maduracion de celulas_t
TRANSCRIPT
MADURACIÓN DE CÉLULAS T

INTRODUCCIÓN
Los linfocitos expresan receptores del antígeno muy variados, que reconocen una amplia variedad de sustancias extrañas.
Esta diversidad se origina a partir de células troncales hematopoyéticas que no expresan receptores de antígeno.
Durante el proceso de maduración se activan receptores específicos formados a partir de un programa de expresión genética secuencial que culmina en la reorganización de los genes de los mismos.

Los precursores de los linfocitos T salen del hígado fetal antes del nacimiento y de la medula ósea de forma posterior; circulan hacia el timo donde finalizan su maduración.
El desarrollo temprano de los linfocitos T al igual que los B se caracteriza por la proliferación de los progenitores, estimulado de forma inicial por la IL-7 producida por las células del estroma de la medula ósea y del timo.

Cada clon de linfocitos T produce un receptor del antígeno con una estructura de unión única.
La reorganización de los genes que codifican las cadenas de los receptores de los linfocitos T, suponen la adición o eliminación aleatorias de nucleótidos al azar.
La supervivencia y proliferación posteriores de los linfocitos en maduración dependen de las señales procedentes del receptor pre-T.
Posteriormente por el proceso de selección clonal se escogen a aquellas clonas que son capaces de reconocer las propias moléculas de MHC y al mismo tiempo son especificas de epítopos antigénicos (positiva/negativa)
Así mismo existen mecanismos que emparejan a los linfocitos T con las moléculas CD4+ que reconocen MHC II y CD8+ que reconocen MHC I.

