Womersley,
1934).
El mayor número de especies de ácaros se ha registrado en Araneae con 36 spp., seguido de
Opiliones con 32 spp., Scorpiones con 24 spp., Acari con 6 spp., Pseudoscorpiones, Amblypygi,
Ricinulei y Solifugae con una especie para cada uno; hasta el momento se desconoce si existen
asociaciones con los otros órdenes, Uropygi, Schizomida y Palpigradi. En México, es escasa la
información sobre los ácaros parásitos de arácnidos, a la fecha sólo se han documentado 7
familias, 8 géneros y 11 especies; de estas últimas siete corresponden a formas foréticas
(hipopodios), tres parásitos protélianos, es decir, que sólo durante la etapa larval están sobre el
huésped, ya que las ninfas y adultos son de vida libre (Hoffmann, 1990), y una especie parásita
permanente. Los huéspedes registrados corresponden a 18 especies de arácnidos de los ordenes:

36
Scorpiones (con 9 spp.), Opiliones (con 13 spp.), Acari (con 2 spp.), Amblypygi (con una sp.) y
Ricinulei (con una sp.) (Cokendolpher, 1993; Ebermann y Palacios-Vargas, 1988; Hoffmann y
López-Campos, 1995; 2000; Martínez y Morales, 1980; Montiel-Parra y González, 2001;
Montiel-Parra et al., 2006; Paredes-León et al., 2005; Peláez, 1962; Welbourn, 1983) (Tabla 1).
Adicionalmente se tiene conocimiento de hipopodios de la familia Acaridae sobre los
pseudoscorpiones Lustrochernes sp., de Chiapas (Villegas-Guzman, com. pers.), así como larvas
de la familia Erythraeidae sobre otros ácaros de Morelos (Montiel-Parra, obs. pers.). Por lo
anterior, el objetivo del presente trabajo es dar a conocer los primeros registros de ácaros
parásitos de arañas en el país, y adicionar nuevos datos de las especies asociadas los
amblipígidos.
Cuadro 1. Ácaros asociados a los arácnidos de México
Ácaro Huésped Estado
MESOSTIGMATA
Uropodidae
Oplithis sp. Hypoaspis sp. ♦ Chis 8
PROSTIGMATA
Cheyletidae
Hoffmannita mexicana Centruroides flavopictus * Oax 9
Erythraeidae
Leptus indianensis Heterovonones incrassatus Chis7
Leptus ca. indianensis Acromares vittatum Chis7
Geay sp. Chis 7
Prionostemma sp. Chis 7
Leptus lomani
Kukulcania sp. ► Gro
Leptus onnae Phrynus garridoi ■
Gro
Paraphrynus pococki ■
Ver 4,11
Leptus ruginus Centruroides limpidus limpidus * Gro 6
Hadrurus gerstchi * Gro 6
Holovonones compressus Chis 7
Metopilio sp. Oax 7
Paramitraceras granulatus Oax 7
Paranelima sp. Oax, Pue7
Prionostemma sp . Oax 7
Vaejovis variegatus * Gro 6
Vonones pelaezi Oax7
Leptus sp. Centruroides limpidus * Gro, Edo. Méx, Mor 4
Cynorta sp. Ver 4
Krusa sp. Mex 1
Leiobunum nycticorpum Hgo, SLP 4

37
Leiobunum sp. DF, Edo. Méx, Mich, Gro, Mor,
Hgo, SLP, Tamps 1,4
Opilion Mich, Mor, Nay, Oax, Ver 4
Leptus sp. Scorpionida * Mex 10
Vejovis (sic) mexicanus * DF 4
Vejovis (sic) spinigerus * Gro, Edo. Méx, Mor 4
Sin determinar Opiliones Mex 5
Ácaro Huésped Estado
Pterygosomatidae
Pimeliaphilus rapax Vejovis (sic) nitudulus nitudulus* Edo. Méx 4
Vejovis (sic) intrepidus cristimanus* Gto 4
Vejovis (sic) punctatus punctatus* Oax 4
Trombidiidae
Allothrombium sp. Modisimus sp. ► Chis 11
Miagrammopes sp. ► Chis 11
Scutacaridae
Imparipes (Imparipes) tocatlphilus Pseudocellus boneti ▲ Gro 2
ASTIGMATA
Acaridae
Sancassania berlesei Centruroides limpidus limpidus * Gro 6
Vaejovis sp. nov * Gro 6
Sancassania ca. ojibwa Acromares vittatum Chis 7
Schwiebea ca. nova Paranelima sp. Oax 7
Sin determinar Opilion Mor 4
Histiostomatidae
Histiostoma sp. Chapalania cifuentesi ♦ Jal 3
* alacrán; ● opilion, ■ amblipígido, ▲ ricinúlido, ♦ ácaro, ► araña. 1) Cokendolpher, 1993; 2) Ebermann y Palacios-Vargas, 1988; 3) Hoffmann y López-Campos, 1995; 4) Hoffmann y López-Campos, 2000; 5) Martínez y Morales, 1980; 6) Montiel-Parra y González, 2001; 7) Montiel-Parra
et al., 2006; 8) Paredes-León et al., 1995; 9) Peláez, 1962; 10) Welbourn, 1983; 11) Este trabajo.
Materiales y Método
Las arañas y amblipígidos se capturaron manualmente durante colectas diurnas y
nocturnas realizadas como parte de los proyectos: “Inventario de los arácnidos en la Selva
Lacandona, Chiapas” y “Estudio de los alacranes de la familia Vaejovidae en México”.
En el estado de Chiapas se colectaron dos arañas con ácaros: Modisimus sp. (Pholcidae),
en Camino hacia "La Cojolita", Mpio. Ocosingo, 15° 45.4833' N, 91° 1.000' W, 215 msnm, 29
abril 2004, J. Castelo, col., y un Miagrammopes sp. (Uloboridae) en Anexo Cofolasa, Mpio.
Ocosingo, 16° 38.462' N 90° 46.881' W, 181 msnm, 8 mayo 2006, colecta diurna, G. Montiel,
col.
En el estado de Guerrero se halló una araña de Kukulcania sp. (Filistatidae), cerca de
Chilpancingo, 11 abril 1999, E. González, col., así como, dos amblipígidos de la especie Phrynus
garridoi (Phrynidae) en Cumbres del Llano Largo, Mpio. Puerto Márquez 16° 49.505’N, 99°
49.999’ W, 371 msnm, 19 junio 2007, O. Francke, H. Montaño y A. Ballesteros, cols. Los
ejemplares se preservaron individualmente en viales con alcohol al 80% y fueron examinados

38
bajo el miscroscopio estereoscopico para extraer a los ácaros, registrando en un catálogo los datos
de colecta y su ubicación en el cuerpo de cada huésped.
Frecuentemente los ácaros de las familias Erythraeidae, Trombiculidae y Trombidiidae, se
rompen cuando se utiliza el lactofenol para digerir su contenido (Montiel-Parra, obs. pers.), razón
por la cual, los ácaros extraídos de las arañas y amblipígidos fueron aclarados con la técnica de
pancreatina (Álvarez-Padilla y Hormiga, 2007(2008)). Posteriormente se procesaron en laminillas
microscópicas con Hoyer y para su estudio se utilizó un microscopio Nikon Optiphot-2. El
material biológico se encuentra depositado en la Colección Nacional de Ácaros (CNAC) y
Colección Nacional de Arácnidos (CNAN) del Instituto de Biología, UNAM.
Por otra parte, se examino una laminilla del género Leptus sp. depositada en la CNAC,
correspondiente a una larva colectada en el amblipígido Paraphrynus pococki del estado de
Veracruz (Hoffmann y López-Campos, 2000).
Resultados y Discusión
De las tres especies de arañas con ácaros, se obtuvieron cinco larvas de las familias
Trombidiidae y Erythraeidae (Cuadro 1). Cuatro ejemplares pertenecen a la especie
Allotrombium sp., una larva se localizo en la parte dorsal del opistosoma en Modisimus sp.
(Pholcidae); y los otros tres ejemplares se ubicaron en el tejido blando que se encuentra en la base
de las coxas I, III y IV en Miagrammopes sp. (Uloboridae). Sólo una larva de la especie Leptus
lomani se encontró en los pliegues del epiginio de Kukulcania sp. (Filistatidae).
En los dos ejemplares de Phrynus garridoi se hallaron tres larvas, una y dos ejemplares,
respectivamente, pertenecientes a la especie Leptus onnae (Cuadro 1). Estas larvas se ubicaron en
la quela del pedipalpo, dorso del caparacho y en la parte ventral de la tibia III. La larva de
Paraphryhus pococki también corresponde a L. onnae (Cuadro 1). Welbourn y Young (1988)
mencionan que las larvas de la familia Trombidiidae se ubican frecuentemente en las líneas de
separación (suturas de muda), en estas áreas conocidas como pleuron, los ácaros se sitúan porque
es una región suave y flexible, lo que facilita la penetración de sus quelíceros, así como su
supervivencia durante la muda. Esto coincide con la ubicación que presentaron las tres larvas de
Allothrombium sp. en Miagrammopes sp. Por otra parte, Zhang (1998) señala que en la mayoría
de las especies de trombídidos se desconocen sus abundancias en sus huéspedes. Welbourn y
Young (1988) encontraron que es más frecuente la presencia de uno a tres ácaros de
Eutrombicula lockelii en las arañas Oxyopidae y Linyphiidae, y que pocos son los casos en los
que se obtuvieron de seis a nueve ácaros. En las arañas aquí estudiadas, la presencia de ácaros
también oscilo de 1 a 3 ejemplares por huésped.
Las larvas de Allotrombium son frecuentes en áfidos (Welbourn y Young, 1998), sin
embargo, también han sido registradas en arañas de la familia Lycosidae; Parker (1965) halló la
especie A. fuliginosum en Lycosa amentata en Inglaterra, y Moss (1960) la especie A. lerouxi en
Trochosa terricola en Canadá. En México, la especie Allotrombium sp., únicamente había sido
registrada en muestras de suelo del estado de Veracruz (Hoffmann y López-Campos, 2000). Por
lo cual, las arañas Miagrammopes sp. y Modisimus sp., son nuevos huéspedes para Allotrombium
sp., además su presencia en el estado de Chiapas, representa un nuevo registro para el estado.
La familia Erythraeidae, con su género cosmopolita Leptus (Latreille), contiene la mayor
parte de las especies parásitas de arácnidos (Welbourn y Young, 1988). Los sitios de ataque de
estos eritraeídos, sólo han sido documentados con detalle en opiliones (Welbourn, 1983; Mcaloon
y Durden, 2000). La especie L. lomani hallada en Kukulcania sp., ha sido registrada previamente

39
en el opilion, Sadocus funestus en Chile (Cokendolpher, 1993). Por tal motivo, Kukulcania sp. es
un nuevo huésped para la especie, además el eritraeído se registra por vez primera en el país.
Los ácaros parásitos de amblipígidos son menos frecuentes, se han encontrado larvas sin
determinar en Phrynus marginemaculatus de Florida y a Leptus sp., en P. kennidae de República
Dominicana; de la segunda especie se registraron como sitios de ataque el caparacho, patas y
pedipalpos (De Armas y Prieto, 2003; Weygoldt, 2000), dichos sitios coinciden con la ubicación
que presentaron las larvas de L. onnae colectadas en Phrynus garreroi, pero se desconoce en
Paraphryhus pococki, porque en las etiquetas de la laminilla, no está señalado la posición de la
larva en el huésped. La especie L. onnae ya había sido registrada en Chiapas (Haitlinger, 2000),
por lo tanto, los ejemplares colectados en Guerrero y Veracruz, incrementan la distribución de la
especie en México.
Conclusiones
Con este trabajo se dan a conocer para México, los primeros registros de ácaros asociados a
las arañas; de las tres especies estudiadas, sólo se conocían los ácaros para ejemplares de la
familia Uloboridae, en las cuales se han registrado los eritraeídos, Leptus hidakai en Singapur
(Baker y Selden, 1997) y Charletonia sp., en Estados Unidos (Welbourn y Young, 1998). Por lo
tanto, además este trabajo contribuye con los primeros aportes de ácaros para las familias
Pholcidae y Filistatidae. También se registra la cuarta especie de amblipígido con presencia de
ácaros.
En el país existe una gran diversidad de arañas y amblipígidos, sin embargo, es escasa la
información acerca de sus parásitos, por ello, es necesario continuar con el estudio en éstos
arácnidos e incluso con el resto de los órdenes de la Clase Arachnida.
Agradecimientos
A los Biól. Alejandro Valdez Mondragón y Alfonso Ballesteros por la identificación de las
arañas y amblípigidos, respectivamente. Al M. en C. Ricardo Paredes León por sus comentarios y
sugerencias al manuscrito.
Literatura Citada Álvarez-Padilla, F. y G. Hormiga. 2007 (2008). A protocol for digesting internal soft tisues and mounting
spiders for scanning electron microscopy. The Journal of Arachnology, 35: 538-542.
André, M. 1949. Nouvelle forme larvarire de Thrombidion (Isothrombium) oparbellae (n. g., n. sp.)
parasite d’un solifugae. Bulletin du Museum, 21(3): 354-357.
Baker, A. S. y P. A. Selden. 1997. New morphological and host data for the ectoparasitic larva of Leptus
hidakai Kawashima (Acari, Acariformes, Erythraeidae). Systematic Parasitology, 36: 183-191.
Cokendolpher, J. C. 1993. Pathogens and parasites of opiliones (Arthropoda: Arachnida). The Journal of
Arachnology, 21:120-146.
Cokendolpher, J. C. y P. G. Mitov. 2007. Natural Enemies, pg 339-373. En: Harvestmen. The Biology of
Opiliones (Pinto da Rocha, R., G. Machado y G. Giribet, eds.). Harvard University Press,
Cambridge, 597 pp.
De Armas, L. F. y D. Prieto T. 2003. Primer registro de ácaros parásitos de amblipígidos (Arachnida:
Amblypygi). Revista Ibérica de Aracnología, 7: 133-134.
Ebermann, E. y J. G. Palacios-Vargas. 1988. Imparipes (Imparipes) tocatlphilus n. sp. (Acari,
Tarsonemina, Scutacaridae) from Mexico and Brazil: First record of ricinuleids as phoresy hosts for
scutacarid mites. Acarologia, 29 (4): 347-354.