RECEPTORES DE LINFOCITOS T El receptor de los linfocitos T que es
capaz de reconocer complejos péptido-MHC se conoce como RLT; el cual es de distribución clonal compuesto por : Unidades α Unidades β CD3 Proteinas ζ
Algunos linfocitos expresan cadenas γδ.
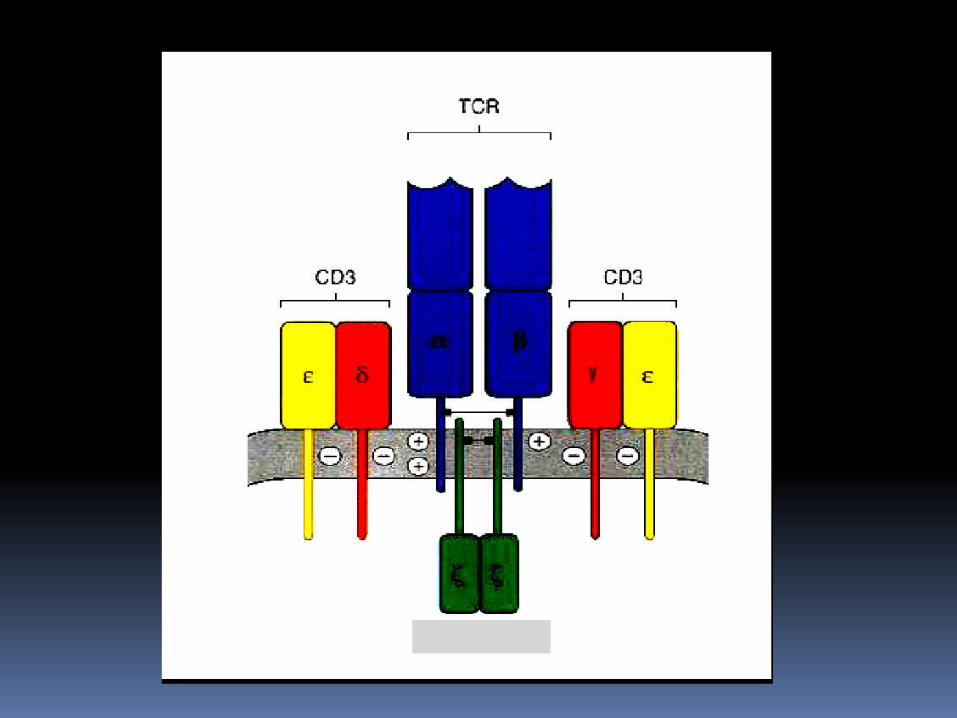
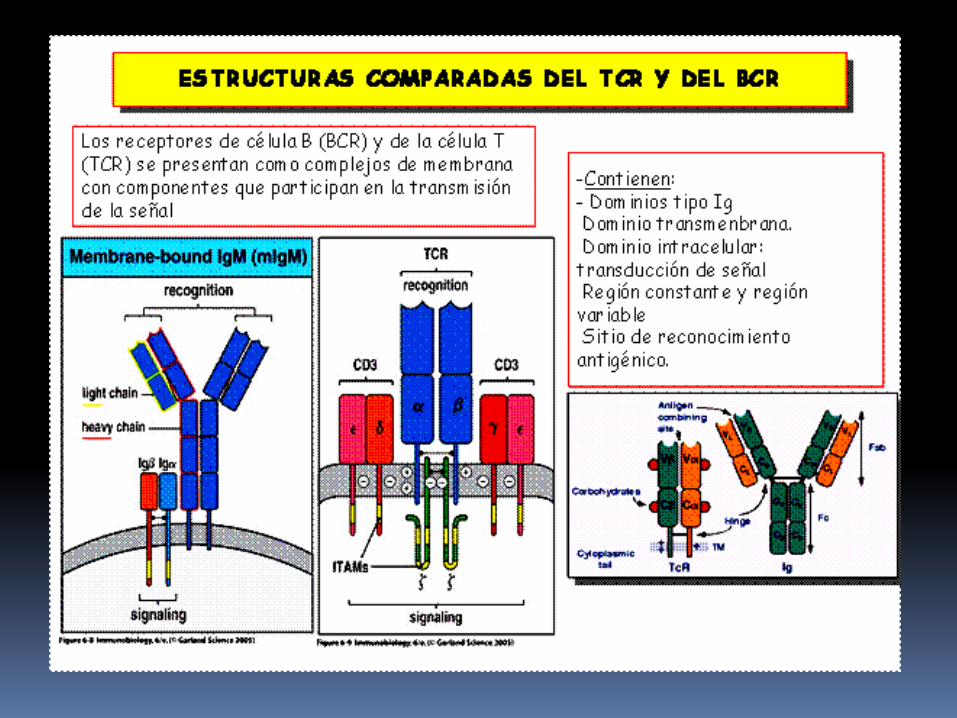
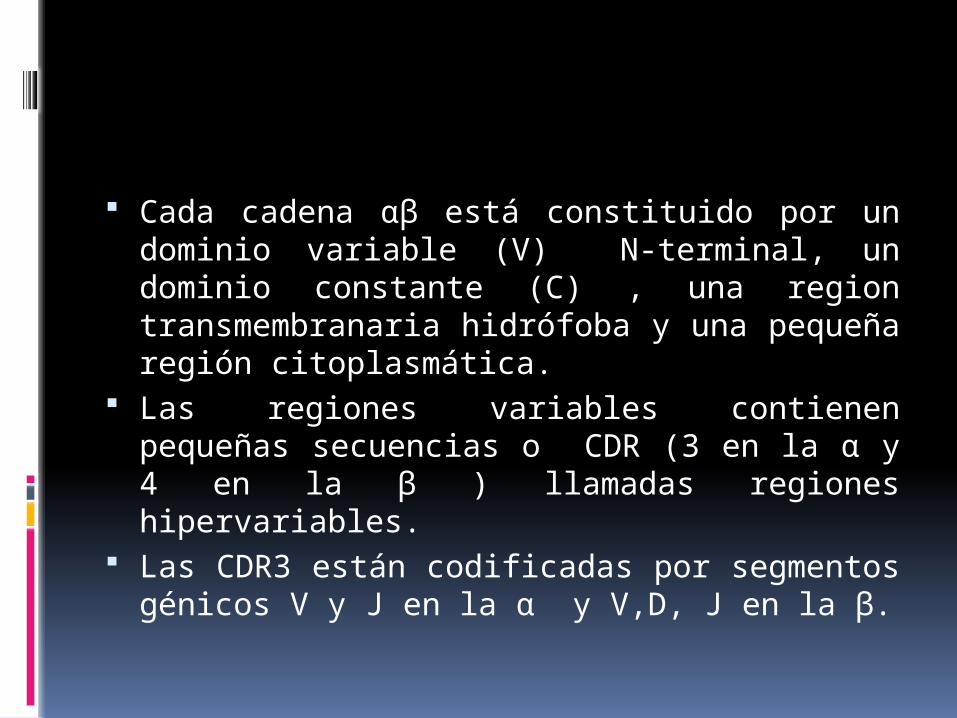
Cada cadena αβ está constituido por un dominio variable (V) N-terminal, un dominio constante (C) , una region transmembranaria hidrófoba y una pequeña región citoplasmática.
Las regiones variables contienen pequeñas secuencias o CDR (3 en la α y 4 en la β ) llamadas regiones hipervariables.
Las CDR3 están codificadas por segmentos génicos V y J en la α y V,D, J en la β.
Las regiones C de las cadenas α y β se continuan con pequeñas regiones bisagra, que consisten en cisteinas que forman un puente disulfuro que une a las cadenas.
La bisagra se sigue de porciones transmembranarias hidrófobas con aminoácidos de cargas positivas (lisina α y lisina/arginina en la β) los cuales interaccionan con los aminoácidos de cargas negativas de otros polipeptidos tranmembranarios (CD3 y ζ).
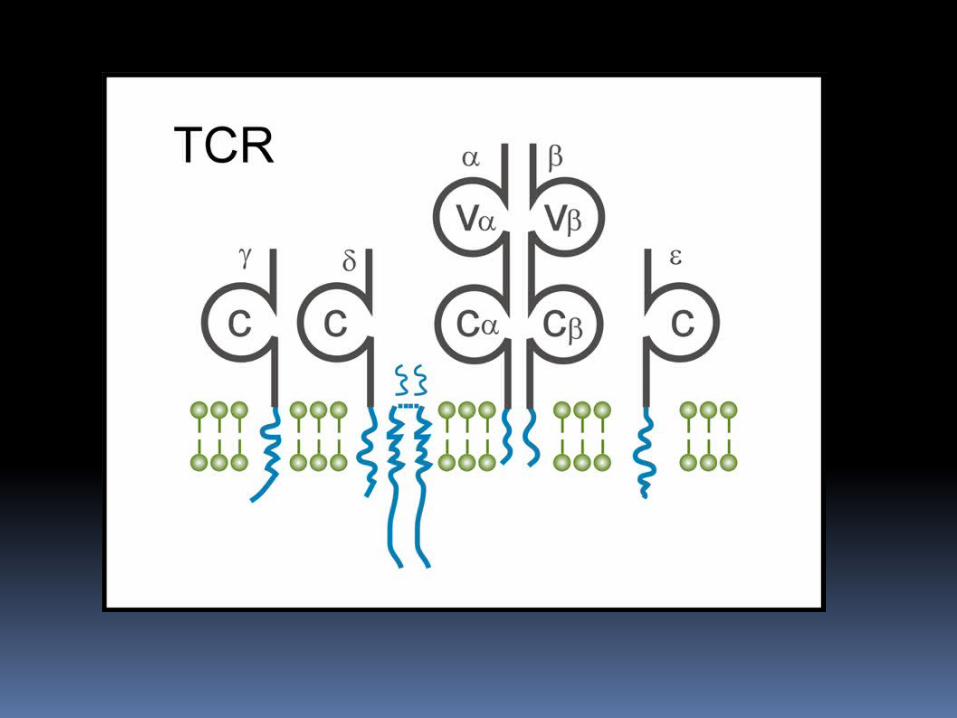