40
Haitlinger, R. 2000. Four new species of Leptus Latreille, 1796 (Acari: Prostigmata: Erythraeidae) from
Central America. Systematic and Applied Acarology, 5: 13-142.
Hoffmann, A. 1990. Los trombicúlidos de México (Acarida: Trombiculidae). Publicaciones especiales 2,
Instituto de Biología, Universidad Nacional Autónoma de México, 275 pp.
Hoffmann, A. y G. López-Campos. 1995. Género y especie nuevos de Hypoaspidinae (Acarida:
Laelapidae) en un caso de foresia múltiple. Anales del Instituto de Biología, Universidad Nacional
Autónoma de México, Serie Zoología, 66(1): 33-46.
Hoffmann, A. y G. López-Campos. 2000. Biodiversidad de los ácaros en México. CONABIO, UNAM,
México, 230 pp.
Martínez C., A. C. y M. Morales S. 1980. Los opilliones (sic) (Arachnida) y sus ectoparásittos (sic)
eritreídos (Acarida, Erythraeidae). Folia Entomológica Mexicana, 43: 44-45.
Mcaloon, F. M. y L. A. Durden. 2000. Attachment sites and frequency distribution of erythraeid mites,
Leptus indianensis (Acari: Prostigmata), ectoparasitic on harvestmen, Leiobunum formosum
(Opiliones). Experimental and Applied Acarology, 24: 561-567.
Montiel-Parra, G. y E. González. 2001. Acarofauna asociada a los alacranes de Atenango del Río,
Guerrero, México, pp E-141. En: Memorias del XXXVI Congreso Nacional de Entomología y
XXVIII Congreso Nacional de Fitopatología (Stanford, C., S. G., A. Morales, M., J. Padilla R. y M.
Ibarra G., eds.). Querétaro, Qro. F-157.
Montiel-Parra, G., C. Guzmán-Cornejo, J. L. Castelo-Calvillo y T. M. Pérez. 2006. Parasitic mites on
harvestmen (Arachnida: Opiliones) from Mexico, p. 128. En: Abstract book, 12 th International
Congress of Acarology, (J. Bruin, ed.). Amsterdam, 226 pp.
Moss, W. W. 1960. Descriptions and mating behavior of Allothrombium lerouxi, new species (Acarina:
Trombidiidae), a predator of small arthropodos in Quebec apple orchards. Canadian Entomologist,
92: 898-905.
Parker, J. R. 1965. More records of mites as ectoparasites in spiders. British Spider Study Group Bulletin,
25: 6.
Paredes-León, R., G. Montiel-Parra, G. A. Villegas-Guzmán, T. M. Pérez-Ortiz y S. Zaragoza-Caballero.
2005. Resultados preliminares de los ácaros asociados a insectos (Insecta: Melolonthidae,
Scarabaeidae) de la Región Selva Lacandona, Chiapas, Pp 84-89. En: Entomología Mexicana,
(Morales, M. M. A., A. Mendoza E., M. P. Ibarra G. y S. Stanford C., eds.), Vol. IV, México,
Memorias del XL Congreso Nacional de Entomología, Tapachula, Chiapas, 1028 pp.
Peláez, D. 1962. Hoffmannita mexicana gen. et sp. nov., Cheyletidae parásito de un alacrán del género
Centruroides (Acarina, Prostigmata). Anales de la Escuela Nacional de Ciencias Biológicas,
México, 11(1-4): 71-83.
Southcott, R. V. 1992. Revision of the larvae of Leptus Latreille (Acarina: Erythraeidae) of Europa and
North America, with descriptions of post-larval. Zoological Journal of the Linnean Society, 105: 1-
153.
Walter, D. E. y H. C. Proctor. 1999. Mites: Ecology, Evolution and Behavior. CABI Publishing , New
York, 322 pp.
Welbourn, W. C. 1983. Potential use of trombidioid and erythraeoid mites as biological control agents of
insect pestes, 103-140 pg. En: Hoy, M. A., Cunningham, G. L. y L. Knutson (eds.). Biological
control of pests by mites. Agricultural Experiment Station, Division of Agriculture y Natural
Resources, University of California, Special Publication 3304, 185 pp.
Welbourn, W. C. y O. P. Young. 1988. Mites parasitic on spiders, with a description of a new species of
Eutrombidium (Acari, Eutrombidiidae). The Journal of Arachnology, 16: 373-385.
Weygolt, P. 2000. Whip Spiders (Chelicerata: Amblypygi). Their Biology, Morphology and Systematics.
Apollo Books, Stenstrup, Dinamarca, 163 pp.
Womersley, H. 1934. A revision of the trombid and erythraeid mites of Australia with descriptions of new
genera and species. Records of the South Australian Museum, 5: 179-254.

41
Zhang, Z-Q. 1998. Biology and ecology of trombidii mites (Acari: Trombidiodea). Experimental and
Applied Acarology, 22: 139-155.
ÁCAROS EN SUELOS DE UN BOSQUE MIXTO EN SANTA JUANA, MICHOACÁN
(ACARIDA: MESOSTIGMATA, PROSTIGMATA, CRYPTOSTIGMATA).
Mites in soils from a mixed wood in Santa Juana, Michoacán (Acarida: Mesostigmata,
Prostigmata, Cryptostigmata).
Ignacio M. Vázquez y M. Guadalupe López-Campos. Laboratorio de Acarología, “Anita
Hoffmann” Departamento de Biología Comparada, Facultad de Ciencias, UNAM. 04510,
México, D. F. [email protected]; [email protected]
Palabras Clave: Ácaros, suelos, Michoacán, México.
Introducción
La biodiversidad de ácaros que albergan los suelos de diferentes ecosistemas forestales ha
sido analizada en otros sitios, como parte del proyecto “El efecto de la diversidad de los suelos en
la biodiversidad de los ecosistemas forestales de México” (Vázquez & López-Campos, 2008). En
el trabajo citado solo se estudiaron los ácaros Prostigmata extraídos de muestras de suelo de una
Finca Cafetalera en Oaxaca. De ellos se destaca la abundancia y diversidad de Cunaxidae con una
densidad de 83.15 Ind. /m2. Cuatro familias más: Eupodidae, Tydeidae, Stigmaeidae y Bdellidae
se distinguieron como abundantes del total de 22 familias que se registraron para la localidad.
En esta ocasión se presenta la abundancia de las familias de tres órdenes de ácaros
(Mesostigmata, Prostigmata y Cryptostigmata) encontrados en muestras de suelo de un bosque
mixto de pino-encino, en Santa Juana Michoacán. Se determinó que el tipo de suelo en el sitio 1
es Andosol lo mismo que en el sitio 2 aunque en el perfil 5 del sitio 2 el suelo tiene morfología de
Luvisol y Andosol; la hojarasca en todos los perfiles esta compuesta de acículas de pino y hojas
de encino principalmente, el terreno tiene una pendiente de entre 40 y 45 grados (P. Krasilnikov,
com. per.).
El tipo de suelo Andosol es altamente poroso, de color oscuro, desarrollado a partir de
material de origen volcánico tal como cenizas volcánicas, piedra pómez (vidrio volcánico) y otros
materiales porosos y de naturaleza vítrea o cristalina. Se le encuentra desde Islandia hasta
Indonesia pero ocurre típicamente en áreas boscosas altas de las tierras continentales que bordean
al océano Pacífico. Su extensión mundial se estima a menos de 1% del total del área del suelo
sobre La Tierra.
En América, el Andosol se encuentra en Canadá, México (a lo largo del Eje
Neovolcánico), Centroamérica y Sudamérica (Colombia, Ecuador, Perú, Bolivia y Chile) (WBR,
2009). En nuestro país se ha registrado este tipo de suelo en lo que llaman la región V Centro,
que incluye el sur de Zacatecas, Aguascalientes, D. F., norte de Michoacán, Estado de México,

42
Tlaxcala y Puebla, con un 8% de la superficie. Además de las características anteriores, el tipo
Andosol consta de una capa superficial muy suelta con abundante materia orgánica y con una
capa endurecida subyacente como límite conocida como tepetate (Sommer & Cram, 2009).
El propósito de este trabajo es observar la diversidad de tres órdenes de ácaros
(Mesostigmata, Prostigmata y Cryptostigmata) en un bosque mixto de pino-encino, en Santa Juana,
Opopeo, Michoacán, México.
Materiales y Método
Se tomaron 2 muestras, una de suelo y una de hojarasca de cada uno de los cinco perfiles
de suelo excavados en 2 sitios de un bosque mixto de pino-encino. La toma de muestras se hizo
del modo habitual, en un recipiente de 10 x 10 x 5 cm, donde se coloco por separado la hojarasca
y el suelo; se trabajo con un total de 20 muestras.
La localidad donde se excavaron los perfiles se conoce como “La Ladera”, esta ubicada en
el Cerro Santa Juana, municipio de Opopeo, Michoacán (Fig. 1). Se registraron las coordenadas
19° 25’ 317’’ N y 101° 33’ 791’’ W en los sitios de muestreo.
La vegetación circundante en los sitios la componen pinos de las especies Pinus lawsonii,
y P. herrerai (Pinaceae); Ternstroemia pringuei (Theaceae) crece como arbusto y Quercus sp.
(Fagaceae) es el otro elemento dominante del bosque mixto. Según los lugareños el sitio se plantó
hace aproximadamente 25 años, después de que estuvo cultivado (P. Krasilnikov, com. pers.).
Los ácaros fueron separados, contados y algunos transparentados y montados para su
determinación. Para calcular la densidad y el porcentaje de individuos por familia se utilizó el
mismo método que en Vázquez & López-Campos, 2008, considerando una superficie de muestra
de 95 cm2.
Resultados y Discusión
Se estudiaron un total de 4770 ejemplares de los que 2321 se ubicaron en el sitio 1 y 2449
en el sitio 2, considerando los tres órdenes. De Mesostigmata se separaron 172 del sitio 1 y 184
del sitio 2, mientras que de Prostigmata se extrajeron 1715 individuos del sitio 1 y 1971 del sitio
2. Finalmente de Cryptostigmata se contaron 340 ejemplares del sitio 1 y 294 del sitio 2; para
cada orden se presentan también los datos al nivel de familia (Cuadro 1).
Dentro del estudio realizado se puede observar que los ácaros del orden Prostigmata
fueron los mejor representados con un total de 3686 ejemplares, el segundo lugar en abundancia
lo representó el orden Cryptostigmata con 728 ejemplares y el menos representado el orden
Mesostigmata con 356. La mayor densidad se encontró en la Familia Tydeidae del orden
Prostigmata con un total de 3105.26 ind/m2. Como nuevas citas para el estado de Michoacán
encontramos seis familias de Mesostigmata, siete de Prostigmata y nueve de Cryptostigmata, de
este último orden solo había sido citada la familia Scheloribatidae.
En la Cuadro 1 pueden distinguirse las familias Laelapidae y Ascidae (Mesostigmata),
Eupodidae y Tydeidae (Prostigmata) así como Oppiidae y Scheloribatidae (Cryptostigmata)
como las más abundantes y por eso representativas de cada orden. En la misma tabla podemos
ver que las formas juveniles de Mesostigmata y Cryptostigmata son muy numerosas, más de 100
individuos por metro cuadrado. Esto señala que las condiciones del suelo son propicias para la
actividad reproductora de las especies en esta época de secas.
Los Mesostigmata de la familia Ascidae han sido registrados como abundantes en otros
ambientes de bosque, señalándose que los ácaros pueden migrar hacia capas subyacentes de la

43
hojarasca mientras la superficie se mantiene seca y vuelven a la hojarasca cuando inicia la época
de lluvias (Price, 1973 citado por Moreno-Moreno, 2008).
De los oribátidos se han definido seis grupos morfológicos adaptativos para los estudios
de ecología y, de acuerdo con eso, los Oppiidae ocupan pequeños espacios en el suelo. Esto
concuerda con nuestros hallazgos pues es en el suelo de los dos sitios muestreados donde mayor
número de ejemplares de la familia se encontraron.
Los sitios en los que se encontraron los representantes de las familias más numerosas de
Prostigmata se presentan en la Cuadro 2. Algunas de ellas están en casi todos los perfiles de los
dos sitios muestreados como es el caso de Tydeidae, Tarsonemidae, Eupodidae, Cunaxidae y
Bdellidae, en orden de mayor a menor abundancia. En vista de que las condiciones del suelo no
son drásticamente diferentes en cuanto a constitución y pH, podemos suponer que los ácaros
encuentran condiciones propicias para desarrollarse en ambos sitios.
En muy notorio que los miembros de la familia Tydeidae (Prostigmata) son los más
numerosos de los ácaros extraídos de las muestras, particularmente en la hojarasca del sitio 1 con
828 ejemplares y el suelo del sitio 2 con 925. De dicha familia Kethley (1982) señala que, al
menos el género Lorryia se considera cosmopolita y prácticamente omnívoro. De este género se
encontraron ejemplares del grupo Bedfordensis.
Otros géneros identificados son Microtydeus, Paralorryia, Paratriophtydeus,
Triophtydeus y Tydeus (Cuadro 3). Entre dichos géneros se ha encontrado que Triophtydeus es el
más numeroso por muestra y con ejemplares machos, hembras y ninfas en una de las muestras de
hojarasca del sitio 1. Esto nos indica que la época de reproducción estaba en plena ejecución
durante la fecha de colecta de las muestras; además, varias de las hembras contenían huevecillos
en sus cuerpos (la duración del ciclo de vida de un tideido dura aproximadamente 8.4 días)
Algunas de las especies de la familia Tydeidae han sido consideradas depredadoras de
ácaros plaga como los tenuipalpidos con quienes comparten el hábitat, sin embargo se ha
comprobado experimentalmente que suelen alimentarse de las hojas de las plantas, de cadáveres
de otros ácaros o bien, quizás lo más común, de los hongos que crecen sobre las hojas de las
plantas en descomposición (McCoy, et al., 1970). De esta manera podríamos suponer que ocurre
algo similar con otras especies o géneros de la familia como Tydeus y Lorryia, pero aún hay
mucho que conocer acerca de los hábitos de alimentación de estos ácaros, tanto en la naturaleza
como en ambientes controlados en el laboratorio.
El conocer los hábitos de alimentación de los ácaros nos ayudaría a explicar, en parte, el o
los motivos por los cuales encontramos tal abundancia y variedad de formas en estos hábitats. Sin
embargo, algo importante que considerar es la estrategia de ciclo de vida que siguen las especies
de tideidos pues se ha visto que mientras las fases ninfales y adultas de especies de oribátidos
mantienen una densidad baja de manera estacional, la densidad de las fases larvales esta sujeta a
cambios de temperatura también estacionales en ambientes de hojarasca de bosque (Lebrun, et
al., 1991)
Consideramos que es necesario completar el estudio de todas las muestras que contengan
ejemplares de la familia Tydeidae y, una vez hecho esto, analizar la presencia de las especies y
los estados de desarrollo para esta familia.
Agradecimientos
Agradecemos el apoyo en la obtención, revisión y separación de las muestras al Biol.
Rafael Gaviño Rojas. Se agradece también la ayuda en la revisión de los Mesostigmata a la M. en