Tanto las cadenas α como las β tienen colas citoplasmaticas carboxiterminales de 5 a 12 aminoácidos de longitud.
El RLT contacta el complejo péptido MHC en una orientación diagonal ajustando los puntos superiores de las helices α del MHC; los bucles CDR1 de las cadenas α y β están colocados sobre los extremos del peptido unido, los bucles CDR2 sobre las helices del MHC y el bucle CDR3 sobre el centro del peptido asociado.
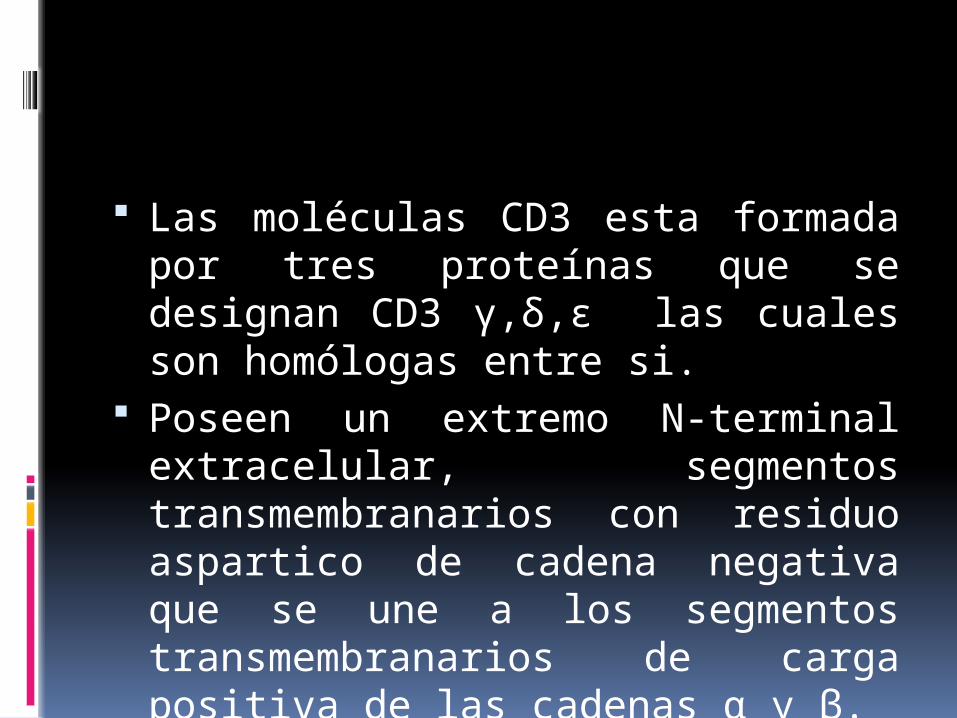
Las moléculas CD3 esta formada por tres proteínas que se designan CD3 γ,δ,ε las cuales son homólogas entre si.
Poseen un extremo N-terminal extracelular, segmentos transmembranarios con residuo aspartico de cadena negativa que se une a los segmentos transmembranarios de carga positiva de las cadenas α y β.
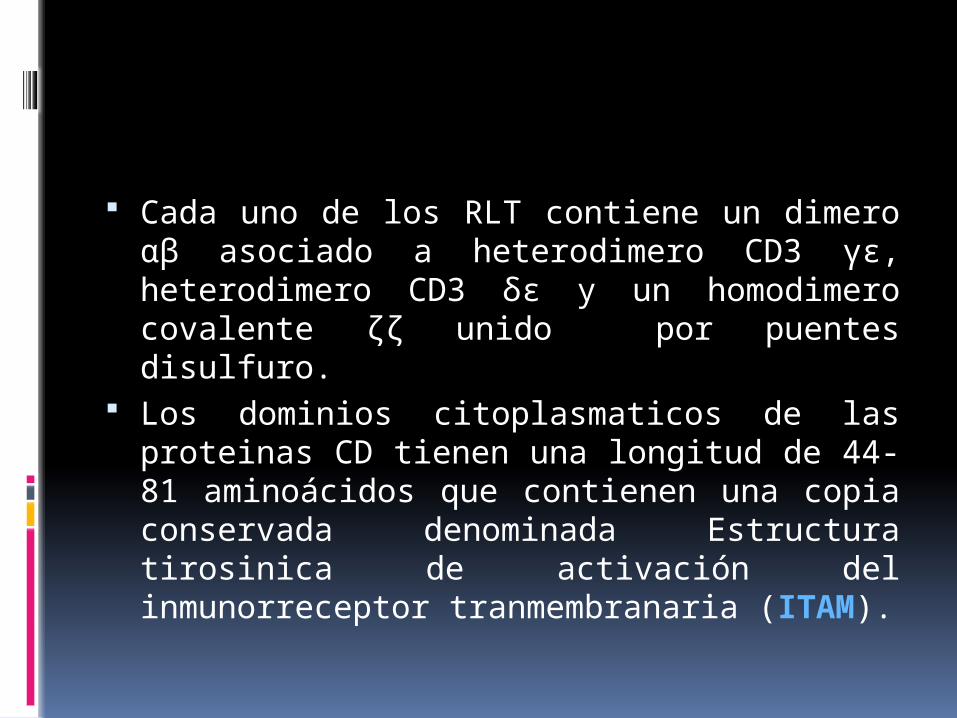
Cada uno de los RLT contiene un dimero αβ asociado a heterodimero CD3 γε, heterodimero CD3 δε y un homodimero covalente ζζ unido por puentes disulfuro.
Los dominios citoplasmaticos de las proteinas CD tienen una longitud de 44-81 aminoácidos que contienen una copia conservada denominada Estructura tirosinica de activación del inmunorreceptor tranmembranaria (ITAM).
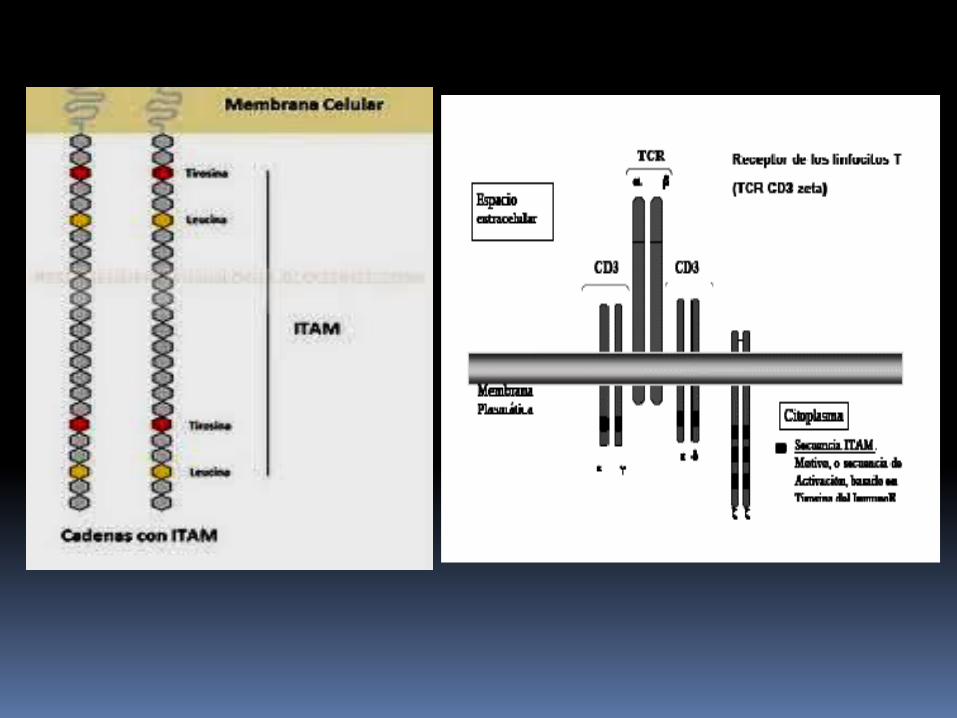

La cadena ζ tiene una región extracelular corta de nueve aminoácidos, una región tranmembranaria con un residuo de ácido aspartico con carga negativa y una región citoplasmatica larga de 113 aminoácidos que contiene ITAM; expresandose como homodimero.

Por otro lado el RLT γδ se expresa en un pequeño número de linfocitos T (5%), presentes en los tejidos epiteliales; los cuales están formados por regiones extracelulares V y C , regiones conectoras o bisagras, segmentos transmembranarios hidrófobos y colas citoplasmáticas cortas.
El heterodimero γδ se asocia a las proteínas CD3 y ζ de igual manera que las anteriores.

Los linfocitos Tγδ no reconocen antígenos peptídicos asociados a MHC ni estan restringidos por MHC ; reconociendo moléculas fosforiladas pequeñas, alquilaminas, lipidos y proteínas que no han sido procesadas por CPA ; poseen una diversidad limitada y pueden reconocer moléculas de MHC I.

ORGANIZACIÓN DE LOS LOCUS GENICOS RLT. Los genes que codifican la cadena αβ
y γ se localizan en tres locus separados y el de la δ esta contenido dentro del locus de la cadena α.
Cada uno de los locus contienen segmentos V, D, C, ademas los locus de la cadena β tiene segmentos D.

En el extremo 5’ de los locus de los RLT hay un grupo de varios segmentos V acompañados de un péptido guia y un promotor.
A distancias variables en dirrección 3’ de los genes V están los genes de la región C de los cuales hay dos en cada locus en las β y γ y solo uno de los α y δ.
Cada uno de los genes de la region C esta formado por cuatro exones que codifican la region C extracelular, una pequeña región bisagra, el segmento transmembranario y la cola citoplasmatica.
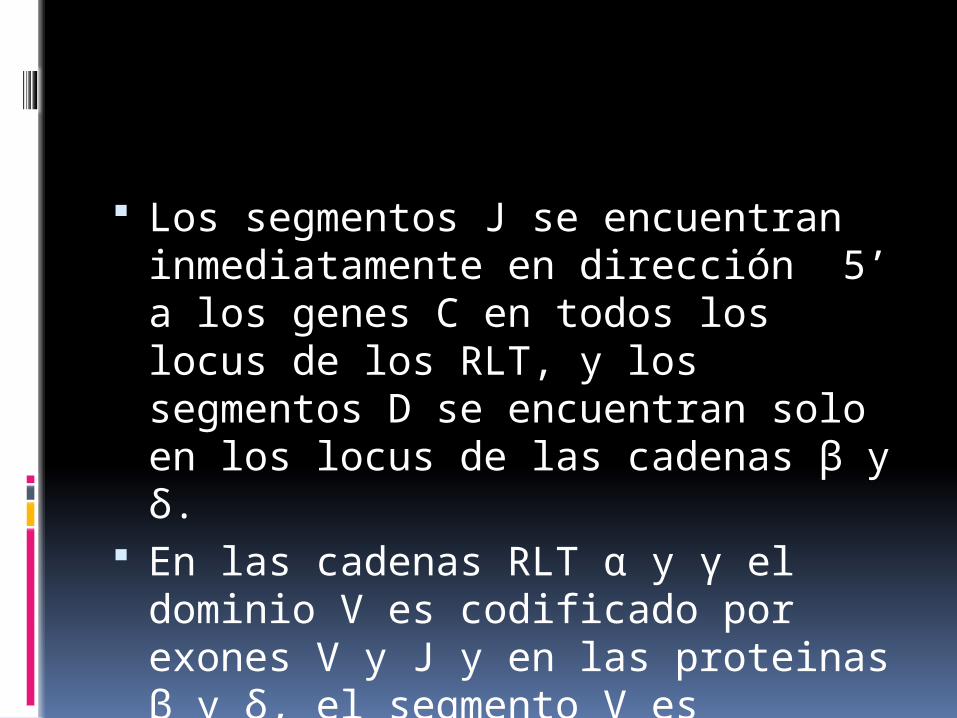
Los segmentos J se encuentran inmediatamente en dirección 5’ a los genes C en todos los locus de los RLT, y los segmentos D se encuentran solo en los locus de las cadenas β y δ.
En las cadenas RLT α y γ el dominio V es codificado por exones V y J y en las proteinas β y δ, el segmento V es codificado por segmentos V,D,J.