44
C. Patricia Chaires y a la Dra. Edith G. Estrada por la revisión de los Cryptostigmata. Este trabajo
se llevo a cabo gracias al apoyo del proyecto PAPIIT-IN104807 bajo la responsabilidad del Dr.
Pavel Krasilnikov, Laboratorio de Edafología, Facultad de Ciencias, UNAM.
Literatura Citada
Hoffmann, A. y M.G. López-Campos. Biodiversidad de los ácaros en México. CONABIO-
UNAM. Jiménez Ed. MéxicoD.F. 230p.
Lebrun, Ph., G. Van Impe, D. De Saint Georges-Gridelet, G. Wauthy & H. M. Andre. 1991. The
life strategies of mites. In: Schuster, R. & P. W. Murphy (Eds.). The Acari. Reproduction,
development and life-history strategies. Chapman & Hall. London. Chap. 1: 3-22.
McCoy, C. W., A. G. Selhime & R. F. Kanavel. 1970. The Feeding behaviour and biology of
Parapronematus acaciae (Acarina: Tydeidae). The Florida Entomologist. 52 (1): 13-19.
Momen F.M. & L. Lundqvist. 1995.The genus Tydeus (Acari: Prostigmata: Tydeidae). In
Southern Sweden; six new species. Acarologia 34 (1): 42-56.
Momen F.M. & T. Solhoy. 1996. A First record of the genus Tydeus in Himalaya, Tydeus
Lundqvisti nov. spec. (Acari: actinedida:Tydeidae). Acarologia 37 (1): 23-25.
Moreno-Moreno, J. A. & M. Mayagoitia-Penagos. 2008. Estudio de Ácaros mesostigmados del
suelo y su estado actual en México. En: Fauna del Suelo I. Micro, meso y macrofauna,
Colegio de Postgraduados. 33-49.
Palácios-Vargas, J. G. & R. Iglesias. 2004. Oribatei (Acari). En: Llorente-Bousquets, J., J.
Morrone, O. Yañez, e I. Várgas.(Eds.). Biodiversidad, Taxonomía y Biogeografía de
Artrópodos de México: Hacia una Síntesis de su Conocimiento. Vol. IV. CONABIO,
Facultad de Ciencias, IBUNAM, Las Prensas de Ciencias. 431-468.
Sommer, C. I. y S. Cram. Suelos. En: http://www.union.org.mx/guia/tesorosdelplaneta/
Suelos.htm, recuperado el 26 de marzo de 2009.
Vázquez I. M. y M. G. López-Campos. 2008. Ácaros Prostigmata (Acariformes: Prostigmata) de
tres tipos de suelo en una finca cafetalera en Huatulco, Oaxaca. En: Entomología
Mexicana, Vol. 7: 66-71. Colegio de Postgraduados, Primera Edición, 2008. Texcoco,
Estado de México. ISBN: 968-839-357-6
Vázquez, M. M. 1999. Catálogo de los ácaros oribátidos edáficos de Sian Ka’an, Q. Roo,
México. CONABIO, Universidad de Quintana Roo. 126 pp.

45
Fig. 1. Localización de los sitios de muestreo en el Cerro Santa Juana, Municipio de Opopeo, Michoacán.
Cuadro 1. Resumen del número de ácaros, por orden y por familia, en Santa Juana, Michoacán. SUE, suelo; HOJ,
hojarasca; Ind, individuos; Fam, familia. Aunque no se determinó la familia, se distingue un solo morfotipo
correspondiente a la superfamilias Crotonoidea, Damaeoidea así como al Cohorte Macropilina.
FAMILIAS SITIO 1 SITIO 2
Totales/Fam
Ind/Fam
(%)
Densidad
Ind/m² SUE HOJ SUE HOJ
MESOSTIGMATA
AMEROSEIDAE 0 0 1 9 10 2.80 10.53
ASCIIDAE 15 20 5 25 65 18.20 68.42
EVIPHIDIDAE 1 0 0 0 1 0.30 1.05
LAELAPIDAE 47 24 32 49 152 42.70 160.0
MACROCHELIDAE 0 0 0 5 5 1.40 5.26
RHODACARIDAE 2 0 0 0 2 0.56 2.10
SEJIDAE 1 0 0 0 1 0.30 1.05
VEIGAEIDAE 0 0 3 4 7 2.00 7.36
ZERCONIDAE 1 0 3 0 4 1.12 4.21
JUVENILES NO DET 12 49 16 32 109 30.62 114.73
PROSTIGMATA
BDELLIDAE 12 20 3 4 39 1.05 41.05
BIMICHAELIDAE 1 0 0 0 1 0.02 1.05
CALIGONELLIDAE 0 0 0 1 1 0.02 1.05
CUNAXIDAE 4 43 7 50 104 2.82 109.47
ERYTHRAEIDAE 1 0 2 0 3 0.08 3.15
EUPODIDAE 54 135 59 174 422 11.45 444.21
NANORCHESTIDAE 2 0 1 2 5 0.14 5.26
PENTHALODIDAE 0 0 0 1 1 0.02 1.05
PYGMEPHORIDAE 3 0 0 0 3 0.08 3.15
RHAGIDIIDAE 6 1 0 1 8 0.22 8.42
SCUTACARIDAE 0 0 1 0 1 0.02 1.05
SMARIDIDAE 0 0 0 1 1 0.02 1.05
TARSONEMIDAE 17 57 14 47 135 3.66 142.10
TROMBIDIDAE 6 1 0 5 12 0.32 12.63
TYDEIDAE 524 828 925 673 2950 80.03 3105.26
CRYPTOSTIGMATA
CEPHEIDAE 1 11 7 4 23 3.16 24.21
CROTONOIDEA 0 0 5 0 5 0.70 5.26
DAMAEOIDEA 0 3 1 6 10 1.40 10.52
EUPHTHIRACARIDAE 0 1 1 4 6 0.82 6.31
GALUMNIDAE 5 1 4 4 14 1.92 14.73
GYMNODAMAEIDAE 2 10 0 0 12 1.65 12.63
MICROZETIDAE 0 0 2 0 2 0.30 2.1

46
OPPIIDAE 155 18 75 15 263 36.13 276.84
PHTHIRACARIDAE 1 1 0 1 3 0.41 3.15
SCHELORIBATIDAE 40 100 38 87 265 36.40 278.94
MACROPILINA 0 0 5 0 5 0.70 5.26
JUVENILES NO DET 52 33 7 28 120 16.50 126.31
Cuadro 2. Número de ejemplares de las Familias más abundantes de ácaros Prostigmata entre los perfiles de cada
sitio. P1 – P5 = Perfil 1 – Perfil 5.
SITIO 1 SITIO 2
P1 P2 P3 P4 P5 P1 P2 P3 P4 P5
BDELLIDAE 7 9 4 5 12 3 1 1 2 1
CUNAXIDAE 2 2 7 24 17 17 5 19 9 7
EUPODIDAE 13 19 7 77 103 71 14 89 41 18
TARSONEMIDAE 21 31 1 6 17 5 12 14 28 2
TYDEIDAE 183 419 118 429 233 203 240 234 387 534
Cuadro 3. Géneros de la familia Tydeidae de una de las muestras del Sitio 1. S/ P / H, Sitio/Perfil/Habitat; AD,
Adulto; PN, Protoninfa; Num Ejem, Número de ejemplares; LV, Larva.
Género Edo. Des. S/ P/ H Num Ejem
Lorryia gpo. bedfordensis AD S1 P4 S 1
Lorryia gpo. bedfordensis PN S2 P4 Hj 1
Lorryia gpo. bedfordensis 3DN S2 P5 Hj 3
Microtydeus PN S2 P4 Hj 1
Microtydeus AD S2 P5 Hj 1
Paralorryia 1AD S2 P3 S 1
Paratriophtydeus ♀ S1 P5 Hj 1
Paratriophtydeus DN, AD S2 P3 S 2
Paratriophtydeus AD S2 P4 Hj 1
Paratriophtydeus PN S2 P5 Hj 1
Paratriophtydeus AD S2 P3 Hj 1
Triophtydeus 2DN, ♀ S1 P1 Hj 3
Triophtydeus 2♂, PN S1 P2 Hj 3
Triophtydeus 2♀ S1 P5 Hj 2
Triophtydeus AD S2 P2 S 1
Triophtydeus PN S2 P5 S 1
Triophtydeus LV S1 P1 Hj 1
Tydeus ♀ S1 P5 Hj 1
Tydeus 3♀, ♂ S1 P1 Hj 4

47
LAS ARAÑAS DEL SUELO DEL BOSQUE MESÓFILO DE MONTAÑA DE LA RESERVA
DE LA BIOSFERA VOLCÁN TACANÁ, CHIAPAS: EFECTOS DE LAS
PERTURBACIONES ANTROPOGÉNICAS
Soil spiders of the Montane Cloud Forest from the Biosphere Reserve Volcán Tacaná, Chiapas:
effects of human disturbances.
Guillermo Ibarra-Núñez1 y David Chamé-Vázquez
2.
1El Colegio de la Frontera Sur, Unidad
Tapachula. Carr. Antiguo Aeropuerto km. 2.5, Tapachula, Chiapas, 30700 México, 2Universidad
de Ciencias y Artes de Chiapas, Escuela de Biología. Tuxtla Gutierrez, Chiapas, México.
Palabras Clave: Araneae, hojarasca, Soconusco, bosque de niebla, impacto humano.
Introducción
Las arañas (Arachnida: Araneae) son uno de los siete ordenes animales más diversos en el
mundo (después de los coleópteros, himenópteros, lepidópteros, dípteros, hemípteros, y los
ácaros) con 40,700 especies, agrupadas en 3,733 géneros y 109 familias. Además, tienen una alta
capacidad de distribución y de adaptación, por lo que están presentes en casi todos los
ecosistemas terrestres (Coddington y Levi 1991; Platnick 2009). En 1996 Jiménez hizo el más
reciente análisis de la fauna de arañas de México, anotando 2,506 especies citadas, lo que
corresponde solo al 6.16 % del total mundial, y a solo un 67.7% de las especies conocidas
actualmente para el resto de Norteamérica (3,700 especies para Canadá y los Estados Unidos en
conjunto) (Ubick et al. 2005). Esto evidencia que aún falta por conocer una parte importante de la
fauna de arañas, pues por ser México un país megadiverso, debería tener entre 3 y 3.5 veces el
numero de artrópodos que el resto de Norteamérica (Llorente et al 1996). Para Chiapas, Jiménez
(1996) registró 281 especies de arañas, equivalente a un 11.2% del total nacional y solo un 7.6%
del resto de Norteamérica. Los estudios sobre la fauna de arañas de Chiapas se han realizado en
muy diversos hábitats ya sean naturales como selvas altas perennifolias, manglares, selvas bajas
subcaducifolias, encinares, como otros manejados por el hombre como cafetales, cacaotales,
potreros, maizales, y otros, pero ninguno de esos trabajos abordó el estudio sistemático de las
arañas del bosque mesófilo de montaña.
El bosque mesófilo de montaña (BMM) o bosque de niebla se desarrolla en áreas
tropicales con clima templado-húmedo, donde las lluvias frecuentes, la presencia constante de
nubosidad y niebla a lo largo del año producen una elevada humedad ambiental, favoreciendo el
desarrollo de una vegetación perenne y abundante, con la presencia de árboles en varios estratos,
numerosos arbustos, helechos y una gran variedad y cantidad de epífitas (Bubb et al. 2004;
Williams-Linera 2007). En México los BMM ocupan de 0.8 a 0.9% de la superficie del territorio
nacional (Williams-Linera 2007). Además de ser un tipo de vegetación raro, los BMM se
encuentran amenazados por diferentes factores: deforestación y conversión (terrenos agrícolas o

48
de pastoreo), como terrenos de caza y de colecta de madera para combustible, por la construcción
de caminos, que fragmentan los bosques y facilitan algunas de las otras amenazas. A estas
amenazas hay que agregar el cambio climático, ya que esto afecta las particulares condiciones
ambientales en que se desarrollan los BMM (Bubb et al. 2004; Williams-Linera 2007). En el
estado de Chiapas se encuentra cerca de la cuarta parte de la superficie total de BMM de México
(Arriaga et al. 2000), aún cuando en los últimos 30 años, en ese estado los bosques de niebla
perdieron 78% de su cobertura original (Solórzano y Oyama 2002). Sin embargo, a pesar de que
ocupan una superficie reducida, se estima que estos bosques albergan entre 10 y 12% de las
especies vegetales registradas para el país (Williams-Linera 2007), y un 30% de esas son especies
endémicas (Bubb et al. 2004). Algo similar ocurre con los grupos animales que se han estudiado
en los BMM, por lo cual estos hábitats son considerados de alta diversidad, particularmente por
su elevada proporción de especies endémicas (de 30 a 60 %), y porque continúan siendo fuente
de especies nuevas para la ciencia, aún de grupos considerados bien estudiados como las aves
(Bubb et al. 2004). De la fauna que habita en los BMM, los vertebrados han sido los más
estudiados, y solo recientemente algunos grupos de insectos (Bubb et al. 2004; Williams-Linera
2007), sin embargo no existe ningún estudio sobre las arañas habitantes de los BMM.
Materiales y Método
Se seleccionaron dos áreas de BMM dentro del polígono de la Reserva de la Biosfera
Volcán Tacaná con diferente grado de impacto antropogénico, una relativamente conservada y la
otra parcialmente perturbada, en función de la intensidad de su aprovechamiento (extracción de
madera y pastoreo de ganado) y separadas por unos 800 m. Ambas áreas localizadas en el Ejido
Talquián, Municipio de Unión Juárez, Chiapas (15°05′15″ - 15°05′37″ N; 92°05′55″ - 92°06′06″
W; 2010 - 2058 m snm), con un clima templado-húmedo C(m). En cada una de las áreas se
establecieron dos transectos de 50 m de largo, que se muestrearon 2 veces por mes, durante seis
meses (diciembre 2006 a mayo 2007). En cada transecto se emplearon dos métodos de colecta: 1)
se colocaron 10 trampas de caída (con solución de anticongelante al 50%), separadas 5 m entre sí,
para recuperarse a los 4 días, y 2) se colectaron 5 muestras de hojarasca de una superficie de 50 x
50 cm en bolsas de plástico, para procesarse el mismo día en el laboratorio en embudos de
Berlese durante dos a tres días. Los ejemplares colectados se preservaron en alcohol al 80% y se
determinaron a especie o en su defecto a morfoespecie, quedando depositados en la Colección de
Arácnidos de El Colegio de la Frontera Sur, Unidad Tapachula (ECOTAAR).Para las
estimaciones de diversidad y de similitud entre áreas se uso el programa EstimateS (Colwell
2006)
Resultados
Se colectaron 1794 ejemplares, de los cuales 13 no pudieron identificarse por ser ninfas
muy jóvenes. Los restantes 1781 ejemplares corresponden a 60 especies distintas, de 51 géneros
y 22 familias de arañas (Tabla 1). A la fecha se tienen plenamente identificadas 11 especies, de
las restantes 49 se ha determinado que al menos 22 son especies nuevas (incluyendo un género
nuevo), y las restantes 27 se han determinado a género, pero aún no se han determinado a
especie. De los 51 géneros, 47 tienen sólo una especie y de las 22 familias, 12 tienen un solo
género.
En el BMM conservado se detectaron 48 especies, 42 géneros y 20 familias, en cambio en
el BMM perturbado se detectaron 37 especies, 30 géneros y 15 familias. De las 48 especies de