PROCESO DE RECOMBINACIÓN VDJ
Sinapsis- algunas porciones del cromosoma del receptor al antígeno son accesibles a la maquinaria de recombinación y dos segmentos codificadores son aproximados por la formación de un bucle.
Escisión- consiste en la generación de roturas enzimáticas en el ADN en las uniones heptámetros y secuencias codificadoras.
Codificación y procesamiento- los extremos codificadores rotos son modificados por adición o eliminación de bases generando mayor diversidad.
Unión- los extremos codificadores y los de señal son aproximados y ligados por las ligasas.
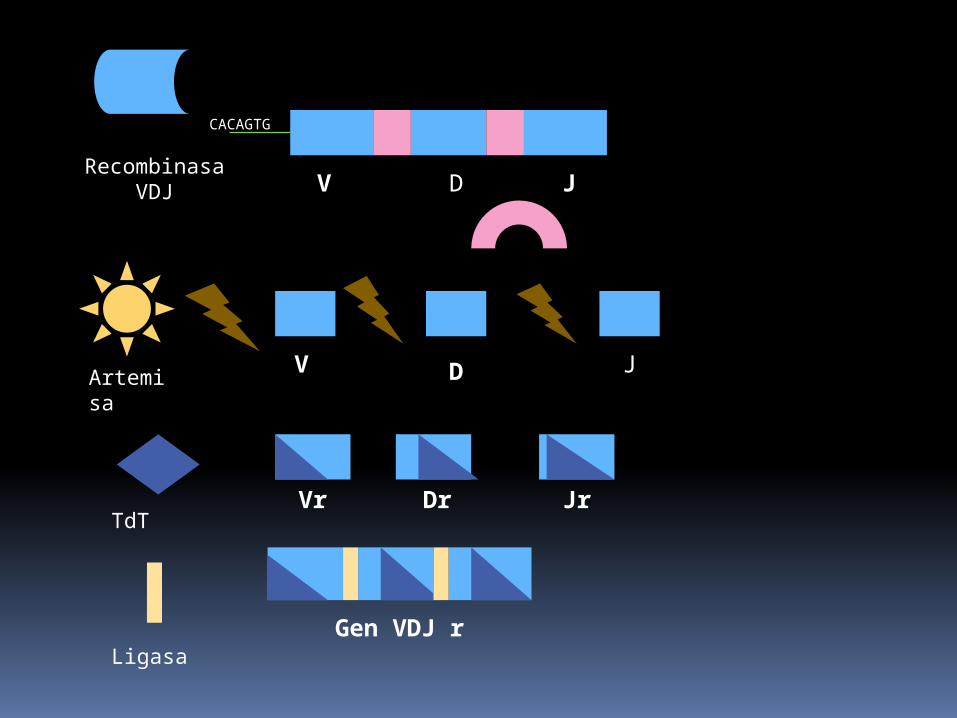
Recombinasa VDJ
CACAGTG
V D J
Artemisa
V D J
TdTVr Dr Jr
LigasaGen VDJ r
La recombinasa de VDJ formada por Rag 1 (reconoce la unión entre heptámero y segmentos codificadores) y Rag 2 (que une la anterior a factores de accesibilidad que llevan a locus específicos abiertos) formando horquillas.
Otras enzimas importantes son las Ku 70,Ku 80, que unen los extremos de ADN uniéndose a las roturas y reclutando a la proteína cinasa dependiente del ADN (ADN-PK) cuya función es reparar el ADN.
La ADN-PK, fosforila a la Artemisa que abre las horquillas en los extremos codificadores.
La adición de nuevos nucleótidos esta mediada por la desoxinucleotidil transferasa terminal(TdT).
finalmente se ligan los extremos rotos mediante la ADN ligasa IV.

MADURACION DE LINFOCITOS T La maduración de los linfocitos T
supone: La reorganización secuencial y la
expresión de los genes del RLT La proliferación celular La selección inducida por el antígeno La adquisición de capacidades
funcionales.

FUNCION DEL TIMO
El timo es el principal punto de maduración de los linfocitos T.
Los linfocitos Tm tienen una esperanza de vida de unos 20 años.
Los linfocitos T se expresan por primera vez en el timo a la semana 7-8 de la gestación.
Los timocitos inmaduros que no expresan receptores de superficie, se encuentran en el seno subcapsular y la región cortical externa del timo.

En la corteza los timocitos expresan por primera vez los RLT γδ ó αβ; con la posterior maduración en CD4 o CD8, migran de la corteza hasta la médula y despues salen del timo a la circulación.
El entorno tímico proporciona estimulos necesarios para la proliferación y maduración procedentes de células endoteliales, macrófagos y células dendríticas.
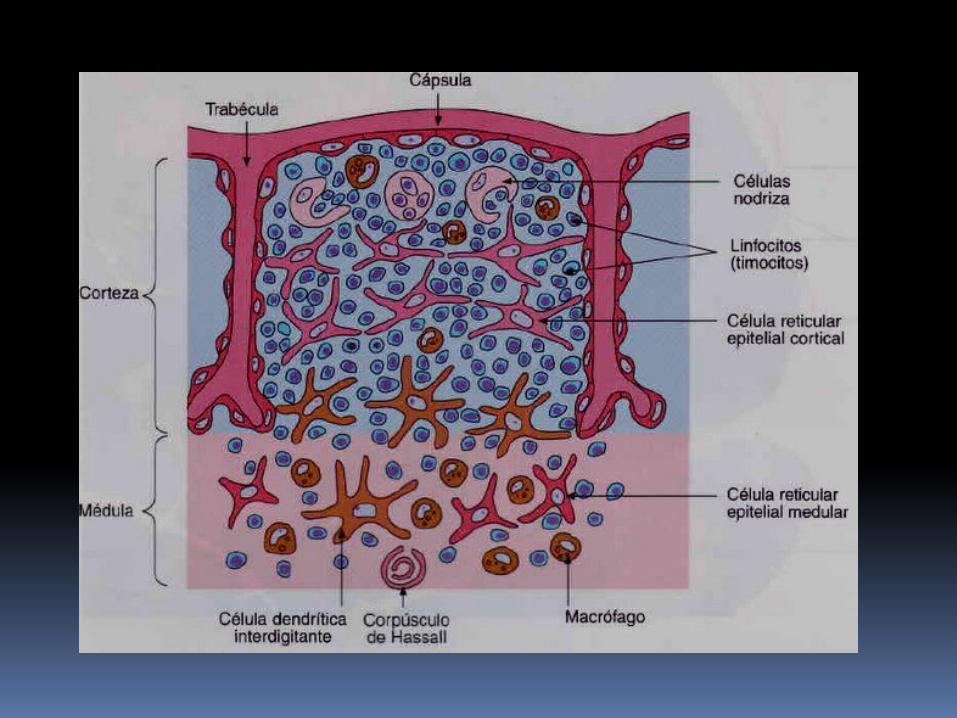
Dentro de la corteza, las células epiteliales forman una red de prolongaciones citoplasmáticas largas alrededor de las cuales deben pasar los timocitos.
Las células dendriticas localizadas en la unión corticomedular y dentro de la medula; finalmente, los macrófagos presentes mayormente en la medula.
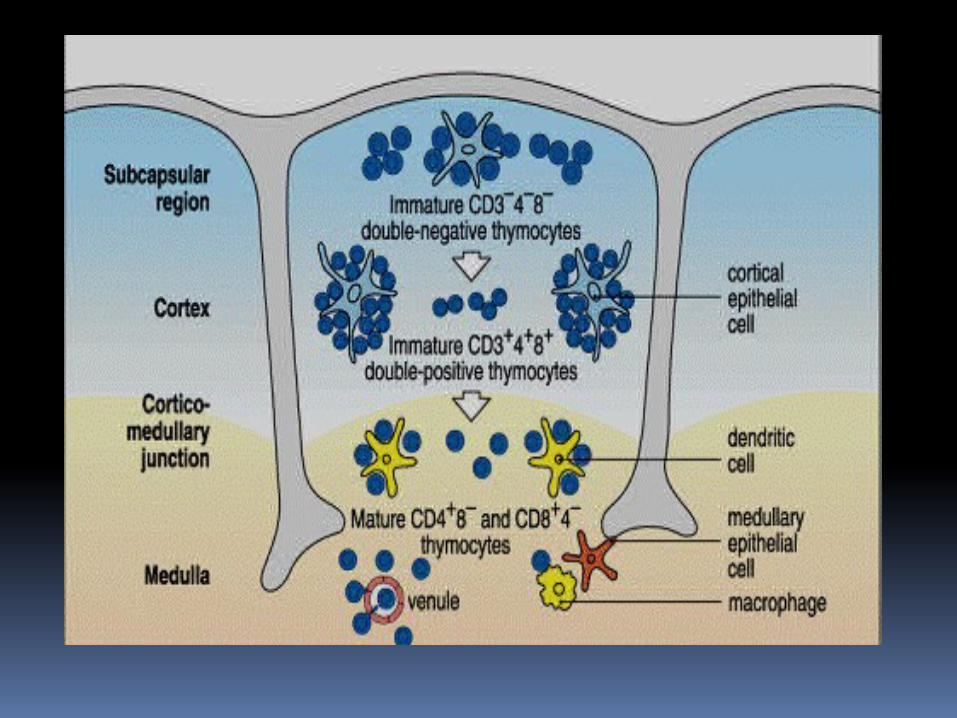