49
BMM conservado, 23 (47.9%) se encontraron solo en esa área, mientras que de las 37 especies de
BMM perturbado, 11 (29.7%) se encontraron solo en esa área. De los 42 géneros encontrados en
BMM conservado, 38 (90.5%) tienen una sola especie, y de las 20 familias, 11 (55%) tienen un
solo género. De los 30 géneros encontrados en BMM perturbado, 27 (90%) tienen una sola
especie, y de las 15 familias, 9 (60%) tienen un solo género.
El empleo de dos estimadores de riqueza de especies basados en datos de abundancia,
ACE y Chao1, indican un valor estimado, como densidad de especies (Gotteli y Colwell 2001),
de 72 especies para el BMM conservado (24 especies más que las observadas), y de 42 a 43
especies para el BMM perturbado (5 a 6 especies más que las observadas) (Fig. 1).
Cuadro 1. Lista de especies de arañas colectadas en el suelo del bosque mesófilo de Montaña de Talquián, Municipio
de Unión Juárez, Chiapas.
FAMILIA GÉNERO ESPECIE FAMILIA GÉNERO ESPECIE
Agelenidae Melpomene transversa Mysmenidae Calodipoena colima
Novalena sp. Ochyroceratidae Ochyrocera sp.
Anyphaenidae Anyphaena trifida Speocera sp.
Anyphaena sp. 2 Oecobiidae Oecobius concinnus
Anyphaena sp. 3 Oonopidae Dysderina sp.
Anyphaena sp. 4 Oonops sp.
Anyphaena sp. 5 Scaphiella sp.
Araneidae Mangora ixtapan Stenoonops sp.
Micrathena sp. Género 1 sp.
Clubionidae Elaver sp. Género 2 sp.
Corinnidae Creugas sp. Pholcidae Anopsicus sp.
Megalostrata sp. Metagonia sp.
Piabuna sp. Salticidae Corythalia sp.
Trachelas sp. Ilargus sp.
Ctenidae Ctenus sp. Lyssomanes sp.
Dipluridae Euagrus carlos Naphrys sp.
Gnaphosidae Zimiromus sp. Género 1 sp.
Leptonetidae Archoleptoneta sp. Género 2 sp.
Linyphiidae Ceratinopsis sp. Scytodidae Scytodes sp.
Erigone sp. Tetragnathidae Chrysometa heredia
Fissiscapus sp. Leucauge sp.
Linyphia linguatula Theraphosidae Género 1 sp.
Meioneta sp. Theridiidae Phoroncidia sp.
Mermessus sp. 1 Stemmops questus
Mermessus sp. 2 Stemmops sp2
Mermessus sp. 3 Theridion sp. 1
Psilocymbium sp. Theridion sp. 2
Walckenaeria iviei Theridion sp. 3
Género 1 sp. Thymoites puer
Lycosidae Trochosa sp. Zodariidae Tenedos sp.

50
Del total de 60 especies registradas, 25 fueron detectadas en ambos sitios (especies
compartidas), sin embargo el número estimado de especies compartidas (Chao-shared, basado en
datos de abundancia) se eleva a 30 especies. Las estimaciones de similitud entre las dos parcelas,
indican una similitud cualitativa moderada (índice de Sorensen = 0.588, índice de Jaccard =
0.417), en tanto que la similitud cuantitativa va de moderada (índice de Bray-Curtis = 0.441) a
elevada (índice de Morisita-Horn = 0.804). De las 22 especies nuevas 10 se comparten entre los 2
sitios; de las no compartidas hay 6 en BMM conservado, y 6 en BMM perturbado.
Fig. 1. Curvas de acumulación de especies observadas y estimadas para los dos sitios de estudio. C = BMM
conservado, P = BMM perturbado, Sobs = especies observadas, ACE = Estimador de Cobertura basado en datos de
Abundancia, Chao1 = Estimador 1 de Chao (Colwell, 2006).
Del total de ejemplares (1781), el 74.6% (1328) se colectaron en la parcela conservada, y
el 25.4% (453) en la parcela perturbada, evidenciando una diferencia significativa en abundancia
(chi cuadrada, p<0.0001). Esta marcada diferencia se presentó incluso en cada una de las dos
técnicas de muestreo por separado: conservado-hojarasca 847 (47.6%) vs. perturbado-hojarasca
221 (12.4%) (p<0.0001); conservado-trampas de caída 481 (27.0%) vs. perturbado-trampas de
caída 232 (13.0%) (p<0.0001).
Discusión
En un estudio sobre la fauna arañas de un bosque de pino-encino en el Estado de México,
Medina (2002) encontró 26 especies de arañas del suelo correspondientes a 22 géneros y 14
familias. Por otra parte Ruiz (2004) e Ibarra et al. (2004) estudiaron la fauna de arañas del suelo
en un cacaotal de la región del Soconusco, Chiapas encontrando 52 especies de 43 géneros y 17
familias. Los resultados del presente estudio revelan que el BMM, independientemente de su
estado de conservación, alberga una elevada riqueza de arañas, sobre todo considerando que los
valores observados corresponden solamente a un par de sitios estudiados durante un periodo de 6
meses. El número de especies no descritas, y en consecuencia de especies endémicas,
0
10
20
30
40
50
60
70
80
90
1 2 3 4 5 6 7 8 9 10 11 12
esp
ecie
s
muestras
C Sobs
C ACE
C
Chao1P Sobs

51
corresponde a un 36.7% del total de especies de ese hábitat, sin embargo es posible que este
porcentaje sea mayor sí algunas de las especies aún no determinadas también resultan especies
nuevas. El porcentaje de endemismos observado es similar a los que señalan Bubb et al. (2004)
para otros grupos de organismos que habitan en este tipo de vegetación y que se considera se
debe en parte a su condición de “islas ecológicas”.
Las elevadas proporciones de especies únicas (de un género), o géneros únicos (de una
familia) revelan una elevada diversidad taxonómica (Magurran 2004) para ambos sitios
estudiados, lo cual revela la importancia de estos hábitats como sitios de utilidad en la
conservación de la biodiversidad.
Las curvas derivadas de los estimadores de riqueza indican que el muestreo resultó más
completo en el sitio perturbado que en el conservado, lo que se evidencia por la mayor
aproximación a la asíntota de las curvas del BMM perturbado, de manera que no se puede esperar
encontrar muchas más especies aún cuando se incremente el esfuerzo de muestreo. En cambio,
para el BMM conservado, las dos curvas estiman que un mayor esfuerzo de muestreo puede
revelar hasta un 50% más de especies que las ya detectadas. Así estos indicadores evidencian una
diferencia en la riqueza estimada para las dos condiciones estudiadas. Por otra parte los datos
sobre la riqueza taxonómica y los de similitud no muestran diferencias importantes entre las dos
condiciones de BMM.
A pesar de haber realizado el mismo esfuerzo de muestreo en los dos sitios, se observa
una significativa mayor abundancia en el BMM conservado, diferencia que se sostiene para cada
uno de los dos métodos de muestreo empleados, y que según el método de colecta va del doble
(trampas de caída) al cuádruple (hojarasca-Berlese) de abundancia, revelando un fuerte impacto a
la densidad de las poblaciones de arañas, muy probablemente relacionado con las diferencias en
la condiciones ambientales de la hojarasca entre los 2 sitios.
Estos resultados evidencian una similitud moderada en la composición de las especies,
pero al mismo tiempo una clara diferencia en la abundancia y la riqueza de las comunidades de
arañas entre los dos sitios estudiados, a pesar de que se encuentran sólo a 800 m de distancia una
de otra y prácticamente a la misma altitud. Las diferencias entre las comunidades de los 2 sitios,
muy posiblemente reflejan las alteraciones de condiciones ambientales del suelo, que se tornan
menos favorables para muchas especies de arañas en el BMM perturbado
Agradecimientos
Al CONACYT por el apoyo al proyecto 067254- AC-2006-52923 del Fondo Institucional.
Al Tec. Académico José Álvaro García Ballinas (ECOSUR), por su participación en las colectas
y procesamiento del material colectado. Al Dr. Jeremy Miller (California Academy of Sciences,
EEUU) por la corroboración de un taxon.
Literatura Citada
Arriaga L., J. M. Espinoza, C. Aguilar, E. Martínez, L. Gómez y E. Loa (coordinadores). 2000.
Regiones terrestres prioritarias de México. Comisión Nacional para el Conocimiento y uso
de la Biodiversidad, México.
Bubb P., I. May, L. Miles, J. Sayer. 2004. Cloud Forest Agenda. UNEP-WCMC, Cambridge,
U.K. http://www.unep-wcmc.org/resources/publications/UNEP_WCMC_bio_series/20.htm
Coddington J. A. y H. W. Levi 1991. Systematics and the evolution of spiders (Araneae). Annu.
Rev. Ecol. Syst., 22: 565-592.

52
Colwell R. K. 2006. EstimateS: Statistical estimation of species richness and shared species from
samples. Version 8. Persistent URL <purl.oclc.org/estimates>
Gotteli N.J., Colwell R. K. 2001. Quantifying biodiversity: procedures and pitfalls in the
measurement and comparison of species richness. Ecol. Lett. 4:379–339
Ibarra-Núñez G., E. B. Moreno Molina, A. Ruiz Colemares, M. Trujillo Olivera y J. A. García
Ballinas. 2004. Las arañas tejedoras (Araneidae, Tetragnathidae, Theridiidae y Uloboridae)
de una plantación de cacao en Chiapas, México. En: Morales M. A., Ibarra G. M., Rivera
G. A. P. y Stanford C. S. (Eds.). Entomología Mexicana. Vol. 3. (ISBN: 968-839-411-4). El
Colegio de Postgraduados, Montecillo, Edo. de México. pp. 38-41.
Jiménez M. L. 1996. Araneae. En: J. Llorente Bousquets, A. N. García Aldrete y E. González
Soriano (Eds.), Biodiversidad, taxonomía y biogeografía de artrópodos de México: hacia
una síntesis de su conocimiento. Instituto de Biología, UNAM.: 83-101.
Llorente Bousquets J., E. González S., A. N. García Aldrete y C. Cordero. 1996. Breve Panorama
de la taxonomía de artrópodos en México.En: J. Llorente Bousquets, A. N. García Aldrete y
E. González Soriano (Eds.), Biodiversidad, taxonomía y biogeografía de artrópodos de
México: hacia una síntesis de su conocimiento. Instituto de Biología, UNAM.: 3-14.
Magurran A. E. 2004. Measuring Biological Diversity. Blackwell Science Ltd. Oxford, U.K. 256
pp.
Medina Soriano F: J. 2002. Las arañas y su distribución temporal en un bosque de San Martín
Cachihuapan, Municipio de Villa del Carbón, Estado de México. Tesis de Licenciatura en
Biología. UNAM, Facultad de Estudios Superiores, Campus Iztacala. 129 pp.
Platnick N. I. 2009. The world spider catalog, version 9.5. American Museum of Natural History,
online at: http://research.amnh.org/entomology/spiders/catalog/index.html.
Ruiz Colmenares A. 2004. Diversidad de arañas de suelo en una plantación de cacao, Mpio.
Tuxtla Chico, Soconusco, Chiapas, México. Tesis de Licenciatura en Biología. Universidad
de Ciencias y Artes de Chiapas, Escuela de Biología; Tuxtla Gutiérrez, Chiapas. 67 pp.
Solórzano S y K. Oyama. 2002. El Quetzal, una especie en peligro de extinción. Biodiversitas,
45: 2-6.
Ubick D., P. Paquin, P.E. Cushing, y V. Roth (eds.). 2005. Spiders of North America: an
identification manual. American Arachnological Society. 377 pp.
Williams-Linera G. 2007. El bosque de niebla del centro de Veracruz: ecología, historia y destino
en tiempos de fragmentación y cambio climático. CONABIO – Instituto de Ecología A. C.,
Xalapa, Veracruz, México. 208 pp.

53
ESTIMADORES DE DENSIDAD POR KERNEL PARA ANALIZAR DISTRIBUCIONES DE
FRECUENCIA DE TALLA EN ARAÑAS JUVENILES.
The use of kernel density estimators to analyze juvenile length-frequency distributions of spiders
Irma Gisela Nieto-Castañeda1,2
, María Luisa Jiménez-Jiménez1 e Isaías Hazarmabeth Salgado-
Ugarte2.
1Centro de Investigaciones Biológicas del Noroeste, Mar Bermejo 195, Col. Playa Palo
de Santa Rita, La Paz, B.C.S. 23096, México 2Facultad de Estudios Superiores Zaragoza,
UNAM, Batalla 5 de mayo s/n Esq. Fuerte de Loreto. Col. Ejército de Oriente, Iztapalapa, C.P.
09230 México D.F., México. [email protected], [email protected]
Palabras Clave: EDKs, histogramas, distribución de frecuencias de tallas.
Introducción
La mayoría de los aracnólogos han utilizado la estimación directa del número de estadios
para describir el ciclo de vida de las araña; sin embargo existen otros métodos basados en el
análisis de la longitud de frecuencia de tallas a lo largo del tiempo (métodos indirectos). Estas
distribuciones de frecuencia han sido comúnmente analizadas mediante histogramas de
frecuencia (Wright & Coyle 2000). Sin embargo este procedimiento presenta varios problemas
incluyendo la dependencia del origen y del ancho del intervalo, la discontinuidad y el uso de
intervalos de ancho fijo. Estos problemas han motivado el interés de los estadísticos en métodos
más eficientes aunque computacionales intensivos. En este sentido, los estimadores de densidad
por kernel (EDKs) no dependen de la posición del origen y son estimadores continuos (Silverman
1986). Además, existen diferentes métodos para elegir el ancho del intervalo (Härdle 1991; Scott
1992). Estos estimadores no paramétricos resultan en figuras que son más suavizadas que los
histogramas de frecuencia, permitiendo el fácil reconocimiento de características tales como
valores extraordinarios, sesgo y multimodalidad (Salgado-Ugarte et al. 1993, 1995). La mayoría
de estos métodos han sido empleados para describir el crecimiento de peces adultos y juveniles;
sin embargo no han sido utilizados para describir tallas de clases en arañas durante su ciclo de
vida. En este trabajo se presenta el uso del EDKs como una técnica moderna para analizar la
distribución de frecuencias de tallas en arañas, tomando como modelo a los juveniles de la
especie Syspira tigrina Simon, 1885.
Materiales y Método
El área de llamada “El Comitán”, La Paz, se localiza al oeste de la ciudad de La Paz en
Baja California Sur, México (24°7′N, 110°25′W, 20 m a.s.l.). Se trazaron dos transectos de 100 m
de longitud, en cada uno se colocaron 10 trampas de caída con una separación de 10m entre sí,
utilizando anticongelante como preservador. Las trampas permanecieron activas de julio del 2005
a julio del 2006, removiendo el contenido de las mismas mensualmente y preservándolos en
etanol al 70%. Todas las arañas pertenecientes a S. tigrina, se les midió la longitud de la tibia I.
Con las medidas obtenidas, se construyeron EDKs mensuales utilizando los programas
propuestos por Salgado-Ugarte et al. 1995, 1997. El ancho de banda crítica se basó en la prueba

54
bootstrap de multimodalidad (Silverman, 1981, 1986). La distribución de las densidades
resultantes fueron transformadas a frecuencias suavizadas y descompuestas en sus componentes
gaussianos con el programa Stata v. 9.1 (Salgado-Ugarte et al. 1994), utilizando el método
propuesto por Bhattacharya (1967).
Resultados
Durante este estudio se midieron y analizaron un total de 660 tibias de juveniles de S.
tigrina. Las distribuciones de frecuencia suavizada mensuales o EDKs, y los histogramas de
frecuencia pueden observarse en la figura 1. El ancho de banda empleado (indicado por la prueba
de multimodalidad) para construir de los EDKs varió entre 0.09 y 0.32 mm dependiendo del
número de observaciones y su variación. En la mayoría de los casos se observó una moda
dominante con varias modas menores.
Fig. 1. EDKs e histogramas de frecuencia mensuales, donde se observan las frecuencias de tallas de S. tigrina
durante un ciclo anual.