Dos tipos de moléculas son importantes para la maduración: Las moléculas de MHC clase I y II,
expresadas por células epiteliales y dendríticas.
Citocinas y quimiocinas de las células estromales (sobretodo IL-7 y las quimiocinas CCL21 y CCL19 que son reconocidas por el receptor CCR7); mediando el movimiento de los linfocitos T en desarrollo en el timo.

Un único precursor da lugar a muchas progenies de las cuales un 95% muere por apoptosis debido a : Imposibilidad de reorganizar de forma
productiva el gen de la cadena β del RLT. La imposibilidad de ser seleccionado
positivamente por la CPA. La selección negativa inducida por autoantigenos.

FASES DE LA MADURACION DEL LINFOCITO T
TIMOCITO PRO-T (negativos dobles) Son los timocitos corticales mas
inmaduros , no expresan RLT, CD3, o cadenas ζ, ni CD4,CD8.
La mayoria >90% dará en ultimo término a las células maduras CD4 o CD8 restringidas por CPA.
Las proteínas Rag 1 y Rag 2 se expresan por primera vez en esta fase, reorganizandose los genes de la cadena β.
Primero se reorganiza D a J y después con los segmentos V.

LINFOCITO PRE-T La reorganización de los genes V a D, J la caracterizan
formando los transcriptos nucleares primarios de las cadenas β del RLT y se adicionan las colas de poli-A.
Si se produce una reorganización productiva la proteina de la cadena β se expresará en la superficie celular asociada a una proteina invariable pre-Tα y las proteinas CD3 y ζ para formar el pre-RLT.
Su función es similar al Pre RLB y contribuyen a : La mediación de la supervivencia de los linfocitos pre-T. Expansión proliferativa maxima. Inicio de la recombinación del locus de la cadena α. Inhiben la reorganización posterior de las cadenas β.

TIMOCITOS POSITIVOS DOBLES- Son timocitos que expresan tanto CD4 como CD8 . Inducen la expresión de CCR7 que guian a la célula
desde la corteza hasta la medula. En ellos se verifica la reorganización de los genes de la
cadena α (segmentos V y J) con la expresión de los heterodimeros αβ del RLT.
Existe una exclusion alélica escaza o nula en el locus de las cadenas α con reorganizaciones productivas en ambos cromosomas de esta cadena (30% de los linfocitos maduros).
La reorganización del gen α da lugar a la eliminación del locus δ.
Estos linfocitos se hacen reactivos a los antigenos y están sometidos a selección positiva y negativa.

TIMOCITOS POSITIVOS SIMPLES- Constituyen los linfocitos que pasan con
éxito los procesos de selección expresando solo CD4 o CD8
CD4 son capaces de producir citocinas en respuesta a la estimulación antigénica posterior y expresan moléculas efectoras tipo CD40 cooperando con los linfocitos B y macrófagos.
CD8 que son capaces de producir moléculas que matan a otras células.

PROCESOS DE SELECCIÓN DE LOS LINFOCITOS T
El principal factor que determina la selección de los linfocitos T de forma positiva o negativa, es la intensidad del reconocimiento del antigeno; de modo que el reconocimiento con avidez baja produce selección positiva y el reconocimiento de avidez elevada induce la selección negativa.
En el caso de los linfocitos T inmaduros pueden reconocer cualquier antígeno (propio o extraño) mostrado por cualquier molécula del MHC (propia o extraña).

Los linfocitos T útiles son los que son específicos frente a péptidos extraños presentado por las moléculas del MHC propios.
Cuando los linfocitos positivos dobles expresan RLT αβ, entran en contacto con péptidos propios, presentado por MHC propios de células epiteliales del timo en la corteza.
La selección positiva involucra a aquellos timocitos cuyos RLT se unen con avidez baja estimulandose para sobrevivir, ademas fijandolo a restricción por MHC I o la II (areas corticales).

La selección negativa elimina los timocitos cuyos RLT se unen intensamente a antigenos peptidicos propios asociados a MHC propios (zona medular).
Si el RLT de una célula reconoce moléculas de MHC I cargadas con péptidos y al mismo tiempo se verifica la interacción con CD8 puede perder la expresión de CD4 con la posterior maduración y supervivencia de la misma hacia una T CD8+ y viceversa.
PROXIMA CLASE:ACTIVACION DE LA
CELULA T