55
Discusión y Conclusiones
En este trabajo, los EDKs mostraron que durante los meses de noviembre a diciembre, es
probable que ocurra la época principal de reclutamiento, porque se encontraron las tallas más
pequeñas y abundantes (< 0.7 mm de longitud tibial, N ≥ 69) las cuales están presentes en menor
frecuencia durante los meses de agosto a enero. Esto sugiere que debe existir un continuo
reclutamiento, lo que se traduce en que las hembras deben ovispositar en diferentes meses del año
o más de una vez en el año, al menos. Por su parte, la baja frecuencia de tallas mayores sugiere
que estén continuamente muriendo, quizá por depredación o alguna otra causa, o bien modifiquen
un poco sus hábitos errantes y no sean capturadas con frecuencia en las trampas. A partir enero-
febrero, se observa un aumento en el crecimiento de las arañas, donde las tallas dominantes están
siempre por arriba de los 0.7 mm de longitud tibial, y continúan incrementando entre los meses
de mayo a julio, lo que sugiere, que se aproxime el periodo de madurez sexual y por ende el
periodo reproductivo.
Esta información hubiese sido imposible de detectar mediante los histogramas de
frecuencia tradicionales, debido a que dependiendo de la amplitud del ancho del intervalo y el
origen, los grupos de tallas resultantes varían considerablemente (Salgado-Ugarte 2002), es por
ello que los EDKs se proponen como una alternativa más robusta en la interpretación de
fenologías de arañas por métodos indirectos.
Los trabajos previos, donde se ha determinado la estructura de tallas empleando
histogramas de frecuencia, fueron realizados solo con arañas tejedoras o arañas errantes que
habitan en la vegetación en zonas holárticas (Toft 1976, Aiken y Coyle 2000, Wrigth y Coyle
2000). Estos trabajos muestran una clara sincronía del ciclo de vida de los organismos, lo que se
traduce en gráficas con modas marcadas y huecos entre las mismas, que los autores interpretan
como estadios y límites entre los mismos, respectivamente; sin embargo, estas conclusiones
carecen de algún soporte estadístico.
Agradecimientos
Se agradece al Dr. Frederick A. Coyle, por sus sugerencias al trabajo; a Miguel Mauricio
Correa-Ramírez y Carlos Palacios-Cardiel por su poyo durante el trabajo de campo. Al Consejo
Nacional de Ciencia y Tecnología (SEMARNAT-CONACYT C01-0052) por los recursos
otorgados para este proyecto.
Literatura Citada
Aiken, M. & F.A. Coyle. 2000. Habitat distribution, life history and behavior of Tetragnatha
spider species in the Great Smoky Mountains National Park. Journal of Arachnology
28:97–106.
Bhattacharya, C.G. 1967. A simple method of resolution of a distribution into Gaussian
components. Biometrics 23: 115-135.
Härdle,W.K. 1991. Smoothing techniques.With implementation in S. Springer Verlag, New
York.
Salgado-Ugarte, I.H. 2002. Suavización no paramétrica para análisis de datos. FES Zaragoza-
DGAPA UNAM (PAPIIT IN217596; PAPIME 192031): 139p.

56
Salgado-Ugarte, I.H., M. Shimizu and T. Taniuchi. 1993. Exploring the shape of univariate data
using kernel density estimators. Stata Technical Bulletin 16: 8-19.
Salgado-Ugarte, I.H., M. Shimizu and T.Taniuchi. 1994. Semi-graphical determination of
Gaussian components in mixed distributions. Stata Technical Bulletin 18: 15-27.
Salgado-Ugarte, I.H., M. Shimizu and T. Taniuchi. 1995. Practical rules for bandwidth selection
in univariate density estimation. Stata Technical Bulletin 27: 5-19.
Salgado-Ugarte, I.H., M. Shimizu and T. Taniuchi. 1997. Nonparametric assessment of
multimodality for univariate data. Stata Technical Bulletin 38: 27-35.
Scott, D.W. 1992.Multivariate density estimation: theory, practice and visualization. John Wiley
& Sons,New York.
Silverman, B.W. 1981. Using kernel density estimates to investigate multimodality. Journal of
the Royal Statistical Society, B 43: 97-99.
Silverman, B.W. 1986. Density estimation for statistics and data analysis. Chapman and Hall,
London.
Toft, S. 1976. Life histories of spiders in a Danish beech wood. Natura Jutlandica 19:5–40.
Wright, R.L, F.A. Coyle. 2000. Habitat distribution, life history and behaviour of Neriene species
in the Great Smoky Mountains National Park (Araneae, Linyphiidae). Bulletin of the
British Arachnological Society 11:293–304.

57
NUEVOS REGISTROS Y DISTRIBUCIÓN ACTUALIZADA DE Diplocentrus zacatecanus
Hoffmann, 1931 (SCORPIONES: DIPLOCENTRIDAE)
New records and updated distribution of Diplocentrus zacatecanus Hoffmann, 1931 (Scorpiones:
Diplocentridae)
Javier Ponce-Saavedra1, Oscar F. Francke B.
2 y Ana F. Quijano-Ravell
1.
1Laboratorio de
Entomología “Biol. Sócrates Cisneros Paz” Facultad de Biología. Universidad Michoacana de
San Nicolás de Hidalgo. Morelia, Michoacán. C. P. 58060. [email protected]. 2Departamento de Zoología, Instituto de Biología, Universidad Nacional Autónoma de México,
Apartado Postal 70-153, 04510, México, D. F.
Palabras Clave: Alacranes, Diplocentridae, distribución geográfica.
Introducción
El género Diplocentrus Peters, 1861 con aproximadamente 50 especies descritas a la
fecha, se encuentra registrado desde el Sur de los Estados Unidos (Arizona, Nuevo México y
Texas) hasta Honduras (Sissom y Fet, 2000). México representa el área geográfica de mayor
riqueza para el género con 45 especies registradas, incluyendo cinco especies compartidas en el
norte del país con los Estados Unidos de Norteamérica. El género ha sido trabajado ampliamente
por el Dr. Oscar Francke atendiendo aspectos de taxonomía e historias de vida, conocimiento
necesario de comprender la variabilidad ontogenética del grupo (Francke, 1975; 1977a; 1977b;
1978a; 1978b; 1981; 1982; Sissom y Francke, 1998; Francke y Ponce, 2005). A pesar de ser un
género de amplia distribución en México, hay varios estados de la República Mexicana en los
que no hay registros para esta familia; por ejemplo en Michoacán Ponce y Beutelspacher (2001)
registran los primeros Diplocentrus con base en dos ejemplares determinados erróneamente como
D. keyserlingii Karsch, taxón que actualmente se considera restringido al estado de Oaxaca,
estado que actualmente tiene el mayor número de especies registradas para la familia con 10 (Fet
et al., 2000), a las que se deben adicionar otras dos que ya se tienen descritas y enviadas para su
publicación y al menos 2 taxones específicos más que esperan ser trabajados (Santibañez, 2009).
El género requiere de trabajo ya que se tienen al menos otras 18 poblaciones de diferentes partes
de México que probablemente representen nuevas especies. Otros estados con registros para este
género son: Aguascalientes (1 especie), Nuevo León (3), Yucatán (2) Guerrero (2),Tamaulipas
(1) Jalisco (1), Nayarit (1), Sinaloa (1), Sonora (3), Quintana Roo (1), Distrito Federal, Campeche
(1), Veracruz (1), Chiapas (1), Morelos (1), Puebla(1), Coahuila (1), Zacatecas (1) (Fet et al.,
2000, Lourenço y Sissom, 2000).
Diplocentrus zacatecanus fue primeramente descrito por Hoffmann (1931) como una
subespecie de Diplocentrus keyserlingii Karsch y elevado a nivel específico por Sissom y Walker
(1992). Tiene como localidad tipo registrada a Tepezalá en el estado de Aguascalientes, aunque
Beutelspacher (2000) refiere también una localidad en Zacatecas. El epíteto subespecífico se debe
a malas determinaciones de material proveniente de Zacatecas y que posteriormente fueron
aclaradas dejando a la fecha a D. keyserlingii con distribución en Oaxaca, D. tehuacanus

58
Hoffmann en Morelos, Oaxaca y Puebla y D. zacatecanus en Aguascalientes (Francke, 1977a,
Fet et al., 2000; Lourenço y Sissom, 2000, Santibañez, 2009), con la observación de que éste
complejo de especies y de hecho todo el género, deberá ser estudiado sistemáticamente para
clarificar el estatus de las especies actualmente reconocidas y varias poblaciones que presentan
problemas para su determinación.
El objetivo de este trabajo fue el de actualizar la información sobre Diplocentrus
zacatecanus en lo que refiere a su distribución geográfica para ayudar a clarificar la situación
taxonómica de estas poblaciones y tener mayor conocimiento sobre este género y en particular
sobre el grupo de especies afines a D. keyserlingii que aún representa un problema taxonómico
importante en la sistemática de la familia Diplocentridae.
Materiales y Métodos
Se revisó material procedente de 16 localidades ubicadas en los estados de Aguascalientes
(2), Zacatecas (3), Durango (1), Hidalgo (1), Estado de México (2), Michoacán (1), Querétaro
(4), San Luis Potosí (1) y Guanajuato (1) (Cuadro 1).
Se tuvo información correspondiente a 71 ejemplares machos y 41 hembras de los que se
obtuvo una vez revisada la descripción y determinada su pertenencia a la especie de interés, se
hizo el conteo pectinal para 47 machos encontrando que en esta especie puede variar de 12
dientes (22 peines equivalentes al 23.5% de la muestra) a 17 (2) con moda= 13 (35 peines
equivalentes al 37% % del material revisado); también se encontraron 22 peines con 14 dientes
por lo que el 84% de la muestra tuvo conteo pectinal entre 12 y 14 dientes; mientras que para las
hembras se pudieron revisar los 41 ejemplares en los que el conteo fue de 8 (2 peines) a 14
dientes pectíneos (2 peines), con una moda de 11 y 12 dientes (29 peines para cada conteo que
representa el 34.5% de la muestra, 69% de los peines con estos valores).
También se revisó el conteo de las espinas en los tarsómeros, característica que junto con
el conteo pectinal representan buena evidencia para la identificación de la especie. Una vez
seguros de que el material corresponde a Diplocentrus zacatecanus, se procedió a ubicar las
localidades por medio de sus coordenadas geográficas y se obtuvo la altitud sobre el nivel del
mar para hacer una primera generalización sobre el hábitat de esta especie en el área de
distribución así determinada, elaborando un mapa con los puntos registrados usando Google
Earth© 2009 y posteriormente haciendo el dibujo correspondiente.
Resultados y Conclusiones
Se registra formalmente a Diplocentrus zacatecanus Karsch por primera ocasión para los
estados de: Durango, Estado de México, Guanajuato, Hidalgo, Michoacán, Querétaro y San Luis
Potosí, ampliando a nueve estados la distribución hasta ahora reconocida para la especie sólo en
el estado de Aguascalientes y Zacatecas.
Se puede establecer que la especie se distribuye en las zonas altas (1560 a 2514 msnm)
del centro de la república Mexicana en climas templado fríos (Figura 1) y se puede distinguir de
otros taxones similares por el conteo pectinal de los machos entre 12 y 14 dientes y 11 a 12 en las
hembras, en combinación con una fórmula de espinas en los tarsómeros de 5/6; 5(6)/7; 7(6)/7;
7(6)/7, en las patas 1 a 4 respectivamente.

59
Cuadro1. Localidades registradas para Diplocentrus zacatecanus.
Estado Localidad (es) * Coordenadas geográficas Altitud
(msnm) Longitud Latitud
Aguascalientes Tepezala (L1) 22° 13' 20.94'' 102° 10' 17.53'' 2100
Durango Loma alta, Mpio. de San Juan de
Guadalupe (L2)
24°37'58.05''
102°45'2.80"
1560
Vicente Guerrero (L3) 23° 43' 53.99'' 103° 59' 14.55'' 1922
Estado de México La Loma, Mpio. de Aculco (L4) 20° 06’ 07.03'' 99° 51' 25.71'' 2450
San Jose Deguedo, Mpio. de
Soyaniquilpan (L5)
20° 05' 09.04''
99° 33' 48.78''
2514
Guanajuato Coroneo (L6) 20° 11' 54.81'' 100° 21' 56.60'' 2280
Hidalgo Michimaloya, Mpio. de Tula de
Allende (L7)
20° 05' 34.88''
99° 23' 57.93''
2078
Michoacán Contepec (L8) 19° 57' 10.44'' 100° 09' 48.90'' 2480
Querétaro Cadereyta (L9) 20° 41' 38.29'' 99° 48' 36.21'' 2037
El organal Mpio de Cadereyta (L10) 20° 37' 09.49'' 99° 38' 27.98'' 1733
Huimilpan (L11) 20° 22' 22.23'' 100° 16' 27.48'' 2288
Vizarrón de Montes, Cadereyta de
Montes (L12)
20° 50' 03.63''
99° 43' 09.27''
2055
San Luis Potosí Villa de Ramos (L13) 22° 49' 53.21'' 101° 54' 36.01'' 2200
Zacatecas Emiliano Zapata, Mpio. Sain Alto
(L14) 23° 34' 16.66'' 103° 18' 02.75'' 2067
Cañitas de Felipe Pescador (L15) 23° 36' 20.10'' 102° 43' 47.50'' 2027
Zacatecas (L16) 22° 46' 19.52'' 102° 34' 32.01'' 2440
* La clave entre paréntesis, corresponde a la etiqueta en el mapa de la fig. 1.
Fig. 1. Mapa de distribución de Diplocentrus zacatecanus. Para el nombre y coordenadas de las localidades, ver el
cuadro 1.
Agradecimientos
Se agradece a Víctor S. Mondragón N. de la Universidad Michoacana de San Nicolás de
Hidalgo, las facilidades para recolectar los ejemplares de Coroneo, Guanajuato en los terrenos de
su propiedad.
Literatura Citada

60
Fet, V., W. D. Sissom, G. Lowe y M. E. Braunwalder. 2000. Catalog of the Scorpions of the
World (1758-1998). New York Entomol. Soc., New York., 690 pp.
Francke, O. F. 1975. A new species of Diplocentrus from New Mexico and Arizona
(Scorpionida: Diplocentridae). Journal of Arachnology 2: 107- 118.
Francke, O. F. 1977a. The genus Diplocentrus in the Yucatan Peninsula with description of two
new troglobites. Bulletin of the Association for Mexican cave studies., 6:49-61
Francke, O. F. 1977b. Scorpions of the genus Diplocentrus from Oaxaca, México. Journal of
Arachnology., 4:145-200.
Francke, O. F. 1978a. New Troglobite scorpion of Genus Diplocentrus (Scorpionida:
Diplocentridae). Ent. News 89 (1-2): 39 – 45.
Francke, O. F. 1978b. Systematic revision of Diplocentrid Scorpions (Diplocentridae) from
circum Caribbean Lands. Special Publications of the Museum, Texas Tech University,
Lubbock, Texas, E.U.A.14:1-92
Francke, O. F. 1981. Birth behavior and life history of Diplocentrus spitzeri (Scorpiones:
Diplocentridae). Southwestern Naturalist., 25:517-524.
Francke, O. F. 1982. Parturition in scorpions (Arachnida, Scorpiones): A review of the ideas.
Revue arachnologique., 4:27-37.
Francke O. F. y Ponce S. J. 2005. A new species of Diplocentrus (Arachnida: Scorpiones) from
Michoacan, Mexico. Revista Mexicana de Biodiversidad vol. 76: 49- 53
Hoffmann, C. C. 1931. Monografías para la Entomología Médica de México. Monografía No. 2,
Los Escorpiones de México (primera parte) Diplocentridae, Chactidae, Vejovidae. Anales
del Instituto de Biología, Tomo II. México., pp. 291-408.
Lourenço, W. R. y W. D. Sissom. 2000. Scorpiones. En: J. Llorente Bousquets, E. González
Soriano y N. Papavero (Eds.). Biodiversidad, taxonomía y biogeografía de artrópodos de
México. Hacia una síntesis de su conocimiento. Universidad Nacional Autónoma de
México, México., 2:115-135.
Ponce, S. J y C. R. Beutelspacher B. 2001. Alacranes de Michoacán UMSNH. Ediciones
Michoacanas. Morelia, Michoacán, México., 103 pp.
Prendini, L. 2000. Phylogeny and classification of the Superfamily Scorpionoidea Latreille 1802
(Chelicerata, Scorpiones): An exemplar approach. Cladistics, 16, 1–78.
Prendini, L. y W. Wheeler. 2005. Scorpion higher phylogeny and classification, taxonomic
anarchy, and standards for peer review in online publishing. Cladistics, 21: 446–494.
Santibañez-López, C. E. 2009. Escorpiofauna (Arachnida: Scorpiones) de la Sierra Norte del
Estado de Oaxaca. Tesis de Maestría. Instituto de Biología. Universidad Nacional
Autónoma de México. 122 pp.
Sissom, W. D. y O. F. Francke. 1998. Taxonomic comments on the scorpion Diplocentrus
mexicanus Peters and its subspecies (Scorpiones: Diplocentridae). Mitteilungen aus dem
Museum für Naturkunde (Berlin), Zoologische Reihe 74: 259- 260.
Sissom, W. D. y V. Fet. 2000. Family Diplocentridae Karsch, 1880. In Catalog of the Scorpions
of the World (1758-1998), V. Fet et al (eds.). The New York Entomological Society. p.
329- 354.

61
DISTRIBUCIÓN GEOGRÁFICA DE Vaejovis variegatus Pocock, 1898 (SCORPIONES:
VAEJOVIDAE) EN EL ESTADO DE MICHOACÁN, MEXICO.
Geographic distribution of Vaejovis variegatus Pocock, 1898 (Scorpiones: Vaejovidae) in Michoacan
state, Mexico.
Ana F. Quijano-Ravell, Javier Ponce-Saavedra. Laboratorio de Entomología “Biol. Sócrates
Cisneros Paz” Facultad de Biología. Universidad Michoacana de San Nicolás de Hidalgo.
Morelia, Michoacán. C. P. 58060. [email protected].
Palabras Clave: Vaejovidae, V. variegatus, distribución, Michoacán.
Introducción
La familia Vaejovidae cuenta con 10 géneros, 157 especies y 19 subespecies (Coddintong
et al., 2004; Prendini y Wheeler, 2005; Rein, 2009). Esta familia incluye sólo especies de
América del Norte (en el suroeste de Canadá, Estados Unidos, Mexico) y Guatemala en
Centroamérica. Vaejovis Koch, 1836, es el género de escorpiones mejor representado en México
y hasta 1998 se registraban 70 especies (Sissom, 2000) que en los últimos años 74 especies
(Coddintong et al., 2004; Prendini y Wheeler, 2005; Rein, 2009).
El género no es monofilético y de acuerdo con Sissom (2000), para su estudio se
reconocen cinco grupos: el grupo “eusthenura” diagnosticado por Williams (1970), transferido
recientemente al género Hoffmannius Soleglad y Fet, 2008; el grupo “intrepidus” (propuesto por
Hoffmann, 1931 y que contiene al escorpión más grande del género, recientemente transferido al
género Thorellius Soleglad y Fet, 2008; el grupo “mexicanus” (también propuesto por Hoffmann,
1931), el grupo “nitidulus” (diagnosticado por Sissom y Francke (1985), 6 especies de este grupo
han sido transferidas al género Franckeus Soleglad y Fet, 2005, en tanto que 12 permanecen en el
grupo “nigrescens”) y finalmente el grupo “punctipalpi” (propuesto por Williams en 1971) y
transferido recientemente al género Kochius Soleglad y Fet, 2008).
Vaejovis exhibe una amplia gama de preferencias de hábitat: algunos viven en arena
(psammofilos), otros son habitantes de grietas en roca o acantilados (rupícolas), algunos habitan
bajo piedras (lapidícolas), y también los hay que viven en cuevas (troglobios) (Lourenço y
Sissom, 2000).
Vaejovis variegatus Pocock, especie que se ubicaría en el grupo “mexicanus”. El grupo
puede reconocerse con base en el margen interno del dedo fijo de la quela del pedipalpo con seis
hileras de gránulos divididas por 6 grandes gránulos; la serie de tricobotrias ib-it ubicada en la
base del dedo fijo y no sobre la palma de la quela (Sissom, 2000). Las especies del grupo
comparten un patrón de color distintivo el cual consiste en manchas oscuras en todo el cuerpo,
sobre un color de fondo obscuro a claro (con algunas excepciones en la parte ventral del
mesosoma) y que al menos, en los segmentos III y IV del metasoma, hay una carinación ventral
más o menos granular (Sissom, 1989).
Vaejovis variegatus fue considerada una subespecie de V. punctatus Karsch por Hoffmann
(1931) y la mayoría de los autores posteriores. Fue devuelta a la condición de especie válida por
Beutelspacher (2000) sin ninguna justificación para el cambio y lo registra para Tacámbaro en

62
Michoacán y diferentes localidades de Guerrero, Edo. De México, Morelos, Oaxaca y Puebla. En
el mismo año, Fet et al. en el Catálogo de los Escorpiones del Mundo reconocen la especie como
válida registrándola para Guerrero, Edo. de México, Morelos y Puebla, pero no hay registro de
esta especie para el Estado de Michoacán. Posteriormente, Ponce y Beutelspacher (2001)
mencionan la presencia de la especie en varias localidades del estado incluyendo la parte más
baja de la Depresión del Balsas y finalmente Ponce (2003) menciona nuevas localidades en los
municipios de Apatzingán, Arteaga, Carácuaro, J. Mújica y Nocupétaro.
En el presente trabajo se actualiza la información sobre los registros de Vaejovis
variegatus y sus formas cercanas las cuales hemos etiquetado como Vaejovis aff. variegatus
conformando un área de distribución muy amplia en el estado de Michoacán (Figura 1), con
características ecológicas variadas, corroborando la suposición de que se trata de un complejo de
especies (com. per. W. David Sissom, 2009)1.
Materiales y Método
Se revisaron 108 ejemplares procedentes de 35 localidades (Cuadro 1) ubicadas en el
estado Michoacán, depositados en la Colección Aracnológica del Laboratorio de Entomología
“Biol. Sócrates Cisneros Paz” de la Facultad de Biología de la Universidad Michoacana de San
Nicolás de Hidalgo. Se identificó el material determinando si correspondía o no a la descripción
de la especie típica, procediendo a etiquetarlos y separarlos en dos grupos, los “típicos” y los que
no correspondieron exactamente con la descripción, los cuales fueron examinados con ayuda del
Dr. W. David Sissom de I&M Texas University.
Resultados y Discusión
Se determinó la existencia de tres taxones además de la especie típica, dos que
representan especies nuevas y que ya están en descripción y una que se identificó como Vaejovis
aff. variegatus y que requiere de mayor estudio para determinar su correcta identificación. De las
35 poblaciones, 18 se identificaron como V. aff. variegatus, 13 como la forma típica (V.
variegatus), 4 para Vaejovis sp. nov. 1 y 1 población para la otra especie nueva (Vaejovis sp. nov.
2), la cual corresponde a una localidad del municipio de Tancítaro. En la localidad de Tacámbaro,
se registraron dos especies: la “típica” y la especie nueva 1.
Las poblaciones determinadas como Vaejovis aff. variegatus son alacranes que
corresponden a la descripción de esta especie pero de coloración clara, la cual no se había
registrado anteriormente. Las poblaciones de Vaejovis variegatus y V. aff variegatus muestran
distribución geográfica distinta, ya que la mayoría de las poblaciones de Vaejovis aff. variegatus
han sido encontrados en vegetación de selva baja caducifolia en altitudes menores a los 1000
msnm con excepción de dos registros, uno en el área de Tzitzio y otro en Nocupétaro; mientras
que la especie típica se encuentra en selva baja y también en zonas con bosque de pino-encino en
altitudes mayores a los 1000 msnm.
La especie nueva 1, tiene poblaciones en zonas con vegetación de bosque de pino y pino-
encino, ocupando las zonas más altas de la distribución hasta ahora reconocida para este
complejo de poblaciones morfológicamente similares; mientras que la segunda especie nueva,
hasta ahora sólo se conoce de Tancítaro, a una altitud superior a los 2000 msnm (Fig. 1).
1 W. David Sissom. Reconocido investigador especialista en la familia Vaejovidae. Universidad Texas I&M en
Canyon, Texas

63
Cuadro 1. Localidades registradas actualmente para Vaejovis variegatus y Vaejovis aff. variegatus
en Michoacán.
LOCALIDAD (ES)* ALTITUD COORDENADAS
(msnm) Longitud Latitud Vaejovis aff. Variegatus El Ahuijote, Mpio. Churumuco (L1) 390 18° 42' 26'' 101° 45' 58''
Altamira (El Tesorero), Apatzingán
(L2) 420 18° 58' 04'' 102° 23' 11''
Camino a Baztán, El Rodeo, Huetamo
(L3) 767 18° 38' 15" 101° 09' 04"
Carácuaro (L4) 540 18° 59' 49" 101° 01 '47"
Cerro de Turitzio, cerca de Arúa,
Huetamo (L5) 480 18° 31' 55" 100° 55' 53"
Churumuco (L6) 240 18° 39' 50" 101° 37' 59"
Km 128 Carretera Morelia-L.C, J.
Mujica (L7) 650 18° 59' 48" 102° 14' 04"
Km 153 Carretera Huacana-4 Caminos
(L8) 365 18° 53' 36" 102° 01' 05"
Km 6 carretera Carácuaro-Nocupétaro
(L9) 770 18° 58' 00” 101° 06' 08"
Las Cañas, Arteaga (L10) 370 18° 33' 05" 101° 58' 09"
Paso de Núñez, Carácuaro (L11) 690 18° 53' 02" 100° 56' 44"
3km de Tzitzio (L12) 1540 19° 33' 53" 101° 55' 36"
3.8 Km Carretera Puruarán-Turicato
(L13) 1000 19° 05' 11" 101° 30' 02"
ECA, Apatzingán (L14) 300 19° 05' 01" 102° 21' 03"
El Chauz, Mpio. de la Huacana (L15) 225 18°53' 28" 102° 02' 49"
San José de la Peña, Mpio. de La
Huacana (L16) 378 19° 02' 02" 102° 02' 18"
Tres Puentes, Mpio. de Tzitzio (L17) 1700 19° 29' 20" 100° 53' 52"
Tzitzio (L18) 19° 35' 08" 100° 55' 29"
Vaejovis sp. Nov. 1 Santa Clara del Cobre (L19) 2186 19° 24' 02" 101° 38' 28"
Tacámbaro (L20) 1640 19° 13' 59" 101° 27' 38"
Nahuatzen (L21) 2421 19° 39' 36" 101° 54' 23"
Carapan (L22) 1967 19° 52' 29" 102° 02' 06"
Vaejovis sp. nov 2.*** El Aguacate, Mpio. de Tancítaro (L23) 2150 19° 23' 03" 102° 13' 07"
Vaejovis variegatus
(Típico)
Arroyo Frío, Pedernales, Mpio de
Tacámbaro (L24) 1240 19° 10' 10" 101° 28' 07"
Bosque Lázaro Cárdenas, Morelia
(L25) 1919 19° 40' 49" 101° 10' 40"
Chorros del Varal, Mpio. de los Reyes
(L26) 1092 19° 30' 53" 102° 34' 14"
El Carrizal, Mpio. de Carácuaro (L27) 607 19° 00' 32" 101° 06' 31"
El Carrizal, Tarímbaro (L28) 1943 19° 49' 58" 101° 09' 49"
Filtros viejos, Morelia (L29) 1961 19° 40' 47" 101° 09' 31"
Grutas de Ziranda, Cd. Hidalgo (L30) 1836 19° 38' 20" 100° 29' 54"
Km 4 de San Antonio de las Huertas,
Nocupétaro (L31) 1040 19° 04' 59" 101° 17' 15"
La Caratacua, Mpio. de Zacapu (L32) 1836 19° 44' 12" 101° 39' 24"
La Magdalena, Tarímbaro (L33) 1880 19° 48' 36” 101° 08' 42"
Mesón Nuevo, Tarímbaro (L34) 1927 19° 49' 21" 101° 10' 03"
Morelia (L35) 1912 19° 42' 10" 101° 11' 09"
Tacámbaro (L20) 1640 19° 13' 59" 101° 27' 38"
* La clave entre paréntesis, corresponde a la etiqueta en el mapa de la figura 1
** Especie ya descrita y el artículo en proceso de revisión
*** Especie en proceso de descripción
El grupo de poblaciones que hasta ahora se han determinado como Vaejovis variegatus
actualmente se encuentra en revisión ya que probablemente se trate de un complejo de especies
del que actualmente solo está descrita la especie típica y las dos que están en proceso, pero es

64
probable que la que estamos presentando en este trabajo como Vaejovis aff. variegatus, también
corresponda a un nuevo taxón específico.
Figura 1. Ubicación de los diferentes taxones del complejo V. variegatus en Michoacán
Conclusiones
En Michoacán, al menos hay dos taxones hasta ahora determinados como Vaejovis
variegatus, representados por varias poblaciones con diferencias de apariencia y distribución
geográfica, además de dos especies nuevas morfológicamente distinguibles de la especie típica:
una en la región de bosque de pino en la Meseta Tarasca, la cual ya está descrita y en proceso de
publicación y otra en el área del Tancítaro y que está en el proceso de descripción (Com. per. W.
David Sissom)1.
Literatura Citada
Beutelspacher, B. C. R., 2000. Catálogo de los alacranes de México. Universidad Michoacana de
San Nicolás de Hidalgo, Morelia, Michoacán, México., 175 pp.
Coddington, J.A., G. Giribet, M.S. Harvey, L. Prendini y D.E. Walter. 2004. Arachnida. pp. 296–
318 In: Cracraft, J. and Donoghue, M. (Eds.) Assembling the Tree of Life. Oxford
University Press, Oxford. Inglaterra.
Fet, V., W. D. Sissom, G. Lowe y M. E. Braunwalder. 2000. Catalog of the Scorpions of the
World (1758-1998). New York Entomological Society. 690 pp.
Hoffmann, C. C. 1931. Monografías para la entomología médica de México. Monografía Num. 2,
Los escorpiones de México. Primera parte: Diplocentridae, Chactidae, Vejovidae. Anales
del Instituto de Biología Universidad Nacional Autónoma de México, 2(4): 291 – 408

65
Lourenço, W. y D. W. Sissom. 2000. Scorpiones. En: Llorenre-Bousquets, J.E., A. N. García A.
y E. González S. (Eds.) 2000, Biodiversidad, Taxonomía y Biogeografía de artrópodos de
México: Hacia una síntesis de su conocimiento.Vol. II. CONABIO, UNAM y BAYER.
115-136.
Ponce, S. J y C. R. Beutelspacher B. 2001. Alacranes de Michoacán UMSNH. Ediciones
Michoacanas. Morelia, Michoacán, México. 103 pp
Ponce, S. J., 2003. Ecología y Distribución del Género Centruroides Marx 1890 (Scorpiones:
Buthidae), en la Depresión del Balsas del Estado de Michoacán. Tesis de Doctorado en
Ciencias. Universidad Autónoma de Querétaro, Facultad de Ciencias Naturales. Santiago
de Querétaro., 276 pp.
Prendini, L. y W. Wheeler. 2005. Scorpion higher phylogeny and classification, taxonomic
anarchy, and standards for peer review in online publishing. Cladistics, 21: 446–494 Rein J. O. 2009, http://www.ub.ntnu.no/scorpion-files/scorpionidae.php [Actualización 11.03.09]
Sissom, W. D. 2000. Family Vaejovidae, Thorell, 1876. pp. 503-553. En: Fet V., W. D. Sissom,
G. Lowe & M. E. Braunwalder (Eds.). Catalog of the Scorpions of the World (1758-
1998). New York Entomological Society, New York, E.U.A.,690 pp.
Sissom, W. D. y O. F. FRANCKE. 1985. Redescriptions of some poorly known species of the
nitidulus group of the genus Vaejovis (Scorpiones, Vaejovidae). Journal of Arachnology,
13(2): 243-266.
Soleglad, M.E. y V. Fet. 2005. A new scorpion genus (Scorpiones: Vaejovidae) from Mexico.
Euscorpius, 24: 1–13.
Soleglad, M.E. y V. Fet. 2008. Contributions to scorpion systematics. III. Subfamilies
Smeringurinae and Syntropinae (Scorpiones: Vaejovidae). Euscorpius, 71: 1–115.

66
SOLÍFUGOS DEPOSITADOS EN LA COLECCIÓN DE ARÁCNIDOS DE LA UNICACH,
CHIAPAS, MÉXICO (ARACHNIDA: SOLIFUGAE).
Solifugids deposited in the arachnids collection of UNICACH, Chiapas, México (Arachnida:
Solifugae).
Kaleb Zárate-Gálvez1 y Rafael Gaviño-Rojas
2.
1Colección de Arácnidos, Facultad de Ciencias
Biológicas, Universidad de Ciencias y Artes de Chiapas. Libramiento norte poniente s/n, colonia
Lajas Maciel, C.P. 29039. Tuxtla Gutiérrez, Chiapas, México. 2Laboratorio de Acarología “Anita
Hoffmann”, Facultad de Ciencias, Universidad Nacional Autónoma de México. Coyoacán 04510,
D. F. México. [email protected]
Palabras Clave: Solifugae, Chiapas, México.
Introducción
Los solífugos son comúnmente llamados como arañas camello “camel-spiders”, arañas
del viento “wind-spiders” y arañas del sol “sun-spiders”, referidos a diversos aspectos de su
morfología o biología. Este grupo constituye uno de los “ordenes menores” de los arácnidos, es
considerado como meso-diverso ya que mundialmente está representado por poco más de 1100
especies repartidas en 12 familias, cuatro de las cuales se encuentran en el continente americano
en donde se distribuyen desde los EUA hasta Argentina, incluyendo algunas islas antillanas
(Armas, 2004; Harvey, 2002; 2003).
Morfológicamente se distinguen de los demás arácnidos por la presencia de maleolos
(estructuras en forma de raqueta) localizados en la parte ventral de los segmentos basales de las
patas IV, se desconoce la función exacta de estas estructuras aunque se cree que podría ser
principalmente sensorial. Además, poseen discos adhesivos en la parte distal de los pedipalpos, lo
que les sirve para escalar superficies verticales y, un par de quelíceros que resultan de gran
tamaño en comparación con su cuerpo, los cuales en la gran mayoría de las especies carecen de
glándulas venenosas. Los miembros adultos presentan tamaños que varían de 1 a 10 cm
dependiendo de la especie (Harvey, 2003; Muma, 1951).
La velocidad, agresividad y voracidad también resultan características de este grupo. Se
alimentan principalmente de invertebrados y lagartijas, aunque las especies de mayor tamaño
también pueden llegar a matar algunas aves y pequeños mamíferos. Al igual que otros arácnidos,
el canibalismo ocurre en este orden (Muma, 1951).
La mayoría de las especies son de hábitos nocturnos, aunque algunos miembros de este
grupo presentan conductas diurnas, lo que resulta poco común entre los arácnidos a excepción del
orden Araneae. Habitan en las regiones tropicales y subtropicales del mundo, siendo más diversos
y mayormente restringidos hacia los climas calientes y áridos de estas regiones. A diferencia de
otros ordenes de arácnidos, no se han encontrado en el ambiente cavernícola (Harvey, 2003;
Muma, 1951).
En México se tienen registradas alrededor de 66 especies, incluidas en 11 géneros y 2
familias (Vázquez, 2004). De forma general, Eremobatidae presenta una distribución Neártica,
ocupando la parte norte y centro del país; mientras que Ammotrechidae se encuentra mayormente
distribuido en el sureste mexicano, aunque también existen representantes de esta familia en la

67
península de Baja California, Jalisco, Sinaloa y Sonora. El conocimiento que se tiene en el país
de este orden es desproporcionado ya que la mayoría de los datos provienen de los lugares mas
trabajados, referidos a la parte norte y centro del país (Vázquez-Rojas, 1995; Ballesteros &
Francke, 2007), en tanto que aún se conoce poco del sureste del país; aunado a esto, son pocos
los investigadores en este país que se dedican a su estudio. El objetivo de este trabajo fue
determinar los ejemplares depositados en la Colección de Arácnidos de la UNICACH.
Materiales y Método
Los ejemplares estudiados se encuentran depositados en la Colección de Arácnidos,
Facultad de Ciencias Biológicas de la Universidad de Ciencias y Artes de Chiapas, Tuxtla
Gutiérrez, Chiapas, México. Estos ejemplares son representativos de varias partes del estado de
Chiapas, localizado en el sureste de México y colindante con los estados de Oaxaca, Tabasco y
Veracruz, además de ser fronterizo con la republica de Guatemala. De igual manera se revisó y
tomaron en cuenta los registros de Muma (1986), Brookhart y Brookhart (2006) y Vázquez
(2004). La taxonomía está de acuerdo con Harvey (2003) y Muma (1970).
Resultados y Discusión
Se revisaron un total de 34 ejemplares pertenecientes a una sola familia (Ammotrechidae),
dos géneros y tres especies confirmadas, además de 2 géneros (Ammotrechesta y Branchia) y sus
especies por corroborar; en la representatividad de los sexos se encontró lo siguiente: 19♀, 13♂,
un juvenil y un ejemplar indeterminado (Cuadro 1).
Cuadro 1. Solífugos depositados en la Colección de Arácnidos de la UNICACH.
Especie Núm.
Ejemp Localidades Sexo
Ammotrecha stolli 2 Cerro Mactumatzá, Tuxtla Gutiérrez ♀
Ammotrecha stolli 2 Tuxtla Gutiérrez 1♂, 1♀
Ammotrecha stolli 1 Rivera Santa Cruz, Pantepec ♀
Ammotrecha stolli 1 Tiltpec, Jiquipilas ♂
Ammotrecha stolli 1 Parque ecológico “Laguna Verde”, Coapilla ♂
Ammotrecha limbata 1 Tuxtla Gutiérrez ¿?
Ammotrecha sp. 7 Tuxtla Gutiérrez; Pacú, Suchiapa; Pijijiapan 4♀, 3♂
Ammotrechella
stimpsoni 2
Colonia Bienestar Social, ciudad de Tuxtla
Gutiérrez ♂
Ammotrechella
stimpsoni 1 Cerro Mactumatzá, Tuxtla Gutiérrez ♂
Ammotrechella
stimpsoni 1 Tuxtla Gutiérrez ♂
Ammotrechella sp. 6 Parque Nacional Cañón del Sumidero; Cerro
Mactumatzá, Tuxtla Gutiérrez. 2♂, 4♀
Ammotrechesta sp. 7 Tuxtla Gutiérrez 5♀, 1♂ y
1 juvenil
Branchia angustus 1 Mirador “La Atalaya”, Cañón del Sumidero ♀
Branchia sp. 1 Tuxtla Gutiérrez ♀

68
Los ejemplares determinados solamente hasta nivel de género (Ammotrecha sp. y
Ammotrechella sp.) no concuerdan completamente con los caracteres específicos diagnósticos,
por lo que es necesario tener más material para verificar las distintas variaciones, como lo son: el
color y la espinación de patas y pedipalpos. Esto mismo sucede con los géneros Ammotrechesta y
Branchia, para los cuáles sería ideal verificar con el material tipo del género, además de verificar
con la descripción original. De ser confirmado la determinación de estos dos últimos géneros
representarían registros importantes. El género centroamericano Ammotrechesta sería registrado
por primera vez para México y el género Branchia ampliaría su distribución hacia el sureste del
país.
Excluyendo los géneros inciertos y los ejemplares indeterminados, la colección tiene
representada más del 50% de las especies registradas para el estado (Cuadro 2), siendo
Ammotrecha chiapasi y Ammotrechella bolivari las especies faltantes. Esto puede deberse a las
pocas colectas realizadas y a la rareza de estas especies. Debido a sus hábitos pocos ejemplares
de este orden de arácnidos están representados en la colecciones (Muma, 1951), lo que también
se ve reflejado en esta colección.
En concordancia con Brookhart y Brookhart (2006) los datos de la distribución presentan
un sesgo hacia las localidades más colectadas, como lo es el centro del estado. Futuras
prospecciones y colectas aracnológicas en otros sitios, con particular interés en este grupo,
ayudarán a clarificar las especies ya registradas o podrían revelar especies aún desconocidas para
la ciencia.
Cuadro 2. Solífugos registrados en la literatura para Chiapas, México.
Orden Solifugae
Familia Ammotrechidae Roewer, 1934
Subfamilia Ammotrechinae Roewer, 1934
Género Ammotrecha Banks, 1900
Ammotrecha stolli (Pocock), 1895
Ammotrecha limbata (Lucas), 1835
Ammotrecha chiapasi Muma, 1986
Género Ammotrechella
Ammotrechella stimpsoni Putnam, 1883
Ammotrechella bolivari Mello-Leitão, 1942
Agradecimientos
Agradecemos la ayuda en la revisión y determinación de muchos de los ejemplares al Dr.
Ignacio Vázquez de la Facultad de Ciencias, UNAM.
Literatura Citada
Armas, L. F. De 2004. Arácnidos de República Dominicana. I. Palpigradi, Schizomida,
Solifugae, Thelyphonida (Arthropoda: Arachnida). Revista Ibérica de Aracnología, Vol.
Especial Monográfico 2: 1-63.
Ballesteros, A. & O. F. Francke. 2007. A new species of sun-spider from sand dunes in Coahuila,
Mexico, (Arachnida: Solifugae: Eremobatidae). Zootaxa, 1665: 61-68.

69
Brookhart J.O. & I. P.Brookhart. 2006. An annotated checklist of continental North American
solifugae with type depositories, abundance, and notes on their zoogeography. The
Journal of Arachbologi, 34:299-329
Muma, M. H. 1951. The arachnid order Solpugida in the United States. Bulletin of the American
Museum of Natural History, 97(2): 35-141.
Muma, M. H. 1970. A synoptic review of North American, Central American, and West Indian
Solpugida (Arthropoda: Arachnida). Arthropods of Florida and Neighboring Land Areas,
5: 1-62.
Muma, M. H. 1986. New species and records of Solpugida (Arachnida) from Mexico, Central
America and the West Indies. Novitates Arthropodae 2(2): 1-23.
Harvey, M. S. 2002. The neglected cousins: what do we known about the smaller arachnids
orders?. The Journal of Arachnology, 30: 357-372.
Harvey, M. S. 2003. Catalogue of the smaller arachnid orders of the World: Amblypygi, Uropygi,
Schizomida, Palpigradi, Ricinulei and Solifugae. CSIRO Publishing. Collingwood
Victoria, Australia. i-xi + 385 pp.
Vázquez-Rojas, I. 1995. Los Arácnidos de México parte I: Ricinulei, Amblypygi, Solifugae,
Palpigradi, Schizomida, Uropygi. Dugesiana 2(1): 15-36.
Vázquez R., I. 1996. Solifugae. Pp. 75-78. En: Biodiversidad, taxonomía y biogeografía de
artrópodos de México: hacia una síntesis de su conocimiento. J. Llorente Bousquets, A. N.
García Aldrete y E. González Soriano (Eds.). 1ª edición. Universidad Nacional Autónoma
de México. 660 pp.
Vázquez, I. M. 2004. Los solífugos (Arachnida: Solifugae) mexicanos, datos actuales. Pp. 10-19.
En: 1er
Simposio Nacional de Aracnología. R. Rojo y F. Medina (compiladores). Sociedad
Mexicana de Entomología. Mazatlán, Sinaloa.

70
ÁCAROS MESOSTIGMADOS (ACARI: MESOSTIGMATA) DE LA SIERRA GORDA DE
QUERÉTARO, MÉXICO.
Mesostigmatid mites (Acari: Mesostigmata) from, Sierra Gorda, Querétaro, Mexico.
Lucia Georgina Pastrana-Ruiz. Ecología y Sistemática de Microartrópodos, Departamento de
Ecología y Recursos Naturales, Facultad de Ciencias, Universidad Nacional Autónoma de
México, México D. F. 04510. E- mail: [email protected]
Palabras Clave: Mesostigmados, suelo, hojarasca, México.
Introducción
Los mesostigmados constituyen un numeroso grupo de ácaros que se han adaptado
exitosamente a una gran variedad de hábitats. La mayoría son depredadores de vida libre, que se
encuentran en el suelo, en la hojarasca o sobre plantas donde se alimentan de invertebrados
pequeños. También existen las formas parásitas de vertebrados o invertebrados, las especies que
son fungívoras, detritívoras, polinívoras y algunas especies que han establecido relaciones
foréticas con otros animales (Krantz y Ainscough, 1990) Desempeñan un papel muy importante
dentro de la red alimentaria del suelo y son determinantes en los mecanismos de control de
poblaciones dentro del ecosistema edáfico (Koehler, 1999; Krebs, 1978). Se les asocia con la
recirculación de los nutrimentos del suelo, pues las diversas formas de alimentación que
presentan, ayudan a la rápida descomposición de la materia orgánica y a la diseminación de la
misma. Además, el desplazamiento de este tipo de organismos promueve la aeración y el cambio
físico de la estructura edáfica (Huhta et al. 1988, Evans 1992, Juma 1993, Koehler 1999,
Hoffmann y López-Campos 2002).
En el ambiente edáfico, los mesostigmados se establecen de acuerdo a los recursos que
ofrece el suelo, el cual puede definirse como un ecosistema compuesto por minerales, materia
orgánica y seres vivos, cuyas interacciones dan como resultado las propiedades químicas y físicas
que lo componen, así como, el establecimiento de las comunidades, tanto vegetales como
animales (Wolters 1991, Jordana 1996).
Físicamente, el suelo sufre procesos complejos de génesis y desarrollo continuamente, de
tal forma que es dinámico y muy heterogéneo en espacio y tiempo (Siebe et al. 1997). Debido a
ésto, se han hecho enormes esfuerzos para clasificar la superficie terrestre, con base en
características como profundidad, contenido de materia orgánica, color, textura, comportamiento
químico, pH, estructura, drenaje, densidad, concreciones, salinidad, etc. Sin embargo, a pesar de
que un suelo puede clasificarse dentro de uno de los grupos edafológicos principales, a escala
local presenta variaciones en sus propiedades, de tal forma que dentro de una misma unidad
edáfica existe cierto grado de heterogeneidad espacial (Aguilera 1989).
La heterogeneidad en las características edáficas, juega un papel muy importante en el
establecimiento y desarrollo de las comunidades de organismos, así como en sus funciones y
distribución específica (Dindal 1990, Castaño-Meneses 2002, Hodkinson 2005). Es por ello, que
la presente investigación se llevó a cabo en dos zonas que presentan un suelo con propiedades
particulares distintas; así mismo, se tomaron en cuenta diferentes épocas del año, para poder
registrar las variaciones e influencia de los factores ambientales en el establecimiento de los

71
ácaros. De esta manera, se obtuvieron patrones de distribución espacial y temporal que hacen
evidente la estructura de la comunidad de los Mesostigmata en diversas clases de suelos.
Materiales y Método
El área de estudio se ubica dentro de la Reserva de la Biosfera Sierra Gorda, donde se
estudiaron dos zonas con características bióticas y abióticas similares, siendo la única variable, la
diferencia en las propiedades físicas del suelo.
Ambas zonas se eligieron dentro de bosques mesófilos, con vegetación predominante de
pino, encino y liquidámbar, donde existe un clima cálido sub-húmedo. En todos los casos se
trabajo en laderas con pendientes de aproximadamente 45º, orientadas hacia el Este. El suelo de
acuerdo con sus propiedades generales se determinó como Leptosol (Semarnat 2002).
La localidad 1 se encuentra cerca del poblado conocido como Puerto San Agustín,
Municipio de Landa de Matamoros (21º 14’ 31.33’’ N y 99º 04’28.16’’O) a una altitud de 1592
msnm. Las propiedades características del suelo en la localidad se derivan de la textura
predominantemente arcillosa.
La localidad 2 corresponde al Municipio de Jalpan de Serra en las cercanías del poblado
La Yerbabuena (21º 08’ 00.87’’ N y 99º 27’ 29.05’’ O) a una altitud de 1506 msnm. Este sitio se
caracteriza por una textura limosa del suelo de la cual se derivan las propiedades particulares del
suelo.
Se realizaron muestreos trimestrales de Junio de 2007 a Junio de 2008, a fin de
monitorear la estructura de la comunidad de ácaros, para ésta primera etapa de análisis se
consideraron los muestreos de Junio, Septiembre y Diciembre de 2007. En cada localidad se
localizaron, en forma aleatoria, 10 sitios en los cuales se colectó una muestra de suelo y una de
hojarasca. Las muestras se tomaron mediante nucleadores de 15 x 15 x 5 cm, directamente en la
superficie del suelo y se transportaron al laboratorio.
Para extraer la fauna, se utilizaron embudos de Berlese-Tullgren. Una vez extraídos los
organismos, se separaron los ácaros mesostigmados y se procedió al conteo y determinación de
especies mediante el montaje del material en preparaciones semipermanentes con líquido de
Hoyer.
Con la contabilización de ácaros mesostigmados se obtuvieron los datos de abundancia y
riqueza de especies con los que se analizó la variación espacial y temporal en las comunidades de
éste grupo de ácaros.
Resultados
La revisión del material colectado proporcionó un inventario inédito de la artropodofauna
en la Reserva de la Biósfera Sierra Gorda, en el estado de Querétaro, dicho inventario incluye 13
clases y 24 ordenes de artrópodos edáficos durante la primera etapa de revisión de material
(Cuadro 1).
Se encontró que en todas las colectas, los mesostigmados conforman el tercer grupo más
abundante y el primero en cuanto a depredadores. Este patrón se siguió en ambas zonas de
estudio y en ambos biotopos a lo largo de todas las colectas.
En total, el Orden Mesostigmata presentó 5 familias y 12 morfoespecies, distribuidas tal
como se presenta en el cuadro 2.

72
Cuadro 1. Inventario de la artropodofauna encontrada en las localidades de San Agustín y La Yerbabuena, en el
estado de Querétaro, México.
Clase Orden
Pseudoscorpiones
Palpigradi
Araneae
Opiliones
Acari Astigmata
Prostigmata
Mesostigmata
Cryptostigmata
Pauropoda
Diplopoda
Chilopod
Symphyla
Collembola Poduromorpha
Entomobryomorpha
Symphypleona
Protura
Diplura
Insecta Tysanoptera
Hemiptera
Homoptera
Coleoptera
Diptera
Trichoptera
Hymenoptera
Cuadro 2. Distribución de las familias y morfoespecies de mesostigmados, en las localidades y biotopos de estudio. Familia
Morfoespecie San Agustin Yerbabuena
Suelo Hojarasca Suelo Hojarasca
Ascidae Sp. A1 X X X X
Sp. A2 X X X X
Sp. A3 X X
Uropodidae Sp. U1 X X X
Polyaspis sp. X X X
Polyaspis (Dipolyaspis) sp. X X
Sp. U4 X X
Sp. U5 X
Podocinidae Sp. P1 X X X
Laelapidae Sp. L1 X X X X
Sp. L2 X X
Macrochelidae Sp. M1 X X
Se presentó una marcada variación en la abundancia entre las diferentes zonas de estudio.
La localidad de Puerto San Agustín, tuvo la mayor abundancia con 5021 mesostigmados,
mientras que la localidad de La Yerbabuena solamente presentó 1440 individuos (Cuadro 3).
Las observaciones por biotopo, indicaron que en San Agustín, existe mayor abundancia en
la hojarasca, mientras que en la localidad de La Yerbabuena la mayor abundancia se encuentra en
el suelo (Cuadro 3).

73
Cuadro 3. Abundancia total y relativa de ácaros mesostigmados en las dos localidades bajo estudio.
Biotopo San Agustín La Yerbabuena
Abundancia A. Relativa Abundancia A. Relativa
Suelo 1671 33.28 1378 95.69
Hojarasca 3350 66.72 62 4.31
Total 5021 100 1440 100
Discusión y Conclusiones
El estado de Querétaro es muy complejo en cuanto a su fisiografía y sus climas, lo cual
permite la existencia de múltiples ecosistemas en los que se encuentran gran diversidad de
especies (Rzedowski y de Rzedowski 1991). Es por ello que muchos investigadores han centrado
sus esfuerzos en esta área, sobre todo enfocados hacia la caracterización de la flora, sin embargo,
en cuanto a fauna se refiere, no se han dado grandes avances; en el caso de los artrópodos ni
siquiera existe un censo general del grupo, es por ello que el inventario generado a partir de ésta
investigación constituye un importante avance en cuanto a la descripción de ésta área y en cuanto
al conocimiento de los microartrópodos en el país.
En cuanto a los ácaros mesostigmados, los resultados indican que la proporción de
individuos sobrepasa en gran medida la abundancia de otros grupos de artrópodos (insectos,
miriápodos, arácnidos y ácaros prostigmados y astigmados), por lo que se puede concluir que se
trata de un grupo dominante dentro del ecosistema, tal como lo afirman Krebs (1978) y Koehler
(1999), pues tienen gran influencia sobre la dinámica ecológica, ya que la mayoría presenta
hábitos depredadores que limitan el crecimiento en las poblaciones de sus presas.
La mayor diversidad de ácaros mesostigmados se dio en la localidad de Puerto San
Agustín, donde encontramos características edáficas derivadas de un tipo de suelo arcilloso. Esta
alta diversidad se explica por que, propiedades como el tamaño de poro y alto nivel de retención
de agua permiten que exista una alta disponibilidad de recursos tales como el espacio y los
nutrientes biológicamente útiles (Aguilera 1989, Siebe et al. 1997).
Por el contrario, en la localidad de La Yerbabuena, el suelo presenta características
particulares provenientes de un suelo con alto nivel de arcillas. En este caso las partículas
edáficas son más pequeñas y por lo tanto los espacios intersticiales también, este tipo de
características limita en gran medida la retención de agua e incrementa la intemperización del
suelo. De manera que la disponibilidad de espacio para los organismos es menor, así como la
abundancia de nutrientes útiles y por ello la diversidad de mesostigmados se ve reducida
(Aguilera 1989, Siebe et al. 1997).
Es importante señalar que además de la influencia directa de las propiedades del suelo
sobre la variación en las comunidades de mesostigmados, existe influencia indirecta dada por la
variación de la vegetación secundaria que puede adaptarse a dichas características edáficas. De
esta variación se derivan diferencias en la abundancia y composición de la hojarasca de cada
localidad, la cual se traduce en diferencias de disponibilidad de recursos de cada zona.
Agradecimientos
A la Dra. Gabriela Castaño Meneses por dirigir este trabajo, en coordinación con los Dres.
Oscar Francke (Instituto de Biología, UNAM) y Pavel Krassilnikov (Facultad de Ciencias,
UNAM). Al proyecto PAPIT-IN 104807 “El efecto de la diversidad de suelos sobre la
biodiversidad de los ecosistemas forestales de México” mediante el cual se obtuvieron los

74
recursos económicos de la investigación. A la Dra Magdalena Vázquez González (Universidad de
Quintana Roo) por la identificación de los ejemplares de Uropodina.
Literatura Citada
Aguilera, H. 1989. Tratado de edafología de México. Tomo I. Facultad de Ciencias, UNAM.
México, 222 pp.
Castaño-Meneses, R. G. 2002. Estructura de la comunidad de artrópodos epífitos y su papel en el
crecimiento de Tillandsia violacea (Bromeliacea) en un bosque templado de Hidalgo,
México. Tesis Doctorado. Universidad Nacional Autónoma de México, México, 143 pp.
Dindal, D. L. (Ed.) 1990. Soil Biology Guide. Wiley Interscience Publication, New York, 1349
pp.
Evans, G. O. 1992. Principles of Acarology. CAB Internacional, Wallingford, UK, 565 pp.
Hodkinson, I. D. 2005. Terrestrial insects along elevation gradients: species and community
responses to altitude. Biol. Rev. 80: 489-513.
Hoffmann, A. y M. G. López-Campos. 2002. Acari. pp. 223-276. En: (J. Llorente & J. J.
Morrone, ed.) Biodiversidad, taxonomía y biogeografía de artrópodos de México: Hacia
una síntesis de su conocimiento. Universidad Nacional Autónoma de México, México.
Huhta, V., H. Setala y J. Haimi. 1988. Leaching of N and C from brich leaf litter and raw humus
with special emphasis on the influence of soil fauna. Soil Biol. Biochem. 20: 875-878.
Jordana, R. 1996. Ecología y aspectos funcionales de la biodiversidad en el suelo. II Congreso de
la Sociedad Española de Agricultura Ecológica. Pamplona-Iruña, España, 225-240.
Juma, N. G. 1993. Interrelationships between soil structure/textura, soil biota/soil organic matter
and crop production. Geoderma. 57:3-30.
Krantz, G. W. y B. D. Ainscough. 1990. Acarina: Mesostigmata (Gamasida). Pp 583-593. En:
D.L. Dindal (ed.) Soil Biology Guide.
Koehler, H. 1999. Predatory mites (Gamasina, Mesostigmata). Agric., Ecosyst. Environ. 74: 395-
410.
Krebs, C. J. 1978. Ecology, The experimental Análisis of Distribution and Abundante. Harper &
Row, USA. 694 pp.
Rzedowski, J. y G. C. de Rzedowski. 1991. Flora del Bajío y regiones adyacentes. Centro
Regional Bajío. Michoacán, Mexico. 134 pp.
Semarnat, 2002. Inventario Nacional de Suelos. Secretaría del Medio Ambiente y Recursos
Naturales, México.
Siebe, C., P. Palacios y A. Cortés. 1997. Variabilidad y distribución espaciales de algunos
parámetros físicos y químicos del suelo en el distrito de riego 03. Estado de Hidalgo. Rev.
Mex. Cienc. Geol. 11: 62 -67.
Wolters, V. 1991. Soil invertebrates-effects on nutrient turnover and soil structure, a review. Z.
Pflanzenernahr. Bodenk. 154:389-402.